 Edoardo Zaccaria
Edoardo Zaccaria Elianne van der Valk2
Elianne van der Valk2 Soumya K. Kar
Soumya K. Kar Johanna M. J. Rebel
Johanna M. J. Rebel Dirkjan Schokker
Dirkjan Schokker- 1Wageningen Livestock Research, Animal Nutrition, Wageningen, Netherlands
- 2Wageningen Livestock Research, Animal Breeding and Genomics, Wageningen, Netherlands
- 3Wageningen Bioveterinary Research, Epidemiology, Bioinformatics, Animal Models, and Vaccine Development, Lelystad, Netherlands
- 4Adaptation Physiology, Wageningen University, Wageningen, Netherlands
Epigenomics, a field that studies epigenetic changes on a genome-wide scale, has gained prominence because of its potential to reveal biological mechanisms underlying phenotypes in livestock. Animal production is highly dependent on the interaction between animal genetics, physiology, environment, and management practises. Many of these factors have a bidirectional relationship with the epigenome, as they influence and are influenced by it. This article focuses on the role of epigenetics in the adaptation of livestock to environment, particularly heat stress. Epigenetic changes induced by heat stress have been observed in livestock, resulting in short- and long-term alterations that generally affect production performance and health. Research provides strong evidence that gene expression in livestock is also influenced by epigenetic processes such as DNA methylation, histone modifications, chromatin remodelling, and non-coding RNAs to cope with heat stress. Nutritional interventions are a promising way to mitigate the epigenetic changes induced by heat stress. A better understanding of the molecular mechanisms involved in the regulation of gene expression during heat stress is crucial to identify strategies and interventions that can maintain or even improve the health and productivity of monogastric livestock and adapt their resilience and efficiency to different environmental conditions.
Introduction
The use of high-throughput technologies in livestock research has increased interest in defining cellular signalling pathways through advances in molecular technology. These include nucleic acid and protein sequencing, as well as metabolite profiling and analysis, with the goal of better understanding the various biological molecules-DNA, RNA, proteins, and metabolites. Often referred to as “omics” studies, this approach provides a better understanding of the mechanisms regulating an animal’s actual physiology and interprets biological mechanisms underlying complex phenotypes, like health and feed efficiency. Enabling omics technology, livestock researchers can estimate breeding values more accurately, thereby assisting the selection of animals (Chakraborty et al., 2022) and finding nutritional strategies or other interventions (Kar, 2017) that enhance animal health and productivity during environmental challenges.
Epigenomics is a field of study that explores the study of epigenetic modifications on a genome-wide scale. From the Aristotelian word epigenesis, the term ‘epigenetics’ was derived, and it was first coined by Conrad Hal Waddington (Waddington, 1942). Epigenetics refers to changes in gene expression or cellular phenotype that occur without altering the underlying DNA sequence. This involves DNA methylation, post-translational modification of histones, but also linked to the regulation of gene expression by non-coding RNAs, genome instabilities or any other force that could modify a phenotype. Enabling the rapid advancements in next-generation sequencing technology, livestock researchers often generate an enormous amount of epigenomic sequencing data that help them to identify and gather epigenomic biomarkers that reveal biological mechanisms underlying complex health phenotypes in livestock. Increasing lines of scientific evidence support the concept that certain acquired traits are derived from environmental exposure during early embryonic and foetal development, i.e., foetal programming, and can even be “memorized” in the germline as epigenetic information and be transmitted to future generations as a non-genetic inherited factors (Zhu et al., 2021; David et al., 2019).
Epigenetic modifications play a crucial role in regulating gene expression and can be influenced by various factors, including developmental processes and environmental stimuli. Heat stress is one of the main environmental factors that negatively affect animal health and welfare, and it will certainly continue to threaten food security (Ross et al., 2015). Specifically, heat stress-induced economic losses result to poor performance, reduced and inconsistent growth, decreased carcass quality, and increased mortality and morbidity (Ross et al., 2015). Despite the fact that all farm animal species are susceptible to heat stress, birds and pigs are particularly sensitive to heat stress due to either lacking or non-functional sweat glands and continues to threaten global sources of animal protein (Ringseis and Eder, 2022; Guo et al., 2018; Nawab et al., 2018; Saeed et al., 2019).
Animal production is highly dependent on the interaction between animal genetics, physiology, environment, and management practices, which include housing and feeding, many of which have a bidirectional relationship with the epigenome, as they influence and are influenced by it. Therefore, it is imperative to consider epigenetics as one of the factors responsible for phenotypic variation and resilience in livestock, and it should be considered in livestock breeding, health and disease, management including housing, and provision of nutrition, especially in response to heat stress. While emerging studies indicate that RNA modifications, or RNA epigenetics, also contribute significantly to stress responses in livestock (Xing et al., 2024), only few studies have examined this aspect in monogastrics. In this article, we review the epigenetic coping mechanisms in poultry and pigs (see Figure 1 for a schematic representation). In addition, the impact of changes during embryogenesis and the putative impact of nutrients on animal physiology are discussed.
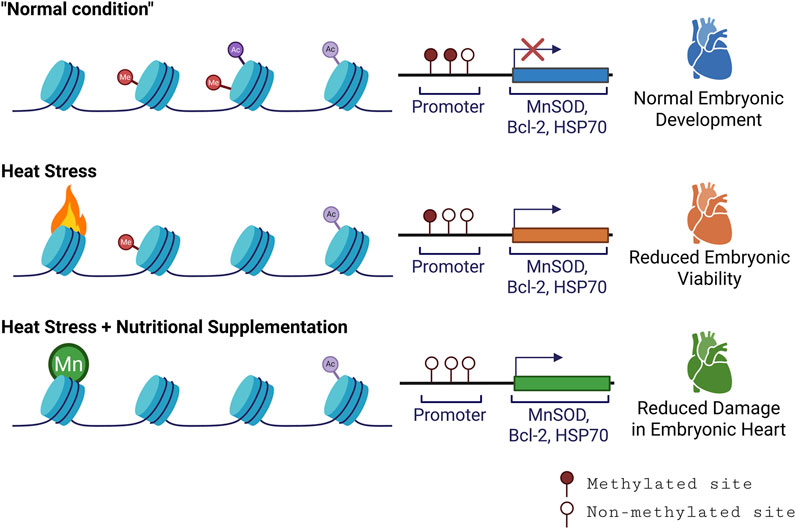
Figure 1. Schematic overview of epigenetic markers, modification, and their effect on host phenotype. Here we highlight that heat stress affects the epigenetic landscape of the embryonic chicken heart. Under the normal condition (baseline methylation), genes involved in antioxidant defence and cell survival (i.e., MnSOD, Bcl-2, HSP70) remain silenced due to DNA methylation and histone modification. Heat stress partially relaxes chromatin structure, but is insufficient alone, leading to reduced embryonic viability. Manganese (Mn) supplementation promotes histone H3K9 hypomethylation and gene activation, leading to improved antioxidant responses and reduced embryonic mortality (Created with BioRender).
