 Alejandro Reinares-Sebastián1,2,3
Alejandro Reinares-Sebastián1,2,3 Noelia Esteban-García1,2,4
Noelia Esteban-García1,2,4 Masahiko Takada5,6
Masahiko Takada5,6 Inés Trigo-Damas1,2,3,7*
Inés Trigo-Damas1,2,3,7*- 1HM Centro Integral de Neurociencias Abarca Campal (CINAC), Hospital Universitario HM Puerta del Sur, HM Hospitales, Madrid, Spain
- 2Instituto de Investigación Sanitaria HM Hospitales, Madrid, Spain
- 3CIBERNED (Center for Networked Biomedical Research on Neurodegenerative Diseases), Instituto Carlos III, Madrid, Spain
- 4PhD Program in Neuroscience Autónoma de Madrid University–Cajal Institute, Madrid, Spain
- 5Center for the Evolutionary Origins of Human Behavior, Kyoto University, Inuyama, Aichi, Japan
- 6Department of Neurology, Graduate School of Medicine, Osaka University, Osaka, Japan
- 7Centro Universitario HM Hospitales de Ciencias de la Salud (CUHMED), Universidad Camilo José Cela, Madrid, Spain
Parkinson’s disease (PD) remains a major challenge for translational neuroscience, with an increasing global prevalence and persistent unmet therapeutic needs. While its classical motor symptoms, such as bradykinesia, rigidity, and tremor, are well characterized, the clinical spectrum extends to diverse and often disabling non-motor manifestations, including hyposmia, constipation, and sleep disturbances. These features typically precede motor deficits and may dominate the late stages of disease. Despite decades of research, existing treatments remain primarily symptomatic and fail to halt disease progression. This situation has driven the development of a broad repertoire of preclinical models—ranging from in vitro cellular systems to complex animal models—to better understand pathogenesis and identify disease-modifying strategies. However, significant translational gaps persist, partly due to limitations in how well these models recapitulate the heterogeneity and complexity of human PD. In this review, we critically examine the main preclinical models available for PD, assessing their strengths and weaknesses for modeling both motor and non-motor features. We discuss recent advances, persistent challenges, and highlight key considerations for improving the predictive value of experimental models in drug discovery for Parkinson’s disease.
Introduction
Parkinson’s disease (PD) is the second most common neurodegenerative disease after Alzheimer’s disease (Reichmann et al., 2009). The number of people affected by PD has been estimated at 70 million and with the progressive aging of the population over the next 10 years the number could double. In Europe alone, it represents 14 billion euro in direct and indirect costs and this amount is only expected to gradually increase (Bach et al., 2011; Chaudhuri et al., 2024). In most cases, PD is a sporadic idiopathic disease that could be related to environmental factors, and only 10–15% of cases could be related to familiar PD such as SNCA, LRRK2, VPS35, PRKN, PINK1, GBA, and DJ-1 mutations (Blesa et al., 2016a,b; Shadrina and Slominsky, 2021).
The cardinal motor symptoms of PD are mainly associated with the degeneration of dopaminergic neurons, starting in the ventrolateral region of the substantia nigra pars compacta (SNc) (Blesa et al., 2022). When the loss of nigrostriatal dopaminergic neurons reaches 50% and the striatal dopamine (DA) deficit increased up to 80%, cardinal motor symptoms (akinesia, rigidity, and tremor) appear (Marsden, 1990; Fearnley and Lees, 1991; Armstrong and Okun, 2020). The development of the first motor symptoms is preceded by a series of non-motor symptoms (hyposmia, alterations of the rapid eye movement (REM) phase, sleep behavior disorders (RBD) and constipation) (Postuma and Berg, 2016; Schapira et al., 2017; Berg et al., 2018; Bloem et al., 2021). Interestingly, these symptoms may end up dominating the clinical profile in the last years of the disease (Blesa et al., 2022). The duration of the preclinical phase is unknown; however, it could be extended from 10 to 15 years before diagnosis (Blesa et al., 2017; Tolosa et al., 2021). Another neuropathological hallmark of PD is the presence of Lewy bodies (LB) in the dopaminergic neurons, whose main component is the α-synuclein (α-syn) protein. The accumulation of α-syn becomes widespread in the brain during the progression of PD (Braak et al., 2003). Although it is considered a hallmark, the presence of α-syn protein may also occur in the natural aging process of the brain and in other synucleinopathies (Greffard et al., 2010; Shahmoradian et al., 2019).
Traditionally, pharmacological treatment of PD is symptomatic and based on L-DOPA because DA or other exogenous catecholamines cannot cross the blood–brain barrier (BBB) (Rautio et al., 2008; Lafuente et al., 2019). However, chronic treatment with L-DOPA causes motor complications (on–off phenomenon) in most patients, associated with abnormal involuntary movements (dyskinesia) (Ahlskog and Muenter, 2001; Porras et al., 2015). Despite the difficulties in PD treatment, there are currently 136 agents in different trial phases and therapeutic categories according to the study by McFarthing et al. (2024), and this number has not greatly varied throughout recent years. Although only a few drugs have reached clinical trials for proving their effectiveness, none of them have been shown to fundamentally slow the pathology (McFarthing et al., 2022, 2024).
The clinical need for new PD-modifying strategies has led to the development of a wide variety of experimental models, from in vitro to animal models, and focused on improving pathogenic understanding of PD and validating potential therapies (Blesa et al., 2016a,b; Chia et al., 2020; Balzano et al., 2023). The translatability of experiments is based on the correct choice of experimental models that can simulate human pathology. This means that they must have features of human PD, theoretical rationales, and responses to treatments comparable to the PD patients (Blesa and Przedborski, 2014; Kin et al., 2019). However, most drug candidates that seem promising in the preclinical phases fail in clinical trials, making clear the gap in the transformation of preclinical to clinical outcomes (Del Rey N. L.-G. et al., 2018; Del Rey N. L. et al., 2018; Trigo-Damas et al., 2018). This manuscript aims to review the strengths and limitations of different experimental models available for PD and discuss their usefulness in drug discovery and development.
Preclinical platforms: from yeast, cells, and organoids to animals
In the last 20 years, yeast, mainly Saccharomyces cerevisiae, have been extensively used for the development of experimental models for neurodegenerative diseases. In essence, this type of model is based on two strategies: (1) study of heterologous expression of genes of monogenic diseases; (2) analysis of the functional activity of pathologically significant human orthologous genes (Dunham and Fowler, 2013). In the case of PD, both strategies have been used indistinctly, however, the first strategy is worth highlighting since it has been used for the analysis of the genes that encode the proteins parkin, PINK1, LRRK2, and α-syn (this last protein being the most studied in this models) (Dunham and Fowler, 2013; Shadrina and Slominsky, 2021).
Definitely, yeast models have allowed to expand our knowledge on the toxicity of α-syn inclusions (Auluck et al., 2010; Büttner et al., 2013). One of the most remarkable result obtained in this models have been the use of expression of genes encoding heat shock proteins to reduce α-syn toxicity; while deletion of genes encoding chaperones increased its toxicity (Chen et al., 2005; Bandres-Ciga et al., 2019; Billingsley et al., 2019). Thus, these types of models have opened the door to their use as a powerful screening tool to identify and optimize new compounds or drugs that reduce α-syn aggregation, such us flavonoids (Griffioen et al., 2006; Kritzer et al., 2009), which have been tested in rats and mice with promising results, reducing muscle tremor, reversed behavior deficits, striatal dopamine depletion, and neuronal cell loss (Figure 1; Mu et al., 2016; Ay et al., 2017; de Andrade Teles et al., 2018).
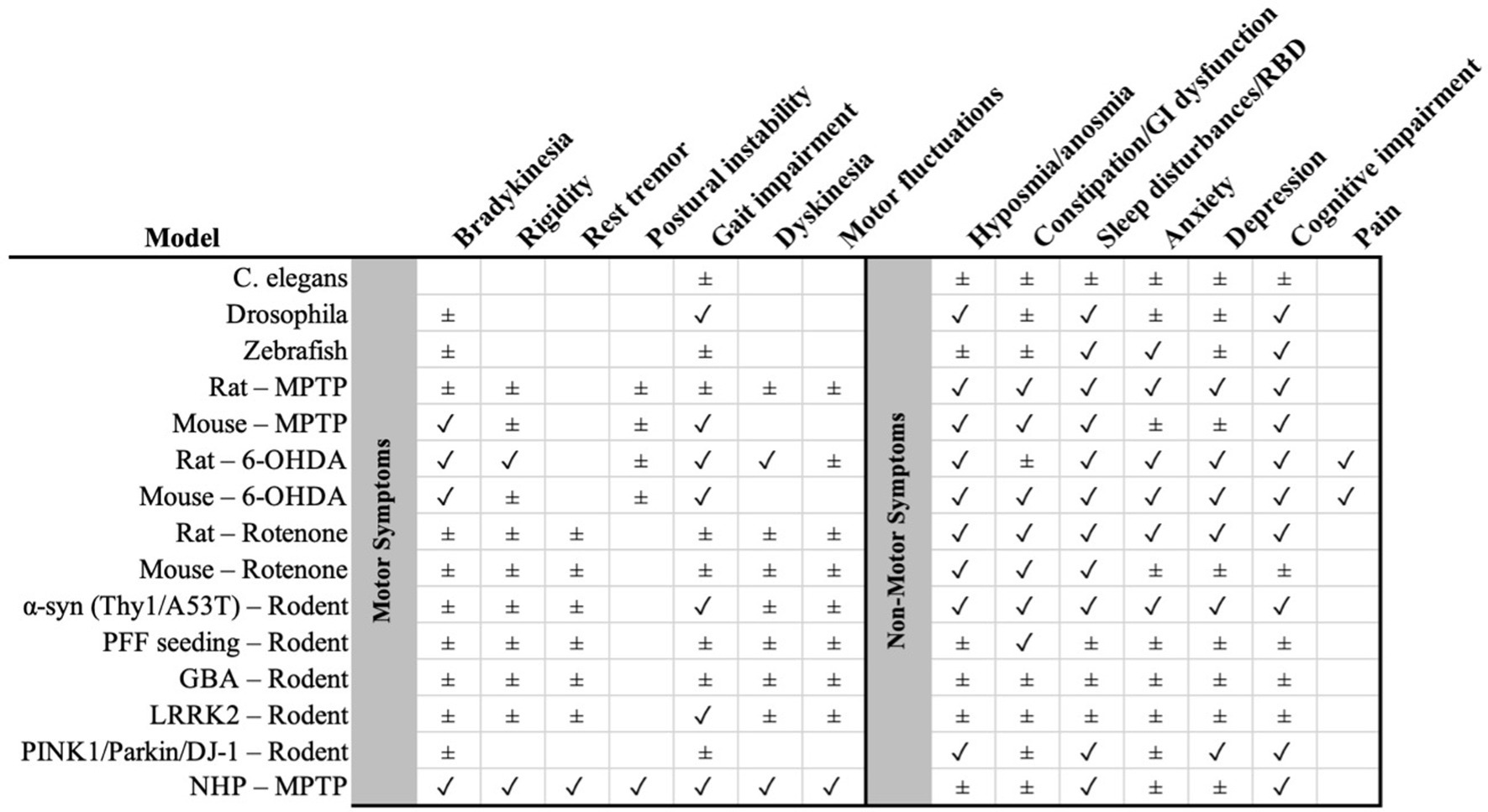
Figure 1. Summary matrix of preclinical Parkinson’s disease models (rows) versus clinical features (columns), with motor symptoms on the left and non-motor symptoms on the right (separated by the gray divider). Symbols indicate the level of support: ✓ consistent evidence that the feature is reproduced by the model; ± partial/heterogeneous or indirect evidence. NHP, non-human primate; α-syn, alpha-synuclein; PFF, pre-formed fibrils; 6-OHDA, 6-hydroxydopamine; MPTP, 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine; GBA, glucocerebrosidase; LRRK2, leucine-rich repeat kinase 2; PINK1/Parkin/DJ-1 as indicated. GI, gastrointestinal; RBD, REM sleep behavior disorder.
Despite yeast models having been the first approximation to shedding a light on metabolic pathways and /or proteins, these models should not be considered as ideal platforms for clinical trials due to their natural limitations, as a simplified model system and, therefore, need deeper confirmation in more complex models (Surguchov, 2021).
Cell models, due to their easy handling, rapid growth and simplicity usually become the first approach for modeling neurodegenerative diseases such as PD. Some examples are still being used in laboratories as the neuroblastoma cell line (SH-SY5Y), the pheochromocytoma cell line (PC12), the Neuro-2a cell line or the MN9D cell line (Falkenburger et al., 2016; Lázaro et al., 2017). At the same time, it is worth mentioning the human mesencephalic cells (LUHMES), that have a stable dopaminergic phenotype even with similar electrical properties (Falkenburger et al., 2016; Lázaro et al., 2017).
Cell models provide opportunities to develop pharmacologic or genetic studies in a relatively simple way, offering strategies to modulate the disease pathways. Noteworthy, one relevant example of the potential of these models is exenatide, a glucagon-like peptide-1 (GLP-1) receptor agonist resistant to enzymatic degradation, which demonstrated neuroprotective effects in vitro. It was tested in rat PC12 cells, which possess synthesis, metabolism and transporter systems of dopamine, and it was capable of inducing neurite growth and promoting neuronal differentiation. Following its promising results in preclinical PD models, the drug has advanced to phase III clinical trial (NCT04232969) (see Supplementary Table 1; Foltynie and Aviles-Olmos, 2014; Athauda et al., 2017; Charvin et al., 2018).
Interestingly, the search for personalized treatments has made the use of other cell culture approaches of growing relevance. An example of this is induced pluripotent stem cells (iPSCs), which can be reprogrammed from patient-derived somatic cells and differentiated into dopaminergic neurons (Torrent et al., 2015). A pioneering case in the USA demonstrated that the autologous transplantation of iPSC-derived dopaminergic progenitors, resulted in graft survival, functional evidence of dopamine production, and clinical improvement during follow-up (Figure 1; Schweitzer et al., 2020). In parallel, the phase 1/2 Kyoto trial (jRCT2090220384) (see Supplementary Table 1), initiated in 2018, employed cryopreserved allogeneic iPSC-derived dopaminergic progenitors with varying degrees of HLA compatibility to evaluate the safety and efficacy of this approach in PD patients (Takahashi, 2020). Although these strategies confirm the feasibility of iPSC-bases therapies, challenges remain regarding their high cost, complex production processes and, the need for immunosuppression. As an alternative, dopaminergic progenitors derived from human embryonic stem cells (hESCs) have been explored as an allogenic source. The first clinical trial with hESCs, reported by Wang et al., 2017 (NCT083119636) (see Supplementary Table 1), demonstrated the feasibility of such transplants in PD patients (Wang et al., 2018). Subsequently, two additional phase 1 clinical trials (NCT04802733 and STEM-PD) (see Supplementary Table 1) were initiated, confirming the safety and functional integration of the grafts in patients with PD (Kirkeby et al., 2017; Tabar et al., 2025).
In addition, CRISPR/cas9 has emerged as a novel technique in cell cultures and it has expanded the possibility of genome editing (Soldner et al., 2016). In summary, these two approaches provide closer alignment with the characteristics of PD patients, thereby facilitating studies designed to evaluate personalized treatments or compounds. However, they have not been able to establish pathological phenotypes of PD, since these are based on both mutations and environmental factors. One solution to this is the use of organoids 3D, allowing to investigate the genotype–phenotype relationships in PD, maintaining translational aspects of the disease (Jo et al., 2021). Organoid methodology allows the development of more accurate human cellular models for the investigation of neurodegenerative disorders, where biological processes- such as cell–cell interactions, pathological cascades, or spatial organization- that are not adequately reproduced in two-dimensional cultures (Smits et al., 2019). Furthermore, by imitating physiological conditions it allows testing new drugs or evaluating existing ones (Zhang et al., 2024; Frattini et al., 2025) Additionally, it provides a realistic reproduction of the blood brain barrier (BBB), which is crucial for advancing treatment strategies for PD (Cui et al., 2024; Dao et al., 2024). In conclusion, 3D cell culture systems serve as a bridge between traditional in vitro experiments and in vivo animal models (Smits and Schwamborn, 2020).
Nevertheless, this model has inherent limitations, and data derived from cell lines provide an important contribution to research by validating and refining the interpretation of results when applied alongside more advanced animal models such as the Drosophila, murine or other mammal model.
Modeling motor symptoms of Parkinson’s disease
Experimental models for PD have been pivotal in mimicking the hallmark motor symptoms—bradykinesia, rigidity, and tremor—providing platforms for understanding disease mechanisms and testing therapeutic approaches (He et al., 2024). These models include invertebrate organisms, and vertebrate animals, each contributing uniquely to the translational pipeline. These models—ranging from neurotoxin-induced lesions in rodents and primates to genetically engineered animals—aim to replicate key aspects of the disease, particularly its motor manifestations (Dovonou et al., 2023). While no single model fully recapitulates all facets of PD, each provides valuable insights into specific disease mechanisms and treatment responses (Barker and Björklund, 2020). The use of such models has been pivotal in elucidating dopaminergic pathways, evaluating pharmacological interventions like levodopa and dopamine agonists, and exploring novel approaches including gene and cell therapies (Chen et al., 2023).
