 Muhammad Ghazanfar Abbas1
Muhammad Ghazanfar Abbas1 Muhammad Binyameen1†
Muhammad Binyameen1† Muhammad Azeem2*†
Muhammad Azeem2*† Shahid Majeed3Zahid Mehmood Sarwar1Abdul Nazir4Mahar Muhammad Imran Sharif1Amna Parveen2
Shahid Majeed3Zahid Mehmood Sarwar1Abdul Nazir4Mahar Muhammad Imran Sharif1Amna Parveen2 Raimondas Mozūratis5,6*†
Raimondas Mozūratis5,6*†- 1Laboratory of Insect Chemical Ecology, Department of Entomology, Faculty of Agricultural Sciences & Technology, Bahauddin Zakariya University, Multan, Pakistan
- 2Department of Chemistry, COMSATS University Islamabad, Abbottabad, Pakistan
- 3Department of Entomology, University of Agriculture, Faisalabad, Pakistan
- 4Department of Environmental Sciences, COMSATS University Islamabad, Abbottabad, Pakistan
- 5Laboratory of Chemical and Behavioural Ecology, Institute of Ecology, State Scientific Research Institute Nature Research Centre, Vilnius, Lithuania
- 6Department of Zoology, Stockholm University, Stockholm, Sweden
Plant-based essential oils have gained attention as a natural alternative for controlling mosquitoes due to their repellent, larvicidal and oviposition deterrent properties. We tested repellent, larvicidal, and oviposition deterrent effects of essential oils (EOs) of Mentha spicata (L.), Ocimum basilicum (L.), and Abutilon indicum (L.) against three mosquito species (Diptera: Culicidae) including Aedes aegypti (L.), Anopheles gambiae s. l. Giles, and Culex quinquefasciatus Say by using contact-based technique. In screening bioassays, M. spicata I, M. spicata II, O. basilicum I, O. basilicum II, and A. indicum EOs showed higher repellency against Cx. quinquefasciatus as compared to Ae. aegypti and An. gambiae when tested at 33.3 μg/cm2. In time-span bioassays performed at 33.3 μg/cm2, EO of M. spicata I exhibited 100% repellence up to 45, 30, and 75 min against Ae. aegypti, An. gambiae, and Cx. quinquefasciatus, respectively. Interestingly, at this tested dose, M. spicata I and M. spicata II showed higher repellence compared to DEET against Ae. aegypti and Cx. quinquefasciatus after 45 and 75 min, respectively. Their repellency was observed up to 150 and 210 min against Ae. aegypti and Cx. quinquefasciatus, respectively. In larvicidal bioassays, M. spicata I EO proved more toxic against 2nd instar larvae of Ae. aegypti, An. gambiae, and Cx. quinquefasciatus (LC50 = 11.0, 42.9, and 12.6 mg/L, respectively) compared to other tested EOs. In oviposition bioassays, M. spicata I exhibited the highest activity, showing 60%, 46%, and 79% oviposition deterrence against Ae. aegypti, An. gambiae, and Cx. quinquefasciatus, respectively, tested at a dose of 600 µg/cm2. Major compounds of M. spicata I, M. spicata II, O. basilicum I, and O. basilicum II EOs were piperitenone oxide (38.8%), piperitone oxide (35.4%), estragole (55.3%), and linalool (43.8%), respectively. In conclusion, M. spicata EO could be used to control mosquitoes and their bites.
1 Introduction
Mosquito-borne diseases are widespread in tropical and subtropical regions of the world, ranging from asymptomatic to severe, and can even be fatal (1). Chikungunya, for example, can cause chronic joint pain that can last for years. Similarly, Zika infection has been associated with neurological disorders and fatal abnormalities during pregnancy (2). Malaria can have a devastating impact on the socioeconomic development of affected communities by lowering workforce productivity and raising healthcare costs. Children are especially susceptible to these vector-borne diseases. Filariasis affects over 120 million people worldwide, causing debilitating symptoms like elephantiasis (3). Every year, over one million people worldwide die as a result of mosquito-borne diseases (4, 5). Keeping in mind the harmful effects of diseases associated with mosquitoes, there is a need to control the population of mosquitoes and use personal protection means against their biting.
