 Andrés Felipe Leal
Andrés Felipe Leal Harry Pachajoa
Harry Pachajoa Shunji Tomatsu
Shunji Tomatsu- 1Centro de Investigaciones en Anomalías Congénitas y Enfermedades Raras, Universidad Icesi, Cali, Colombia
- 2Centro de Investigaciones Clínicas, Fundación Valle de Lili, Cali, Colombia
- 3Nemours Children’s Health, Wilmington, DE, United States
- 4Institute for the Study of Inborn Errors of Metabolism, Faculty of Science, Pontificia Universidad Javeriana, Bogotá, Colombia
- 5Departamento de Genética Clínica, Fundación Valle de Lili, Cali, Colombia
- 6Faculty of Arts and Sciences, University of Delaware, Newark, DE, United States
- 7Department of Pediatrics, Thomas Jefferson University, Philadelphia, PA, United States
Lysosomal storage disorders (LSDs) are rare substrate-accumulating diseases primarily characterized by mutations in genes encoding proteins involved in lysosomal function, most of which have enzymatic activity. Resulting lysosomal dysfunction leads to the overaccumulation of non- or partially degraded substrates. While it is true that enzyme deficiency is the primary cause of LSDs, the epigenetic alterations in DNA methylation, miRNA expression, and histone modifications appear to be critical mechanisms involved in the pathogenesis of LSDs. As epigenetic marks are, in most cases, reversible, their study becomes vital to developing strategies aimed at reversing epigenome alterations. Although classical therapeutic alternatives aim to recover the lysosomal function by restoring the protein expression lost, the use of modifiers able to repair the epigenetic modifications in LSDs may become a promising strategy. This manuscript explores the most recent evidence on the epigenetic alterations in LSDs. It also discusses their modulation through epigenetic modulators, a novel and intriguing approach to treat LSDs, as well as the potential of the CRISPR/Cas9 system.
1 Introduction
Belonging to the inborn errors of metabolism (IEM), the lysosomal storage disorders (LSDs) encompass over 70 genetically inherited metabolic diseases characterized by the accumulation of undegraded substrates due to lysosomal dysfunction (Leal et al., 2020a; Ballabio, 2016; Parenti et al., 2021). LSDs are multisystemic pathologies, often presenting with neurological decline, skeletal abnormalities, hepatosplenomegaly, along with pulmonary and cardiac involvement (Parenti et al., 2021).
Resulting substrate accumulation leads to disturbances in cell homeostasis, which ultimately promote pro-oxidant, pro-inflammatory, and pro-apoptotic profiles (Leal et al., 2022). Beyond lysosomal substrate accumulation, as the primary cause of the LSDs, some studies suggest that epigenetic dysregulation may contribute to LSD pathogenesis (Hassan et al., 2017). For instance, aberrant DNA methylation, histone modification patterns, and disrupted non-coding RNA expression have been documented in LSDs (Fu et al., 2022; Shen et al., 2022; Morena et al., 2019; Vargas-López and Alméciga-Díaz, 2022; Xiao et al., 2019; Kunkel et al., 2023; Üzen et al., 2025; Tarallo et al., 2019; Tomatsu et al., 2005; Dasgu et al., 2015), further supporting their role in the pathomechanisms of these disorders.
Given that epigenetic modifications can be reversible, they emerge as attractive targets for therapeutic modulation (Lossi et al., 2024). Some molecules, such as GSK-J4, a histone demethylase inhibitor (KDMi) (Kunkel et al., 2023), RVX-208, which targets epigenetic readers (Fu et al., 2022), and miRNA overexpression (Inamura et al., 2021), have been evaluated in LSD models with encouraging results. Similarly, the use of histone deacetylase (HDAC) and DNA methyltransferase (DNMT) inhibitors already approved for use in oncology and neurodegeneration may also open a new avenue for rapidly moving into their repurposing for LSD treatment. The potential of these molecules is exciting, but equally thrilling is the use of advanced CRISPR/Cas9-based tools. This innovative strategy may open a promising opportunity for modifying epigenetic marks in LSDs.
In this manuscript, we review recent evidence on the role of epigenetic dysregulation in LSDs and the potential of treatments emerging from classical- and CRISPR/Cas9-based epigenetic mark-modifier approaches. The paper search was conducted using PubMed, Web of Science, and Google Scholar databases, and the following Boolean terms were applied: “lysosomal storage disorder” OR “LSD” AND “epigenetics” OR “dCas9” OR “epigenome” OR “DNA methylation” OR “histones” OR “chromatin”. Peer-reviewed papers from 2015 to 2025 were included, with a focus on those published between 2018 and 2025. Studies up to 2015 were systematically reviewed by Hassan et al. (2017), and we encourage interested readers to consult their work.
2 Molecular mechanisms of epigenetic signatures
2.1 DNA methylation
DNA methylation is mediated by the covalent addition of a methyl group to the 5′position of cytosine residues, typically within CpG-rich regions (Figure 1) (Mattei et al., 2022; Li and Tollefsbol, 2021). DNA methylation is regulated by cellular metabolism since it requires the methionine cycle to maintain the levels of S-adenosylmethionine (SAM), the universal source of the methyl group for DNA and histone methyltransferases (HMTs), and to control S-adenosylhomocysteine (SAH), a competitive inhibitor of HMTs (Bernasocchi and Mostoslavsky, 2024). Hypermethylation silences the transcription of genes by blocking the transcriptional machinery from binding to the genome; conversely, hypomethylation may result in the overexpression of genes (Mattei et al., 2022; Angeloni and Bogdanovic, 2019). DNA methylation is an essential physiological mechanism and a key epigenetic modification that underpins X-chromosome inactivation, genomic imprinting, and tissue-specific gene expression (Li and Tollefsbol, 2021). Aberrant DNA methylation is observed in pathological scenarios such as cancer (Li and Tollefsbol, 2021; Nishiyama and Nakanishi, 2021), autoimmune disorders (Ali et al., 2022; Danieli et al., 2024), neurodegenerative conditions (Ghosh and Saadat, 2023; De Plano et al., 2024), as well as some LSDs (Shen et al., 2022; Vargas-López and Alméciga-Díaz, 2022; Tomatsu et al., 2005). Importantly, oxidative stress has been documented in multiple LSDs (Ago et al., 2024; Lee and Hong, 2023; Leal and Alméciga-Díaz, 2022; Simoncini et al., 2020; Stepien et al., 2020) and may influence the activity of methyltransferases, thereby disrupting the SAM/SAH balance. Indeed, an increased accumulation of SAH can reduce the adequate availability of methyl donors, ultimately contributing to both global and gene-specific DNA hypomethylation. These findings support the idea that metabolic stress and impaired lysosomal function converge on the epigenome, linking classical metabolic dysfunction with transcriptional dysregulation in LSDs.