Invertebrate models
These organisms share many conserved molecular pathways and cellular processes with humans and have been successfully used to study environmental and genetic factors (Fatima et al., 2018). Their rapid growth, low-cost maintenance, easy manipulation and well-known genome have made them a useful and ethically less problematic pharmacological screening tool compared to other animal models (Surguchov, 2021). Unfortunately, the absence of a complex physiological network, and the lack of the orthologous α-syn gene in most invertebrate models, have shown differences in both the pathogenic pathway and the effects on it. These disparities among models may arise from inherent variations between species or differences in the methodologies employed (Shadrina and Slominsky, 2021; Surguchov, 2021).
Caenorhabditis elegans
C. elegans is a microscopic nematode characterized by a short shelf life, many progeny and easy cultivation. This nematode has a small and well-studied genome with many orthologous PD-related genes, however it lacks the genes that encode for α-syn protein in humans (Shaye and Greenwald, 2011; Cooper and Van Raamsdonk, 2018).
Caenorhabditis elegans—toxic-based
This model allows the use of different neurotoxins including 6-hydroxydopamine (6-OHDA), paraquat, rotenone, and 1-methyl-4-phenylpyridinium (MPP+), resulting in selective neurodegeneration of dopaminergic neurons (around 30%). Similarly, the combination of genetic models with environmental neurotoxicity exposure led to an 85% of death (Zeng et al., 2018). Accordingly, this model has been used to test novel factors with the potential of reducing the neuronal toxicity of α-syn in dopaminergic neurons and epigenetic factors (Gaeta et al., 2019; Goya et al., 2020) and to evaluate the “prion-like” α-syn hypothesis owing to its transparency which allows the visualization of neuron-to-neuron transfer of α-syn (Sandhof et al., 2020).
Caenorhabditis elegan—genetic
Its transparency and easy of genetic manipulation have allowed genes to be silenced in specific neuronal subtypes- such as GABAergic, serotonergic, dopaminergic, cholinergic or glutamatergic neurons- using interfering RNA (RNAi), a method that is stable during at least 3 generations (Cooper and Van Raamsdonk, 2018). One example of this, is the work by Sohrabi et al., where they screened 50 drugs approved by the Food and Drugs Administration (FDA) in the C. elegans neuronal bcat-1 knockdown, which causes abnormal behavior with age (Figure 1; Sohrabi et al., 2021).
Lymnaea stagnalis
Moreover, L. stagnalis, offers a relatively long lifespan and high conservation of aging-related molecules (Fodor et al., 2020). Classic studies have shown that intermittent exposure of this snail to rotenone induced significant reduction of locomotion, feeding and tyrosine hydroxylase (TH) expression in the central nervous system, suggesting that this model could be suitable for PD research (Vehovszky et al., 2007; Maasz et al., 2017; Zeng et al., 2018). The potential benefit of this model in testing new drugs and compounds becomes evident with the compound Carnosine, a natural endogenous dipeptide that has shown benefits in clinical trials (Rivi et al., 2024; O’Toole et al., 2025).
Drosophila melanogaster
D. melanogaster or fruit fly is another powerful invertebrate model that shares approximately 75% of human diseases-related genes (Marsh and Thompson, 2006). Just like the rest of the invertebrate models, the fruit fly has numerous advantages related to handling: short life cycle, large progeny, a well-known small genome that allows edition and easy manipulation, as well as the conservation of brain structures and metabolic pathways (Aryal and Lee, 2019).
Drosophila melanogaster—toxic-based
Toxins can be used for modeling in Drosophila; however, these toxic models have not been widely adopted due to difficulties in estimation of the optimal dose to obtain comparable models. Nevertheless, some studies have been successful in reproducing the main characteristics of PD in toxic models (Figure 1; Coulom and Birman, 2004; Abolaji et al., 2018; Farombi et al., 2018).
Drosophila melanogaster—genetic
Drosophila’s nervous system is not completely homologous to the human, and one example is the case of the gene encoding α-syn in humans which has not been found in the Drosophila. Thus, the characteristics associated with α-syn are usually promoted by the inclusion of SNCA human gene in transgenic (TG) flies (Shadrina and Slominsky, 2021). TG-flies overexpressing α-syn display a progressive loss of dopaminergic neurons, α-syn inclusion formation, and loss of climbing ability. This has allowed to differentiate the presymptomatic and symptomatic phases characteristic of PD (Xiong and Yu, 2018). In addition, there are some drugs tested in the genetic Drosophila model that have managed to reach clinical trials, this is the case of Resveratrol (Adedara et al., 2022), Curcumin (Siddique et al., 2014) or Pomalidomide (Palmas et al., 2022). In the preclinical study of the Pomalidomide, using the LRRK2 genetic model, treated fruit flies managed to show improved climbing activity (Figure 1; Casu et al., 2020).
Although invertebrate models do not recapitulate the full anatomical and physiological complexity of the human nervous system, they are a powerful strategy for studying fundamental disease mechanisms, testing gene–environment interactions, and conducting large-scale drug screening. Their value resides in their capacity to yield robust and reproducible results during early-stage research, providing a foundation for validation in more complex models.
Vertebrate models
Vertebrate models are crucial in PD research, as they closely replicate key anatomical and physiological features of the human brain. This similarity allows us to study drug efficacy and delivery under conditions that more accurately reflect the human disease. However, this type of organisms is generally less recommended for drugs and genes screenings due to their long-life span and their complex genome, compared to the invertebrate models. Furthermore, these models can be expensive to maintain, and there are higher ethical restrictions, which limits the number and severity of the experimental procedures. These models encompass a wide phylogenetic spectrum, ranging from fish to primates, each with distinct advantages and limitations that shape their translational relevance (Blesa et al., 2018).
Zebrafish
The Zebrafish is distinguished by its small size, a rapid breed (3 months per generation) and a relatively simple maintenance. This model possesses a dopaminergic system located in the posterior tuberculum, functionally analogous to the human dopaminergic system (Shadrina and Slominsky, 2021). Their genomic and physiological organization makes them closer to the humans’ compared with Drosophila, C. elegans or L. stagnalis.
Zebrafish—toxic-based
Zebrafish exhibit susceptibility to various specific dopamine neurotoxins that selectively target dopaminergic neurons, including 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP), 6-OHDA, paraquat, and rotenone (Feng et al., 2014; Sarath Babu et al., 2016; Zeng et al., 2018; Zhang et al., 2017). For example, the intraperitoneal administration of MPTP in adult zebrafish resulted in reduced locomotor activity, decreased crossing frequency and swimming distance, increased freeze bouts, and extended freeze duration (Figure 1). Additionally, this treatment led to an upregulation of γ1- and γ2-synuclein expression (Sarath Babu et al., 2016; Zeng et al., 2018). Thus, this model has been used to assess the potential effect of Rasagiline (Cronin and Grealy, 2017), a selective inhibitor of the monoamine oxidase B (MAO-B), which clinical studies have demonstrated its potential neuroprotective effect in humans (NCT02789020) (see Supplementary Table 1).
Zebrafish—genetic
Moreover, 70% of human genes have orthologs in the zebrafish, including those which cause familiar PD (LRRK2, PRKN, DJ1, and PINK1), though there is no evidence of orthologs of α-syn genes (Wang et al., 2021). However, there are other orthologous genes which deletion leads to motor deficits (Figure 1) and decreased dopamine (DA) levels, such as β-, γ1-, and γ2-synuclein (Barnhill et al., 2020; Goldsmith and Jobin, 2012; Stewart et al., 2014; Doyle and Croll, 2022). However, restoring gene expression or overexpressing human α-syn these deficits are solved (Milanese et al., 2012) indicating that endogenous forms of synuclein in zebrafish could perform a function comparable to the human α-syn.
On the other hand, zebrafish have some shortcomings. There are less TG-models developed compared with other animals, the number of validated reagents is limited, and they have a natural capability of regenerating cell injuries that can mask the progression of neurodegeneration (Barbosa and Ninkovic, 2016; Caldwell et al., 2019; Clarke et al., 1987). Nonetheless, zebrafish offer a rapid and cost-effective system for early-stage drug screening and toxicological studies (Razali et al., 2021; Zambusi and Ninkovic, 2020).
Rodents
Rodents, especially mice and rats, are the most commonly used animal model in PD, being the preferred selection in 80% of all PD studies because they entail easy and well-established experimental protocols, standardized behavioral assays and a wide range of possible TG-models (Kin et al., 2019; Chia et al., 2020).
Rodent—toxic-based
From the beginning of the century, the use of toxic models in rodents became widespread (Blandini and Armentero, 2012; Kin et al., 2019; Shadrina and Slominsky, 2021), been the most frequently used MPTP, rotenone, 6-OHDA and paraquat (Blesa and Przedborski, 2014; Trigo-Damas et al., 2018). These models reproduce most of the key features of PD, such as dopaminergic neuronal loss and motor impairments (Figure 1), being 6-OHDA lesions-based model is the most commonly used in pharmacological research (Hernandez-Baltazar et al., 2017; Zeng et al., 2018). Alternatively, there are other less commonly used toxin-induced PD models in mice, such as reserpine (acting on presynaptic terminals), aminochrome (promoting mitochondrial dysfunction), or trichloroethylene or β-sitosterol β-d-glucoside (BSSG) (both inducing loss of DA neurons) and titanium dioxide nanoparticles (Van Kampen et al., 2015; Segura-Aguilar, 2017; Razali et al., 2021). Also some studies are focused in the role of protofibrils and fibrils, that represent a toxic component for TH neurons (Gómez-Benito et al., 2020; Izco et al., 2021; Dovonou et al., 2023).
Two prominent pharmacological examples where the 6-OHDA model has been used as a preclinical model and have made it to clinical phases: (I) the drug CVT-301, an inhaled formulation of levodopa (Lipp et al., 2016), increased plasma levels compared to oral administration (Bartus et al., 2004). This treatment was approved in 2018 for PD patients by the U. S. Food and Drug Administration (FDA) (Grosset et al., 2013; Lewitt et al., 2016; LeWitt and Fahn, 2016; Lipp et al., 2016; Isaacson et al., 2018). (II) Apomorphine has limited oral bioavailability, which has led to the development of different routes of parenteral administration. The rapid clinical response of apomorphine (7–10 min) has made intermittent subcutaneous injections a suitable therapy for “on/off” fluctuations (Borkar et al., 2017; Katzenschlager et al., 2018).
The MPTP mice model is another important toxic model used in the study of PD. This model has been used to develop drugs that target not only the dopaminergic pathways affected in PD, but also other aspects such as mitochondrial dysfunction, protein degradation, neuroinflammation, and the accumulation of oxidative species (Blesa et al., 2015; Belarbi et al., 2017; Balzano et al., 2023). In this sense, one of the drugs that has given promising results with neurotrophic and anti-inflammatory properties is exenatide. It is a peptide receptor agonist like glucagon 1 (GLP-1) and resistant to enzyme degradation. In MPTP mouse model, exenatide has been shown to attenuate toxicity, preserving neurons in the SNc combined with a decrease in activated microglia (Li et al., 2009; Belarbi et al., 2017). Also, the synthetic androstenetriol HE3286 has been tested in the murine MPTP model demonstrating a neuroprotective effect and reduction of motor impairment and neuroinflammation (Nicoletti et al., 2012). The clinical trial of HE3286 was completed in 2023 for PD (NCT05083260) (see Supplementary Table 1).
Finally, one of the most relevant approaches of the last decade is to promote neuronal survival using neurotrophic factors to help affected neurons in PD (Heiss et al., 2019; Lindholm and Saarma, 2022; Moriarty et al., 2022). Cerebral dopaminergic neurotrophic factor (CDNF) and glia cell line-derived neurotrophic factor (GDNF) have been shown to have a strong result in restoring the function and neuroprotection of midbrain DA neurons in murine neurotoxic models (6-OHDA and MPTP) (Kirik et al., 2000; Oiwa et al., 2006; Smith and Cass, 2007). As a result of these type of preclinical studies, clinical trials have been developed in 2019 and 2020 to test the safety of CDNF by brain infusion in patients with PD (NCT03295786 or NCT03775538) (see Supplementary Table 1). One of the solutions that has been launched to solve the handicap of the direct administration of GDNF in the brain, is the use of gene therapy by viral vectors, such as adenovirus, lentivirus, and adeno-associated virus (AAV) vectors (Enterría-Morales et al., 2020; Terse et al., 2021). This route of administration has been validated with respect to direct administration studies, demonstrating that AAV-GDNF administration in the SN or striatum prevent cell death in the rat 6-OHDA unilateral model (Kordower et al., 2000; Connor, 2001; Wang et al., 2002). Other alternative to the viral-vectors, it is a neurotensin-poliplex method, which achieves by transfection the overexpression of GDNF encapsulated nanoparticles in dopaminergic cells. This allows the expression of GDNF in a controlled and prolonged manner (Espadas-Alvarez et al., 2017). Furthermore, Yurek et al. have administered nanopolysine particles and a plasmid encoding GDNF to the midbrain of rats, improving the number of TH + cells and the rotational behavior of animals (Figure 1; Yurek et al., 2017). In this sense, in 2020 started a new clinical trial to evaluate the safety and the potential effect of the AAV2-GDNF delivered to the long-lasting PD putamen patients (NCT04167540) (see Supplementary Table 1).
Rodent—genetic
In parallel, genetic rodent models have been used for the study of personalized modifications in the genome, replicating key pathogenic axes of PD (α-synuclein aggregation, lysosomal and mitochondrial dysfunction). These models have consequently driven significant advances in understanding PD pathophysiological process of this disorder (Blesa and Przedborski, 2014). However, different models of α-syn overexpression and mutations of the principal genes (A53T, A30P, and E46K) have been created, reporting progressive motor decline, neuritic dystrophy and intraneuronal inclusions (Giasson et al., 2002; Zarranz et al., 2004; Aniszewska et al., 2022, respectively). One example of the potential therapeutic effectiveness of these models is demonstrated in studies on: B-312 active immunization (Phase 1, NCT04075318), ABBV-0805 passive immunization (Phase 1, NCT04127695), buntanetap/ANVS401 translational inhibitor (Phase 2a, NCT04524351), and UCB0599/minzasolmin (Phase 1 NCT04875962; Phase 2 NCT04658186; extension NCT05543252), together with the anti-inflammatory hypoestoxide (NCT04858074) (see Supplementary Table 1; Kim et al., 2015; Kuo et al., 2019; Nordström et al., 2021; Nimmo et al., 2022). These data illustrate the models’ value for mechanism-based pharmacology and biomarker alignment, showing significant improvements in pathological and behavioral outcomes.
TG-mice overexpressing human wild-type α-syn (SNCA) have been pivotal for advancing immunotherapies in PD. One example is prasinezumab (RO7046015) reducing α-syn pathology and improving motor performance in two complementary overexpression lines, both expressing human WT (not A53T/A30P) SNCA (Pagano et al., 2021; Richter et al., 2023). These convergent mechanistic and behavioral effects provided the basis for clinical translation (Pagano et al., 2021, 2024). Also using one of these models, a small molecule misfolding inhibitor (UCB0599/minzasolmin; predecessor NPT200-11) has been tested reducing α-syn pathology and improved PD-relevant endpoints (Price et al., 2018), leading to Phase 1 clinical studies (NCT05543252) (see Supplementary Table 1).
Additionally, TG-rodents carrying the LRRK2 mutation, which mimics dysfunction in nigrostriatal dopaminergic neurotransmission, exhibit variable locomotor changes (Figure 1) and increased tau expression and/or phosphorylation (Daniel and Moore, 2015; Seegobin et al., 2020). However, more recent constructs featuring LRRK2-G2019S have shown increased efficiency, causing approximately 50% neuronal loss in the substantia nigra pars compacta (SNc) in mice (Lee et al., 2010; Xiong et al., 2018). However, many knock-in and several overexpression lines do not show a notably SNc dopaminergic cell loss (Li et al., 2010; Tsika et al., 2014; West, 2017). As a result, preclinical models featuring LRRK2 have been instrumental in the development of novel drugs aimed at improving the pathogenesis of genetic PD. These efforts have subsequently led to clinical studies in patients (Jennings et al., 2022, 2023). In the same line for the TG-rodent model but different mutation, it is important to briefly mention the mutations in parkin and DJ-1 that seldom show frank nigrostriatal dopaminergic cell loss at baseline, despite progressive behavioral or biochemical changes (Yamaguchi and Shen, 2007; Chandran et al., 2008), but they consistently reveal mitochondrial/oxidative stress and proteostatic disturbances (Malkus et al., 2009; Lopert and Patel, 2014). Loss of parkin impairs mitophagy and can depress mitochondrial biogenesis (Stevens et al., 2015), positioning these lines as tractable platforms to interrogate proteostatic-mitophagic pathways and antioxidant strategies (Shin et al., 2020). While single-gene transgenic models have provided substantial insights into disease mechanisms, they often fail to reproduce the full spectrum of PD pathology. In this regard, “double-hit or “multi-hit” approaches, in which genetic susceptibility (e.g., LRRK2, parkin or other mutations) is combined with an environmental exposure such as rotenone, MPTP or inflammatory microenvironment may offer a more accurate representation of the complex and heterogeneous pathophysiology of PD (Rudyk and Hayley, 2019; Dwyer et al., 2020).
In relation with these TG models, drugs regulating other pathological pathways, such as protein degradation or reactive species, are being investigated. Of note here is ambroxol, which can regulate the activity of Gcase (an enzyme involved in lysosomal function). In the study by Migdalska-Richards et al., ambroxol increased Gcase activity and reduced the level of α-syn in L444P TG-mice in the Gba1 gene and in overexpressing α-syn TG-mice (Migdalska-Richards et al., 2016). Their promising preclinical findings prompted the launch of different clinical trials focused on different aspects of PD (NCT02941822 and NCT02914366) (see Supplementary Table 1).
