1
Molecular Brain Research Group, Department of Physiology and Pharmacology, Robarts Research Institute, University of Western Ontario, London, Ontario, Canada
2
Institute of Neuroscience, Department of Psychology, Carleton University, Ottawa, Ontario, Canada
3
University of Ottawa Institute of Mental Health Research, Royal Ottawa Hospital and Departments of Psychology and Psychiatry, University of Ottawa, Ottawa, Ontario, Canada
4
Neuromorphological and Neuroendocrine Research Laboratory, Hungarian Academy of Sciences and Semmelweis University, Budapest, Hungary
5
Department of Clinical and Theoretical Mental Health, Semmelweis University, Budapest, Hungary
Inter-relationships ordinarily exist between mRNA expression of GABAA subunits in the frontopolar cortex (FPC) of individuals that had died suddenly from causes other than suicide. However, these correlations were largely absent in persons that had died by suicide. In the present investigation, these findings were extended by examining GABAA receptor expression patterns (of controls and depressed individuals that died by suicide) in the orbital frontal cortex (OFC), hippocampus, amygdala. locus coeruleus (LC) and paraventricular nucleus (PVN), all of which have been implicated in either depression, anxiety or stress responsivity. Using QPCR analysis, we found that in controls the inter-relations between GABAA subunits varied across brain regions, being high in the hippocampus and amygdala, intermediate in the LC, and low in the OFC and PVN. The GABAA subunit inter-relations were markedly different in persons that died by suicide, being reduced in hippocampus and amygdala, stable in the LC, but more coordinated in the OFC and to some extent in the PVN. It seems that altered brain region-specific inhibitory signaling, stemming from altered GABAA subunit coordination, are associated with depression/suicide. Although, it is unknown whether GABAA subunit re-organization was specifically tied to depression, suicide, or the accompanying distress, these data show that the coordinated expression of this transcriptome does vary depending on brain region and is plastic.
Major depressive disorder (MDD) is a complex psychiatric illness that likely involves genetic, neurochemical and experiential factors (Caspi et al., 2003
; Kendler et al., 2005
; Millan, 2006
). Although considerable evidence has pointed to a role for serotonergic processes in subserving MDD (Maes and Meltzer, 1995
; Pineyro and Blier, 1999
), several other neurochemical mechanisms have been implicated in this disorder. These have included growth factors, such as brain-derived neurotrophic factor (BDNF) (Duman and Monteggia, 2006
; Shaltiel et al., 2007
), corticotropin releasing hormone (CRH) and other peptides, including neuromedin B and somatostatin (Nemeroff, 1996
; Reul and Holsboer, 2002
; Merali et al., 2004
, 2006
). As well, there has been a resurgence in the notion that GABAA functioning may be associated with depressive illness (Tunnicliff and Malatynska, 2003
; Rupprecht et al., 2006
; Sequeira and Turecki, 2006
; Sanacora and Saricicek, 2007
; Sequeira et al., 2007
).
GABAA receptors are present in virtually all neurons of the central nervous systems, and play a fundamental role in controlling brain excitability (Fritschy and Brunig, 2003
). These receptors are constructed of pentameric protein complexes (subunits) derived from a repertoire (cassette) of 21 different proteins/genes. Commensurate with the view that GABA functioning may contribute to MDD, it has been demonstrated that GABAA receptor subunit mRNAs are either up or down regulated in association with MDD, depending on the brain region (Merali et al., 2004
; Choudary et al., 2005
; Rupprecht et al., 2006
; Sequeira and Turecki, 2006
). We have also demonstrated that in addition to frank changes of mRNA expression of GABAA subunits, it may be important to consider the expressional organization of this gene cassette in relation to pathology. Specifically, in examining the biological variability of the mRNA abundance of these subunits, we found that in the frontopolar cortex (FPC) of individuals that died suddenly of causes other than suicide, strong, positive inter-relations existed between subunit mRNA expression. The correlated expression implies that even though expression may vary, it does so in a coordinated manner, implying that the control in the relative levels of mRNA abundance is important to regulate GABAA receptor mediated inhibitory function (Merali et al., 2004
). In contrast, among depressed individuals that had died by suicide the inter-relations between these subunits within the FPC was greatly disturbed (Merali et al., 2004
). It was suggested that changes of the expression of GABAA receptor subunits might influence the timing of neuronal firing patterns, which ostensibly could have functional consequences associated with psychopathology.
In our previous work assessing “re-organization” of GABAA subunits, the analyses were restricted primarily to regions of the FPC (Merali et al., 2004
). Yet, MDD and suicide likely involves multiple brain regions. In order to determine the breadth of the apparent expression in the brain of depressed suicides, in the present investigation we expanded our analysis to several brain regions implicated in depression and/or stressor responsivity [i.e., hippocampus, paraventricular nucleus (PVN), amygdala, locus coeruleus (LC) and the dorsal vagal complex (DVC)]. As well, an analysis of GABA subunit expression and organization was undertaken within neurons of the orbital frontal cortex (OFC), as this region has been implicated as part of the neural circuitry associated with depression (Drevets, 2000
; Leyton et al., 2006
) and neurochemical functioning within this regions is altered by stressor exposure (Anisman et al., 2007
). Moreover, the OFC is thought to contribute to cognitive processes and impulsivity (Best et al., 2002
), and emotional rather than cognitive perspective-taking (Hynes et al., 2006
), which might be factors associated with suicidality. Consistent with our previous finings in the FPC, we now report that high coordination between GABAA subunits occurs in the hippocampus and amygdala of individuals that died through through causes other than suicide, whereas these inter-relations are markedly reduced in those individuals that died by suicide. However, within the OFC and PVN of individuals that died by suicide, this gene cassette’s expression may become more organized. In effect, the data indicate that inhibitory plasticity is complex and not uniformally disturbed in MDD suicide brains.
