 Leandro Castañeyra-Ruiz1*
Leandro Castañeyra-Ruiz1* Ibrahim González-Marrero2Luis H. García-Abad2Emilio Gonzalez-Arnay2Marcial Camacho3Emilia Ma Carmona-Calero2,4Seunghyun Lee1Celine Thao-Quyen Tran1Brian W. Hanak1,5Michael Muhonen1,5Agustín Castañeyra-Perdomo2,4
Ibrahim González-Marrero2Luis H. García-Abad2Emilio Gonzalez-Arnay2Marcial Camacho3Emilia Ma Carmona-Calero2,4Seunghyun Lee1Celine Thao-Quyen Tran1Brian W. Hanak1,5Michael Muhonen1,5Agustín Castañeyra-Perdomo2,4- 1Children’s Hospital of Orange County Research Institute, Orange, CA, United States
- 2Unidad de Anatomía, Departamento de Ciencias Médicas Básicas, Facultad de Medicina, Universidad de La Laguna, San Cristóbal de La Laguna, Spain
- 3Unidad de Farmacología, Departamento de Medicina Física y Farmacología, Facultad de Medicina Universidad de La Laguna, San Cristóbal de La Laguna, Spain
- 4Instituto de Investigación y Ciencias de Puerto del Rosario, Puerto del Rosario, Spain
- 5Department of Neurological Surgery, University of California Irvine, Orange, CA, United States
Glioblastoma (GBM) progression is linked to aquaporin-4 (AQP4), whose functions extend beyond water transport to influence perivascular architecture, immune modulation, edema, and treatment response. In the healthy brain, AQP4 is highly polarized at astrocytic endfeet, supporting perivascular fluid exchange and glymphatic clearance. In GBM, AQP4 is frequently upregulated and mislocalized, correlating with blood–brain barrier (BBB) disruption, impaired directional fluid movement, and peritumoral edema. Peritumoral astrocytic mislocalization of AQP4, together with tumor mass effect, compromises glymphatic function by distorting perivascular spaces and compressing cerebrospinal fluid (CSF)-Interstitial fluid (ISF) exchange zones. We review evidence that AQP4 isoforms (M1 vs. M23) differentially shape motility and membrane organization, and we outline how AQP4-linked signaling axes (e.g., indoleamine 2,3-dioxygenase 1 (IDO1)/tryptophan 2,3-dioxygenase (TDO)-kynurenine–aryl hydrocarbon receptor (AhR) can bias pro-invasive states and immunosuppressive niches enriched with M2-like macrophages). We integrate a four-zone perivascular framework to localize where GBM most perturbs periarterial and perivenous pathways, as well as meningeal lymphatic outflow. Finally, we discuss therapeutic directions spanning AQP4 modulation, isoform balance, and BBB-bypassing delivery strategies. Overall, AQP4 emerges as a mechanistic hub connecting BBB instability, glymphatic impairment, edema, immune evasion, and invasion in GBM.
Introduction
The Glymphatic system (GS) is a brain-wide fluid clearance network responsible for the exchange of CSF and ISF. It facilitates the removal of metabolic waste and contributes to fluid homeostasis and solute trafficking within the central nervous system (CNS) (Xu et al., 2022). The GS comprises a multi-segmented anatomical continuum, including periarterial influx pathways, CSF–ISF exchange zones, and perivenous efflux routes that ultimately drain into the meningeal lymphatic vessels (MLVs) (Xu et al., 2022; Klostranec et al., 2021; Jessen et al., 2015; Hablitz and Nedergaard, 2021). Central to the function of this system is aquaporin-4 (AQP4), a water channel protein abundantly expressed at astrocytic endfeet, which enables directional fluid movement along perivascular routes (Plog and Nedergaard, 2018; Gomolka et al., 2023).
Aquaporins have increasingly been associated with the pathophysiology of several conditions, including hydrocephalus and systematic hypertension, since these pathologies have characteristic nervous system water transport alterations (Castaneyra-Ruiz et al., 2016a; Gonzalez-Marrero et al., 2022; Castaneyra-Ruiz et al., 2013; Castaneyra-Ruiz et al., 2016b; Castaneyra-Ruiz et al., 2019; Carmona-Calero et al., 2015); however, beyond its canonical role, specially AQP4 has emerged as an astrocytic lineage marker (Castaneyra-Ruiz et al., 2022a; Castaneyra-Ruiz et al., 2022b; Holst et al., 2019) and a mediator of diverse astrocytic functions, including cellular migration, cytoskeletal remodeling and microenvironmental interactions. These functions are often associated with the dominant AQP4-isoforms expressed in an astrocyte (Pisani et al., 2021; Smith et al., 2014) (Figure 1). These non-canonical roles of AQP4 have generated increasing interest in its contribution to neuropathological states, particularly brain tumors of glial origin, including gliomas and astrocytomas.
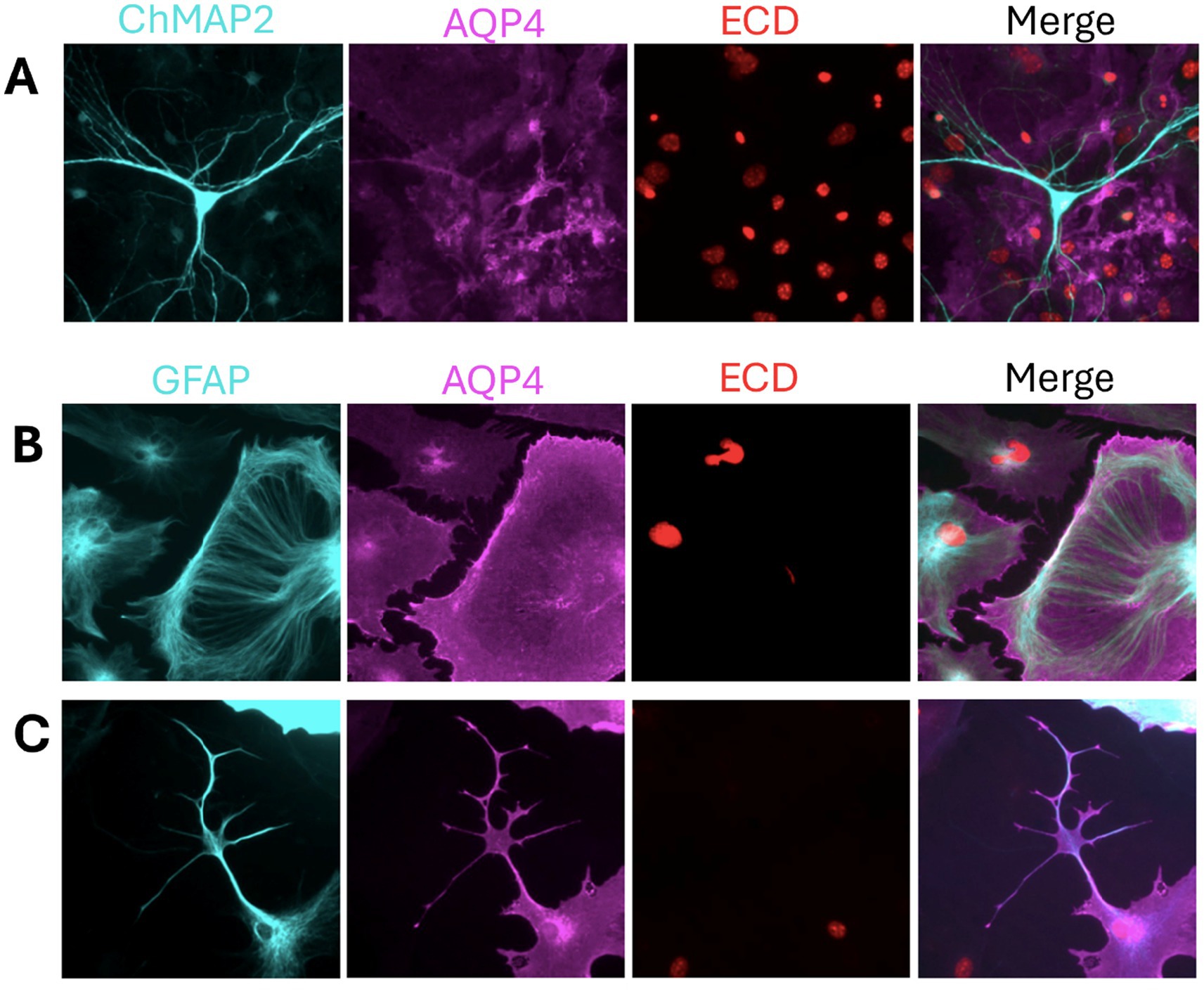
Figure 1. AQP4 glia lineage marker and indicator of morphological diversity. (A) Mixed neuron–astrocyte cultures stained with ChMAP2 (neurons, cyan), AQP4 (magenta), and ECD (nuclei, red) show AQP4 expression restricted to astrocytes, confirming its cell-type specificity. (B,C) Astrocyte monocultures stained with GFAP (cyan), AQP4, and ECD reveal distinct morphologies. (B) Cells with broad lamellipodia and radial GFAP arrays reflect a migratory phenotype, potentially linked to M1-AQP4. (C) Stellate astrocytes with membrane-enriched AQP4 suggest a stabilized phenotype, consistent with M23-AQP4 and OAP formation. Samples were fixed with 4% PFA, permeabilized with 0.1% Tween-20, and stained using anti-mouse GFAP (rhodamine red) and anti-rabbit AQP4 (alexa fluor 488). Unpublished image from our laboratory.
