1
Graduate School of Science, Nagoya University, Nagoya, Japan
2
Department of Pharmacology, The School of Pharmacy, London, UK
3
Department of Molecular, Cellular and Developmental Biology, University of Michigan, Ann Arbor, MI, USA
Glycine is a major inhibitory neurotransmitter in the spinal cord and brainstem. Recently, in vivo analysis of glycinergic synaptic transmission has been pursued in zebrafish using molecular genetics. An ENU mutagenesis screen identified two behavioral mutants that are defective in glycinergic synaptic transmission. Zebrafish bandoneon (beo) mutants have a defect in glrbb, one of the duplicated glycine receptor (GlyR) β subunit genes. These mutants exhibit a loss of glycinergic synaptic transmission due to a lack of synaptic aggregation of GlyRs. Due to the consequent loss of reciprocal inhibition of motor circuits between the two sides of the spinal cord, motor neurons activate simultaneously on both sides resulting in bilateral contraction of axial muscles of beo mutants, eliciting the so-called ‘accordion’ phenotype. Similar defects in GlyR subunit genes have been observed in several mammals and are the basis for human hyperekplexia/startle disease. By contrast, zebrafish shocked (sho) mutants have a defect in slc6a9, encoding GlyT1, a glycine transporter that is expressed by astroglial cells surrounding the glycinergic synapse in the hindbrain and spinal cord. GlyT1 mediates rapid uptake of glycine from the synaptic cleft, terminating synaptic transmission. In zebrafish sho mutants, there appears to be elevated extracellular glycine resulting in persistent inhibition of postsynaptic neurons and subsequent reduced motility, causing the ‘twitch-once’ phenotype. We review current knowledge regarding zebrafish ‘accordion’ and ‘twitch-once’ mutants, including beo and sho, and report the identification of a new α2 subunit that revises the phylogeny of zebrafish GlyRs.
Glycine receptors (GlyRs) and GABAA receptors are pentameric ligand-gated chloride channels (reviewed in Moss and Smart, 2001
) that mediate inhibitory synaptic transmission. In the vertebrate CNS, GABAergic synaptic transmission is mainly used in the brain, while glycinergic synaptic transmission operates in the brainstem and spinal cord to regulate motor systems. Although GlyRs are found in all vertebrates and perhaps selected invertebrates (Kehoe et al., 2009
), mammalian GlyRs have been studied most extensively. Inherited defects in genes encoding the major adult α1β GlyR causes startle disease/hyperekplexia in humans, characterized by noise or touch-induced seizures that result in muscle stiffness and life-threatening neonatal apnea episodes ( Sament and Schwartz, 1957
; Kirstein and Silfverskiold, 1958
; Bakker et al., 2006
; Harvey et al., 2008
). The biological roles of GlyRs containing the α2, α3 and α4 subunits are less clear, although GlyR α3 is clearly linked to inflammatory pain pathways (Harvey et al., 2004
). Glycine also binds to the NR1 subunit of the NMDA receptor, acting as an essential coagonist for excitatory synaptic transmission mediated by glutamate (Johnson and Ascher, 1987
; Moriyoshi et al., 1991
; Kuryatov et al., 1994
). Glycine can also activate the NR3B-containing NMDA receptor in the absence of L-glutamate in heterologous expression systems (Chatterton et al., 2002
). However, the in vivo physiological significance of glycine-mediated excitatory synaptic transmission is unclear.
Two glycine transporters (GlyTs) also regulate glycinergic synaptic transmission (Eulenburg et al., 2005
). GlyTs are thought to take up glycine from the synaptic cleft to terminate glycine-mediated synaptic transmission (GlyT1) and resupply glycine to glycinergic presynaptic terminals (GlyT2). Mouse models of GlyT1 dysfunction exhibit severe motor deficits accompanied by lethargy, hypotonia and hyporesponsivity, and die within 6–14 h after birth as a result of respiratory failure, although wasting and dehydration caused by an inability to suckle may also play a role (Gomeza et al., 2003a
; Tsai et al., 2004
). Curiously, these symptoms resemble glycine encephalopathy, a disease associated with disruption of the mitochondrial glycine cleavage system, which degrades excess glycine (Applegarth and Toone, 2006
). Mutations in the GlyT2 gene cause startle disease/hyperekplexia in humans and congenital muscular dystonia type 2 (CMD2) in cattle (Rees et al., 2006
; Charlier et al., 2008
; Harvey et al., 2008
).
Intensive examination of the small freshwater fish, zebrafish, has occurred in the past several decades due to the optical clarity and accessibility of zebrafish embryos and amenability to genetic strategies (Streisinger et al., 1981
; Eisen et al., 1986
; Driever et al., 1996
; Haffter et al., 1996
). These studies have enhanced our understanding of the in vivo function of genes involved in the regulation of glycinergic synaptic transmission. Two zebrafish mutations, one defective in GlyR function (bandoneon) and the other in GlyT1 function (shocked), were isolated from behavioral mutagenesis screens (Granato et al., 1996
; Cui et al., 2004
, 2005
; Luna et al., 2004
; Hirata et al., 2005
; Masino and Fetcho, 2005
; Mongeon et al., 2008
). Importantly, these zebrafish mutants show physiological and behavioral defects similar to non-human mammalian mutants as well as humans with startle disease and glycine encephalopathy. Thus, zebrafish mutants serve as attractive vertebrate models for childhood neurological disorders. In this review, we discuss the history of the zebrafish mutants bandoneon and shocked and report the identification of new GlyR cDNA sequences that revise the phylogeny of zebrafish GlyRs.
Zebrafish (Danio rerio) have several advantages for the analysis of vertebrate development (Grunwald and Eisen, 2002
). First, raising zebrafish is easy. A pair of adult zebrafish can generate 100–200 fertilized eggs in a single spawn. The generation time is 3 months, which is comparable to other vertebrate models such as mice. It is neither expensive nor difficult to maintain thousands of zebrafish in a laboratory. Second, all stages of development occur externally and rapidly, with most organs formed by 5 days of fertilization (Kimmel et al., 1995
). The fast pace of development allows one to analyze development in living zebrafish. Third, the embryos are transparent, which makes them amenable for live imaging of individual cells deep within the body such as neurons in the brain. In fact, a number of transgenic zebrafish that express GFP (green fluorescent protein), RFP (red fluorescent protein) or calcium indicators under control of tissue-specific, Gal4-inducible or stress-inducible promoters has been generated to visually monitor the development of tissues and stress response as well as neuronal activity (Amsterdam et al., 1995
; Peters et al., 1995
; Higashijima et al., 1997
, 2003
; Long et al., 1997
; Scheer and Campos-Ortega, 1999
; Halloran et al., 2000
). Fourth, the electrophysiological activity of neurons and muscles in zebrafish embryos can be recorded using standard current-clamp and voltage-clamp methods (Prugh et al., 1982
; Grunwald et al., 1988
; Legendre and Korn, 1995
; Drapeau et al., 1999
; Fetcho, 2007
). Using these methods, the properties of neural circuits that underlie the earliest zebrafish behaviors are beginning to be clarified (Legendre and Korn, 1994
; Ribera and Nüsslein-Volhard, 1998
; Neuhauss et al., 1999
; Ono et al., 2001
; Saint-Amant and Drapeau, 2001
; Sidi et al., 2003
; Kimura et al., 2006
; McLean et al., 2007
; Tanimoto et al., 2009
).