Synthetic insecticides such as temephos, deltamethrin, metofluthrin, acetamiprid, and cypermethrin effectively control mosquitoes (6–8). However, using these traditional insecticides can have negative consequences, including resurgence, resistance development, residual effects on the environment, and negative impacts on non-target organisms (9). Insecticides can harm humans, causing reproductive, carcinogenic, and endocrine problems (10, 11). Besides controlling mosquitoes through insecticides, personal protection is considered a suitable approach to prevent the bite of mosquitoes. N, N-diethyl-m-toluamide (DEET) and IR3535 are synthetic repellent compounds used against blood-sucking insects and effectively deter mosquitoes. However, continuous and excessive use of these synthetic repellents could harm human health, i.e., swelling, eye irritation and rashes (12–14). The alternative approach could be a way forward to combat mosquitoes and mosquito-borne diseases.
As a result, scientists have been focused on developing plant-based pest-control solutions. The products extracted from plants have proved effective in controlling insect pests for an extended period without harming the environment (15–17). Essential oils (EOs) derived from plants have been traditionally used in controlling insect pests (18, 19). These have a considerable share of the pesticide market, accounting for around $700 million with 45,000 tons of the world’s total pesticide output (20). EOs are being considered alternatives to synthetic insecticides in controlling mosquitoes due to their selective action in controlling target pests, as well as their minimal effects on non-target organisms and high environmental degradations (21). There are a few plant-based mosquito repellents available in the market. For example, the United States Environmental Protection Agency has approved Java citronella oil as a blood-sucking insect repellent (22). The p-menthane-3,8-diol (PMD) is a monoterpene, another plant-based natural product used as a mosquito repellent. It is a spent product of the distillation of leaves of the Australian lemon-scented gum tree, Corymbia citriodora (Hook.) (Myrtales: Myrtacea), commonly known by the synonym Eucalyptus citriodora. The U.S. Centers for Disease Control and Prevention (CDC) endorsed two non-DEET mosquito repellents, including PMD, in April 2005 (20, 23).
In our previous study, we screened seven EOs for repellent activity against Aedes aegypti L. (Diptera: Culicidae) mosquitoes and revealed that the EO of Mentha spicata L. (Lamiales: Lamiaceae) was the most efficient repellent (24). In the proposed study, two different chemotypes of M. spicata and Ocimum basilicum L (Lamiales: Lamiaceae) were used to test their bioactivity against three different mosquito species. Although several studies have documented the mosquito repellent and larvicidal properties of M. spicata and O. basilicum, there is a notable gap in the biological activities of their various chemotypes. While the carvone and piperitenone oxide chemotypes of M. spicata are extensively reported (25–30) the piperitone oxide chemotype of M. spicata is seldom mentioned. Besides M. spicata and O. basilicum, Abutilon indicum L. was also studied. To our best knowledge, only a few publications reported the larvicidal effect of EOs derived from Indian mallow Abutilon indicum (L.) (Malvales: Malavaceae) (31–33), while repellent and oviposition deterrent activities remain unknown. Moreover, to evaluate the potential use of a natural product, its efficiency and longevity of action are commonly compared with the most efficient positive control. Keeping in mind the importance of EOs in controlling mosquitoes, the current study was performed to comprehensively evaluate the most important bioactivities concerning mosquito control, i.e., repellent, larvicidal, and oviposition deterrent activities of selected EOs against three species of mosquitoes (Diptera: Culicidae): yellow fever mosquito, Ae. aegypti, Anopheles gambiae s. l., and southern house mosquito Culex quinquefasciatus Say.