Definitely, murine models, whether toxin-induced or transgenic, have been instrumental in advancing pharmacological studies for PD, enabling some drugs to progress to clinical phases.
Minipigs
Minipigs have gained popularity in recent years due to their anatomical and physiological similarities with humans, particularly with regards to the brain. The large size of the SNc and striatum in minipigs make them particularly suitable for neuroimaging and electrophysiological studies. The study of PD in minipigs started to surface back in 1999 when M. Mikkelsen et al. administered MPTP to nine pigs, which caused the animals to develop muscle rigidity, hypokinesia and impaired coordination (Figure 1). Also, the histological assessment revealed a reduction of the striatal amount of DA and a decrease of the number of DA cells in the SNc, 3 months after MPTP administration (Mikkelsen et al., 1999). More recent studies have administered different toxins such as 6-OHDA in the nigrostriatal pathway, or have inoculated lactacystin in the forebrain inhibiting the ubiquitin proteasome system (Christensen et al., 2018; Lillethorup et al., 2018; Zhu et al., 2018; Del Rey et al., 2021). In the past few years, genetic models based on this animal with human SNCA or simultaneous mutations in different genes using CRISPR/Cas9-mediated gene editing have been created (Mehmood et al., 2021; Shadrina and Slominsky, 2021). As in murine models, drugs of new formulations of L-DOPA, such as ND0612, have been tested. ND0612 is a liquid formulation of levodopa and carbidopa that can be administered subcutaneously through a patch device. It has completed phase III trials and has shown promising results (Ramot et al., 2017; McFarthing et al., 2021). Moreover, research utilizing subcutaneous apomorphine and emerging treatments such as GDNF has been undertaken in preclinical trials employing the minipig model. These studies, highlighted by Ramot et al. (2018), Ramot et al. (2017), and Wahlberg et al. (2020), underscore the adaptability and efficacy of this model in advancing novel therapies for Parkinson’s disease. These instances illustrate merely a fraction of the potential contributions the minipig.
Dogs
Dogs have been categorized as a natural model of PD due to significant similarities in diet, microbiomes and neuropathological features (Ambrosini et al., 2019). Furthermore, these models have been used to test pharmacology treatments, such as the CV-301, where maximum absorption times were studied in a dog model (beagle) (Watanabe et al., 2015; Jeong et al., 2016; Lipp et al., 2016). MPTP-intoxicated dogs exhibit clinical symptoms resembling those of PD, including akinesia, head tremor, and reduced oculovestibular reflex activity (Figure 1), along with other pathophysiological characteristics like abnormal concentrations or decreases of certain substances (Podell et al., 2003; van Vliet et al., 2008; Choi et al., 2011; Lee et al., 2013; Yang Y. et al., 2020). However, despite their potential, the utilization of dogs in PD research remains relatively uncommon due to their high cost, ethical considerations and limited availability of genetically modified lines.
Non-human primates
Finally, non-human primate (NHP) models have played a crucial role in elucidating the pathophysiology of PD, as their genetic, physiological, and behavioral similarities to humans enable the reproduction of distinct disease stages that closely parallel those observed in patients (Blesa et al., 2018).
Non-human primate—toxic-based
The most extended toxic model of NHPs is the MPTP model. This MPTP model simulates the topographic pattern of dopaminergic degeneration in the striatum characteristic of PD (Blesa et al., 2012; Del Rey et al., 2024).
Depending on the toxic administration protocol, the acute and chronic stages of the pathological process can be reproduced (Figure 1; Blesa et al., 2011, 2012; Seo et al., 2019; Molinet-Dronda et al., 2022). However, the MPTP-NHP model has the same problem as other drug-induced models: the lack of α-syn aggregates (Halliday et al., 2009). Even so, this model has been essential in PD neuropharmacology to discern the side effects of L-DOPA and characterize the motor fluctuations induced by L-DOPA typical of patients (Cenci and Crossman, 2018; Hamadjida et al., 2020). In addition, in the pharmacological field, NHPs have tended to be used as a last step toward the clinic due to the advantages and disadvantages exposed above. Therefore, there are only a few drugs tested in this model. One example of the use of NHP could be trophic factors, administered intraparenchymal freely or in AAV. Mainly, toxic models have been used for research with trophic factors such as CDNF and GDNF, capable of promoting neuronal survival and repair (Garea-Rodríguez et al., 2016; Kordower and Burke, 2018). Such is the efficacy that several clinical trials tested these compounds (NCT01621581 and NCT06285643) (see Supplementary Table 1). One example is the effectiveness of AA2-GDNF, which is now in Phase II of a clinical trial after successful completion of Phase I (NCT06285643) (see Supplementary Table 1).
Non-human primate—genetic
Due to the absence of α-syn aggregates in the MPTP-NHP model, studies like that of Kirik et al. explored advancements in viral vector technology, enabling the unilateral overexpression of human wild-type α-syn and the A53T mutant α-syn in the SNc of adult marmosets (Kirik et al., 2003). Both wild-type and mutant A53T animals developed PD-like pathological manifestation (Figure 1) and histopathological studies revealed α-syn cytoplasmatic inclusions, granular deposits, and dystrophic neurites as well as 30–60% degeneration of DA neurons in the SNc (Kirik et al., 2003). However, subsequent studies using different viral vectors in NHPs showed variability in results between species (Eslamboli et al., 2007; Bourdenx et al., 2015; Yang et al., 2015; Marmion and Kordower, 2018). Precisely, Bourdenx et al. (2015) did not find any aging effects on the nigrostriatal pathway due to overexpression of mutant A53T α-syn via a recombinant AAV2/9 vector, which had previously been effective in rats. Recently, Koprich et al. (2016) conducted a series of experiments testing an AAV1/2 vector with overexpression of mutant A53T α-syn, which showed a 50% loss of nigral DA neurons and a 60% reduction in striatal DA content after 4 months (Koprich et al., 2016).
On the other hand, TG-NHPs are not abundant due to ethical concerns, cost and long gestation and maturation. However, the study of Niu et al. (2015) developed the first progressive transgenic model of PD. They used a lentiviral vector expressing A53T α-syn under human ubiquitin promoter in a rhesus macaque. This model showed an age-dependent non-motor symptoms such as cognitive impairments and some fine motor coordination deficits, though no clear evidence of neuronal degeneration in the magnetic resonance imaging (MRI) was observed (Niu et al., 2015).
In summary, vertebrate models provide a tiered framework that integrates increasing experimental complexity with enhanced translational relevance. Lower vertebrates like zebrafish enable rapid screening, and rodents offer a standardized stage. On the other hand, larger mammals support therapy development and pharmacokinetic testing, and NHPs provide the most clinically relevant models for evaluating safety and efficacy prior to human trials. A clear understanding of the strengths and limitations of each model is critical for aligning model selection with the specific research question and phase of drug development.
Modeling non-motor symptoms of Parkinson’s disease
As outlined above, non-motor symptoms constitute a critical component of PD, emerging well before dopaminergic degeneration and underscoring the need for their early detection. Because of that, animal models focused on this aspect are essential to develop new therapeutical strategies to treat early symptoms such as: hyposmia, constipation, alteration in REM, RBD, anxiety/depression, and cognitive decline, as the most common ones in PD (Postuma et al., 2015; Heinzel et al., 2019).
Animal models for prodromal PD are mainly based on toxins, genetics, and overexpression of α-syn. It should also be noted that not all animal models described above can reproduce this phase of PD, because of either the type of species used or the poor reproducibility of the model (Taguchi et al., 2020). Furthermore, many non-motor symptoms of PD observed in animal models—as in patients—can arise from distinct underlying mechanisms, and their manifestation may vary considerably not only between different studies but also within the same model. This variability highlights the complexity of accurately replicating non-motor features of PD in preclinical research (Wichmann et al., 2025).
Invertebrate models
Drosophila melanogaster
Invertebrate models have also contributed to the study of certain non-motor symptoms-related mechanisms. In Drosophila, transgenic lines have shown certain life activities which depends on normal DA levels: as olfactory conditioning, sleep and arousal regulation, and memory and learning process (Figure 1; Selcho et al., 2009; Berry et al., 2012; Ueno et al., 2012; Dung and Thao, 2018). Specifically, PINK1 and parkin null Drosophila models have reported changes in learning and memory and in circadian rhythms (Julienne et al., 2017). Moreover, disruptions of light/dark behavior have been corroborated though electrophysiological studies in l-LNv clock neurons (responsible for circadian rhythms synchronization) which have shown an abnormal activity (Parisky et al., 2008; Hirsh et al., 2010; Shang et al., 2011; Julienne et al., 2017). The number of drugs tested in this non-motor model are few, however, Lawal et al., conducted unbiased Drosophila neuropsychiatric screens that identified aminergic agents and additional candidates affecting dopaminergic and sleep/circadian phenotypes, illustrating the platform’s capacity to uncover repurposing leads for PD (Lawal et al., 2014).
Vertebrate models
Zebrafish
Although less explored for non-motor symptoms, zebrafish models are increasingly used in this area. Zebrafish possess a pool of cognitive processes comprising learning, memory, fear, anxiety, perception, social skills and sleep patterns which can provide the opportunity of non-motor studies and the research of the prodromic phase of PD (Bailey et al., 2013). For example, rotenone-exposed adults display PD-like motor and non-motor phenotypes (Wang et al., 2017); cognitive readouts such as short-term avoidance memory and lateralization are robust (Figure 1; Fontana et al., 2018); and standardized adult pipelines allow concurrent quantification of motor and non-motor behaviors (Selvaraj et al., 2019). However, the absence of higher cortical structures and the limited repertoire of measurable affective behavior restrict the depth of non-motor symptoms modeling in this type of species.
Rodents
Rodent—toxic-based
In rodents, it has been demonstrated that olfactory dysfunction exists after the first injection with MPTP in a mouse model (Schintu et al., 2009), after intranasal administration of MPTP in rats, and also administering rotenone in rats and mice (Figure 1; Prediger et al., 2009; Babatunde et al., 2023; Yuan et al., 2023). Similarly, another of the non-motor symptoms studies from which results have been obtained in most traditional toxin models is constipation; MPTP exposure in mice has been reported to decrease the population of dopaminergic neurons and produce gastrointestinal impairments with an increment of intestinal motility and relaxation of the colon (Anderson and Gibbons, 2007). Also, rotenone-treated rats have been shown to show abnormal gastrointestinal function (Drolet et al., 2009). Moreover, rotenone-treated rats and in general MPTP-treated rodents shown sleep and circadian rhythm alterations (Laloux et al., 2008; Chen et al., 2013). Finally, in the 6-OHDA rat model, the reduction of dopaminergic neurons that is considered responsible for the RBD expressed in this model has been reported several times. However, there were no pathological changes in the most affected regions in RBD (Vo et al., 2014; Medeiros et al., 2019). In these cases, Ceftriaxone and intranasal insulin have been tested in MPTP rat models and 6-OHDA rat model demonstrating good effects in working memory, object recognition and cognitive deficits (Figure 1; Huang et al., 2015; Hsieh et al., 2017; Yang L. et al., 2020). Both drugs are ongoing in clinical trials with non-motor symptoms PD patients (NCT03413384 and NCT02064166) (see Supplementary Table 1; Degirmenci et al., 2023).
Rodent—genetic
Various strategies have been employed to generate different non-motor phenotypes in TG models. One example involves prion promoter-mice, where they exhibit reduced intestinal motility at 3 months, accompanied by the emergence of α-syn aggregates in the mesenteric and submucosal plexus of the colon (Rota et al., 2019; Taguchi et al., 2020). Another approach utilizes the Thy1-promoter, effectively replicating many non-motor symptoms in mice. This includes hyposmia at 3–5 months, anxiety at 4 months, sleep disturbances at 10 months, and intestinal dysfunction at 12 months (Fleming et al., 2008; Taylor et al., 2009; Chesselet et al., 2012; Taguchi et al., 2020). A third strategy employs endogenous α-syn promoters using bacterial artificial chromosome (BAC) and P1-derived artificial chromosome (PAC). A53T-SNCA BAC models have reported partial replication of the early PD process, encompassing non-motor symptoms like hyposmia and RBD (Taguchi et al., 2020). Conversely, A53T SNCA mice with PAC demonstrated reduced intestinal motility and prolonged intestinal transit time (Kuo et al., 2010). Rasagiline in this case has shown improvements in olfactory deficits in an α-syn-mouse model, overexpressing wild-type human α-syn (Petit et al., 2013), and has been tested in PD patients with different non-motor symptoms as hyposmia, apathy, cognition or sleep (NCT00902941, NCT00755027, NCT01382342, NCT01032486, respectively) (see Supplementary Table 1). Another example, is Buntanetap (also known as posiphen or ANVS-40), an enantiomer traditionally inhibitor that reduces amyloid precursor protein and α-synuclein expression in brain and gut of A53T transgenic mice (Kuo et al., 2019). The clinical trials were completed in December of 2021 with positive results in terms of improvements of motor function (especially in patients with a longer disease duration) and a maintenance or improvement in cognitive state in certain subgroups, with an acceptable safety and tolerability profile, and side effects mainly attributed to CSF catheterization for the SILK analysis (NCT05357989); full peer-reviewed efficacy results are pending (see Supplementary Table 1; Galasko et al., 2024).
Mitochondrial models such as Mitopark mice, in which the mitochondrial transcription factor TFAM in specifically deleted (Westerlund et al., 2008), have described in the studies certain non-motor deficits; circadian desynchronization, early cognitive impairments (including olfactory disfunction, learning and memory deficits), anxiety or depression (Figure 1; Li et al., 2013; Fifel and Cooper, 2014; Langley et al., 2021).
Finally, it is important to consider the route of administration when modeling α-synucleinopathies and non-motor features in mice. Injections of α-syn fibrils into the olfactory bulb or the gastrointestinal tract reproduce non-motor symptoms alongside α-syn pathology. By contrast, subcutaneous administration has been reported to elicit non-motor readouts without robust central α-syn pathology. A recent alternative is intravenous delivery using viral vectors: in addition to inducing α-syn pathology across multiple brain regions and the small intestine, this approach yields measurable non-motor phenotypes—for example, olfactory-bulb seeding drives pathology within olfactory circuits with early neuronal loss; gastrointestinal inoculation produces enteric α-syn pathology with impaired gastrointestinal transit; and intravenous AAV-mediated α-syn overexpression leads to widespread central and enteric pathology accompanied by anxiety-like behavior and mild deficits in attention/working-memory paradigms (Manfredsson et al., 2018; Del Rey N. L. et al., 2018; Kuan et al., 2021).
Dogs
Interestingly, dog gut microbiome more closely resembles the human gut microbiome in composition and functional overlap, compared with rodent models. Therefore, it may be a potential model for the study of gut-brain interactions in PD. However, the canine models represent a minority in case of PD research (Ambrosini et al., 2019).
Non-human primates
Non-human primate—toxic-based
The MPTP-NHP model has revealed loss of REM sleep and increased napping after the first intravenous injection of MPTP (0.5 mg/kg) (Barraud et al., 2009). Following studies have monitored that electroencephalographic activity shows suppression of REM sleep after first MPTP injection and chronic MPTP administration results in progressive sleep deterioration, fragmentation, and reduced sleep efficacy with a corresponding increased sleepiness during the day by about 50% (Barraud et al., 2009; Fox and Brotchie, 2010). Moreover, studies in MPTP marmosets have reported an increased muscle tone during REM sleep (Fox and Brotchie, 2010; Verhave et al., 2011; Bezard and Fernagut, 2014; Bao et al., 2023). Additionally, MPTP-NHP model has also been used to describe cognitive deficits during attentional and executive task performance, by measuring visual discrimination, delayed response or matching-to-sample, object retrieval/detour tasks that are impaired even in MPTP-treated NHPs with minimal motor deficits (Figure 1; Bao et al., 2023). Furthermore, assessments of self-initiated and visually-triggered saccades in MPTP monkeys have revealed increased errors, including an increased tendency to initiate inappropriate or premature responses (Schneider and Kovelowski, 1990; Taylor et al., 1990; Slovin et al., 1999; Pessiglione et al., 2008; Fox and Brotchie, 2010). A good example of preclinical study in MPTP models is the NYX-458, a NMDAR modulator which improves cognitive performance in terms of working memory, attention and executive function (Barth et al., 2020; Degirmenci et al., 2023). NYX-458 effects has been demonstrating in an active Phase II clinical trial on mild cognitive impairment, mild dementia associated with PD or prodromal or manifest Lewy Body dementia (NCT04148391) (see Supplementary Table 1). Indeed, the MPTP-NHP model stands as the gold standard when it comes to study cognitive and sleep deficits in PD. This model has proven to be invaluable in enhancing outcome measurements, facilitating the translation of preclinical findings into potentially beneficial drug interventions for PD.
In summary, although modeling non-motor symptoms remains a challenge due to their heterogeneity and complexity, significant progress has been made across species. Rodents and NHPs currently allow the most robust approaches to assessing cognitive, olfactory, and gastrointestinal symptoms of PD, while invertebrates and fish models provide understanding of individual physiological processes. A comprehensive understanding of non-motor symptoms requires integrative approaches that combine behavioral, histological, and molecular strategies in different models.