Participants
Brains from suicides and from control participants of approximately equal age, who died suddenly from causes not directly involving any diseases of the central nervous system (Age = 51.3 ± 4.00 and 45.0 ± 3.40, respectively; F(1,22) = 1.40, p = 0.24), were obtained at autopsy at the Department of Forensic Medicine of the Semmelweis University Medical School in Budapest (as described in Merali et al., 2004
). All controls and suicide victims were Caucasian from Hungary (Budapest region). In the present study we assayed expression in brains of both male and female persons who had died by suicide or who had died suddenly as a result of causes other than suicide. Table 1
provides a description of cause of death for each cohort and the N for each group (Poulter et al., 2008
).
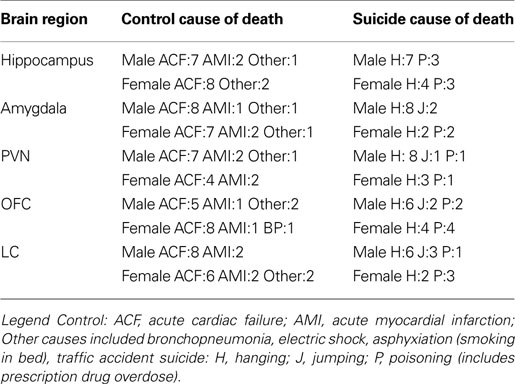
Table 1. Summary of brain regions assayed as a function of cause of death and sex in control and suicide samples.
Tissue harvesting occurred after written informed consent was obtained from next of kin, which included the request to consult the medical chart and to conduct neurochemical and/or biochemical analyses. The local ethics committee at Semmelweis and the Ethics Committees of Carleton University and the University of Western Ontario approved harvesting and analyses of the tissue samples. The ethical rules for dissecting human brains vary across countries. In some of the European countries, as in Hungary, once death is confirmed by three physicians/pathologist, the removal of the brain may proceed. In the cases of persons who were suicides or died in traffic accident, pathological sectioning, as “medicolegal cases”, is ordinarily obligatory. These brains may be removed from the skull as soon as 1–2 h post mortem, frozen and stored until the pathological sectioning. The dissection (microdissection) of the brain can be performed after pathological diagnosis has been obtained, including tests for HIV, tuberculosis, syphilis, hepatitis, alcohol and other drugs).
Suicide victims died by hanging, drug overdose or jump from height. Medical, psychiatric and drug history of suicides were obtained through chart review coupled with interviews with the attending physician/psychiatrist and family members, as previously described (Merali et al., 2004
). These interviews were semi- structured and focused on issues such as previous psychiatric history, family history of mental illness, and recent stressful experiences. In each instance a psychiatric diagnosis of depressive disorder was previously on record. The diagnoses were conducted and/or confirmed by experienced psychiatrists on the basis of DSM-IV criteria. Insofar as could be determined, the participants had not used antidepressant medication for at least 2 months prior to death and did not have a history of either drug or alcohol abuse. Toxicological tests of blood samples confirmed that drugs or alcohol were not present in cases of death by hanging or jump from height.
Examination of medical records of controls confirmed the absence of a history of psychiatric illness, alcohol or drug abuse during the last 10 years. Moreover, interviews with family members indicated that control participants had never been treated for depression, and did not have a history of alcohol abuse. Causes of death in control subjects were acute cardiac failure, myocardial infarction or traffic accident there was no other chronic diseases reported. In all instances death was sudden and did not involve a prolonged agonal state.
Tissue Collection and Dissection
Brains were obtained 1–6 h after death (M ± SEM = 3.50 ± 0.46 and 2.75 ± 0.39 h for suicide and controls; F(1, 22) = 1.88, p = 0.18). (1) the orbital frontal cortex (OFC-Brodmann area 11) (2) hippocampus (hippo) (3) Amygdala (Amg) (4) Locus Coeruleus (LC), and (5) Paraventricular nucleus (PVN). For a description of the dissection procedure used please see Supplemental Material.
Tissue Preparation and Quality Assurance
After thawing, Trizol was used to isolate total cellular RNA from cellular protein and genomic DNA as described by the manufacturers protocol (Invitrogen, Canada). The samples were verified to be free of contaminating DNA as no signal originated from genomic DNA among the no reverse transcription controls. Isolated RNA was checked for purity by ensuring that the OD 260/280 ratio was greater than 1.8. An analysis of the RNA quality using an Agilent BioAnalyzer showed the RNA integrity number (RIN) for Control was 5.6 ± 0.3 (range 5.2–6.9), whereas suicide samples the RIN was 5.8 ± 0.2 (F < 1.0) (range 5.2–6.3). Although these values indicate some degradation, it has been shown that this quality of RIN stills yields valid data particularly when normalized to a reference such as has been done here (Fleige and Pfaffl, 2006
; Fleige et al., 2006
). A correlation analysis of the RIN versus the cycle threshold (Ct) of synaptophysin was –0.26 and 0.23 for control and suicide, respectively. Brain pH was found to be 6.45 ± 0.26 for controls and 6.58 ± 0.26 for individuals that died by suicide (mean ± SEM, F < 1). In addition, the Budapest brain bank, as a member of the European Brain Bank Consortium (BrainNet II Europe) had two member institutions (Imperial College, London and Universitat de Barcelona) perform nucleic acid stability tests on 50 tissue samples from the Budapest brain bank. These samples, taken from 16 different human brains that included five brains represented in the present sample, showed little degradation and were termed highly suitable for studies of this kind.