Glioblastoma multiforme (GBM) is the most common and lethal form of primary brain tumor in adults, characterized by diffuse infiltration, genetic heterogeneity, and limited responsiveness to current therapies (Batash et al., 2017; Ghosh et al., 2018). Despite multimodal treatment involving surgery, radiation, and chemotherapy, the median survival remains dismal—approximately 15 months (Alifieris and Trafalis, 2015; Wu et al., 2021; Pearson et al., 2020). This poor prognosis is largely attributed to the tumor’s aggressive invasion into surrounding brain parenchyma, its ability to resist conventional therapies, and its manipulation of the immune and fluid microenvironments (van Linde et al., 2017; Yalamarty et al., 2023).
Importantly, AQP4 has been implicated in several aspects of GBM progression, including enhanced cell migration, resistance to apoptosis, disruption of the blood–brain barrier (BBB), and the polarization of tumor-associated macrophages (TAMs) toward an immunosuppressive phenotype (Amiry-Moghaddam, 2019; Simone et al., 2019; Lan et al., 2022). These functions associate AQP4 with tumor-promoting functions.
This review provides a comprehensive synthesis of current knowledge linking glymphatic anatomy with GBM pathophysiology and AQP4 dysfunction. We propose a refined anatomical model of the glymphatic system, including four functional zones, and evaluate the isoform-specific contributions of AQP4 to glioma cell behavior. By integrating molecular, anatomical, and clinical insights, we aim to define the AQP4–glymphatic axis as a critical node in GBM progression and a potential target for future diagnostic and therapeutic strategies.
Anatomical organization of the glymphatic system
The GS facilitates directional CSF influx, ISF exchange, and metabolic waste clearance from the brain parenchyma (Xu et al., 2022; Iliff et al., 2012). Traditionally divided into three segments—periarterial influx (S1), CSF–ISF exchange (S2), and perivenous efflux (S3) (Xu et al., 2022). Here, we propose a fourth anatomical segment (S4), corresponding to the meningeal perivenous drainage compartment, which is structurally and functionally distinct from its parenchymal counterpart, as described below.
S1: Periarterial influx region
This region surrounds arteries that penetrate the brain from the subarachnoid space. The Virchow–Robin space (VRS), located between the arterial vascular adventitia (AVA) and the pia mater (PM), forms the main conduit for CSF entry into the brain interstitium (Hablitz and Nedergaard, 2021). This segment establishes the foundation of glymphatic flow (Figure 2).
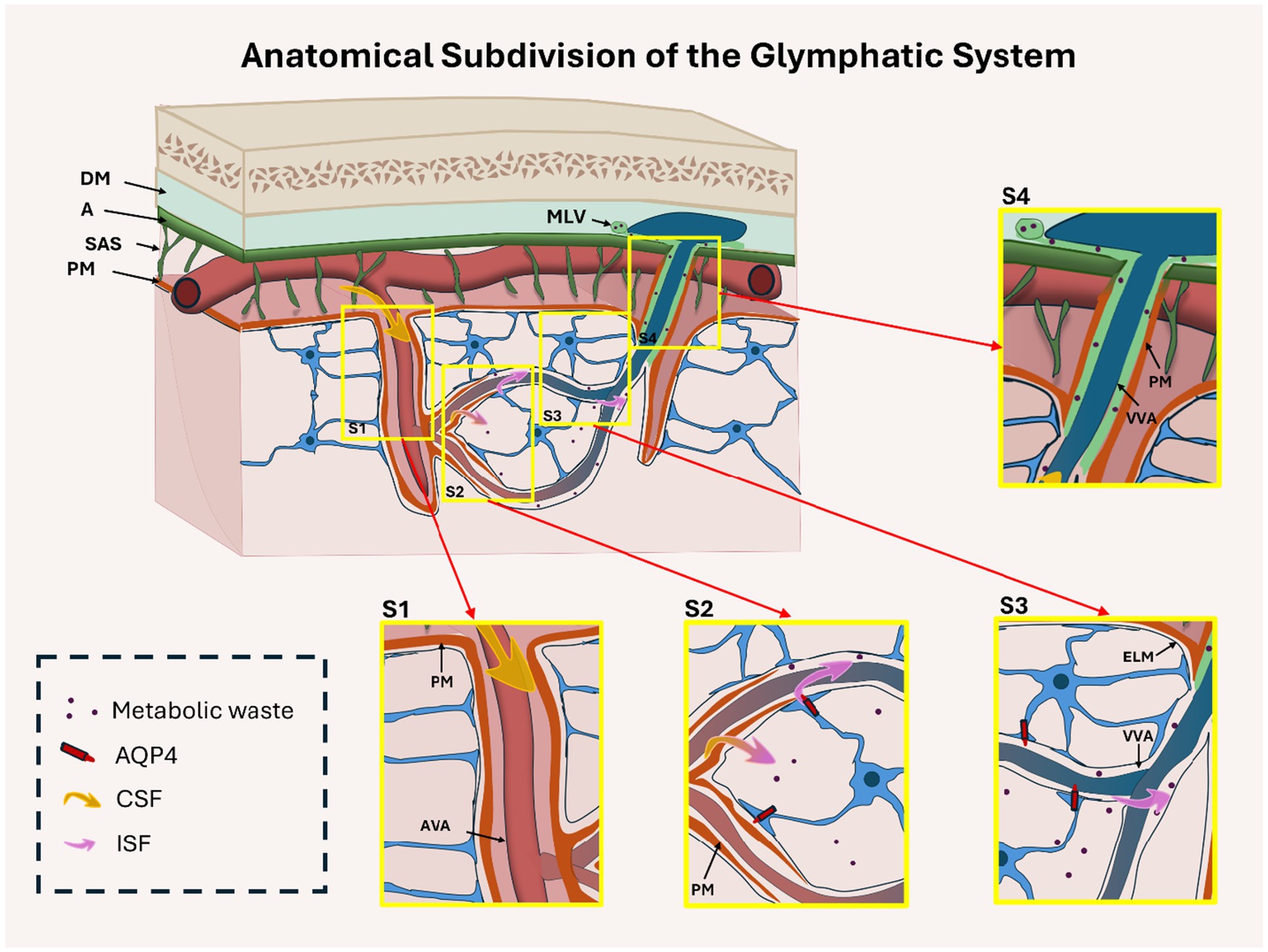
Figure 2. Anatomical organization of the glymphatic system, subdivided into four functional regions: periarterial influx (S1), CSF–interstitial fluid (ISF) exchange (S2), perivenous parenchymal efflux (S3), and meningeal efflux (S4). This cross-sectional diagram shows the brain cortex, subarachnoid space (SAS), and meninges (dura mater [DM], arachnoid [A], pia mater [PM]). In S1, CSF (yellow arrows) enters periarterial spaces through the Virchow-Robin space, located between the arterial vascular adventitia (AVA) and the PM. In S2, the PM becomes discontinuous, enabling CSF–ISF exchange mediated by astrocytic AQP4 channels. In S3, glymphatic fluid flows along perivenous spaces between the external limiting membrane (ELM) and venous vascular adventitia (VVA) within the parenchyma. In S4, flow continues in the meningeal compartment between the VVA and PM toward meningeal lymphatic vessels (MLV). AQP4 is relevant only in the parenchymal segments (S2–S3), but not in S1 or S4. Insets S1–S4 highlight key anatomical features of each region.
S2: CSF–ISF exchange zone
Here, the pia mater thins and gradually disappears, allowing for the direct interaction of CSF with the interstitial compartment. In this section, AQP4, highly expressed at the astrocytic endfeet, facilitates bidirectional water transport between CSF and ISF, making this region central to solute exchange and fluid balance (Gomolka et al., 2023; Giannetto et al., 2024) (Figure 2).
S3: Perivenous efflux region (parenchymal)
As glymphatic flow transitions to venous outflow, CSF–ISF passes along perivenous spaces bordered by the external limiting membrane (ELM) and venous vascular adventitia (VVA). AQP4 expression persists in this segment, enabling waste clearance toward the brain surface (Rasmussen et al., 2018; Iliff et al., 2015). Structural disruption of this segment is frequently observed in pathological conditions, including GBM (Figure 2).
S4: Perivenous efflux region (meningeal)
Beyond the parenchyma, fluid continues along subarachnoid perivenous spaces bordered by the VVA and PM, eventually draining into the meningeal lymphatic vessels (MLVs). AQP4 is not functionally involved in this segment (Figure 2). However, anatomical alterations in this zone—such as compression or obstruction by tumor masses—may still indirectly influence glymphatic function.
This four-zone anatomical model offers a refined framework for investigating region-specific vulnerabilities in glymphatic transport, particularly in pathologies such as GBM, in which edema and stasis predominantly arise in CSF–ISF exchange (S2) and parenchymal perivenous efflux (S3), whereas diminished out flow affects meningeal lymphatic regions (S4) (Ma et al., 2019).
AQP4 localization and blood–brain barrier integrity
AQP4 not only facilitates glymphatic transport but is also essential for maintaining the structural and functional integrity of the blood–brain barrier (BBB). Under physiological conditions, AQP4 is highly polarized at perivascular astrocytic endfeet, where it mediates bidirectional water flux and contributes to homeostatic exchange between vascular and parenchymal compartments (Warth et al., 2007; Warth et al., 2004; Valente et al., 2024; Salman et al., 2022). Its perivascular localization further supports endothelial tight junction stability and BBB ultrastructure (e.g., astrocytic endfeet abut microvessels, influencing endothelial morphology) (Wolburg et al., 2012; Mueller et al., 2023). The BBB is anatomically adjacent to the perivascular spaces engaged by glymphatic flow. Therefore, AQP4 at the astrocyte provides a molecular interface for exchange, between vascular and perivascular/glymphatic compartments. Alterations in AQP4 expression, or loss of perivascular polarity can disturb endfoot–vascular coupling, degrade tight-junction integrity, increase BBB permeability, and exacerbate vasogenic edema in injury or disease states (Mueller et al., 2023; Jeon et al., 2021; Zhou et al., 2008). In GBM, loss of AQP4 polarity and disruption of its perivascular organization correlate with increased blood–brain barrier permeability, as demonstrated by elevated sodium fluorescein leakage and higher MRI-derived edema indices in patient specimens (Valente et al., 2022; Solar et al., 2022; Abbrescia et al., 2024). This breakdown allows plasma proteins and water to enter the brain parenchyma, resulting in vasogenic edema, and perivascular disorganization (Wolburg et al., 2012) that in turn impairs glymphatic clearance (Valente et al., 2022; Gao et al., 2024)—two coupled yet separable processes (Warth et al., 2007; Warth et al., 2004; Valente et al., 2024; Wolburg et al., 2012; Noell et al., 2012).