The aforementioned advantages make the zebrafish an excellent system for examining vertebrate development. However, it is the genetic manipulability of zebrafish that has attracted the most attention of biologists (Grunwald and Eisen, 2002
; Amsterdam and Hopkins, 2006
). Methods for mutagenesis of zebrafish were established in the early 1990’s (Mullins et al., 1994
) with two large-scale mutant screens completed in Tübingen, Germany and Boston, USA by 1996 (Driever et al., 1996
; Haffter et al., 1996
). These screens used chemical mutagens and identified more than 4,000 mutants to kick-start large-scale mutagenesis analysis of a vertebrate embryo. The advent of the zebrafish genome sequencing project in 2001
1
has greatly improved the molecular identification of chemically-induced mutations. Retrovirus- and transposon- mediated gene disruption was also developed to generate zebrafish mutants in a systematic manner (Lin et al., 1994
; Gaiano et al., 1996a
,b
; Amsterdam et al., 1999
, 2004
; Kawakami et al., 2000
, 2004
; Golling et al., 2002
; Sivasubbu et al., 2006
; Wang et al., 2007
). In addition to these forward genetic approaches, targeting-induced local lesion in genome, combining ENU mutagenesis with large-scale exon sequencing has made it possible to inactivate selected zebrafish genes (Wienholds et al., 2002
). More recently, a gene-targeting technique using designed sequence-specific zinc-finger proteins has been demonstrated to be effective in disrupting key genes in zebrafish (Doyon et al., 2008
; Meng et al., 2008
). Genes can also be knocked down during embryonic stages by the injection of antisense morpholino oligonucleotides into recently fertilized embryos (Nasevicius and Ekker, 2000
). These highly stable oligonucleotides can effectively block translation or splicing of a target mRNA to interfere with gene function in vivo. Splice-site morpholinos have the major advantage that their efficacy can be monitored by RT-PCR.
Transgenesis is also a powerful tool for analysis of development and gene function in zebrafish. The first zebrafish transgenic lines were generated by injection of DNA into embryos (Stuart et al., 1988
, 1990
; Culp et al., 1991
; Bayer and Campos-Ortega, 1992
). Transgenic zebrafish exhibiting cell-specific or inducible gene expression have proven extremely useful for in vivo analysis of gene function and developmental processes (Higashijima et al., 1997
; Long et al., 1997
; Halloran et al., 2000
). More recently, virus- and transposon-mediated transgenesis methods have greatly improved the efficiency of generating transgenic zebrafish (Davidson et al., 2003
; Kawakami et al., 2004
; Ellingsen et al., 2005
; Sivasubbu et al., 2006
; Kwan et al., 2007
; Villefranc et al., 2007
). The increased efficiency has significantly enhanced the application of powerful controlled expression methods such as the Gal4/UAS system (Scheer and Campos-Ortega, 1999
; Inbal et al., 2006
; Scott et al., 2007
; Asakawa et al., 2008
; Halpern et al., 2008
).
Recent technical advances make zebrafish an attractive complement to invertebrate systems such as C. elegans and Drosophila on the one hand and the transgenic, knockout and knock-in mice on the other. Indeed, zebrafish mutant and transgenic embryos are useful for in vivo high-throughput chemical screening, thereby enabling discovery of novel pharmaceutical reagents that may be useful for mitigating human diseases (Peterson et al., 2000
, 2004
; Stern et al., 2005
).
Zebrafish exhibit three distinct behaviors during embryogenesis; spontaneous coiling, touch-evoked escape contractions and swimming. Spontaneous coiling appears after 17 hours postfertilization (hpf) and consists of side-to-side alternating contractions of the axial muscles in the trunk and tail (Figure 1
A; Saint-Amant and Drapeau, 1998
; Downes and Granato, 2006
; Pietri et al., 2009
). This relatively slow coiling is independent of sensory stimulation, with the frequency of spontaneous coiling peaking at 0.3∼1 Hz at 19 hpf and gradually declining to less than 0.1 Hz by 26 hpf. Thus, locomotor circuits are functional as early as 17 hpf. Interestingly, the isolated trunk and tail following transections between somites 5 and 7 also displays spontaneous coiling with a similar time course and frequency compared to intact embryos (Downes and Granato, 2006
). However, transections removing the first 10 somites eliminate all spontaneous activity (Pietri et al., 2009
). These experiments suggest that the neural network triggering spontaneous coiling is located in the rostral spinal cord between somites 5 and 10.
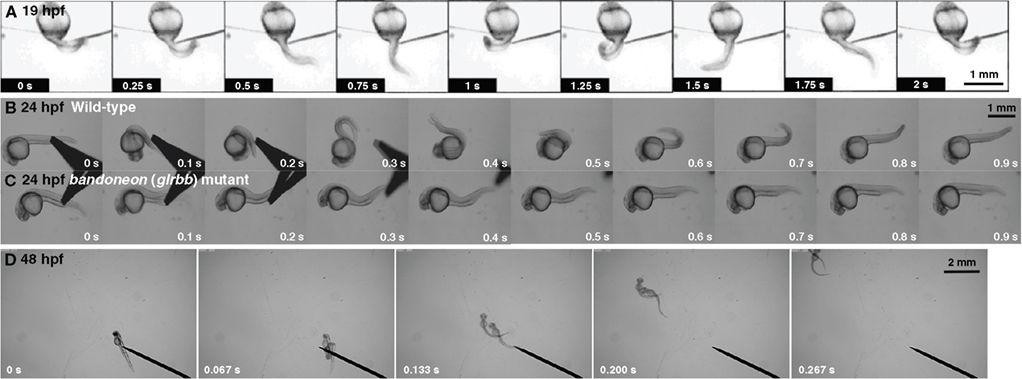
Figure 1. Zebrafish embryos display three early behaviors. (A) A 19 hpf wild-type embryo exhibits spontaneous coiling. (B) At 24 hpf, a wild-type embryo responds to mechanosensory stimulation with two fast, alternating contractions. (C) A bandoneon (beo) mutant embryo with a defect in glrbb responds to mechanosensory stimulation with bilateral axial muscle contractions that causes the trunk to shorten and bend dorsally. (D) At 48 hpf, wild-type embryos swim away following tactile stimulation.
After 21 hpf, zebrafish embryos respond to touch with escape contractions that typically consists of two to three rapid, alternating contractions of the tail, with muscles contralateral to the side of tactile stimulation contracting first to turn the embryo away from the stimulus (Figure 1
B; Saint-Amant and Drapeau, 1998
; Hirata et al., 2005
). Head and yolk stimulation activates trigeminal sensory neurons (Drapeau et al., 2002
), whereas trunk and tail stimulation activates Rohon-Beard neurons, which are primary sensory neurons located within the spinal cord and hindbrain of embryonic fish and amphibians. Thus, the neural circuitry responsible for locomotor responses to external stimuli is functional shortly after the appearance of spontaneous coiling. Applying tactile stimuli to spinalized embryos, which were transected rostral to somite 1, evokes the normal touch response (Pietri et al., 2009
). By contrast, transections at more caudal locations (somites 1–10) result in progressively weaker responses in progressively caudal locations (Downes and Granato, 2006
; Pietri et al., 2009
). Thus, the rostral hindbrain is necessary for the full touch-evoked escape response. It has also been reported that the touch response is dependent on AMPA-type glutamate receptor activation (Pietri et al., 2009
).
By 28 hpf, tactile stimulation initiates swimming (Figure 1
D; Saint-Amant and Drapeau, 1998
). The frequency of alternating contractions during swimming reaches 30 Hz at 36 hpf, which is comparable with the frequency in adult zebrafish (Buss and Drapeau, 2001
). Although spinalized embryos transected between somites 5–7 respond to touch with an initial tail flip, swimming does not follow the initial response in most cases (Downes and Granato, 2006
). Thus, it appears that the spinal cord can initiate a touch response, but that supraspinal input is necessary for swimming.
