 Ricardo Figueroa-Huitrón
Ricardo Figueroa-Huitrón Hibraim Adán Pérez-Mendoza
Hibraim Adán Pérez-Mendoza- 1Laboratorio de Ecología Evolutiva y Conservación de Anfibios y Reptiles, Facultad de Estudios Superiores Iztacala, Universidad Nacional Autónoma de México (UNAM), Tlalnepantla, Estado de México, Mexico
- 2Posgrado en Ciencias Biológicas, Unidad de Posgrado, Universidad Nacional Autónoma de México, Ciudad de México, Mexico
Introduction: Thermal tolerance and performance breadth can define the suitable areas where reptiles can survive and perform at an optimal physiological level. It is important to determine the distribution of suitable areas for reptile physiology and to assess how they could change in future climate change scenarios. In this paper, we utilized a mechanistic approach to model current and future physiological suitability of three snakes endemic to Mexico: Crotalus polystictus, Conopsis lineata and Thamnophis melanogaster.
Methods: Utilizing thermal physiology data previously generated, we modeled operative temperatures in the present and in 2070, under SSP3-7.0 and SSP5-8.5. We utilized performance breadth as the suitability threshold and calculated the amount of daily and annual hours that fall within this range in each grid cell of the accessibility areas of the three species
Results: We found that suitability is at its highest on the pine-oak forests in mountainous regions. Future scenarios will provide wider windows with favorable temperatures, and the suitability of the three species will increase in most of their accessible areas. On average, annual hours of physiological suitability will increase by 103% and 133% (under SSP3-7.0 and SSP5-8.5, respectively) for C. polystictus, 12% and 14% for C. lineata, and 17% and 20% for T. melanogaster
Discussion: These results show that the thermal environments of the mountainous regions of Mexico are very important for the sustenance of these species in a changing world. These areas are hotspots for reptile biodiversity, but they face problems of area reduction. Further research considering the effect of temperature on reproduction or the change in precipitation, would provide a broader picture about the physiological responses of these species. Integrating physiological information like this in a geographical context is important to designing adequate conservation strategies for reptiles.
1 Introduction
Climate change is rapidly altering environmental conditions on a global scale. Reptiles are one of the groups believed to experience these alterations more drastically, since their physiology is directly linked to environmental temperatures (Sinervo et al., 2010). However, reptiles have developed diverse behavioral strategies to mitigate the negative effects of thermal variation, including changes in activity periods or microhabitat selection (Kearney et al., 2009). This capacity of behavioral adjustments to thermoregulation is related to the availability of suitable microhabitats and the structure of the thermal landscape (Årevall et al., 2018; Sears and Angilletta Jr., 2015). For instance, some species which experience variable thermal conditions within their distribution can adjust their activity periods along the day to better exploit the thermal landscape (Weatherhead et al., 2012).
In addition to behavior, differences in physiological sensitivity play a key role in how reptiles respond to climate change (Aubret and Shine, 2010; Huey et al., 2012). Adjustments in physiological traits such as metabolic rates or locomotor performance have been reported to aid in thriving across thermally variable environments (Plasman et al., 2020; Stellatelli et al., 2022). Analyzing the performance breadth, which is the range of temperatures over which organisms can perform adequately (Taylor et al., 2021), is a reliable way to address the extent of thermal tolerance of organisms and assess their ability to withstand temperature variation (e.g. Gómez-Alés et al., 2018; Vicenzi et al., 2019).
However, not all species are plastic enough to cope with rising temperatures, and there can be a limit to how much they can adapt in short periods of time (van Heerwaarden et al., 2016; Zhang et al., 2023). On the other hand, if thermal conditions are not optimal at present, some species may benefit from climate change, since rising temperatures can improve the thermal landscape and provide more suitable conditions for their physiological processes (Cabezas-Cartes et al., 2019; Youngblood et al., 2023). Physiological performance is notoriously temperature dependent, so as temperature increases so does performance. However, after reaching the thermal optimum, there is a threshold beyond which performance decreases rapidly and overheating risk is higher (Angilletta Jr., 2006; Tomlinson, 2019). For species whose optimal or preferred temperatures are close to this thermal threshold, the room for adjustments can be too thin. These species are said to have a low thermal safety margin and often can experience greater risk of overheating (Sunday et al., 2014; Sánchez-Fernández et al., 2016). Taking this into consideration, it is important to assess how the conditions for optimal physiological performance could change.
Mechanistic models are ideal for this endeavor, since they incorporate biological and physiological processes to explain the relationship between the environment and organismal performance (Buckley et al., 2010; Taylor et al., 2021). A common approach to translating this relationship into an evaluation of suitability is to estimate the amount of time when favorable temperatures are available for organisms. Several studies have utilized the calculation of hours of activity and restriction to assess the effects of climate change on the distribution of reptiles (Ceia-Hasse et al., 2014; Mizsei et al., 2024; Sinervo et al., 2010). A caveat, however, of mechanistic models is that they often rely on very precise biological information for calibration, which can be difficult to obtain for inconspicuous species (Buckley et al., 2010; Evans et al., 2015).
In Mexico, studies on the thermal biology of snakes have been increasing in recent years (Arnaud et al., 2019; Jaramillo-Alba et al., 2020; Venegas-Barrera et al., 2025). However, assessments studies on the potential effects of climate change and risk projections have relied solely on correlative niche models (González-Fernández et al., 2018; Rubio-Blanco et al., 2024; Sunny et al., 2023). It is therefore relevant to use the physiological information being generated to develop further estimates of the effects of climate change. With this in mind, we gathered physiological information from three model snake species and tailored mechanistic models that assess the impact of climate change on their thermal physiology. We selected three snake species endemic to Mexico: Crotalus polystictus, Conopsis lineata and Thamnophis melanogaster. These species are important and abundant representatives of the herpetofauna of central and western Mexico, and in recent years their thermal ecology and physiology have been studied in detail (Figueroa-Huitrón et al., 2024a, 2024b). In this study, we carried out microclimatic and biophysical models within the NicheMapR framework (Kearney and Porter, 2017, 2020) to estimate body temperatures in the present and under different climate change scenarios (year 2070, under SSP3-7.0 and SSP5-8.5). With that data we used thermal performance breadth as the threshold to evaluate physiological suitability by calculating how much time the snakes experience temperatures that enable optimal performance. Since these species have wide intervals of thermal preference and performance breadth, we expect them to be able to buffer the rise of temperatures and thus maintain adequate levels of physiological suitability in the future scenarios.
2 Materials and methods
2.1 Study species
Conopsis lineata is a small-sized, fossorial snake with an average snout-vent length (SVL) of around 18 cm. It spends most of its time within burrows or under rocks and logs, where it feeds on larvae, insects, and other arthropods (García-Balderas et al., 2014; Ramírez-Bautista et al., 2009). It is distributed in central and southern Mexico, inhabiting pine-oak forests, montane cloud forests, xerophytic shrubs, and fir forests in an altitude range from 1,700 to 3,100 m above sea level (Goyenechea and Flores-Villela, 2006; Ramírez-Bautista et al., 2009).
Crotalus polystictus is a medium-sized, terrestrial viper with an average SVL of about 60 to 70 cm (Campbell and Lamar, 2004). It inhabits pine-oak forests, shrublands, and both dry and humid grasslands in central and western Mexico, between 1,450 and 2,739 m of elevation (Meik et al., 2012). This species is a sit-and-wait predator that feeds mostly on small rodents (Mackessy et al., 2018).
Thamnophis melanogaster is a medium-sized, semi-aquatic colubrid with an average SVL of about 40 cm (Manjarrez et al., 2017). It inhabits riverbanks and lake shores at an elevation range from 1,158 to 2,545m above sea level and is widely distributed throughout Mexico (Ramírez-Bautista et al., 2009; Rossman et al., 1996). It uses shelters on land, near aquatic bodies, and forages actively underwater, feeding on aquatic prey (Manjarrez et al., 2013).
2.2 Field work and ecophysiological experiments
We collected data of these species in three locations in Central Mexico where they are locally abundant. We sampled C. lineata in a forest clearing at “Los Dinamos,” an ecotourism park within the Magdalena River basin located in the western mountainous region of Mexico City, during the period from 2019 to 2023. We sampled C. polystictus in a cropland area surrounded by grasslands and patches of oak forest in San Bartolo Morelos, located in the northwest region of Estado de México, over the period from 2017 to 2023. We sampled T. melanogaster on the banks of the Arcos del Sitio River within the Sierra de Tepotzotlán State Park in Estado de México, during the period from 2021 to 2023.
With the snakes we encountered, we first conducted a study on thermal ecology (Figueroa-Huitrón et al., 2024b). We registered the field body temperature (Tb) of each snake we encountered (sample size was 221 for C. polystictus, 66 for C. lineata and 38 for T. melanogaster) and determined the thermal intervals of activity. Using thermal gradients, both on semi-captivity and in the laboratory, we obtained data on the average preferred temperature (Tpref) and the set-point temperature range (Tset) of the species (sample size was 92 for C. polystictus, 35 for C. lineata and 28 for T. melanogaster, we made between 6 and 12 measurements per individual), as defined by Hertz et al. (1993). Afterwards, we conducted a study on the thermal ecophysiology of these species (Figueroa-Huitrón et al., 2024a), in which we analyzed the effect of five thermal treatments (15°C, 25°C, 30°C, 33°C and 36°C) on locomotor performance (sample size was 33 for C. polystictus, 31 for C. lineata and 38 for T. melanogaster, we made one measurement per individual for each treatment). We obtained the critical temperatures (CTmax and CTmin), at which performance equals zero, and calculated thermal performance curves (TPC’s) for each species. The full methods for both the thermal ecology and ecophysiology procedures are detailed in Figueroa-Huitrón et al. (2024a, 2024b).