Conclusion
Animal models are indispensable tools for investigating the complex pathophysiology of PD and for evaluating novel therapeutic strategies. They have significantly advanced our understanding of the disease and facilitated the identification of promising new compounds. Nevertheless, despite decades of research with a broad spectrum of preclinical models—from cellular models and rodent platforms to NHPs—PD still lacks a disease modifying therapy. This unresolved challenge underscores a critical gap: while current models reproduce many motor and non-motor features of PD, none of them fully capture its complexity, heterogeneity, or progressive nature of the disorder (Figure 2).
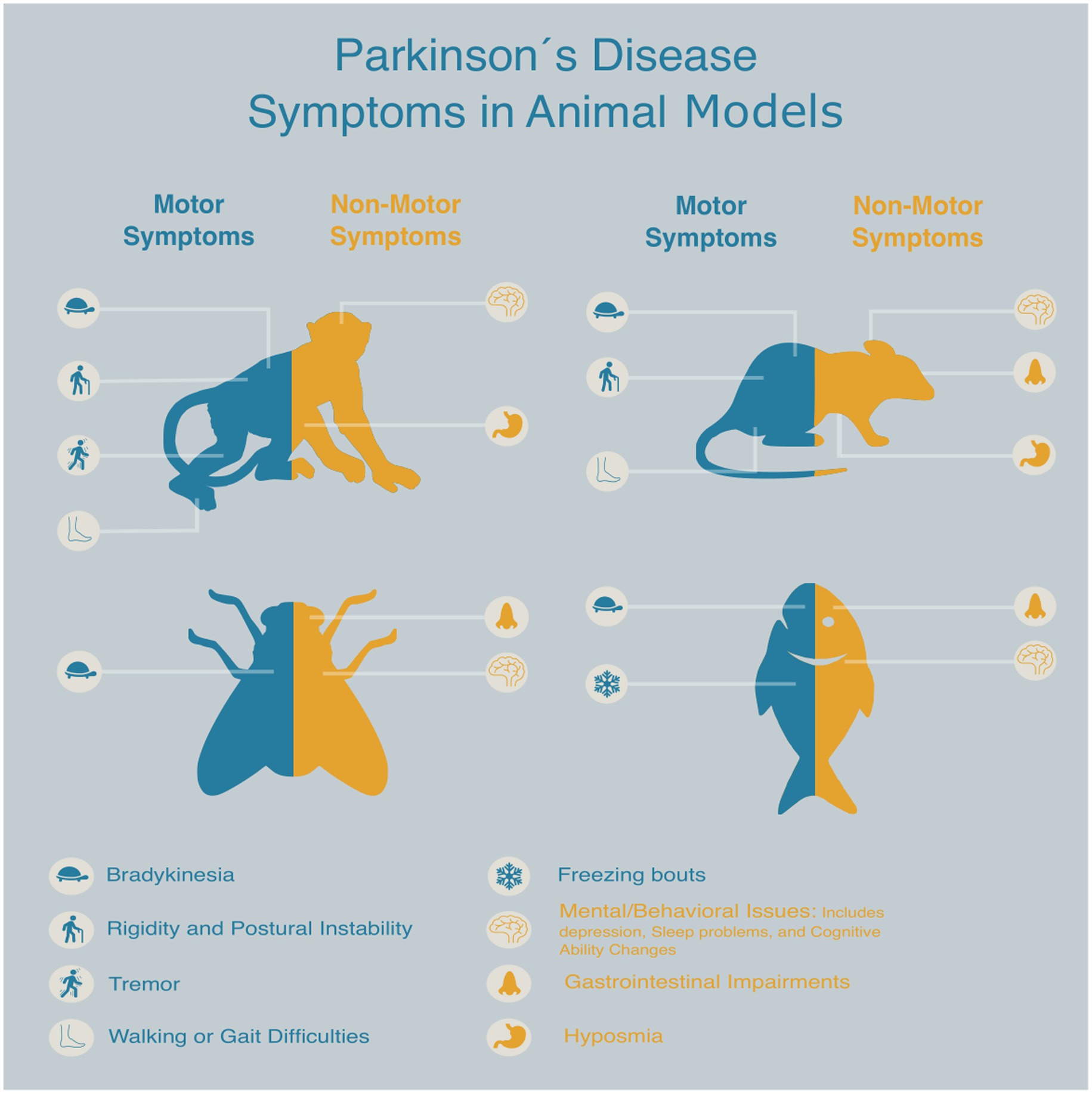
Figure 2. Schematic representation of key animal models commonly used in Parkinson’s disease drug discovery, highlighting the presence and treatment of both motor and non-motor symptoms associated with each model.
Each model contributes to our understanding of PD pathophysiology, yet none is sufficient on its own. A clear example of this limitation is the repeated failure of many promising compounds that perform well in preclinical stages but do not translate into successful clinical outcomes, highlighting the restricted predictive validity of existing systems. Cellular and invertebrate models enable mechanistic studies and pharmacological screening rodent models provide reproducible platforms for behavioral or molecular analyses; then NHP models offer high translational value for testing therapeutic strategies. Moreover, most models have traditionally focused on motor deficits, although by the time these appear neurodegeneration has already begun; this realization has shifted attention toward non-motor symptoms and prodromal stages as earlier and more predictive indicators of the disease (Table 1).
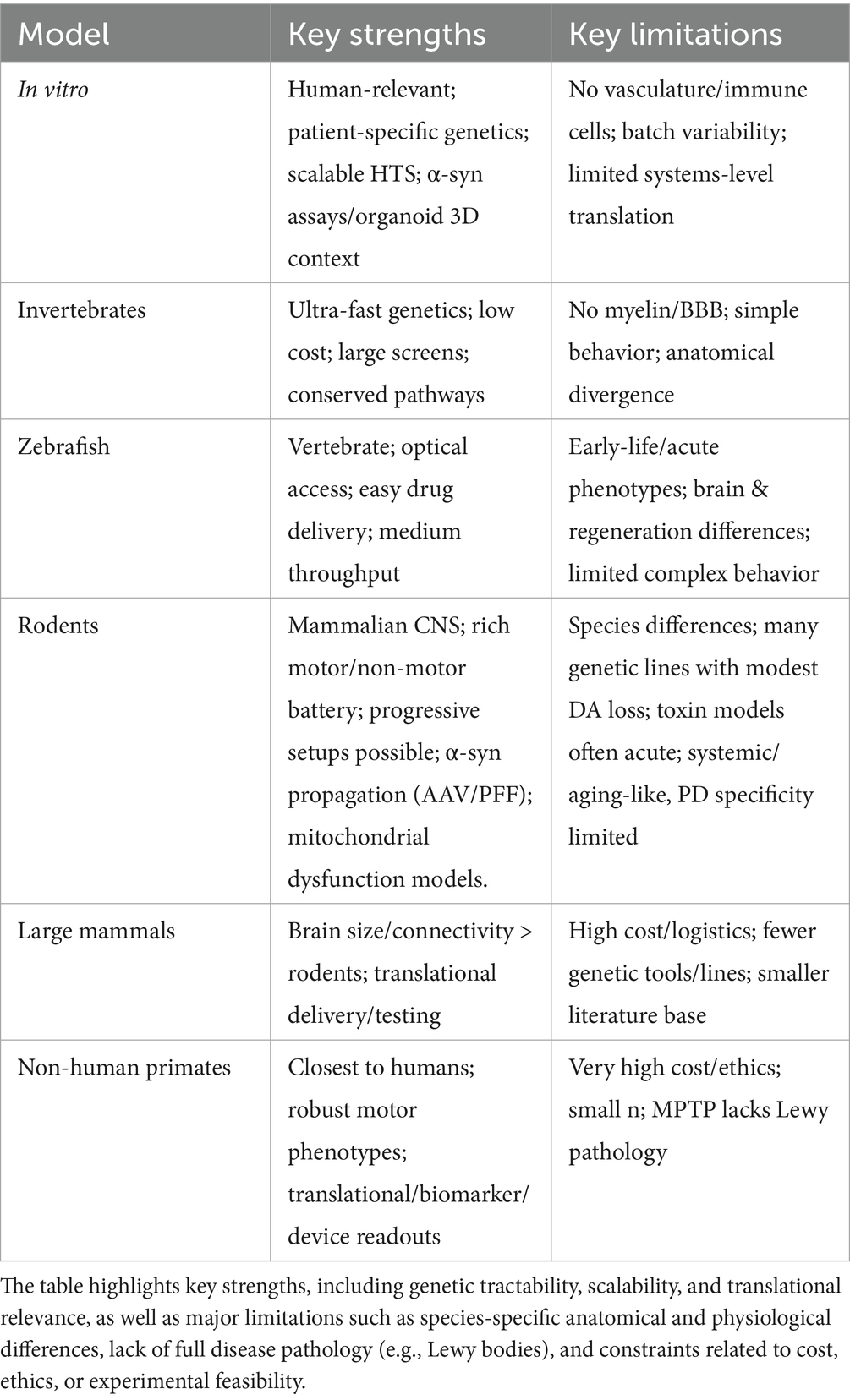
Table 1. Summary of experimental models commonly employed in Parkinson’s disease (PD) research.
Looking ahead, improving the predictive power of PD models will require an integrative approach that incorporates genetic, environmental, and age-related factors reflecting both motor and non-motor aspects across disease progression. Greater alignment between preclinical endpoints and clinical outcomes, together with stronger emphasis on inter-individual variability, will also be a key to boosting translational applicability. Ultimately, the selection and refinement of existing models, alongside the development of new and more representative platforms, will be key to more accurately evaluating novel therapies, identifying reliable biomarkers for early intervention, and effectively bridging the gap between promising preclinical findings and meaningful clinical benefits for patients with PD.
Author contributions
AR-S: Methodology, Investigation, Writing – review & editing, Conceptualization, Writing – original draft. NE-G: Writing – original draft, Conceptualization, Writing – review & editing, Investigation, Methodology. MT: Writing – review & editing. IT-D: Validation, Conceptualization, Writing – review & editing, Formal analysis, Supervision, Writing – original draft, Funding acquisition.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. Instituto de salud carlos III PFIS (FI21/000919) (N.E-G.) XI Convocatoria de Investigación de la Universidad Camilo José Cela (NeuroMelNHP) (I. T-D).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnagi.2025.1692592/full#supplementary-material
References
Abolaji, A. O., Adedara, A. O., Adie, M. A., Vicente-Crespo, M., and Farombi, E. O. (2018). Resveratrol prolongs lifespan and improves 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced oxidative damage and behavioural deficits in Drosophila melanogaster. Biochem. Biophys. Res. Commun. 503, 1042–1048. doi: 10.1016/j.bbrc.2018.06.114
Adedara, A. O., Babalola, A. D., Stephano, F., Awogbindin, I. O., Olopade, J. O., Rocha, J. B. T., et al. (2022). An assessment of the rescue action of resveratrol in parkin loss of function-induced oxidative stress in Drosophila melanogaster. Sci. Rep. 12:3922. doi: 10.1038/s41598-022-07909-7
Ahlskog, J. E., and Muenter, M. D. (2001). Frequency of levodopa-related dyskinesias and motor fluctuations as estimated from the cumulative literature. Mov. Disord. 16, 448–458. doi: 10.1002/mds.1090
Ambrosini, Y. M., Borcherding, D., Kanthasamy, A., Kim, H. J., Willette, A. A., Jergens, A., et al. (2019). The gut-brain axis in neurodegenerative diseases and relevance of the canine model: a review. Front. Aging Neurosci. 11:130. doi: 10.3389/fnagi.2019.00130
Anderson, L. M., and Gibbons, G. H. (2007). Notch: a mastermind of vascular morphogenesis. J. Clin. Invest. 117, 299–302. doi: 10.1172/JCI31288
Aniszewska, A., Bergström, J., Ingelsson, M., and Ekmark-Lewén, S. (2022). Modeling Parkinson’s disease-related symptoms in alpha-synuclein overexpressing mice. Brain Behav. 12:e2628. doi: 10.1002/brb3.2628
Armstrong, M. J., and Okun, M. S. (2020). Diagnosis and treatment of Parkinson disease: a review. JAMA 323, 548–560. doi: 10.1001/jama.2019.22360
Aryal, B., and Lee, Y. (2019). Disease model organism for Parkinson disease: Drosophila melanogaster. BMB Rep. 52, 250–258. doi: 10.5483/BMBRep.2019.52.4.204
Athauda, D., Maclagan, K., Skene, S. S., Bajwa-Joseph, M., Letchford, D., Chowdhury, K., et al. (2017). Exenatide once weekly versus placebo in Parkinson’s disease: a randomised, double-blind, placebo-controlled trial. Lancet 390, 1664–1675. doi: 10.1016/S0140-6736(17)31585-4
Auluck, P. K., Caraveo, G., and Lindquist, S. (2010). α-Synuclein: membrane interactions and toxicity in parkinson’s disease. Annu. Rev. Cell Dev. Biol. 26, 211–233. doi: 10.1146/annurev.cellbio.042308.113313
Ay, M., Luo, J., Langley, M., Jin, H., Anantharam, V., Kanthasamy, A., et al. (2017). Molecular mechanisms underlying protective effects of quercetin against mitochondrial dysfunction and progressive dopaminergic neurodegeneration in cell culture and MitoPark transgenic mouse models of Parkinson’s disease. J. Neurochem. 141, 766–782. doi: 10.1111/jnc.14033
Babatunde, B. R., Adeyeye, T. A., Johnson, V. F., and Shallie, P. D. (2023). Rotenone induced olfactory deficit in Parkinson’s disease rat model: the protective role of adenosine A2A receptors antagonist. J. Chem. Neuroanat. 127:102188. doi: 10.1016/j.jchemneu.2022.102188
Bach, J.-P., Ziegler, U., Deuschl, G., Dodel, R., and Doblhammer-Reiter, G. (2011). Projected numbers of people with movement disorders in the years 2030 and 2050. Mov. Disord. 26, 2286–2290. doi: 10.1002/mds.23878
Bailey, J., Oliveri, A., and Levin, E. D. (2013). Zebrafish model systems for developmental neurobehavioral toxicology. Birth Defects Res. C Embryo Today Rev. 99, 14–23. doi: 10.1002/bdrc.21027
Balzano, T., Esteban-García, N., and Blesa, J. (2023). Neuroinflammation, immune response and α-synuclein pathology: how animal models are helping us to connect dots. Expert Opin. Drug Discov. 18, 13–23. doi: 10.1080/17460441.2023.2160440
Bandres-Ciga, S., Saez-Atienzar, S., Bonet-Ponce, L., Billingsley, K., Vitale, D., Blauwendraat, C., et al. (2019). The endocytic membrane trafficking pathway plays a major role in the risk of Parkinson’s disease. Mov. Disord. 34, 460–468. doi: 10.1002/mds.27614
Bao, Y., Gan, C., Chen, Z., Qi, Z., Meng, Z., and Yue, F. (2023). Quantification of non-motor symptoms in parkinsonian Cynomolgus monkeys. Brain Sci. 13:1153. doi: 10.3390/brainsci13081153
Barbosa, J. S., and Ninkovic, J. (2016). Adult neural stem cell behavior underlying constitutive and restorative neurogenesis in zebrafish. Neurogenesis 3:e1148101. doi: 10.1080/23262133.2016.1148101
Barker, R. A., and Björklund, A. (2020). Animal models of Parkinson’s disease: are they useful or not? J. Parkinsons Dis. 10, 1335–1342. doi: 10.3233/JPD-202200
Barnhill, L. M., Murata, H., and Bronstein, J. M. (2020). Studying the pathophysiology of Parkinson’s disease using zebrafish. Biomedicine 8:197. doi: 10.3390/biomedicines8070197
Barraud, Q., Lambrecq, V., Forni, C., McGuire, S., Hill, M., Bioulac, B., et al. (2009). Sleep disorders in Parkinson’s disease: the contribution of the MPTP non-human primate model. Exp. Neurol. 219, 574–582. doi: 10.1016/j.expneurol.2009.07.019
Barth, A. L., Schneider, J. S., Johnston, T. H., Hill, M. P., Brotchie, J. M., Moskal, J. R., et al. (2020). NYX-458 improves cognitive performance in a primate Parkinson’s disease model. Mov. Disord. 35, 640–649. doi: 10.1002/mds.27962
Bartus, R. T., Emerich, D., Snodgrass-Belt, P., Fu, K., Salzberg-Brenhouse, H., Lafreniere, D., et al. (2004). A pulmonary formulation of L-dopa enhances its effectiveness in a rat model of Parkinson’s disease. J. Pharmacol. Exp. Ther. 310, 828–835. doi: 10.1124/jpet.103.064121
Belarbi, K., Cuvelier, E., Destée, A., Gressier, B., and Chartier-Harlin, M. C. (2017). NADPH oxidases in Parkinson’s disease: a systematic review. Mol. Neurodegener. 12, 1–18. doi: 10.1186/s13024-017-0225-5
Berg, D., Adler, C. H., Bloem, B. R., Chan, P., Gasser, T., Goetz, C. G., et al. (2018). Movement disorder society criteria for clinically established early Parkinson’s disease. Mov. Disord. 33, 1643–1646. doi: 10.1002/mds.27431
Berry, J. A., Cervantes-Sandoval, I., Nicholas, E. P., and Davis, R. L. (2012). Dopamine is required for learning and forgetting in Drosophila. Neuron 74, 530–542. doi: 10.1016/j.neuron.2012.04.007
Bezard, E., and Fernagut, P.-O. (2014). Premotor Parkinsonism models. Parkinsonism Relat. Disord. 20, S17–S19. doi: 10.1016/S1353-8020(13)70007-5
Billingsley, K. J., Barbosa, I. A., Bandrés-Ciga, S., Quinn, J. P., Bubb, V. J., Deshpande, C., et al. (2019). Mitochondria function associated genes contribute to Parkinson’s disease risk and later age at onset. NPJ Parkinsons Dis. 5:1. doi: 10.1038/s41531-019-0080-x
Blandini, F., and Armentero, M.-T. (2012). Animal models of Parkinson’s disease. FEBS J. 279, 1156–1166. doi: 10.1111/j.1742-4658.2012.08491.x
Blesa, J., Foffani, G., Dehay, B., Bezard, E., and Obeso, J. A. (2022). Motor and non-motor circuit disturbances in early Parkinson disease: which happens first? Nat. Rev. Neurosci. 23, 115–128. doi: 10.1038/s41583-021-00542-9
Blesa, J., García-Cabezas, A., and Sánchez-González, C. (2011). Inter-hemispheric asymmetry of nigrostriatal dopaminergic lesion: a possible compensatory mechanism in Parkinson’s disease. Front. Syst. Neurosci. 5:92. doi: 10.3389/fnsys.2011.00092
Blesa, J., Pifl, C., Sánchez-González, M. A., Juri, C., García-Cabezas, M. A., Adánez, R., et al. (2012). The nigrostriatal system in the presymptomatic and symptomatic stages in the MPTP monkey model: a PET, histological and biochemical study. Neurobiol. Dis. 48, 79–91. doi: 10.1016/j.nbd.2012.05.018
Blesa, J., and Przedborski, S. (2014). Parkinson’s disease: animal models and dopaminergic cell vulnerability. Front. Neuroanat. 8:155. doi: 10.3389/fnana.2014.00155
Blesa, J., Trigo-Damas, I., Del Rey, N. L. G., and Obeso, J. A. (2018). The use of nonhuman primate models to understand processes in Parkinson’s disease. J. Neural Transm. 125, 325–335. doi: 10.1007/s00702-017-1715-x
Blesa, J., Trigo-Damas, I., Dileone, M., Del Rey, N. L. G., Hernandez, L. F., and Obeso, J. A. (2017). Compensatory mechanisms in Parkinson’s disease: circuits adaptations and role in disease modification. Exp. Neurol. 298, 148–161. doi: 10.1016/j.expneurol.2017.10.002
Blesa, J., Trigo-Damas, I., Quiroga-Varela, A., and Jackson-Lewis, V. R. (2015). Oxidative stress and Parkinson’s disease. Front. Neuroanat. 9:91. doi: 10.3389/fnana.2015.00091
Blesa, J., Trigo-Damas, L., Quiroga-Varela, L., and Jackson-Lewis, V. R. (2016a). Animal models of Parkinson’s disease, en J. Dorszewska and W. Kozubski (eds.) Challenges in Parkinson’s disease. Cham, Switzerland: Springer International Publishing AG.