Epigenetic processes
By epigenome, we refer to all epigenetic marks on DNA and RNA in a single cell that are generated by epigenetic processes that regulate gene expression and play an important role in genome function and stability. These epigenetic processes are important to the livestock (Pandey et al., 2025), which include DNA and RNA methylation, histone modification, chromatin remodelling, and non-coding RNAs. These processes, including DNA methylation, histone modifications, chromatin remodelling, and non-coding RNAs, play a central role in cellular identity, genome integrity, and environmental adaptation (Sindhu et al., 2024; Ibeagha-Awemu et al., 2022). Epigenetic mechanisms are increasingly recognised as crucial regulators of phenotype in livestock species, impacting traits such as growth, reproduction, immunity, and stress responsiveness (Pandey et al., 2025). In recent years, major technological breakthroughs have transformed our ability to study the animal epigenome with greater resolution and biological relevance. Approaches such as whole-genome bisulfite sequencing (WGBS) for DNA methylation (Yang et al., 2025; Brito et al., 2025), CUT&RUN and CUT&TAG for histone mark profiling (Zhou et al., 2024), and single-cell transcriptomic and chromatin accessibility mapping (e.g., single-cell ATAC-seq, multi-omics platforms) now allow researchers to simultaneously map transcriptional activity, chromatin accessibility, and epigenomic state at the single-cell level in livestock tissues (Li et al., 2024; Tan et al., 2025). These advances have led to unprecedented insights into development, stress responses, and disease in agriculturally important animals. Additionally, the emergence of ideas like epigenetic memory, transgenerational inheritance, and the discovery of functional epitranscriptomic marks (e.g., m6A) in stress responses have shifted the view from static to dynamic and reversible models of gene regulation (Laporta et al., 2024; Henikoff et al., 2025; Sherif et al., 2024; Sajjanar et al., 2025; Xie et al., 2025). With epigenetic memory, we refer to a transient stress that can cause stable changes in gene expression through persistent chromatin marks, such as H3K27ac or DNA methylation, at regulatory regions. It has been demonstrated in chickens conditioned to elevated embryonic incubation temperatures, leading to sustained HSP downregulation and thermotolerance (David et al., 2019; Rosenberg et al., 2022; Halli et al., 2025). Transgenerational epigenetic inheritance, where stress-induced epigenetic marks (e.g., altered methylation in germ cells or early embryos) are passed to offspring, has been observed in poultry and other relevant livestock species (Halli et al., 2025; He et al., 2023). The rise of epitranscriptomics implicates RNA modifications, especially N6-methyladenosine (m6A), as key regulators of mRNA stability, translation, and stress signalling in livestock cells (Chen et al., 2023). A livestock-wide review by Ren et al. (Ren et al., 2024) consolidates evidence that m6A machinery operates across target tissues, liver, muscle, reproductive organs, mediating environmental adaptation and productivity traits.
DNA methylation
DNA methylation, an addition of a methyl group to a cytosine adjacent to guanine (CpG), is a critical and most studied epigenetic process in mammalian models, catalysed by DNA methyltransferases and demethylated by ten-eleven translocation methylcytosine dioxygenases (TETs) (Unoki, 2019; Wu and Zhang, 2017). Methylation in promoter regions often leads to gene silencing, probably achieved via direct inhibition of transcription factor (TF) binding (Becker et al., 1987; Dantas Machado et al., 2015), while intragenic methylation can enhance transcription and affect splicing (L et al., 2009; Gelfman et al., 2013). This process modulates gene expression in response to environmental stimuli, such as heat stress, across various animal taxa, like worms (Caenorhabditis elegans), fish (Gasterosteus aculeatus) and chicken (Gallus gallus) (Wan et al., 2021; Metzger and Schulte, 2017; Vinoth et al., 2018). DNA methylation’s role in transcriptional regulation suggests a significant potential contribution to inherited phenotypes, although research predominantly focuses on parent-to-offspring inheritance (Fitz-James and Cavalli, 2022).
Histone modifications
Histone modification is another epigenetic mechanism with important implications for altering gene expression in response to external stimuli. Histones are a family of proteins (H1-H5) that form structures called nucleosomes via the ordering and packing of the DNA molecule into structural units. Usually, the post-translational modification encompasses methylation, phosphorylation, acetylation, ubiquitylation, or sumoylation of the histone N-terminal tail (Bannister and Kouzarides, 2011; Swygert and Peterson, 2014). While these modifications often upregulate gene expression, the effect is complex, depending on the modification’s nature and location (Jambhekar et al., 2019). Histone modifications, governed by enzymes like histone acetylases and deacetylases, regulate chromatin structure and, thus, transcription factors’ access (Huang et al., 2015; Dose et al., 2011). In heat stress studies, the focus has been on H2B and H3 histones, noting changes in methylation patterns (Marinova et al., 2011; Wu et al., 2020), and multigenerational inheritance of modifications (Klosin et al., 2017).
Chromatin remodelling (3D genome organisation)
In recent years, more importance has been given to the role of the genome structural organisation and DNA folding in gene regulation, DNA repair, chromosome translocation and cell development (Therizols et al., 2014). Despite DNA folding into nucleosome being well described in the literature (Luger et al., 1997), little is known about how nucleosome interacts with each other or how chromatin folds within the nucleus (Bonev and Cavalli, 2016). Major chromatin remodelling has also been observed upon heat shock, mediated by the Heat Shock Protein 70 (HSP70) and aimed to activate the heat shock response (Khanna et al., 2014). Moreover, local, and quick chromatin changes have been observed within 60 s from temperature elevation, as well as displacement of nucleosomes followed by extensive transcriptional activation of several HSP genes (Chowdhary et al., 2019; Kainth et al., 2021). Finally, chromatin remodelling seems to play a pivotal role in animal acclimatisation, increasing resilience to future heat exposures upon moderate heat over long periods (Murray et al., 2022).
Non-coding RNAs
Non-coding RNAs (ncRNAs) are RNA molecules not translated into proteins but perform various molecular functions. Among the different types of ncRNAs, PIWI-interacting RNA (piRNA), small interfering RNA (siRNA), Long non-coding RNAs (lncRNA), and microRNAs (miRNA) regulate gene expression at the transcriptional and post-transcriptional level. In recent years, a growing interest has been posed on their role in stress response, adaptation, and epigenetic information inheritance (Duempelmann et al., 2020; Czech et al., 2018; O'Brien et al., 2018; Raza et al., 2021; Rossnerova et al., 2020).
RNA modification
RNA modifications, such as N6-methyladenosine (m6A), represent a dynamic and reversible layer of gene regulation, also referred to as RNA epigenetics. Beyond m6A, more than 160 distinct RNA chemical modifications have been identified, influencing RNA stability, splicing, localization, and translation. Recent breakthroughs reveal that m6A and its associated enzymes modulate the heat stress response in livestock (Cerneckis et al., 2024). For example, maternal heat exposure in pigs alters m6A methylation patterns in neonatal liver and adipose tissue, affecting early fat deposition and metabolic programming (Heng et al., 2019). Similar regulatory roles for m6A have been demonstrated in sheep, where heat stress influences both lipid metabolism and heat shock protein gene expression via m6A-dependent mechanisms (Chen et al., 2023). Although direct evidence in poultry remains limited, cell and a few animal studies indicate that m6A methylation dynamically regulates heat-shock protein pathways in response to acute temperature shifts, suggesting a conserved function across species (Cao et al., 2024). These insights position RNA epigenetics as a promising frontier for enhancing thermotolerance and resilience in monogastric livestock.
Coping with the changing environment, heat stress and epigenetic modifications
Heat stress affects livestock productivity, disrupting growth rates, fertility, meat quality, and overall health and welfare. Such stress reprioritizes metabolism, physiology, and behaviour, emphasizing thermoregulation over other activities (Wu et al., 2020; Negrón-Pérez et al., 2019). Over time, heat stress triggers metabolic adaptations and epigenetic modifications that aid in the altered physiological states, from structural and behavioural adjustments to physiological and immunoregulatory changes (Gupta et al., 2022; Goel et al., 2021). Epigenetics plays a major role in this response and eventual adaptation to heat and other stressors, inducing a molecular memory of past experiences (see also Figure 2).