2 Materials and methods
2.1 Collection and maintenance of plant material
The fresh aerial parts of selected plants were hand-picked in the flowering season. The samples of M. spicata and O. basilicum were collected from different locations in the district of Bhakkar, Pakistan whereas a sample of A. indicum was collected from Multan, Pakistan (Table 1). Some plants from each sample were isolated, placed in blotting sheets, and pressed using a standard process. Each plant species specimen was mounted on a standard herbarium sheet and deposited in the herbarium of the Department of Environmental Sciences, COMSATS University Islamabad, Abbottabad Campus, Abbottabad, Pakistan for record, and a voucher number was assigned (34). The identification of the plants was authenticated by a plant taxonomist at the Department of Environmental Sciences, COMSATS University Islamabad, Abbottabad Campus, Abbottabad, Pakistan. The plant samples to be used for EO extraction were gently washed with tap water and then rinsed with distilled water. The plant material was spread on a white cotton cloth in a shady area and air dried using a ceiling fan. All plant samples were processed in the same way but in separate rooms. The shade-dried plant material was stored in airtight poly bags at room temperature for about 2 weeks until used for EOs extraction.
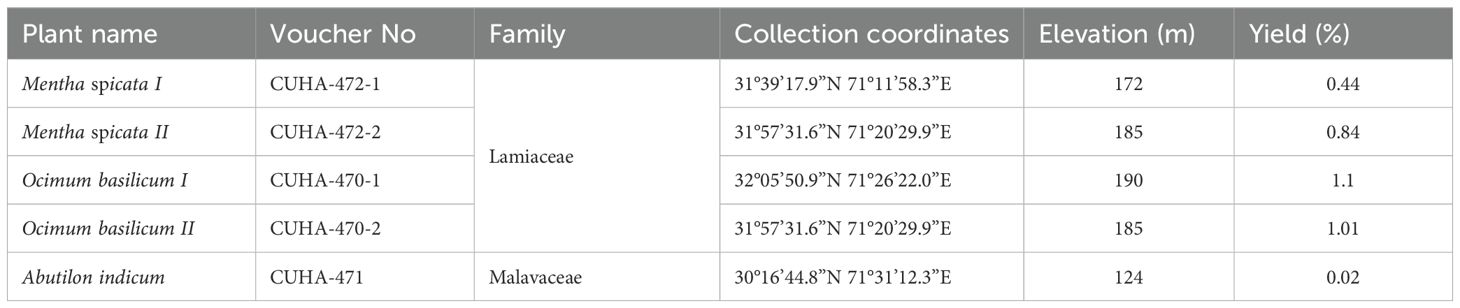
Table 1. Plant material used in the study and percentage yield of EOs.
2.2 Extraction of EOs
The steam distillation method was used to extract EOs from the collected plant material using a Clevenger-type apparatus, as described in the previous study (24). A stainless-steel vessel was loaded with plant material (300g), and two litres of distilled water were added to the bottom of the vessel. Water had no direct contact with the plant material. The distillation vessel was heated by using an electric hotplate. Volatile compounds released from the plant material and steam were cooled using a condenser fitted on the head of the vessel, and the distillate was collected in a separating funnel for three hours. The EO layer formed above the water layer was decanted and dried over anhydrous MgSO4. The extracted EO was weighed, and the percentage yield was calculated using the dry mass of the plant. The samples of EOs were stored at -20 °C until used for bioassays and chemical analysis.
2.3 Rearing of mosquitoes
Ae. aegypti, An. gambiae s. l., and Cx. quinquefasciatus mosquitoes were reared in the laboratory using methods described in previous studies (24, 35–38). The mosquitoes at the larval stage were obtained from the Punjab Health Department, Multan, Pakistan. Larvae were placed in a plastic container (20 × 16 × 4 cm) filled with 1 L water and fed with fish food (Osaka green fish food, India) containing 3% crude fat, 4% crude fibre, and 28% crude protein. Pupae were collected daily from the larval container and transferred to plastic cups containing 200 mL of tap water. The plastic cups were placed in Plexi-glass cages (30 × 30 × 30 cm) for the emergence of adults. Cotton soaked with 10% sucrose solution was placed in cages as an adult diet. After 4–5 days, females (Ae. aegypti and Cx. quinquefasciatus) were fed blood from a constrained pigeon placed in the adult cage while An. gambiae were fed on human arm blood. Wax paper was wrapped on the inner walls of the plastic jar, filled with water and placed in the adult cage for oviposition. After oviposition, the wax paper with eggs was transferred to the larval container with 1000 mL of tap water for hatching. The rearing process continued until enough adults and larvae were obtained for the repellence, oviposition and larvicidal bioassays, respectively. The rearing of three mosquito species was carried out in separate rooms. All rearing was carried out in a controlled room maintained at 25 ± 2°C for Ae. aegypti and Cx. quinquefasciatus while for An. gambiae room was maintained at 30 ± 2°C. Relative humidity was maintained at 80 ± 10% with a photoperiod of 12h:12h light:dark.