Blesa, J., Trigo-Damas, L., Quiroga-Varela, A., and Jackson-Lewis, V. R. (2016b). Parkinson’s disease-associated mutations affect mitochondrial function, 139–158.
Bloem, B. R., Okun, M. S., and Klein, C. (2021). Parkinson’s disease. Lancet 397, 2284–2303. doi: 10.1016/S0140-6736(21)00218-X
Borkar, N., Andersson, D. R., Yang, M., Müllertz, A., Holm, R., and Mu, H. (2017). Efficacy of oral lipid-based formulations of apomorphine and its diester in a Parkinson’s disease rat model. J. Pharm. Pharmacol. 69, 1110–1115. doi: 10.1111/jphp.12758
Bourdenx, M., Dovero, S., Engeln, M., Bido, S., Bastide, M. F., Dutheil, N., et al. (2015). Lack of additive role of ageing in nigrostriatal neurodegeneration triggered by α-synuclein overexpression. Acta Neuropathol. Commun. 3:46. doi: 10.1186/s40478-015-0222-2
Braak, H., Tredici, K. D., Rüb, U., de Vos, R. A. I., Jansen Steur, E. N. H., and Braak, E. (2003). Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol. Aging 24, 197–211. doi: 10.1016/S0197-4580(02)00065-9
Büttner, S., Faes, L., Reichelt, W. N., Broeskamp, F., Habernig, L., Benke, S., et al. (2013). The Ca2+/Mn2+ ion-pump PMR1 links elevation of cytosolic Ca(2+) levels to α-synuclein toxicity in Parkinson’s disease models. Cell Death Differ. 20, 465–477. doi: 10.1038/cdd.2012.142
Caldwell, L. J., Davies, N. O., Cavone, L., Mysiak, K. S., Semenova, S. A., Panula, P., et al. (2019). Regeneration of dopaminergic neurons in adult zebrafish depends on immune system activation and differs for distinct populations. J. Neurosci. Off. J. Soc. Neurosci. 39, 4694–4713. doi: 10.1523/JNEUROSCI.2706-18.2019
Casu, M. A., Mocci, I., Isola, R., Pisanu, A., Boi, L., Mulas, G., et al. (2020). Neuroprotection by the immunomodulatory drug Pomalidomide in the Drosophila LRRK2WD40 genetic model of Parkinson’s disease. Front. Aging Neurosci. 12:31. doi: 10.3389/fnagi.2020.00031
Cenci, M. A., and Crossman, A. R. (2018). Animal models of l-dopa-induced dyskinesia in Parkinson’s disease. Mov. Disord. 33, 889–899. doi: 10.1002/mds.27337
Chandran, J. S., Lin, X., Zapata, A., Höke, A., Shimoji, M., Moore, S. O., et al. (2008). Progressive behavioral deficits in DJ-1 deficient mice are associated with normal nigrostriatal function. Neurobiol. Dis. 29, 505–514. doi: 10.1016/j.nbd.2007.11.011
Charvin, D., Medori, R., Hauser, R. A., and Rascol, O. (2018). Therapeutic strategies for Parkinson disease: beyond dopaminergic drugs. Nat. Rev. Drug Discov. 17, 804–822. doi: 10.1038/nrd.2018.136
Chaudhuri, K. R., Azulay, J. P., Odin, P., Lindvall, S., Domingos, J., Alobaidi, A., et al. (2024). Economic burden of Parkinson’s disease: a multinational, real-world, cost-of-illness study. Drug. Real Wor. Outc. 11, 1–11. doi: 10.1007/s40801-023-00410-1
Chen, H., Burton, E. A., Ross, G. W., Huang, X., Savica, R., Abbott, R. D., et al. (2013). Research on the premotor symptoms of Parkinson’s disease: clinical and etiological implications. Environ. Health Perspect. 121, 1245–1252. doi: 10.1289/ehp.1306967
Chen, Q., Thorpe, J., and Keller, J. N. (2005). α-Synuclein alters proteasome function, protein synthesis, and stationary phase viability*. J. Biol. Chem. 280, 30009–30017. doi: 10.1074/jbc.M501308200
Chen, Y., Hong, Z., Wang, J., Liu, K., Liu, J., Lin, J., et al. (2023). Circuit-specific gene therapy reverses core symptoms in a primate Parkinson’s disease model. Cell 186, 5394–5410.e18. doi: 10.1016/j.cell.2023.10.004
Chesselet, M.-F., Richter, F., Zhu, C., Magen, I., Watson, M. B., and Subramaniam, S. R. (2012). A progressive mouse model of Parkinson’s disease: the Thy1-aSyn (“line 61”) mice. Neurotherapeutics 9, 297–314. doi: 10.1007/s13311-012-0104-2
Chia, S. J., Tan, E. K., and Chao, Y. X. (2020). Historical perspective: models of Parkinson’s disease. Int. J. Mol. Sci. 21, 1–14. doi: 10.3390/ijms21072464
Choi, C.-B., Kim, S. Y., Lee, S. H., Jahng, G. H., Kim, H. Y., Choe, B. Y., et al. (2011). Assessment of metabolic changes in the striatum of a MPTP-intoxicated canine model: in vivo 1H-MRS study of an animal model for Parkinson’s disease. Magn. Reson. Imaging 29, 32–39. doi: 10.1016/j.mri.2010.03.043
Christensen, A. B., Sørensen, J. C. H., Ettrup, K. S., Orlowski, D., and Bjarkam, C. R. (2018). Pirouetting pigs: a large non-primate animal model based on unilateral 6-hydroxydopamine lesioning of the nigrostriatal pathway. Brain Res. Bull. 139, 167–173. doi: 10.1016/j.brainresbull.2018.02.010
Clarke, D. W., Mohtadi, C., and Tuffs, P. S. (1987). Generalized predictive control—part I. The basic algorithm. Automatica 23, 137–148. doi: 10.1016/0005-1098(87)90087-2
Connor, B. (2001). Annual scientific meeting of ASCEPT 2000: new investigator symposium adenoviral vector-mediated delivery of glial cell line-derived neurotrophic factor provides neuroprotection in the aged parkinsonian rat. Clin. Exp. Pharmacol. Physiol. 28, 896–900. doi: 10.1046/j.1440-1681.2001.03544.x
Cooper, J. F., and Van Raamsdonk, J. M. (2018). Modeling Parkinson’s disease in C. elegans. J. Parkinsons Dis. 8, 17–32. doi: 10.3233/JPD-171258
Coulom, H., and Birman, S. (2004). Chronic exposure to rotenone models sporadic Parkinson’s disease in Drosophila melanogaster. J. Neurosci. 24, 10993–10998. doi: 10.1523/JNEUROSCI.2993-04.2004
Cronin, A., and Grealy, M. (2017). Neuroprotective and neuro-restorative effects of minocycline and rasagiline in a zebrafish 6-hydroxydopamine model of Parkinson’s disease. Neuroscience 367, 34–46. doi: 10.1016/j.neuroscience.2017.10.018
Cui, X., Li, X., Zheng, H., Su, Y., Zhang, S., Li, M., et al. (2024). Human midbrain organoids: a powerful tool for advanced Parkinson’s disease modeling and therapy exploration. npj Parkinson’s Disease 10:189. doi: 10.1038/s41531-024-00799-8
Daniel, G., and Moore, D. J. (2015). Modeling LRRK2 pathobiology in Parkinson’s disease: from yeast to rodents. Curr. Top. Behav. Neurosci. 22, 331–368. doi: 10.1007/7854_2014_311
Dao, L., You, Z., Lu, L., Xu, T., Sarkar, A. K., Zhu, H., et al. (2024). Modeling blood-brain barrier formation and cerebral cavernous malformations in human PSC-derived organoids. Cell Stem Cell 31, 818–833.e11. doi: 10.1016/j.stem.2024.04.019
de Andrade Teles, R. B., Diniz, T. C., Costa Pinto, T. C., de Oliveira Júnior, R. G., Gama e Silva, M., de Lavor, É. M., et al. (2018). Flavonoids as therapeutic agents in Alzheimer’s and Parkinson’s diseases: a systematic review of preclinical evidences. Oxidative Med. Cell. Longev. 2018:7043213. doi: 10.1155/2018/7043213
Degirmenci, Y., Angelopoulou, E., Georgakopoulou, V. E., and Bougea, A. (2023). Cognitive impairment in Parkinson’s disease: an updated overview focusing on emerging pharmaceutical treatment approaches. Medicina 59. doi: 10.3390/medicina59101756
Del Rey, N. L.-G., Balzano, T., Martin-Rodriguez, L., Salinas-Rebolledo, C., Trigo-Damas, I., Rojas-Fernandez, A., et al. (2021). Lack of parkinsonian pathology and neurodegeneration in mice after long-term injections of a proteasome inhibitor in olfactory bulb and amygdala. Front. Aging Neurosci. 13:698979. doi: 10.3389/fnagi.2021.698979
Del Rey, N. L.-G., Del Rey, N. L., Quiroga-Varela, A., Garbayo, E., Carballo-Carbajal, I., Fernández-Santiago, R., et al. (2018). Advances in Parkinson’s disease: 200 years later. Front. Neuroanat. 12:113. doi: 10.3389/fnana.2018.00113
Del Rey, N. L., Hernández-Pinedo, N., Carrillo, M., Del Cerro, M., Esteban-García, N., Trigo-Damas, I., et al. (2024). Calbindin and Girk2/Aldh1a1 define resilient vs vulnerable dopaminergic neurons in a primate Parkinson’s disease model. NPJ Parkinsons Dis. 10:165. doi: 10.1038/s41531-024-00777-0
Del Rey, N. L., Quiroga-Varela, N., Hernández-Pinedo, N., Hernández-Pinedo, N., et al. (2018). Spread of aggregates after olfactory bulb injection of α-synuclein fibrils is associated with early neuronal loss and is reduced long term. Acta Neuropathol. 135, 65–83. doi: 10.1007/s00401-017-1792-9
Dovonou, A., Bolduc, C., Soto Linan, V., Gora, C., Peralta III, M. R., and Lévesque, M. (2023). Animal models of Parkinson’s disease: bridging the gap between disease hallmarks and research questions. Transl. Neurodegener. 12:36. doi: 10.1186/s40035-023-00368-8
Doyle, J. M., and Croll, R. P. (2022). A critical review of zebrafish models of Parkinson’s disease. Front. Pharmacol. 13:835827. doi: 10.3389/fphar.2022.835827
Drolet, R. E., Cannon, J. R., Montero, L., and Greenamyre, J. T. (2009). Chronic rotenone exposure reproduces Parkinson’s disease gastrointestinal neuropathology. Neurobiol. Dis. 36, 96–102. doi: 10.1016/j.nbd.2009.06.017
Dung, V. M., and Thao, D. T. P. (2018). “Parkinson’s disease model” in Drosophila models for human diseases. ed. M. Yamaguchi (Singapore: Springer (Advances in Experimental Medicine and Biology)), 41–61.
Dunham, M. J., and Fowler, D. M. (2013). Contemporary, yeast-based approaches to understanding human genetic variation. Curr. Opin. Genet. Dev. 23, 658–664. doi: 10.1016/j.gde.2013.10.001
Dwyer, Z., Rudyk, C., Situt, D., Beauchamp, S., Abdali, J., Dinesh, A., et al. (2020). Microglia depletion prior to lipopolysaccharide and paraquat treatment differentially modulates behavioral and neuronal outcomes in wild type and G2019S LRRK2 knock-in mice. Brain Behav. Immun. Health 5:100079. doi: 10.1016/j.bbih.2020.100079
Enterría-Morales, D., Del Rey, N. L. G., Blesa, J., López-López, I., Gallet, S., Prévot, V., et al. (2020). Molecular targets for endogenous glial cell line-derived neurotrophic factor modulation in striatal parvalbumin interneurons. Brain Commun. 2:fcaa105. doi: 10.1093/braincomms/fcaa105
Eslamboli, A., Romero-Ramos, M., Burger, C., Bjorklund, T., Muzyczka, N., Mandel, R. J., et al. (2007). Long-term consequences of human alpha-synuclein overexpression in the primate ventral midbrain. Brain 130, 799–815. doi: 10.1093/brain/awl382
Espadas-Alvarez, A. J., Bannon, M. J., Orozco-Barrios, C. E., Escobedo-Sanchez, L., Ayala-Davila, J., Reyes-Corona, D., et al. (2017). Regulation of human GDNF gene expression in nigral dopaminergic neurons using a new doxycycline-regulated NTS-polyplex nanoparticle system. Nanomedicine 13, 1363–1375. doi: 10.1016/j.nano.2017.02.006
Falkenburger, B. H., Saridaki, T., and Dinter, E. (2016). Cellular models for Parkinson’s disease. J. Neurochem. 139, 121–130. doi: 10.1111/JNC.13618
Farombi, E. O., Abolaji, A. O., Farombi, T. H., Oropo, A. S., Owoje, O. A., and Awunah, M. T. (2018). Garcinia kola seed biflavonoid fraction (Kolaviron), increases longevity and attenuates rotenone-induced toxicity in Drosophila melanogaster. Pestic. Biochem. Physiol. 145, 39–45. doi: 10.1016/j.pestbp.2018.01.002
Fatima, A., Jyoti, S., and Siddique, Y. H. (2018). Models of Parkinson’s disease with special emphasis on Drosophila melanogaster. CNS Neurol. Disord. Drug Targets 17, 757–766. doi: 10.2174/1871527317666180820164250
Fearnley, J. M., and Lees, A. J. (1991). Ageing and Parkinson’s disease: substantia nigra regional selectivity. Brain 114, 2283–2301. doi: 10.1093/brain/114.5.2283
Feng, C. W., Wen, Z. H., Huang, S. Y., Hung, H. C., Chen, C. H., Yang, S. N., et al. (2014). Egects of 6-hydroxydopamine exposure on motor activity and biochemical expression in zebrafish (Danio rerio) larvae. Zebrafish, 11, 227–239. doi: 10.1089/zeb.2013.0950
Fifel, K., and Cooper, H. M. (2014). Loss of dopamine disrupts circadian rhythms in a mouse model of Parkinson’s disease. Neurobiol. Dis. 71, 359–369. doi: 10.1016/j.nbd.2014.08.024
Fleming, S. M., Tetreault, N. A., Mulligan, C. K., Hutson, C. B., Masliah, E., and Chesselet, M. F. (2008). Olfactory deficits in mice overexpressing human wildtype alpha-synuclein. Eur. J. Neurosci. 28, 247–256. doi: 10.1111/j.1460-9568.2008.06346.x
Fodor, I., Urbán, P., Kemenes, G., Koene, J. M., and Pirger, Z. (2020). Aging and disease-relevant gene products in the neuronal transcriptome of the great pond snail (Lymnaea stagnalis): a potential model of aging, age-related memory loss, and neurodegenerative diseases. Invert. Neurosci. 20:9. doi: 10.1007/s10158-020-00242-6
Foltynie, T., and Aviles-Olmos, I. (2014). Exenatide as a potential treatment for patients with Parkinson’s disease: first steps into the clinic. Alzheimers Dement. 10. doi: 10.1016/j.jalz.2013.12.005
Fontana, B. D., Mezzomo, N. J., Kalueff, A. V., and Rosemberg, D. B. (2018). The developing utility of zebrafish models of neurological and neuropsychiatric disorders: a critical review. Exp. Neurol. 299, 157–171. doi: 10.1016/j.expneurol.2017.10.004
Fox, S. H., and Brotchie, J. M. (2010). “The MPTP-lesioned non-human primate models of Parkinson’s disease. Past, present, and future” in Progress in brain research. Amsterdam, The Netherlands: Elsevier., vol. 184, 133–157.
