 María Aránzazu Penedo
María Aránzazu Penedo Héctor Cañeque-Rufo
Héctor Cañeque-Rufo Esther Gramage
Esther Gramage Gonzalo Herradón
Gonzalo Herradón- 1Department of Health and Pharmaceutical Sciences, School of Pharmacy, Universidad San Pablo-CEU, CEU Universities, Madrid, Spain
- 2Red de Investigación en Atención Primaria de Adicciones, Instituto de Salud Carlos III, MICINN and FEDER, Madrid, Spain
- 3Department of Chemistry and Biochemistry, School of Pharmacy, Universidad San Pablo-CEU, CEU Universities, Madrid, Spain
- 4Instituto de Estudios de las Adicciones, Universidad San Pablo-CEU, CEU Universities, Madrid, Spain
Microglial cells are key mediators of ethanol-induced neuroinflammation through the release of proinflammatory cytokines and activation of Toll-like receptors. Recently, the signaling pathway initiated by the interaction of the neurotrophic factors pleiotrophin (PTN) and midkine (MK) with receptor-type protein tyrosine phosphatase β/ζ (RPTPβ/ζ) has emerged as a pharmacological target in ethanol-induced neuroinflammatory and neurodegenerative processes. However, the underlying molecular mechanisms remain unclear. In this study, we developed a human co-culture system composed of differentiated SH-SY5Y neuronal cells and HMC3 microglial cells to simulate microglial-neuronal interactions during ethanol exposure. In HMC3 cells, PTN mRNA expression levels were significantly upregulated by ethanol exposure, whereas MK levels were not altered. In contrast, ethanol exposure caused a significant downregulation of MK expression in co-cultures. In general, ethanol increased the expression of inflammatory genes in monocultures of HMC3 cells but not in SH-SY5Y cells. In addition, ethanol exposure caused a highly significant upregulation of TLR3 and TLR4 in HMC3 cells, which was absent in co-cultures. We also observed a significant attenuation of ethanol-induced increases of inflammatory markers such as IL-1β and CCL2 in co-cultures, indicating the regulatory role of neuronal-microglial interactions. In conclusion, our study provides novel insights into the modulatory actions of microglial-neuronal interactions in ethanol-induced neuroimmune responses and suggests the therapeutic potential of the PTN/RPTPβ/ζ signaling pathway to prevent the deleterious effects of alcohol on the brain.
1 Introduction
Alcohol consumption remains a critical public health concern due to its profound and lasting effects on the brain. Excessive intake has been shown to exert direct neurotoxic effects and to trigger persistent neuroinflammatory processes that contribute to neurodegeneration (Pascual et al., 2021). Neuroimmune responses, particularly those mediated by sustained microglial activation and the synthesis and release of pro-inflammatory cytokines, play a central role in this chronic inflammatory state, which can persist even after alcohol withdrawal (Crews et al., 2024; Crews et al., 2011). Over time, this environment may disrupt synaptic plasticity and neurogenesis, leading to cognitive and motor impairments, and contributing to the onset and progression of various neurodegenerative disorders, including alcohol-related cognitive decline (Crews, 2012; Pascual et al., 2018; Walter and Crews, 2017).
Understanding the molecular mechanisms involved in alcohol-induced neuroinflammation in humans presents considerable ethical and methodological challenges. Numerous studies have reported alterations at both the cellular and microenvironmental levels of the central nervous system (CNS), as well as behavioral changes associated with alcohol use disorder (AUD) (Pascual et al., 2018; Hartung, 2009; Walter and Crews, 2017). However, most of these investigations have focused on descriptive characterizations, and the specific mechanisms driving these changes are still poorly understood. In this context, in vitro cellular models offer a valuable alternative for exploring molecular pathways and evaluating novel therapeutic strategies (Goshi et al., 2020). Particularly promising are co-culture systems, which recreate physiologically relevant interactions between key CNS cell types (Gresa-Arribas et al., 2012; Roqué and Costa, 2017). Among them, the interplay between dopaminergic neurons and microglia appears to be especially important in mediating the inflammatory and neurotoxic responses to ethanol exposure (Alfonso-Loeches et al., 2010; Correa et al., 2013; Kraft and Harry, 2011).
Recently, the signaling pathway initiated by the interaction of pleiotrophin (PTN) and midkine (MK) with receptor-type protein tyrosine phosphatase β/ζ (RPTPβ/ζ), also known as PTPRZ1, has emerged as a molecular axis of interest in neuroinflammatory and neurodegenerative processes. PTN and MK are neurotrophic factors and endogenous inhibitors of the tyrosine phosphatase activity of RPTPβ/ζ (Herradon et al., 2019). This receptor regulates interactions between glial and neuronal cells (Del Campo et al., 2021), and its pharmacological modulation is being explored as a promising strategy to mitigate alcohol-induced neuropathology (Fernández-Calle et al., 2018; Galán-LLario et al., 2023a; Galán-LLario et al., 2023b; Galán-LLario et al., 2024; Rodríguez-Zapata et al., 2023). To further characterize the functional roles of RPTPβ/ζ, our group designed and synthesized MY10, a selective inhibitor that interacts with its intracellular PD1 domain, effectively inactivating the tyrosine phosphatase activity of RPTPβ/ζ (Pastor et al., 2018).
Preclinical studies have shown that treatment with MY10 reduces alcohol consumption and blocks conditioned place preference, suggesting a targeted action on alcohol-related reward mechanisms (Calleja-Conde et al., 2020; Fernández-Calle et al., 2018). Additionally, MY10 modulates gene expression in the prefrontal cortex, likely through the regulation of PTN and MK signaling pathways (Fernández-Calle et al., 2017; Fernández-Calle et al., 2020; Herradón and Pérez-García, 2014). In addition, MY10 has demonstrated significant neuroprotective effects against ethanol-induced neuroinflammation. Recent findings demonstrate that MY10 also prevents ethanol-induced impairments in adolescent hippocampal neurogenesis, attenuates microglial reactivity, and modulates the organization of perineuronal nets (PNNs), with some effects showing sex-dependent variability (Galán-Llario et al., 2023a; Galán-LLario et al., 2023b; Galán-LLario et al., 2024). Further supporting its mechanism of action, PTN overexpression has been found to reproduce both the neuroprotective and ethanol-suppressive effects of MY10 (Galán-Llario et al., 2024).
The present study uses a human in vitro co-culture model composed of dopaminergic neurons and microglia, designed to simulate ethanol-induced neuroinflammation and to characterize the role of RPTPβ/ζ in this process. This model enables the study of neuron–glia crosstalk, a fundamental component of ethanol-driven neuroinflammatory processes, and allows the evaluation of pharmacological candidates such as MY10. The findings provide substantial evidence that neuron–microglia interactions critically influence the responses to ethanol, representing a meaningful advancement in the development of physiologically relevant platforms for investigating alcohol-induced neuropathology.