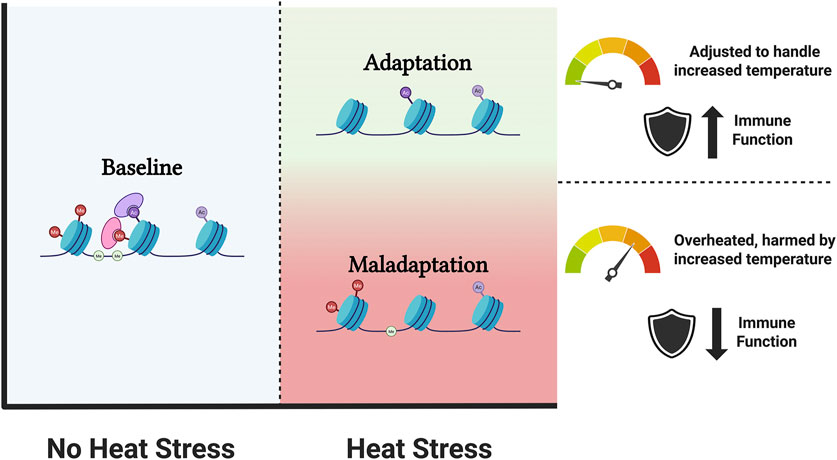
Figure 2. Schematic representation of adaptation to heat stress. The blue zone indicates conditions without heat stress, characterized by a baseline epigenetic landscape. When the animal is experiencing heat stress, adaptation (green zone) or maladaptation (red zone) of the epigenetic landscape occurs. Adapted animal are adjusted to handle increased temperatures often with improved immune function, whereas maladapted animals show overheating, are harmed by the increased temperatures and show a detrimental effect on their immune function. (Created with BioRender).
DNA methylation and histone modifications are fundamental regulatory mechanisms found across all biological states, but changes in these epigenetic marks have been observed in animals experiencing heat stress (Wu et al., 2020; Murray et al., 2022; Wang et al., 2019), and can contribute to both short-term adaptation and long-term health consequences. Long and moderate exposure promotes alteration to suboptimal ambient temperature (acclimation to heat or adaptation), leading to easier and faster activation of the heat shock response in the future (Murray et al., 2022). On the other hand, intense heat stress or severe heat injury is associated with long-term health consequences, affecting several metabolic and physiological processes, i.e., immunosuppression, altered heat shock protein (HSP) response, impair cell morphology and apoptosis, which can lead to maladaptation (Wang et al., 2019; Murray et al., 2021; Argaud et al., 2007).
The heat shock response is crucial in cellular and animal survival under stress. This response triggers the rapid increase of HSP that function mainly as molecular chaperones, providing thermotolerance and protection from other stressors (Hietakan et al., 2006). Importantly, HSP expression can be regulated through epigenetic mechanisms, providing an important link between stress adaptation and epigenetics (Tetievsky and Horowitz, 1985; Kisliouk et al., 2017). Temperature-dependent histone-phosphorylation induces the active state of the chromatin that will also be maintained when the stressors are removed, allowing a rapid re-acclimation and promoting a more heat-resilient phenotype (Murray et al., 2022), forming epigenetic memory and affecting HSP expression for several cell cycles (Kisliouk et al., 2017; Dor and Cedar, 2018). For livestock, poultry researchers have shown that by increasing the egg shell temperature during incubation, reduces HSP gene expression post-hatch when encountering a high ambient temperature (Rajkumar et al., 2017).
Dietary strategies and nutritional interventions, including increased dietary fat content and reduced protein intake, have been shown to mitigate heat stress, leading to improved egg production, growth performance, and metabolic efficiency (Poorghasemi et al., 2013; Spencer et al., 2003; Moallem et al., 2010; Kolani et al., 2018). Additives rich in phenolic compounds, flavonoids, and antioxidants also enhance animal welfare during thermal stress (Zimbelman et al., 2010; Oke et al., 2021; Oke, 2018). Moreover, the overall micronutrient composition of the feed must be formulated based on the environmental condition to help the animal to mitigate stress effects. Vitamin C, E, minerals (Na+, K+, and Mg+), and methionine have been reported to produce beneficial effects in goats and chicken (Nwunuji et al., 2014; Saiz del Barrio et al., 2020).
Furthermore, an intriguing concept known as cross-tolerance has been observed in mice and chicken, where early-life heat stress exposure results in increased resilience to other forms of stress (Ben-Nun et al., 2022; Shein et al., 2005). This response is likely due to shared molecular mechanisms across different stress types, leading to a faster and more robust response to subsequent stressors (Ben-Nun et al., 2022; Shein et al., 2005). Rosenberg et al. (Rosenberg et al., 2022; Rosenberg et al., 2020) showed that chicks exposed to high temperatures during embryonic stages displayed transgenerational heat and immunological resilience, emphasizing the potential of epigenetic processes in modulating stress responses (Rosenberg et al., 2022). This transgenerational adaptation, the potential for dietary intervention, and understanding of the heat shock response highlight the importance of multi-disciplinary approaches to tackle the challenges climate change presents to monogastric livestock production.
Epigenetic impact of heat stress in chicken embryo
During embryogenesis, careful epigenetic remodelling is necessary to avoid developmental defects and ensure healthy development of the embryo. Genomic imprinting involves genes only expressed from the maternal or paternal chromosome in diploid cells and plays crucial roles in early vertebrate development (Barlow, 2011). Heat stress during the early embryonic period can result in lifelong consequences, altering physiological processes, for example, in mouse embryos aberrant methylation imprinting resulted in developmental failure (Zhu et al., 2008). Moreover, these changes may significantly affect embryonic growth, as well as the potential to pass down DNA methylation errors to subsequent generations of livestock (Ross et al., 2017; Ayo et al., 2011; Huber et al., 2020). Previous research has focused primarily on the negative effects of heat stress during embryogenesis (Ross et al., 2017; Ayo et al., 2011; Wettemann and Bazer, 1985). Recent studies have shown that controlled exposure to heat stress during this period can induce epigenetic changes that enhance an animal’s ability to adapt to higher temperatures later in life (Rocha et al., 2022; Piestun et al., 2011).
Host response to heat stress
Heat shock proteins provide cellular protection, have anti-apoptotic effects, and are synthesized under stress. They play a key role in the heat stress response and adaptation of an animal’ (Parsell et al., 1994). Their expression can be modulated by embryonic heat exposure, and epigenetic processes, such as histone modifications or DNA methylation of HPS promoter regions, may have subsequent effects on heat stress resistance in adulthood (Vinoth et al., 2018; Tetievsky and Horowitz, 1985; Kisliouk et al., 2017).
Epigenomic alteration results shed light on coping and adaptive abilities during heat stress in livestock. Plasticity of the preoptic anterior hypothalamus (POAH) plays an important role in thermoregulation of an animal, and its response may vary depending on heat stress during embryogenesis’ (Tzschentke and Basta, 2002; Griffin et al., 2001). Changes in thermoregulatory activity due to embryonic heat stress may differ between vertebrates. Piglets born from heat-stressed sows had less skeletal muscle and more adipose tissue (Johnson et al., 2015), whereas heat-stressed chickens demonstrated reduced metabolic rates and core temperatures (Piestun et al., 2011; Piestun et al., 2009; Piestun et al., 2008). The plasticity of the POAH in response to environmental stressors has increased the interest in the hypothalamus’s underlying epigenetic changes. Heat stress-induced epigenetic alterations in the hypothalamus may impact the whole body thermoregulation and metabolism. Reduced thyroid gland activity and thyroid hormone T3 levels in heat-stressed chickens emphasize the metabolic shifts, potentially optimizing the animals’ energy efficiency (Vinoth et al., 2018; Piestun et al., 2008; McNabb, 1995).