2.4 Mosquito repellency bioassay
A human bait technique was used to test the repellence potential of EOs against Ae. aegypti, An. gambiae s. l., and Cx. quinquefasciatus females (24, 35, 39). The positive control DEET (99% purity, Sigma-Aldrich, St. Louis, MO, USA) and EOs solutions, 1% (10 mg/mL) and 10% (100 mg/mL), were prepared by dissolving the respective substances in absolute ethanol (Daejung, Korea). Ethanol was used as a negative control in repellency bioassays. Twenty mated and blood-starved 4–5 days old female Ae. aegypti (strains 10) were released from the laboratory-reared colony in the experimental cage (30 × 30 × 30 cm) about 12 h before the start of the repellency bioassay. The hands of the subjects (2 volunteers) were washed with scent-free liquid soap and allowed to dry for about 10 min before starting the bioassay. Plastic gloves were used to cover the subject’s hand except for the 30 cm2 circular area on the dorsal side of the hand. A 100 μL aliquot solution of the test substance (1% or 10% equivalent to 33.3 µg/cm2 and 333 µg/cm2, respectively) or pure solvent as a negative control was evenly applied on the exposed area of the hand and dried in air for three min before exposing the hand to Ae. aegypti females. The subject’s hand was exposed to the females in the experimental cage, and mosquito landings were counted for 5 min. The experiment was repeated randomly five times for both the test samples and the negative control. The same procedure was followed to evaluate the repellency of EOs against An. gambiae and Cx. quinquefasciatus. The human volunteers were informed about the test procedure, and consent was obtained before conducting repellence bioassays. The repellency percentage was calculated using the formula: percentage repellency = [(Mc – Mt)/Mc] ×100, where Mc is the number of mosquito landings on the negative control (solvent) treated hand and Mt is the number of mosquito landings on the test substance treated hand. All volunteers followed standardised procedures to minimise the variability.
2.5 Time span bioassays
Plant EOs that showed at least 50% repellence were further investigated to determine their repellent longevity. Time-span repellent bioassays were performed by following the same protocol as mentioned above in the repellency bioassay, except for the exposure of the same treated hand to the females of Ae. aegypti, An. gambiae s. l., and Cx. quinquefasciatus for 5 min after each 15 min interval until the number of landings on control and treatment didn’t differ significantly. Time span bioassays were conducted using test samples at the dosages of 33.3 μg/cm2 and 333 μg/cm2. The experiments were repeated five times, and fresh female mosquitoes were employed for each replicate. A repellency bioassay for each mosquito species was conducted in separate climate-controlled rooms.
2.6 Larvicidal bioassays
Larvicidal bioassays against 2nd instar larvae of Ae. aegypti, An. gambiae, and Cx. quinquefasciatus were conducted through a modified protocol described in previous studies (38, 40). Briefly, ice cube trays (50 mL well size) were cleaned with tap water, and each well of an ice cube tray was filled with 20 mL of water. Fifteen 2nd instar larvae of Cx. quinquefasciatus, or Ae. aegypti, or An. gambiae were added through a pipet to each well of the ice cube tray. DMSO was used to dilute the tested EOs, and 50 µL of the resulting solution was added to each well, with a final concentration ranging from 6.25 mg/L to 1600 mg/L. The 2nd instar larvae of tested mosquitoes were exposed to EOs or DMSO for 24 and 48 hours to assess their susceptibility. A fish diet was used to feed larvae during the exposure time. The larvae that did not show movement after the exposure period were considered dead. DMSO was used as a negative control, and its concentration in test media never exceeded 0.25%. Chlorpyriphos was used as the positive control. Each experiment was repeated at least six times to ensure the reliability and reproducibility of the results.