2 Materials and methods
2.1 Cell cultures
Human SH-SY5Y neuroblastoma cells and HMC3 human microglial cells (generously provided by Dr. Marta del Campo, ATCC) were used in this study.
2.1.1 SH-SY5Y cells
SH-SY5Y cells were cultured in T-75 flasks at 37°C in a humidified atmosphere containing 5% CO2. To induce dopaminergic neuronal differentiation (Figure 1), cells were treated with 1:500 µM retinoic acid (RA, Thermo Fisher, United States) for 3–4 days in proliferation medium consisting of Dulbecco’s Modified Eagle’s Medium (DMEM, Biowest, France) supplemented with 10% Fetal Bovine Serum (FBS, Sigma-Aldrich, United States) and 1% penicillin/streptomycin (100 U/ml-100 μg/mL; Gibco, United States). After this initial phase, 5 × 105 SH-SY5Y cells/well were seeded into 6-well treated culture plates (WVR, United States) and maintained additionally for 7 days in DMEM proliferation medium with RA. Medium was replaced every 2–3 days to maintain optimal differentiation conditions. Neuronal maturation was subsequently achieved by incubating cells for 14 days in maturation medium composed of Neurobasal (Gibco, United States), B-27 supplement (Gibco, United States), and 1% penicillin/streptomycin (Sigma-Aldrich, United States) with RA.
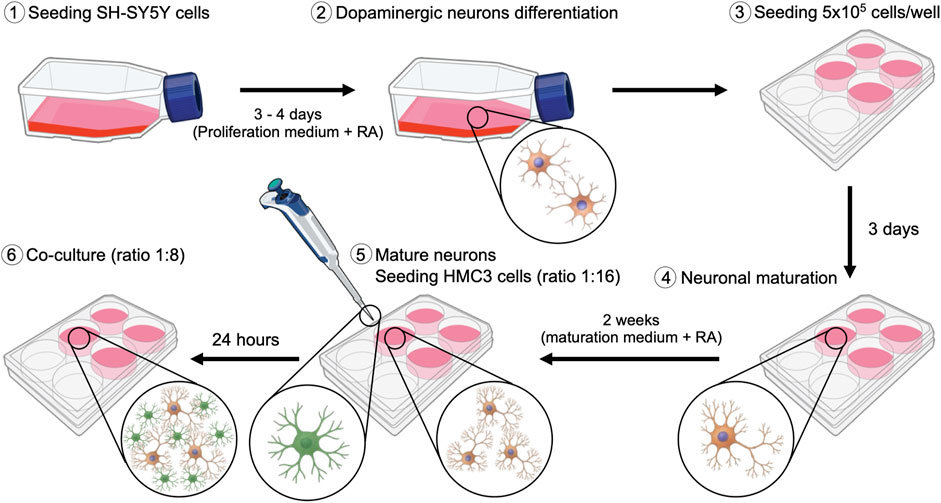
Figure 1. Schematic representation of the protocol implemented for the establishment of the SH-SY5Y and HMC3 co-culture.
2.1.2 HMC3 cells
5 × 105 HMC3 microglial cells/well were cultured in 6-well plates using proliferation medium and maintained at 37°C in a humidified incubator with 5% CO2.
2.1.3 SH-SY5Y and HMC3 co-culture
Once SH-SY5Y cells had ceased proliferation and exhibited mature neuronal morphology, HMC3 microglial cells were added to the culture at an initial ratio of 1:16 (HMC3:SH-SY5Y). The co-culture was maintained in neuronal maturation medium for 24 h to allow HMC3 cells to adhere and establish cellular contacts and to acclimate to promote adaptation. Subsequently, HMC3 cells were permitted to proliferate within the co-culture until a final ratio of 1:8 (HMC3:SH-SY5Y) was reached.
2.2 Immunofluorescence
Immunofluorescence staining assays were performed to confirm the SH-SY5Y and HMC3 co-culture (see Supplementary Figure S1). Cells were washed with phosphate-buffered saline (PBS, pH 7.4) and fixed with 4% paraformaldehyde for 5 min at room temperature. Cells were then permeabilized and blocked using a solution containing 10% Normal Goat Serum (NGS, Abcam, United States) and 10% Bovine Serum Albumin (BSA, Sigma-Aldrich, United States), and 0,2% Triton X-100 in PBS for 1 h. Neuronal cells were identified using a primary antibody against MAP-2 (Ab221693, Abcam, United Kingdom), while microglial cells were labeled with an anti-IBA1 antibody (Ab5076, Abcam, United Kingdom). Secondary antibody incubation was carried out using Alexa Fluor 488-conjugated anti-rabbit IgG (A21206, Thermo Fisher, United States) and Alexa Fluor 555-conjugated anti-goat IgG (705-565-147, Jackson Immuno Research, United Kingdom). Nuclear staining was performed with DAPI (D1306, Invitrogen, United States). Coverslips were mounted with mounting medium and imaged using a fluorescence microscope (Leica DM5500).
2.3 Determination of the expression levels of neurotrophic and immune genes
Cultures were exposed to 100 mM ethanol for 24 h, using a 4% ethanol chamber to maintain ethanol concentration (Coleman, et al., 2017). Three experimental groups were evaluated: SH-SY5Y monoculture, HMC3 monoculture, and SH-SY5Y + HMC3 co-culture. To evaluate the potential effects of RPTPβ/ζ inhibition, cultures were treated with MY10 at concentrations of 0.1 µM, 1 μM, and 10 μM, diluted in DMSO (final concentrations range: 0.0005%–0.05%) (Sigma-Aldrich, United States). Control groups received as vehicle 0.05% DMSO. Each experimental condition was tested in five independent biological replicates, with three technical replicates per group.
Following treatment, adherent cells were washed with PBS (pH 7.4) and lysed using TRIzol Reagent (Thermo Fisher, United States). Total RNA was isolated using the RNeasy Mini Kit (Qiagen, Germany), following the manufacturers protocol. First-strand cDNA was synthesized using the First-Strand cDNA Synthesis Kit (NZYTech, Portugal). Quantitative real-time PCR (qPCR) analysis was performed using the SYBR Green detection method (#1725272, Bio-Rad, United States) on a CFX96 Real-Time PCR Detection System (Bio-Rad, United States). The relative expression of each target gene was normalized using three housekeeping genes: RPL30, HPRT, and PKM. Primer sequences designed and used for amplification are listed in Table 1.