Results from studies indicate that embryonic heat stress causes epigenetic reprogramming in the hypothalamus, which changes molecular pathways involved in thermoregulation and metabolism. David et al. reported epigenomic modification while investigating the genome-wide distribution of histone modifications in the hypothalamic tissue of heat-stressed chicken embryos. They found modifications in genes related to neurodevelopment, metabolism, and gene regulation that may contribute to environmental stress response (David et al., 2019). These epigenetic changes may contribute to thermal acclimation later in life. Furthermore, the genes like the mammalian ortholog for CREB Binding Protein (Cbp-1) and the gene encoding SWI/SNF complex subunit SMARCC2 (Swsn-1), are vital for acetylation and chromatin remodelling, respectively, play central roles in heat stress adaptation (Zhou et al., 2019). In Caenorhabditis elegans, inhibiting these genes disrupted the memory of heat adaptation, suggesting their pivotal role in long-term thermoregulatory responses (Zhou et al., 2019; Booth and Brunet, 2016; Benayoun et al., 2015).In addition, embryonic heat stress boosts levels of both corticotropin-releasing hormone (CRH) (Cramer et al., 2019) and brain-derived neurotrophic factor (BDNF) in the hypothalamus (Yossifoff et al., 2008). In fact, extreme heat stress in early postnatal chickens caused an increase in CRH protein levels in the hypothalamus compared to moderate heat stress and control chicken. This increase was associated with hypermethylation in the intron site of the Crh, silencing the gene expression (Cramer et al., 2019). Moreover, the expression of the BDNF, an activator of the biochemical pathway involved in heat adaptation (Labunskay and Meiri, 2006; Meiri, 2008) and also controlled by epigenetic processes, in the hypothalamus was increased 3 days after hatching, suggesting a potentially long-lasting effect of embryonic heat stress on several neuronal changes (Yossifoff et al., 2008).
A better understanding of the molecular mechanisms involved in the regulation of gene expression during heat stress is crucial to identify strategies and interventions that can maintain or even improve the health and productivity of livestock and adapt their resilience and efficiency to different environmental conditions.
Combating heat stress and research opportunities
Heat stress can impact the epigenome during embryonic development, adulthood, and even across generations. As a result, the agricultural sector is searching for ways to lower the impact of the epigenomic changes caused by embryonic heat stress or develop genetic variants of monogastric livestock that are better adapted to heat stress as adult.
Chickens have emerged as pivotal models for understanding the epigenetic repercussions of heat stress (Kisliouk et al., 2010). Exposure to heat, particularly ‘conditioning’ embryos via elevated incubation temperatures, bestows an adaptive epigenetic memory in chickens, thus enhancing their resilience against subsequent heat stress (Zaboli et al., 2017; Yahav and Hurwitz, 1996). This adaptive mechanism speeds up the expression of HSPs and heat shock factor genes and bolsters interleukin and ROS-scavenging protein production (Yahav et al., 2004). Such responses are intricately tied to epigenetic mechanisms, including DNA methylation, histone modifications, and miRNA activity (Ramiah et al., 2022; Kisliouk et al., 2011). Notably, genomic regions associated with chicken domestication display differential methylation patterns, suggesting alterations stemming from selection pressures (Li et al., 2015; Nätt et al., 2012). Conditioning through modified embryonic incubation holds promise as a method to produce poultry that are better equipped to endure heat stress.
In parallel to conditioning embryos with heat, there’s growing interest in “epi-nutrients”. Epi-nutrients are essential nutrients that modulate the epigenome, especially DNA methylation during embryonic development (Kussmann and Bladeren, 2011; Delcurto et al., 2013). These include vital nutrients like vitamin B12, choline, and folate (Mazzio and Soliman, 2014). It is evident that epi-nutrient like folate is present in the bovine oviductal fluid during the oestrous cycle and bovine oviduct epithelial cells express folate receptors and transporters (Garcia et al., 2018). Research has shown that folic acid supplementation in pregnant mice can counter heat stress-induced neural tube defects and alter imprinted gene methylation (Shin and Shiota, 1999). A similar effect has been observed in human embryos when the expecting mothers consumed folic acid (Haggarty et al., 2013). In chickens, manganese-supplemented diets have demonstrated increased hatchability ratios and decreased mortality when exposed to heat stress. This protective effect was tied to heightened expression of the antioxidant enzyme manganese superoxide dismutase, which correlated with global DNA hypomethylation of histone H3 lysine 9 (H3K9) deacetylation in the embryonic heart. The reduced mortality is believed to stem from decreased cell death in the embryonic heart, potentially driven by elevated expression of the anti-apoptotic B-cell lymphoma 2 (Bcl-2) (Zhuo et al., 2017).
While conditioning and epi-nutrients offer encouraging outcomes for countering the epigenetic alterations caused by heat stress, understanding their long-term implications and mechanisms remains an ongoing pursuit. Further studies will help ascertain their full potential in increasing livestock resilience against heat stress, thus aiding in more sustainable agricultural practices.
Author contributions
EZ: Writing – original draft, Writing – review and editing. EV: Writing – original draft, Writing – review and editing. SK: Writing – review and editing. JR: Writing – review and editing. DS: Writing – original draft, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was funded by the WUR internal program KB34 Towards a Circular and Climate Neutral Society (2019-2022), project KB-34-2A-3 Epigenetics 2022.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Argaud, L., Ferry, T., Le, Q. H., Marfisi, A., Ciorba, D., Achache, P., et al. (2007). Short- and long-term outcomes of heatstroke following the 2003 heat wave in Lyon, France. Arch. Intern Med. 167 (20), 2177–2183. doi:10.1001/archinte.167.20.ioi70147
Ayo, J. O., Obidi, J. A., and Rekwot, P. I. (2011). Effects of heat stress on the well-being, fertility, and hatchability of chickens in the northern Guinea Savannah zone of Nigeria: a review. ISRN Veterinary Sci. 2011, 838606. doi:10.5402/2011/838606
Bannister, A. J., and Kouzarides, T. (2011). Regulation of chromatin by histone modifications. Cell Res. 21 (3), 381–395. doi:10.1038/cr.2011.22
Barlow, D. P. (2011). Genomic imprinting: a mammalian epigenetic discovery model. Annu. Rev. Genet. 45 (1), 379–403. doi:10.1146/annurev-genet-110410-132459
Becker, P. B., Ruppert, S., and Schütz, G. (1987). Genomic footprinting reveals cell type-specific DNA binding of ubiquitous factors. Cell 51 (3), 435–443. doi:10.1016/0092-8674(87)90639-8
Benayoun, B. A., Pollina, E. A., and Brunet, A. (2015). Epigenetic regulation of ageing: linking environmental inputs to genomic stability. Nat. Rev. Mol. Cell Biol. 16 (10), 593–610. doi:10.1038/nrm4048
Ben-Nun, O., Kisliouk, T., Marco, A., Rosenberg, T., and Meiri, N. (2022). Early-life thermal stress mediates long-term alterations in hypothalamic microglia. Glia 70 (4), 619–633. doi:10.1002/glia.24129
Bonev, B., and Cavalli, G. (2016). Organization and function of the 3D genome. Nat. Rev. Genet. 17 (11), 661–678. doi:10.1038/nrg.2016.112
Booth, L. N., and Brunet, A. (2016). The aging epigenome. Mol. Cell 62 (5), 728–744. doi:10.1016/j.molcel.2016.05.013
Brito, L. F., Maskal, J. M., Chen, S. Y., Oliveira, H. R., Graham, J. R., Schinckel, A. P., et al. (2025). 164 genome-wide DNA methylation profiling in in utero heat-stressed pigs as measured by whole-genome bisulfite sequencing.