2.7 Oviposition deterrence
The oviposition deterrence bioassay was conducted by adopting a method described by previous studies (38, 41). Briefly, sixty 5–7 days old and blood-fed female mosquitoes were released in a bioassay cage. Two plastic cups filled with 100 mL of distilled water were placed diagonally in the corners of the bioassay cage. One cup was a test treatment, while the other was a control. An aliquot of 600 µL of 1% or 10% ethanolic solution of an EO (w/v) was evenly sprayed on half of the wax paper strip (10 × 20 cm), air dried for 2 min, and then wrapped along the inner walls of the water-filled plastic cup in such a way that the EO treated area (10 × 10 cm) remained above the water level. The overall concentration on the treated wax paper was 60 µg/cm2 for 1% and 600 µg/cm2 for 10% ethanolic solution of an EO. In the control cup, the solvent-treated filter paper was wrapped in the same way described for the test cup. After applying the sample or solvent, the cups were left outside the cages for 5 min so that the solvent could evaporate before the commencement of the experiment. The control and sample-treated cups were left in the adult mosquito cage for 48 hours for oviposition. Afterwards, the eggs laid in each cup were counted. The positions of the control and test cups were changed randomly to avoid the position effects on oviposition. We conducted oviposition tests in each experiment, five times using a fresh mosquito population.
2.8 Chemical analysis of EOs
EOs that showed about 50% or higher repellence against all tested species of mosquitoes were analysed using a Hewlett Packard gas chromatograph connected to a mass spectrometer (GC–MS) by adopting the method described in earlier studies (24, 38). The GC had a 30 m capillary column (DB-5, Agilent Technologies Inc., Santa Clara, CA, USA) with a 0.25 mm internal diameter and a stationary phase film thickness of 0.25 µm. The GC injector temperature was maintained isothermally at 235°C throughout the sample analysis. The GC oven temperature was programmed as follows: initial temperature of 40°C for 2 min, then increased to 240°C at a rate of 4°C per min, and finally maintained at 240°C for 8 min. Helium was used as the mobile phase at a constant flow rate of 1 mL/min through the column. An aliquot of 1 µL of dilute EO solution was injected into the GC, and the injector was operated in a splitless mode for 30 sec. The MS was operated using the following parameters: an electron energy of 70 eV for ionisation, an ion source temperature of 180°C, and a mass spectrum scan range of 30–400 m/z. The total ion chromatogram was used to calculate the percent composition of compounds in EOs. A solution of a series of straight-chain alkanes (C9-C24) was injected into the GC-MS using the same parameters as the EOs analyses. The retention times of unknown compounds and alkanes were used to calculate the retention indices of separated compounds. Mass spectra and retention indices of separated compounds were initially compared to those available in the NIST-2008 MS library and webbook NIST online library to identify the separated compounds. Finally, identifications were verified by injecting available pure standard compounds purchased from Sigma–Aldrich (St. Louis, MO, USA) and Alfa Aesar (Haverhill, MA, USA).
2.9 Statistical analysis
To evaluate the statistical difference between the repellence and oviposition deterrent activity data of different EOs, one-way ANOVA was used, followed by the Tukey test at a significant threshold of alpha = 0.05 for pairwise comparisons of group means. The statistical analysis was performed on the Statistica 8.1 software version 14.0.1.25 (TIBCO Software Inc, Palo Alto, CA, USA). The observed larvicidal activities (LC50) were determined using the computer software SPSS 20 (IBM, USA). The lethal concentration estimates for tested essential oils were considered significantly different (p < 0.05) from the baseline essential oil if confidence limits for relative median potency ratios did not overlap with the value 1 (35, 38).
3 Results
3.1 Yield (%) of EOs
The aerial parts of O. basilicum I contained the highest amount of EO, yielding 1.10%, while the leaves of A. indicum contained the lowest amount of EO, i.e. 0.02% (Table 1).