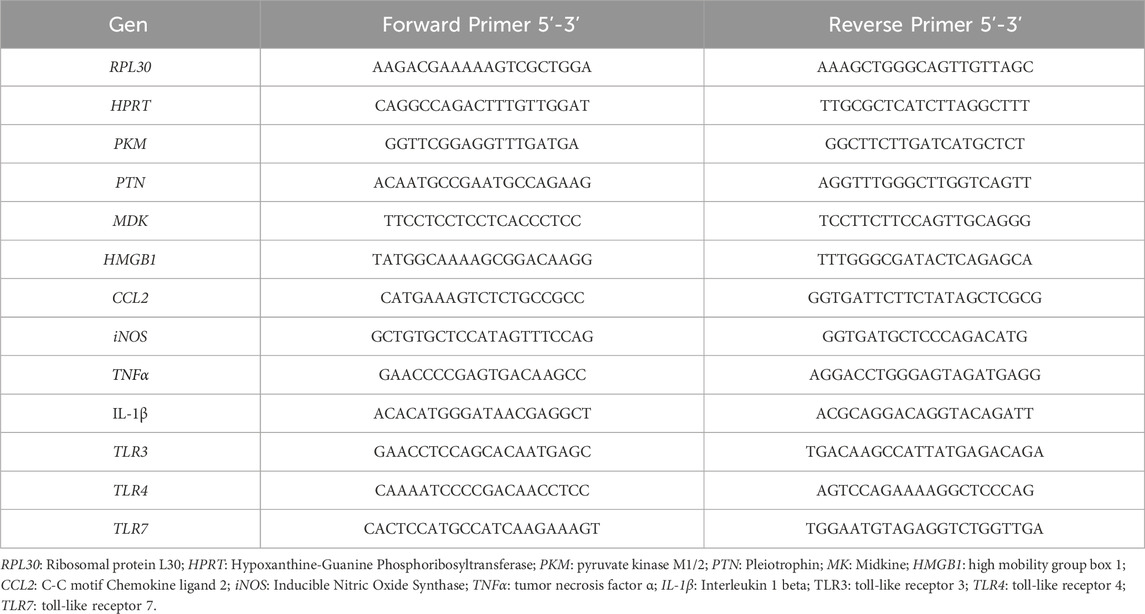
Table 1. Primer sequences used for qPCR analysis.
2.4 Statistical analysis
All statistical analyses were performed using GraphPad Prism version 10 (GraphPad Software, United States). Data are presented as mean ± standard error of the mean (SEM). T-Student tests were used for targeted analysis of genes associated with the RPTPβ/ζ pathway. For experiments involving multiple conditions, two-way ANOVA were conducted considering ethanol exposure and MY10 treatment as variables, followed by Bonferroni’s Post-hoc correction for multiple comparisons. A P-value <0.05 was considered statistically significant.
3 Results
3.1 Modulation of the components of the PTN/MK/RPTPβζ axis following ethanol exposure
We evaluated the transcriptional response of the RPTPβζ endogenous ligands PTN and MK, after 24-h exposure to 100 mM ethanol in three in vitro cellular models (Figure 2). Our results revealed that ethanol did not induce significant changes in the expression levels of these cytokines in SH-SY5Y dopaminergic neurons (Figures 2a,d). However, in HMC3 cells, PTN mRNA expression levels were significantly upregulated compared to untreated controls (Figure 2b; t = 3.149; P = 0.0042). In contrast, MK mRNA expression levels were not altered by ethanol exposure in microglial cells (Figure 2e). Interestingly, in the neuron-microglia co-culture system, ethanol exposure caused a significant downregulation of MK expression (Figure 2f; t = 2.162; P = 0.04), without altering PTN levels (Figure 2c). The data suggest that neuron-microglia interactions modulate the microglial response to ethanol and highlight the importance of cellular crosstalk in shaping proinflammatory signaling.
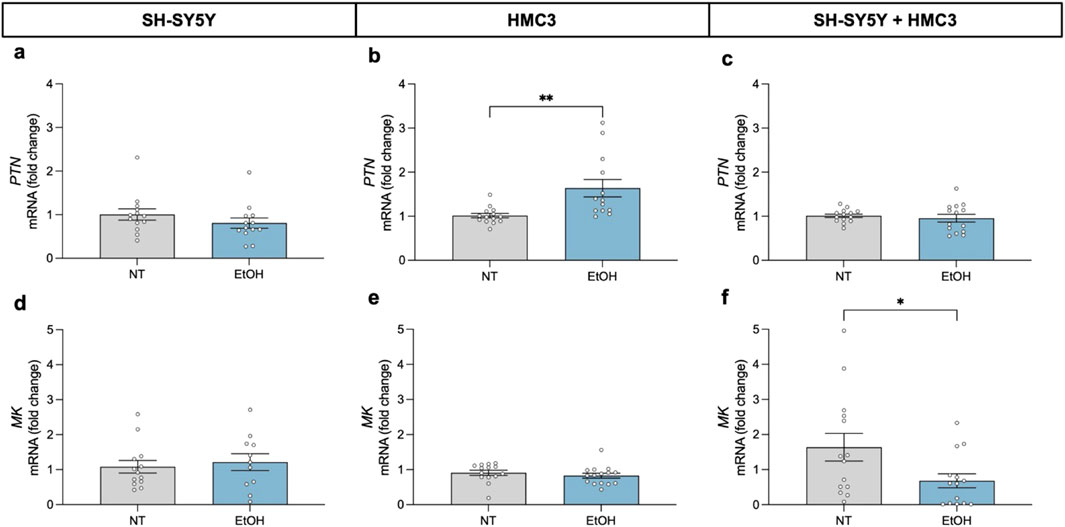
Figure 2. Effect of 24-h ethanol exposure (100 mM) on PTN and MK expression in neurons, microglia and co-cultures. PTN mRNA expression in SH-SY5Y cells (a), HMC3 cells (b), and co-culture (c). MK mRNA expression in SH-SY5Y cells (d), HMC3 cells (e), and co-culture (f). Data are presented as mean ± SEM. *P < 0,05; **P < 0,01. NT: no treatment; EtOH: ethanol.