Cao, Y., Ren, Q., Chang, S., Cui, W., Zhao, P., and Wang, Y. (2024). N6-methyladenosine RNA methylation modification regulates the transcription of viral-derived E (XSR) miRNAs to promote ALV-J replication. Vet. Microbiol. 298, 110218. doi:10.1016/j.vetmic.2024.110218
Cerneckis, J., Ming, G. L., Song, H., He, C., and Shi, Y. (2024). The rise of epitranscriptomics: recent developments and future directions. Trends Pharmacol. Sci. 45 (1), 24–38. doi:10.1016/j.tips.2023.11.002
Chakraborty, D., Sharma, N., Kour, S., Sodhi, S. S., Gupta, M. K., Lee, S. J., et al. (2022). Applications of omics technology for livestock selection and improvement. Front. Genet. 13, 774113. doi:10.3389/fgene.2022.774113
Chen, B., Yuan, C., Guo, T., Liu, J., Yang, B., and Lu, Z. (2023). Molecular mechanism of m6A methylation modification genes METTL3 and FTO in regulating heat stress in sheep. Int. J. Mol. Sci. 24 (15), 11926. doi:10.3390/ijms241511926
Chowdhary, S., Kainth, A. S., Pincus, D., and Gross, D. S. (2019). Heat shock factor 1 drives intergenic association of its target gene loci upon heat shock. Cell Rep. 26 (1), 18–28. doi:10.1016/j.celrep.2018.12.034
Cramer, T., Rosenberg, T., Kisliouk, T., and Meiri, N. (2019). Early-life epigenetic changes along the corticotropin-releasing hormone (CRH) gene influence resilience or vulnerability to heat stress later in life. Mol. Psychiatry 24 (7), 1013–1026. doi:10.1038/s41380-018-0280-5
Czech, B., Munafò, M., Ciabrelli, F., Eastwood, E. L., Fabry, M. H., Kneuss, E., et al. (2018). piRNA-Guided genome defense: from biogenesis to silencing. Annu. Rev. Genet. 52 (1), 131–157. doi:10.1146/annurev-genet-120417-031441
Dantas Machado, A. C., Zhou, T., Rao, S., Goel, P., Rastogi, C., Lazarovici, A., et al. (2015). Evolving insights on how cytosine methylation affects protein–DNA binding. Briefings Funct. Genomics. 14 (1), 61–73. doi:10.1093/bfgp/elu040
David, S.-A., Vitorino Carvalho, A., Gimonnet, C., Brionne, A., Hennequet-Antier, C., Piégu, B., et al. (2019). Thermal manipulation during embryogenesis impacts H3K4me3 and H3K27me3 histone marks in chicken hypothalamus. Front. Genet. 10, 1207. doi:10.3389/fgene.2019.01207
Delcurto, H., Wu, G., and Satterfield, M. C. (2013). Nutrition and reproduction: links to epigenetics and metabolic syndrome in offspring. Curr. Opin. Clin. Nutr. Metabolic Care 16 (4), 385–391. doi:10.1097/MCO.0b013e328361f96d
Dor, Y., and Cedar, H. (2018). Principles of DNA methylation and their implications for biology and medicine. Lancet 392 (10149), 777–786. doi:10.1016/S0140-6736(18)31268-6
Dose, A., Liokatis, S., Theillet, F.-X., Selenko, P., and Schwarzer, D. (2011). NMR profiling of histone deacetylase and acetyl-transferase activities in real time. ACS Chem. Biol. 6 (5), 419–424. doi:10.1021/cb1003866
Duempelmann, L., Skribbe, M., and Bühler, M. (2020). Small RNAs in the transgenerational inheritance of epigenetic information. Trends Genet. 36 (3), 203–214. doi:10.1016/j.tig.2019.12.001
Fitz-James, M. H., and Cavalli, G. (2022). Molecular mechanisms of transgenerational epigenetic inheritance. Nat. Rev. Genet. 23 (6), 325–341. doi:10.1038/s41576-021-00438-5
Garcia, E. V., Oliva, M. E., LeBlanc, J. G., and Barrera, A. D. (2018). Epi-nutrients in the oviductal environment: folate levels and differential gene expression of its receptors and transporters in the bovine oviduct. Theriogenology 119, 189–197. doi:10.1016/j.theriogenology.2018.07.008
Gelfman, S., Cohen, N., Yearim, A., and Ast, G. (2013). DNA-Methylation effect on cotranscriptional splicing is dependent on GC architecture of the exon–intron structure. Genome Res. 23 (5), 789–799. doi:10.1101/gr.143503.112
Goel, A., Ncho, C. M., and Choi, Y. H. (2021). Regulation of gene expression in chickens by heat stress. J. animal Sci. Biotechnol. 12 (1), 11. doi:10.1186/s40104-020-00523-5
Griffin, J. D., Saper, C. B., and Boulant, J. A. (2001). Synaptic and morphological characteristics of temperature-sensitive and -insensitive rat hypothalamic neurones. J. Physiology 537 (2), 521–535. doi:10.1111/j.1469-7793.2001.00521.x
Guo, Z., Lv, L., Liu, D., and Fu, B. (2018). Effects of heat stress on piglet production/performance parameters. Trop. Animal Health Prod. 50 (6), 1203–1208. doi:10.1007/s11250-018-1633-4
Gupta, S., Sharma, A., Joy, A., Dunshea, F. R., and Chauhan, S. S. (2022). The impact of heat stress on immune status of dairy cattle and strategies to ameliorate the negative effects. Animals open access J. MDPI. 13 (1), 107. doi:10.3390/ani13010107
Haggarty, P., Hoad, G., Campbell, D. M., Horgan, G. W., Piyathilake, C., and McNeill, G. (2013). Folate in pregnancy and imprinted gene and repeat element methylation in the offspring. Am. J. Clin. Nutr. 97 (1), 94–99. doi:10.3945/ajcn.112.042572
Halli, K., Yin, T., Koch, C., Krebs, S., and Konig, S. (2025). Heat stress induces specific methylation, transcriptomic and metabolic pattern in dairy cows and their female progeny. Sci. Rep. 15 (1), 17021. doi:10.1038/s41598-025-01082-3
He, Y., Taylor, R. L., Bai, H., Ashwell, C. M., Zhao, K., Li, Y., et al. (2023). Transgenerational epigenetic inheritance and immunity in chickens that vary in Marek's disease resistance. Poult. Sci. 102 (12), 103036. doi:10.1016/j.psj.2023.103036
Heng, J., Tian, M., Zhang, W., Chen, F., Guan, W., and Zhang, S. (2019). Maternal heat stress regulates the early fat deposition partly through modification of m6A RNA methylation in neonatal piglets. Cell Stress and Chaperones 24 (3), 635–645. doi:10.1007/s12192-019-01002-1
Henikoff, S., Dent, S. Y. R., Mostoslavsky, R., Wang, T., and Comai, L. (2025). Editorial: current insights in epigenetics and epigenomics. Front. Epigenetics Epigenomics 3, 3–2025. doi:10.3389/freae.2025.1566501
Hietakangas, V., and Sistonen, L. (2006). “Regulation of the heat shock response by heat shock transcription factors,” in Topics in current genetics (Berlin, Heidelberg: Springer), 1–34.
Huang, H., Lin, S., Garcia, B. A., and Zhao, Y. (2015). Quantitative proteomic analysis of histone modifications. Chem. Rev. 115 (6), 2376–2418. doi:10.1021/cr500491u
Huber, E., Notaro, U. S., Recce, S., Rodríguez, F. M., Ortega, H. H., Salvetti, N. R., et al. (2020). Fetal programming in dairy cows: effect of heat stress on progeny fertility and associations with the hypothalamic-pituitary-adrenal axis functions. Animal Reproduction Sci. 216, 106348. doi:10.1016/j.anireprosci.2020.106348
Ibeagha-Awemu, E. M., Kiefer, H., McKay, S., and Liu, G. E. (2022). Editorial: epigenetic variation influences on livestock production and disease traits. Front. Genet. 13, 942747. doi:10.3389/fgene.2022.942747
Jambhekar, A., Dhall, A., and Shi, Y. (2019). Roles and regulation of histone methylation in animal development. Nat. Rev. Mol. Cell Biol. 20 (10), 625–641. doi:10.1038/s41580-019-0151-1
Johnson, J. S., Sanz Fernandez, M. V., Patience, J. F., Ross, J. W., Gabler, N. K., Lucy, M. C., et al. (2015). Effects of in utero heat stress on postnatal body composition in pigs: II. Finishing phase. J. Animal Sci. 93 (1), 82–92. doi:10.2527/jas.2014-8355
Kainth, A. S., Chowdhary, S., Pincus, D., and Gross, D. S. (2021). Primordial super-enhancers: heat shock-induced chromatin organization in yeast. Trends Cell Biol. 31 (10), 801–813. doi:10.1016/j.tcb.2021.04.004
Kar, S. K. (2017). FeedOmics, an approach to evaluate the functional properties of protein containing feed ingredients.