Frattini, E., Faustini, G., Lopez, G., Carsana, E. V., Tosi, M., Trezzi, I., et al. (2025). Lewy pathology formation in patient-derived GBA1 Parkinson’s disease midbrain organoids. Brain 148, 1242–1257. doi: 10.1093/brain/awae365
Gaeta, A. L., Caldwell, K. A., and Caldwell, G. A. (2019). Found in translation: the utility of C. elegans Alpha-Synuclein models of Parkinson’s disease. Brain Sci. 9. doi: 10.3390/brainsci9040073
Galasko, D., Farlow, M. R., Lucey, B. P., Honig, L. S., Elbert, D., Bateman, R., et al. (2024). A multicenter, randomized, double-blind, placebo-controlled ascending dose study to evaluate the safety, tolerability, pharmacokinetics (PK) and pharmacodynamic (PD) effects of Posiphen in subjects with early Alzheimer’s disease. Alzheimer's Res Ther 16:151. doi: 10.1186/s13195-024-01490-z
Garea-Rodríguez, E., Eesmaa, A., Lindholm, P., Schlumbohm, C., König, J., Meller, B., et al. (2016). Comparative analysis of the effects of neurotrophic factors CDNF and GDNF in a nonhuman primate model of Parkinson’s disease. PLoS One 11:e0149776. doi: 10.1371/journal.pone.0149776
Giasson, B. I., Duda, J. E., Quinn, S. M., Zhang, B., Trojanowski, J. Q., and Lee, V. M. Y. (2002). Neuronal alpha-synucleinopathy with severe movement disorder in mice expressing A53T human alpha-synuclein. Neuron 34, 521–533. doi: 10.1016/s0896-6273(02)00682-7
Goldsmith, J. R., and Jobin, C. (2012). Think small: zebrafish as a model system of human pathology. J. Biomed. Biotechnol. 2012:817341. doi: 10.1155/2012/817341
Gómez-Benito, M., Granado, N., García-Sanz, P., Michel, A., Dumoulin, M., and Moratalla, R. (2020). Modeling Parkinson’s disease with the alpha-synuclein protein. Front. Pharmacol. 11:356. doi: 10.3389/fphar.2020.00356
Goya, M. E., Xue, F., Sampedro-Torres-Quevedo, C., Arnaouteli, S., Riquelme-Dominguez, L., Romanowski, A., et al. (2020). Probiotic Bacillus subtilis protects against α-Synuclein aggregation in C. elegans. Cell Rep. 30, 367–380.e7. doi: 10.1016/j.celrep.2019.12.078
Greffard, S., Verny, M., Bonnet, A. M., Seilhean, D., Hauw, J. J., and Duyckaerts, C. (2010). A stable proportion of Lewy body bearing neurons in the substantia nigra suggests a model in which the Lewy body causes neuronal death. Neurobiol. Aging 31, 99–103. doi: 10.1016/j.neurobiolaging.2008.03.015
Griffioen, G., Duhamel, H., van Damme, N., Pellens, K., Zabrocki, P., Pannecouque, C., et al. (2006). A yeast-based model of α-synucleinopathy identifies compounds with therapeutic potential. Biochim. Biophys. Acta Mol. basis Dis. 1762, 312–318. doi: 10.1016/j.bbadis.2005.11.009
Grosset, K. A., Malek, N., Morgan, F., and Grosset, D. G. (2013). Inhaled dry powder apomorphine (VR040) for ‘off’ periods in Parkinson’s disease: an in-clinic double-blind dose ranging study. Acta Neurol. Scand. 128, 166–171. doi: 10.1111/ane.12107
Halliday, G., Herrero, M. T., Murphy, K., McCann, H., Ros-Bernal, F., Barcia, C., et al. (2009). No Lewy pathology in monkeys with over 10 years of severe MPTP parkinsonism. Mov. Disord. 24, 1519–1523. doi: 10.1002/mds.22481
Hamadjida, A., Nuara, S. G., Kwan, C., Frouni, I., Bédard, D., Gourdon, J. C., et al. (2020). Monoamine oxidase a inhibition with moclobemide enhances the anti-parkinsonian effect of L-DOPA in the MPTP-lesioned marmoset. Naunyn Schmiedeberg's Arch. Pharmacol. 393, 2157–2164. doi: 10.1007/S00210-020-01933-Y
Heinzel, S., Berg, D., Gasser, T., Chen, H., Yao, C., Postuma, R. B., et al. (2019). Update of the MDS research criteria for prodromal Parkinson’s disease. Mov. Disord. 34, 1464–1470. doi: 10.1002/mds.27802
Heiss, J. D., Lungu, C., Hammoud, D. A., Herscovitch, P., Ehrlich, D. J., Argersinger, D. P., et al. (2019). Trial of magnetic resonance–guided putaminal gene therapy for advanced Parkinson’s disease. Mov. Disord. 34, 1073–1078. doi: 10.1002/mds.27724
Hernandez-Baltazar, D., Zavala-Flores, L. M., and Villanueva-Olivo, A. (2017). El modelo de 6-hidroxidopamina y la fisiopatología parkinsoniana: nuevos hallazgos en un viejo modelo. Neurologia 32, 533–539. doi: 10.1016/j.nrl.2015.06.011
He, S., Ru, Q., Chen, L., Xu, G., and Wu, Y. (2024). Advances in animal models of Parkinson’s disease. Brain Res. Bull. 215:111024. doi: 10.1016/j.brainresbull.2024.111024
Hirsh, J., Riemensperger, T., Coulom, H., Iché, M., Coupar, J., and Birman, S. (2010). Roles of dopamine in circadian rhythmicity and extreme light sensitivity of circadian entrainment. Curr. Biol. 20, 209–214. doi: 10.1016/j.cub.2009.11.037
Hsieh, M.-H., Meng, W. Y., Liao, W. C., Weng, J. C., Li, H. H., Su, H. L., et al. (2017). Ceftriaxone reverses deficits of behavior and neurogenesis in an MPTP-induced rat model of Parkinson’s disease dementia. Brain Res. Bull. 132, 129–138. doi: 10.1016/j.brainresbull.2017.05.015
Huang, C.-K., Chang, Y. T., Amstislavskaya, T. G., Tikhonova, M. A., Lin, C. L., Hung, C. S., et al. (2015). Synergistic effects of ceftriaxone and erythropoietin on neuronal and behavioral deficits in an MPTP-induced animal model of Parkinson’s disease dementia. Behav. Brain Res. 294, 198–207. doi: 10.1016/j.bbr.2015.08.011
Isaacson, S. H., Fahn, S., Pahwa, R., Tanner, C. M., Espay, A. J., Trenkwalder, C., et al. (2018). Parkinson’s patients with dyskinesia switched from immediate release amantadine to open-label ADS-5102. Mov. Disord. Clin. Pract. 5, 183–190. doi: 10.1002/mdc3.12595
Izco, M., Blesa, J., Verona, G., Cooper, J. M., and Alvarez-Erviti, L. (2021). Glial activation precedes alpha-synuclein pathology in a mouse model of Parkinson’s disease. Neurosci. Res. 170, 330–340. doi: 10.1016/j.neures.2020.11.004
Jennings, D., Huntwork-Rodriguez, S., Henry, A. G., Sasaki, J. C., Meisner, R., Diaz, D., et al. (2022). Preclinical and clinical evaluation of the LRRK2 inhibitor DNL201 for Parkinson’s disease. Sci. Transl. Med. 14:eabj2658. doi: 10.1126/scitranslmed.abj2658
Jennings, D., Huntwork-Rodriguez, S., Vissers, M. F. J. M., Daryani, V. M., Diaz, D., Goo, M. S., et al. (2023). LRRK2 inhibition by BIIB122 in healthy participants and patients with Parkinson’s disease. Mov. Disord. 38, 386–398. doi: 10.1002/mds.29297
Jeong, J.-W., Yu, C., Lee, J. H., Moon, K. S., Kim, E., Yoo, S. E., et al. (2016). Subacute toxicity evaluation of KR-33493, FAF1 inhibitor for a new anti-parkinson’s disease agent, after oral administration in rats and dogs. Regul. Toxicol. Pharmacol. 81, 387–396. doi: 10.1016/j.yrtph.2016.09.022
Jo, J., Yang, L., Tran, H. D., Yu, W., Sun, A. X., Chang, Y. Y., et al. (2021). Lewy body–like inclusions in human midbrain organoids carrying Glucocerebrosidase and α-Synuclein mutations. Ann. Neurol. 90, 490–505. doi: 10.1002/ANA.26166
Julienne, H., Buhl, E., Leslie, D. S., and Hodge, J. J. L. (2017). Drosophila PINK1 and parkin loss-of-function mutants display a range of non-motor Parkinson’s disease phenotypes. Neurobiol. Dis. 104, 15–23. doi: 10.1016/j.nbd.2017.04.014
Katzenschlager, R., Poewe, W., Rascol, O., Trenkwalder, C., Deuschl, G., Chaudhuri, K. R., et al. (2018). Apomorphine subcutaneous infusion in patients with Parkinson’s disease with persistent motor fluctuations (TOLEDO): a multicentre, double-blind, randomised, placebo-controlled trial. Lancet Neurol. 17, 749–759. doi: 10.1016/S1474-4422(18)30239-4
Kim, C., Ojo-Amaize, E., Spencer, B., Rockenstein, E., Mante, M., Desplats, P., et al. (2015). Hypoestoxide reduces neuroinflammation and α-synuclein accumulation in a mouse model of Parkinson’s disease. J. Neuroinflammation 12:236. doi: 10.1186/s12974-015-0455-9
Kin, K., Yasuhara, T., Kameda, M., and Date, I. (2019). Animal models for Parkinson’s disease research: trends in the 2000s. Int. J. Mol. Sci. 20. doi: 10.3390/ijms20215402
Kirik, D., Annett, L. E., Burger, C., Muzyczka, N., Mandel, R. J., Björklund, A., et al. (2003). Nigrostriatal alpha-synucleinopathy induced by viral vector-mediated overexpression of human alpha-synuclein: a new primate model of Parkinson’s disease. Proc. Natl. Acad. Sci. USA 100, 2884–2889. doi: 10.1073/pnas.0536383100
Kirik, D., Rosenblad, C., and Bjorklund, A. (2000). Preservation of a functional nigrostriatal dopamine pathway by GDNF in the intrastriatal 6-OHDA lesion model depends on the site of administration of the trophic factor. Eur. J. Neurosci. 12, 3871–3882. doi: 10.1046/j.1460-9568.2000.00274.x
Kirkeby, A., Parmar, M., and Barker, R. A. (2017). Strategies for bringing stem cell-derived dopamine neurons to the clinic: a European approach (STEM-PD). Prog. Brain Res. 230, 165–190. doi: 10.1016/bs.pbr.2016.11.011
Koprich, J. B., Johnston, T. H., Reyes, G., Omana, V., and Brotchie, J. M. (2016). Towards a non-human primate model of alpha-Synucleinopathy for development of therapeutics for Parkinson’s disease: optimization of AAV1/2 delivery parameters to drive sustained expression of alpha Synuclein and dopaminergic degeneration in macaque. PLoS One 11, e0167235–e0167213. doi: 10.1371/journal.pone.0167235
Kordower, J. H., and Burke, R. E. (2018). Disease modification for Parkinson’s disease: axonal regeneration and trophic factors. Mov. Disord. 33, 678–683. doi: 10.1002/mds.27383
Kordower, J. H., Emborg, M. E., Bloch, J., Ma, S. Y., Chu, Y., Leventhal, L., et al. (2000). Neurodegeneration prevented by lentiviral vector delivery of GDNF in primate models of Parkinson's disease. science 290, 767–773. doi: 10.1126/science.290.5492.767
Kritzer, J. A., Hamamichi, S., McCaffery, J. M., Santagata, S., Naumann, T. A., Caldwell, K. A., et al. (2009). Rapid selection of cyclic peptides that reduce α-Synuclein toxicity in yeast and animal models HHS public access author manuscript. Nat. Chem. Biol. 5, 655–663. doi: 10.1038/nchembio.193
Kuan, W.-L., Stott, K., He, X., Wood, T. C., Yang, S., Kwok, J. C. F., et al. (2021). Systemic α-synuclein injection triggers selective neuronal pathology as seen in patients with Parkinson’s disease. Mol. Psychiatry 26, 556–567. doi: 10.1038/s41380-019-0608-9
Kuo, Y.-M., Li, Z., Jiao, Y., Gaborit, N., Pani, A. K., Orrison, B. M., et al. (2010). Extensive enteric nervous system abnormalities in mice transgenic for artificial chromosomes containing Parkinson disease-associated α-synuclein gene mutations precede central nervous system changes. Hum. Mol. Genet. 19, 1633–1650. doi: 10.1093/hmg/ddq038
Kuo, Y.-M., Nwankwo, N., and Rogers, M. (2019). Translational inhibition of α-synuclein by Posiphen normalizes distal colon motility in transgenic Parkinson mice. Am. J. Neurodegener. Dis. 8, 1–15.
Lafuente, J. V., Requejo, C., and Ugedo, L. (2019) Chapter 8 - Nanodelivery of therapeutic agents in Parkinson’s disease, in A. Sharma and H. S. Sharma (eds.) Nanoneuroprotection and Nanoneurotoxicology. Amsterdam, The Netherlands: Elsevier (Progress in Brain Research), pp. 263–279.