3.2 Impact of pharmacological inhibition of RPTPβζ by MY10 on ethanol-induced neuroinflammation
To investigate the potential modulatory actions of MY10 on ethanol effects, we analyzed the expression of key proinflammatory genes in SH-SY5Y cells, HMC3 cells, and the co-culture of both in response to ethanol and different concentrations of MY10 (0.1 μM, 1 μM, and 10 μM) (Figure 3). Two-way ANOVA did not reveal significant effects of ethanol or the treatment with MY10 on the mRNA expression levels of HMGB1 in any of the cell systems evaluated (Figures 3a–c). Interestingly, ethanol exposure significantly affected CCL2 mRNA levels (Figures 3d–f). Specifically, ethanol caused a significant upregulation of CCL2 levels in HMC3 microglial cells (F (1,107) = 29.62; P < 0.0001; Figure 3e). However, a significant downregulation was observed in SH-SY5Y neuronal cells (F (1,110) = 9.931; P = 0.0021; Figure 3d), and in the co-culture of neurons and microglia (F (1,106) = 10.72; P = 0.0014; Figure 3f). The two-way ANOVA performed with iNOS mRNA levels data only rendered a significant effect of ethanol exposure in SH-SY5Y cells (F (1,104) = 12.9; P = 0.0005), in which iNOS expression was upregulated by ethanol exposure (Figure 3g).
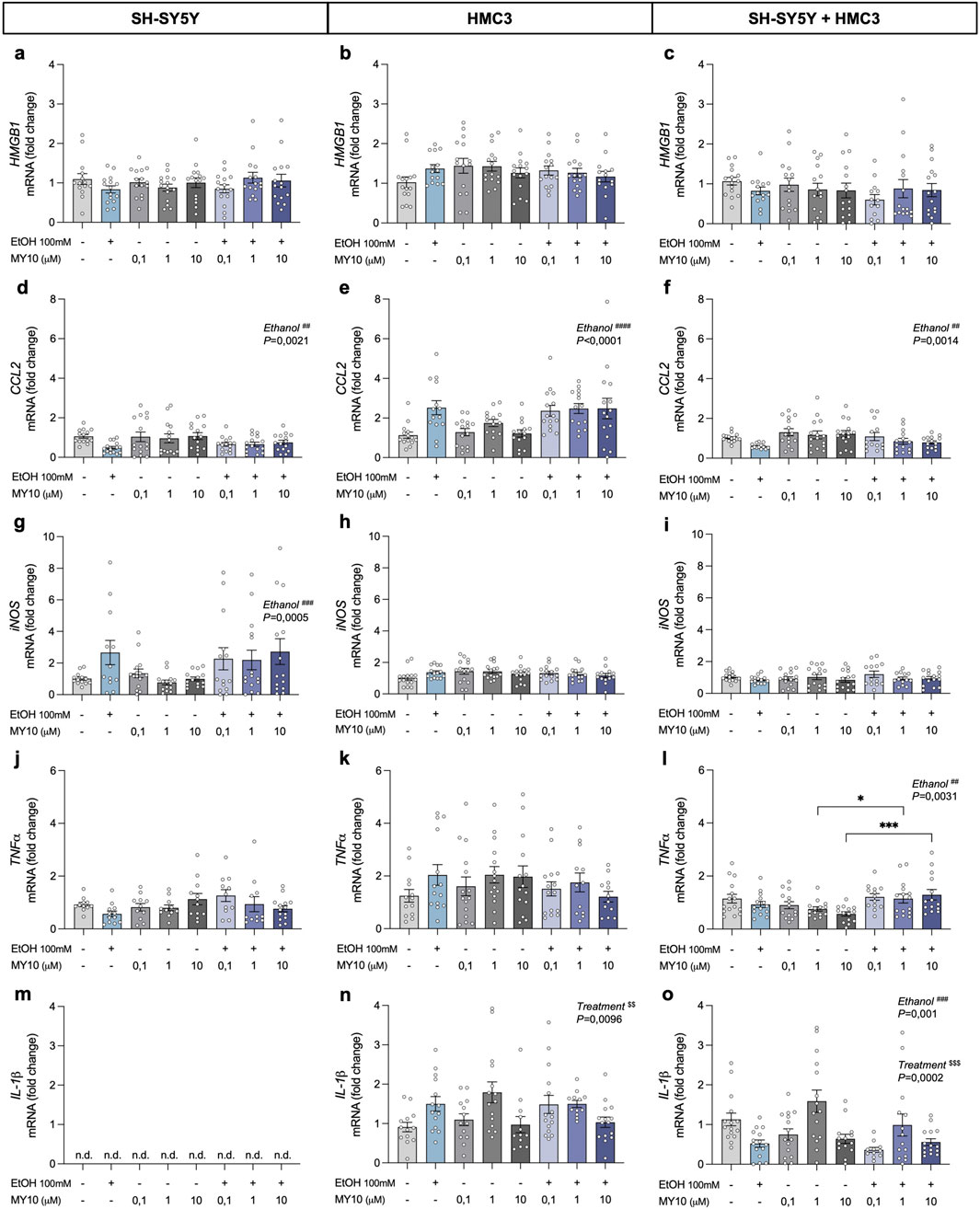
Figure 3. Effect of 24-h ethanol exposure (100 mM) and MY10 treatment on the expression of inflammatory genes in neurons, microglia and co-cultures. HMGB1 mRNA expression in SH-SY5Y cells (a), HMC3 cells (b), and co-culture (c). CCL2 mRNA expression in SH-SY5Y cells (d), HMC3 cells (e), and co-culture (f). iNOS mRNA expression in SH-SY5Y cells (g), HMC3 cells (h), and co-culture (i). TNFα mRNA expression in SH-SY5Y cells (j), HMC3 cells (k), and co-culture (l). IL-1β mRNA expression in SH-SY5Y cells (m), HMC3 cells (n), and co-culture (o). Data are presented as mean ± SEM. *P < 0.05, ***P < 0.001. ##P < 0.01, ###P ≤ 0.001, ####P < 0.0001 for a significant effect of EtOH exposure. $$P < 0.01, $$$P < 0.001 for significant effect of MY10 treatment. EtOH: ethanol.
Additionally, we did not observe significant effects of ethanol exposure or the treatment with MY10 on TNFα mRNA levels in monocultures (Figures 3j,k). However, in the co-culture system (Figure 3l), two-way ANOVA showed a significant effect of ethanol exposure on the mRNA expression levels of TNFα (F (1,105) = 9.192; P = 0.0031), together with a significant interaction between ethanol and MY10 treatment (F (3,105) = 3.902; P = 0.0109). Treatment with MY10 tended to reduce the mRNA expression levels of TNFα, which was not observed in the groups treated concomitantly with MY10 and ethanol (Figure 3l).
Importantly, IL-1β, which was only expressed in microglia cells, showed contrasting profiles (Figures 3m–o). IL-1β mRNA expression in HMC3 cells (Figure 3n) was significantly upregulated by treatment with MY10 (F (3,101) = 4.015; P = 0.0096) and the same tendency was observed after ethanol exposure. In contrast, the two-way ANOVA performed with the data from the co-cultures of neurons and microglia revealed significant effects of the treatment with MY10 (F (3,106) = 7.307; P = 0.0002) and of ethanol exposure (F (1,106) = 11.42; P = 0.001). In the co-cultures, ethanol exposure decreased IL-1β mRNA expression levels, which seemed to be only reverted by 1 μM MY10 (Figure 3o).