Since zebrafish embryos display organized behaviors within the first several days of development, behavioral mutagenesis screens are an efficient way to isolate mutants that have defects in the formation and function of neural circuits, including neuronal excitability and synaptic transmission. In the Tübingen screen, Granato and his colleagues reported 166 mutants that showed defective motility at 48–60 hpf (Granato et al., 1996
). Mutations that induced obvious developmental defects such as abnormal morphology and increased degeneration were eliminated, since they would also lead to defective responses. Some of the behavioral mutations were linked to muscle defects by simple visual inspection of muscle striation using polarized light. The actin-myosin structure of normal muscle fibers resulted in birefringence (double refraction) when viewed this way, while muscles with defective actin-myosin organization resulted in decreased birefringence (Felsenfeld et al., 1990
). Indeed, mutations in dystrophin, laminin, titin, Hsp90 and the cognate cochaperone Unc45b were identified in this manner (Bassett et al., 2003
; Etard et al., 2007
; Hall et al., 2007
; Steffen et al., 2007
; Hawkins et al., 2008
; Guyon et al., 2009
). Several other muscle mutations exhibited defects in excitation-contraction (EC) coupling or muscle structure that resembled human myopathies (Schredelseker et al., 2005
, 2009
; Zhou et al., 2006
; Hirata et al., 2007
; Dowling et al., 2009
). Since some of these zebrafish mutants exhibit muscle degeneration similar to human diseases, they could be useful for the biological and therapeutic analysis of these diseases (Kunkel et al., 2006
; Lieschke and Currie, 2007
).
Among the 166 motility mutations isolated from the Tübingen screen, 103 mutants displayed normal birefringence suggesting impairments in the nervous system, neuromuscular junction (NMJ) or functional components of muscle such as EC coupling. These mutants have been further classified into several classes by their responses to touch such as no response, normal but reduced response, vigorous but abnormal response, or simultaneous, bilateral contractions. The latter were named ‘accordion’ class mutants (Table 1
), because they respond to tactile stimulation with apparent simultaneous, bilateral contractions of axial muscles, resulting in shortening of the body rather than the normal alternating contractions. Molecular genetic studies have revealed that the ‘accordion’ phenotype can arise from numerous distinct mechanisms. For example, accordion (acc) mutants have slow muscle relaxation due to defective clearance of cytosolic Ca2+ caused by mutations in atp2a1, encoding the sarcoplasmic reticulum Ca2+-ATPase SERCA1, resulting in overlap of contractions by axial muscles on the two sides of mutant fish (Gleason et al., 2004
; Hirata et al., 2004
). Other ‘accordion’ class mutants show defects in cholinergic transmission, such as zeihharmonika (zim) which harbors either missense or nonsense mutations in ache, encoding acetylcholinesterase (Behra et al., 2002
; Downes and Granato, 2004
) and bajan (baj) which harbors a splice acceptor site mutation in chat, encoding choline acetyltransferase (Wang et al., 2008
). By contrast, diwanka (diw) mutants show defective primary motoneuron pathfinding as a result of nonsense mutations in plod3, encoding the procollagen lysine 2-oxoglutarate 5-dioygenase 3 (Zeller and Granato, 1999
; Zeller et al., 2002
; Schneider and Granato, 2006
).
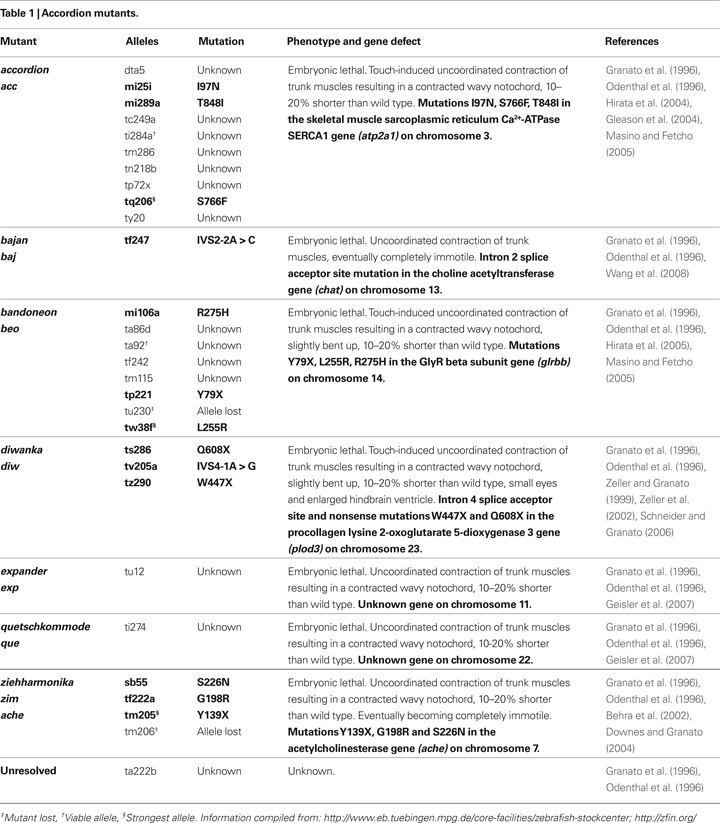
Simultaneous contraction of antagonistic muscles was also attributable to bilateral activation of motor neurons caused by impaired reciprocal inhibition in glrbb, encoding one of two zebrafish GlyR β subunits (Figure 1
C; Hirata et al., 2005
). Thus, defects in muscle Ca2+ storage, cholinergic transmission, motor projection and glycinergic transmission lead to very similar phenotypes in zebrafish embyros. Two additional ‘accordion’ class mutants, expander (exp) and quetschkommode (que) remain to be characterized (Table 1
; Granato et al., 1996
), and it will be intriguing to determine whether the underlying defects fit into one or other of the functional classes above. In order to distinguish between neuronal and muscle defects, one approach is to record the electrophysiological responses of muscles to sensory stimulation (Figure 2
A). The output of the nervous system and status of the NMJ can be assayed by membrane potential and voltage-clamp recordings following tactile stimulation of zebrafish embryos. For example, recordings from the muscles of accordion (atp2a1) mutants show normal rhythmic activity corresponding to fictive swimming, indicating that the nervous system is unaffected in accordion (Figures 2
B,C; Hirata et al., 2004
). However, highly aberrant, arrhythmic responses can be recorded from the muscles of bandoneon (glrbb) larvae, demonstrating that the nervous system output is defective in these mutants (Figure 2
D; Hirata et al., 2005
).
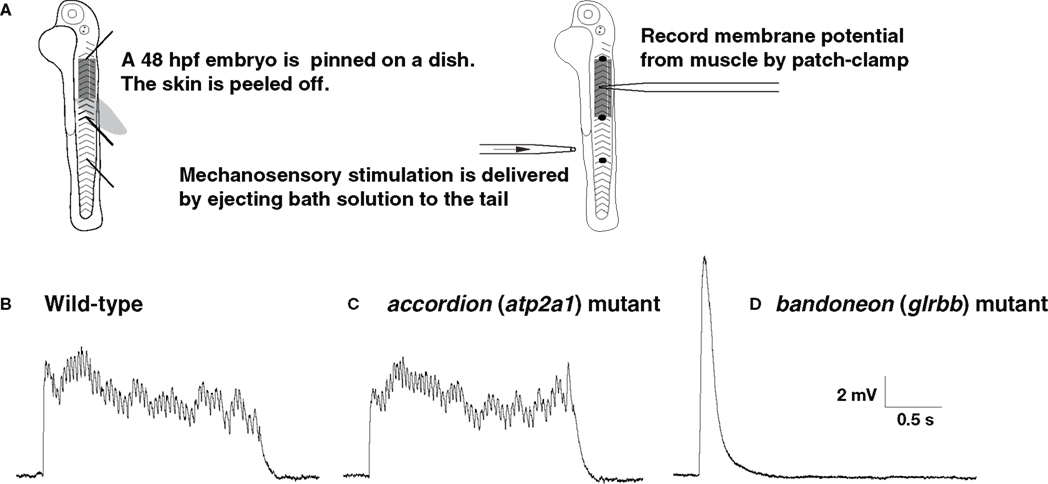
Figure 2. The output of the CNS is normal in accordion (atp2a1) mutants but abnormal in bandoneon (glrbb) mutants. (A) Schematic summary of the experimental procedure. (B) Wild-type muscle at 48 hpf responds to tactile stimulation with rhythmic depolarizations, representing cyclic muscle contractions during fictive swimming. (C) accordion (atp2a1) mutant muscle respond to tactile stimulation with a similar rhythmic pattern, indicating that the outputs from the CNS are normal. (D) bandoneon (glrbb) mutant muscle responds to tactile stimulation with a short and large voltage response devoid of rhythmicity, suggesting that the CNS outputs are aberrant.