2.3 Physiological modelling
To design the accessible area for each species we first downloaded the geographical records of the three species from the GBIF database (available at https://zenodo.org/records/17438062). After eliminating duplicates and records with missing or wrong coordinates, we worked with 61 records for C. polystictus, 350 for C. lineata and 159 for T. melanogaster. We took into consideration the Neotropical biogeographic provinces (Morrone et al., 2022) and the WWF terrestrial ecoregions (Dinerstein et al., 2017). We selected the provinces and ecoregions that matched with the species records and then pruned this selection taking into consideration biogeographical aspects of the species (location of historical records, presence of geographical barriers, vagility of the organisms) and the distribution maps proposed by the IUCN (IUCN, 2024). A graphical visualization of this process is available in Supplementary Figure S1, on the Supplementary Material. We then transformed each accessible area’s vector into a raster at a resolution of 5 arcminutes and extracted the coordinates of each cell to subsequently run the mechanistic models, one at each site. While this resolution may reduce detectable microhabitat heterogeneity, computational limitations and the extent of the accessibility ranges of these species prohibited the use of a finer scale. These analyses were made with QGis 3.40 and R version 4.41 (R Core Team, 2024) with the package terra (Hijmans, 2024).
With the physiological data that we previously estimated, we tailored mechanistic models for each species at each pixel of their accessibility areas (1607 models for C. polystictus, 3363 for C. lineata and 5627 for T. melanogaster). We used the R package NicheMapR which, based on terrain features and climate conditions as well as calculations of energy exchange processes, calculates the microclimates to which an organism is exposed (Kearney and Porter, 2017) and the resulting body temperatures (Kearney and Porter, 2020).
First, we adjusted the microclimate models with the function “micro_global”, which relies on the global climate database developed by New et al. (2002). We set the models to calculate hourly data at a time interval of 365 days over a continuous year. Since none of our species are arboreal, we set the “Usrhyt” parameter to 10 cm, so that air temperature would be calculated near ground level. We left the “DEP” parameter at default, so that the microhabitat characteristics of the soil would be calculated at 0, 2.5, 5, 10, 15, 20, 30, 50, 100 and 200 cm of depth.
We then applied the biophysical model with the “ectotherm” function. This function runs with the results generated by the microclimate function and calculates the operative temperatures that the animals would experience in the same period and conditions set for the microclimate models (Kearney and Porter, 2020). In relation to the DEP parameter from micro_global, the ectotherm function determines that an organism that reaches unfavorable temperatures can burrow to avoid overheating. We defined the maximum borrowing depths as 50 cm for C. polystictus and T. melanogaster, and 100 cm for C. lineata. We defined the other basic parameters of the ectotherm function with relevant physiological information derived from our previous studies (Table 1). Since Tset is a measure of the preferred thermal threshold in which organisms operate, we used the lower and upper limits of Tset as the minimum and maximum foraging temperatures, respectively. The minimum temperatures at which organisms move away from retreats and start basking were derived from our Tb observations during fieldwork (the lowest Tb recorded in individuals found in the open, early in the morning during the beginning of their activity period).
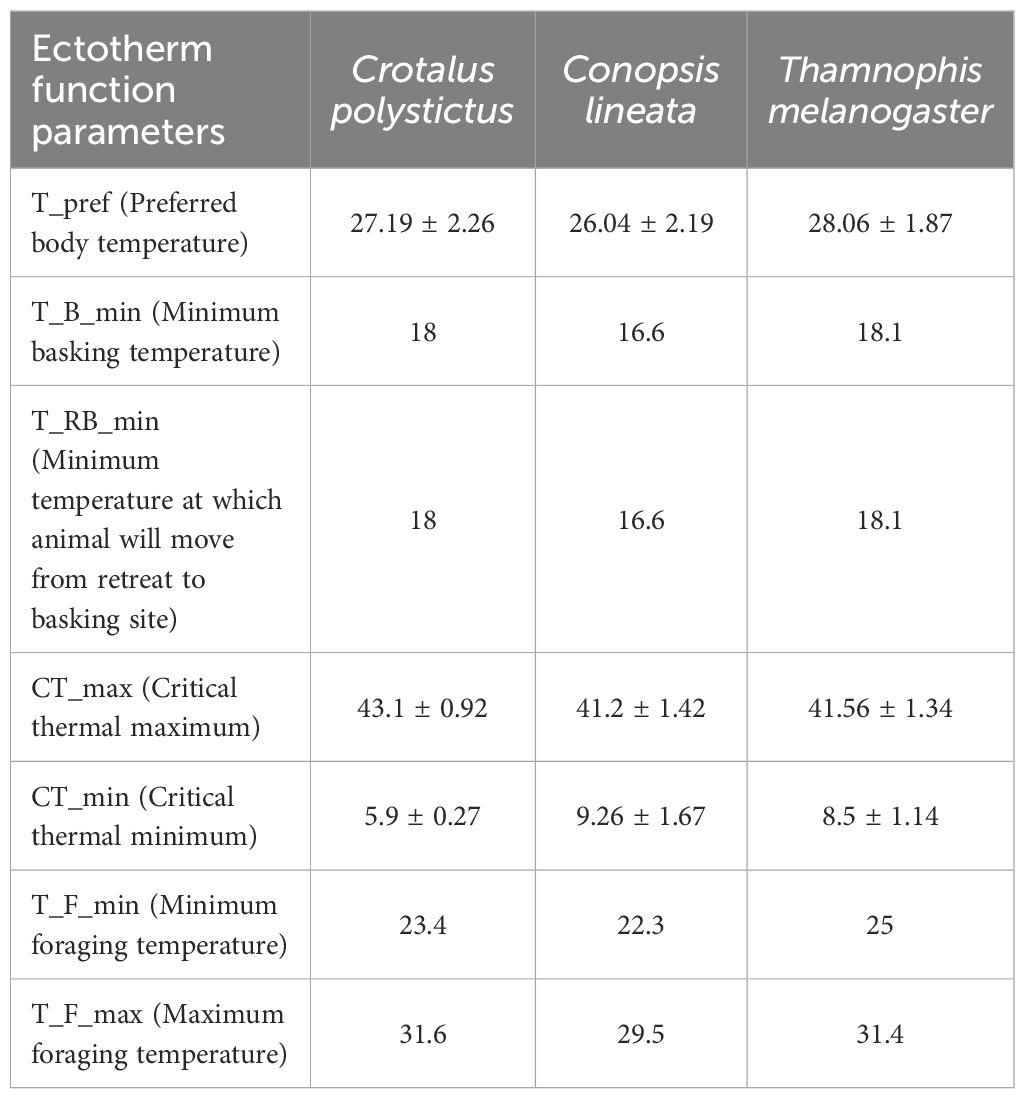
Table 1. Custom input values (in °C, ± SD) used in the “ectotherm” function of NicheMapR for each species.
To determine the thermal threshold of physiological optimum, we calculated the thermal performance breadth (B90) from each species’ TPC. B90 refers to the thermal interval in which performance is at least 90% of the maximum value (Supplementary Figure S1). We consider this is an adequate threshold for physiological optimum since it encompasses most of the extension of the TPC in which performance is adequate, and excludes the tails of the curve, where performance levels drop significantly. In comparison to other measurements of performance breadth, B90 is a more restrictive measure than B80 but does not come as close to critical limits like B95. After obtaining estimates of operative temperatures with NicheMapR, we made physiological suitability calculations in two time scales: 1) We counted how many hours through the entire year fell within B90 to determine a suitability value for each pixel (measured in suitable hours per year). We then attached each pixel’s count into the accessible areas and plotted the resulting raster using QGis 3.40. 2) We counted how many hours per day fell within B90 and averaged them across all sites to get a daily suitability value; this gave us information about the phenological change of physiological suitability.
2.4 Climate change projections
To estimate the change in physiological suitability for each species, we calculated the projected increase in temperature and reran the NicheMapR models. We downloaded historical (1970-2000) maximum temperature data from WorldClim version 2.1 (Fick and Hijmans, 2017) to set the reference for “present” thermal conditions. Regarding future conditions, we first followed the method and application (GCM compareR, downloaded from https://github.com/marquetlab/GCM_compareR) developed by Fajardo et al. (2020) to select the general circulation models (GCM’s). We compared the GCM’s available (Supplementary Table S1) and selected three: IPSL.CM6A.LR, MRI.ESM2.0 and CNRM.CM6.1. The results from GCM compareR showed that these three GCM’s do not deviate extremely from the average of the general comparison, but their projections represent environmental changes in different directions. For each GCM, we downloaded monthly averages of maximum air temperatures for the year 2070 under the 3-7.0 and 5-8.5 shared socioeconomic pathways (SSP’s) from WorldClim version 2.1. These SSP’s represent different emissions scenarios driven by different socioeconomic assumptions: 3-7.0 represents high greenhouse gas emissions and low climate change mitigation policies, while 5-8.5 represents the worst-case scenario, with the highest emissions. We chose these scenarios because we wanted to see the responses of the species in the medium term, considering challenging conditions for their physiology. We then extracted the thermal data in each of the pixels in the accessible areas of the three species and subtracted the current values from the future ones. We averaged the monthly differences to obtain a single thermal change value per pixel, which we used as input for the “warm” argument of the “micro_global” function. This argument represents offset values that are added to the baseline temperature values used by NicheMapR, thus allowing the models to be run considering the projected temperature rise. We reran the microclimate and ectotherm models with the offset values and computed the hours of optimal physiology for each future scenario, in the same way as we did for the present. Finally, we calculated the differences in the number of annual hours of optimal physiology between each future scenario and the present, for each pixel. This allowed us to determine whether physiological suitability increases or decreases at each pixel and thus compare the different scenarios with the present.