QPCR Analysis
For QPCR, SYBR green detection was used according to the manufacturer’s protocol (BioRad QPCR kit). A Stratagene MX-4000 real time thermocycler was used to collect the data. All PCR primer pairs used generated amplicons between 90 and 110 base pairs. Amplicon identity was checked by restriction analysis. Primer efficiency was measured from the slope relation between absolute copy number or RNA quantity and the Ct using the MX-4000 software. All human primer pairs had a minimum of 93% percent efficiency. Human primers have been previously published (Merali et al., 2004
). We chose synaptophysin as a reference gene since it is expressed only in neurons, thus subunit expression is normalized to the neuronal content of the sample. Furthermore, there is no report that has shown in the mature brain that synaptophysin expression is labile. In fact one study has shown that even after severe brain perturbations synaptophysin is stably expressed (Chen et al., 2001
). Nevertheless, a second reference gene, cyclophylin-A (cyp) was used to confirm that differences in the GABAA subunit mRNAs within the OFC were, in fact, not related to non-specific factors. Regardless of whether synaptophysin or cyp was used as a reference gene, the observed outcomes were the same.
The expression of each species was normalized by subtracting its Ct from the synaptophysin Ct. The normalized Ct values (denoted as Ctn) for each mRNA species were averaged for each brain region in both Control and Suicide groups. Because synaptophysin is a higher abundance mRNA species compared to the GABA receptor transcripts, the Ctn is usually negative. Therefore, a Ctn value that is less (more negative) than another Ctn, indicates a relatively lower abundance of that mRNA species and vice versa. Although the changes in some species were less than a cycle, where statistically significant these differences were still larger than the standard error of the average Ctn. We have previously shown the subject-to-subject variability of the synaptophysin Ct’s was unrelated to the post-mortem harvest time (Merali et al., 2004
), and this was again found to be the case in the present investigation using a larger N. We reverse transcribed an equivalent amount of total RNA for both groups (2 µg). Then an aliquot of this RT reaction was used for QPCR analysis (10 nanogram equivalent). These reactions were run in parallel, with internal standards, for all transcripts.
Statistical Analyses
Data were analyzed by 2 (Suicide vs. Control) × 2 (Male vs. Female) between-groups analyses of variance (ANOVA) independently for each brain region and GABAA subunit. In some brain regions, where the N for females was less than 5, males and females were pooled for analysis. Follow-up tests comprised t-tests using a Bonferonni correction to maintain α at 0.05. Where linear correlations were conducted to assess the relationship between outcome variables, or between dependent and independent variables (e.g., age, time to autopsy), these were done by Pearson product moment correlations. As many correlations were conducted, the α level for statistical significance was adjusted to <0.025. Since the GABAA subunits were not found to vary as a function of sex, data for the two sexes were pooled for the correlational analyses. χ2 analyses were conducted to determine whether the frequency of significant subunit correlations differed between groups. As several analyses involved outliers that might have influenced the outcomes, separate Spearman rank order correlations were conducted. Irrespective of whether parametric or nonparametric analyses were performed, the outcomes were the same.
mRNA Content and Interrelations in Depressed Suicides vs. Control
Hippocampal receptor subunit mRNA expression did not vary as a function of the “Cause of death”, Sex or the interaction between these variables. However, the correlations evident between the subunits varied appreciably in the controls and suicides. Specifically, in control hippocampus, 12 transcript pairs were significantly correlated, all of which are highly expressed in the hippocampus (Figure 1
C). In contrast, the frequency of significant correlations in the hippocampus (below the diagonal in Figure 1
C) was lower in suicides, as only 6 correlations were significant (χ2 = 6.22; p < 0.025). The reduction of subunit correlations in individuals that died by suicide was most notable in the case of the α2 and α3 subunit interrelations, which declined from 3 to 1 and 5 to 1, respectively. Although the relations between subunits were lower in the suicide brain, those correlations that were significant in the suicide hippocampus were also been found to be significant in control samples (see Figure 1
). Thus, the change of significant correlations was not simply a random rearrangement of significant outcomes.