Khanna, N., Hu, Y., and Belmont, A. S. (2014). HSP70 transgene directed motion to nuclear speckles facilitates heat shock activation. Curr. Biol. CB 24 (10), 1138–1144. doi:10.1016/j.cub.2014.03.053
Kisliouk, T., Cramer, T., and Meiri, N. (2017). Methyl CpG level at distal part of heat-shock protein promoter HSP70 exhibits epigenetic memory for heat stress by modulating recruitment of POU2F1-associated nucleosome-remodeling deacetylase (NuRD) complex. J. Neurochem. 141 (3), 358–372. doi:10.1111/jnc.14014
Kisliouk, T., Yosefi, S., and Meiri, N. (2011). MiR-138 inhibits EZH2 methyltransferase expression and methylation of histone H3 at lysine 27, and affects thermotolerance acquisition. Eur. J. Neurosci. 33 (2), 224–235. doi:10.1111/j.1460-9568.2010.07493.x
Kisliouk, T., Ziv, M., and Meiri, N. (2010). Epigenetic control of translation regulation: alterations in histone H3 lysine 9 post-translation modifications are correlated with the expression of the translation initiation factor 2B (Eif2b5) during thermal control establishment. Dev. Neurobiol. 70 (2), 100–113. doi:10.1002/dneu.20763
Klosin, A., Casas, E., Hidalgo-Carcedo, C., Vavouri, T., and Lehner, B. (2017). Transgenerational transmission of environmental information in C. elegans. Science. 356 (6335), 320–323. doi:10.1126/science.aah6412
Kolani, A., Adjrah, Y., Eklou-Lawson, M., Teteh, A., and Tona, K. (2018). Effects of dietary palm oil on production performance and serum parameters of laying hens. Int. J. Poult. Sci. 18 (1), 1–6. doi:10.3923/ijps.2019.1.6
Kussmann, M., and Bladeren, P. J. V. (2011). The extended nutrigenomics - understanding the interplay between the genomes of food, gut microbes, and human host. Front. Genet. 2 (MAY), 21. doi:10.3389/fgene.2011.00021
Labunskay, G., and Meiri, N. (2006). R-Ras3/(M-Ras) is involved in thermal adaptation in the critical period of thermal control establishment. J. Neurobiol. 66 (1), 56–70. doi:10.1002/neu.20191
Laporta, J., Khatib, H., and Zachut, M. (2024). Review: phenotypic and molecular evidence of inter- and trans-generational effects of heat stress in livestock mammals and humans. Animal 18 (Suppl. 2), 101121. doi:10.1016/j.animal.2024.101121
Li, Q., Wang, Y., Hu, X., Zhao, Y., and Li, N. (2015). Genome-wide mapping reveals conservation of promoter DNA methylation following chicken domestication. Sci. Rep. 5, 8748. doi:10.1038/srep08748
Li, W., Ma, X., Li, X., Zhang, X., Sun, Y., Ning, C., et al. (2024). Integrating single-cell RNA-seq and ATAC-seq analysis reveals uterine cell heterogeneity and regulatory networks linked to pimpled eggs in chickens. Int. J. Mol. Sci. 25 (24), 13431. doi:10.3390/ijms252413431
Lister, R., Pelizzola, M., Dowen, R. H., Hawkins, R. D., Hon, G., Tonti-Filippini, J., et al. (2009). Human DNA methylomes at base resolution show widespread epigenomic differences. Nature 462 (7271), 315–322. doi:10.1038/nature08514
Luger, K., Mäder, A. W., Richmond, R. K., Sargent, D. F., and Richmond, T. J. (1997). Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature 389 (6648), 251–260. doi:10.1038/38444
Marinova, Z., Leng, Y., Leeds, P., and Chuang, D.-M. (2011). Histone deacetylase inhibition alters histone methylation associated with heat shock protein 70 promoter modifications in astrocytes and neurons. Neuropharmacology 60 (7), 1109–1115. doi:10.1016/j.neuropharm.2010.09.022
E. A. Mazzio, and K. F. A. Soliman (2014). Epigenetics and nutritional environmental signals2014 (Oxford University Press).
McNabb, F. M. (1995). Thyroid hormones, their activation, degradation and effects on metabolism. J. Nutr. 125 (6 Suppl. l), 1773S-1776S–6S. doi:10.1093/jn/125.suppl_6.1773S
Meiri, N. (2008). 14-3-3epsilon expression is induced during the critical period of thermal control establishment. Dev. Neurobiol. 68 (1), 62–72. doi:10.1002/dneu.20571
Metzger, D. C. H., and Schulte, P. M. (2017). Persistent and plastic effects of temperature on DNA methylation across the genome of threespine stickleback (Gasterosteus aculeatus). Proc. R. Soc. B Biol. Sci. 284 (1864), 20171667. doi:10.1098/rspb.2017.1667
Moallem, U., Altmark, G., Lehrer, H., and Arieli, A. (2010). Performance of high-yielding dairy cows supplemented with fat or concentrate under hot and humid climates. J. Dairy Sci. 93 (7), 3192–3202. doi:10.3168/jds.2009-2979
Murray, K. O., Brant, J. O., Iwaniec, J. D., Sheikh, L. H., de Carvalho, L., Garcia, C. K., et al. (2021). Exertional heat stroke leads to concurrent long-term epigenetic memory, immunosuppression and altered heat shock response in female mice. J. Physiology 599 (1), 119–141. doi:10.1113/JP280518
Murray, K. O., Clanton, T. L., and Horowitz, M. (2022). Epigenetic responses to heat: from adaptation to maladaptation. Exp. Physiol. 107 (10), 1144–1158. doi:10.1113/EP090143
Nätt, D., Rubin, C.-J., Wright, D., Johnsson, M., Beltéky, J., Andersson, L., et al. (2012). Heritable genome-wide variation of gene expression and promoter methylation between wild and domesticated chickens. BMC Genomics 13, 59. doi:10.1186/1471-2164-13-59
Nawab, A., Ibtisham, F., Li, G., Kieser, B., Wu, J., Liu, W., et al. (2018). Heat stress in poultry production: mitigation strategies to overcome the future challenges facing the global poultry industry. J. Therm. Biol. 78, 131–139. doi:10.1016/j.jtherbio.2018.08.010
Negrón-Pérez, V. M., Fausnacht, D. W., and Rhoads, M. L. (2019). Invited review: management strategies capable of improving the reproductive performance of heat-stressed dairy cattle. J. Dairy Sci. 102 (12), 10695–10710. doi:10.3168/jds.2019-16718
Nwunuji, T. P., Mayowa, O. O., Yusoff, S. M., Bejo, S.-K., Salisi, S., and Mohd, E. A. W. (2014). The ameliorative effect of ascorbic acid on the oxidative status, live weight and recovery rate in road transport stressed goats in a hot humid tropical environment. Animal Sci. J. 85 (5), 611–616. doi:10.1111/asj.12174
O'Brien, J., Hayder, H., Zayed, Y., and Peng, C. (2018). Overview of MicroRNA biogenesis, mechanisms of actions, and circulation. Front. Endocrinol. 9, 402. doi:10.3389/fendo.2018.00402
Oke, O. E. (2018). Evaluation of physiological response and performance by supplementation of Curcuma longa in broiler feed under hot humid tropical climate. Trop. Animal Health Prod. 50 (5), 1071–1077. doi:10.1007/s11250-018-1532-8
Oke, O. E., Oyelola, O. B., Iyasere, O. S., Njoku, C. P., Oso, A. O., Oso, O. M., et al. (2021). In ovo injection of Black cumin (Nigella sativa) extract on hatching and post hatch performance of thermally challenged broiler chickens during incubation. Poult. Sci. 100 (3), 100831. doi:10.1016/j.psj.2020.10.072
Pandey, N., and Yadav, A. K. (2025). “Epigenomic insights into model animals and their application in understanding livestock systems in veterinary science,” in Bioinformatics in veterinary science: vetinformatics. Editors J.-M. Kim, and R. K. Pathak (Singapore: Springer Nature Singapore), 63–85.