3.3 Effect of MY10 on toll-like receptors expression (TLRs) after ethanol exposure
To further explore the pathways involved in ethanol-induced neuroinflammation, we evaluated the transcriptional profiles of Toll-like receptors (TLRs) (Figure 4). The two-way ANOVA revealed a significant effect of ethanol exposure on HMC3 cells (F (1,103) = 10.8; P = 0.0014) and a significant interaction between ethanol and treatment with MY10 (F (3,103) = 2.705; P < 0.05) on TLR3 mRNA expression. Ethanol exposure upregulated TLR3 mRNA expression levels in microglial cells, an effect that was attenuated by treatment with MY10 (Figure 4a). In contrast, we did not find significant effects of ethanol exposure or treatment with MY10 in the expression of TLR3 in the co-culture of microglia and neurons (Figure 4b). Ethanol exposure increased TLR4 mRNA expression levels in HMC3 cells with a significant interaction between ethanol and treatment with MY10 (F (3,100) = 3.455; P = 0.0193) (Figure 4c). As in the case of TLR3, this effect was not observed in the co-cultures (Figure 4d). We did not find significant effects of ethanol exposure or treatment with MY10 in the expression of TLR7 in HMC3 cells (Figure 4e). In the co-cultures, the two-way ANOVA performed revealed a significant effect of the treatment with MY10 (F (3,109) = 3.757; P = 0.013), which caused a modest but consistent increase on TLR7 mRNA expression levels in all groups treated with MY10 independently of the concentration (Figure 4f).
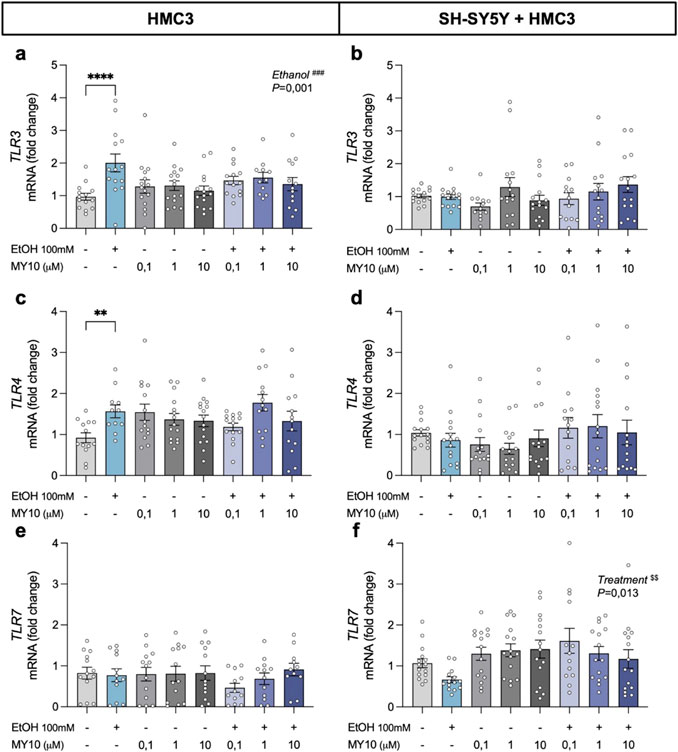
Figure 4. Effect of 24-h ethanol exposure (100 mM) and MY10 treatment on the expression of Toll-like receptor (TLRs) genes in microglia and co-cultures of neurons and microglia. TLR3 mRNA expression in HMC3 cells (a) and microglial-neuron co-culture (b). TLR4 mRNA expression in HMC3 cells (c) and microglial-neuron co-culture (d). TLR7 mRNA expression in HMC3 cells (e) and microglial-neuron co-culture (f). Data are presented as mean ± SEM. **P < 0.01, ****P < 0.0001. ###P ≤ 0.001 for a significant effect of EtOH exposure. $$P ≤ 0.01 for significant effect of MY10 treatment. EtOH: ethanol.
4 Discussion
In this study, we used a human co-culture model that combines HMC3 microglial cells with SH-SY5Y cells differentiated into neurons. This model was developed to create a physiologically relevant platform (Goshi et al., 2020) for studying ethanol-induced neuroinflammatory responses and the possible modulatory role of RPTPβ/ζ.
We observed that ethanol selectively modulates the expression of PTN and MK depending on the cellular context. Interestingly, PTN expression was upregulated in HMC3 monocultures following ethanol exposure, whereas MK was downregulated in co-cultures. These differences suggest that microglia respond to ethanol by increasing the expression of a potent neurotrophic factor, PTN, suggesting a stress-adaptive response in these critical immune cells. In contrast, the decreased expression of MK induced by ethanol exposure in co-cultures suggests a complex regulation of this neurotrophic factor expression involving neuronal-microglial interactions. These results are important since the cerebral expression of both PTN and MK is regulated by ethanol in both humans and animal models (Herradon et al., 2019). Our data indicate for the first time that ethanol exposure differentially regulated PTN and MK expression depending on the cell type and neuron-microglia communication.
In addition, we observed a robust microglial response induced by ethanol exposure, characterized by increased expression of proinflammatory genes such as CCL2 and TLRs, consistent with previous reports (Crews et al., 2011; Qin and Crews, 2012; Coleman et al., 2017; Walter and Crews, 2017; Lawrimore and Crews, 2017; Lawrimore et al., 2019). In contrast, neurons in monoculture exhibited a downregulation of CCL2 expression alongside a pronounced upregulation of iNOS, suggesting a limited but specific response to ethanol exposure. These neurons appear to engage selective stress-related signaling pathways in response to the neurotoxic stimulus (Lawrimore and Crews, 2017).
When both cell populations were integrated into a co-culture system, we observed a substantial attenuation of ethanol-induced inflammatory responses compared to microglia monocultures (Abellanas et al., 2019; Haenseler et al., 2017), reflected in the expression of key proinflammatory markers, such as CCL2, IL-1β, as well as TLR3 and TLR4. These findings suggest that neurons exert a dampening effect on microglial reactivity, potentially through direct cell-to-cell signaling. This interaction may have important implications for regulating neuroimmune responses and preserving neuronal viability (Alfonso-Loeches et al., 2010; Crews et al., 2017; Correa et al., 2013; Kraft and Harry, 2011). Our results align with previous co-culture studies that have demonstrated the cross-regulation of glial reactivity via neuronal signals (Crews et al., 2024; Zou and Crews, 2014; Lawrimore et al., 2019; Boyadjieva and Sarkar, 2010; Arzua et al., 2020).