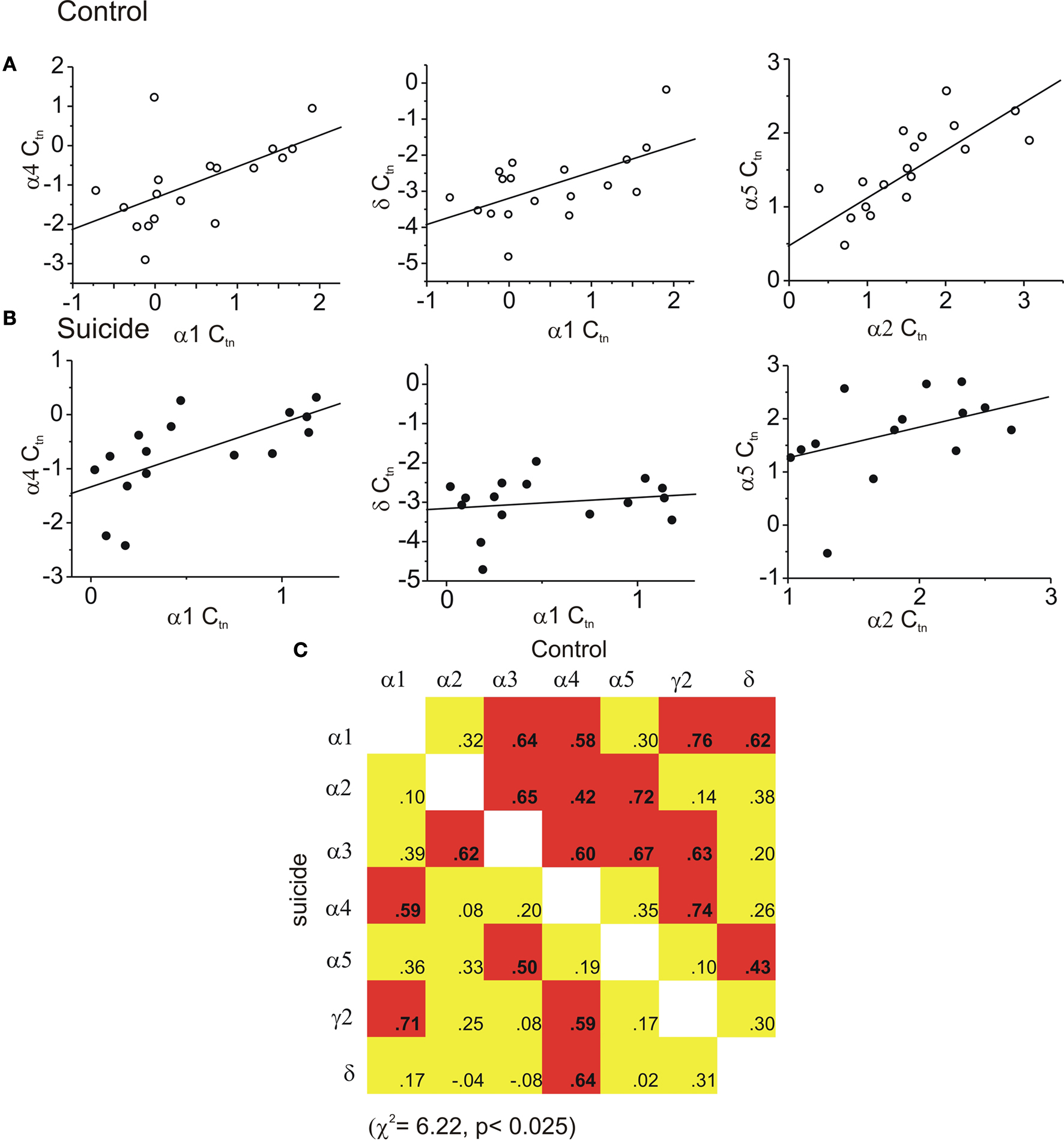
Figure 1. Coordinated expression decreased in the hippocampus of suicides. In (A) regression graphs show examples of three significant correlated subunit pairs of controls that were not significantly correlated in suicide hippocampus shown in (B). In suicide hippocampus (lower portion of the correlation matrix; the frequency of inter-relations was diminished. In the matrix (C) all possible correlations are shown. In control brain (upper portion of the correlation matrix) thirteen subunit pairs were significantly correlated. (red indicates a statistically significant positive correlation was found, yellow indicates a non significant correlation).
Figures 1
A,B also shows several representative scatter plots between GABA subunits, in this case between the α1 and δ, α1 and α4, and the α2 and α5 subunits. As shown in the figure, the relation between the α1 and α4 subunits was high in the controls (upper panel) and was equally high in the suicide brain. However, the relations between the α1 and δ, and the α2 and α5 subunits evident in control subjects was markedly lower in those that died by suicide (lower panels).
In the amygdala, with the exception of the δ subunit, neither the Cause of death, Sex, nor the Cause of death × Sex interaction was associated with subunit changes. The δ subunit of suicides was moderately, although significantly, down-regulated by 0.6 cycles in the suicides, F(1,29) = 5.73 p < 0.05 (Control −1.8 ± 0.2 versus suicide −2.4 ± 0.1). The correlational analyses indicated that within the amygdala of controls, eight regressions were significantly related. Of these, the α4 subunit was correlated to four other subunits’ expression, including α1, α2,γ2 and δ (Figure 2
C), and the α1 subunit was related to α4, γ2 and δ subunits. In persons that died by suicide, significant relations appeared less frequently than in controls (χ2 = 4.75; p < 0.05), as only 2 of the correlations were significant. The scatter plots shown in Figures 2
A,B and again depict several correlations that were significant in controls, but were not evident in suicide samples.