Parsell, D. A., Kowal, A. S., Singer, M. A., and Lindquist, S. (1994). Protein disaggregation mediated by heat-shock protein Hsp104. Nature 372 (6505), 475–478. doi:10.1038/372475a0
Piestun, Y., Halevy, O., Shinder, D., Ruzal, M., Druyan, S., and Yahav, S. (2011). Thermal manipulations during broiler embryogenesis improves post-hatch performance under hot conditions. J. Therm. Biol. 36 (7), 469–474. doi:10.1016/j.jtherbio.2011.08.003
Piestun, Y., Halevy, O., and Yahav, S. (2009). Thermal manipulations of broiler embryos--the effect on thermoregulation and development during embryogenesis. Poult. Sci. 88 (12), 2677–2688. doi:10.3382/ps.2009-00231
Piestun, Y., Shinder, D., Ruzal, M., Halevy, O., Brake, J., and Yahav, S. (2008). Thermal manipulations during broiler embryogenesis: effect on the acquisition of thermotolerance. Poult. Sci. 87 (8), 1516–1525. doi:10.3382/ps.2008-00030
Poorghasemi, M., Seidavi, A., Qotbi, A. A. A., Laudadio, V., and Tufarelli, V. (2013). Influence of dietary fat source on growth performance responses and carcass traits of broiler chicks. Asian-Australasian J. Animal Sci. 26 (5), 705–710. doi:10.5713/ajas.2012.12633
Rajkumar, U., Vinoth, A., Shanmugam, M., Rajaravindra, K. S., and Rama Rao, S. V. (2017). Effect of increased incubation temperature on hsp 90 and 60 gene expressions in coloured broiler chickens. J. Appl. Animal Res. 45 (1), 298–303. doi:10.1080/09712119.2016.1174128
Ramiah, S. K., Balakrishnan, K. N., Subramaniam, Y., Iyasere, O. S., and Idrus, Z. (2022). Effects of thermal manipulation on mRNA regulation of response genes regarding improvement of thermotolerance adaptation in chickens during embryogenesis. Animals 12 (23), 3354. doi:10.3390/ani12233354
Raza, S. H. A., Abdelnour, S. A., Dhshan, A. I. M., Hassanin, A. A., Noreldin, A. E., Albadrani, G. M., et al. (2021). Potential role of specific microRNAs in the regulation of thermal stress response in livestock. J. Therm. Biol. 96, 102859. doi:10.1016/j.jtherbio.2021.102859
Ren, T., Xu, M., Du, X., Wang, Y., Loor, J. J., Lei, L., et al. (2024). Research progress on the role of M6A in regulating economic traits in livestock. Int. J. Mol. Sci. 25 (15), 8365. doi:10.3390/ijms25158365
Ringseis, R., and Eder, K. (2022). Heat stress in pigs and broilers: role of gut dysbiosis in the impairment of the gut-liver axis and restoration of these effects by probiotics, prebiotics and synbiotics. J. Animal Sci. Biotechnol. 13 (1), 126–16. doi:10.1186/s40104-022-00783-3
Rocha, A. C. G., Patrone, L. G. A., Cristina-Silva, C., Silva, K. S. C., Bícego, K. C., Szawka, R. E., et al. (2022). Metabolic and respiratory chemosensitivity and brain monoaminergic responses to cold exposure in chicks subjected to thermal manipulation during incubation. J. Therm. Biol. 109, 103317. doi:10.1016/j.jtherbio.2022.103317
Rosenberg, T., Kisliouk, T., Cramer, T., Shinder, D., Druyan, S., and Meiri, N. (2020). Embryonic heat conditioning induces TET-dependent cross-tolerance to hypothalamic inflammation later in life. Front. Genet. 11, 767. doi:10.3389/fgene.2020.00767
Rosenberg, T., Marco, A., Kisliouk, T., Haron, A., Shinder, D., Druyan, S., et al. (2022). Embryonic heat conditioning in chicks induces transgenerational heat/immunological resilience via methylation on regulatory elements. FASEB J. 36 (7), e22406. doi:10.1096/fj.202101948R
Ross, J. W., Hale, B. J., Gabler, N. K., Rhoads, R. P., Keating, A. F., Baumgard, L. H., et al. (2015). Physiological consequences of heat stress in pigs. Animal Prod. Sci. 55 (12), 1381–1390. doi:10.1071/an15267
Ross, J. W., Hale, B. J., Seibert, J. T., Romoser, M. R., Adur, M. K., Keating, A. F., et al. (2017). Physiological mechanisms through which heat stress compromises reproduction in pigs. Mol. Reproduction Dev. 84 (9), 934–945. doi:10.1002/mrd.22859
Rossnerova, A., Izzotti, A., Pulliero, A., Bast, A., Rattan, S. I. S., and Rossner, P. (2020). The molecular mechanisms of adaptive response related to environmental stress. Int. J. Mol. Sci. 21 (19), 7053. doi:10.3390/ijms21197053
Saeed, M., Abbas, G., Alagawany, M., Kamboh, A. A., Abd El-Hack, M. E., Khafaga, A. F., et al. (2019). Heat stress management in poultry farms: a comprehensive overview. J. Therm. Biol. 84, 414–425. doi:10.1016/j.jtherbio.2019.07.025
Saiz del Barrio, A., Mansilla, W. D., Navarro-Villa, A., Mica, J. H., Smeets, J. H., den Hartog, L. A., et al. (2020). Effect of mineral and vitamin C mix on growth performance and blood corticosterone concentrations in heat-stressed broilers. J. Appl. Poult. Res. 29 (1), 23–33. doi:10.1016/j.japr.2019.11.001
Sajjanar, B., Krishnaswamy, N., Saxena, V. K., and Dhara, S. K. (2025). Stress responses to changing environmental factors in the domestic animals: an epigenetic perspective. J. Anim. Physiol. Anim. Nutr. Berl. 109, 1000–1016. doi:10.1111/jpn.14115
Shein, N. A., Horowitz, M., Alexandrovich, A. G., Tsenter, J., and Shohami, E. (2005). Heat acclimation increases hypoxia-inducible factor 1alpha and erythropoietin receptor expression: implication for neuroprotection after closed head injury in mice. J. Cereb. Blood Flow. Metab. 25 (11), 1456–1465. doi:10.1038/sj.jcbfm.9600142
Sherif, Z. A., Ogunwobi, O. O., and Ressom, H. W. (2024). Mechanisms and technologies in cancer epigenetics. Front. Oncol. 14, 1513654. doi:10.3389/fonc.2024.1513654
Shin, J. H., and Shiota, K. (1999). Folic acid supplementation of pregnant mice suppresses heat-induced neural tube defects in the offspring. J. Nutr. 129, 2070–3. doi:10.1093/jn/129.11.2070
Sindhu, P., Magotra, A., Sindhu, V., and Chaudhary, P. (2024). Unravelling the impact of epigenetic mechanisms on offspring growth, production, reproduction and disease susceptibility. Zygote 32 (3), 190–206. doi:10.1017/S0967199424000224
Spencer, J. D., Boyd, R. D., Cabrera, R., and Allee, G. L. (2003). Early weaning to reduce tissue mobilization in lactating sows and milk supplementation to enhance pig weaning weight during extreme heat stress. J. Animal Sci. 81 (8), 2041–2052. doi:10.2527/2003.8182041x
Swygert, S. G., and Peterson, C. L. (2014). Chromatin dynamics: interplay between remodeling enzymes and histone modifications. Biochimica Biophysica Acta 1839 (8), 728–736. doi:10.1016/j.bbagrm.2014.02.013
Tan, B., Hong, L., Xiao, L., Wu, J., Lu, G., Wang, S., et al. (2025). Rewiring of 3D chromatin topology orchestrates transcriptional reprogramming in muscle fiber-type specification and transformation. Nat. Commun. 16 (1), 5833. doi:10.1038/s41467-025-60866-3
Tetievsky, A., and Horowitz, M. (1985). “Posttranslational modifications in histones underlie heat acclimation-mediated cytoprotective memory,”J. Appl. Phys. 109. 1552–1561. doi:10.1152/japplphysiol.00469.2010
Therizols, P., Illingworth, R. S., Courilleau, C., Boyle, S., Wood, A. J., and Bickmore, W. A. (2014). Chromatin decondensation is sufficient to alter nuclear organization in embryonic stem cells. Sci. (New York, NY) 346 (6214), 1238–1242. doi:10.1126/science.1259587
Tzschentke, B., and Basta, D. (2002). Early development of neuronal hypothalamic thermosensitivity in birds: influence of epigenetic temperature adaptation. Comp. Biochem. Physiology Part A Mol. and Integr. Physiology 131 (4), 825–832. doi:10.1016/s1095-6433(02)00020-x
Unoki, M. (2019). Recent insights into the mechanisms of <Em>De novo</Em> and maintenance of DNA methylation in mammals. IntechOpen.