3 Results
3.1 Physiological suitability in the present
The B90 interval derived from the TPC’s was 30.2 - 38.7°C for C. polystictus, 22.5 - 32.5°C for C. lineata and 23.2 – 33.2°C for T. melanogaster. The shapes of the TPC’s and the B90 limits within them are represented on Supplementary Figure S2. Our analysis of suitable hours counts across each species’ distribution shows that all three experience a wide range of suitability. Crotalus polystictus has the widest interval, ranging from just 20 to 3990 hours of optimal physiology (Figure 1A). On the other hand, C. lineata and T. melanogaster have narrower intervals, that go from 2720 to 6320 and from 2540 to 6840 hours, respectively (Figures 2A and 3A). Overall, the three species share Jalisco, central Michoacán and southern Estado de México as some of the areas of greater physiological suitability (Figures 1A, 2A, 3A). C. lineata and T. melanogaster also share southern San Luis Potosí and northern Querétaro and Hidalgo. C. lineata also has highly suitable areas in Oaxaca, whereas T. melanogaster has some at the southern part of the Sierra Madre Occidental. Through the year, C. polystictus experiences the highest suitability in spring, from April to June (Figure 4A), when daily hours of suitability rise beyond two. Conopsis lineata and T. melanogaster experience high suitability during summer, from May to September, when daily hours of suitability range from 10 to 15 (Figures 4C, E).
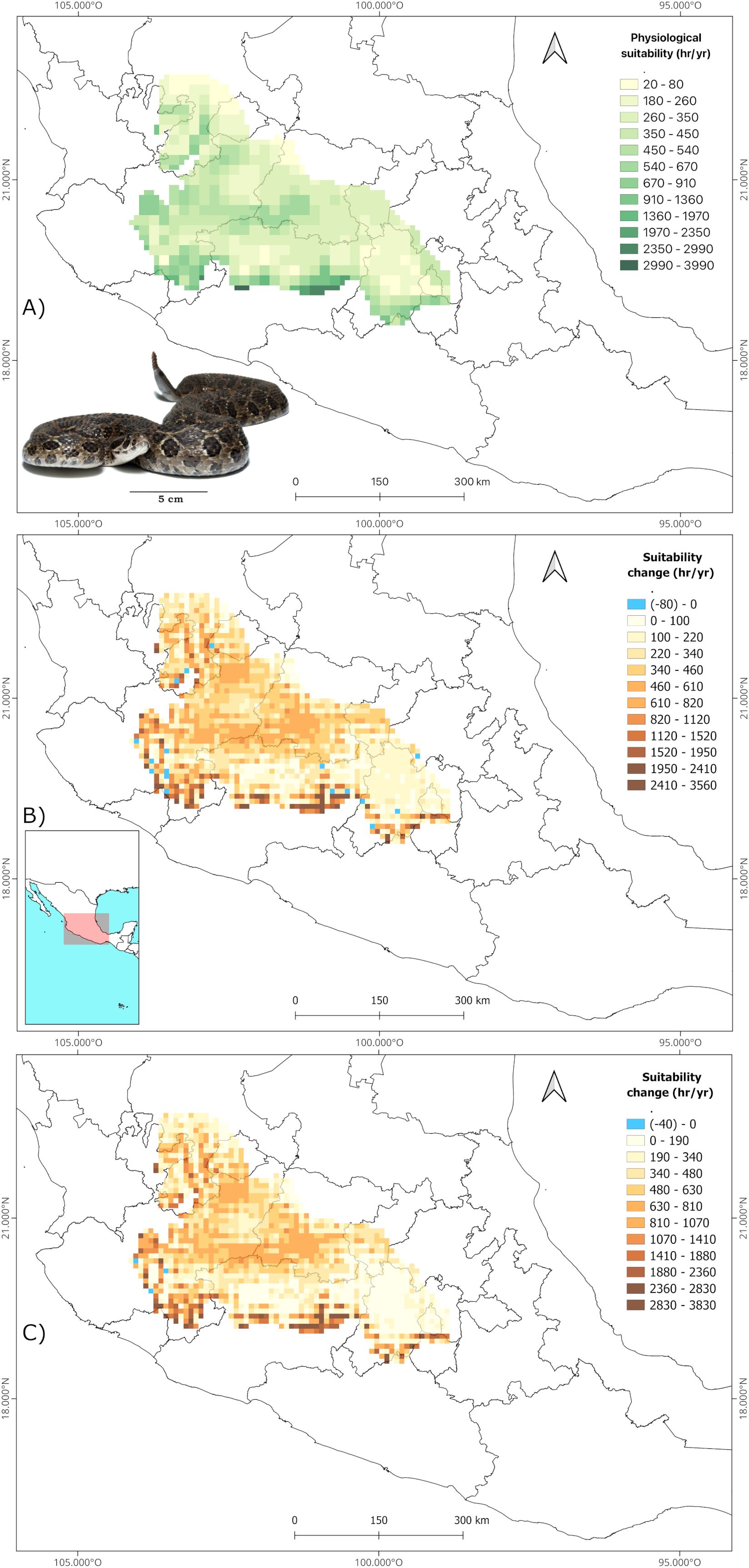
Figure 1. Estimation of annual hours of physiological suitability for Crotalus polystictus in the present (A) and change predictions in future scenarios 2070 SSP 3-7.0 (B) and 2070 SSP 5-8.5 (C). Photograph credits: L. E. Bucio-Jiménez.
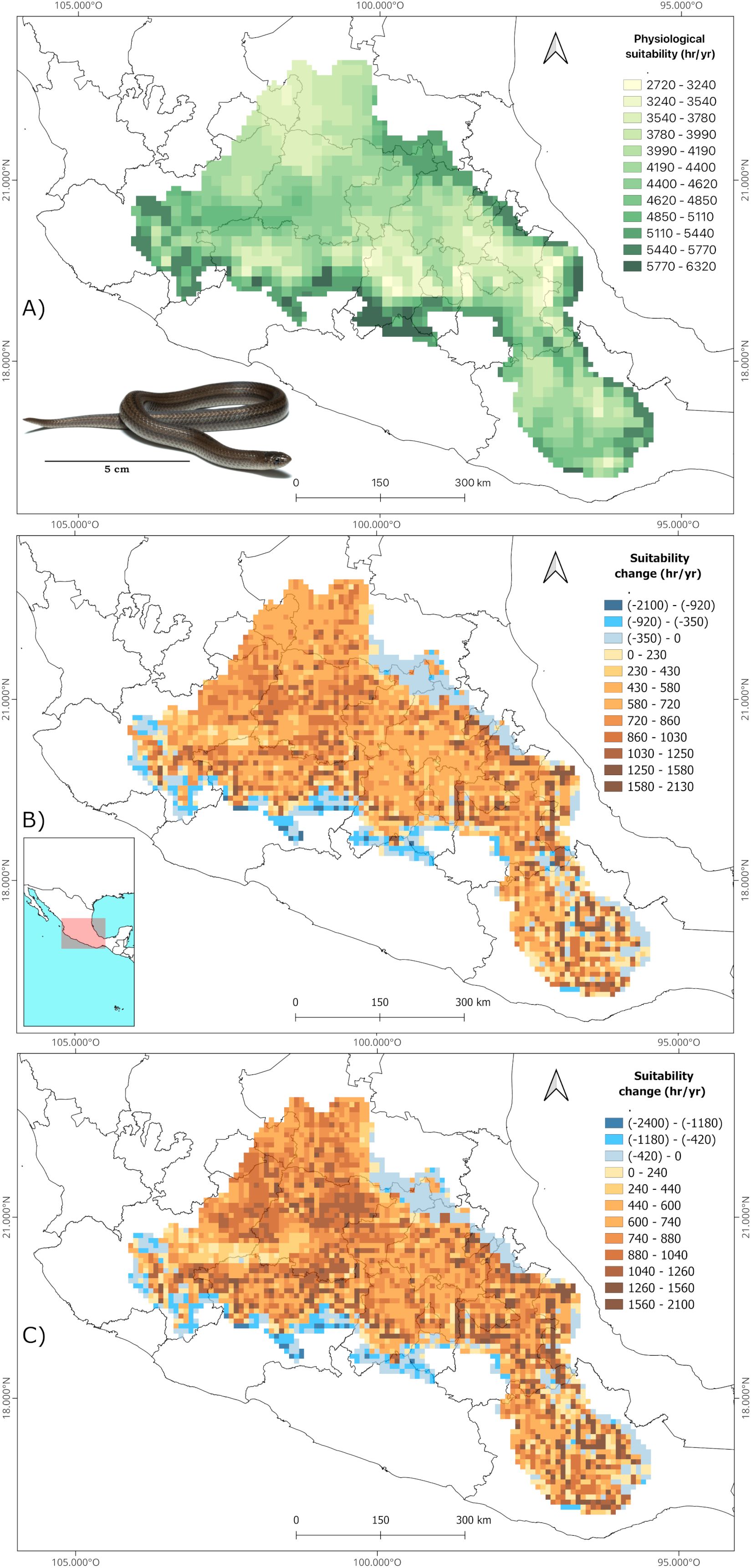
Figure 2. Estimation of annual hours of physiological suitability for Conopsis lineata in the present (A) and change predictions in future scenarios 2070 SSP 3-7.0 (B) and 2070 SSP 5-8.5 (C). Photograph credits: L. E. Bucio-Jiménez.