The pharmacological inhibition of RPTPβ/ζ with MY10 revealed a selective and cell-dependent modulation of neuroinflammatory signaling. In both SH-SY5Y and HMC3 monocultures, MY10 treatment alone did not alter the expression of most proinflammatory genes, suggesting that basal activation of this pathway may be limited in cellular resting states. However, in cells exposed to ethanol, MY10 partially influenced the inflammatory responses by regulating the expression of specific genes, such as IL-1β and TNFα, in both microglial cells and co-cultures. Moreover, MY10 significantly altered the TLR7 expression in co-cultures, indicating that its effects are not limited to cytokine modulation but may extend to upstream components of innate immune signaling (Coleman and Crews, 2018). These results underscore the importance of studying pharmacological interventions in models that more accurately reflect in vivo cellular communication (Coleman et al., 2017; Coleman and Crews, 2018). Furthermore, these findings support previous studies using animal models regarding the neuroprotective and anti-inflammatory properties of MY10, validating the PTN/MK–RPTPβ/ζ signaling axis as a promising therapeutic target in ethanol-induced brain injury (Calleja-Conde et al., 2020; Fernández-Calle et al., 2018; Fernández-Calle et al., 2019; Fernández-Calle et al., 2020; Galán-Llario et al., 2023a; Galán-Llario et al., 2023b; Herradon et al., 2019).
A point to consider in the present work is the lack of protein-level validation of the transcriptional changes observed. Although gene expression analysis provides a valuable overview of regulatory trends, post-transcriptional mechanisms may alter final protein levels. Future studies quantifying key neuroimmune mediators, such as CX3CL1 (Fractalkine), should be useful to validate and complement the transcriptomic data and further elucidate the bidirectional signaling between neurons and microglia in response to alcohol and MY10. In addition, it is important to note that a limitation of this study is the inability to determine the specific cellular origin of gene expression changes in the co-culture system. However, as in many in vivo murine models that analyze entire brain regions without isolating specific cell types, this approach reflects the complexity of the brain environment and captures coordinated neuroimmune responses. It provides biologically significant, translationally relevant insights and facilitates the future application of cell-specific techniques such as immunocytochemistry, flow cytometry or single-cell RNA sequencing (scRNA-seq), which allow higher-resolution analysis of heterogeneous systems. Recent studies using scRNA-seq in alcohol dependence models (Salem et al., 2024; Warden et al., 2024) have demonstrated the utility of this approach in resolving cell-type and region-specific transcriptomic changes. In fact, it has allowed us to interpret more accurately the data collected here in human cells in the context of the relevant previous studies that defined the effects of MY10 and ethanol in different rodent models (Calleja-Conde et al., 2020; Fernández-Calle et al., 2018; Galán-Llario et al., 2023a; Galán-Llario et al., 2023b). In this sense, previous studies using primary rodent cultures of both cortical and hippocampal neurons and microglia have demonstrated strong inflammatory responses to ethanol and other neurotoxic agents (Crews et al., 2011; Coleman et al., 2017). However, species-specific differences in aspects such as alcohol metabolism, gene expression, and immune response limit the translational relevance of these models (Pascual et al., 2018; Walter and Crews, 2017; Seok et al., 2013). Additionally, reliance exclusively on murine models may fail to fully identify human-specific regulation of genes in response to alcohol, as revealed by comparative transcriptomic analyses of postmortem human and mouse brain tissue (Pascual et al., 2018; van der Worp et al., 2010). Our study suggests that human co-culture systems derived from immortalized cell lines offer a level of standardization that is difficult to achieve with primary cultures, often limited by heterogeneity and availability (Abud et al., 2017; Dello Russo et al., 2018; Pascual et al., 2018; Walter and Crews, 2017; Szabo and Gulya, 2013).
In conclusion, our study provides novel insights into the modulatory actions of microglial-neuronal interactions in ethanol-induced neuroimmune responses. These findings strongly support the utility of the human neuronal-microglial co-culture model as a physiologically relevant in vitro platform for studying ethanol-induced immune responses. Furthermore, the pharmacological assessment of the RPTPβ/ζ inhibitor MY10 in this context highlights the potential of this signaling pathway as an innovative therapeutic target in the deleterious effects of ethanol in the brain. While the model has inherent limitations due to the use of 2D systems and immortalized cell lines, it offers a valuable approach for advancing translational research on alcohol pathophysiology and the development of novel neuroprotective strategies.
Data availability statement
The datasets are available from the corresponding author upon request.
Author contributions
MP: Formal Analysis, Data curation, Writing – review and editing, Conceptualization, Writing – original draft, Methodology, Investigation. HC-R: Formal Analysis, Writing – original draft, Data curation, Writing – review and editing, Conceptualization. EG: Formal Analysis, Supervision, Data curation, Funding acquisition, Conceptualization, Writing – original draft, Writing – review and editing. GH: Data curation, Resources, Project administration, Writing – original draft, Conceptualization, Writing – review and editing, Formal Analysis, Funding acquisition, Supervision.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by National Plan on Drug abuse, Ministerio de Sanidad of Spain (grants 2023I018 to GH and 2024I044 to EG) and by ISCIII Redes de Investigación Cooperativa Orientadas a Resultados en Salud (RICORS), Red de Investigación en Atención Primaria de Adicciones (RIAPAd; grants RD21/0009/0013 and RD24/0003/0011 to GH). MAP was supported by Red de Investigación en Atención Primaria de Adicciones (RIAPAd). HC-R was supported by a fellowship from Fundación Universitaria San Pablo CEU-Santander.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The handling editor MSGG declared a past co-authorship with the author GH.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2025.1634202/full#supplementary-material