Vinoth, A., Thirunalasundari, T., Shanmugam, M., Uthrakumar, A., Suji, S., and Rajkumar, U. (2018). Evaluation of DNA methylation and mRNA expression of heat shock proteins in thermal manipulated chicken. Cell Stress Chaperones 23 (2), 235–252. doi:10.1007/s12192-017-0837-2
Wan, Q.-L., Meng, X., Dai, W., Luo, Z., Wang, C., Fu, X., et al. (2021). N 6 -methyldeoxyadenine and histone methylation mediate transgenerational survival advantages induced by hormetic heat stress. Sci. Adv. 7 (1), eabc3026. doi:10.1126/sciadv.abc3026
Wang, S.-H., Cheng, C.-Y., Chen, C.-J., Chan, H.-L., Chen, H.-H., Tang, P.-C., et al. (2019). Acute heat stress changes protein expression in the testes of a broiler-type strain of Taiwan country chickens. Anim. Biotechnol. 30 (2), 129–145. doi:10.1080/10495398.2018.1446972
Wettemann, R. P., and Bazer, F. W. (1985). Influence of environmental temperature on prolificacy of pigs. J. reproduction Fertil. Suppl. 33, 199–208.
Wu, J., Zhang, W., and Li, C. (2020). Recent advances in genetic and epigenetic modulation of animal exposure to high temperature. Front. Genet. 11, 653. doi:10.3389/fgene.2020.00653
Wu, X., and Zhang, Y. (2017). TET-Mediated active DNA demethylation: mechanism, function and beyond. Nat. Rev. Genet. 18 (9), 517–534. doi:10.1038/nrg.2017.33
Xie, S., Hagen, D., Becker, G. M., Davenport, K. M., Shira, K. A., Stegemiller, M. R., et al. (2025). Analyzing the relationship of RNA and DNA methylation with gene expression. Genome Biol. 26 (1), 140. doi:10.1186/s13059-025-03617-3
Xing, Y., Tang, Y., Chen, Q., Chen, S., Li, W., Mi, S., et al. (2024). The role of RNA epigenetic modification-related genes in the immune response of cattle to mastitis induced by Staphylococcus aureus. Anim. Biosci. 37 (7), 1141–1155. doi:10.5713/ab.23.0323
Yahav, S., and Hurwitz, S. (1996). Induction of thermotolerance in Male broiler chickens by temperature conditioning at an early age. Poult. Sci. 75 (3), 402–406. doi:10.3382/ps.0750402
Yahav, S., Straschnow, A., Luger, D., Shinder, D., Tanny, J., and Cohen, S. (2004). Ventilation, sensible heat loss, broiler energy, and water balance under harsh environmental conditions. Poult. Sci. 83 (2), 253–258. doi:10.1093/ps/83.2.253
Yang, Y., Chen, Y., Hu, L., Zhang, C., Chen, G., Hou, L., et al. (2025). Molecular regulation of whole genome DNA methylation in heat stress response of dairy cows. BMC Genomics 26 (1), 464. doi:10.1186/s12864-025-11683-x
Yossifoff, M., Kisliouk, T., and Meiri, N. (2008). Dynamic changes in DNA methylation during thermal control establishment affect CREB binding to the brain-derived neurotrophic factor promoter. Eur. J. Neurosci. 28 (11), 2267–2277. doi:10.1111/j.1460-9568.2008.06532.x
Zaboli, G.-R., Rahimi, S., Shariatmadari, F., Torshizi, M. A. K., Baghbanzadeh, A., and Mehri, M. (2017). Thermal manipulation during pre and post-hatch on thermotolerance of Male broiler chickens exposed to chronic heat stress. Poult. Sci. 96 (2), 478–485. doi:10.3382/ps/pew344
Zhou, L., He, B., Deng, J., Pang, S., and Tang, H. (2019). Histone acetylation promotes long-lasting defense responses and longevity following early life heat stress. PLoS Genet. 15 (4), e1008122. doi:10.1371/journal.pgen.1008122
Zhou, P., Wang, W., Li, J., Zheng, Z., Du, X., Fu, L., et al. (2024). Identification of novel transcription factors regulated by H3K27 acetylation in myogenic differentiation of porcine skeletal muscle satellite cells. FASEB J. 38 (21), e70144. doi:10.1096/fj.202401285RR
Zhu, J.-Q., Liu, J.-H., Liang, X.-W., Xu, B.-Z., Hou, Y., Zhao, X.-X., et al. (2008). Heat stress causes aberrant DNA methylation of H19 and Igf-2r in mouse blastocysts. Mol. Cells 25 (2), 211–215. doi:10.1016/s1016-8478(23)17572-8
Zhu, L., Marjani, S. L., and Jiang, Z. (2021). The epigenetics of gametes and early embryos and potential long-range consequences in livestock species—filling in the picture with epigenomic analyses. Front. Genet. 12, 557934. doi:10.3389/fgene.2021.557934
Zhuo, Z., Lamont, S. J., and Abasht, B. (2017). RNA-seq analyses identify frequent allele specific expression and no evidence of genomic imprinting in specific embryonic tissues of chicken. Sci. Rep. 7 (1), 11944. doi:10.1038/s41598-017-12179-9
Keywords: heat stress, epigenome, livestock, chicken, pig
Citation: Zaccaria E, van der Valk E, Kar SK, Rebel JMJ and Schokker D (2025) Mini review: Studying epigenomic alterations can shed light on coping and adaptive abilities during heat stress in monogastric livestock. Front. Genet. 16:1561804. doi: 10.3389/fgene.2025.1561804
Received: 16 January 2025; Accepted: 18 July 2025;
Published: 01 August 2025.
Edited by:
Eveline M. Ibeagha-Awemu, Agriculture and Agri-Food Canada (AAFC), CanadaReviewed by:
Geoffrey E. Dahl, University of Florida, United StatesCopyright © 2025 Zaccaria, van der Valk, Kar, Rebel and Schokker. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dirkjan Schokker, ZGlya2phbi5zY2hva2tlckB3dXIubmw=