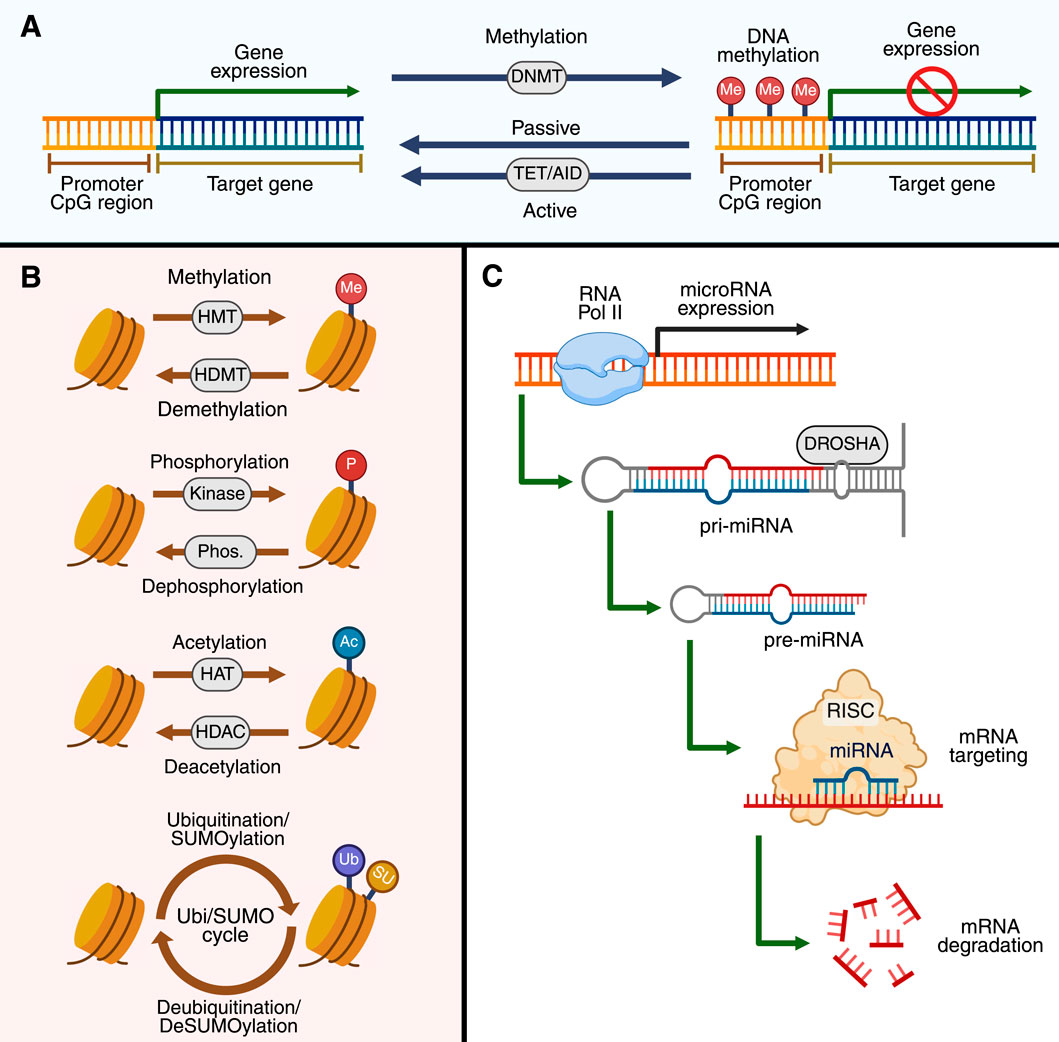
Figure 1. Molecular mechanism in epigenetics. In (A), the processes of DNA methylation and demethylation control gene expression at CpG-rich promoter regions are shown. DNA methyltransferases (DNMTs) mediate the addition of methyl groups (Me) to cytosines, leading to gene silencing. Passive demethylation occurs during DNA replication, whereas active demethylation is catalyzed by the ten-eleven translocation (TET) enzymes and activation-induced cytidine deaminase (AID). In (B), the post-translational modifications of histones are shown. Histone methyltransferases (HMTs) and histone demethylases (HDMTs) add and remove methyl groups (Me), respectively. Likewise, histones can also be phosphorylated (P) by kinases, while phosphorylation is removed by phosphatases (Phos.). Similarly, acetylation (Ac) of histones occurs through histone acetyltransferases (HATs) and is deacetylated by histone deacetylases (HDACs). Finally, histones can also undergo ubiquitination and SUMOylation (Ub/SUMO). In (C), the mechanism of non-coding RNA expression is illustrated. DROSHA first processes primary microRNAs (pri-miRNAs) into precursor microRNAs (pre-miRNAs), which are then loaded onto RISC. In the RISC, mature miRNAs direct the complex to specific mRNAs, resulting in the degradation or translational inhibition of these mRNAs. The image was created by BioRender.com.
2.2 Histone modifications
Histones are basic proteins that package DNA, forming a structure known as nucleosomes (Knapp et al., 2023; Zhang et al., 2021). Their significance lies in their ability to undergo a variety of post-translational modifications (PTMs) at their N-terminal tails, such as acetylation, methylation, phosphorylation, ubiquitination, and sumoylation. These modifications play a pivotal role in shaping the chromatin structure and influencing gene expression (Figure 1) (Zhang et al., 2021; Millán-Zambrano et al., 2022). For instance, histone acetylation of lysine residues (e.g., H3K9ac) typically induces transcription by opening the chromatin. Conversely, certain methylation marks (H3K27me3, H3K9me3) are associated with transcriptional repression (Zhang et al., 2021; Jain and Epstein, 2024). The activity of histone acetyltransferases (HATs), histone deacetylases (HDACs), HMTs, and histone demethylases (KDMs), which contribute to chromatin dynamics, is tightly regulated at the transcriptional, post-translational, and cellular levels (Jain and Epstein, 2024; Ramaiah et al., 2021). The urgency of understanding histone modifications is further highlighted by their role in disease, with dysregulation of these processes being linked to several human diseases.
2.3 Non-coding RNAs
Non-coding RNAs (ncRNAs) form a vast and functionally diverse group of RNAs that are not translated into proteins but rather are key regulators of gene expression (Figure 1) (Nemeth et al., 2024; Chen and Kim, 2024). This diversity is evident in the fact that microRNAs (miRNAs), approximately 22 nucleotides in length, typically bind to complementary sequences in the 3′untranslated regions (3′UTRs) of target mRNAs, promoting mRNA degradation or translation repression. Similarly, long non-coding RNAs (lncRNAs), which are greater than 200 nt in length, are frequently implicated in chromatin modification, transcriptional interference, and post-transcriptional control via RNA stabilization or degradation (Chen and Kim, 2024; Virciglio et al., 2021). The complexity and diversity of ncRNA underscore their importance in molecular biology, and dysregulation of these molecules has been associated with several human diseases, including LSDs (Morena et al., 2019; Xiao et al., 2019; Üzen et al., 2025; Tarallo et al., 2019; Dasgu et al., 2015; Virciglio et al., 2021).
3 Epigenetics and LSDs
Recent evidence strongly supports that epigenetic dysregulation in LSDs may offer additional pathogenic mechanisms beyond the lysosomal accumulation. This underscores the promising potential of certain epigenetic signatures in the treatment, diagnosis, and prognosis of LSD patients. In this section, we delve into the latest developments in the field of epigenome alterations in LSDs, which have been succinctly summarized in Table 1.
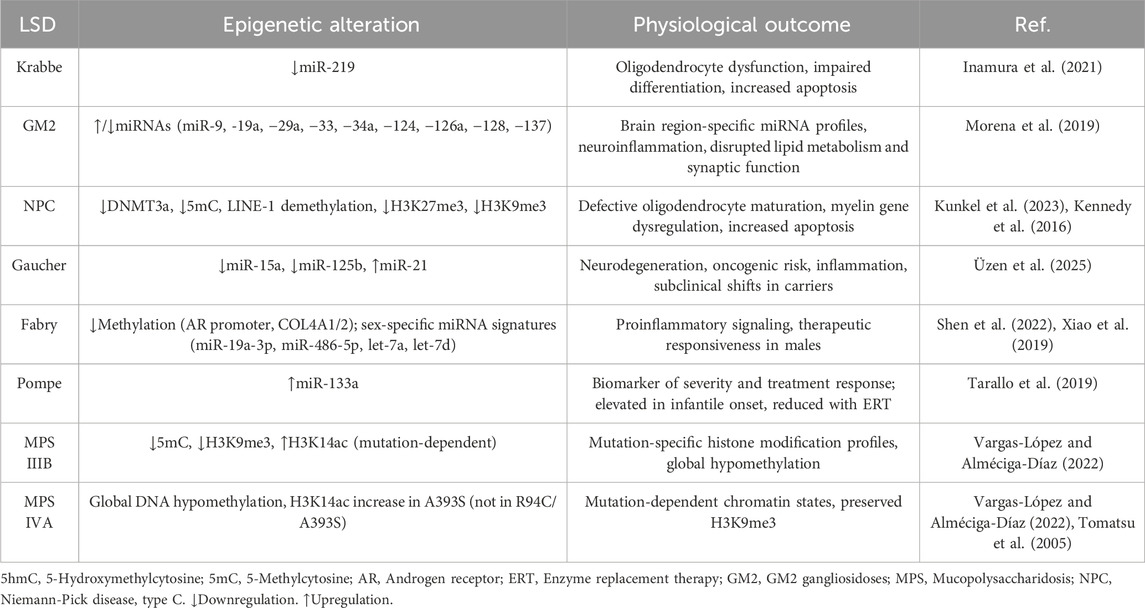
Table 1. Main epigenetic findings in LSDs.
3.1 Krabbe
Krabbe disease (KD, OMIM # 245200) is an LSD caused by mutations in the GALC gene, leading to a deficiency of galactocerebrosidase (GALC) and accumulation of the glycolipids galactosylceramide (GalCer) and sulfatide (Maghazachi, 2023). GALC deficiency results in severe demyelination in the central nervous system (CNS), along with early-onset forms causing rapid neurodegeneration and early death (Maghazachi, 2023). Regarding epigenetics, Inamura et al. (2021) identified an epigenetic mechanism involving miR-219 in the oligodendrocyte pathology of KD using the twitcher mouse model (Inamura et al., 2021). The authors observed a significant decrease in the miR-219 expression in developing oligodendrocytes in vivo and in vitro. Interestingly, oligodendrocyte precursor cells (OPCs) isolated from twitcher mice failed to differentiate and exhibited increased caspase-3-mediated apoptotic cell death when cultured (Inamura et al., 2021). Likewise, functional assays using luciferase reporters confirmed that miR-219-mediated mRNA repression was diminished in KD oligodendrocytes (Inamura et al., 2021), suggesting that miR-219 downregulation contributes to oligodendrocyte dysfunction and demyelination in KD.