3.2 Screening repellent bioassays
There were significant differences in the repellency of M. spicata I, M. spicata II, O. basilicum I, O. basilicum II, and A. indicum EOs against females of Ae. aegypti (df = 5, F = 1048, p < 0.001), An. gambiae (df = 5, F = 399, p < 0.001), and Cx. quinquefasciatus (df = 5, F = 1741, p < 0.001) at a tested dose of 33.3 µg/cm2 (Figure 1). DEET showed a similar repellency (100%) against Ae. aegypti, An. gambiae, and Cx. quinquefasciatus. Importantly, M. spicata I and M. spicata II EOs proved most effective and showed similar patterns of repellence against all tested mosquito species (Figure 1). M. spicata I and M. spicata II EOs displayed 100% repellency against Ae. aegypti and Cx. quinquefasciatus while against An. gambiae displayed 96% and 94% repellence, respectively. O. basilicum I and O. basilicum II EOs provided complete protection (100%) against Cx. quinquefasciatus. EO distilled from A. indicum revealed the least repellence against tested species of mosquitoes as compared to other tested EOs. Comparatively, more repellence of tested EOs was observed against Cx. quinquefasciatus compared to Ae. aegypti and An. gambiae (Figure 1).
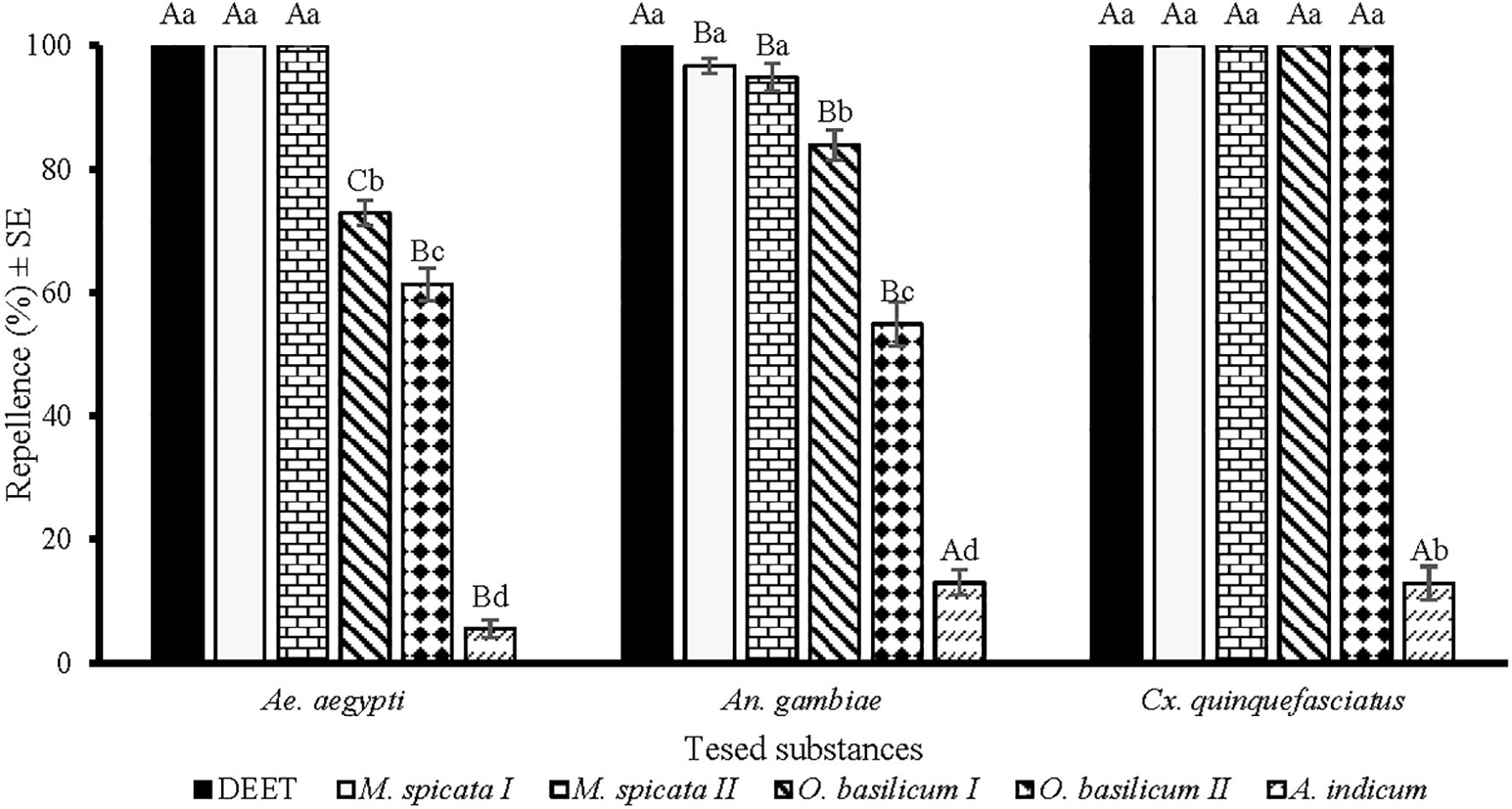
Figure 1. Repellency of positive control DEET and EOs at 33.3 µg/cm2 against Aedes aegypti, Anopheles gambiae s. l., and Culex quinquefasciatus mosquito females. Lower case letters above the columns represent significant differences (ANOVA post-hoc Tukey, p < 0.05) among