Additional touch-insensitive mutants from the Tübingen screen, such as alligator (ali), macho (mao) and steifftier (ste), have defects in the excitability of sensory Rohon-Beard neurons (Granato et al., 1996
; Ribera and Nüsslein-Volhard, 1998
; Gnuegge et al., 2001
; Pineda et al., 2005
). Although the genes responsible for these mutations have not yet been identified, these mutants show reduced Na+ current amplitudes. By contrast, the zebrafish twitch twice (twt) mutant showing an aberrant unidirectional startle response was found to harbor nonsense mutations in robo3, encoding roundabout 3, a Slit ligand receptor essential for Mauthner cell axon guidance (Burgess et al., 2009
). Unsurprisingly, other mutations that affect axon outgrowth also exhibit abnormal behavior (Zeller and Granato, 1999
; Zhang and Granato, 2000
; Zhang et al., 2004
; Schneider and Granato, 2006
; Palaisa and Granato, 2007
; Tanaka et al., 2007
). For example, the mutant unplugged (unp) is defective in muscle-specific receptor tyrosine kinase (MuSK) and exhibits defective initial outgrowth of motor axons (Lefebvre et al., 2007
; Jing et al., 2009
). Other behavioral mutants exhibiting decreased synaptic transmission and clustering of nAChRs at the NMJ have also been identified including nicotinic receptor (nic) and sofa potato (sop) which harbor mutations in the nAChR α and δ subunit genes, respectively. By contrast, mutants unp and twitch-once (two) have mutations in the genes encoding MuSK and the AChR clustering factor rapsyn, respectively (Westerfield et al., 1990
; Sepich et al., 1998
; Ono et al., 2001
, 2002
, 2004
; Saint-Amant et al., 2008
). Curiously, another mutant in the ‘twitch-once’ class of motility mutants (Granato et al., 1996
), shocked (sho), was recently shown to result from mutations in slc6a9 encoding GlyT1 (Table 2
; Cui et al., 2005
; Mongeon et al., 2008
). Once again, this highlights that central nervous system and muscle defects can result in phenocopying in zebrafish.

Inhibitory GlyRs belong to a superfamily of ligand-gated ion channels that includes nAChRs, serotonin (5HT3) receptors and GABAA receptors (Lynch, 2004
). GlyRs are heteromultimers consisting of ligand-binding α and structural β subunits (Grenningloh et al., 1987
, 1990a
; Langosch et al., 1988
), the latter contain a binding site for gephyrin, a multifunctional cytoplasmic linker protein that clusters αβ GlyRs at synapses (Kirsch et al., 1993
; Meyer et al., 1995
; Feng et al., 1998
; Kim et al., 2006
). Four α subunit genes (GLRA1, GLRA2, GLRA3 and GLRA4) and a single β subunit gene (GLRB) have been identified in mammals (Grenningloh et al., 1987
, 1990a
,b
; Kuhse et al., 1990
, 1991
; Akagi et al., 1991
). In humans, however, GLRA4 is a pseudogene. Several variants are also created by alternative splicing and RNA editing (Meier et al., 2005
), which may modify functional properties such as agonist specificity, affinity and desensitization kinetics.
In zebrafish, four GlyR α subunits (αZ1, αZ2, αZ3 and αZ4) and two β subunits genes (βa (= βZ) and βb encoded by glrba and glrbb, respectively; Table 3
) were initially reported (David-Watine et al., 1999
; Imboden et al., 2001a
,b
,c
; Hirata et al., 2005
). Phylogenetic analysis suggested that αZ1, αZ3 and αZ4 showed high sequence similarity to the mammalian GlyR α1, α3 and α4 subunits, respectively (Imboden et al., 2001a
), and were referred to as zebrafish GlyR α1, GlyR α3, and GlyR α4 (Figures 3
A,C,D). A sequence reported as αZ2 was originally thought to encode a GlyR α2 subunit (Imboden et al., 2001b
), but Imboden and colleagues subsequently reclassified this protein as a second α4 subunit based on a more detailed phylogenetic analysis (Imboden et al., 2001a
). Thus, αZ2 was renamed α4a, and αZ4 was renamed α4b and the genes were renamed glra4a and glra4b, respectively. The existence of two distinct orthologs of a mammalian gene is not uncommon in the zebrafish genome, due to the suspected duplication of the whole genome during fish evolution (Amores et al., 1998
). Using the most recent zebrafish genome assembly Zv8
2
, we identified a novel zebrafish GlyR α subunit gene on chromosome 9 that is likely to encode α2 based on our own phylogenetic and sequence analysis (GenBank GQ406228; Figure 3
B). We also amplified new zebrafish cDNAs encoding the correct N-terminus of α4a, containing a cleavable signal peptide sequence (GenBank GQ406229), which may explain why the originally isolated α4a (αZ2) required the signal peptide of α1 in functional expression experiments (Imboden et al., 2001b
). Therefore, zebrafish have five α subunit (glra1, glra2, glra3, glra4a and glra4b) and two β subunit (glrba and glrbb) genes (Figures 3
A–E). As well as considering overall sequence identity and similarity, we considered other diagnostic criteria in our assignment of orthologs. For example, exon 3a and 3b are typically alternatively spliced in the mouse, rat and human GlyR α2 transcripts. But, RT-PCR and genome analysis suggests that zebrafish glra2, glra4a and glra4b do not show alternative splicing of exon 3. However, examining diagnostic residues in the M1-M3 domains proved more useful. The second residue in M2 (also called the 2’ residue) is typically glycine in GlyR α1/α4 and alanine in GlyR α2/α3 subunits. This residue has been suggested to influence the conductance of GlyR channels (Bormann et al., 1993
) and picrotoxin/picrotin blockade (Wang et al., 2007
; Yang et al., 2007
). From this point of view, our revised orthology appears to be more accurate, since the zebrafish GlyR α2 subunit has an alanine residue at the 2’ position, whereas both zebrafish GlyR α4 subunits harbor a glycine residue at this position.
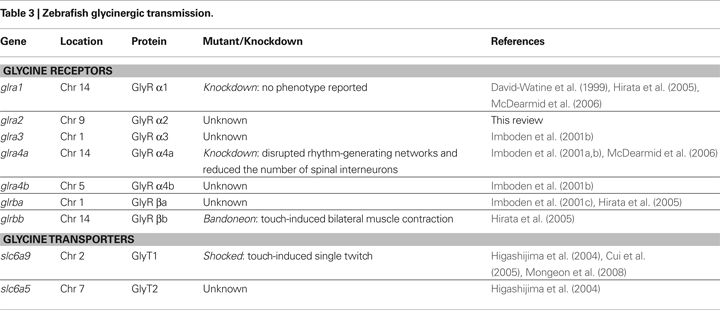

Figure 3. Sequence alignments of zebrafish GlyR subunits with avian and mammalian counterparts. (A) Sequence alignment of human (GenBank accession: NP_000162), rat (NP_037265), mouse (NP_065238) GlyR α1 with zebrafish (NP_571477) GlyR α1. The four membrane-spanning domains are represented as M1-M4. The 2’ residues in M2 are highlighted by grey box. Signal peptides are denoted by negative numbering. (B) Protein sequence alignment of human (NP_002054), rat (NP_036700), mouse (NP_906272), chick (XP_001234291) GlyR α2 with zebrafish (GQ406228) GlyR α2. (C) Protein sequence alignment of human (NP_006520), rat (NP_446176), mouse (NP_536686), chick (XP_420527) GlyR α3 with zebrafish (NP_694497) GlyR α3. (D) Protein sequence alignment of rat (XP_346351), mouse (NP_034427), chick (XP_001232995) with zebrafish GlyR GlyR α4a (GQ406229) and GlyR α4b (AAH85599). (E) Protein sequence alignment of human (NP_000815), rat (NP_445748), mouse (NP_034428) and chick (XP_420379) GlyR β with zebrafish GlyR βb (NP_001003587) and GlyR βa (XP_683646). Position of mutations identified in the three beo alleles are represented by arrowheads.