3.2 GM2 gangliosidoses
GM2 gangliosidoses are a group of three related LSDs, as follows: Tay-Sach (OMIM # 272800), Sandhoff (OMIM # 268800), and AB variant (OMIM # 272750), characterized by impaired degradation of the GM2 ganglioside (Leal et al., 2020b). The lysosomal accumulation of GM2 in neurons leads to progressive neurodegeneration, with early-onset forms typically causing motor decline, seizures, and early death in infancy or childhood (Leal et al., 2020b; Toro et al., 2021). A study reported by Morena et al. (2019) provided, for the first time, an in-depth analysis of miRNA dysregulation in mouse models of Tay-Sachs and Sandhoff diseases (Morena et al., 2019). This study conducted miRNA analysis of 12 candidates involved in lipid metabolism, neural development, and neuroinflammatory processes, specifically in the subventricular zone (SVZ) and the striatum region (STR) of the brain (Morena et al., 2019). Interestingly, the comparative analysis revealed distinct brain region- and disease-specific patterns of miRNA expression. Nine miRNAs (including miR-9, −19a, −29a, −33, −34a, −124, −126a, −128, −137) showed altered levels in SVZ and STR across models, with some miRNAs uniquely dysregulated in SVZ or STR (Morena et al., 2019). The computational miRNA-mRNA network analysis further predicted downstream effects on pathways central to lysosomal dysfunction, lipid trafficking, axon guidance, synaptic signaling, inflammation, and cell survival (Morena et al., 2019), suggesting that miRNA-mediated post-transcriptional regulation may play a critical, region-specific role in the epigenetic mechanisms of GM2 gangliosidosis.
3.3 Niemann-pick type C
Niemann–Pick type C (NPC) is an LSD caused by mutations in the NPC1 (NPC type C1, OMIM # 257220) or NPC2 (NPC type C2, OMIM # 607625) genes, leading to defective cholesterol and lipid transport out of lysosomes (Lee and Hong, 2023). Early studies conducted by Kennedy et al. (2016) in a murine model of NPC revealed DNA methylation disruption, accompanied by a significant decrease in DNMT3a and methyl-CpG binding protein expression, decreased 5-methylcytosine staining, global LINE-1 demethylation, and promoter-specific hypermethylation of single-copy genes, supporting the alteration of epigenetic markers (Kennedy et al., 2016). Most recently, Kunkel et al. (2023) have brought to light that NPC1 is essential for maintaining repressive histone methylation during oligodendrocyte maturation (Kunkel et al., 2023). In Npc1−/− mice, RNA sequencing revealed disrupted myelin gene expression and aberrant activation of neuronal programs in oligodendrocyte lineage cells, along with reduced numbers of OLIG2+ and MBP+ cells, increased apoptosis, and a loss of H3K27me3 and H3K9me3 marks (Kunkel et al., 2023), further uncovering pathogenic mechanisms in NPC.
3.4 Gaucher
Gaucher disease (GD) is an LSD characterized by a deficiency of the glucocerebrosidase (GCase), leading to the lysosomal accumulation of glucocerebroside. Clinically, three types of GD are distinguished as follows: type 1 (non-neuronopathic, OMIM # 230800), type 2 (acute neuronopathic, OMIM # 230900), and type 3 (chronic neuronopathic, OMIM # 231000) (Minervini et al., 2023). Epigenetic alterations, such as the dysregulation of miR-181c-5p, miR-34b-5p, and miR-10a-5p, have been identified in early studies using the neuronopathic GD mouse model and are attributed to the pathogenesis of GD by impairing axonal guidance, synaptic plasticity, and mitochondrial function (Dasgu et al., 2015). Altered miRNA profiles have been widely identified in cancer (Wilson and Scaffidi, 2025). Interestingly, a study conducted by Üzen et al. (2025) assessed the oncogenic miRNA expression profile in type I GD patients (Üzen et al., 2025). The blood-based profiling revealed significant downregulation of miR-15a and miR-125b, both known tumor suppressors (Üzen et al., 2025), and upregulation of the oncogenic miR-21 in patients undergoing enzyme replacement therapy (ERT) (Üzen et al., 2025), suggesting a sustained pro-oncogenic molecular state that could predispose GD patients to increased risks for multiple myeloma, hepatocellular carcinoma, and hematologic malignancies. Importantly, heterozygous carriers also exhibited reduced levels of miR-15a, miR-150, and miR-181b, indicating subclinical epigenetic shifts (Üzen et al., 2025). These findings significantly contribute to our understanding of GD, suggesting that epigenetic dysregulation may play a crucial role in neurodegeneration in GD and an increased oncogenic risk.
3.5 Fabry
Fabry disease (FD, OMIM # 301500) is an X-linked LSD caused by mutations in the GLA gene, leading to a deficiency of the α-galactosidase A (α-Gal A), resulting in pathological accumulation of globotriaosylceramide (Gb3) (Lenders and Brand, 2021). Lysosomal accumulation of Gb3 in FD patients particularly affects endothelial cells, the kidneys, the heart, and the CNS (Lenders and Brand, 2021). Epigenetic alterations in FD have been described in several studies. For instance, Shen et al. (2022) demonstrated that the α-Gal A defect leads to DNA hypomethylation, thereby modifying the methylation pattern of genes, including the androgen receptor promoter, and hypomethylation of the COL4A1 and COL4A2 genes, resulting in their overexpression (Shen et al., 2022). These findings were accompanied by elevated methionine levels in both patient-derived endothelial cells and FD mouse models (Shen et al., 2022), linking lysosomal glycosphingolipid accumulation to epigenetically driven transcriptional remodeling.
At the post-transcriptional level, several studies have revealed miRNA-based dysregulation in FD. For instance, Xiao et al. (2019) identified distinct serum miRNA signatures that correlate with treatment response (Xiao et al., 2019). Most importantly, as FD primarily affects males, the authors also included analysis based on sex differences. In this context, the levels of miR-19a-3p and miR-486-5p expression were significantly reduced in ERT-treated males compared to untreated males (Xiao et al., 2019), suggesting that an epigenetic response to therapy is quantifiable and measurable. Conversely, female patients showed greater diversity in the expression of the miRNAs (Xiao et al., 2019), possibly due to the larger clinical and molecular heterogeneity associated with X-linked transmission. These results suggest that individual miRNAs may serve as biomarkers to monitor treatment responsiveness in male Fabry patients, highlighting the importance of considering gender in epigenetic marker investigations. Other miRNAs predicting the occurrence of a proinflammatory process, such as let-7a and let-7d, were identified in FD (Maier et al., 2021), which may represent potential biomarkers. This discovery offers optimism for the future of disease monitoring in FD.
3.6 Pompe
Pompe disease (PD, OMIM # 232300) is an LSD caused by mutations in the GAA gene. GAA encodes acid α-glucosidase (GAA). GAA deficiency results in the progressive accumulation of glycogen (George et al., 2024). In PD, an analysis of miRNA conducted by Tarallo et al. (2019) in mouse models (at 3 and 9 months) and plasma from Pompe patients identified a specific miRNA profile associated with disease severity and treatment response (Tarallo et al., 2019). In mice, 211 miRNAs were dysregulated in skeletal muscle and 66 in the heart, with distinct patterns at different ages. Relevantly, in plasma from six patients, 55 miRNAs were differentially expressed, with 16 overlapping those altered in mouse tissues (Tarallo et al., 2019). The miR-133a was proposed as a key biomarker, with levels significantly elevated in Pompe patients, especially in infantile-onset cases, compared to late-onset cases, and correlating with clinical severity (Tarallo et al., 2019). Interestingly, in three infantile patients, miR-133a levels decreased upon ERT, coinciding with clinical improvement (Tarallo et al., 2019). These findings suggest that circulating miRNAs, particularly miR-133a, may serve as non-invasive biomarkers for monitoring disease progression and therapeutic efficacy in PD, offering hope for improved disease management.
3.7 Mucopolysaccharidosis IIIB
MPS IIIB (OMIM # 252920) is an LSD characterized by dysfunction of the lysosomal enzyme α-N-acetylglucosaminidase (NAGLU) (Rouse et al., 2024). Impaired NAGLU enzyme activity leads to the accumulation of heparan sulfate, primarily in the CNS (Rouse et al., 2024). Recently, Vargas-López et al. (2024) identified pronounced epigenetic alterations in two MPS IIIB patient-derived fibroblasts carrying different mutations in the NAGLU gene (Vargas-López et al., 2024). Although a global reduction in DNA methylation levels (5-methylcytosine) and the heterochromatin marker H3K9me3 was reported in both cell lines, the authors found that histone H3K14 acetylation was differentially altered in both MPS IIIB fibroblasts (Vargas-López et al., 2024). In fibroblasts carrying the P358L mutation, H3K14 acetylation was significantly increased. At the same time, it was found to be unaffected in E153K-containing MPS IIIB fibroblast models, supporting the notion that specific NAGLU mutations may differentially impact the epigenetic landscape in MPS IIIB.