Mechanistically, the mislocalization of AQP4 is associated with the degradation of anchoring proteins such as agrin and components of the dystrophin–dystroglycan complex, which are responsible for tethering AQP4 to the astrocytic endfoot membrane (Warth et al., 2004; Wolburg et al., 2012; Noell et al., 2012). The detachment of AQP4 from these complexes not only disrupts water channel function but also undermines the scaffold supporting the BBB (Figure 3).
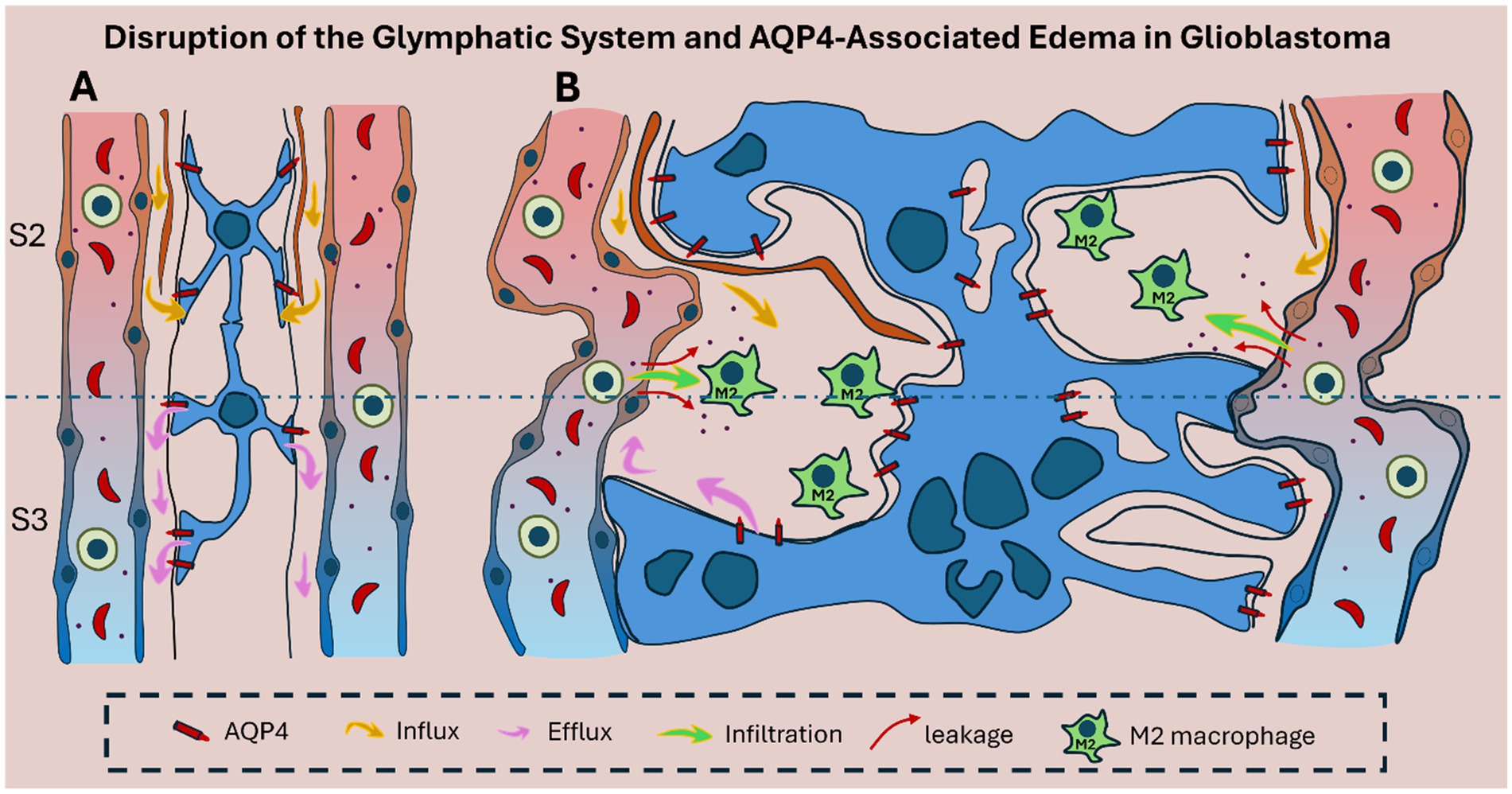
Figure 3. Schematic representation of glymphatic system disruption in glioblastoma. (A) In healthy brain tissue, the glymphatic system supports directional CSF–ISF exchange through intact Virchow–Robin spaces in zones S2 and S3. Polarized AQP4 is concentrated at astrocytic endfeet, enabling efficient perivascular fluid movement and metabolic waste clearance. (B) In glioblastoma, glymphatic architecture is disrupted by glial proliferation, vascular alteration, and tumor-induced structural disorganization. AQP4 becomes mislocalized, reducing directional flow and promoting peritumoral edema. CSF influx (yellow arrows) and impaired efflux (pink arrows) cause fluid accumulation, while increased vascular permeability allows leakage of immune cells (green) and blood-derived products (red arrows), reflecting local BBB disruption that further amplifies edema and inflammation. In addition, M2-polarized macrophages sustain a tumor-permissive microenvironment.
Understanding the link between AQP4 localization and BBB stability could affect therapeutic approaches aimed at restoring vascular integrity. By reestablishing the polarized expression of AQP4 and preserving the dystrophin–dystroglycan complex, it may be possible to reinforce BBB function and attenuate tumor-associated edema and invasion.
Glymphatic dysfunction, AQP4 dysregulation, and peritumoral edema in glioblastoma
Anatomical and functional disturbances in the GS are increasingly recognized as contributors to the pathophysiology of glioblastoma. In particular, tumor-induced compression, glial proliferation, and vascular disorganization disrupt directional perivascular flow and glymphatic clearance, changes that are most pronounced in the CSF–ISF exchange (S2) and perivenous efflux (S3) regions, with reduced meningeal outflow in the newly defined S4 zone as reported in glioma (Rasmussen et al., 2018; Gao et al., 2024; Scalia et al., 2024) (Figure 3).
Advanced MRI techniques have revealed significantly reduced glymphatic activity in GBM patients, correlating with interstitial fluid retention and peritumoral edema (Mueller et al., 2023). These disruptions are closely associated with changes in AQP4 expression and distribution. In healthy brain tissue, AQP4 is highly polarized at astrocytic end-feet lining the perivascular spaces, facilitating the exchange of CSF and ISF, as well as the clearance of metabolic waste (McCoy et al., 2010; Mou et al., 2010). In GBM, however, AQP4 becomes disorganized—often redistributed throughout the astrocytic membrane or even overexpressed in tumor cells—thereby compromising tumor-associated perivascular fluid transport (Noell et al., 2012; Mou et al., 2010) as illustrated schematically in Figure 3 and histologically in Figure 4.
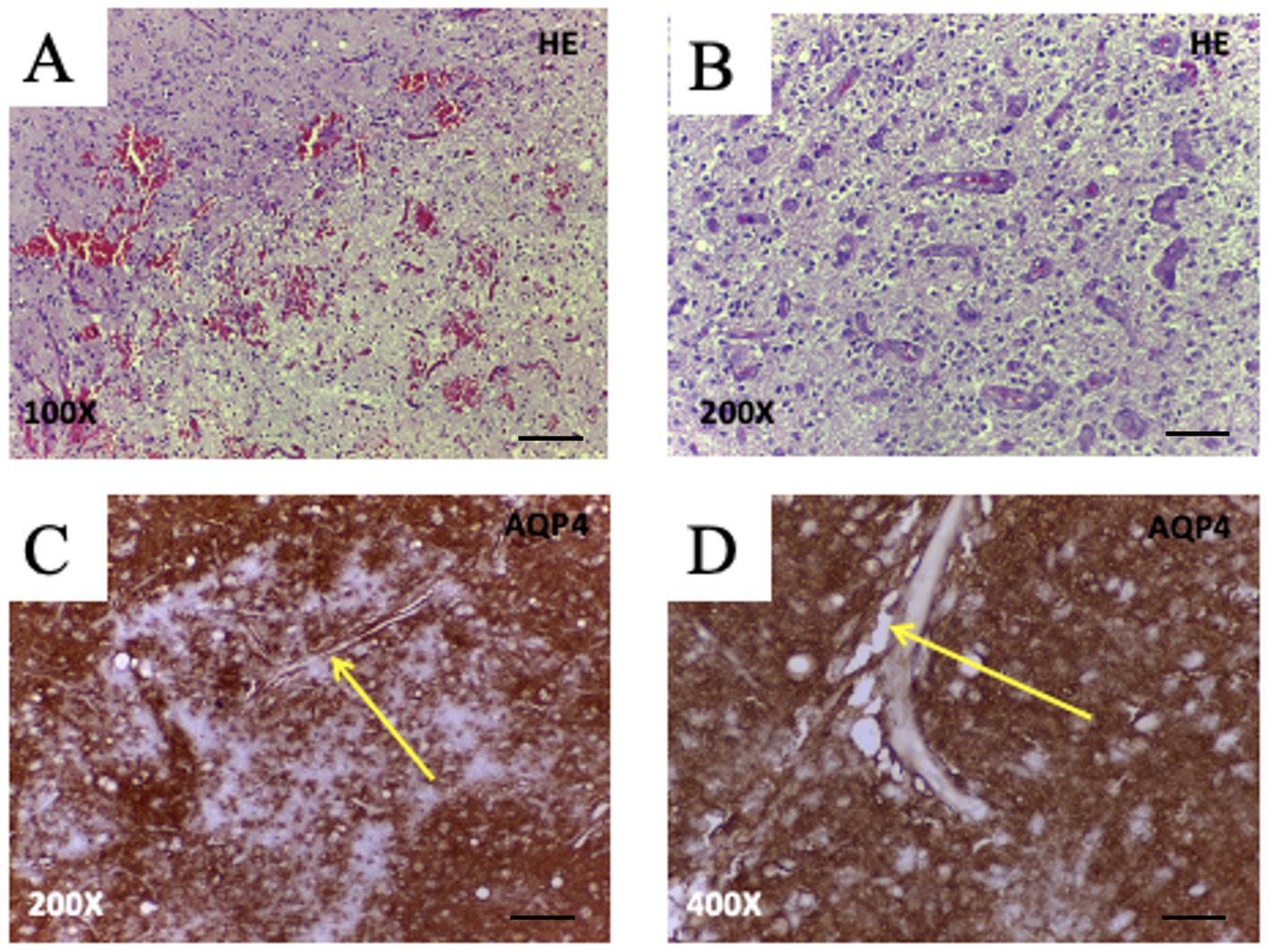
Figure 4. Histopathological features and AQP4 expression in glioblastoma (WHO Grade IV). (A,B) Hematoxylin and eosin (H&E) staining of glioblastoma tissue reveals hallmark features, including moderate hypercellularity and geographic necrosis (A, 100X; bar 200 μm), and prominent microvascular proliferation with glomeruloid-like vessels (B, 200X; bar 100 μm). (C,D) Immunohistochemistry for AQP4 shows strong perivascular labeling at astrocytic endfeet (arrows), highlighting pathological vessels surrounded by AQP4 + glial processes (C, 200X; bar 100 μm) and the presence of irregular, dilated perivascular spaces (D, 400X; bar 50 μm). These findings support AQP4’s role in tumor-associated vascular remodeling, glial proliferation, and potentially impaired glymphatic clearance in glioblastoma. Images from unpublished clinical case from our group.