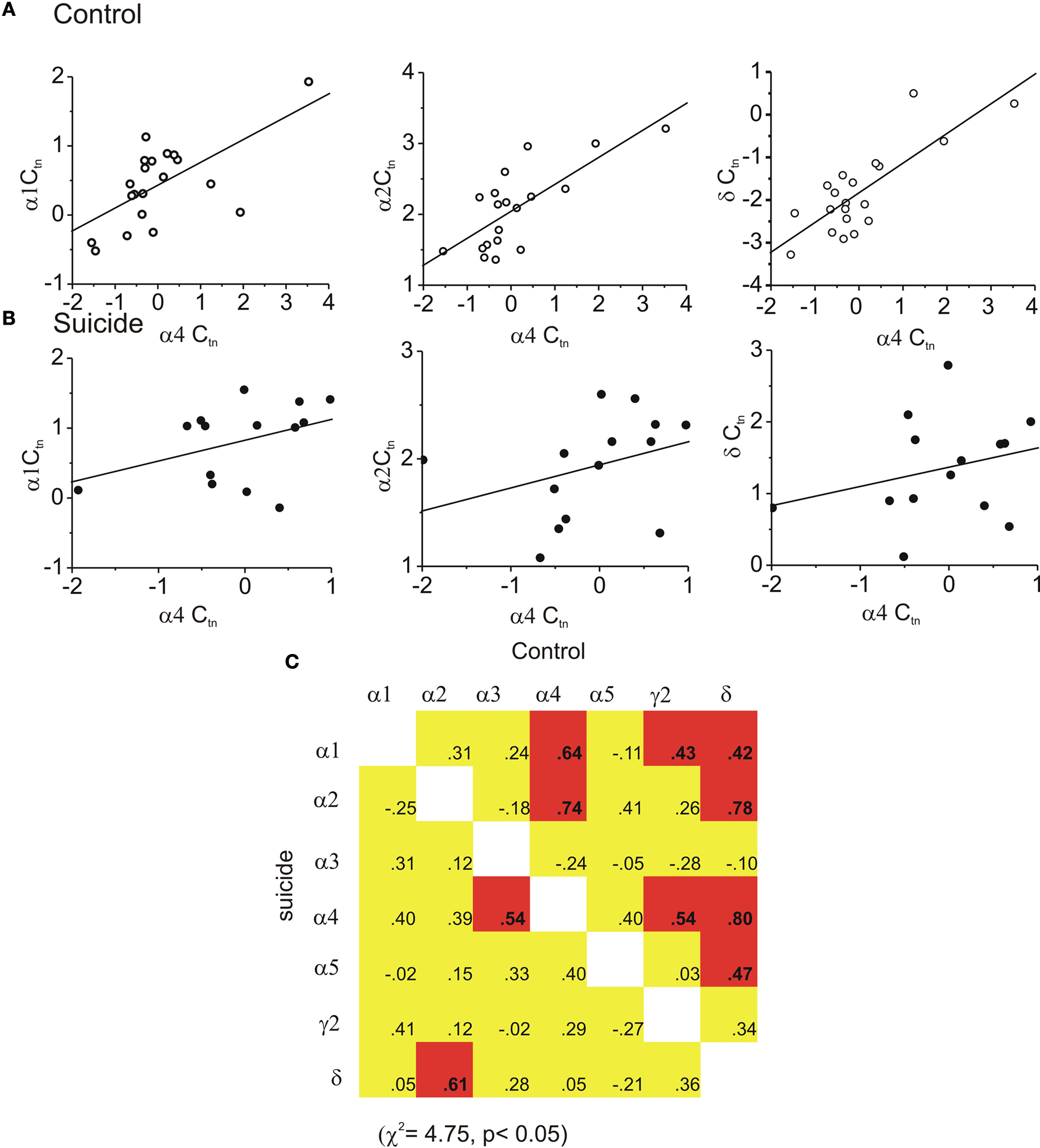
Figure 2. Coordinated expression is decreased in the amygdala of suicides. In (A) the regression graphs show examples of three significantly correlated subunit pairs among controls that were not significantly correlated in suicide amygdala shown in (B). In (C) the matrix of all possible correlations are shown. In control brain upper portion of matrix eight subunit pairs were significantly correlated. In suicide hippocampus the frequency of inter-relations was diminished. (Red indicates a statistically significant positive correlation was found yellow indicates a non significant correlation).
Within the OFC only two statistically significant differences in subunit expression were found. First, α4 subunit expression was higher in female suicides (by approximately 1.0 cycle) than in female controls, F(1,29) = 5.27, p < 0.03 (Control female −0.6 ± 0.2 versus Suicide female 0.5 ± 0.3). The δ subunit was also modestly, but significantly, increased in both male and female suicides by about 0.5 cycles F(1,29) = 5.10, p < 0.04 (Control −2.3 ± 0.2 versus Suicide −1.8 ± 0.1). Unlike the effects seen in the hippocampus and amygdala, within the OFC the frequency of significant correlations in control brains was relatively low (3 of 21 were significant) and increased in the suicides (Figure 3
C, 8 of 21 were significant, χ2 = 5.45; p < 0.025). Figures 3
A,B shows examples of the regressions between three pairs of subunits (α1, α2and γ2), in control brain where only the α1 versus γ2 combination were correlated. These relationships were markedly increased in suicide brain. In effect, unlike the previous findings in the FPC, and the present hippocampal and amygdala findings, expression patterns were more organized in suicide OFC. The pattern of correlations evident was not likely a reflection of spurious relations, as those correlations that were significant in controls remained so in the suicide samples.
Figure 3. Coordinated expression of GABAA subunits was increased in the OFC of suicides. In (A) three regression graphs are shown, for control tissue, where only the α1 versus γ2 subunits are correlated (middle panel). In (B) (suicide tissue) each of these relationships were significantly related. In (C) the matrix of all possible correlations are shown. In control brain only three subunit pairs were significantly correlated (upper portion of the correlation matrix). However, the frequency of inter-relations was significantly higher in suicide brain (lower portion of the correlation matrix). (Red indicates a statistically significant positive correlation was found yellow indicates a non significant correlation while green indicates a statistically relevant negative correlation was found).