However, assuming orthology based on sequence identity alone is unwise, and it is also important to consider patterns of gene expression. Hindbrain neurons likely to be reticulospinal interneurons and spinal neurons express both glra1 and glra4a by 24 hpf (Imboden et al., 2001a
; Hirata et al., 2005
; McDearmid et al., 2006
) but glra4a was lost by 48 hpf. By contrast, Imboden and colleagues showed robust expression of glra4a at 52 hpf in the olfactory pits, mesencephalon, rhombencephalon and somites (Imboden et al., 2001a
). However, glra4b expression appears to be restricted to the retina in 52 hpf embryos (Imboden et al., 2001a
) and the expression patterns of glra2 and glra3 remain to be determined. Interestingly, glrbb is also expressed by reticulospinal and spinal neurons by 24 hpf, while glrba is not expressed in zebrafish until 72 hpf (Hirata et al., 2005
). Thus, whilst some data appear contradictory, it is likely that early embryonic GlyRs have the capacity to be heteromeric, with GlyR βa and GlyR βb forming different heteromeric GlyRs based on developmental expression profiles, and the apparent lack of compensation shown in bandoneon (beo).
In the Tübingen screen, seven alleles of the ‘accordion’ class beo mutation (tp221, tw38f, ta86d, ta92, tm115, tf242 and tu230) were isolated (Granato et al., 1996
; Table 1
) and named after the South American accordion-like instrument. From our behavioral screen, we generated another allele (mi106a) and showed that it results from missense and nonsense mutations in glrbb, encoding the GlyR βb subunit (Hirata et al., 2005
). As mentioned previously, at 24 hpf wild-type embryos responded to touch with multiple coils of the body, which was achieved by alternating trunk contractions (Figure 1
B). By contrast, beo mutants displayed simultaneous contraction of the bilateral axial muscles that resulted in a dorsal flexure and shortening of the body following a tactile stimulation (Figure 1
C). Later, when wild-type embryos swam in response to touch, beo mutants contracted the trunk muscle simultaneously on both sides and failed to swim in response to touch. Although beo mutants exhibited an abnormal tactile response, spontaneous coiling was not affected. In addition to behavioral perturbation, beo mutants exhibited secondary morphological defects in the notochord and axial muscles that were common to zebrafish behavioral mutants showing excessive contraction of the musculature (Hirata et al., 2004
; Lefebvre et al., 2004
). In fact, suppression of motor behavior either by a sodium channel inhibitor (tricaine) or a muscle myosin inhibitor (N-benzyl-p-toluene sulfonamide) prevented the morphological perturbations of mutant muscle. The beo larvae typically died at 7 days postfertilization (dpf), presumably due to their inability to swim and feed effectively, but cumulative notochord damage may have also contributed to lethality.
Alternation of muscle contractions on the left and the right side of animals requires reciprocal inhibition between left and right sides of the hindbrain and spinal cord (reviewed in Grillner, 2003
). Disruption of this inhibition can lead to simultaneous activation of bilateral motor neurons and thus simultaneous muscle contractions on both sides. Normally, tactile stimulation delivered to one side of the body leads to contraction of the contralateral side followed by contraction of the ipsilateral side. To see whether bilateral muscles are simultaneously activated in beo, we measured the latency of muscle depolarization following mechanosensory stimulation to the contralateral side and the ipsilateral side. We found that in wild-type muscles, contralateral tactile stimulation results in a muscle response 25 ms faster than ipsilateral stimulation, corresponding to initial touch-induced activation of contralateral muscles followed by ipsilateral muscles (Figure 4
A). By contrast, the latencies of response to ipsilateral and contralateral stimulation are comparable in beo, indicating that tactile stimulation activates both contralateral and ipsilateral muscles simultaneously. Thus, reciprocal inhibition appears to be deficient in beo.
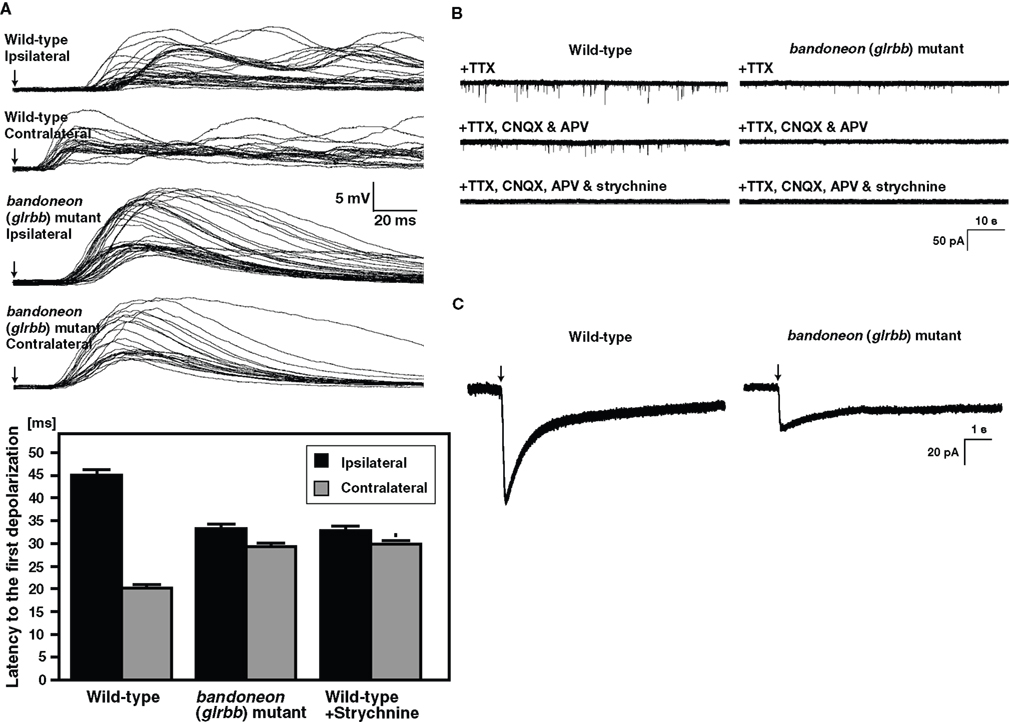
Figure 4. Simultaneously contraction of bilateral axial muscles in bandoneon (glrbb) mutants due to the loss of glycinergic synaptic transmission. (A) Superimposed voltage responses of muscles evoked by mechanosensory stimulation. Arrows indicate the time of stimulation. The latency of the muscle response to contralateral stimulation was shorter than that to ipsilateral stimulation in wild-type, whereas the latency to ipsilateral and contralateral stimulation was comparable in beo mutants. Histograms show that the latency to half-maximal amplitude of the first depolarization was shorter in contralateral stimulation compared to ipsilateral stimulation in wild-type. The latency of the response to tactile stimulation in strychnine-treated wild-type muscles was comparable to latency in beo mutants. (B) Spontaneous synaptic currents recorded from a wild-type motor neuron in the presence of TTX were decreased in frequency following block of NMDA and AMPA receptors by application of CNQX and APV, respectively. The non-glutamatergic currents in wild-type are eliminated by further application of strychnine, showing that they are glycinergic currents. In beo, non-glutamatergic currents in the presence of CNQX and APV are missing, indicating that glycinergic synaptic currents are absent. (C) A puff of exogenous glycine induced a current in a wile-type motor neuron and a smaller current in a beo mutant motor neuron.