tested substances against each mosquito species, while upper case letters represent the significant difference (ANOVA post-hoc Tukey, p < 0.05) among different species of mosquitoes towards a particular test substance. Error bars represent the standard error (n =5).
In screening bioassays, M. spicata I, M. spicata II, O. basilicum I, and O. basilicum II EOs showed more than 50% repellence against all tested species of mosquitoes and were further tested to evaluate the maximum period of repellency.
3.3 Time-span repellency of tested substances against Ae. aegypti
In time span bioassays, statistical data analysis revealed a significant impact of EOs repellency against Ae. aegypti at 33.3 µg/cm2 (df = 4, F = 254, p < 0.001). M. spicata I and M. spicata II EOs displayed 100% repellency against Ae. aegypti when tested immediately after application at a dose of 33.3 µg/cm2 (Figure 2A). However, their repellent potential decreased over time and reached 22% and 8%, respectively, at 45 min post-treatment. EOs of O. basilicum I and O. basilicum II showed 71% and 61% repellency, respectively, at 33.3 µg/cm2, and their repellency was observed only for up to 15 min (Figure 2A).
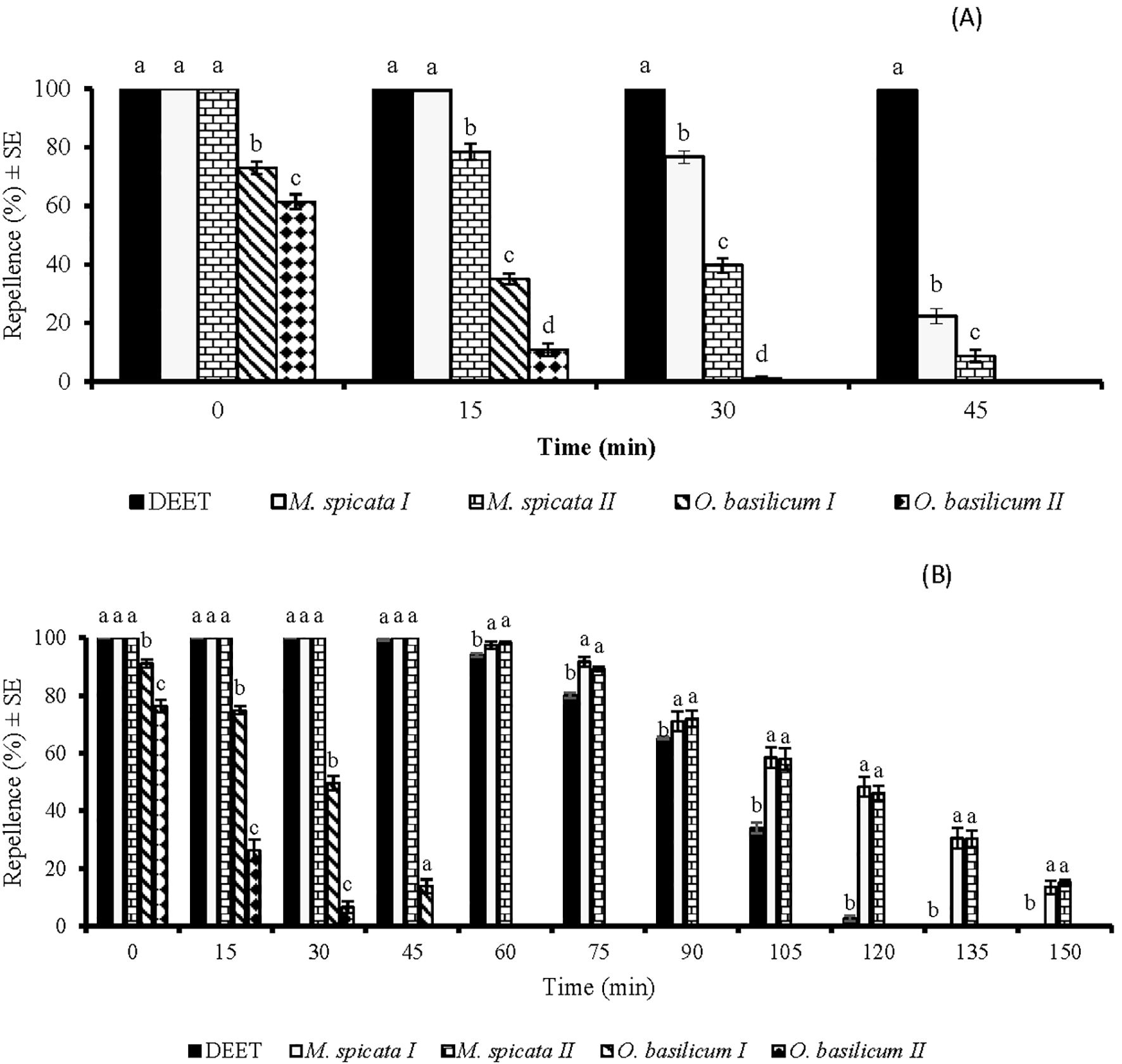
Figure 2. Time span repellent effect of DEET and four EOs against Aedes aegypti females at doses of 33.3 µg/cm2 (A) and 333 µ g/cm2 (B). Different letters indicate significant differences (p < 0.05) in repellency between different tested samples within each time interval, according to the ANOVA post-hoc Tukey test. “SE” stands for standard error (n = 5).