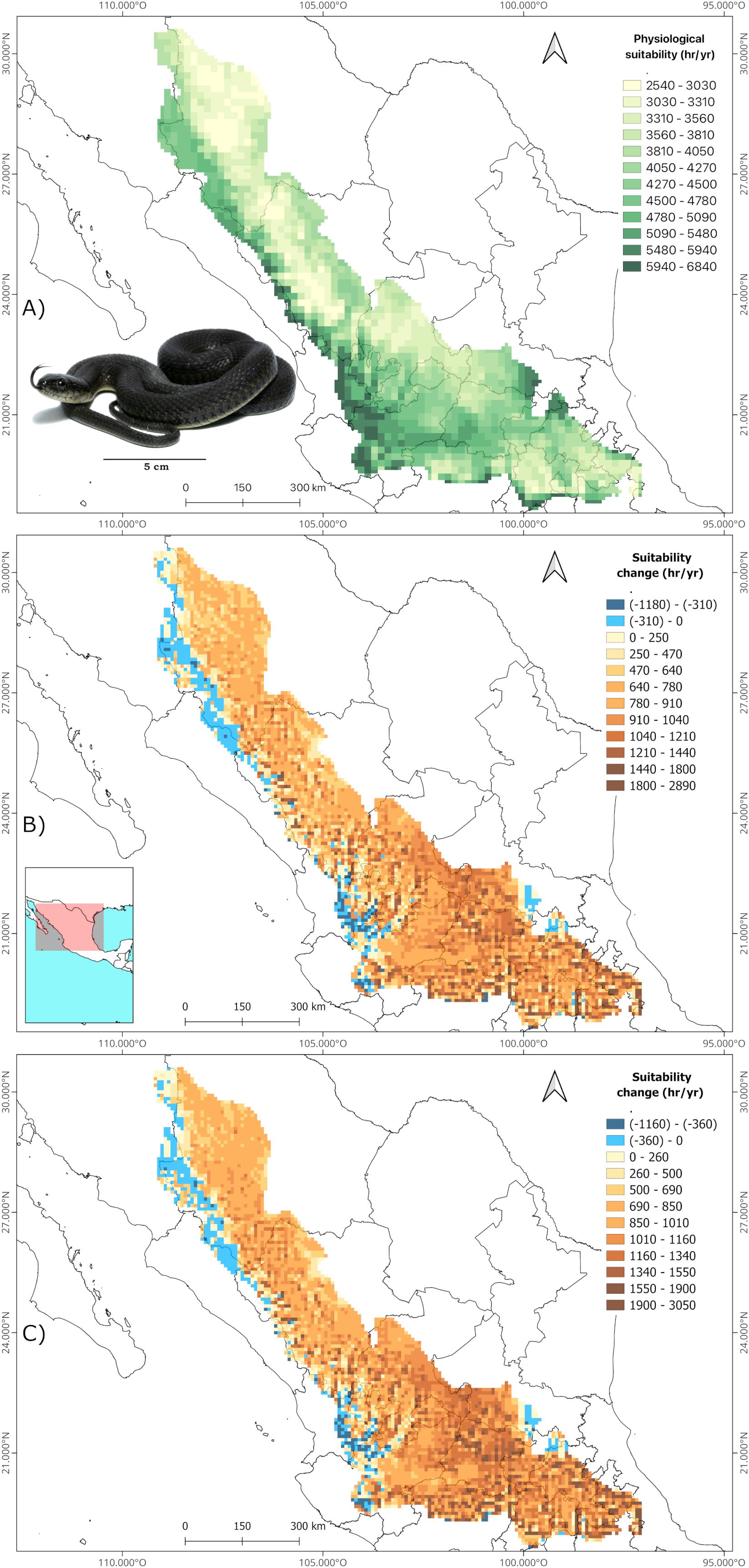
Figure 3. Estimation of annual hours of physiological suitability for Thamnophis melanogaster in the present (A) and change predictions in future scenarios 2070 SSP 3-7.0 (B) and 2070 SSP 5-8.5 (C). Photograph credits: L. E. Bucio-Jiménez.
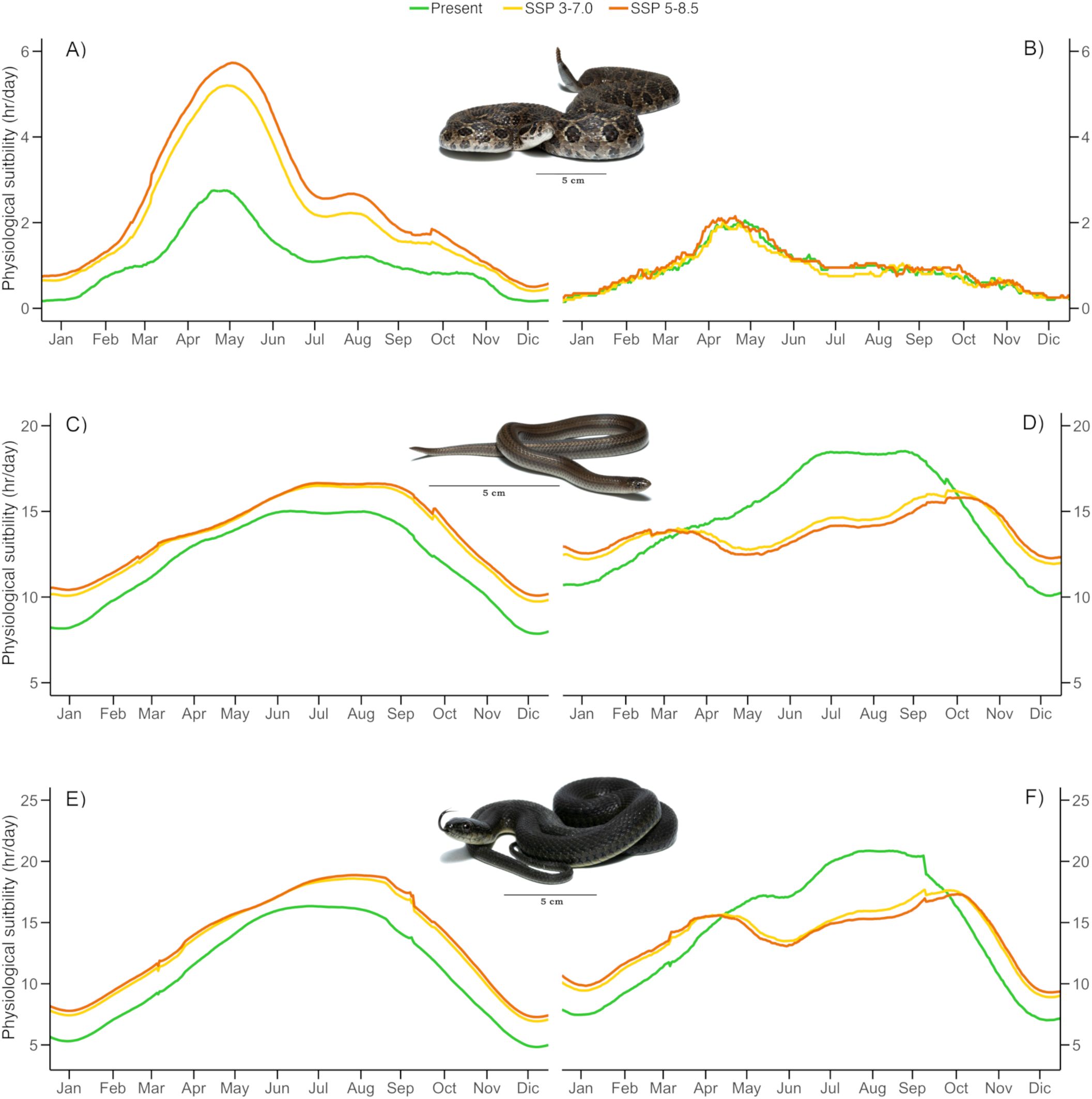
Figure 4. Mean daily physiological suitability throughout a continuous year for present and future scenarios. Panels (A, C, E) represent calculations for all sites, whereas panels (B, D, F) represent calculations only for the sites of each species where suitability decreases in future scenarios. (A, B): Crotalus polystictus, (C, D): Conopsis lineata, (E, F): Thamnophis melanogaster. Photographs credits: L. E. Bucio-Jiménez.
3.2 Future scenarios
We found that the singular values of annual suitable hours per cell vary according to each GCM, however, the amplitude of such variation is little across the GCM’s, and the results of the three coincide greatly in the direction and amplitude of the change in suitability per pixel. Our focus was on the geographical array of the physiological suitability, which is very similar across all GCM’s. Therefore, we will focus only on the projections derived from MRI.ESM2.0, which is the one that yielded the most contrasting results in comparison to present suitability. The maps of the other GCM’s are provided on the Supplementary Material (Supplementary Figures S3-S11). It is projected that all three species will experience an increase in physiological suitability in most of their accessible areas in both future scenarios (Figures 1B, C, 2B, C, 3B, C). Crotalus polystictus will experience the largest increase, by a wide margin, being more than 100% on average in both future scenarios (Table 2). Considering the SSP3-7.0 scenario, average suitability will increase by 12.1% and 17.3% for C. lineata and T. melanogaster, respectively. Under the warmer SSP 5-8.5 scenario, average suitability will increase at a higher rate, by 14% for C. lineata and by 20% for T. melanogaster.