References
Abellanas, M. A., Zamarbide, M., Basurco, L., Luquin, E., Garcia-Granero, M., Clavero, P., et al. (2019). Midbrain microglia mediate a specific immunosuppressive response under inflammatory conditions. J. Neuroinflammation 16, 233. doi:10.1186/s12974-019-1628-8
Abud, E. M., Ramirez, R. N., Martinez, E. S., Healy, L. M., Nguyen, C. H. H., Newman, S. A., et al. (2017). iPSC-Derived human microglia-like cells to study neurological diseases. Neuron 94 (2), 278–293. doi:10.1016/j.neuron.2017.03.042
Alfonso-Loeches, S., Pascual-Lucas, M., Blanco, A. M., Sanchez-Vera, I., and Guerri, C. (2010). Pivotal role of TLR4 receptors in alcohol-induced neuroinflammation and brain damage. J. Neurosci. official J. Soc. Neurosci. 30 (24), 8285–8295. doi:10.1523/JNEUROSCI.0976-10.2010
Arzua, T., Yan, Y., Jiang, C., Logan, S., Allison, R. L., Wells, C., et al. (2020). Modeling alcohol-induced neurotoxicity using human induced pluripotent stem cell-derived three-dimensional cerebral organoids. Transl. psychiatry 10 (1), 347. doi:10.1038/s41398-020-01029-4
Boyadjieva, N. I., and Sarkar, D. K. (2010). Role of microglia in ethanol's apoptotic action on hypothalamic neuronal cells in primary cultures. Alcohol. Clin. Exp. Res. 34 (11), 1835–1842. doi:10.1111/j.1530-0277.2010.01271.x
Calleja-Conde, J., Fernández-Calle, R., Zapico, J. M., Ramos, A., de Pascual-Teresa, B., Bühler, K. M., et al. (2020). Inhibition of receptor protein tyrosine phosphatase β/ζ reduces alcohol intake in rats. Alcohol. Clin. Exp. Res. 44 (5), 1037–1045. doi:10.1111/acer.14321
Coleman, L. G., and Crews, F. T. (2018). Innate immune signaling and alcohol use disorders. Handb. Exp. Pharmacol. 248, 369–396. doi:10.1007/164_2018_92
Coleman, L. G., Zou, J., and Crews, F. T. (2017). Microglial-derived miRNA let-7 and HMGB1 contribute to ethanol-induced neurotoxicity via TLR7. J. neuroinflammation 14 (1), 22. doi:10.1186/s12974-017-0799-4
Coleman, L. G., Zou, J., Qin, L., and Crews, F. T. (2018). HMGB1/IL-1β complexes regulate neuroimmune responses in alcoholism. Brain, Behav. Immun. 72, 61–77. doi:10.1016/j.bbi.2017.10.027
Correa, F. G., Hernangómez, M., and Guaza, C. (2013). Understanding microglia-neuron cross talk: relevance of the microglia-neuron cocultures. Methods Mol. Biol. Clift. N.J. 1041, 215–229. doi:10.1007/978-1-62703-520-0_20
Crews, F. T. (2012). Immune function genes, genetics, and the neurobiology of addiction. Alcohol Res. Curr. Rev. 34 (3), 355–361. doi:10.17615/3882-nr29
Crews, F. T., Coleman, L. G., Macht, V. A., and Vetreno, R. P. (2024). Alcohol, HMGB1, and innate immune signaling in the brain. Alcohol Res. Curr. Rev. 44 (1), 04. doi:10.35946/arcr.v44.1.04
Crews, F. T., Walter, T. J., Coleman, L. G., and Vetreno, R. P. (2017). Toll-like receptor signaling and stages of addiction. Psychopharmacology 234 (9-10), 1483–1498. doi:10.1007/s00213-017-4560-6
Crews, F. T., Zou, J., and Qin, L. (2011). Induction of innate immune genes in brain create the neurobiology of addiction. Brain, Behav. Immun. 25 (Suppl. 1), S4-S12–S12. doi:10.1016/j.bbi.2011.03.003
Del Campo, M., Fernández-Calle, R., Vicente-Rodríguez, M., Martín Martínez, S., Gramage, E., Zapico, J. M., et al. (2021). Role of receptor protein tyrosine phosphatase β/ζ in neuron-microglia communication in a cellular model of parkinson's disease. Int. J. Mol. Sci. 22 (13), 6646. doi:10.3390/ijms22136646
Dello Russo, C., Cappoli, N., Coletta, I., Mezzogori, D., Paciello, F., Pozzoli, G., et al. (2018). The human microglial HMC3 cell line: where do we stand? A systematic literature review. J. neuroinflammation 15 (1), 259. doi:10.1186/s12974-018-1288-0
Fernández-Calle, R., Galán-Llario, M., Gramage, E., Zapatería, B., Vicente-Rodríguez, M., Zapico, J. M., et al. (2020). Role of RPTPβ/ζ in neuroinflammation and microglia-neuron communication. Sci. Rep. 10 (1), 20259. doi:10.1038/s41598-020-76415-5
Fernández-Calle, R., Gramage, E., Zapico, J. M., de Pascual-Teresa, B., Ramos, A., and Herradón, G. (2019). Inhibition of RPTPβ/ζ blocks ethanol-induced conditioned place preference in pleiotrophin knockout mice. Behav. brain Res. 369, 111933. doi:10.1016/j.bbr.2019.111933
Fernández-Calle, R., Vicente-Rodríguez, M., Gramage, E., Pita, J., Pérez-García, C., Ferrer-Alcón, M., et al. (2017). Pleiotrophin regulates microglia-mediated neuroinflammation. J. neuroinflammation 14 (1), 46. doi:10.1186/s12974-017-0823-8
Fernández-Calle, R., Vicente-Rodríguez, M., Pastor, M., Gramage, E., Di Geronimo, B., Zapico, J. M., et al. (2018). Pharmacological inhibition of receptor protein tyrosine phosphatase β/ζ (PTPRZ1) modulates behavioral responses to ethanol. Neuropharmacology 137, 86–95. doi:10.1016/j.neuropharm.2018.04.027
Galán-Llario, M., Gramage, E., García-Guerra, A., Torregrosa, A. B., Gasparyan, A., Navarro, D., et al. (2024). Adolescent intermittent ethanol exposure decreases perineuronal nets in the hippocampus in a sex dependent manner: modulation through pharmacological inhibition of RPTPβ/ζ. Neuropharmacology 247, 109850. doi:10.1016/j.neuropharm.2024.109850
Galán-Llario, M., Rodríguez-Zapata, M., Fontán-Baselga, T., Gramage, E., Vicente-Rodríguez, M., Zapico, J. M., et al. (2023b). Inhibition of RPTPβ/ζ reduces chronic ethanol intake in adolescent mice and modulates ethanol effects on hippocampal neurogenesis and glial responses in a sex-dependent manner. Neuropharmacology 227, 109438. doi:10.1016/j.neuropharm.2023.109438