There were no differences between suicides and controls in the abundance of any subunit mRNAs in the PVN. As in the OFC, within the PVN significant inter-relations between the subunits was infrequent (3 of 21 correlations were significant) in individuals that had died by causes unrelated to suicide (see Figure 4
C). As well, unlike the relation between GABAA subunits previously observed within other prefrontal cortical structures, the interrelationships of the subunit expression in the samples of persons who died by suicide actually were more coordinated, with 8 of 21 inter- relations correlating (positively) significantly with one another. This included the 3 correlations that had been significant in controls, plus 5 others that had not been significant previously. The difference of significant correlations between the suicides and controls was statistically significant (χ2 = 13.80; p < 0.01). Figure 4
also shows that several of the subunit relations that had not been significant in controls but were highly significant in samples from individuals that died by suicide.
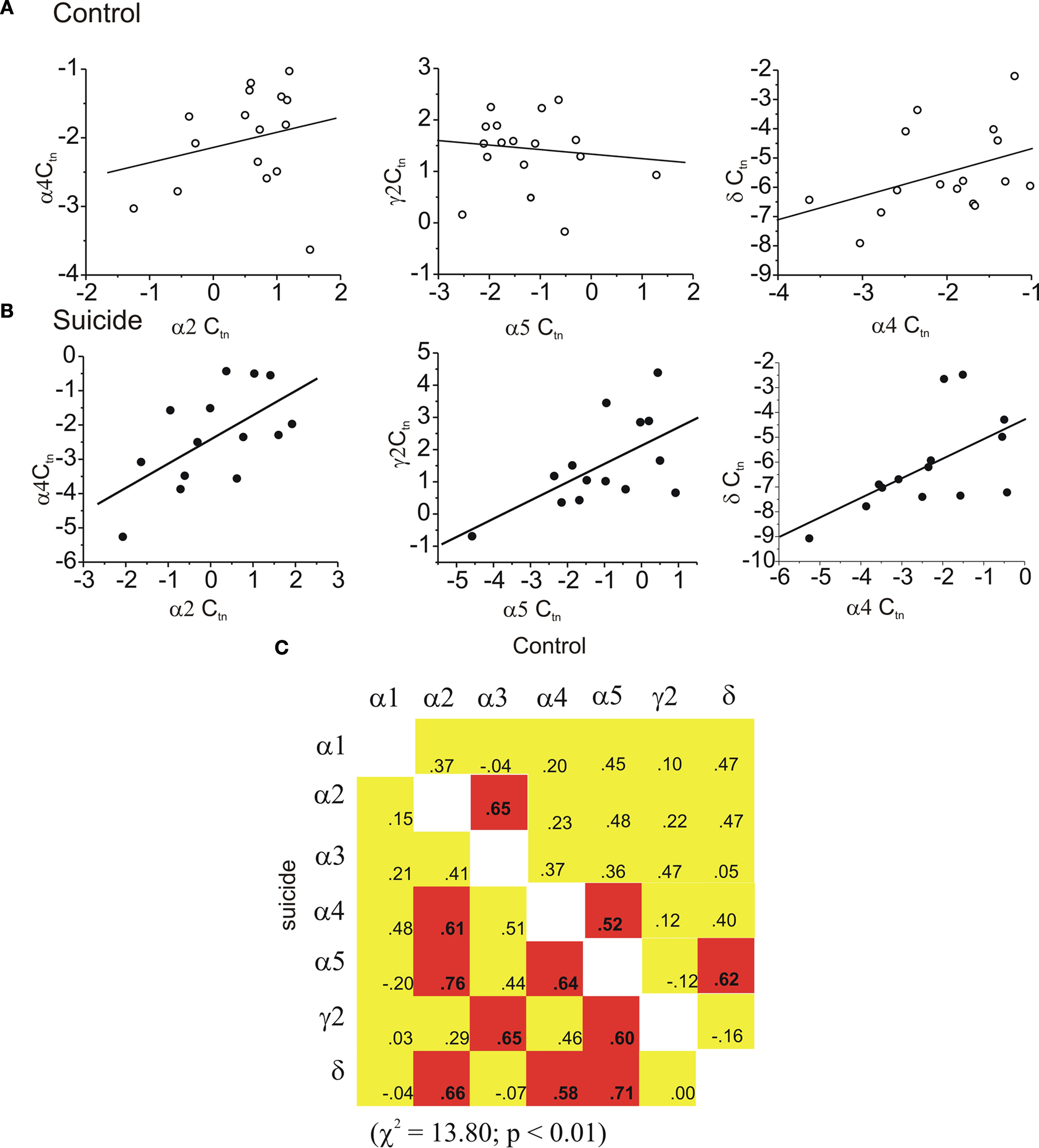
Figure 4. Coordinated expression was increased in the PVN of suicides. In (A) regression graphs show examples of three uncorrelated subunit pairs in the PVN of controls that were significantly correlated in suicide PVN shown in (B). In (C) the matrix of all possible correlations are shown. In control brain three pairs were found to be significantly correlated (upper portion of the correlation matrix). In suicide PVN the frequency of inter-relations was increased to 8. (red indicates a statistically significant positive correlation was found yellow indicates a non significant correlation).
Finally, the subunit expression of controls and persons that died by suicide did not differ within the LC. Furthermore, the correlations between subunits indicated that of the 8 significant interrelations found in controls, 7 were stable, also being evident in the suicide samples. One relation between expression patterns was lost, namely α4 versus α5 between control and suicide, whereas the α4 versus γ2 relation became significant in suicides.