3.8 Mucopolysaccharidosis IVA
MPS IVA (OMIM # 253000) is an LSD caused by mutations in the GALNS gene, which encodes for the lysosomal enzyme N-acetylgalactosamine-6-sulfatase (GALNS) (Leal et al., 2023). GALNS deficiency leads to the accumulation of keratan sulfate and chondroitin 6-sulfate, resulting in skeletal dysplasia and systemic manifestations in MPS IVA patients (Leal et al., 2023). Early studies conducted by Tomatsu et al. (2005) revealed methylation epigenetic alteration patterns at CpG dinucleotides in the GALNS gene (Tomatsu et al., 2005). Most recently, in vitro analyses of patient-derived fibroblasts have demonstrated a pattern of global DNA hypomethylation (Vargas-López et al., 2024). While the heterochromatin marker H3K9me3 remained preserved in MPS IVA, the H3K14 acetylation pattern in MPS IVA fibroblasts was found to be mutation-dependent, similar to that observed in MPS IIIB fibroblasts (Vargas-López et al., 2024). Remarkably, patients with MPS IVA harboring the A393S mutation had enhanced H3K14 acetylation. In contrast, the double mutation R94C/A393S did not affect the H3K14 acetylation profile (Vargas-López and Alméciga-Díaz, 2022), indicating that mutations can differentially influence epigenetic marks. These novel findings open new avenues for understanding and potentially treating MPS IVA.
4 Targeting epigenetic alterations in LSDs
Current evidence strongly supports the epigenetic dysregulation in several LSD. In consequence, seeking novel alternatives to restore the epigenetic alterations may offer a promising alternative for treating LSDs along with classical interventions, including ERT and GT. This last section explores classical and advanced approaches aimed at rescuing the altered epigenetic signature in LSDs.
4.1 Histone-modifying alternatives
The histone modification-based strategies rely on histone-modifying enzymes, including HDACs, HMTs, and KDMs (Yang et al., 2022). In NPC disease, loss of transcriptional silencing is observed, which impairs the proper function of oligodendrocytes (Kunkel et al., 2023). Notably, GSK-J4, a KDMi, has been reported to restore the maturation of NPC oligodendrocyte progenitor cells (Kunkel et al., 2023), supporting the concept that correcting the epigenetic signature could represent a valid therapeutic approach. Interestingly, HATi, such as garcinol, may be more suitable in conditions where histone hyperacetylation has been observed, for example, in MPS IIIB and MPS IVA, where specific mutations are associated with increased H3K14 acetylation (Vargas-López and Alméciga-Díaz, 2022). Taken together, these considerations highlight that the use of epigenetic modulators in LSDs should be individualized, guided by mutation-specific epigenetic signatures and the underlying chromatin contexts.
4.2 Epigenetic readers
Epigenetic readers are a class of proteins that bind to domains selectively recognizing covalent modifications on DNA, histones, and non-histone proteins deposited by epigenetic writers (Damiani et al., 2020). Upon binding, epigenetic readers recruit effector complexes that modify chromatin architecture and influence gene expression (Damiani et al., 2020). Consequently, targeting epigenetic reader proteins could be a promising alternative in epigenome modulation of LSDs. For instance, Apabetalone (RVX-208), which binds to the BD2 domain of the bromodomain and extraterminal-containing protein family (BET), has been shown to suppress proinflammatory transcriptional programs in innate immune cells from patients with FD undergoing ERT (Fu et al., 2022). Interestingly, RVX-208 led to a decrease in proinflammatory markers, including TNF-α, IL-12, MCP-1, and IL-6, as well as reduced oxidative stress (Fu et al., 2022), further supporting the notion that epigenetic modulators can positively impact the pathogenic features in LSD, such as FD.
4.3 microRNA-targeted approaches
miRNAs are the longest ncRNAs studied in LSD and are widely recognized to be altered (Morena et al., 2019; Xiao et al., 2019; Üzen et al., 2025; Tarallo et al., 2019; Dasgu et al., 2015). Since miRNA tightly regulates post-transcriptional gene expression of several factors involved in the pathogenesis of LSDs, such as lysosomal dysfunction, autophagy, immune activation, and oxidative stress, among others, miRNA are potential targets for treating LSDs. For instance, in KD, the downregulation of miR-219 has been identified as a key trigger of OPC dysfunction, which limits their maturation into oligodendrocytes (Inamura et al., 2021). Interestingly, treatment with exogenous miR-219 has shown potential in restoring maturation markers (MBP and PLP), while reducing caspase-3 activation and psychosine levels (Inamura et al., 2021), highlighting that targeting dysregulated miRNA may be an alternative for treating KD. Further research and clinical trials are necessary to fully explore the potential of miRNA-targeted approaches in LSDs; however, the initial results are promising.
In GD, miRNA profiling has consistently detected dysregulated miRNAs, which likely play a role in inflammation, synaptic dysfunction, and mitochondrial stress, ultimately contributing to the development of certain cancers (Üzen et al., 2025; Dasgu et al., 2015). Although there are no studies directly targeting miRNA in GD, the use of isofagomine, a pharmacological chaperone for GCase, normalizes approximately 40%–60% of altered miRNA in the brains of GD mouse models, suggesting that classical approaches may influence the epigenome in some LSDs (Dasgu et al., 2015). This finding underscores the potential of miRNA as a therapeutic target in LSDs, including GD, and warrants further investigation into the role of miRNA dysregulation in the pathogenesis of these diseases.
4.4 CRISPR/Cas9-based epigenome editing
Early reports demonstrated that the Clustered Regularly Interspaced Short Palindromic Repeats and CRISPR-associated protein 9 (CRISPR/Cas9) system is an immune system that protects prokaryotes from re-infection by phages (Barrangou et al., 2007; Jiang and Doudna, 2017). Later, studies conducted by Doudna and Charpentier demonstrated the suitability of CRISPR/Cas9 in recognizing, binding to, and cutting genomic DNA with high precision (Jiang and Doudna, 2017; Leal et al., 2024; Zhang, 2021), leading to development of a promising genome editing approach, which is currently used to induce knockouts and knock-ins (Figure 2), and resulting in its application for innovative gene therapies for treating human diseases (Sharma et al., 2021; Wan et al., 2023).
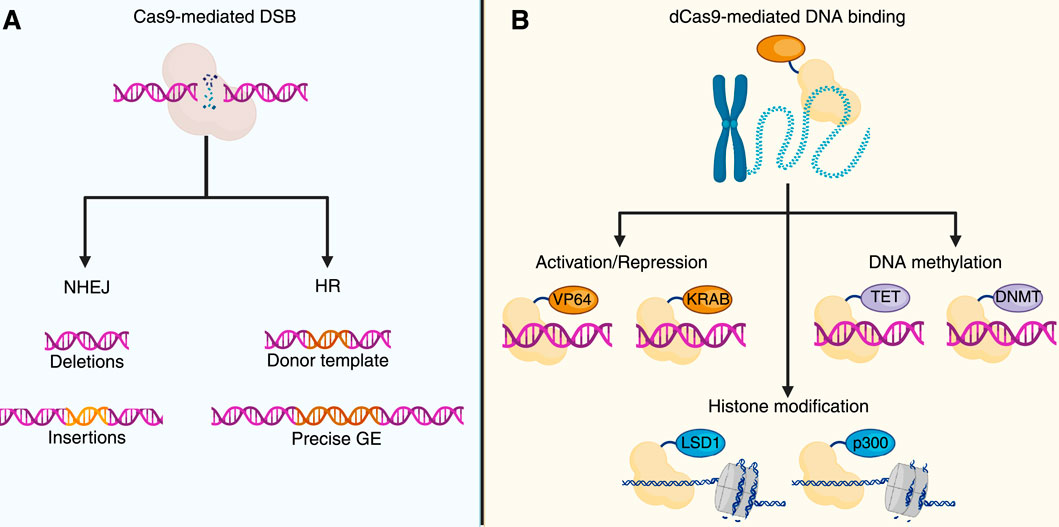
Figure 2. CRISPR/Cas and genome and epigenome editing. (A) CRISPR/Cas9 genome editing uses a catalytically active Cas9 that induces double-strand breaks (DSBs) at target loci. DSB repair by NHEJ may introduce small indels, while, in the presence of a donor DNA template, HR facilitates targeted genome modification. (B) CRISPR/Cas9-based epigenome editing (GE) utilizes a catalytically dead Cas9 (dCas9), which allows for the targeted recruitment of effector domains without inducing DNA cleavage. dCas9 fused to transcriptional activators (e.g., VP64) or repressors (e.g., KRAB) modulates gene expression. Joining with ten-eleven translocation enzymes (TET) or DNA methyltransferases (DNMT) allows locus-specific demethylation or methylation of the DNA. Histone modifications can also be mediated by dCas9-fused effectors, such as lysine-specific demethylase 1 (LSD1) or histone acetyltransferase p300. The image was created by BioRender.com.