Examination of glycinergic synaptic transmission in beo by patch-clamp recordings of motor neurons showed that glycinergic, but not glutamatergic synaptic transmission was absent in beo (Figure 4
B). Concordantly, immunolabeling with an anti-GlyR α antibody confirmed that GlyRs were not clustered in beo spinal cord as they were in wild-type zebrafish. Interestingly, application of exogenous glycine directly onto motor neurons elicited currents in beo motor neurons, suggesting that non-clustered extrasynaptic GlyRs, which may represent homomeric α subunit GlyRs, existed in beo mutants (Figure 4
C). In fact, fetal extrasynaptic GlyRs were thought to be homopentamers of GlyR α2 in rodents (Becker et al., 1988
; Malosio et al., 1991
; Watanabe and Akagi, 1995
). Taken together, these results demonstrated that the GlyR βb subunit was required for synaptic aggregation of GlyRs, corroborating previous findings showing that GlyR β interacted with gephyrin (Meyer et al., 1995
; Sola et al., 2004
; Kim et al., 2006
) a multifunctional cytoplasmic protein that is crucial for the synaptic localization of GlyRs (Kirsch et al., 1993
; Feng et al., 1998
). The synaptic GlyRs that were eliminated in beo could contain either GlyR α1 or α4a, since the corresponding genes appeared to be expressed by hindbrain and spinal neurons during early development. Antisense knockdown of GlyR α4a (but not GlyR α1) reduced glycinergic synaptic transmission and disrupted activity of circuits underlying swimming (McDearmid et al., 2006
), suggesting that the early synaptic GlyRs could be α4a/βb heteromers. A complication in this study was that GlyR α4a was referred to as α2 based upon the initial designation of this cDNA as αZ2 (Imboden et al., 2001a
,b
). The behavioral phenotype associated with α1 and α4a knockdown was not reported, and the translation blocking α4a morpholino used in these studies also caused a reduction in the number of spinal interneurones (McDearmid et al., 2006
), a phenotype not examined in beo (Hirata et al., 2005
). However, the sequences of multiple ESTs and our own αZ4a cDNA cloning suggested that the α4a morpholino used by McDearmid et al. (2006)
may have been directed against a mis-spliced intronic sequence upstream of glra4a exon 2. This does not preclude gene knockdown by interference with glra4a splicing, but further studies with other GlyR-directed morpholinos may be warranted to uncover the exact biological roles of the zebrafish GlyR genes.
In humans defects in glycinergic synaptic transmission lead to hyperekplexia. Hyperekplexia is a rare neurological syndrome that is characterized by an exaggerated startle response accompanied by transient muscle rigidity in response to unexpected acoustic or tactile stimuli (Gastaut, 1967
; Bakker et al., 2006
). More than 20 distinct missense mutations and several nonsense and frameshift mutations have been identified in the GlyR α1 subunit gene (GLRA1) to date (Shiang et al., 1993
; Bakker, 2006
; Harvey et al., 2008
). Most missense mutations in the second membrane-spanning domain and its neighboring loops are dominant mutations, whereas most point mutations in the N-terminal extracellular domain and M3-M4 intracellular loop are recessive. Missense and nonsense mutations in GLRA1 that lead to exaggerated startle reflexes are also found in spontaneous and induced mouse mutants and bovine congenital myoclonus (Gundlach et al., 1988
; Buckwalter et al., 1994
; Ryan et al., 1994
; Pierce et al., 2001
; Holland et al., 2006
; Traka et al., 2006
). To date, no mutations in the other functional GlyR α subunit genes (GLRA2 and GLRA3) have been reported, but in one family compound heterozygous mutations in GLRB were associated with hyperekplexia (Rees et al., 2002
). Similarly in mice, GlyR β hypomorphs (due to a LINE1 transposable element insertion causing mis-splicing of Glrb transcripts) also exhibit abnormal startle responses (Becker et al., 1992
; Kingsmore et al., 1994
; Mülhardt et al., 1994
; Hartenstein et al., 1996
).
The mutations underlying three alleles of beo have been identified to date (Hirata et al., 2005
; Table 1
). In the tp221 allele, a nonsense mutation (Y79X) is predicted to cause truncation of GlyR βb. By contrast, in tw38f and mi106a, missense mutations L255R and R275H, respectively, were found in the first membrane-spanning domain (M1) and in the intracellular loop between the first (M1) and second (M2) membrane-spanning domains. The R275H mutation in zebrafish GlyR βb affects a highly conserved arginine residue prior to M2 and in fact in human GlyR α1, the corresponding mutation R252H is known to accelerate degradation of GlyR α1 (Rea et al., 2002
), suggesting that mi106a mutation is a hypomorph of GlyR βb. Characterization of the remaining beo alleles is underway, and may reveal key residues of involved in the function of GlyR βb.
Glycine transporters are 12 membrane-spanning domain proteins that belong to Na+/Cl–-dependent transporter superfamily. In vertebrates, two glycine transporters, GlyT1 and GlyT2, mediate the uptake of glycine from the extracellular space to the cytosol driven by an electrogenic gradient (Eulenburg et al., 2005
). GlyT1 is enriched in astrocytes and some excitatory neurons, whereas GlyT2 is enriched in inhibitory glycinergic neurons (Adams et al., 1995
; Zafra et al., 1995
; Jursky and Nelson, 1996
; Cubelos et al., 2005
). At glycinergic synapses, astroglial GlyT1 is thought to clear glycine from the synaptic cleft, so terminating neurotransmission. Since glycine serves both as ligand for GlyR activation and as a coagonist and possibly primary ligand for NMDA receptors (Johnson and Ascher, 1987
; Kuryatov et al., 1994
; Chatterton et al., 2002
), GlyT1 regulates both inhibitory and excitatory synaptic transmission. By contrast, GlyT2 is localized to presynaptic terminals of glycinergic neurons and is essential for glycine reuptake, replenishing the pool of releasable transmitter (Zafra et al., 1995
; Gomeza et al., 2003b
; Mahendrasingam et al., 2003
).
Zebrafish sho mutants fail to initiate swimming following tactile stimulation at 3 dpf (Granato et al., 1996
; Cui et al., 2004
; Luna et al., 2004
). This phenotype is caused by missense mutations in slc6a9, which encodes GlyT1 (Cui et al., 2005
; Mongeon et al., 2008
). The ta229g allele, which displays the strongest phenotype, results from a G81D missense mutation in the second membrane-spanning domain. Expression of the recombinant zGlyT1 G81D mutant in Xenopus oocytes revealed that GlyT1 function is abolished by this mutation (Cui et al., 2005
). The milder te301 allele harbors a C305Y missense mutation that is located next to a deduced glycine-binding residue in the pore-forming sixth membrane-spanning domain (Yamashita et al., 2005
; Rees et al., 2006
; Harvey et al., 2008
; Mongeon et al., 2008
). Both sho mutant alleles display severely compromised tactile-induced locomotion and the frequency of spontaneous coiling is reduced. Tactile stimuli do not evoke escape contractions in sho mutants at 24 hpf, unlike wild-type siblings. At later stages, when tactile stimuli induce swimming in wild-type fish, sho mutants respond with a few uncoordinated trunk contractions (or not at all) and do not display typical swimming behavior. While shota229g mutants normally die within 2 weeks of development, importantly they can be maintained to adulthood by careful feeding. These adult shota229g homozygous fish are less active than wild-type zebrafish but fertile, indicating some degree of functional recovery. Interestingly, the weaker shote301 mutants recover by around 4–5 dpf (Mongeon et al., 2008
), suggesting that there is a degree of compensation for the loss of functional GlyT1 in zebrafish.