GABAA receptor expression is highly heterogeneous, varying across brain regions as well as at the cellular and subcellular level (Fritschy and Brunig, 2003
). In the present investigation, we demonstrated that the expression of few GABAA receptor subunits differed between tissue samples that came from individuals that had died by suicide vs. those that died quickly from factors unrelated to suicide. The GABAA receptor subunit mRNA abundance in the hippocampus and amygdala of controls and individuals that died by suicide differed modestly, but were evident with respect to the α4 and δ subunits. Interestingly, these two subunits are thought to play a central role in tonic (as opposed to phasic) inhibition, suggesting that the balance between these two modes of inhibition may be altered.
Just as we previously observed in the FPC [10], in the present investigation it was found that the GABAA subunit coordination was disturbed in both the hippocampus and amygdala. In contrast, however, in the OFC and the PVN, the modest number of significant inter-relations ordinarily evident in control samples was significantly increased in samples from persons that had died by suicide. These finding expand on our original observations, but indicate that regulation of GABAA subunit coordination across brain regions may involve different processes and/or have diverse implications for depression/suicide.
The presence of coordinated GABA subunit expression was previously observed in studies of epilepsy in both rats and humans, in which GABAA receptor mRNA abundance were coordinately expressed, particularly α1 and γ2 as well as α4 and δ (Brooks-Kayal et al., 1998
, 1999
). The biological significance of coordinated regulation of receptors and ion channels (like gap junctions and potassium channels) that are constructed from a heterogeneous family of proteins is not clear, but a few speculative interpretations are warranted. Transcriptionally regulating relative mRNA abundance would be a mechanism that ensures a proportional abundance of protein. It would, after all, be energetically wasteful to produce 10 times more of one protein than another if the stoichiometry of their assembly is 1:1. Specifically, for heteromeric proteins, like GABAA receptors, that have highly variable functionality, it may be particularly important to have the correct balance of receptors involved in, for example, phasic versus tonic inhibition. Another reason may be to have the correct balance of synaptic timing as some receptors give rise to fast synaptic currents (α1) while others are relatively slow (α5). So, the expression of one subunit with a particular physiological characteristic may be balanced by the expression of another with differing behaviour. In agreement with this speculation, a genome wide analysis of both mRNA and protein expression in S. cerevisiae showed that functionally related proteins were coordinately expressed and that mRNA expression was correlated with protein abundance (Washburn et al., 2003
). Thus, the alterations in expressional patterns of a functionally related gene cassette, as shown here, is not unprecedented and it is certainly possible that changing subunit patterns might have an impact on behaviour.
It is uncertain what processes govern the coordinated subunit expression, especially as the genes for various GABAA subunits appear on different chromosomes, making it unlikely that chromosomal location (linkage) plays a role in this respect. However, it seems likely that a complex array of factors are responsible for the observed region-specific differences in protein expression. Why in some regions these relations decreased in persons that died by suicide, whereas in other regions the frequency of these relations actually increased is not clear. However, it is unlikely that the many correlations found between the subunits reflect an unidentified systematic error (related, perhaps, to primer specificity or RNA quality) and/or merely random correlations that would arise given the large number of correlations conducted. Had some systematic errors been present, then the same outcome ought to have been evident in all brains regions, which was clearly not the case. Moreover, in brain regions in which suicide was associated with reduced correlations, those relations that had been non-significant in controls remained non-significant in suicides, and conversely, in regions in which the correlations were more frequent in suicide than control conditions, those subunit relations that had been significant in control samples remained significant in suicide samples. Thus the correlations do not seem to arise sporadically.
The present findings updates our previous report that described changes in subunit coordination (from frequent to infrequent coordinated expression) in controls vs individuals that died by suicide. In that report, we used the pejorative term “dysregulated” Merali et al. (2004)
. However, given that expression of the subunits within the OFC and PVN actually became more organized in persons who died by suicide, we should probably describe these changes as “re-organized” rather than “dysregulated”. In effect, this report corrects the assertion that normal healthy brain has tightly controlled expression (a high frequency of significant correlations), whereas few correlations are present in the diseased brain. Re-organisation better describes a situation where the level of expression may transition from being highly divergent (very few correlation patterns) to one where there is an apparent high degree of coordination, or vice versa. Such transitions may reflect the plasticity associated with induction of disease or it may reflect the brain’s “attempts” to compensate for some other imbalance.
Defining the regulation of GABA receptor promoters is still at an early stage, but the basic transcriptional “machinery” for several subunits (α1,α2,α3,α4,α5,α6,β1,β3,γ2 and δ) has been described in several reports (Kang et al., 1994
; Kim et al., 1997
; Penschuck et al., 1997
; Mu and Burt, 1999
; McLean et al., 2000
; Russek et al., 2000
; Fuchs and Celepirovic, 2002
; Ma et al., 2004
; Urak et al., 2006
). Interestingly, using a computational analyses, several potential common transcriptional factor(s) have been identified for the α subunits (Steiger and Russek, 2004
). These included early growth response factors 1 and 3 (Egr 1,3), nerve growth factor protein C (NGFIC), Myc associated growth factor (MAZ) and zinc binding protein factors 89 and 9 (ZBPF 89 and ZF9, respectively). Although many of these sites were clustered, raising the possibility that they might function together, only Egr 3, which is able to activate the expression of the α4 subunit was shown to be functional. The finding of the present investigation that the expression of this subunit was diminished in the OFC of persons that died by suicide, and was also previously found to be low in the FPC, is consistent with the speculation the Egr might be related to depression/suicide. Parenthetically, the finding that Erg 3 is influenced by BDNF (Roberts et al., 2006
), and BDNF expression may be reduced among depressed individuals (Dwivedi et al., 2003
), is consistent with this perspective.