Histological analyses have shown that AQP4 is frequently upregulated around aberrant tumor vasculature and accumulates near dilated or disorganized perivascular spaces (Warth et al., 2007; Gao et al., 2024; Noell et al., 2012; Scalia et al., 2024) (Figure 4). Such alterations not only disrupt local water homeostasis but also impair perivascular fluid transport, contributing to tissue swelling and elevated intracranial pressure. Consistent with this, Scalia et al. (2024) reported that GBM-associated glymphatic dysfunction involves reduced meningeal lymphatic drainage and loss of directional fluid clearance. In glioblastoma, AQP4 is often mislocalized or overexpressed within tumor cells, resulting in loss of perivascular polarization and impaired coordination of perivascular flow. Experimental models demonstrate reduced glymphatic influx and clearance in tumor-bearing hemispheres (Kaur et al., 2023) while diffusion MRI studies in patients reveal diminished perivascular flow indices (lower DTI-ALPS) in tumor-affected regions (Liang et al., 2025). Given that polarized AQP4 expression at astrocytic endfeet is essential for CSF–interstitial fluid exchange (Simon et al., 2022; Peng et al., 2023) its disruption in GBM likely compromises glymphatic transport not only within the tumor core but also in surrounding brain tissue, thereby promoting peritumoral fluid accumulation and impaired metabolic waste clearance.
The edema is a hallmark of glioblastoma and contributes substantially to the clinical symptoms and poor prognosis associated with the disease. Traditionally considered a passive consequence of tumor growth, peritumoral edema is now recognized as an active process involving glymphatic dysfunction and astrocytic activation. The breakdown of glymphatic flow due to tumor expansion and vascular alteration results in the accumulation of interstitial and cerebrospinal fluid in the peritumoral region (Ma et al., 2019; Scalia et al., 2024; Lan et al., 2023; Siri et al., 2024).
AQP4 plays a central role in this process. Elevated expression and mislocalization of AQP4 around tumor boundaries are strongly correlated with the extent and severity of peritumoral edema (Gao et al., 2024; Noell et al., 2012; Mou et al., 2010). Rather than facilitating efficient fluid clearance, disorganized AQP4 expression disrupts osmotic gradients and contributes to aberrant water influx into the extracellular space, thereby worsening local swelling.
Therapeutically, strategies aimed at modulating AQP4 distribution—either through pharmacologic means or gene-targeted approaches—may offer a way to reduce edema and improve clinical outcomes. Additionally, restoring the integrity of the glymphatic system may help re-establish fluid balance and relieve peritumoral mass effect. As such, understanding the bidirectional relationship between AQP4 function and glymphatic flow is essential for the development of targeted interventions to mitigate edema in GBM.
AQP4 in immune modulation and the tumor microenvironment
Beyond water transport, AQP4 contributes to cellular dynamics and to the immune architecture of glioblastoma; higher AQP4 expression is associated with M2-like tumor-associated macrophage (TAM) enrichment and immune evasion (Lan et al., 2022; Mou et al., 2010). One of its critical functions lies in driving TAM polarization toward the M2 phenotype—a state characterized by immune suppression, angiogenesis, and tissue remodeling (Lan et al., 2022; Wang et al., 2023) (Figure 5). M2-polarized TAMs secrete anti-inflammatory cytokines such as IL-10 and TGF-β, support extracellular matrix reorganization, and suppress antigen presentation, thereby contributing to immune evasion and tumor growth. High AQP4 expression in glioma tissues has been associated with a higher prevalence of M2-like TAMs, linking AQP4 to an immunosuppressive tumor niche (Pisani et al., 2021; Lan et al., 2022; Wang et al., 2023; Seifert et al., 2015; Simone et al., 2022; Du et al., 2020).
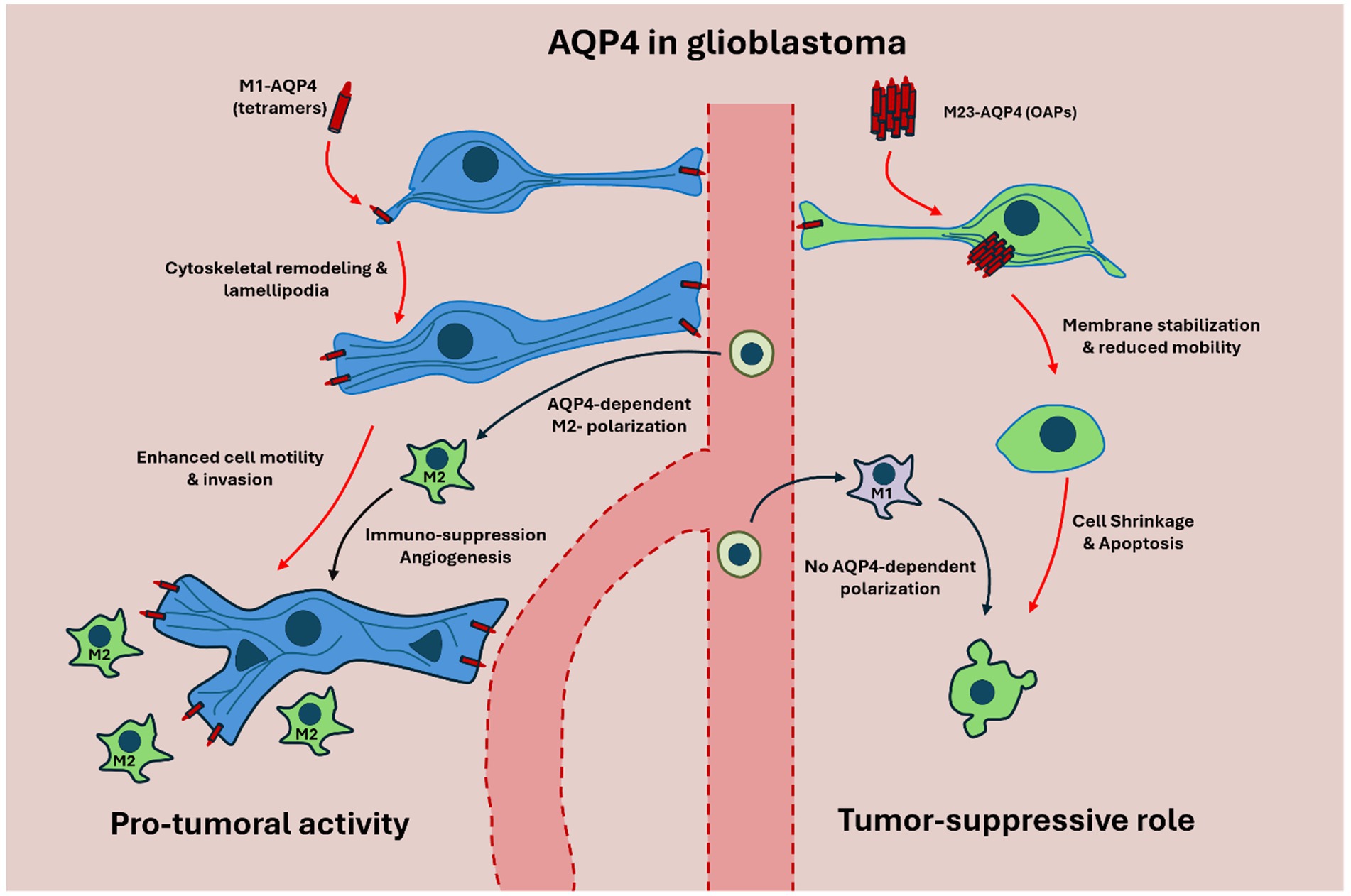
Figure 5. Schematic representation of M1-AQP4 and M23-AQP4 isoforms in glioblastoma (GBM). On the left, M1-AQP4, which forms non-OAP tetramers, predominates in GBM and drives cytoskeletal remodeling and lamellipodia formation, thereby enhancing glioma cell motility, invasion, and extracellular-matrix remodeling. This M1-dominated state is further associated with polarization of tumor-associated macrophages (TAMs) toward the immunosuppressive M2 phenotype, promoting angiogenesis, immune evasion, and tumor progression. On the right, M23-AQP4 assembles into OAPs that stabilize astrocytic membranes and maintain polarized perivascular water flux under physiological conditions. In this configuration, AQP4 supports glymphatic flow, limits cell migration, and favors apoptosis, reflecting a tumor-suppressive and homeostatic profile. Together, these isoform-specific mechanisms illustrate how the M1/M23 balance governs both normal astrocytic physiology and the malignant as well as immunological behavior of GBM.