The effects of the loss of GlyT1 function on the nervous system and/or muscles were examined by electrophysiology. Voltage recordings from muscle of wild-type embryos show sustained episodes of rhythmic depolarizations corresponding to swimming following tactile stimulation, whereas sho muscle responds with one or two short, arrhythmic depolarizations corresponding to the uncoordinated ‘twitch-once’ muscle contractions exhibited by sho mutants (Figures 5
A,B; Cui et al., 2004
). Similarly, wild-type spinal motor neurons respond to touch with a long burst of action potentials, while sho motor neurons responded with only a short burst. Thus, the nervous system output of the CNS is aberrant in sho, signifying that the loss of GlyT1 disturbs the CNS function at glycinergic synapses (Cui et al., 2005
). Interestingly, the sho mutants also exhibit aberrant electrical coupling between axial muscle fibers at 3 dpf (Luna et al., 2004
), but this may be a secondary consequence of the CNS defects, since GlyT1 is not expressed in muscle. If the defect in CNS signaling in sho is attributable to increased extracellular glycine levels in the synaptic cleft due to the loss of GlyT1 function, one would expect normal responses to be restored in mutants when excess glycine is washed out. To examine this possibility, the hindbrain, which contains many of the neurons mediating responses to tactile stimulation, was exposed to various solutions by perfusion following removal of dorsal roof of the fourth ventricle and responses to tactile stimulation assayed by muscle recordings (Cui et al., 2005
). When mutants were perfused with glycine-free solution, they respond with normal swimming behavior (Figure 5
C) and this functional recovery is inhibited by addition of exogenous glycine (Figure 5
D). Furthermore, sho mutants are more sensitive to the deleterious effects of exogenous glycine on touch-induced swimming compared to wild-type fish (Mongeon et al., 2008
). Thus, the putative increase in extracellular glycine due to the loss of GlyT1 is likely to mediate aberrant signaling within the mutant CNS.
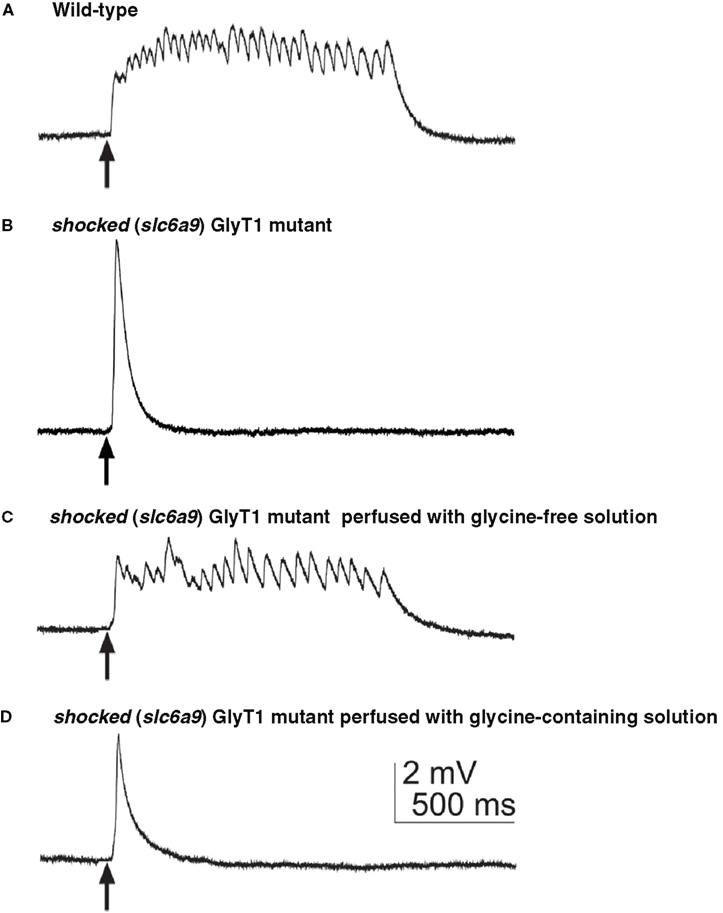
Figure 5. The aberrant motor response of shocked (slc6a5) mutants defective in GlyT1 is due to high external glycine. (A) Muscle voltage recording from a wild-type embryo showed normal fictive swimming in response to mechanosensory stimulation. Arrows indicate the time of stimulation. (B) Muscle recording from a sho mutant embryo displaying a large, nonrhythmic depolarization. (C) Muscle recording from a sho mutant after the hindbrain was exposed and perfused with glycine-free solution demonstrated rhythmic depolarizations similar to fictive swimming. (D) Muscle recording from the same sho embryo as in (C) after switching the perfusion from glycine-free solution to saline containing 0.2 mM glycine again exhibited the aberrant response characteristic of sho mutants.
One presumption of these perfusion experiments was that extracellular glycine levels were high in the synaptic cleft of sho mutants. This excess glycine is predicted to lead to increased inhibition of neurons receiving glycinergic inputs. If this presumption is correct, then blocking glycinergic inhibition should ameliorate the effects of high glycine in sho mutants. In fact, application of low concentrations of strychnine to sho mutants led to partial recovery of spontaneous coiling in 21 hpf embryos and normal swimming responses in older (40–46 hpf) embryos (Cui et al., 2005
). However, the partially recovered response might also be attributed to increased excitatory transmission via NMDA receptors in sho mutants. Taken together, it appears that the defective signaling in sho is consistent with abnormally high glycine in the synaptic cleft.
GlyT1 knockout mice showed many features that resembled those exhibited by zebrafish sho mutants, most notably motor deficits including those involving respiratory neural circuits (Gomeza et al., 2003a
; Tsai et al., 2004
). Recordings of spontaneous neuronal activity from hypoglossal motor neurons revealed that inspiratory cycling of the respiratory network of the brain stem was nearly eliminated in GlyT1 mutant mice. Much like the palliative effect of low concentration strychnine on the swimming circuit of zebrafish sho mutants, normal rhythmic activity was restored in hypoglossal motor neurons upon application of low strychnine to brainstem slices from GlyT1 knockout mice. Furthermore, voltage-clamp analyses of hypoglossal neurons were consistent with an increase in extracellular glycine. Thus, the neural defects seen in GlyT1-deficient mice are also likely to be due to increased levels of synaptic glycine, leading to suppression of neural networks. So far, no human GlyT1 defects have been associated with any disease. However, GlyT1 dysfunction has been suggested to play a possible role in glycine encephalopathy (Gomeza et al., 2003a
; Harvey et al., 2008
), as well as the psychiatric disorder schizophrenia (Freedman, 2003
; Tsai et al., 2004
), where NMDA receptor hypofunction is suspected. GlyT1 inhibitors elicit activation of NMDA receptors by increasing synaptic glycine levels, thus accelerating the co-agonist action of glycine may be useful pharmacological tools to mitigate some features of schizophrenia (Le Pen et al., 2003
). However, whether sequence variations in the GlyT1 gene (SLC6A9) are linked to glycine encephalopathy or schizophrenia and whether GlyT1 inhibitors are useful pharmacotherapies remains to be determined. Interestingly, SNPs in the human GlyT1 gene were recently proposed to be associated with methamphetamine-use disorder (Morita et al., 2008
; Bousman et al., 2009
). Animal models are clearly required for investigating the biological roles of GlyT1 and for the identification of therapeutic agents for treatment of human disorders related to GlyT1. In this respect, mouse GlyT1 knockouts may be less than ideal, since they die on the day of birth (Gomeza et al., 2003a
; Tsai et al., 2004
), whereas zebrafish sho mutants are accessible and viable.