The CRISPR/Cas9 system includes a single guide RNA (sgRNA) that binds to a desired DNA sequence and recruits the Cas9 protein. Interaction of Cas9 with a protospacer adjacent motif (PAM) in DNA precedes sgRNA-mediated DNA binding (Jiang and Doudna, 2017; Jinek et al., 2012). Moreover, Cas9 introduces a double-strand break (DSB) in the DNA, ultimately promoting the activation of the DSB repair. Cas9 has two nuclease domains (HNH and RuvC) that generate the DSBs (Jiang and Doudna, 2017). If a donor template is co-delivered into the nucleus, homologous recombination takes place, and exogenous DNA information is inserted (knock-in). Instead, when no donor is available, the non-homologous end-joining (NHEJ) pathway is activated, introducing random insertions and deletions (indels), which can lead to knockouts (Leal et al., 2024; Chen et al., 2024). Interestingly, novel approaches have combined the extraordinary ability of CRISPR/Cas9 with the use of enzymes capable of modifying epigenetic signatures, resulting in a novel approach termed CRISPR/Cas9-based epigenome editing (EE) (Villiger et al., 2024; Fadul et al., 2023). This innovative approach utilizes the recognizing and binding properties of the CRISPR/Cas9 system while avoiding Cas9-mediated DSBs by employing a catalytically inactive Cas9 enzyme, also known as dead Cas9 (dCas9) (Cai et al., 2023). dCas9 carries two mutations in the HNH (H840A) and RuvC (D10A) domains that prevent DSBs within the DNA (Liu et al., 2020; Whinn et al., 2019). Along with dCas9, the CRISPR/Cas9-based EE leverages epigenetic effectors that can inhibit or activate gene transcription (Figure 2) (Koonin et al., 2023; Sar and Dalai, 2021). Table 2 summarizes common effectors used in CRISPR/Cas-based EE.
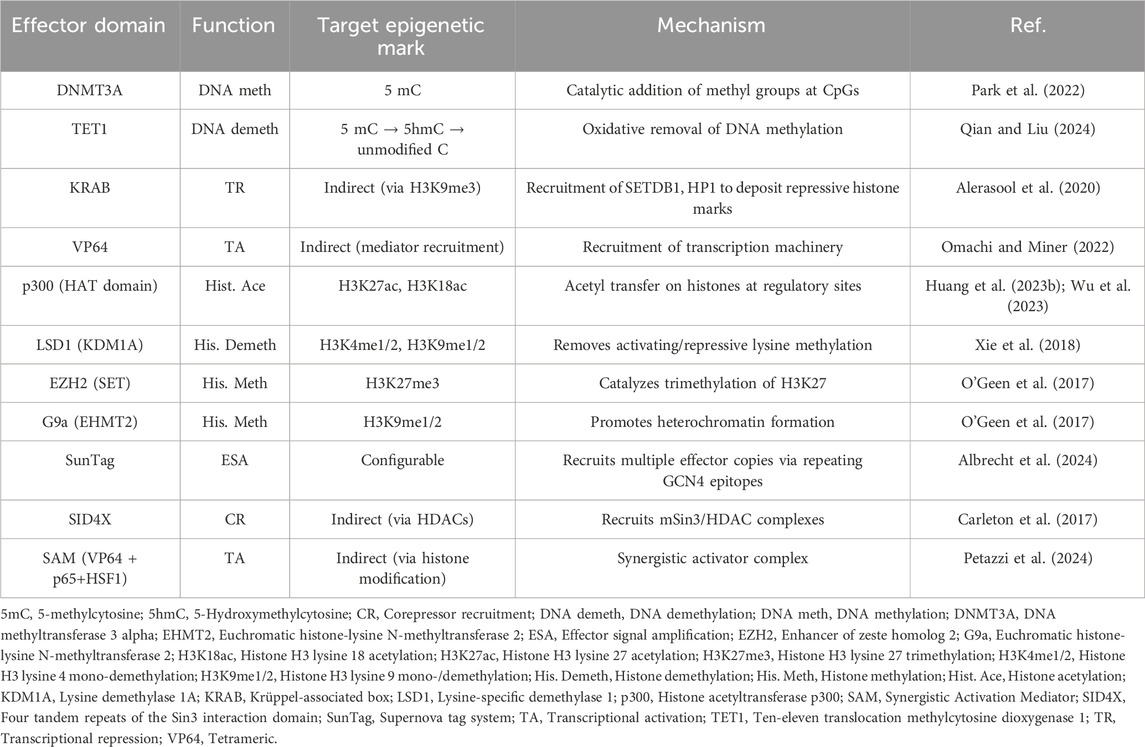
Table 2. Common epigenetic effectors evaluated in CRISPR/Cas9-based EE approaches.
CRISPR/Cas9-based EE has been evaluated in several rare diseases, including Prader–Willi Syndrome (Rohm et al., 2025), Facioscapulohumeral Muscular Dystrophy (Mariot and Dumonceaux, 2022), and Fragile X Syndrome (Liu et al., 2018), among others. However, its true potential lies in its application to LSDs, where it has not yet been tested. Indeed, the epigenetic landscape observed in multiple LSD models strongly suggests that epigenome profile restoration may be within reach through CRISPR/Cas9-based EE, thereby offering a new avenue for treating LSDs (Figure 2).
5 Future perspectives
The understanding of the epigenetic landscape in LSDs provides a new paradigm in the LSD pathogenesis, from monogenic, substrate-accumulating diseases to complex disorders involving transcriptional and chromatin-level dysregulation, which contribute to the disturbance of cell homeostasis. Importantly, as epigenetic modifications can be reversed, several therapeutic strategies hold enormous potential to ameliorate the epigenetic alterations in LSDs (Figure 3). In this regard, the use of HAT inhibitors, epigenetic readers modulators, miRNA mimics, or antagonists may provide a further direction in treating LSDs. Beyond pharmacological approaches, the interplay between metabolism and the epigenome opens new opportunities for nutritional and metabolic interventions in LSDs. Strategies aimed at supporting one-carbon metabolism, including supplementation with methyl donors such as folate, choline, or betaine, may help restore the SAM/SAH ratio and partially correct DNA hypomethylation (Bernasocchi and Mostoslavsky, 2024; Inoue-Choi et al., 2012). Evidence from metabolic and neurodegenerative disorders suggests that such interventions may improve epigenetic homeostasis (Bekdash, 2021; McKee et al., 2017; Araki et al., 2022). Although these approaches remain largely unexplored in LSDs, metabolic adjuvants could represent safe, accessible, and potentially synergistic options to complement ERT, GT, or emerging epigenetic treatments.
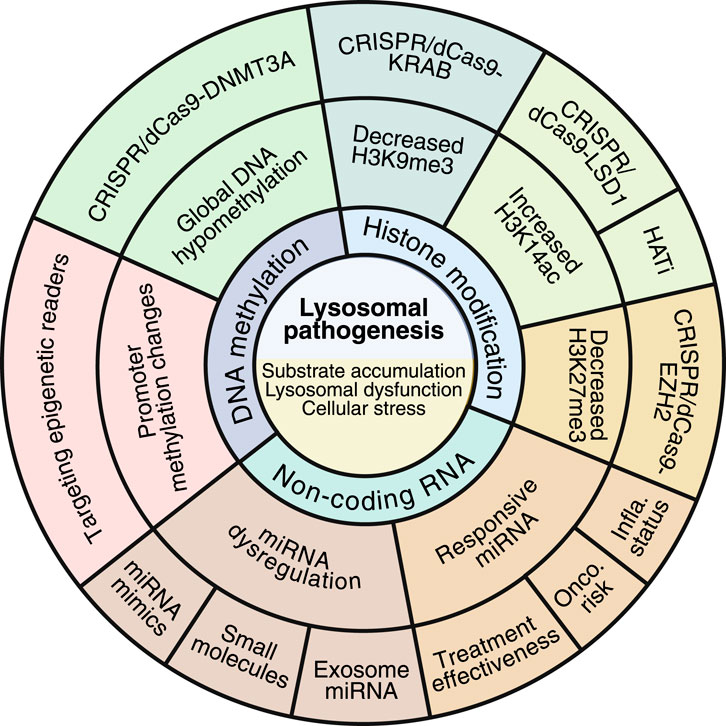
Figure 3. Kaleidoscope view of the epigenetic landscape in LSDs. The epigenetic landscape in LSDs has switched the classical view of these diseases from substrate-accumulating diseases to complex inherited metabolic disorders in which several epigenetic actors are actively contributing to the global cell homeostasis impairment. While epigenome alterations are gaining importance in the pathogenesis of LSDs, novel therapeutic alternatives are emerging to rescue these alterations. Likewise, some epigenetic marks have been recognized as playing a role in monitoring disease progression, prognosis, and oncogenic risk. The image was created by BioRender.com.