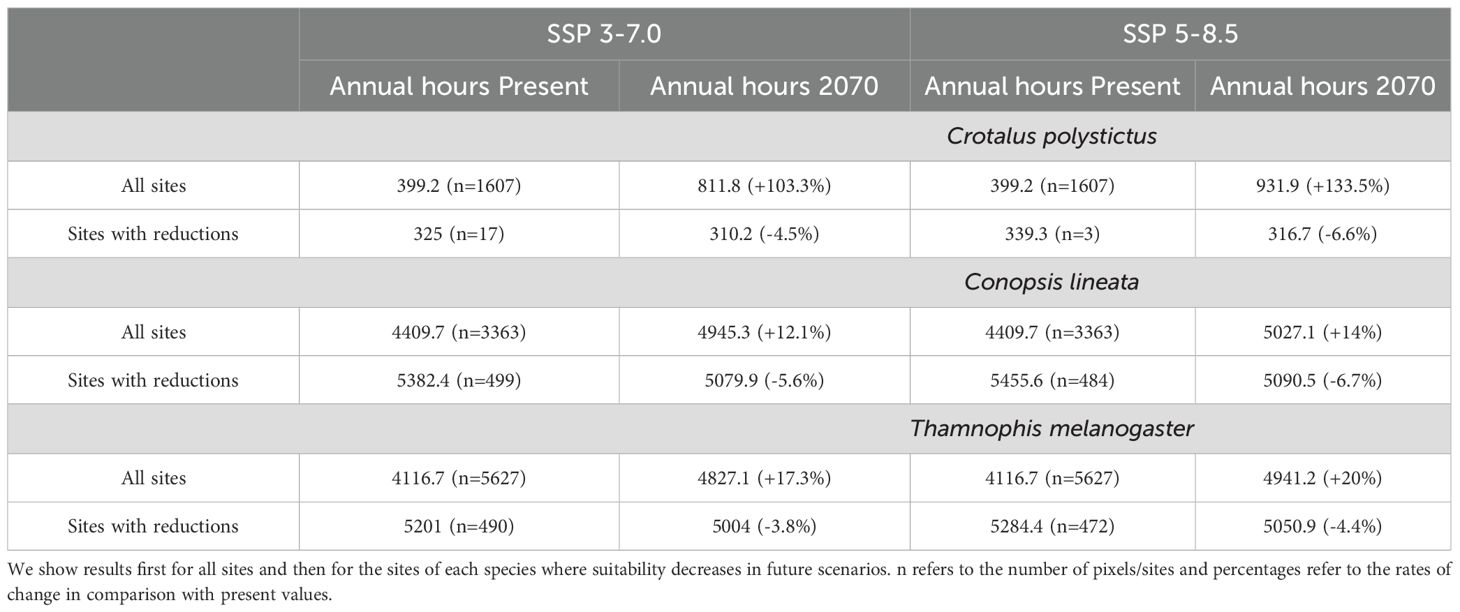
Table 2. Mean values of annual hours of physiological suitability for each species at the present and two climate change scenarios.
While the average physiological suitability will increase, there are some sites for the three species where it will decrease. Crotalus polystictus will experience the least reductions. In the SSP3-7.0 scenario, 17 of its sites will experience a 4.5% reduction, and only three sites on the SSP5-8.5 scenario will experience a 6.6% reduction (Figures 1B, C, Table 2). Conopsis lineata and T. melanogaster will experience reductions in several areas, although these overlap with some of their most suitable sites at present, where average suitability is more than 5000 hours (Figures 2B, C, 3B, C, Table 2). Considering the SSP3-7.0 scenario, there are 499 sites for C. lineata and 490 sites for T. melanogaster in which annual hours of suitability will be reduced by 5.6 and 3.8%, respectively (Table 2). In the SSP5-8.5 scenario the reductions increase: there are 484 sites for C. lineata and 472 for T. melanogaster whose suitability will be reduced by 6.7 and 4.4%, respectively (Table 2).
Regarding phenological variation, all three species will experience an increase in daily physiological suitability throughout the year (Figures 4A, C, E). While the increase will be relatively uniform for C. lineata and T. melanogaster, C. polystictus will experience a more pronounced increase from March to June, when suitability will approximately double (Figure 4A). In the sites that will suffer reductions, such decreases will be during April to October for C. lineata and T. melanogaster. During winter, suitability will still be higher than that of the present (Figures 4D, F). For C. polystictus, only a small decrease will happen from May to August, on the SSP 3-7.0 scenario (Figure 4B).
4 Discussion
Our results indicate that the mountainous regions of Mexico harbor the areas that present the highest physiological suitability for the three species. Our study species, having wide intervals of thermal preference and performance breadth, can physiologically cope with the environmental variability of these regions. The three species have high suitability levels across the Trans-Mexican Volcanic Belt (TMVB), which is a very important biogeographic region and a hotspot for reptile biodiversity (Bryson et al., 2011; González-Fernández et al., 2018). In an ecological niche study of rattlesnakes from central Mexico, Rubio-Blanco et al. (2024) found similar results for C. polystictus, noting that the highest suitability is found in central states along the TMVB. For C. lineata, other mountainous areas with high physiological suitability are the Sierra Madre Oriental and the Sierra Madre del Sur, while T. melanogaster has high suitability on the Sierra Madre Occidental. Sunny et al. (2023) found similar suitable areas for C. lineata, however, their results show more suitability on the Mexican plateau and less on the Sierra Madre Oriental. There is also an ecological niche study that modeled T. melanogaster, and their results are very close to ours, with the western TMBV and southern Sierra Madre Occidental being the most suitable areas (González-Fernández et al., 2018). These results highlight the importance of the mountainous regions of Mexico for these species. Notably, these regions represent areas of great reptile diversification in the past and harbor great reptile diversity in the present (Cisneros-Bernal et al., 2022; Mastretta-Yanes et al., 2015; Ramírez-Arce et al., 2025; Ramírez-Bautista and Cruz-Elizalde, 2013).
Across a continuous year, C. polystictus has the highest suitability in spring, whereas C. lineata and T. melanogaster have the highest suitability in summer. This generally aligns with the activity patterns we found previously (Figueroa-Huitrón et al., 2024b). With the high B90 of C. polystictus, it was expected that the most suitable period of the year was that of the highest temperatures. Particularly, this period aligns with the late gestation of females, and it is well documented that pregnant females require more precise thermoregulation during this process (Lourdais et al., 2013; Marinho et al., 2022). High suitability for C. lineata and T. melanogaster aligns with the rainy season in most areas of Mexico, when prey abundance is high and snake parturitions occur (Manjarrez and San-Roman-Apolonio, 2015; Setser et al., 2010).
The three species experience a wide range of suitability across their distribution. However, in this regard, there is a clear difference between C. polystictus and the other two species, since its count of thermally suitable hours, both annually and daily, is much lower. This result is derived from the way in which temperature affects performance in this species. Thermal performance curves have a natural left-skewed shape (Angilletta Jr., 2006; Bulté and Blouin-Demers, 2006), but such skewness is more pronounced on the TPC of C. polystictus (Figueroa-Huitrón et al., 2024a). This means that C. polystictus reaches optimal levels of performance at higher temperatures, resulting in a higher B90 and a low count of physiological suitability in our calculations. However, we have found that this species targets a wide interval of temperatures while thermoregulating and can be considered a thermal generalist (Figueroa-Huitrón et al., 2024b). With such high performance breath, it is likely that C. polystictus has adapted to thermoregulate at suboptimal temperatures. Conopsis lineata and T. melanogaster, on the other hand, have wider and lower performance breath, so they experience higher physiological suitability values. The lifestyles of these two species may allow them to maintain performance on a wider set of environments. Fossorial species like C. lineata can benefit from burrow temperatures as a buffer to achieve optimal temperatures (Camacho et al., 2015). In a similar way, semiaquatic species like T. melanogaster can benefit from the more thermally stable aquatic environments (McConnachie et al., 2011).
Our results from the mechanistic models suggest that physiological suitability for the three species will increase within the climate change context. We have found that the three species can maintain physiological performance and thermoregulation on wide thermal intervals (Figueroa-Huitrón et al., 2024a, 2024), and current conditions do not appear to provide snakes with the full spectrum of temperatures to maximize their physiological performance. Having wider temporal windows to perform at optimal levels would be beneficial at first instance, but taking into consideration the shape of TPC’s, operating at higher, more optimal temperatures would also mean operating closer to the threshold beyond which performance decreases, and closer to CTmax. If the thermal limits are not plastic enough, this would lead to a reduction in the thermal safety margin, which increases the risk of overheating (Seebacher et al., 2023; Sunday et al., 2014). This is most evident with C. polystictus, which already has the narrowest thermal safety margin of our species. However, the variability of habitats found on the areas these species inhabit can incentivize behavioral thermoregulation that would buffer exposure to critical temperatures (Goller et al., 2014; Sears and Angilletta Jr., 2015). A more holistic analysis that incorporates the changes that climate change could impose into habitat heterogeneity would allow us to see a bigger picture of this matter.
Focusing on the sites that are projected to experience reductions in suitability, the period of highest activity for C. lineata and T. melanogaster is when suitability will decrease. These reductions go from 15–20 to 10–15 daily hours of physiological suitability. We found previously that the daily activity pattern of these snakes is roughly from 9 am to 8 pm, just above 10 hours (Figueroa-Huitrón et al., 2024b). This could suggest that, on sites that are already warm enough to offer high suitability, potential future reductions may not represent highly risky scenarios. We also found that such sites with projected reductions are located within some of the areas that are most suitable in the present. The reduction rates at these sites are low, less than 10% for all species. The vegetation in these sites is mostly pine-oak and tropical dry forests, which are some of the most important ecosystems for reptile diversity (Hernández-Salinas et al., 2023; Sánchez-López et al., 2025). These places must be prioritized in conservation efforts, since pine-oak and tropical dry forests will face important threats for their distribution and biodiversity, according with various climate change projections (Carrell et al., 2023; Gómez-Mendoza and Arriaga, 2007; Manrique-Ascencio et al., 2024; Siyum, 2020).