Zebrafish bandoneon and shocked mutants are useful models for understanding of glycinergic synaptic transmission and for clarifying the biological consequences of gene disruption that impinge upon glycinergic signaling in vivo. Furthermore, future analysis of other zebrafish mutations may reveal new insights. For example, two ‘accordion’ class mutants (que and exp) remain to be analyzed in depth, as well as the crazy fish mutant techno trousers (tnt). Although these mutants have been suggested to harbor defects in glycinergic transmission because they exhibit exaggerated startle responses in response to touch (Granato et al., 1996
), history has taught us that there are several potential phenocopies of glycinergic defects. Equally, it is unclear why defects in glra1 (encoding GlyR α1) and slc6a5 (encoding GlyT2) were not uncovered in mutagenesis screens to date, since these are highly mutable genes in other species (Harvey et al., 2008
). Importantly, the genetic, developmental, and physiological accessibility of zebrafish make them useful animal models of human syndromes such as hyperekplexia. Small molecule screens using zebrafish mutants have successfully identified several drugs that ameliorate mutant phenotypes (Peterson et al., 2004
; Stern et al., 2005
). Such screens using zebrafish sho mutants have identified several compounds that mitigate the impairment of touch response (Hirata, unpublished). Future comparative and integrative studies using a variety of organisms including zebrafish with defects in glycinergic transmission are a promising strategy for a comprehensive understanding and development of pharmaceutical agents for human diseases defective in glycinergic synaptic transmission.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We apologize to investigators whose work could not be cited in this manuscript owing to space limitations. We thank Drs. Y. Oda and S. Takagi for insightful discussion and encouragement. Research has been supported by Grant-in-Aid for Young Scientists (A) and for Scientific Research on Priority Area “System Genomics” from the Ministry of Education, Culture, Sports, Science and Technology of Japan and the Nakajima Foundation and Career Development Award of the Human Frontier Science Program to Hiromi Hirata; National Science Foundation (0725976) and National Institutes of Neurological Diseases and Strokes (1RO1NS054731) to John Y. Kuwada; Medical Research Council (G0500833, G0601585) to Robert J. Harvey.
Charlier, C., Coppieters, W., Rollin, F., Desmecht, D., Agerholm, J. S., Cambisano, N., Carta, E., Dardano, S., Dive, M., Fasquelle, C., Frennet, J. C., Hanset, R., Hubin, X., Jorgensen, C., Karim, L., Kent, M., Harvey, K., Pearce, B. R., Simon, P., Tama, N., Nie, H., Vandeputte, S., Lien, S., Longeri, M., Fredholm, M., Harvey, R. J., and Georges, M. (2008). Highly effective SNP-based association mapping and management of recessive defects in livestock. Nat. Genet. 40, 449–454.
Doyon, Y., McCammon, J. M., Miller, J. C., Faraji, F., Ngo, C., Katibah, G. E., Amora, R., Hocking, T. D., Zhang, L., Rebar, E. J., Gregory, P. D., Urnov, F. D., and Amacher, S. L. (2008). Heritable targeted gene disruption in zebrafish using designed zinc-finger nucleases. Nat. Biotechnol. 26, 702–708.
Geisler, R., Rauch, G. J., Geiger-Rudolph, S., Albrecht, A., van Bebber, F., Berger, A., Busch-Nentwich, E., Dahm, R., Dekens, M. P., Dooley, C., Elli, A. F., Gehring, I., Geiger, H., Geisler, M., Glaser, S., Holley, S., Huber, M., Kerr, A., Kirn, A., Knirsch, M., Konantz, M., Küchler, A. M., Maderspacher, F., Neuhauss, S. C., Nicolson, T., Ober, E. A., Praeg, E., Ray, R., Rentzsch, B., Rick, J. M., Rief, E., Schauerte, H. E., Schepp, C. P., Schönberger, U., Schonthaler, H. B., Seiler, C., Sidi, S., Söllner, C., Wehner, A., Weiler, C., and Nüsslein-Volhard, C. (2007). Large-scale mapping of mutations affecting zebrafish development. BMC Genomics 8, 11.
Golling, G., Amsterdam, A., Sun, Z., Antonelli, M., Maldonado, E., Chen, W., Burgess, S., Haldi, M., Artzt, K., Farrington, S., Lin, S.-Y., Nissen, R. M., and Hopkins, N. (2002). Insertional mutagenesis in zebrafish rapidly identifies genes essential for early vertebrate development. Nat. Genet. 31, 135–140.
Granato, M., van Eeden, F. J. M., Schach, U., Trowe, T., Brand, M., Furutani-Seiki, M., Haffter, P., Hammerschmidt, M., Heisenberg, C.-P., Jiang, Y.-J., Kane, D. A., Kelsh, R. N., Mullins, M. C., Odenthal, J., and Nüsslein-Volhard, C. (1996). Genes controlling and mediating locomotion behavior of the zebrafish embryo and larva. Development 123, 399–413.
Grenningloh, G., Schmieden, V., Schofield, P. R., Seeburg, P. H., Siddique, T., Mohandas, T. K., Becker, C.-M., and Betz, H. (1990b). Alpha subunit variants of the human glycine receptor: primary structures, functional expression and chromosomal localization of the corresponding genes. EMBO J. 9, 771–776.
Haffter, P., Granato, M., Brand, M., Mullins, M. C., Hammerschmidt, M., Kane, D. A., Odenthal, J., van Eeden, F. J., Jiang, Y.-J., Heisenberg, C. P., Kelsh, R. N., and Furutani-Seiki, M., Vogelsang, E., Beuchle, D., Schach, U., Fabian, C., and Nüsslein-Volhard, C. (1996). The identification of genes with unique and essential functions in the development of the zebrafish, Danio rerio. Development 123, 1–36.
Hall, T. E., Bryson-Richardson, R. J., Berger, S., Jacoby, A. S., Cole, N. J., Hollway, G. E., Berger, J., and Currie, P. D. (2007). The zebrafish candyfloss mutant implicates extracellular matrix adhesion failure in laminin α2-deficient congenital muscular dystrophy. Proc. Natl. Acad. Sci. U.S.A. 104, 7092–7097.
Harvey, R. J., Depner, U. B., Wässle, H., Ahmadi, S., Heindl, C., Reinold, H., Smart, T. G., Harvey, K., Schütz, B., Abo-Salem, O. M., Zimmer, A., Poisbeau, P., Welzl, H., Wolfer, D. P., Betz, H., Zeilhofer, H. U., and Müller, U. (2004). GlyR α3: an essential target for spinal PGE2-mediated inflammatory pain sensitization. Science 304, 884–887.
Hawkins, T. A., Haramis, A.-P., Etard, C., Prodromou, C., Vaughan, C. K., Ashworth, R., Ray, S., Behra, M., Holder, N., Talbot, W. S., Pearl, L. H., Strähle, U., and Wilson. S. W. (2008). The ATPase-dependent chaperoning activity of Hsp90a regulates thick filament formation and integration during skeletal muscle myofibrillogenesis. Development 135, 1147–1156.
Hirata, H., Watanabe, T., Hatakeyama, J., Sprague, S. M., Saint-Amant, L., Nagashima, A., Cui, W. W., Zhou, W., and Kuwada, J. Y. (2007). Zebrafish relatively relaxed mutants have a ryanodine receptor defect, show slow swimming and provide a model of multi-minicore disease. Development 134, 2771–2781.
Morita, Y., Ujike, H., Tanaka, Y., Kishimoto, M., Okahisa, Y., Kotaka, T., Harano, M., Inada, T., Komiyama, T., Hori, T., Yamada, M., Sekine, Y., Iwata, N., Iyo, M., Sora, I., Ozaki, N., and Kuroda, S. (2008). The glycine transporter 1 gene (GLYT1) is associated with methamphetamine-use disorder. Am. J. Med. Genet. B 147B, 54–58.
Odenthal, J., Haffter, P., Vogelsang, E., Brand, M., van Eeden, F. J., Furutani-Seiki, M., Granato, M., Hammerschmidt, M., Heisenberg, C. P., Jiang, Y. J., Kane, D. A., Kelsh, R. N., Mullins, M. C., Warga, R. M., Allende, M. L., Weinberg, E. S., and Nüsslein-Volhard, C. (1996). Mutations affecting the formation of the notochord in the zebrafish, Danio rerio. Development 123, 103–115.
Rees, M. I., Harvey, K., Pearce, B. R., Chung, S. K., Duguid, I. C., Thomas, P., Beatty, S., Graham, G. E., Armstrong, L., Shiang, R., Abbott, K. J., Zuberi, S. M., Stephenson, J. B., Owen, M. J., Tijssen, M. A., van den Maagdenberg, A. M., Smart, T. G., Supplisson, S., and Harvey, R. J. (2006). Mutations in the gene encoding GlyT2 (SLC6A5) define a presynaptic component of human startle disease. Nat. Genet. 38, 801–806.