Our data focused primarily on the control of the GABAA receptor gene cassette at the “mRNA level”. However, we have observed that the α1 subunit protein content is correlated with the mRNA amounts in the FPC (r = 0.77 p < 0.03 n = 9 control = 4 and suicide = 5), but obviously, it would be preferable if this could be done for all subunits and their interrelations examined. To do so requires antibodies having identical sensitivity (analogous to PCR primers having similar (high) efficiency) for each subunit, so this type of analysis is not presently possible. Nevertheless, the fact that the coordination of the various subunit transcripts were re-organized, even when the transcript levels were unaltered, raises the possibility that depression/suicide may be more closely aligned with the organization of processes controlling the relative amounts of subunits rather than the actual mRNA expression and corresponding protein abundance.
The fundamental question that arises concerns the potential influence of the subunit changes on neuronal functioning. Ordinarily, the activity of neural networks or firing patterns involves cellular characteristics (for example, channel densities, calcium buffering, cell morphology) and network parameters (distribution of neurons, number and location of synaptic contacts). Based on computational models that varied these and several other parameters, it was shown that the timing of neural networks could function within a wide range circuit parameters (Prinz et al., 2004
). Despite the resiliency of this system, it was recognized that critical interrelationships exist between certain parameters (for example synaptic strength), and being outside of a particular range of values resulted in the generation of aberrant rhythms. (Prinz et al., 2004
) In the context of the present investigation, it might be expected that variations in the relative amounts of α subunits or in their coordination may influence the stoichiometric ratios of α subunits comprising GABAA receptors. The altered subunit composition influences the duration of inhibitory currents (Hájos and Mody, 1997
; Haas and Macdonald, 1999
; Hutcheon et al., 2004
; Ponomarenko et al., 2004
) that are fundamental in regulating network timing patterns (Traub et al., 1996
; Wang and Buzsaki, 1996
; Zhang et al., 1998
; Sohal et al., 2000
; Ponomarenko et al., 2004
) and hence may influence behavioral outcomes. In effect, through re-organized subunit expression inhibitory synaptic behavior may be influenced to the extent that network timing is altered in stress reactive brain regions, culminating in MDD. This hypothesis is similar to one posed for schizophrenia, in which the expression of reelin, a protein important for the formation of inhibitory synapses, has been shown to be down regulated in human schizophrenia as well as in a mouse model of this disorder (Dong et al., 2005
; Grayson et al., 2006
). Thus it was suggested that a miswiring of inhibitory circuits might be an underlying cause of schizophrenia.
It is recognized that conclusions that can be drawn on the basis of the present investigation are subject to several limitations. Among other things, although individuals that had died by suicide had a history of depression, it is not necessarily the case that the present findings are generalizable to depression where suicidal intent was not present, nor is it the case that suicide itself (i.e., either the intent or the act) was without effect. Furthermore, although the sample size in the present investigation was in line with most studies that assessed biological correlates of depression/suicide, the number of participants was still relatively limited. Finally, given that manipulations of mood states were not possible in this type of study (i.e., the data are correlational in nature), and hence causal attributions between depression/suicide and GABAA subunit inter-relations are not possible. In this respect, the present findings, although informative, do not shed light on whether the altered GABAA subunit coordination stems from depression, the stressful experiences that may have provoked MDD (or for that matter the stress associated with being depressed), or as a consequence of the suicide act itself (see discussion in van Heeringen, 2001
).
Despite these limitations, it appears that GABAA receptor expression is coordinated in a region specific manner in human brain, and this organisation is often altered in depressed suicides, particularly in highly stressor-reactive regions, such as the amygdala and hippocampus. The goal now will be to identify, at the molecular level, the signalling pathways/mechanisms that control these gene expression patterns and the processes that lead to altered coordination profiles, and to determine whether altered coordination of the GABAA gene cassette is apparent in depressed persons that had died as a result of factors other than suicide.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
This research was supported by a CIHR grant to Michael O. Poulter, Hymie Anisman and Zul Merali, EU Grant FP6, BNEII No. LSHM-CT-2004-503039 to MP. A NARSAD Independent Researcher Award also supports Michael O. Poulter. Hymie Anisman is a Tier I Canada Research Chair.
The Supplementary Material for this article can be found online at http://www.frontiersin.org/molecularneuroscience/paper/10.3389/neuro.02/003.2010/
Choudary, P. V., Molnar, M., Evans, S. J., Tomita, H., Li, J. Z., Vawter, M. P., Myers, R. M., Bunney, W. E. Jr., Akil, H., Watson, S. J., and Jones, E. G. (2005). Altered cortical glutamatergic and GABAergic signal transmission with glial involvement in depression. Proc. Natl. Acad. Sci U.S.A. 102, 15653–15658.
Roberts, D. S., Hu, Y., Lund, I. V., Brooks-Kayal, A. R., and Russek, S. J. (2006). Brain-derived neurotrophic factor (BDNF)-induced synthesis of early growth response factor 3 (Egr3) controls the levels of type A GABA receptor alpha 4 subunits in hippocampal neurons. J Biol. Chem. 281, 29431–29435.