Novel studies have addressed the use of exosomes for transporting miRNA in non-LSD (Huang et al., 2023a), which may also constitute a promising alternative in the treatment of LSDs. Indeed, the large number of FDA-approved molecules may accelerate the implementation of epigenetic drugs into the classical approaches used in LSDs. Likewise, the latest advances in the CRISPR/Cas9 system, a revolutionary tool for editing the epigenome, may offer a novel and exciting strategy for restoring epigenetic disturbances in LSDs, thereby increasing the treatment options available.
Finally, the use of epigenomic signatures may benefit early diagnosis, monitor therapeutic response, and classify patients, especially in LSDs such as Gaucher and Fabry, where epigenetic perturbations may influence oncogenic risk or inflammatory status (Figure 3). However, it is crucial to emphasize the importance of systematically investigating sex-specific epigenetic variation and mutation-dependent chromatin states, recently highlighted in Fabry and MPS subtypes. These variations could be key factors in the progression and prognosis of LSDs, making this research an urgent and significant area of study.
Author contributions
AL: Conceptualization, Writing – original draft, Writing – review and editing, Formal Analysis, Investigation, Methodology, Software, Visualization. HP: Conceptualization, Investigation, Writing – original draft, Writing – review and editing, Validation. ST: Conceptualization, Writing – original draft, Writing – review and editing, Funding acquisition, Project administration, Resources, Supervision.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. AF and HP were supported by Universidad Icesi and Fundación Valle de Lili (Contract ID: 2025-ART-001). This work was also supported by grants from the Austrian MPS society, A Cure for Robert, Inc., The Carol Ann Foundation, Angelo R. Cali and Mary V. Cali Family Foundation, Inc., The Vain and Harry Fish Foundation, Inc., The Bennett Foundation, Jacob Randall Foundation, and Nemours Funds. ST was supported by an Institutional Development Award from the Eunice Kennedy Shriver National Institute of Child Health and Human Development of the National Institutes of Health (NICHD) (1R01HD102545-01A1).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ago, Y., Rintz, E., Musini, K. S., Ma, Z., and Tomatsu, S. (2024). Molecular mechanisms in pathophysiology of mucopolysaccharidosis and prospects for innovative therapy. Int. J. Mol. Sci. 25 (2), 1113. doi:10.3390/ijms25021113
Albrecht, C., Rajaram, N., Broche, J., Bashtrykov, P., and Jeltsch, A. (2024). Locus-specific and stable DNA demethylation at the H19/IGF2 ICR1 by epigenome editing using a dCas9-SunTag system and the catalytic domain of TET1. Genes (Basel) 15 (1). doi:10.3390/genes15010080
Alerasool, N., Segal, D., Lee, H., and Taipale, M. (2020). An efficient KRAB domain for CRISPRi applications in human cells. Nat. Methods 17 (11), 1093–1096. doi:10.1038/s41592-020-0966-x
Ali, M. M., Naquiallah, D., Qureshi, M., Mirza, M. I., Hassan, C., Masrur, M., et al. (2022). DNA methylation profile of genes involved in inflammation and autoimmunity correlates with vascular function in morbidly obese adults. Epigenetics 17 (1), 93–109. doi:10.1080/15592294.2021.1876285
Angeloni, A., and Bogdanovic, O. (2019). Enhancer DNA methylation: implications for gene regulation. Essays Biochem. 63 (6), 707–715. doi:10.1042/EBC20190030
Araki, R., Nishida, S., Oishi, Y., Tachioka, H., Kita, A., and Yabe, T. (2022). Methyl donor supplementation prevents a folate deficiency-induced depression-like State and neuronal immaturity of the dentate gyrus in mice. Neuroscience 485, 12–22. doi:10.1016/j.neuroscience.2022.01.007
Barrangou, R., Fremaux, C., Deveau, H., Richards, M., Boyaval, P., Moineau, S., et al. (2007). CRISPR provides acquired resistance against viruses in prokaryotes. Science 315 (5819), 1709–1712. doi:10.1126/science.1138140
Bekdash, R. A. (2021). Early life nutrition and mental health: the role of DNA methylation. Nutrients 13 (9), 3111. doi:10.3390/nu13093111
Bernasocchi, T., and Mostoslavsky, R. (2024). Subcellular one carbon metabolism in cancer, aging and epigenetics. Front. Epigenet Epigenom 2, 1451971. doi:10.3389/freae.2024.1451971
Cai, R., Lv, R., Shi, X., Yang, G., and Jin, J. (2023). CRISPR/dCas9 tools: epigenetic mechanism and application in gene transcriptional regulation. Int. J. Mol. Sci. 24 (19), 14865. doi:10.3390/ijms241914865
Carleton, J. B., Berrett, K. C., and Gertz, J. (2017). Multiplex enhancer interference reveals collaborative control of gene regulation by Estrogen receptor α-Bound enhancers. Cell. Syst. 5 (4), 333–44.e5. doi:10.1016/j.cels.2017.08.011
Chen, L. L., and Kim, V. N. (2024). Small and long non-coding RNAs: past, present, and future. Cell. 187 (23), 6451–6485. doi:10.1016/j.cell.2024.10.024
Chen, X., Du, J., Yun, S., Xue, C., Yao, Y., and Rao, S. (2024). Recent advances in CRISPR-Cas9-based genome insertion technologies. Mol. Ther. Nucleic Acids 35 (1), 102138. doi:10.1016/j.omtn.2024.102138
Damiani, E., Duran, M. N., Mohan, N., Rajendran, P., and Dashwood, R. H. (2020). Targeting epigenetic 'Readers' with natural compounds for cancer interception. J. Cancer Prev. 25 (4), 189–203. doi:10.15430/JCP.2020.25.4.189
Danieli, M. G., Casciaro, M., Paladini, A., Bartolucci, M., Sordoni, M., Shoenfeld, Y., et al. (2024). Exposome: epigenetics and autoimmune diseases. Autoimmun. Rev. 23 (6), 103584. doi:10.1016/j.autrev.2024.103584
Dasgupta, N., Xu, Y. H., Li, R., Peng, Y., Pandey, M. K., Tinch, S. L., et al. (2015). Neuronopathic gaucher disease: dysregulated mRNAs and miRNAs in brain pathogenesis and effects of pharmacologic chaperone treatment in a mouse model. Hum. Mol. Genet. 24 (24), 7031–7048. doi:10.1093/hmg/ddv404
De Plano, L. M., Saitta, A., Oddo, S., and Caccamo, A. (2024). Epigenetic changes in alzheimer's disease: DNA methylation and histone modification. Cells 13 (8), 719. doi:10.3390/cells13080719
Fadul, S. M., Arshad, A., and Mehmood, R. (2023). CRISPR-based epigenome editing: mechanisms and applications. Epigenomics 15 (21), 1137–1155. doi:10.2217/epi-2023-0281
Fu, L., Wasiak, S., Tsujikawa, L. M., Rakai, B. D., Stotz, S. C., Wong, N. C. W., et al. (2022). Inhibition of epigenetic reader proteins by apabetalone counters inflammation in activated innate immune cells from fabry disease patients receiving enzyme replacement therapy. Pharmacol. Res. Perspect. 10 (3), e00949. doi:10.1002/prp2.949
George, K. A., Anding, A. L., van der Flier, A., Tomassy, G. S., Berger, K. I., Zhang, T. Y., et al. (2024). Pompe disease: unmet needs and emerging therapies. Mol. Genet. Metab. 143 (3), 108590. doi:10.1016/j.ymgme.2024.108590
Ghosh, P., and Saadat, A. (2023). Neurodegeneration and epigenetics: a review. Neurologia (Engl Ed). 38 (6), e62–e68. doi:10.1016/j.nrleng.2023.05.001
Hassan, S., Sidransky, E., and Tayebi, N. (2017). The role of epigenetics in lysosomal storage disorders: uncharted territory. Mol Genet Metab 122(3), 10–18. doi:10.1016/j.ymgme.2017.07.012
Huang, C. C., Kang, M., Leung, K., Lu, Y., Shirazi, S., Gajendrareddy, P., et al. (2023a). Micro RNA based MSC EV engineering: targeting the BMP2 cascade for bone repair. Front. Cell. Dev. Biol. 11, 1127594. doi:10.3389/fcell.2023.1127594
Huang, H., Zhang, W., Zhang, J., Zhao, A., and Jiang, H. (2023b). Epigenome editing based on CRISPR/dCas9p300 facilitates transdifferentiation of human fibroblasts into Leydig-like cells. Exp. Cell. Res. 425 (2), 113551. doi:10.1016/j.yexcr.2023.113551
Inamura, N., Go, S., Watanabe, T., Takase, H., Takakura, N., Nakayama, A., et al. (2021). Reduction in miR-219 expression underlies cellular pathogenesis of oligodendrocytes in a mouse model of Krabbe disease. Brain Pathol. 31 (5), e12951. doi:10.1111/bpa.12951
Inoue-Choi, M., Nelson, H. H., Robien, K., Arning, E., Bottiglieri, T., Koh, W. P., et al. (2012). One-carbon metabolism nutrient status and plasma S-adenosylmethionine concentrations in middle-aged and older Chinese in Singapore. Int. J. Mol. Epidemiol. Genet. 3 (2), 160–173.