Recent single-cell RNA sequencing studies further support this relationship by demonstrating shifts in macrophage polarization states in AQP4-high gliomas (Wang et al., 2023). Moreover, AQP4-containing extracellular vesicles have been shown to influence the phenotype of surrounding immune cells and contribute to the remodeling of the tumor milieu (Simone et al., 2022).
Taken together, these findings highlight the immunomodulatory potential of AQP4, underscoring its role beyond its function as a water channel. By orchestrating both glioma cell motility and immune evasion, AQP4 emerges as a dual-threat molecule within the GBM microenvironment. Therapies aimed at modulating AQP4 expression or localization may not only impede tumor spread but also recalibrate the immune landscape toward an anti-tumoral state.
AQP4 as a therapeutic target in glioblastoma
One of the most challenging clinical features of glioblastoma is its resistance to therapies, including radiation and chemotherapy. AQP4 has been implicated in maintaining therapy-resistant niches, particularly in regions of hypoxia and low perfusion where slow-cycling, stem-like glioma cells persist (Jia et al., 2023; MacLeod et al., 2019). These microenvironments exhibit altered water-efflux kinetics, suggesting that AQP4 may support cellular survival under cytotoxic stress by preserving ionic balance in hypo-perfused tumor zones, where drug diffusion is limited.
AQP4-enriched astrocytic domains in gliomas often co-localize with regions exhibiting poor therapeutic penetration, hinting at a protective spatial arrangement that shelters vulnerable glioma subpopulations. In such zones, AQP4’s role in maintaining cell polarity and local fluid dynamics may buffer against treatment-induced apoptosis, enabling tumor cell regrowth (Ding et al., 2011).
Beyond its role in water transport, AQP4 contributes to glioma cell migration and invasion through dynamic cytoskeletal remodeling, membrane plasticity, and interactions with ion channels (Ding et al., 2010; Varricchio et al., 2021). Upstream metabolic signaling via the IDO1/TDO–kynurenine (Kyn) pathway activates the AhR, which upregulates AQP4 and biases isoform balance toward invasive phenotypes; AQP4 then acts as a downstream effector that promotes motility, immune tolerance, and survival under therapeutic stress (Du et al., 2020; Varricchio and Yool, 2023).
Pharmacologic inhibition of AQP4—alone or in combination with ion channel modulators—has shown promise in preclinical models by reducing glioma cell motility, sensitizing tumors to standard therapies, and suppressing recurrence (Varricchio et al., 2021; Varricchio and Yool, 2023). For example, TGN-020, a selective AQP4 inhibitor, has demonstrated efficacy in reducing cerebral edema and glioma cell migration in animal models, while AER-271, a more recent compound, has shown similar effects with improved pharmacokinetics and CNS penetration (Behnam et al., 2022). However, both agents face challenges in clinical translation due to limited blood–brain barrier (BBB) permeability and potential off-target effects. To overcome these limitations, novel delivery strategies such as nanoparticle-based platforms and BBB-penetrating antibody conjugates are being explored. Simultaneous inhibition of AQP4 and co-expressed ion channels has shown synergistic effects in reducing glioma cell viability (MacLeod et al., 2019). Targeting upstream regulators like the Kyn–AhR–AQP4 axis also offers a promising approach to disrupt glioma proliferation and immune evasion (Du et al., 2020).
Altogether, AQP4 represents a mechanistically grounded and multifaceted therapeutic target in GBM. Future efforts should prioritize the development of BBB-permeable inhibitors, identification of AQP4-driven patient subgroups, and clinical evaluation of AQP4-directed interventions to improve long-term outcomes.
AQP4 isoforms and functional roles in gliomas
AQP4 exists in multiple isoforms, primarily M1-AQP4 and M23-AQP4, which differ in their N-terminal sequences and in their ability to form supramolecular structures (Pisani et al., 2021; Smith et al., 2014; de Bellis et al., 2021). These isoforms play divergent roles in glioma biology, particularly influencing cellular motility, invasiveness, and the structural integrity of astrocytic networks.
M1-AQP4, typically forming tetramers, is associated with increased astrocyte migration and invasive capacity in high-grade gliomas. Its expression correlates with poorer prognosis, increased recurrence rates, and greater tumor spread (Sun et al., 2020; Engelhorn et al., 2009). M1-AQP4 enhances the formation of lamellipodia—actin-rich membrane protrusions—facilitating dynamic cytoskeletal remodeling required for glioma cell motility (Smith et al., 2014; Lan et al., 2017) (Figure 5).
In contrast, M23-AQP4 forms orthogonal arrays of particles (OAPs) that stabilize the astrocyte plasma membrane and restrict cell motility. These arrays support a more polarized cellular phenotype, associated with membrane rigidity and induction of apoptosis (Amiry-Moghaddam, 2019; Simone et al., 2019; de Bellis et al., 2021; McCoy and Sontheimer, 2007; Hiroaki et al., 2006). Thus, the relative abundance and distribution of AQP4 isoforms can influence the invasive phenotype of glioma cells (Figure 5).
Molecular regulators of AQP4 isoform expression have also been identified. Thus, The Kyn–AhR axis also influences AQP4 expression by favoring the upregulation of the M1-AQP4 isoform, thus reinforcing the pro-invasive and therapy-resistant phenotype of glioma cells (Du et al., 2020). This metabolic-immunologic axis represents a key interface between tumor metabolism and membrane protein regulation. Moreover, non-coding RNAs such as lncRNA LINC00461 and miR-216a modulate AQP4 expression, further linking gene regulation to tumor behavior (Behnam et al., 2022).
These findings emphasize that AQP4 isoform switching is not merely a structural adaptation but may actively drive glioma progression. Understanding and manipulating this isoform balance could represent a novel therapeutic strategy in GBM.
Clinical intersection of AQP4 function: glioblastoma and neuromyelitis optica spectrum disorders (NMOSD)
The therapeutic significance of AQP4 has prompted the investigation of the intersection between glioblastoma and NMOSD, an autoimmune condition characterized by the presence of pathogenic anti-AQP4 antibodies (Castañeyra-Ruiz et al., 2014). Although chronic AQP4 inhibition in NMOSD may hypothetically decrease the risk of glioma development, there is currently insufficient evidence to substantiate this hypothesis.
Numerous case reports have underscored the diagnostic challenges and occasional clinical overlap between NMOSD and gliomas, primarily due to the risk of misdiagnosis in which often NMOSD is erroneously identified as brain tumors located in regions where AQP4 is abundantly expressed (Park and Hwang, 2021; Song et al., 2020; Tomari et al., 2024; Natsis et al., 2021).
A particularly intriguing case highlighted the coexistence of NMOSD and glioblastoma, where the brain tumor emerged while the patient was undergoing AQP4 immunosuppressive treatment for NMOSD. In this situation, the immunosuppression may have prevented the autoimmune system from identifying and targeting the glioblastoma’s AQP4, facilitating its growth. Notably, upon partial withdrawal of immunosuppressive therapy, NMOSD re-emerged while the glioblastoma entered remission (Donovan et al., 2017). This paradoxical outcome suggests a potential immunological conflict between active NMOSD and glioblastoma progression, reinforcing the notion that AQP4-targeted autoimmunity could inherently oppose glioma advancement. Remarkably, Liao et al. (2010) demonstrated that serum from NMOSD patients, rich in anti-AQP4 antibodies, exhibited reactivity against human glioblastomas. While anti-AQP4 antibodies are widely considered pathogenic in NMOSD, some studies suggest they may represent an epiphenomenon rather than a direct driver of disease pathology (Schmetzer et al., 2021). Although this does not undermine the immunological relevance of AQP4, it introduces nuance into the role of recomanti-AQP4 antibodies in potential therapeutic implications for glioblastoma.
One of the most striking features of glioblastoma is its ability to spread through the perivascular spaces of Virchow–Robin, a phenomenon known as perivascular satellitosis or satellitism. This invasive behavior contributes to the tumor’s poor surgical treatability and is mediated by chemokines, such as stromal cell-derived factor 1 alpha (SDF-1α or CXCL12), which is expressed in subpial blood vessels (Wesseling et al., 2011). Glioblastoma cells express corresponding receptors, including CXCR4, which facilitates chemotactic migration (Wesseling et al., 2011; Esencay et al., 2013; Zagzag et al., 2008). Intriguingly, an inverse relationship has been reported between anti-AQP4 autoantibodies and CXCL12 levels in NMOSD patients, suggesting a potential immunological conflict between NMOSD and glioblastoma progression eritumoral astrocytic mislocalization of AQP4, together with tumor mass effect, compromises glymphatic function by distorting perivascular spaces and compressing CSF–ISF exchange zones (Kang et al., 2015). Moreover, homologous aquaporins, such as AQP3, are known to be upregulated at the migrating edge of cancer cells upon stimulation with CXCL12, further implicating aquaporins in chemokine-driven tumor invasion (Satooka and Hara-Chikuma, 2016).