Galán-Llario, M., Rodríguez-Zapata, M., Gramage, E., Vicente-Rodríguez, M., Fontán-Baselga, T., Ovejero-Benito, M. C., et al. (2023a). Receptor protein tyrosine phosphatase β/ζ regulates loss of neurogenesis in the mouse hippocampus following adolescent acute ethanol exposure. Neurotoxicology 94, 98–107. doi:10.1016/j.neuro.2022.11.008
Goshi, N., Morgan, R. K., Lein, P. J., and Seker, E. (2020). A primary neural cell culture model to study neuron, astrocyte, and microglia interactions in neuroinflammation. J. neuroinflammation 17 (1), 155. doi:10.1186/s12974-020-01819-z
Gresa-Arribas, N., Viéitez, C., Dentesano, G., Serratosa, J., Saura, J., and Solà, C. (2012). Modelling neuroinflammation in vitro: a tool to test the potential neuroprotective effect of anti-inflammatory agents. PloS one 7 (9), e45227. doi:10.1371/journal.pone.0045227
Haenseler, W., Sansom, S. N., Buchrieser, J., Newey, S. E., Moore, C. S., Nicholls, F. J., et al. (2017). A highly efficient human pluripotent stem cell microglia model displays a neuronal-co-culture-specific expression profile and inflammatory response. Stem cell Rep. 8 (6), 1727–1742. doi:10.1016/j.stemcr.2017.05.017
Hartung, T. (2009). Toxicology for the twenty-first century. Nature 460 (7252), 208–212. doi:10.1038/460208a
Herradón, G., and Pérez-García, C. (2014). Targeting midkine and pleiotrophin signalling pathways in addiction and neurodegenerative disorders: recent progress and perspectives. Br. J. Pharmacol. 171 (4), 837–848. doi:10.1111/bph.12312
Herradon, G., Ramos-Alvarez, M. P., and Gramage, E. (2019). Connecting metainflammation and neuroinflammation through the PTN-MK-RPTPβ/ζ axis: relevance in therapeutic development. Front. Pharmacol. 10, 377. doi:10.3389/fphar.2019.00377
Kraft, A. D., and Harry, G. J. (2011). Features of microglia and neuroinflammation relevant to environmental exposure and neurotoxicity. Int. J. Environ. Res. public health 8 (7), 2980–3018. doi:10.3390/ijerph8072980
Lawrimore, C. J., Coleman, L. G., Zou, J., and Crews, F. T. (2019). Ethanol induction of innate immune signals across BV2 microglia and SH-SY5Y neuroblastoma involves induction of IL-4 and IL-13. Brain Sci. 9 (9), 228. doi:10.3390/brainsci9090228
Lawrimore, C. J., and Crews, F. T. (2017). Ethanol, TLR3, and TLR4 agonists have unique innate immune responses in neuron-like SH-SY5Y and microglia-like BV2. Alcohol. Clin. Exp. Res. 41 (5), 939–954. doi:10.1111/acer.13368
Pascual, M., López-Hidalgo, R., Montagud-Romero, S., Ureña-Peralta, J. R., Rodríguez-Arias, M., and Guerri, C. (2021). Role of mTOR-regulated autophagy in spine pruning defects and memory impairments induced by binge-like ethanol treatment in adolescent mice. Brain pathol. 31 (1), 174–188. doi:10.1111/bpa.12896
Pascual, M., Montesinos, J., and Guerri, C. (2018). Role of the innate immune system in the neuropathological consequences induced by adolescent binge drinking. J. Neurosci. Res. 96 (5), 765–780. doi:10.1002/jnr.24203
Pastor, M., Fernández-Calle, R., Di Geronimo, B., Vicente-Rodríguez, M., Zapico, J. M., Gramage, E., et al. (2018). Development of inhibitors of receptor protein tyrosine phosphatase β/ζ (PTPRZ1) as candidates for CNS disorders. Eur. J. Med. Chem. 144, 318–329. doi:10.1016/j.ejmech.2017.11.080
Qin, L., and Crews, F. T. (2012). NADPH oxidase and reactive oxygen species contribute to alcohol-induced microglial activation and neurodegeneration. J. neuroinflammation 9, 5. doi:10.1186/1742-2094-9-5
Rodríguez-Zapata, M., Galán-Llario, M., Cañeque-Rufo, H., Sevillano, J., Sánchez-Alonso, M. G., Zapico, J. M., et al. (2023). Implication of the PTN/RPTPβ/ζ signaling pathway in acute ethanol neuroinflammation in both sexes: a comparative study with LPS. Biomedicines 11 (5), 1318. doi:10.3390/biomedicines11051318
Roqué, P. J., and Costa, L. G. (2017). Co-Culture of neurons and microglia. Curr. Protoc. Toxicol. 74, 11.24.1–11.24.17. doi:10.1002/cptx.32
Salem, N. A., Manzano, L., Keist, M. W., Ponomareva, O., Roberts, A. J., Roberto, M., et al. (2024). Cell-type brain-region specific changes in prefrontal cortex of a mouse model of alcohol dependence. Neurobiol. Dis. 190, 106361. doi:10.1016/j.nbd.2023.106361
Seok, J., Warren, H. S., Cuenca, A. G., Mindrinos, M. N., Baker, H. V., Xu, W., et al. (2013). Genomic responses in mouse models poorly mimic human inflammatory diseases. Proc. Natl. Acad. Sci. U. S. A. 110 (9), 3507–3512. doi:10.1073/pnas.1222878110
Szabo, M., and Gulya, K. (2013). Development of the microglial phenotype in culture. Neuroscience 241, 280–295. doi:10.1016/j.neuroscience.2013.03.033
van der Worp, H. B., Howells, D. W., Sena, E. S., Porritt, M. J., Rewell, S., O'Collins, V., et al. (2010). Can animal models of disease reliably inform human studies? PLoS Med. 7 (3), e1000245. doi:10.1371/journal.pmed.1000245
Walter, T. J., and Crews, F. T. (2017). Microglial depletion alters the brain neuroimmune response to acute binge ethanol withdrawal. J. Neuroinflammation 14, 86. doi:10.1186/s12974-017-0856-z
Warden, A. S., Salem, N. A., Brenner, E., Sutherland, G. T., Stevens, J., Kapoor, M., et al. (2024). Integrative genomics approach identifies glial transcriptomic dysregulation and risk in the cortex of individuals with alcohol use disorder. bioRxiv Prepr. Serv. Biol. 2024, 2024.08.16.607185. doi:10.1101/2024.08.16.607185
Keywords: alcohol, RPTPβ/ζ, co-culture, neuroinflammation, pleiotrophin, midkine
Citation: Penedo MA, Cañeque-Rufo H, Gramage E and Herradón G (2025) Ethanol-induced changes in neurotrophic and immune genes are regulated by receptor-type protein tyrosine phosphatase β/ζ (RPTPβ/ζ) and microglial-neuronal interactions. Front. Genet. 16:1634202. doi: 10.3389/fgene.2025.1634202
Received: 23 May 2025; Accepted: 18 July 2025;
Published: 22 August 2025.
Edited by:
Maria S. Garcia-Gutierrez, Miguel Hernández University of Elche, SpainReviewed by:
Fernando Rodriguez De Fonseca, University of Malaga, SpainEsther Caparrós, Miguel Hernández University of Elche, Spain
Copyright © 2025 Penedo, Cañeque-Rufo, Gramage and Herradón. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gonzalo Herradón, aGVycmFkb25AY2V1LmVz