Laloux, C., Derambure, P., Kreisler, A., Houdayer, E., Bruezière, S., Bordet, R., et al. (2008). MPTP-treated mice: long-lasting loss of nigral TH-ir neurons but not paradoxical sleep alterations. Exp. Brain Res. 186, 635–642. doi: 10.1007/s00221-008-1268-1
Langley, M. R., Ghaisas, S., Palanisamy, B. N., Ay, M., Jin, H., Anantharam, V., et al. (2021). Characterization of nonmotor behavioral impairments and their neurochemical mechanisms in the MitoPark mouse model of progressive neurodegeneration in Parkinson’s disease. Exp. Neurol. 341:113716. doi: 10.1016/j.expneurol.2021.113716
Lawal, H. O., Terrell, A., Lam, H. A., Djapri, C., Jang, J., Hadi, R., et al. (2014). Drosophila modifier screens to identify novel neuropsychiatric drugs including aminergic agents for the possible treatment of Parkinson’s disease and depression. Mol. Psychiatry 19, 235–242. doi: 10.1038/mp.2012.170
Lázaro, D. F., Pavlou, M. A. S., and Outeiro, T. F. (2017). Cellular models as tools for the study of the role of alpha-synuclein in Parkinson’s disease. Exp. Neurol. 298, 162–171. doi: 10.1016/j.expneurol.2017.05.007
Lee, B. D., Shin, J. H., VanKampen, J., Petrucelli, L., West, A. B., Ko, H. S., et al. (2010). Inhibitors of leucine-rich repeat kinase-2 protect against models of Parkinson’s disease. Nat. Med. 16, 998–1000. doi: 10.1038/nm.2199
Lee, S.-H., Jahng, G. H., Choe, I. H., Choi, C. B., Kim, D. H., and Kim, H. Y. (2013). Neural pathway interference by retained acupuncture: a functional MRI study of a dog model of Parkinson’s disease. CNS Neurosci. Ther. 19, 585–595. doi: 10.1111/cns.12108
Lewitt, P. A., and Fahn, S. (2016). Levodopa therapy for Parkinson disease: a look backward and forward. Neurology 86, S3–S12. doi: 10.1212/WNL.0000000000002509
Lewitt, P. A., Hauser, R. A., Grosset, D. G., Stocchi, F., Saint-Hilaire, M.-H., Ellenbogen, A., et al. (2016). A randomized trial of inhaled levodopa (CVT-301) for motor fluctuations in Parkinson’s disease. Mov. Disord. 31. doi: 10.1002/mds.26611
Lillethorup, T. P., Glud, A. N., Alstrup, A. K. O., Mikkelsen, T. W., Nielsen, E. H., Zaer, H., et al. (2018). Nigrostriatal proteasome inhibition impairs dopamine neurotransmission and motor function in minipigs. Exp. Neurol. 303, 142–152. doi: 10.1016/j.expneurol.2018.02.005
Lindholm, P., and Saarma, M. (2022). Cerebral dopamine neurotrophic factor protects and repairs dopamine neurons by novel mechanism. Mol. Psychiatry 27, 1310–1321. doi: 10.1038/s41380-021-01394-6
Lipp, M. M., Batycky, R., Moore, J., Leinonen, M., and Freed, M. I. (2016). Preclinical and clinical assessment of inhaled levodopa for OFF episodes in Parkinson’s disease. Sci. Transl. Med. 8:360ra136. doi: 10.1126/scitranslmed.aad8858
Li, X., Patel, J. C., Wang, J., Avshalumov, M. V., Nicholson, C., Buxbaum, J. D., et al. (2010). Enhanced striatal dopamine transmission and motor performance with LRRK2 overexpression in mice is eliminated by familial Parkinson’s disease mutation G2019S. J. Neurosci. 30, 1788–1797. doi: 10.1523/JNEUROSCI.5604-09.2010
Li, X., Redus, L., Chen, C., Martinez, P. A., Strong, R., Li, S., et al. (2013). Cognitive dysfunction precedes the onset of motor symptoms in the MitoPark mouse model of Parkinson’s disease. PLoS One 8, e71341–e71311. doi: 10.1371/journal.pone.0071341
Li, Y., Perry, T., Kindy, M. S., Harvey, B. K., Tweedie, D., Holloway, H. W., et al. (2009). GLP-1 receptor stimulation preserves primary cortical and dopaminergic neurons in cellular and rodent models of stroke and parkinsonism. Proc. Natl. Acad. Sci. USA 106, 1285–1290. doi: 10.1073/pnas.0806720106
Lopert, P., and Patel, M. (2014). Brain mitochondria from DJ-1 knockout mice show increased respiration-dependent hydrogen peroxide consumption. Redox Biol. 2, 667–672. doi: 10.1016/j.redox.2014.04.010
Maasz, G., Zrinyi, Z., Reglodi, D., Petrovics, D., Rivnyak, A., Kiss, T., et al. (2017). Pituitary adenylate cyclase-activating polypeptide (PACAP) has a neuroprotective function in dopamine-based neurodegeneration in rat and snail parkinsonian models. Dis. Model. Mech. 10, 127–139. doi: 10.1242/dmm.027185
Malkus, K. A., Tsika, E., and Ischiropoulos, H. (2009). Oxidative modifications, mitochondrial dysfunction, and impaired protein degradation in Parkinson’s disease: how neurons are lost in the Bermuda triangle. Mol. Neurodegener. 4:24. doi: 10.1186/1750-1326-4-24
Manfredsson, F. P., Luk, K. C., Benskey, M. J., Gezer, A., Garcia, J., Kuhn, N. C., et al. (2018). Induction of alpha-synuclein pathology in the enteric nervous system of the rat and non-human primate results in gastrointestinal dysmotility and transient CNS pathology. Neurobiol. Dis. 112, 106–118. doi: 10.1016/j.nbd.2018.01.008
Marmion, D. J., and Kordower, J. H. (2018). α-Synuclein nonhuman primate models of Parkinson’s disease. J. Neural Transm. 125, 385–400. doi: 10.1007/s00702-017-1720-0
Marsh, J. L., and Thompson, L. M. (2006). Drosophila in the study of neurodegenerative disease. Neuron 52, 169–178. doi: 10.1016/j.neuron.2006.09.025
McFarthing, K., Buff, S., Rafaloff, G., Pitzer, K., Fiske, B., Navangul, A., et al. (2024). Parkinson’s disease drug therapies in the clinical trial pipeline: 2024 update. J. Parkinsons Dis. 14, 899–912. doi: 10.3233/JPD-240272
McFarthing, K., Rafaloff, G., Baptista, M. A. S., Wyse, R. K., and Stott, S. R. W. (2021). Parkinson’s disease drug therapies in the clinical trial pipeline: 2021 update. J. Parkinsons Dis. 11, 891–903. doi: 10.3233/JPD-219006
McFarthing, K., Rafaloff, G., Baptista, M., Mursaleen, L., Fuest, R., Wyse, R. K., et al. (2022). Parkinson’s disease drug therapies in the clinical trial pipeline: 2022 update. J. Parkinsons Dis. 12, 1073–1082. doi: 10.3233/JPD-229002
Medeiros, D. d. C., Lopes Aguiar, C., Moraes, M. F. D., and Fisone, G. (2019). Sleep disorders in rodent models of Parkinson’s disease. Front. Pharmacol. 10:1414. doi: 10.3389/fphar.2019.01414
Mehmood, A., Ali, W., Din, Z. U., Song, S., Sohail, M., Shah, W., et al. (2021). Clustered regularly interspaced short palindromic repeats as an advanced treatment for Parkinson’s disease. Brain Behav. 11:e2280. doi: 10.1002/brb3.2280
Migdalska-Richards, A., Daly, L., Bezard, E., and Schapira, A. H. V. (2016). Ambroxol effects in glucocerebrosidase and α-synuclein transgenic mice. Ann. Neurol. 80, 766–775. doi: 10.1002/ana.24790
Mikkelsen, M., MØller, A., Jensen, L. H., Pedersen, A., Harajehi, J. B., and Pakkenberg, H. (1999). MPTP-induced parkinsonism in Minipigs: a behavioral, biochemical, and histological study. Neurotoxicol. Teratol. 21, 169–175. doi: 10.1016/S0892-0362(98)00037-3
Milanese, C., Sager, J. J., Bai, Q., Farrell, T. C., Cannon, J. R., Greenamyre, J. T., et al. (2012). Hypokinesia and reduced dopamine levels in zebrafish lacking β- and γ1-synucleins. J. Biol. Chem. 287, 2971–2983. doi: 10.1074/jbc.M111.308312
Molinet-Dronda, F., Blesa, J., Del Rey, N. L. G., Juri, C., Collantes, M., Pineda-Pardo, J. A., et al. (2022). Cerebral metabolic pattern associated with progressive parkinsonism in non-human primates reveals early cortical hypometabolism. Neurobiol. Dis. 167:105669. doi: 10.1016/j.nbd.2022.105669
Moriarty, N., Gantner, C. W., Hunt, C. P. J., Ermine, C. M., Frausin, S., Viventi, S., et al. (2022). A combined cell and gene therapy approach for homotopic reconstruction of midbrain dopamine pathways using human pluripotent stem cells. Cell Stem Cell 29, 434–448.e5. doi: 10.1016/j.stem.2022.01.013
Mu, X., Yuan, X., du, L. D., He, G. R., and du, G. H. (2016). Antagonism of quercetin against tremor induced by unilateral striatal lesion of 6-OHDA in rats. J. Asian Nat. Prod. Res. 18, 65–71. doi: 10.1080/10286020.2015.1057576
Nicoletti, F., Philippens, I., Fagone, P., Ahlem, C. N., Reading, C. L., Frincke, J. M., et al. (2012). 17α-Ethynyl-androst-5-ene-3β,7β,17β-triol (HE3286) is neuroprotective and reduces motor impairment and Neuroinflammation in a murine MPTP model of Parkinson’s disease. Parkinson’s Disease 2012:969418. doi: 10.1155/2012/969418
Nimmo, J. T., Smith, H., Wang, C. Y., Teeling, J. L., Nicoll, J. A. R., Verma, A., et al. (2022). Immunisation with UB-312 in the Thy1SNCA mouse prevents motor performance deficits and oligomeric α-synuclein accumulation in the brain and gut. Acta Neuropathol. 143, 55–73. doi: 10.1007/s00401-021-02381-5
Niu, Y., Guo, X., Chen, Y., Wang, C. E., Gao, J., Yang, W., et al. (2015). Early Parkinson’s disease symptoms in α-synuclein transgenic monkeys. Hum. Mol. Genet. 24, 2308–2317. doi: 10.1093/hmg/ddu748
Nordström, E., Eriksson, F., Sigvardson, J., Johannesson, M., Kasrayan, A., Jones-Kostalla, M., et al. (2021). ABBV-0805, a novel antibody selective for soluble aggregated α-synuclein, prolongs lifespan and prevents buildup of α-synuclein pathology in mouse models of Parkinson’s disease. Neurobiol. Dis. 161:105543. doi: 10.1016/j.nbd.2021.105543
O’Toole, T. E., Amraotkar, A. R., Gao, H., Sears, C. G., Rai, S. N., Basner, M., et al. (2025). Carnosine supplementation improves cognitive outcomes in younger participants of the NEAT trial. Neurotherapeutics 22. doi: 10.1016/j.neurot.2025.e00541
Oiwa, Y., Nakai, K., and Itakura, T. (2006). Histological effects of intraputaminal infusion of glial cell line-derived neurotrophic factor in Parkinson disease model macaque monkeys. Neurol. Med. Chir. 46, 267–276. doi: 10.2176/nmc.46.267
Pagano, G., Boess, F. G., Taylor, K. I., Ricci, B., Mollenhauer, B., Poewe, W., et al. (2021). A phase II study to evaluate the safety and efficacy of Prasinezumab in early Parkinson’s disease (PASADENA): rationale, design, and baseline data. Front. Neurol. 12:705407. doi: 10.3389/fneur.2021.705407
Pagano, G., Monnet, A., Reyes, A., Ribba, B., Svoboda, H., Kustermann, T., et al. (2024). Sustained effect of prasinezumab on Parkinson’s disease motor progression in the open-label extension of the PASADENA trial. Nat. Med. 30, 3669–3675. doi: 10.1038/s41591-024-03270-6
Palmas, M. F., Ena, A., Burgaletto, C., Casu, M. A., Cantarella, G., Carboni, E., et al. (2022). Repurposing Pomalidomide as a neuroprotective drug: efficacy in an alpha-Synuclein-based model of Parkinson’s disease. Neurotherapeutics 19, 305–324. doi: 10.1007/s13311-022-01182-2
Parisky, K. M., Agosto, J., Pulver, S. R., Shang, Y., Kuklin, E., Hodge, J. J. L., et al. (2008). PDF cells are a GABA-responsive wake-promoting component of the Drosophila sleep circuit. Neuron 60, 672–682. doi: 10.1016/j.neuron.2008.10.042
Pessiglione, M., Petrovic, P., Daunizeau, J., Palminteri, S., Dolan, R. J., and Frith, C. D. (2008). Subliminal instrumental conditioning demonstrated in the human brain. Neuron 59, 561–567. doi: 10.1016/j.neuron.2008.07.005
Petit, G. H., Berkovich, E., Hickery, M., Kallunki, P., Fog, K., Fitzer-Attas, C., et al. (2013). Rasagiline ameliorates olfactory deficits in an alpha-Synuclein mouse model of Parkinson’s disease. PLoS One 8, 1–13. doi: 10.1371/journal.pone.0060691
Podell, M., Hadjiconstantinou, M., Smith, M. A., and Neff, N. H. (2003). Proton magnetic resonance imaging and spectroscopy identify metabolic changes in the striatum in the MPTP feline model of parkinsonism. Exp. Neurol. 179, 159–166. doi: 10.1016/s0014-4886(02)00015-8
Porras, C., Hildesheim, A., González, P., Schiffman, M., Rodríguez, A. C., Wacholder, S., et al. (2015). Performance of self-collected cervical samples in screening for future precancer using human papillomavirus DNA testing. J. Natl. Cancer Inst. 107:400. doi: 10.1093/jnci/dju400
Postuma, R. B., and Berg, D. (2016). Advances in markers of prodromal Parkinson disease. Nat. Rev. Neurol. 12, 622–634. doi: 10.1038/nrneurol.2016.152
Postuma, R. B., Berg, D., Stern, M., Poewe, W., Olanow, C. W., Oertel, W., et al. (2015). MDS clinical diagnostic criteria for Parkinson’s disease. Mov. Disord. 30, 1591–1601. doi: 10.1002/mds.26424
Prediger, R. D. S., Rial, D., Medeiros, R., Figueiredo, C. P., Doty, R. L., and Takahashi, R. N. (2009). Risk is in the air: an intranasal MPTP (1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine) rat model of Parkinson’s disease. Ann. N. Y. Acad. Sci. 1170, 629–636. doi: 10.1111/j.1749-6632.2009.03885.x
Price, D. L., Koike, M. A., Khan, A., Wrasidlo, W., Rockenstein, E., Masliah, E., et al. (2018). The small molecule alpha-synuclein misfolding inhibitor, NPT200-11, produces multiple benefits in an animal model of Parkinson’s disease. Sci. Rep. 8:16165. doi: 10.1038/s41598-018-34490-9
Ramot, Y., Nyska, A., Adar, L., Durlach, C., Fishelovitch, D., Sacco, G., et al. (2018). ND0701, a novel formulation of apomorphine for subcutaneous infusion, in comparison to a commercial apomorphine formulation: 28-day pharmacokinetic study in minipigs and a phase I study in healthy volunteers to assess the safety, tolerability, pharmacokinetics and relative bioavailability. CNS Drugs, 32, 443–454. doi: 10.1007/s40263-018-0512-x
Ramot, Y., Nyska, A., Maronpot, R. R., Shaltiel-Karyo, R., Tsarfati, Y., Manno, R. A., et al. (2017). Ninety-day local tolerability and toxicity study of ND0612, a novel formulation of levodopa/carbidopa, administered by subcutaneous continuous infusion in Minipigs. Toxicol. Pathol. 45, 764–773. doi: 10.1177/0192623317729891
Rautio, J., Kumpulainen, H., Heimbach, T., Oliyai, R., Oh, D., Järvinen, T., et al. (2008). Prodrugs: design and clinical applications. Nat. Rev. Drug Discov. 7, 255–270. doi: 10.1038/nrd2468
Razali, K., Othman, N., Mohd Nasir, M. H., Doolaanea, A. A., Kumar, J., Ibrahim, W. N., et al. (2021). The promise of the zebrafish model for Parkinson’s disease: today’s science and tomorrow’s treatment. Front. Genet. 12:655550. doi: 10.3389/fgene.2021.655550
Reichmann, H., Schneider, C., and Löhle, M. (2009). Non-motor features of Parkinson’s disease: depression and dementia. Parkinsonism Relat. Disord. 15, S87–S92. doi: 10.1016/S1353-8020(09)70789-8
Richter, F., Stanojlovic, M., Käufer, C., Gericke, B., and Feja, M. (2023). A mouse model to test novel therapeutics for Parkinson’s disease: an update on the Thy1-aSyn (“line 61”) mice. Neurotherapeutics 20, 97–116. doi: 10.1007/s13311-022-01338-0
Rivi, V., Caruso, G., Caraci, F., Alboni, S., Pani, L., Tascedda, F., et al. (2024). Behavioral and transcriptional effects of carnosine in the central ring ganglia of the pond snail Lymnaea stagnalis. J. Neurosci. Res. 102:e25371. doi: 10.1002/jnr.25371
Rota, L., Pellegrini, G., Benvenuti, L., Antonioli, L., Fornai, M., Blandizzi, C., et al. (2019). Constipation, deficit in colon contractions and alpha-synuclein inclusions within the colon precede motor abnormalities and neurodegeneration in the central nervous system in a mouse model of alpha-synucleinopathy. Transl. Neurodegener. 8:5. doi: 10.1186/s40035-019-0146-z
Rudyk, C., and Hayley, S. (2019). Leucine-rich repeat kinase-2 (LRRK2) modulates paraquat-induced inflammatory sickness and stress phenotype. J. Neuroinflammation 16:120. doi: 10.1186/s12974-019-1483-7
Sandhof, C. A., Hoppe, S. O., Tittelmeier, J., and Nussbaum-Krammer, C. (2020). C. elegans models to study the propagation of prions and prion-like proteins. Biomolecules 10:1188. doi: 10.3390/biom10081188
Sarath Babu, N., Murthy, C. L. N., Kakara, S., Sharma, R., Brahmendra Swamy, C. V. y., and Idris, M. M. (2016). 1-Methyl-4-phenyl-1,2,3,6-tetrahydropyridine induced Parkinson’s disease in zebrafish. Proteomics, 16, 1407–1420. doi: 10.1002/pmic.201500291