Given the limited data available, these findings suggest an immunological incompatibility between active NMOSD and glioma progression that warrants further exploration. Systematic reviews and large multicenter patient registries could provide crucial insights into whether NMOSD offers any protective effect against glioma development and whether AQP4-targeted immunity could be therapeutically exploited. These observations should be interpreted with caution, yet they remain valuable for generating new hypotheses.
Conclusions and future directions
This review highlights the central role of aquaporin-4 (AQP4) in glioblastoma pathophysiology, extending beyond its canonical function in water transport to include tumor-cell migration, immune modulation, blood–brain barrier (BBB) disruption, and therapeutic resistance. AQP4 dysfunction—particularly its isoform-specific expression and subcellular mislocalization—emerges as a converging mechanism linking glymphatic system disruption with glioma progression.
We propose a refined anatomical framework for the glymphatic system composed of four segments, each with distinct structural and functional roles in cerebrospinal fluid (CSF)–interstitial fluid (ISF) exchange. This segmentation offers new insights into how specific anatomical zones may be selectively compromised in GBM, resulting in impaired waste clearance, edema formation, and immune evasion.
Therapeutically, AQP4 represents a promising but complex target. Pharmacological inhibitors, isoform-specific regulators, and novel delivery systems (e.g., nanoparticle-conjugated agents) offer avenues for disrupting AQP4-mediated tumor support. However, aquaporins are traditionally considered “undruggable”; therefore, future strategies may focus on modulating their upstream regulatory pathways or selectively interfering with pore function. Moreover, the IDO1/TDO–Kyn–AHR–AQP4 signaling axis, together with AQP4’s interaction with tumor-associated immune components such as macrophages, warrant the exploration of combined therapeutic approaches.
The rare clinical intersection between glioblastoma and NMOSD suggests a potentially exploitable immunological relationship, though further data are required to substantiate therapeutic applications.
Future research should focus on (1) clarifying causal links between AQP4 dysregulation and glymphatic failure in gliomas, (2) identifying biomarkers of AQP4 activity and localization for patient stratification, and (3) developing BBB-penetrant, isoform-selective interventions. Such approaches may improve diagnosis, therapeutic targeting, and clinical outcomes in glioblastoma and related central nervous system tumors.
Author contributions
LC-R: Writing – review & editing, Writing – original draft, Conceptualization. IG-M: Writing – review & editing. LG-A: Writing – review & editing. EG-A: Writing – review & editing. MC: Writing – review & editing. EC-C: Writing – review & editing. SL: Writing – review & editing. CT: Writing – review & editing. BH: Writing – review & editing. MM: Writing – review & editing. AC-P: Writing – review & editing, Writing – original draft, Conceptualization.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that Gen AI was used in the creation of this manuscript. To improve the readability of the manuscript (ChatGPT).
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abbrescia, P., Signorile, G., Valente, O., Palazzo, C., Cibelli, A., Nicchia, G. P., et al. (2024). Crucial role of Aquaporin-4 extended isoform in brain water homeostasis and amyloid-β clearance: implications for edema and neurodegenerative diseases. Acta Neuropathol. Commun. 12:159. doi: 10.1186/s40478-024-01870-4
Alifieris, C., and Trafalis, D. T. (2015). Glioblastoma multiforme: pathogenesis and treatment. Pharmacol. Ther. 152, 63–82. doi: 10.1016/j.pharmthera.2015.05.005
Amiry-Moghaddam, M. (2019). AQP4 and the fate of gliomas. Cancer Res. 79, 2810–2811. doi: 10.1158/0008-5472.CAN-19-1185
Batash, R., Asna, N., Schaffer, P., Francis, N., and Schaffer, M. (2017). Glioblastoma multiforme, diagnosis and treatment; recent literature review. Curr. Med. Chem. 24, 3002–3009. doi: 10.2174/0929867324666170516123206
Behnam, M., Motamedzadeh, A., Aalinezhad, M., Dadgostar, E., Rashidi Noshabad, F. Z., Pourfridoni, M., et al. (2022). The role of aquaporin 4 in brain tumors: implications for pathophysiology, diagnosis and therapy. Mol. Biol. Rep. 49, 10609–10615. doi: 10.1007/s11033-022-07656-y
Carmona-Calero, E. M., Gonzalez-Toledo, J. M., Castañeyra-Ruiz, L., González-Marrero, I., Castañeyra-Ruiz, M., Rancel-Torres, N., et al. (2015). Expression of aquaporin-1 in the choroid plexus in communicating and non-communicating hydrocephalic rats. Eur. J. Anat. 19, 125–129.
Castaneyra-Ruiz, L., Gonzalez-Marrero, I., Carmona-Calero, E. M., Abreu-Gonzalez, P., Lecuona, M., Brage, L., et al. (2016b). Cerebrospinal fluid levels of tumor necrosis factor alpha and aquaporin 1 in patients with mild cognitive impairment and idiopathic normal pressure hydrocephalus. Clin. Neurol. Neurosurg. 146, 76–81. doi: 10.1016/j.clineuro.2016.04.025
Castañeyra-Ruiz, L., González-Marrero, I., Castañeyra-Ruiz, A., González-Toledo, J. M., Castañeyra-Ruiz, M., Perez-Molto, F. J., et al. (2014). The neuroepithelium disruption could generate autoantibodies against AQP4 and cause neuromyelitis optica and hydrocephalus. Int. Scholar. Res. Notices 2014:580572. doi: 10.1155/2014/580572
Castaneyra-Ruiz, L., Gonzalez-Marrero, I., Gonzalez-Toledo, J. M., Castaneyra-Ruiz, A., de Paz-Carmona, H., Castaneyra-Perdomo, A., et al. (2013). Aquaporin-4 expression in the cerebrospinal fluid in congenital human hydrocephalus. Fluids Barriers CNS 10:18. doi: 10.1186/2045-8118-10-18
Castaneyra-Ruiz, L., Gonzalez-Marrero, I., Hernandez-Abad, L. G., Carmona-Calero, E. M., Meyer, G., and Castaneyra-Perdomo, A. (2016a). A distal to proximal gradient of human choroid plexus development, with antagonistic expression of Glut1 and AQP1 in mature cells vs. calbindin and PCNA in proliferative cells. Front. Neuroanat. 10:87. doi: 10.3389/fnana.2016.00087
Castaneyra-Ruiz, L., Gonzalez-Marrero, I., Hernandez-Abad, L. G., Carmona-Calero, E. M., Pardo, M. R., Baz-Davila, R., et al. (2022a). AQP4 labels a subpopulation of white matter-dependent glial radial cells affected by pediatric hydrocephalus, and its expression increased in glial microvesicles released to the cerebrospinal fluid in obstructive hydrocephalus. Acta Neuropathol. Commun. 10:41. doi: 10.1186/s40478-022-01345-4
Castaneyra-Ruiz, L., Gonzalez-Marrero, I., Hernandez-Abad, L. G., Lee, S., Castaneyra-Perdomo, A., and Muhonen, M. (2022b). AQP4, astrogenesis, and hydrocephalus: a new neurological perspective. Int. J. Mol. Sci. 23, 1–18. doi: 10.3390/ijms231810438
Castaneyra-Ruiz, L., Hernandez-Abad, L. G., Carmona-Calero, E. M., Castaneyra-Perdomo, A., and Gonzalez-Marrero, I. (2019). AQP1 overexpression in the CSF of obstructive hydrocephalus and inversion of its polarity in the choroid plexus of a Chiari malformation type II case. J. Neuropathol. Exp. Neurol. 78, 641–647. doi: 10.1093/jnen/nlz033
de Bellis, M., Cibelli, A., Mola, M. G., Pisani, F., Barile, B., Mastrodonato, M., et al. (2021). Orthogonal arrays of particle assembly are essential for normal aquaporin-4 expression level in the brain. Glia 69, 473–488. doi: 10.1002/glia.23909
Ding, T., Gu, F., Fu, L., and Ma, Y. J. (2010). Aquaporin-4 in glioma invasion and an analysis of molecular mechanisms. J. Clin. Neurosci. 17, 1359–1361. doi: 10.1016/j.jocn.2010.02.014
Ding, T., Ma, Y., Li, W., Liu, X., Ying, G., Fu, L., et al. (2011). Role of aquaporin-4 in the regulation of migration and invasion of human glioma cells. Int. J. Oncol. 38, 1521–1531. doi: 10.3892/ijo.2011.983
Donovan, L., Zanazzi, G., Riley, C., Canoll, P., Pereira, M., and Odia, Y. (2017). Concurrent diagnosis of glioblastoma (GBM), neuromyelitis optica (NMO) spectrum disorder and progressive multifocal leukoencephalopathy (PML): imaging, pathology, and clinical course (P5. 161). Neurology 88:161. doi: 10.1212/WNL.88.16_Supplement.P5.161