Overall, our temperature-based mechanistic models project increasing temperatures that could provide these species with more opportunities for optimal physiological performance. Even in the sites where it decreases, it seems that physiological suitability could remain high enough to sustain the activity of these species. Other studies have also reported that, for reptiles currently experiencing suboptimal temperatures, climate change could facilitate performance (Cabezas-Cartes et al., 2019; Gómez-Alés et al., 2018; Guerra-Correa et al., 2020). While these results may be promising, there are still several factors that future research needs to consider for a more robust assessment of the effects of climate change. First of all, there is evidence that high temperatures that benefit performance can also affect sperm morphology and viability in reptiles (Quintero-Pérez et al., 2023; Tourmente et al., 2011; Uriostegui Escoto et al., 2025). This is a very relevant trade-off, since reproductive impediments can condition the survival of the species, even if the conditions for other physiological processes are favorable. Reptiles can overcome this problem through behavioral thermoregulation (Domínguez-Godoy et al., 2024), but it is important to adequately address the thermal sensitivity of sperm quality in these species.
It is also important to note that our estimations are based entirely on the thermal landscape, but precipitation regimes and humidity can also be of importance for the maintenance of these species, especially for T. melanogaster. Water temperature will likely increase in many freshwater systems around the globe (Van Vliet et al., 2013) and while the wide thermal preferences of T. melanogaster might suggest it has the ability to cope with these changes, a direct studio on the matter is necessary. Furthermore, water quality may decrease in the future (Whitehead et al., 2009; Xia et al., 2015), so the examination of potential relations between physiological performance and water quality should be addressed, Another aspect that should be considered in future research is to determine whether there is geographic variation in the attributes of the thermal ecology and physiology of these species. There are studies that have reported that thermal ecology attributes are very similar, not only within species, but also among closely related species (Díaz de la Vega-Pérez et al., 2013; Fierro-Estrada et al., 2019; Mueller and Gienger, 2019; Weatherhead et al., 2012). However, some species living in more challenging environments exhibit variation in thermoregulatory patterns or metabolic rates along altitudinal or latitudinal gradients (Jaramillo-Alba et al., 2020; Plasman et al., 2020, 2025; Rivera-Rea et al., 2023; Stellatelli et al., 2022). Since our three species have wide distributions, a more extensive collection of physiological data from environmentally different sites could strengthen our estimates.
In summary, this study shows that future climates will increase the availability of favorable thermal conditions for the physiological performance of our studied species. The mountainous landscape of Mexico provides many suitable microhabitats for these species, a trend that could continue in the future. These results can complement recent efforts to incorporate physiological studies into conservation endeavours. The role of physiology as a formal support for conservation is relatively recent, providing useful information to conservation topics like understanding how species respond to environmental change or the development of adequate management strategies (Cooke & O’Connor, 2010; Madliger et al., 2018). However, translating physiological data to a geographic scale is complicated. In this sense, the determination of favorable (or unsuitable) areas for physiological performance becomes very relevant, since this information could outline conservation efforts like selecting ideal regions for new natural protected areas, the evaluation of existing ones, or the identification of candidate areas for restoration. This is a valuable first step to further conservation strategies in some of the most important regions in Mexico for reptile biodiversity.
Data availability statement
The datasets generated for this study can be found in the Zenodo repository: https://doi.org/10.5281/zenodo.17527172.
Ethics statement
The animal study was approved by Comité de Ética, Facultad de Estudios Superiores Iztacala. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
RF: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Visualization, Writing – original draft. HP: Conceptualization, Funding acquisition, Investigation, Project administration, Resources, Supervision, Validation, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This work was supported by the “Secretaría de Ciencia, Humanidades, Tecnología e Innovación (SECIHTI)” through the Scholarship No.775220.
Acknowledgments
We thank David Prieto-Torres, Cristian Hernández-Herrera and Marco Solis-Sotelo for their comments and guidance on certain parts of the methods. This article is a requirement of RF-H for obtaining the degree of ‘Doctor en Ciencias’, issued by Posgrado en Ciencias Biológicas, UNAM.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/famrs.2025.1711665/full#supplementary-material
References
Angilletta M. J. Jr. (2006). Estimating and comparing thermal performance curves. J. Thermal Biol. 31, 541–545. doi: 10.1016/j.jtherbio.2006.06.002
Årevall J., Early R., Estrada A., Wennergren U., and Eklöf A. C. (2018). Conditions for successful range shifts under climate change: The role of species dispersal and landscape configuration. Diversity Distrib 24, 1598–1611. doi: 10.1111/ddi.12793
Arnaud G., Sandoval S., Escobar-Flores J. G., Escobar-Flores J. G., Gomez-Muñoz V. M., and Buguete J. L. (2019). Thermal ecology of the rattlesnake Crotalus catalinensis from Santa Catalina Island, Gulf of California. Acta Universitaria 28, 39–46. doi: 10.15174/au.2018.1667
Aubret F. and Shine R. (2010). Thermal plasticity in young snakes: how will climate change affect the thermoregulatory tactics of ectotherms? J. Exp. Biol. 213, 242–248. doi: 10.1242/jeb.035931
Bryson R. W., Murphy R. W., Lathrop A., and Lazcano-Villareal D. (2011). Evolutionary drivers of phylogeographical diversity in the highlands of Mexico: A case study of the Crotalus triseriatus species group of montane rattlesnakes. J. Biogeogr. 38, 697–710. doi: 10.1111/j.1365-2699.2010.02431.x
Buckley L. B., Urban M. C., Angilletta M. J. Jr., Crozier L. G., Rissler L. J., and Sears M. W. (2010). Can mechanism inform species’ distribution models? Ecol. Lett. 13, 1041–1054. doi: 10.1111/j.1461-0248.2010.01479.x
Bulté G. and Blouin-Demers G. (2006). Cautionary notes on the descriptive analysis of performance curves in reptiles. J. Thermal Biol. 31, 287–291. doi: 10.1016/j.jtherbio.2005.11.030
Cabezas-Cartes F., Fernández J. B., Duran F., and Kubisch E. L. (2019). Potential benefits from global warming to the thermal biology and locomotor performance of an endangered Patagonian lizard. PeerJ 7, e7437. doi: 10.7717/peerj.7437
Camacho A., Pavao R., Moreira C. N., Pinto A. C. B. F., Navas C. A., and Rodrigues M. T. (2015). Interaction of morphology, thermal physiology and burrowing performance during the evolution of fossoriality in Gymnophthalmini lizards. Funct. Ecol. 29, 515–521. doi: 10.1111/1365-2435.12355
Campbell J. A. and Lamar W. W. (2004). The venomous reptiles of the Western Hemisphere (Itahaca, NY: Cornell University Press).
Carrell J. D., Phinney A. I., Mueller K., and Bean B. (2023). Multiscale ecological niche modeling exhibits varying climate change impacts on habitat suitability of Madrean Pine-Oak trees. Front. Ecol. Evol. 11. doi: 10.3389/fevo.2023.1086062
Ceia-Hasse A., Sinervo B., Vicente L., and Pereira H. M. (2014). Integrating ecophysiological models into species distribution projections of European reptile range shifts in response to climate change. Ecography 37, 679–688. doi: 10.1111/j.1600-0587.2013.00600.x
Cisneros-Bernal A. Y., Rodríguez-Gómez F., Flores-Villela Ó., Fujita M. K., and Velasco J. A. (2022). Phylogeography supports lineage divergence for an endemic rattlesnake (Crotalus ravus) of the Neotropical montane forest in the Trans-Mexican Volcanic Belt. Biol. J. Linn. Soc. 137, 496–512. doi: 10.1093/biolinnean/blac066
Cooke S. J. and O’Connor C. M.(2010).Making conservation physiology relevant to policy makers and conservation practitioners. Conserv. Lett. 3, 159–166. doi: 10.1111/j.1755-263X.2010.00109.x
Díaz de la Vega-Pérez A. H., Jiménez-Arcos V. H., Manríquez-Morán N. L., and Méndez-de la Cruz F. R. (2013). Conservatism of thermal preferences between parthenogenetic Aspidoscelis cozumela complex (Squamata: Teiidae) and their parental species. Herpetol. J. 23, 93–104.
Dinerstein E., Olson D., Joshi A., Vynne C., Burgess N. D., Wikramanayake E., et al. (2017). An ecoregion-based approach to protecting half the terrestrial realm. BioScience 67, 534–545. doi: 10.1093/biosci/bix014
Domínguez-Godoy M. A., Arenas-Ríos E., Uriostegui-Escoto D., Lucio R. A., Díaz de la Vega-Pérez A. H., and Méndez de la Cruz F. R. (2024). Behavioural thermoregulation prevents thermal stress in lizard sperm fertility. Eur. J. Wildlife Res. 70, 113. doi: 10.1007/s10344-024-01864-6
Evans T. G., Diamond S. E., and Kelly M. W. (2015). Mechanistic species distribution modelling as a link between physiology and conservation. Conserv. Physiol. 3, 1–16. doi: 10.1093/conphys/cov056
Fajardo J., Corcoran D., Roehrdanz P. R., Hannah L., and Marquet P. A. (2020). GCM compareR: A web application to assess differences and assist in the selection of general circulation models for climate change research. Methods Ecol. Evol. 11, 656–663. doi: 10.1111/2041-210X.13360
Fick S. E. and Hijmans R. J. (2017). WorldClim 2: new 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 37, 4302–4315. doi: 10.1002/joc.5086
Fierro-Estrada N., González González Y. G., Miles D. B., Martínez-Gómez M., García A., Salgado-Ugarte I. H., et al. (2019). Thermoregulation of the lizard Barisia imbricata at altitudinal extremes. Amphibia Reptilia 40, 349–360. doi: 10.1163/15685381-20191155
Figueroa-Huitrón R., Díaz de la Vega-Pérez A. H., Plasman M., and Pérez-Mendoza H. A. (2024a). Physiological thermal responses of three Mexican snakes with distinct lifestyles. PeerJ 12, e17705. doi: 10.7717/peerj.17705
Figueroa-Huitrón R., Díaz-Martínez E. F., Méndez-de la Cruz F. R., and Pérez-Mendoza H. A. (2024b). Thermoregulation and activity patterns of three species of snakes with different lifestyles in central Mexico. Amphibia Reptilia 45, 319–332. doi: 10.1163/15685381-bja10190
García-Balderas C. M., Quintero-Díaz G. E., Lucio-Palacio C. R., and Valdez-Jiménez D. (2014). Conopsis lineata (lined tolucan ground snake). Diet. Herpetol. Rev. 45, 337.