Jain, R., and Epstein, J. A. (2024). Epigenetics. Adv. Exp. Med. Biol. 1441, 341–364. doi:10.1007/978-3-031-44087-8_18
Jiang, F., and Doudna, J. A. (2017). CRISPR-Cas9 structures and mechanisms. Annu. Rev. Biophys. 46, 505–529. doi:10.1146/annurev-biophys-062215-010822
Jinek, M., Chylinski, K., Fonfara, I., Hauer, M., Doudna, J. A., and Charpentier, E. (2012). A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 337 (6096), 816–821. doi:10.1126/science.1225829
Kennedy, B. E., Hundert, A. S., Goguen, D., Weaver, I. C., and Karten, B. (2016). Presymptomatic alterations in amino acid metabolism and DNA methylation in the cerebellum of a murine model of niemann-pick type C disease. Am. J. Pathol. 186 (6), 1582–1597. doi:10.1016/j.ajpath.2016.02.012
Knapp, K., Naik, N., Ray, S., van Haaften, G., and Bicknell, L. S. (2023). Histones: coming of age in Mendelian genetic disorders. J. Med. Genet. 60 (3), 213–222. doi:10.1136/jmg-2022-109085
Koonin, E. V., Gootenberg, J. S., and Abudayyeh, O. O. (2023). Discovery of diverse CRISPR-cas systems and expansion of the genome engineering toolbox. Biochemistry 62 (24), 3465–3487. doi:10.1021/acs.biochem.3c00159
Kunkel, T. J., Townsend, A., Sullivan, K. A., Merlet, J., Schuchman, E. H., Jacobson, D. A., et al. (2023). The cholesterol transporter NPC1 is essential for epigenetic regulation and maturation of oligodendrocyte lineage cells. Nat. Commun. 14 (1), 3964. doi:10.1038/s41467-023-39733-6
Leal, A. F., and Alméciga-Díaz, C. J. (2022). Efficient CRISPR/Cas9 nickase-mediated genome editing in an in vitro model of mucopolysaccharidosis IVA. Gene Ther., 1–8.
Leal, A. F., Espejo-Mojica, A. J., Sánchez, O. F., Ramírez, C. M., Reyes, L. H., Cruz, J. C., et al. (2020a). Lysosomal storage diseases: current therapies and future alternatives. J. Mol. Med. (Berl) 98, 931–946. doi:10.1007/s00109-020-01935-6
Leal, A. F., Benincore-Flórez, E., Solano-Galarza, D., Garzón Jaramillo, R. G., Echeverri-Peña, O. Y., Suarez, D. A., et al. (2020b). GM2 gangliosidoses: clinical features, pathophysiological aspects, and current therapies. Int. J. Mol. Sci. 21 (17), 1–27. doi:10.1038/s41434-022-00344-3
Leal, A. F., Benincore-Flórez, E., Rintz, E., Herreño-Pachón, A. M., Celik, B., Ago, Y., et al. (2022). Mucopolysaccharidoses: cellular consequences of glycosaminoglycans accumulation and potential targets. Int. J. Mol. Sci. 24 (1), 477. doi:10.3390/ijms24010477
Leal, A. F., Alméciga-Díaz, C. J., and Tomatsu, S. (2023). Mucopolysaccharidosis IVA: current disease models and drawbacks. Int. J. Mol. Sci. 24 (22), 16148. doi:10.3390/ijms242216148
Leal, A. F., Herreno-Pachón, A. M., Benincore-Flórez, E., Karunathilaka, A., and Tomatsu, S. (2024). Current strategies for increasing Knock-In efficiency in CRISPR/Cas9-Based approaches. Int. J. Mol. Sci. 25 (5), 2456. doi:10.3390/ijms25052456
Lee, D., and Hong, J. H. (2023). Niemann-pick disease type C (NPDC) by mutation of NPC1 and NPC2: aberrant lysosomal cholesterol trafficking and oxidative stress. Antioxidants (Basel) 12 (12), 2021. doi:10.3390/antiox12122021
Lenders, M., and Brand, E. (2021). Precision medicine in fabry disease. Nephrol. Dial. Transpl. 36 (Suppl. 2), 14–23. doi:10.1093/ndt/gfab038
Li, S., and Tollefsbol, T. O. (2021). DNA methylation methods: global DNA methylation and methylomic analyses. Methods. 187, 28–43. doi:10.1016/j.ymeth.2020.10.002
Liu, X. S., Wu, H., Krzisch, M., Wu, X., Graef, J., Muffat, J., et al. (2018). Rescue of fragile X syndrome neurons by DNA methylation editing of the FMR1 gene. Cell. 172 (5), 979–92.e6. doi:10.1016/j.cell.2018.01.012
Liu, Z., Liao, Z., Chen, Y., Han, L., Yin, Q., and Xiao, H. (2020). Application of various delivery methods for CRISPR/dCas9. Mol. Biotechnol. 62 (8), 355–363. doi:10.1007/s12033-020-00258-8
Lossi, L., Castagna, C., and Merighi, A. (2024). An overview of the epigenetic modifications in the brain under normal and pathological conditions. Int. J. Mol. Sci. 25 (7), 3881. doi:10.3390/ijms25073881
Maghazachi, A. A. (2023). Globoid cell leukodystrophy (Krabbe disease): an update. Immunotargets Ther. 12, 105–111. doi:10.2147/ITT.S424622
Maier, N., Gatterer, C., Haider, P., Salzmann, M., Kaun, C., Speidl, W. S., et al. (2021). MiRNA Let-7a and Let-7d are induced by globotriaosylceramide via NF-kB activation in fabry disease. Genes (Basel) 12 (8), 1184. doi:10.3390/genes12081184
Mariot, V., and Dumonceaux, J. (2022). Gene editing to tackle facioscapulohumeral muscular dystrophy. Front. Genome 4, 937879. doi:10.3389/fgeed.2022.937879
Mattei, A. L., Bailly, N., and Meissner, A. (2022). DNA methylation: a historical perspective. Trends Genet. 38 (7), 676–707. doi:10.1016/j.tig.2022.03.010
McKee, S. E., Grissom, N. M., Herdt, C. T., and Reyes, T. M. (2017). Methyl donor supplementation alters cognitive performance and motivation in female offspring from high-fat diet-fed dams. FASEB J. 31 (6), 2352–2363. doi:10.1096/fj.201601172R
Millán-Zambrano, G., Burton, A., Bannister, A. J., and Schneider, R. (2022). Histone post-translational modifications - cause and consequence of genome function. Nat. Rev. Genet. 23 (9), 563–580. doi:10.1038/s41576-022-00468-7
Minervini, G., Franco, R., Marrapodi, M. M., Mehta, V., Fiorillo, L., Badnjević, A., et al. (2023). Gaucher: a systematic review on oral and radiological aspects. Medicina (Kaunas) 59 (4), 670. doi:10.3390/medicina59040670
Morena, F., Oikonomou, V., Argentati, C., Bazzucchi, M., Emiliani, C., Gritti, A., et al. (2019). Integrated computational analysis highlights unique miRNA signatures in the subventricular zone and striatum of GM2 gangliosidosis animal models. Int. J. Mol. Sci. 20 (13), 3179. doi:10.3390/ijms20133179
Nemeth, K., Bayraktar, R., Ferracin, M., and Calin, G. A. (2024). Non-coding RNAs in disease: from mechanisms to therapeutics. Nat. Rev. Genet. 25 (3), 211–232. doi:10.1038/s41576-023-00662-1
Nishiyama, A., and Nakanishi, M. (2021). Navigating the DNA methylation landscape of cancer. Trends Genet. 37 (11), 1012–1027. doi:10.1016/j.tig.2021.05.002
O'Geen, H., Ren, C., Nicolet, C. M., Perez, A. A., Halmai, J., Le, V. M., et al. (2017). dCas9-based epigenome editing suggests acquisition of histone methylation is not sufficient for target gene repression. Nucleic Acids Res. 45 (17), 9901–9916. doi:10.1093/nar/gkx578