Du, L., Xing, Z., Tao, B., Li, T., Yang, D., Li, W., et al. (2020). Both IDO1 and TDO contribute to the malignancy of gliomas via the Kyn–AhR–AQP4 signaling pathway. Signal Transduct. Target. Ther. 5:10. doi: 10.1038/s41392-019-0103-4
Engelhorn, T., Savaskan, N. E., Schwarz, M. A., Kreutzer, J., Meyer, E. P., Hahnen, E., et al. (2009). Cellular characterization of the peritumoral edema zone in malignant brain tumors. Cancer Sci. 100, 1856–1862. doi: 10.1111/j.1349-7006.2009.01259.x
Esencay, M., Sarfraz, Y., and Zagzag, D. (2013). CXCR7 is induced by hypoxia and mediates glioma cell migration towards SDF-1α. BMC Cancer 13:347. doi: 10.1186/1471-2407-13-347
Gao, M., Liu, Z., Zang, H., Wu, X., Yan, Y., Lin, H., et al. (2024). A histopathologic correlation study evaluating Glymphatic function in brain tumors by multiparametric MRI. Clin. Cancer Res. 30, 4876–4886. doi: 10.1158/1078-0432.CCR-24-0150
Ghosh, D., Nandi, S., and Bhattacharjee, S. (2018). Combination therapy to checkmate glioblastoma: clinical challenges and advances. Clin. Transl. Med. 7:33. doi: 10.1186/s40169-018-0211-8
Giannetto, M. J., Gomolka, R. S., Gahn-Martinez, D., Newbold, E. J., Bork, P. A., Chang, E., et al. (2024). Glymphatic fluid transport is suppressed by the aquaporin-4 inhibitor AER-271. Glia 72, 982–998. doi: 10.1002/glia.24515
Gomolka, R. S., Hablitz, L. M., Mestre, H., Giannetto, M., Du, T., Hauglund, N. L., et al. (2023). Loss of aquaporin-4 results in glymphatic system dysfunction via brain-wide interstitial fluid stagnation. eLife 12:e82232. doi: 10.7554/eLife.82232
Gonzalez-Marrero, I., Hernandez-Abad, L. G., Gonzalez-Gomez, M., Soto-Viera, M., Carmona-Calero, E. M., Castaneyra-Ruiz, L., et al. (2022). Altered expression of AQP1 and AQP4 in brain barriers and cerebrospinal fluid may affect cerebral water balance during chronic hypertension. Int. J. Mol. Sci. 23, 1–17. doi: 10.3390/ijms232012277
Hablitz, L. M., and Nedergaard, M. (2021). The glymphatic system: a novel component of fundamental neurobiology. J. Neurosci. 41, 7698–7711. doi: 10.1523/JNEUROSCI.0619-21.2021
Hiroaki, Y., Tani, K., Kamegawa, A., Gyobu, N., Nishikawa, K., Suzuki, H., et al. (2006). Implications of the aquaporin-4 structure on array formation and cell adhesion. J. Mol. Biol. 355, 628–639. doi: 10.1016/j.jmb.2005.10.081
Holst, C. B., Brochner, C. B., Vitting-Seerup, K., and Mollgard, K. (2019). Astrogliogenesis in human fetal brain: complex spatiotemporal immunoreactivity patterns of GFAP, S100, AQP4 and YKL-40. J. Anat. 235, 590–615. doi: 10.1111/joa.12948
Iliff, J. J., Goldman, S. A., and Nedergaard, M. (2015). Implications of the discovery of brain lymphatic pathways. Lancet Neurol. 14, 977–979. doi: 10.1016/S1474-4422(15)00221-5
Iliff, J. J., Wang, M., Liao, Y., Plogg, B. A., Peng, W., Gundersen, G. A., et al. (2012). A paravascular pathway facilitates CSF flow through the brain parenchyma and the clearance of interstitial solutes, including amyloid β. Sci. Transl. Med. 4:147ra111-147ra111. doi: 10.1126/scitranslmed.3003748
Jeon, H., Kim, M., Park, W., Lim, J. S., Lee, E., Cha, H., et al. (2021). Upregulation of AQP4 improves blood–brain barrier integrity and perihematomal edema following intracerebral hemorrhage. Neurotherapeutics 18, 2692–2706. doi: 10.1007/s13311-021-01126-2
Jessen, N. A., Munk, A. S. F., Lundgaard, I., and Nedergaard, M. (2015). The glymphatic system: a beginner’s guide. Neurochem. Res. 40, 2583–2599. doi: 10.1007/s11064-015-1581-6
Jia, Y., Xu, S., Han, G., Wang, B., Wang, Z., Lan, C., et al. (2023). Transmembrane water-efflux rate measured by magnetic resonance imaging as a biomarker of the expression of aquaporin-4 in gliomas. Nat. Biomed. Eng. 7, 236–252. doi: 10.1038/s41551-022-00960-9
Kang, H., Cao, S., Chen, T., Jiang, Z., Liu, Z., Li, Z., et al. (2015). The poor recovery of neuromyelitis optica spectrum disorder is associated with a lower level of CXCL12 in the human brain. J. Neuroimmunol. 289, 56–61. doi: 10.1016/j.jneuroim.2015.10.005
Kaur, J., Ding, G., Zhang, L., Lu, Y., Luo, H., Li, L., et al. (2023). Imaging glymphatic response to glioblastoma. Cancer Imaging 23:107. doi: 10.1186/s40644-023-00628-w
Klostranec, J. M., Vucevic, D., Bhatia, K. D., Kortman, H. G., Krings, T., Murphy, K. P., et al. (2021). Current concepts in intracranial interstitial fluid transport and the glymphatic system: part I—anatomy and physiology. Radiology 301, 502–514. doi: 10.1148/radiol.2021202043
Lan, Y. L., Nie, T., and Zou, S. (2022). Identification of the prognostic and immunological roles of aquaporin 4: a potential target for survival and immunotherapy in glioma patients. Front. Cell. Neurosci. 16:1061428. doi: 10.3389/fncel.2022.1061428
Lan, Y. L., Wang, H., Chen, A., and Zhang, J. (2023). Update on the current knowledge of lymphatic drainage system and its emerging roles in glioma management. Immunology 168, 233–247. doi: 10.1111/imm.13517
Lan, Y. L., Wang, X., Lou, J. C., Ma, X. C., and Zhang, B. (2017). The potential roles of aquaporin 4 in malignant gliomas. Oncotarget 8, 32345–32355. doi: 10.18632/oncotarget.16017
Liang, W., Sun, W., Li, C., Zhou, J., Long, C., Li, H., et al. (2025). Glymphatic system dysfunction and cerebrospinal fluid retention in gliomas: evidence from perivascular space diffusion and volumetric analysis. Cancer Imaging 25:51. doi: 10.1186/s40644-025-00868-y
Liao, Z., Ye, J., Guan, Y., Zhang, Y., You, X., An, J., et al. (2010). Construction and clinical application of lentivirus-AQP4 expressing vector. Zhonghua Yi Xue Za Zhi 90, 208–212
Ma, Q., Schlegel, F., Bachmann, S. B., Schneider, H., Decker, Y., Rudin, M., et al. (2019). Lymphatic outflow of cerebrospinal fluid is reduced in glioma. Sci. Rep. 9:14815. doi: 10.1038/s41598-019-51373-9
MacLeod, G., Bozek, D. A., Rajakulendran, N., Monteiro, V., Ahmadi, M., Steinhart, Z., et al. (2019). Genome-wide CRISPR-Cas9 screens expose genetic vulnerabilities and mechanisms of temozolomide sensitivity in glioblastoma stem cells. Cell Rep. 27:e979, 971–986.
McCoy, E. S., Haas, B. R., and Sontheimer, H. (2010). Water permeability through aquaporin-4 is regulated by protein kinase C and becomes rate-limiting for glioma invasion. Neuroscience 168, 971–981. doi: 10.1016/j.neuroscience.2009.09.020
McCoy, E., and Sontheimer, H. (2007). Expression and function of water channels (aquaporins) in migrating malignant astrocytes. Glia 55, 1034–1043. doi: 10.1002/glia.20524
Mou, K., Chen, M., Mao, Q., Wang, P., Ni, R., Xia, X., et al. (2010). AQP-4 in peritumoral edematous tissue is correlated with the degree of glioma and with expression of VEGF and HIF-alpha. J. Neuro-Oncol. 100, 375–383. doi: 10.1007/s11060-010-0205-x
Mueller, S. M., McFarland White, K., Fass, S. B., Chen, S., Shi, Z., Ge, X., et al. (2023). Evaluation of gliovascular functions of AQP4 readthrough isoforms. Front. Cell. Neurosci. 17:1272391. doi: 10.3389/fncel.2023.1272391
Natsis, K., Kalyvas, A., Theochari, E., Papamichalis, E., and Gerkou, A. (2021). Area postrema syndrome in a patient with brainstem glioblastoma. Acta Neurol. Belg. 121, 1087–1088. doi: 10.1007/s13760-021-01736-9
Noell, S., Wolburg-Buchholz, K., Mack, A. F., Ritz, R., Tatagiba, M., Beschorner, R., et al. (2012). Dynamics of expression patterns of AQP4, dystroglycan, agrin and matrix metalloproteinases in human glioblastoma. Cell Tissue Res. 347, 429–441. doi: 10.1007/s00441-011-1321-4
Park, S.-H., and Hwang, J.-H. (2021). Neuromyelitis Optica Spectrum disorders mimicking multiple brain tumors. Brain Tumor Res. Treatm. 9, 35–39. doi: 10.14791/btrt.2021.9.e1
Pearson, J. R. D., Cuzzubbo, S., McArthur, S., Durrant, L. G., Adhikaree, J., Tinsley, C. J., et al. (2020). Immune escape in glioblastoma multiforme and the adaptation of immunotherapies for treatment. Front. Immunol. 11:582106. doi: 10.3389/fimmu.2020.582106
Peng, S., Liu, J., Liang, C., Yang, L., and Wang, G. (2023). Aquaporin-4 in glymphatic system, and its implication for central nervous system disorders. Neurobiol. Dis. 179:106035. doi: 10.1016/j.nbd.2023.106035
Pisani, F., Simone, L., Mola, M. G., De Bellis, M., Frigeri, A., Nicchia, G. P., et al. (2021). Regulation of aquaporin-4 expression in the central nervous system investigated using M23-AQP4 null mouse. Glia 69, 2235–2251. doi: 10.1002/glia.24032