Schapira, A. H. V., Chaudhuri, K. R., and Jenner, P. (2017). Non-motor features of Parkinson disease. Nat. Rev. Neurosci. 18, 435–450. doi: 10.1038/nrn.2017.62
Schintu, N., Frau, L., Ibba, M., Garau, A., Carboni, E., and Carta, A. R. (2009). Progressive dopaminergic degeneration in the chronic MPTPp mouse model of Parkinson’s disease. Neurotox. Res. 16, 127–139. doi: 10.1007/s12640-009-9061-x
Schneider, J. S., and Kovelowski, C. J. (1990). Chronic exposure to low doses of MPTP: I. Cognitive deficits in motor asymptomatic monkeys. Brain Res. 519, 122–128. doi: 10.1016/0006-8993(90)90069-N
Schweitzer, J. S., Song, B., Herrington, T. M., Park, T. Y., Lee, N., Ko, S., et al. (2020). Personalized iPSC-derived dopamine progenitor cells for Parkinson’s disease. N. Engl. J. Med. 382, 1926–1932. doi: 10.1056/NEJMoa1915872
Seegobin, S. P., Heaton, G. R., Liang, D., Choi, I., Blanca Ramirez, M., Tang, B., et al. (2020). Progress in LRRK2-associated Parkinson’s disease animal models. Front. Neurosci. 14:674. doi: 10.3389/fnins.2020.00674
Segura-Aguilar, J. (2017). Aminochrome as preclinical model for Parkinson’s disease. Oncotarget 8, 45036–45037. doi: 10.18632/oncotarget.18353
Selcho, M., Pauls, D., Han, K. A., Stocker, R. F., and Thum, A. S. (2009). The role of dopamine in Drosophila larval classical olfactory conditioning. PLoS One 4:e5897. doi: 10.1371/journal.pone.0005897
Selvaraj, V., Venkatasubramanian, H., Ilango, K., and Santhakumar, K. (2019). A simple method to study motor and non-motor behaviors in adult zebrafish. J. Neurosci. Methods 320, 16–25. doi: 10.1016/j.jneumeth.2019.03.008
Seo, J., Lee, Y., Kim, B. S., Park, J., Yang, S., Yoon, H. J., et al. (2019). A non-human primate model for stable chronic Parkinson’s disease induced by MPTP administration based on individual behavioral quantification. J. Neurosci. Methods 311, 277–287. doi: 10.1016/j.jneumeth.2018.10.037
Shadrina, M., and Slominsky, P. (2021). Modeling Parkinson’s disease: not only rodents? Front. Aging Neurosci. 13:695718. doi: 10.3389/fnagi.2021.695718
Shahmoradian, S. H., Lewis, A. J., Genoud, C., Hench, J., Moors, T. E., Navarro, P. P., et al. (2019). Lewy pathology in Parkinson’s disease consists of crowded organelles and lipid membranes. Nat. Neurosci. 22, 1099–1109. doi: 10.1038/s41593-019-0423-2
Shang, Y., Haynes, P., Pírez, N., Harrington, K. I., Guo, F., Pollack, J., et al. (2011). Imaging analysis of clock neurons reveals light buffers the wake-promoting effect of dopamine. Nat. Neurosci. 14, 889–895. doi: 10.1038/nn.2860
Shaye, D. D., and Greenwald, I. (2011). OrthoList: a compendium of C. elegans genes with human orthologs. PLoS One 6:e20085. doi: 10.1371/journal.pone.0020085
Shin, W. H., Park, J. H., and Chung, K. C. (2020). The central regulator p62 between ubiquitin proteasome system and autophagy and its role in the mitophagy and Parkinson’s disease. BMB Rep. 53, 56–63. doi: 10.5483/BMBRep.2020.53.1.283
Siddique, Y. H., Naz, F., and Jyoti, S. (2014). Effect of curcumin on lifespan, activity pattern, oxidative stress, and apoptosis in the brains of transgenic Drosophila model of Parkinson’s disease. Biomed. Res. Int. 2014:606928. doi: 10.1155/2014/606928
Slovin, H., Abeles, M., Vaadia, E., Haalman, I., Prut, Y., and Bergman, H. (1999). Frontal cognitive impairments and saccadic deficits in low-dose MPTP-treated monkeys. J. Neurophysiol. 81, 858–874. doi: 10.1152/jn.1999.81.2.858
Smith, M. P., and Cass, W. A. (2007). Gdnf reduces oxidative stress in a 6-hydroxydopamine model of Parkinson’s disease. Neurosci. Lett. 412, 259–263. doi: 10.1016/j.neulet.2006.11.017
Smits, L. M., Reinhardt, L., Reinhardt, P., Glatza, M., Monzel, A. S., Stanslowsky, N., et al. (2019). Modeling Parkinson’s disease in midbrain-like organoids. npj Parkinson’s Disease 5:5. doi: 10.1038/s41531-019-0078-4
Smits, L. M., and Schwamborn, J. C. (2020). Midbrain organoids: a new tool to investigate Parkinson’s disease. Front. Cell Dev. Biol. 8:359. doi: 10.3389/fcell.2020.00359
Sohrabi, S., Mor, D. E., Kaletsky, R., Keyes, W., and Murphy, C. T. (2021). High-throughput behavioral screen in C. elegans reveals Parkinson’s disease drug candidates. Commun. Biol. 4:203. doi: 10.1038/s42003-021-01731-z
Soldner, F., Stelzer, Y., Shivalila, C. S., Abraham, B. J., Latourelle, J. C., Barrasa, M. I., et al. (2016). Parkinson-associated risk variant in enhancer element produces subtle effect on target gene expression HHS public access. Nature 533, 95–99. doi: 10.1038/nature17939
Stevens, D. A., Lee, Y., Kang, H. C., Lee, B. D., Lee, Y. I., Bower, A., et al. (2015). Parkin loss leads to PARIS-dependent declines in mitochondrial mass and respiration. Proc. Natl. Acad. Sci. 112, 11696–11701. doi: 10.1073/pnas.1500624112
Stewart, A. M., Braubach, O., Spitsbergen, J., Gerlai, R., and Kalueff, A. V. (2014). Zebrafish models for translational neuroscience research: from tank to bedside. Trends Neurosci. 37, 264–278. doi: 10.1016/j.tins.2014.02.011
Surguchov, A. (2021). Invertebrate models untangle the mechanism of neurodegeneration in Parkinson’s disease. Cells 10:407. doi: 10.3390/cells10020407
Tabar, V., Sarva, H., Lozano, A. M., Fasano, A., Kalia, S. K., Yu, K. K. H., et al. (2025). Phase I trial of hES cell-derived dopaminergic neurons for Parkinson’s disease. Nature 641, 978–983. doi: 10.1038/s41586-025-08845-y
Taguchi, T., Ikuno, M., Hondo, M., Parajuli, L. K., Taguchi, K., Ueda, J., et al. (2020). α-Synuclein BAC transgenic mice exhibit RBD-like behaviour and hyposmia: a prodromal Parkinson’s disease model. Brain 143, 249–265. doi: 10.1093/brain/awz380
Taguchi, T., Ikuno, M., Yamakado, H., and Takahashi, R. (2020). Animal model for prodromal Parkinson’s disease. Int. J. Mol. Sci. 21:1961. doi: 10.3390/ijms21061961
Takahashi, J. (2020). iPS cell-based therapy for Parkinson’s disease: a Kyoto trial. Regen. Ther. 13, 18–22. doi: 10.1016/j.reth.2020.06.002
Taylor, J. R., Elsworth, J. D., Roth, R. H., Sladek, J. R., and Redmond, D. E. (1990). Cognitive and motor deficits in the acquisition of an object retrieval/detour task in mptp-treated monkeys. Brain 113, 617–637. doi: 10.1093/brain/113.3.617
Taylor, T. N., Caudle, W. M., Shepherd, K. R., Noorian, A., Jackson, C. R., Iuvone, P. M., et al. (2009). Nonmotor symptoms of Parkinson’s disease revealed in an animal model with reduced monoamine storage capacity. J. Neurosci. 29, 8103–8113. doi: 10.1523/JNEUROSCI.1495-09.2009
Terse, P. S., Kells, A. P., Noker, P., Wright, J. F., and Bankiewicz, K. S. (2021). Safety assessment of AAV2-hGDNF administered via intracerebral injection in rats for treatment of Parkinson’s disease. Int. J. Toxicol. 40, 4–14. doi: 10.1177/1091581820966315
Tolosa, E., Garrido, A., Scholz, S. W., and Poewe, W. (2021). Challenges in the diagnosis of Parkinson’s disease. Lancet Neurology 20, 385–397. doi: 10.1016/S1474-4422(21)00030-2
Torrent, R., De Angelis Rigotti, F., Dell’Era, P., Memo, M., Raya, A., and Consiglio, A. (2015). Clinical medicine using iPS cells toward the understanding of Parkinson’s disease. J. Clin. Med. 4, 548–566. doi: 10.3390/jcm4040548
Trigo-Damas, I., Del Rey, N. L.-G., and Blesa, J. (2018). Novel models for Parkinson’s disease and their impact on future drug discovery. Expert Opin. Drug Discov. 13, 229–239. doi: 10.1080/17460441.2018.1428556
Tsika, E., Kannan, M., Foo, C. S. Y., Dikeman, D., Glauser, L., Gellhaar, S., et al. (2014). Conditional expression of Parkinson’s disease-related R1441C LRRK2 in midbrain dopaminergic neurons of mice causes nuclear abnormalities without sneurodegeneration. Neurobiol. Dis. 71, 345–358. doi: 10.1016/j.nbd.2014.08.027
Ueno, T., Tomita, J., Tanimoto, H., Endo, K., Ito, K., Kume, S., et al. (2012). Identification of a dopamine pathway that regulates sleep and arousal in Drosophila. Nat. Neurosci. 15, 1516–1523. doi: 10.1038/nn.3238
Van Kampen, J. M., Baranowski, D. C., Robertson, H. A., Shaw, C. A., and Kay, D. G. (2015). The progressive BSSG rat model of Parkinson’s: recapitulating multiple key features of the human disease. PLoS One 10:e0139694. doi: 10.1371/journal.pone.0139694
Van Vliet, S. A. M., Blezer, E. L. A., Jongsma, M. J., Vanwersch, R. A. P., Olivier, B., Philippens,, et al. (2008). Exploring the neuroprotective effects of modafinil in a marmoset Parkinson model with immunohistochemistry, magnetic resonance imaging and spectroscopy. Brain Res. 1189, 219–228. doi: 10.1016/j.brainres.2007.10.059
Vehovszky, A., Szabó, H., Hiripi, L., Elliott, C. J. H., and Hernádi, L. (2007). Behavioural and neural deficits induced by rotenone in the pond snail Lymnaea stagnalis. A possible model for Parkinson’s disease in an invertebrate. Eur. J. Neurosci. 25, 2123–2130. doi: 10.1111/j.1460-9568.2007.05467.x
Verhave, P. S., Jongsma, M. J., van den Berg, R. M., Vis, J. C., Vanwersch, R. A. P., Smit, A. B., et al. (2011). REM sleep behavior disorder in the marmoset MPTP model of early Parkinson disease. Sleep 34, 1119–1125. doi: 10.5665/SLEEP.1174
Vo, Q., Gilmour, T. P., Venkiteswaran, K., Fang, J., and Subramanian, T. (2014). Polysomnographic features of sleep disturbances and REM sleep behavior disorder in the unilateral 6-OHDA lesioned hemiparkinsonian rat. Parkinson's Disease 2014:852965. doi: 10.1155/2014/852965
Wahlberg, L. U., Emerich, D. F., Kordower, J. H., Bell, W., Fradet, T., Paolone, G., et al. (2020). Long-term, stable, targeted biodelivery and egicacy of GDNF from encapsulated cells in the rat and Goettingen miniature pig brain. Current Research in Pharmacology and Drug Discovery, 1, 19–29. doi: 10.1016/j.crphar.2020.04.001
Wang, L. J., Lu, Y. Y., Muramatsu, S. I., Ikeguchi, K., Fujimoto, K. I., Okada, T., et al. (2002). Neuroprotective effects of glial cell line-derived neurotrophic factor mediated by an adeno-associated virus vector in a transgenic animal model of amyotrophic lateral sclerosis. J. Neurosci. 22, 6920–6928. doi: 10.1523/jneurosci.22-16-06920.2002
Wang, X., Zhang, J. B., He, K. J., Wang, F., and Liu, C. F. (2021). Advances of zebrafish in neurodegenerative disease: from models to drug discovery. Front. Pharmacol. 12. doi: 10.3389/fphar.2021.713963
Wang, Y.-K., Zhu, W. W., Wu, M. H., Wu, Y. H., Liu, Z. X., Liang, L. M., et al. (2018). Human clinical-grade parthenogenetic ESC-derived dopaminergic neurons recover locomotive defects of nonhuman primate models of Parkinson’s disease. Stem Cell Reports 11, 171–182. doi: 10.1016/j.stemcr.2018.05.010
Wang, Y., Liu, W., Yang, J., Wang, F., Sima, Y., Zhong, Z. M., et al. (2017). Parkinson’s disease-like motor and non-motor symptoms in rotenone-treated zebrafish. Neurotoxicology 58, 103–109. doi: 10.1016/j.neuro.2016.11.006
Watanabe, Y., Nakamura, Y., Cao, X., Ohara, H., Yamazaki, Y., Murayama, N., et al. (2015). Intravenous Administration of Apomorphine Does NOT induce long QT syndrome: experimental evidence from in vivo canine models. Basic Clin. Pharmacol. Toxicol. 116, 468–475. doi: 10.1111/bcpt.12343
West, A. B. (2017). Achieving neuroprotection with LRRK2 kinase inhibitors in Parkinson disease. Exp. Neurol. 298, 236–245. doi: 10.1016/j.expneurol.2017.07.019
Westerlund, M., Ran, C., Borgkvist, A., Sterky, F. H., Lindqvist, E., Lundströmer, K., et al. (2008). Lrrk2 and α-synuclein are co-regulated in rodent striatum. Mol. Cell. Neurosci. 39, 586–591. doi: 10.1016/j.mcn.2008.08.001
Wichmann, T., Nelson, A., Torres, E. R. S., Svenningsson, P., and Marongiu, R. (2025). Leveraging animal models to understand non-motor symptoms of Parkinson’s disease. Neurobiol. Dis. 208:106848. doi: 10.1016/j.nbd.2025.106848
Xiong, Y., Neifert, S., Karuppagounder, S. S., Liu, Q., Stankowski, J. N., Lee, B. D., et al. (2018). Robust kinase- and age-dependent dopaminergic and norepinephrine neurodegeneration in LRRK2 G2019S transgenic mice. Proc. Natl. Acad. Sci. 115, 1635–1640. doi: 10.1073/pnas.1712648115
Xiong, Y., and Yu, J. (2018). Modeling Parkinson’s disease in Drosophila: what have we learned for dominant traits? Front. Neurol. 9:228. doi: 10.3389/fneur.2018.00228
Yamaguchi, H., and Shen, J. (2007). Absence of dopaminergic neuronal degeneration and oxidative damage in aged DJ-1-deficient mice. Mol. Neurodegener. 2:10. doi: 10.1186/1750-1326-2-10
Yang, L., Zhang, X., Li, S., Wang, H., Zhang, X., Liu, L., et al. (2020). Intranasal insulin ameliorates cognitive impairment in a rat model of Parkinson’s disease through Akt/GSK3β signaling pathway. Life Sci. 259:118159. doi: 10.1016/j.lfs.2020.118159
Yang, W., Wang, G., Wang, C. E., Guo, X., Yin, P., Gao, J., et al. (2015). Mutant alpha-Synuclein causes age-dependent neuropathology in monkey brain. J. Neurosci. 35:8345. doi: 10.1523/JNEUROSCI.0772-15.2015
Yang, Y., Qian, J., Mei, J., Zhong, K., and Niu, C. (2020). In vivo detection of metabolic changes in the striatum of proteasomal inhibition-induced Parkinson’s disease in rats using proton MR spectroscopy at 9.4 T. Int. J. Neurosci. 130, 153–160. doi: 10.1080/00207454.2019.1667783
Yuan, Y., Ma, X., Mi, X., Qu, L., Liang, M., Li, M., et al. (2023). Dopaminergic neurodegeneration in the substantia nigra is associated with olfactory dysfunction in mice models of Parkinson’s disease. Cell Death Discov. 9:388. doi: 10.1038/s41420-023-01684-8
Yurek, D., Hasselrot, U., Sesenoglu-Laird, O., Padegimas, L., and Cooper, M. (2017). Intracerebral injections of DNA nanoparticles encoding for a therapeutic gene provide partial neuroprotection in an animal model of neurodegeneration. Nanomedicine 13, 2209–2217. doi: 10.1016/j.nano.2017.06.010
Zambusi, A., and Ninkovic, J. (2020). Regeneration of the central nervous system-principles from brain regeneration in adult zebrafish. World J. Stem Cells 12, 8–24. doi: 10.4252/wjsc.v12.i1.8
Zarranz, J. J., Alegre, J., Gómez-Esteban, J. C., Lezcano, E., Ros, R., Ampuero, I., et al. (2004). The new mutation, E46K, of alpha-synuclein causes Parkinson and Lewy body dementia. Ann. Neurol. 55, 164–173. doi: 10.1002/ana.10795
Zeng, X.-S., Geng, W.-S., and Jia, J.-J. (2018). Neurotoxin-induced animal models of Parkinson disease: pathogenic mechanism and assessment. ASN Neuro 10:1759091418777438. doi: 10.1177/1759091418777438
Zhang, C., Li, C., Chen, S., Li, Z., Jia, X., Wang, K., et al. (2017). Berberine protects against 6-OHDA-induced neurotoxicity in PC12 cells and zebrafish through hormetic mechanisms involving PI3K/AKT/Bcl-2 and Nrf2/HO-1 pathways. Redox Biology, 11, 1–11. doi: 10.1016/j.redox.2016.10.019
Zhang, Q., Liu, M., Xu, Y., Lee, J., Jones, B., Li, B., et al. (2024). Tilorone mitigates the propagation of α-synucleinopathy in a midbrain-like organoid model. J. Transl. Med. 22:816. doi: 10.1186/s12967-024-05551-7
Keywords: Parkinson’s disease, models, drug development, motor symptoms, non-motor symptoms, preclinical trials, therapies
Citation: Reinares-Sebastián A, Esteban-García N, Takada M and Trigo-Damas I (2025) Drug discovery and development for Parkinson’s disease: are preclinical models good enough? Front. Aging Neurosci. 17:1692592. doi: 10.3389/fnagi.2025.1692592
Edited by:
Philip Wing-lok Ho, Hong Kong Polytechnic University, Hong Kong SAR, ChinaReviewed by:
Eunice Chang, The Hong Kong Polytechnic University, Hong Kong SAR, ChinaSiqing Cai, Peking University Health Science Center, China
Copyright © 2025 Reinares-Sebastián, Esteban-García, Takada and Trigo-Damas. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Inés Trigo-Damas, aXRyaWdvLmhtY2luYWNAaG1ob3NwaXRhbGVzLmNvbQ==