Omachi, K., and Miner, J. H. (2022). Comparative analysis of dCas9-VP64 variants and multiplexed guide RNAs mediating CRISPR activation. PLoS One 17 (6), e0270008. doi:10.1371/journal.pone.0270008
Parenti, G., Medina, D. L., and Ballabio, A. (2021). The rapidly evolving view of lysosomal storage diseases. EMBO Mol. Med. 13 (2), e12836. doi:10.15252/emmm.202012836
Park, H., Shin, J., Kim, Y., Saito, T., Saido, T. C., and Kim, J. (2022). CRISPR/dCas9-Dnmt3a-mediated targeted DNA methylation of APP rescues brain pathology in a mouse model of alzheimer's disease. Transl. Neurodegener. 11 (1), 41. doi:10.1186/s40035-022-00314-0
Petazzi, P., Gutierrez-Agüera, F., Roca-Ho, H., Castaño, J., Bueno, C., Alvarez, N., et al. (2024). Generation of an inducible dCas9-SAM human PSC line for endogenous gene activation. Front. Cell. Dev. Biol. 12, 1484955. doi:10.3389/fcell.2024.1484955
Qian, J., and Liu, S. X. (2024). CRISPR/dCas9-Tet1-Mediated DNA methylation editing. Bio Protoc. 14 (8), e4976. doi:10.21769/BioProtoc.4976
Ramaiah, M. J., Tangutur, A. D., and Manyam, R. R. (2021). Epigenetic modulation and understanding of HDAC inhibitors in cancer therapy. Life Sci. 277, 119504. doi:10.1016/j.lfs.2021.119504
Rohm, D., Black, J. B., McCutcheon, S. R., Barrera, A., Berry, S. S., Morone, D. J., et al. (2025). Activation of the imprinted Prader-Willi syndrome locus by CRISPR-based epigenome editing. Cell. Genom 5 (2), 100770. doi:10.1016/j.xgen.2025.100770
Rouse, C. J., Jensen, V. N., and Heldermon, C. D. (2024). Mucopolysaccharidosis type IIIB: a current review and exploration of the AAV therapy landscape. Neural Regen. Res. 19 (2), 355–359. doi:10.4103/1673-5374.377606
Sar, P., and Dalai, S. (2021). CRISPR/Cas9 in epigenetics studies of health and disease. Prog. Mol. Biol. Transl. Sci. 181, 309–343. doi:10.1016/bs.pmbts.2021.01.022
Sharma, G., Sharma, A. R., Bhattacharya, M., Lee, S. S., and Chakraborty, C. (2021). CRISPR-Cas9: a preclinical and clinical perspective for the treatment of human diseases. Mol. Ther. 29 (2), 571–586. doi:10.1016/j.ymthe.2020.09.028
Shen, J. S., Balaji, U., Shigeyasu, K., Okugawa, Y., Jabbarzadeh-Tabrizi, S., Day, T. S., et al. (2022). Dysregulated DNA methylation in the pathogenesis of fabry disease. Mol. Genet. Metab. Rep. 33, 100919. doi:10.1016/j.ymgmr.2022.100919
Simoncini, C., Torri, S., Montano, V., Chico, L., Gruosso, F., Tuttolomondo, A., et al. (2020). Oxidative stress biomarkers in fabry disease: is there a room for them? J. Neurol. 267 (12), 3741–3752. doi:10.1007/s00415-020-10044-w
Stepien, K. M., Roncaroli, F., Turton, N., Hendriksz, C. J., Roberts, M., Heaton, R. A., et al. (2020). Mechanisms of mitochondrial dysfunction in lysosomal storage disorders: a review. J. Clin. Med. 9 (8), 2596. doi:10.3390/jcm9082596
Tarallo, A., Carissimo, A., Gatto, F., Nusco, E., Toscano, A., Musumeci, O., et al. (2019). microRNAs as biomarkers in pompe disease. Genet. Med. 21 (3), 591–600. doi:10.1038/s41436-018-0103-8
Tomatsu, S., Montaño, A. M., Nishioka, T., Gutierrez, M. A., Peña, O. M., Tranda Firescu, G. G., et al. (2005). Mutation and polymorphism spectrum of the GALNS gene in mucopolysaccharidosis IVA (Morquio A). Hum. Mutat. 26 (6), 500–512. doi:10.1002/humu.20257
Toro, C., Zainab, M., and Tifft, C. J. (2021). The GM2 gangliosidoses: unlocking the mysteries of pathogenesis and treatment. Neurosci. Lett. 764, 136195–29. doi:10.1016/j.neulet.2021.136195
Üzen, R., Bayram, F., Dursun, H., Kardas, F., Cakir, M., Hasan Sohel, M. M., et al. (2025). Tumor suppressor and oncogenic miRNA expressions in patients with type I gaucher disease and carriers. Microrna 14. doi:10.2174/0122115366342286250216032611
Vargas-López, V., Prada, L. F., and Alméciga-Díaz, C. J. (2024). Evidence of epigenetic landscape shifts in mucopolysaccharidosis IIIB and IVA. Sci. Rep. 14, 3961. doi:10.1038/s41598-024-54626-4
Villiger, L., Joung, J., Koblan, L., Weissman, J., Abudayyeh, O. O., and Gootenberg, J. S. (2024). CRISPR technologies for genome, epigenome and transcriptome editing. Nat. Rev. Mol. Cell. Biol. 25 (6), 464–487. doi:10.1038/s41580-023-00697-6
Virciglio, C., Abel, Y., and Rederstorff, M. (2021). Regulatory non-coding RNAs: an overview. Methods Mol. Biol. 2300, 3–9. doi:10.1007/978-1-0716-1386-3_1
Wang, J. Y., and Doudna, J. A. (2023). CRISPR technology: a decade of genome editing is only the beginning. Science 379 (6629), eadd8643. doi:10.1126/science.add8643
Whinn, K. S., Kaur, G., Lewis, J. S., Schauer, G. D., Mueller, S. H., Jergic, S., et al. (2019). Nuclease dead Cas9 is a programmable roadblock for DNA replication. Sci. Rep. 9 (1), 13292. doi:10.1038/s41598-019-49837-z
Wilson, T. S., and Scaffidi, P. (2025). Compromised epigenetic robustness in cancer: fueling evolution, exposing weakness. Trends Cancer 11 (6), 575–590. doi:10.1016/j.trecan.2025.02.001
Wu, Q., Wu, J., Karim, K., Chen, X., Wang, T., Iwama, S., et al. (2023). Massively parallel characterization of CRISPR activator efficacy in human induced pluripotent stem cells and neurons. Mol. Cell. 83 (7), 1125–39.e8. doi:10.1016/j.molcel.2023.02.011
Xiao, K., Lu, D., Hoepfner, J., Santer, L., Gupta, S., Pfanne, A., et al. (2019). Circulating microRNAs in fabry disease. Sci. Rep. 9 (1), 15277. doi:10.1038/s41598-019-51805-6
Xie, N., Zhou, Y., Sun, Q., and Tang, B. (2018). Novel epigenetic techniques provided by the CRISPR/Cas9 system. Stem Cells Int. 2018, 7834175. doi:10.1155/2018/7834175
Yang, Y., Zhang, M., and Wang, Y. (2022). The roles of histone modifications in tumorigenesis and associated inhibitors in cancer therapy. J. Natl. Cancer Cent. 2 (4), 277–290. doi:10.1016/j.jncc.2022.09.002
Zhang, B. (2021). CRISPR/Cas gene therapy. J. Cell. Physiol. 236 (4), 2459–2481. doi:10.1002/jcp.30064
Keywords: epigenetics, histones, lysosomal storage disorders, methylation, miRNA
Citation: Leal AF, Pachajoa H and Tomatsu S (2025) Epigenetic landscape in lysosomal storage disorders: mechanisms and modulation. Front. Genet. 16:1679497. doi: 10.3389/fgene.2025.1679497
Received: 05 August 2025; Accepted: 20 October 2025;
Published: 11 November 2025.
Edited by:
Qiao Li, University of Ottawa, CanadaReviewed by:
Adriana Mantegazza, Thomas Jefferson University, United StatesCopyright © 2025 Leal, Pachajoa and Tomatsu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shunji Tomatsu, c2h1bmppLnRvbWF0c3VAbmVtb3Vycy5jb20=