Goller M., Goller F., and French S. S. (2014). A heterogeneous thermal environment enables remarkable behavioral thermoregulation in Uta stansburiana. Ecol. Evol. 4, 3319–3329. doi: 10.1002/ece3.1141
Gómez-Alés R., Acosta J. C., Astudillo V., Córdoba M., Blanco G. M., and Miles D. B. (2018). Effect of temperature on the locomotor performance of species in a lizard assemblage in the Puna region of Argentina. J. Comp. Physiol. B: Biochem. Syst. Environ. Physiol. 188, 977–990. doi: 10.1007/s00360-018-1185-y
Gómez-Mendoza L. and Arriaga L. (2007). Modeling the effect of climate change on the distribution of oak and pine species of Mexico. Conserv. Biol. 21, 1545–1555. doi: 10.1111/j.1523-1739.2007.00814.x
González-Fernández A., Manjarrez J., García-Vázquez U. O., D’Addario M., and Sunny A. (2018). Present and future ecological niche modeling of garter snake species from the Trans-Mexican Volcanic Belt. PeerJ 2018, 1–20. doi: 10.7717/peerj.4618
Goyenechea I. and Flores-Villela Ó. (2006). Taxonomic summary of Conopsis, Günther 1858 (Serpentes: Colubridae). Zootaxa 27, 1–27. doi: 10.11646/ZOOTAXA.1271.1.1
Guerra-Correa E. S., Merino-Viteri A., Andrango M. B., and Torres-Carvajal O. (2020). Thermal biology of two tropical lizards from the Ecuadorian Andes and their vulnerability to climate change. PloS One 15, e0228043. doi: 10.1371/journal.pone.0228043
Hernández-Salinas U., Cruz-Elizalde R., Ramírez-Bautista A., Wilson L. D., Berriozabal-Islas C., Johnson J. D., et al. (2023). Taxonomic and functional diversity of the amphibian and reptile communities of the state of Durango, Mexico. Community Ecol. 24, 229–242. doi: 10.1007/s42974-023-00145-7
Hertz P. E., Huey R. B., and Stevenson R. D. (1993). Evaluating temperature regulation by field-active ectotherms: the fallacy of the inappropriate question. Am. Nat. 142, 796–818. doi: 10.1086/285573
Hijmans R. (2024). terra: Spatial Data Analysis (R package version 1.8-6). Available online at: https://rspatial.github.io/terra/.
Huey R. B., Kearney M. R., Krockenberger A. K., Holtum J. A. M., Jess M., and Williams S. E. (2012). Predicting organismal vulnerability to climate warming: Roles of behaviour, physiology and adaptation. Philos. Trans. R. Soc. B: Biol. Sci. 367, 1665–1679. doi: 10.1098/rstb.2012.0005
IUCN (2024). The IUCN Red List of Threatened Species. Version 2024-1. Available online at: https://www.iucnredlist.org (Accessed May, 2024).
Jaramillo-Alba J. L., Díaz de la Vega-Pérez A. H., Bucio-Jiménez L. E., Méndez-de la Cruz F. R., and Pérez-Mendoza H. A. (2020). Comparative thermal ecology parameters of the mexican dusky rattlesnake (Crotalus triseriatus). J. Thermal Biol. 92, 102695. doi: 10.1016/j.jtherbio.2020.102695
Kearney M. R. and Porter W. P. (2017). NicheMapR – an R package for biophysical modelling: the microclimate model. Ecography 40, 664–674. doi: 10.1111/ecog.02360
Kearney M. R. and Porter W. P. (2020). NicheMapR – an R package for biophysical modelling: the ectotherm and Dynamic Energy Budget models. Ecography 43, 85–96. doi: 10.1111/ecog.04680
Kearney M. R., Shine R., and Porter W. P. (2009). The potential for behavioral thermoregulation to buffer “cold-blooded” animals against climate warming. Proc. Natl. Acad. Sci. 10, 3835–3840. doi: 10.1073/pnas.0808913106
Lourdais O., Guillon M., DeNardo D. F., and Blouin-Demers G. (2013). Cold climate specialization: adaptive covariation between metabolic rate and thermoregulation in pregnant vipers. Physiol. Behav. 119, 149–155. doi: 10.1016/j.physbeh.2013.05.041
Mackessy S. P., Leroy J., Mociño-Deloya E., Setser K., Bryson R. W., and Saviola A. J. (2018). Venom ontogeny in the mexican lance-headed rattlesnake (Crotalus polystictus). Toxins 10, 271. doi: 10.3390/toxins10070271
Madliger C. L., Love O. P., Hultine K. R., and Cooke S. J.(2018). The conservation physiology toolbox: Status and opportunities. Conserv. Physiol. 6, 1–16. doi: 10.1093/conphys/coy029
Manjarrez J., García C. M., and Drummond H. (2013). Variation in the diet of the Mexican black-bellied gartersnake Thamnophis melanogaster: importance of prey availability and snake body size. J. Herpetol. 47, 413–420. doi: 10.1670/12-174
Manjarrez J., Macías Garcia C., and Drummond H. (2017). Morphological convergence in a Mexican garter snake associated with the ingestion of a novel prey. Ecol. Evol. 7, 7178–7186. doi: 10.1002/ece3.3265
Manjarrez J. and San-Roman-Apolonio E. (2015). Timing of birth and body condition in neonates of two gartersnake species from central México. Herpetologica 71, 12–18. doi: 10.1655/HERPETOLOGICA-D-13-00098
Manrique-Ascencio A., Prieto-Torres D. A., Villalobos F., and Guevara R. (2024). Climate-driven shifts in the diversity of plants in the Neotropical seasonally dry forest: Evaluating the effectiveness of protected areas. Global Change Biol. 30, e17282. doi: 10.1111/gcb.17282
Marinho P. S., Chinchilla J. E. O., Braz H. B., and Almeida-Santos S. M. (2022). Effects of pregnancy on the body temperature of the South American Rattlesnake, Crotalus durissus, in southeastern Brazil. South Am. J. Herpetol. 23, 67–74. doi: 10.2994/SAJH-D-18-00081.1
Mastretta-Yanes A., Moreno-Letelier A., Piñero D., Jorgensen T. H., and Emerson B. C. (2015). Biodiversity in the Mexican highlands and the interaction of geology, geography and climate within the Trans-Mexican Volcanic Belt. J. Biogeogr. 42, 1586–1600. doi: 10.1111/jbi.12546
McConnachie S., Greene S. N., and Perrin M. R.(2011).Thermoregulation in the semi-aquatic yellow anaconda, Eunectes notaeus. J. Therm. Biol. 36, 71–77. doi: 10.1016/j.jtherbio.2010.11.005
Meik J. M., Streicher J. W., Mociño-Deloya E., Setser K., and Lazcano D. (2012). Shallow phylogeographic structure in the declining Mexican lance-headed rattlesnake, Crotalus polystictus (Serpentes: Viperidae). Phyllomedusa 11, 3–11. doi: 10.11606/issn.2316-9079.v11i1p3-11
Mizsei E., Radovics D., Rák G., Budai M., Bancsik B., Szabolcs M., et al. (2024). Alpine viper in changing climate: thermal ecology and prospects of a cold-adapted reptile in the warming Mediterranean. Sci. Rep. 14, 18988. doi: 10.1038/s41598-024-69378-4
Morrone J. J., Escalante T., Rodriguez-Tapia G., Carmona A., Arana M., and Mercado-Gomez J. D. (2022). Biogeographic regionalization of the Neotropical region: New map and shapefile. Anais Da Academia Bras. Cienc. 94, e20211167. doi: 10.1590/0001-3765202220211167
Mueller A. and Gienger C. M. (2019). Comparative thermal ecology of copperheads (Agkistrodon contortrix) and cottonmouths (Agkistrodon piscivorus). J. Thermal Biol. 79, 73–80. doi: 10.1016/j.jtherbio.2018.12.009
New M., Lister D., Hulme M., and Makin I. (2002). A high-resolution data set of surface climate over global land areas. Climate Res. 21, 1–25. doi: 10.3354/cr021001
Plasman M., Bautista A., McCUE M. D., and Díaz de la Vega-Pérez A. H. (2020). Resting metabolic rates increase with elevation in a mountain-dwelling lizard. Integr. Zool. 15, 363–374. doi: 10.1111/1749-4877.12434
Plasman M., Gonzalez-Voyer A., Bautista A., and Díaz de la Vega-Pérez A. H. (2025). Flexibility in thermal requirements: a comparative analysis of the wide-spread lizard genus Sceloporus. Integr. Zool. 20, 850–866. doi: 10.1111/1749-4877.12860
Quintero-Pérez R. I., Mendez-de la Cruz F. R., Miles D. B., Chávez M. C. V., Lopez-Ramirez Y., Arenas-Moreno D. M., et al. (2023). Trade-off between thermal preference and sperm maturation in a montane lizard. J. Thermal Biol. 113, 103526. doi: 10.1016/j.jtherbio.2023.103526
Ramírez-Arce D. G., Ochoa-Ochoa L. M., Lira-Noriega A., and Martorell C. (2025). Reptile diversity patterns under climate and land use change scenarios in a subtropical montane landscape in Mexico. J. Biogeogr. 52, 108–121. doi: 10.1111/jbi.15017
Ramírez-Bautista A. and Cruz-Elizalde R. (2013). Reptile community structure in two fragments of cloud forest of the Sierra Madre Oriental, Mexico. North-Western J. Zool. 9, 410–17.