Plog, B. A., and Nedergaard, M. (2018). The glymphatic system in central nervous system health and disease: past, present, and future. Ann. Rev. Pathol. 13, 379–394. doi: 10.1146/annurev-pathol-051217-111018
Rasmussen, M. K., Mestre, H., and Nedergaard, M. (2018). The glymphatic pathway in neurological disorders. Lancet Neurol. 17, 1016–1024. doi: 10.1016/S1474-4422(18)30318-1
Salman, M. M., Kitchen, P., Halsey, A., Wang, M. X., Törnroth-Horsefield, S., Conner, A. C., et al. (2022). Emerging roles for dynamic aquaporin-4 subcellular relocalization in CNS water homeostasis. Brain 145, 64–75. doi: 10.1093/brain/awab311
Satooka, H., and Hara-Chikuma, M. (2016). Aquaporin-3 controls breast cancer cell migration by regulating hydrogen peroxide transport and its downstream cell signaling. Mol. Cell. Biol. 36, 1206–1218. doi: 10.1128/MCB.00971-15
Scalia, G., Ferini, G., Silven, M. P., Noto, M., Costanzo, R., Maugeri, R., et al. (2024). Alterations of the glymphatic system in glioblastomas: a systematic review. Anticancer Res. 44, 3223–3230. doi: 10.21873/anticanres.17140
Schmetzer, O., Lakin, E., Roediger, B., Duchow, A., Asseyer, S., Paul, F., et al. (2021). Anti-aquaporin 4 IgG is not associated with any clinical disease characteristics in neuromyelitis optica spectrum disorder. Front. Neurol. 12:635419. doi: 10.3389/fneur.2021.635419
Seifert, M., Garbe, M., Friedrich, B., Mittelbronn, M., and Klink, B. (2015). Comparative transcriptomics reveals similarities and differences between astrocytoma grades. BMC Cancer 15, 1–22. doi: 10.1186/s12885-015-1939-9
Simon, M., Wang, M. X., Ismail, O., Braun, M., Schindler, A. G., Reemmer, J., et al. (2022). Loss of perivascular aquaporin-4 localization impairs glymphatic exchange and promotes amyloid β plaque formation in mice. Alzheimer's Res Ther 14:59. doi: 10.1186/s13195-022-00999-5
Simone, L., Pisani, F., Binda, E., Frigeri, A., Vescovi, A. L., Svelto, M., et al. (2022). AQP4-dependent glioma cell features affect the phenotype of surrounding cells via extracellular vesicles. Cell Biosci. 12:150. doi: 10.1186/s13578-022-00888-2
Simone, L., Pisani, F., Mola, M. G., De Bellis, M., Merla, G., Micale, L., et al. (2019). AQP4 aggregation state is a determinant for glioma cell fate. Cancer Res. 79, 2182–2194. doi: 10.1158/0008-5472.CAN-18-2015
Siri, S., Burchett, A., and Datta, M. (2024). Simulating the impact of tumor mechanical forces on glymphatic networks in the brain parenchyma. Biomech. Model. Mechanobiol. 23, 2229–2241. doi: 10.1007/s10237-024-01890-y
Smith, A. J., Jin, B. J., Ratelade, J., and Verkman, A. S. (2014). Aggregation state determines the localization and function of M1- and M23-aquaporin-4 in astrocytes. J. Cell Biol. 204, 559–573. doi: 10.1083/jcb.201308118
Solar, P., Hendrych, M., Barak, M., Valekova, H., Hermanova, M., and Jancalek, R. (2022). Blood-brain barrier alterations and edema formation in different brain mass lesions. Front. Cell. Neurosci. 16:922181. doi: 10.3389/fncel.2022.922181
Song, D., Xu, D., Gao, Q., Hu, P., and Guo, F. (2020). Intracranial metastases originating from pediatric primary spinal cord glioblastoma multiforme: a case report and literature review. Front. Oncol. 10:99. doi: 10.3389/fonc.2020.00099
Sun, D.-P., Lee, Y.-W., Chen, J.-T., Lin, Y.-W., and Chen, R.-M. (2020). The bradykinin-BDKRB1 axis regulates aquaporin 4 gene expression and consequential migration and invasion of malignant glioblastoma cells via a Ca2+-MEK1-ERK1/2-NF-κB mechanism. Cancer 12:667. doi: 10.3390/cancers12030667
Tomari, Y., Igata, Y., Chong, P. F., Kajiwara, K., Hatai, E., Sonoda, Y., et al. (2024). The etiology and outcome of area postrema syndrome in childhood: two cases and a literature review. Pediatr. Neurol. 152, 11–15. doi: 10.1016/j.pediatrneurol.2023.12.010
Valente, O., Abbrescia, P., Signorile, G., Messina, R., de Gennaro, L., Bozzi, M. T., et al. (2024). Aquaporin-4 expression levels and mis-localization are inversely linked to peritumoral edema in gliomas of varying aggressiveness. bioRxiv. doi: 10.1101/2024.09.15.613106
Valente, O., Messina, R., Ingravallo, G., Bellitti, E., Zimatore, D. S., de Gennaro, L., et al. (2022). Alteration of the translational readthrough isoform AQP4ex induces redistribution and downregulation of AQP4 in human glioblastoma. Cell. Mol. Life Sci. 79:140. doi: 10.1007/s00018-021-04123-y
van Linde, M. E., Brahm, C. G., de Witt Hamer, P. C., Reijneveld, J. C., Bruynzeel, A. M. E., Vandertop, W. P., et al. (2017). Treatment outcome of patients with recurrent glioblastoma multiforme: a retrospective multicenter analysis. J. Neuro-Oncol. 135, 183–192. doi: 10.1007/s11060-017-2564-z
Varricchio, A., Ramesh, S. A., and Yool, A. J. (2021). Novel ion channel targets and drug delivery tools for controlling glioblastoma cell invasiveness. Int. J. Mol. Sci. 22:11909. doi: 10.3390/ijms222111909
Varricchio, A., and Yool, A. J. (2023). Aquaporins and ion channels as dual targets in the design of novel glioblastoma therapeutics to limit invasiveness. Cancer 15:849. doi: 10.3390/cancers15030849
Wang, R., Peng, L., Xiao, Y., Zhou, Q., Wang, Z., Tang, L., et al. (2023). Single-cell RNA sequencing reveals changes in glioma-associated macrophage polarization and cellular states of malignant gliomas with high AQP4 expression. Cancer Gene Ther. 30, 716–726. doi: 10.1038/s41417-022-00582-y
Warth, A., Kröger, S., and Wolburg, H. (2004). Redistribution of aquaporin-4 in human glioblastoma correlates with loss of agrin immunoreactivity from brain capillary basal laminae. Acta Neuropathol. 107, 311–318. doi: 10.1007/s00401-003-0812-0
Warth, A., Simon, P., Capper, D., Goeppert, B., Tabatabai, G., Herzog, H., et al. (2007). Expression pattern of the water channel aquaporin-4 in human gliomas is associated with blood–brain barrier disturbance but not with patient survival. J. Neurosci. Res. 85, 1336–1346. doi: 10.1002/jnr.21224
Wesseling, P., Kros, J. M., and Jeuken, J. W. (2011). The pathological diagnosis of diffuse gliomas: towards a smart synthesis of microscopic and molecular information in a multidisciplinary context. Diagn. Histopathol. 17, 486–494. doi: 10.1016/j.mpdhp.2011.08.005
Wolburg, H., Noell, S., Fallier-Becker, P., Mack, A. F., and Wolburg-Buchholz, K. (2012). The disturbed blood–brain barrier in human glioblastoma. Mol. Asp. Med. 33, 579–589. doi: 10.1016/j.mam.2012.02.003
Wu, W., Klockow, J. L., Zhang, M., Lafortune, F., Chang, E., Jin, L., et al. (2021). Glioblastoma multiforme (GBM): An overview of current therapies and mechanisms of resistance. Pharmacol. Res. 171:105780. doi: 10.1016/j.phrs.2021.105780
Xu, D., Zhou, J., Mei, H., Li, H., Sun, W., and Xu, H. (2022). Impediment of cerebrospinal fluid drainage through glymphatic system in glioma. Front. Oncol. 11:790821. doi: 10.3389/fonc.2021.790821
Yalamarty, S. S. K., Filipczak, N., Li, X., Subhan, M. A., Parveen, F., Ataide, J. A., et al. (2023). Mechanisms of resistance and current treatment options for glioblastoma Multiforme (GBM). Cancers (Basel) 15, 1–29. doi: 10.3390/cancers15072116
Zagzag, D., Esencay, M., Mendez, O., Yee, H., Smirnova, I., Huang, Y., et al. (2008). Hypoxia-and vascular endothelial growth factor-induced stromal cell-derived factor-1α/CXCR4 expression in glioblastomas: one plausible explanation of Scherer's structures. Am. J. Pathol. 173, 545–560. doi: 10.2353/ajpath.2008.071197
Keywords: AQP4, glioblastoma, glioma, glymphatic system, astrocyte
Citation: Castañeyra-Ruiz L, González-Marrero I, García-Abad LH, Gonzalez-Arnay E, Camacho M, Carmona-Calero EM, Lee S, Tran CT-Q, Hanak BW, Muhonen M and Castañeyra-Perdomo A (2025) Aquaporin-4 in glioblastoma: a nexus of glymphatic dysfunction, edema, immune evasion, and treatment resistance. Front. Cell. Neurosci. 19:1685491. doi: 10.3389/fncel.2025.1685491
Edited by:
Bozena Kaminska, Polish Academy of Sciences, PolandReviewed by:
Yanet Karina Gutierrez-Mercado, University of Guadalajara, MexicoAnna Malik, University of Warsaw, Poland
Copyright © 2025 Castañeyra-Ruiz, González-Marrero, García-Abad, Gonzalez-Arnay, Camacho, Carmona-Calero, Lee, Tran, Hanak, Muhonen and Castañeyra-Perdomo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Leandro Castañeyra-Ruiz, TGVhbmRyby5jYXN0YW5leXJhLnJ1aXpAY2hvYy5vcmc=