Ramírez-Bautista A., Hernández-Salinas U., García-Vázquez U. O., Leyte-Manrique A., and Canseco-Márquez L. (2009). Herpetofauna del Valle de México: diversidad y conservación (1a edición) (Universidad Autónoma del Estado de Hidalgo).
R Core Team (2024). R: A language and environment for statistical computing (Vienna, Austria: R Foundation for Statistical Computing). Available online at: https://www.R-project.org/.
Rivera-Rea J., Macotela L., Moreno-Rueda G., Suárez-Varón G., Bastiaans E., Quintana E., et al. (2023). Thermoregulatory behavior varies with altitude and season in the sceloporine mesquite lizard. J. Thermal Biol. 114, 103539. doi: 10.1016/j.jtherbio.2023.103539
Rossman D. A., Ford N. B., and Seigel R. A. (1996). The garter snakes: evolution and ecology. 1st ed (Oklahoma, EUA: University of Oklahoma Press).
Rubio-Blanco T., Martínez-Díaz-González R., Heredia-Bobadilla R. L., Guido-Patiño J. C., Arenas S., Caballero-Viñas C., et al. (2024). Predicting the effects of climate and land use changes on small rattlesnakes in central Mexico: Insights for conservation planning. J. Nat. Conserv. 79, 126607. doi: 10.1016/j.jnc.2024.126607
Sánchez-Fernández D., Rizzo V., Cieslak A., Faille A., Fresneda J., and Ribera I. (2016). Thermal niche estimators and the capability of poor dispersal species to cope with climate change. Sci. Rep. 6, 23381. doi: 10.1038/srep23381
Sánchez-López K. E., Martín-Regalado C. N., Gómez-Ugalde R. M., and Martínez-Ramírez E. (2025). Diversity of amphibians and reptiles in conserved forests and disturbed areas: A comparison in Southern Mexico. Diversity 17, 141. doi: 10.3390/d17030141
Sears M. W. and Angilletta M. J. Jr. (2015). Costs and benefits of thermoregulation revisited: Both the heterogeneity and spatial structure of temperature drive energetic costs. Am. Nat. 185, E94–E102. doi: 10.1086/680008
Seebacher F., Narayan E., Rummer J. L., Tomlinson S., and Cooke S. J. (2023). How can physiology best contribute to wildlife conservation in a warming world? Conserv. Physiol. 11, coad038. doi: 10.1093/conphys/coad038
Setser K., Mociño-Deloya E., Pleguezuelos J. M., Lazcano D., and Kardon A. (2010). Reproductive ecology of female Mexican lance-headed rattlesnakes. J. Zool. 281, 175–182. doi: 10.1111/j.1469-7998.2010.00692.x
Sinervo B., Méndez-de la Cruz F. R., Miles D. B., Heulin B., Bastiaans E., Villagrán-Santa Cruz M., et al. (2010). Erosion of lizard diversity by climate change and altered thermal niches. Science 328, 894–899. doi: 10.1126/science.1184695
Siyum Z. G. (2020). Tropical dry forest dynamics in the context of climate change: syntheses of drivers, gaps, and management perspectives. Ecol. Processes 9, 25. doi: 10.1186/s13717-020-00229-6
Stellatelli O. A., Vega L. E., Block C., Rocca C., Bellagamba P., Dajil J. E., et al. (2022). Latitudinal pattern of the thermal sensitivity of running speed in the endemic lizard Liolaemus multimaculatus. Integr. Zool. 17, 619–637. doi: 10.1111/1749-4877.12579
Sunday J. M., Bates A. E., Kearney M. R., Colwell R. K., Dulvy N. K., Longino J. T., et al. (2014). Thermal-safety margins and the necessity of thermoregulatory behavior across latitude and elevation. Proc. Natl. Acad. Sci. United States America 111, 5610–5615. doi: 10.1073/pnas.1316145111
Sunny A., Manjarrez J., Caballero-Viñas C., Bolom-Huet R., Gómez-Ortiz Y., Domínguez-Vega H., et al. (2023). Modelling the effects of climate and land-cover changes on the potential distribution and landscape connectivity of three earth snakes (Genus Conopsis, Günther 1858) in central Mexico. Sci. Nat. 110, 52. doi: 10.1007/s00114-023-01880-7
Taylor E. N., Diele-Viegas L. M., Gangloff E. J., Hall J. M., Halpern B., Massey M. D., et al. (2021). The thermal ecology and physiology of reptiles and amphibians: A user’s guide. J. Exp. Zool. Part A: Ecol. Integr. Physiol. 335, 13–44. doi: 10.1002/jez.2396
Tomlinson S. (2019). The mathematics of thermal sub-optimality: Nonlinear regression characterization of thermal performance of reptile metabolic rates. J. Thermal Biol. 81, 49–58. doi: 10.1016/j.jtherbio.2019.02.008
Tourmente M., Giojalas L. C., and Chiaraviglio M. (2011). Sperm parameters associated with reproductive ecology in two snake species. Herpetologica 67, 58–70. doi: 10.1655/HERPETOLOGICA-D-10-00052.1
Uriostegui Escoto D., Méndez de la Cruz F. R., García Lorenzana M., López Ramírez Y., and Arenas Ríos E. (2025). Temperatures lower than preferred ones maintain DNA integrity and sperm quality of Lepidophyma gaigeae (SQUAMATA: XANTUSIIDAE). Animals 15, 1784. doi: 10.3390/ani15121784
van Heerwaarden B., Kellermann V., and Sgrò C. M. (2016). Limited scope for plasticity to increase upper thermal limits. Funct. Ecol. 30, 1947–1956. doi: 10.1111/1365-2435.12687
Van Vliet M. T., Franssen W. H., Yearsley J. R., Ludwig F., Haddeland I., Lettenmaier D. P., et al. (2013). Global river discharge and water temperature under climate change. Global. Environ. Change 23, 450–464. doi: 10.1016/j.gloenvcha.2012.11.002
Venegas-Barrera C. S., Sunny A., and Manjarrez J. (2025). Thermal ecology of the Mexican Garter Snake (Thamnophis eques): temporal and spatial variations. PeerJ 13, e18641. doi: 10.7717/peerj.18641
Vicenzi N., Kubisch E. L., Ibargüengoytía N. R., and Corbalán V. (2019). Thermal sensitivity of performance of Phymaturus palluma (Liolaemidae) in the highlands of Aconcagua: vulnerability to global warming in the Andes. Amphibia-Reptilia 40, 207–218. doi: 10.1163/15685381-20181072
Weatherhead P. J., Sperry J. H., Carfagno G. L. F., and Blouin-Demers G. (2012). Latitudinal variation in thermal ecology of North American ratsnakes and its implications for the effect of climate warming on snakes. J. Thermal Biol. 37, 273–281. doi: 10.1016/j.jtherbio.2011.03.008
Whitehead P. G., Wilby R. L., Battarbee R. W., Kernan M., and Wade A. J. (2009). A review of the potential impacts of climate change on surface water quality. Hydrol. Sci. J. 54, 101–123. doi: 10.1623/hysj.54.1.101
Xia X. H., Wu Q., Mou X. L., and Lai Y. J. (2015). Potential impacts of climate change on the water quality of different water bodies. J. Environ. Inf. 25, 85–98. doi: 10.3808/jei.201400263
Youngblood J. P., Cease A. J., Talal S., Copa F., Medina H. E., Rojas J. E., et al. (2023). Climate change expected to improve digestive rate and trigger range expansion in outbreaking locusts. Ecol. Monogr. 93, e1550. doi: 10.1002/ecm.1550
Keywords: climate change, ecophysiology, mechanistic models, thermal ecology, snakes, performance breadth
Citation: Figueroa-Huitrón R and Pérez-Mendoza HA (2025) Climate change may improve physiological suitability for three widespread Mexican snakes. Front. Amphib. Reptile Sci. 3:1711665. doi: 10.3389/famrs.2025.1711665
Received: 23 September 2025; Accepted: 10 November 2025; Revised: 04 November 2025;
Published: 26 November 2025.
Edited by:
Maria Delia Basanta, National Polytechnic Institute of Mexico (CINVESTAV), MexicoReviewed by:
René Bolom-Huet, Universidad Autónoma del Estado de México, MexicoAldo Gomez Benitez, Universidad Autónoma Metropolitana-Lerma (UAM-L), Mexico
Carlos Luna Aranguré, National Autonomous University of Mexico, Mexico
Copyright © 2025 Figueroa-Huitrón and Pérez-Mendoza. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hibraim Adán Pérez-Mendoza, aGlicmFpbXBlcmV6QHVuYW0ubXg=