There was a significant difference (df = 4, F = 131, p < 0.001) in the repellent activities of EOs against Ae. aegypti tested at a dose of 333 µg/cm2. M. spicata I and M. spicata II EOs displayed complete protection against Ae. aegypti for up to 45 min (Figure 2B). Interestingly, these EOs exhibited higher repellency (p < 0.05) against Ae. aegypti at this tested dose compared to DEET after 45 min post-treatment, and their repellency was observed for up to 150 min post-treatment (Figure 2B). EOs of O. basilicum I and O. basilicum II showed 91% and 76% repellency, respectively, against Ae. aegypti at 333 µg/cm2, when applied immediately after application. However, both EOs showed repellence only for 30 and 45 min, respectively and later, no repellency was observed against Ae. aegypti (Figure 2B).
3.4 Time-span repellency of tested substances against An. gambiae
There was a significant difference in the repellency of tested EOs against An. gambiae at 33.3 µg/cm2 (df = 4, F = 113, p < 0.001) (Figure 3A). M. spicata I, M. spicata II EOs, and DEET showed 100% repellence against An. gambiae, when tested immediately after application of 33.3 µg/cm2 (Figure 3A). However, later repellency of M. spicata I and M. spicata II EOs decreased to 37% and 34%, respectively, at 30 min post-treatment. Comparatively, O. basilicum II EO showed the least repellence against An. gambiae at 33.3 µg/cm2 (Figure 3A).
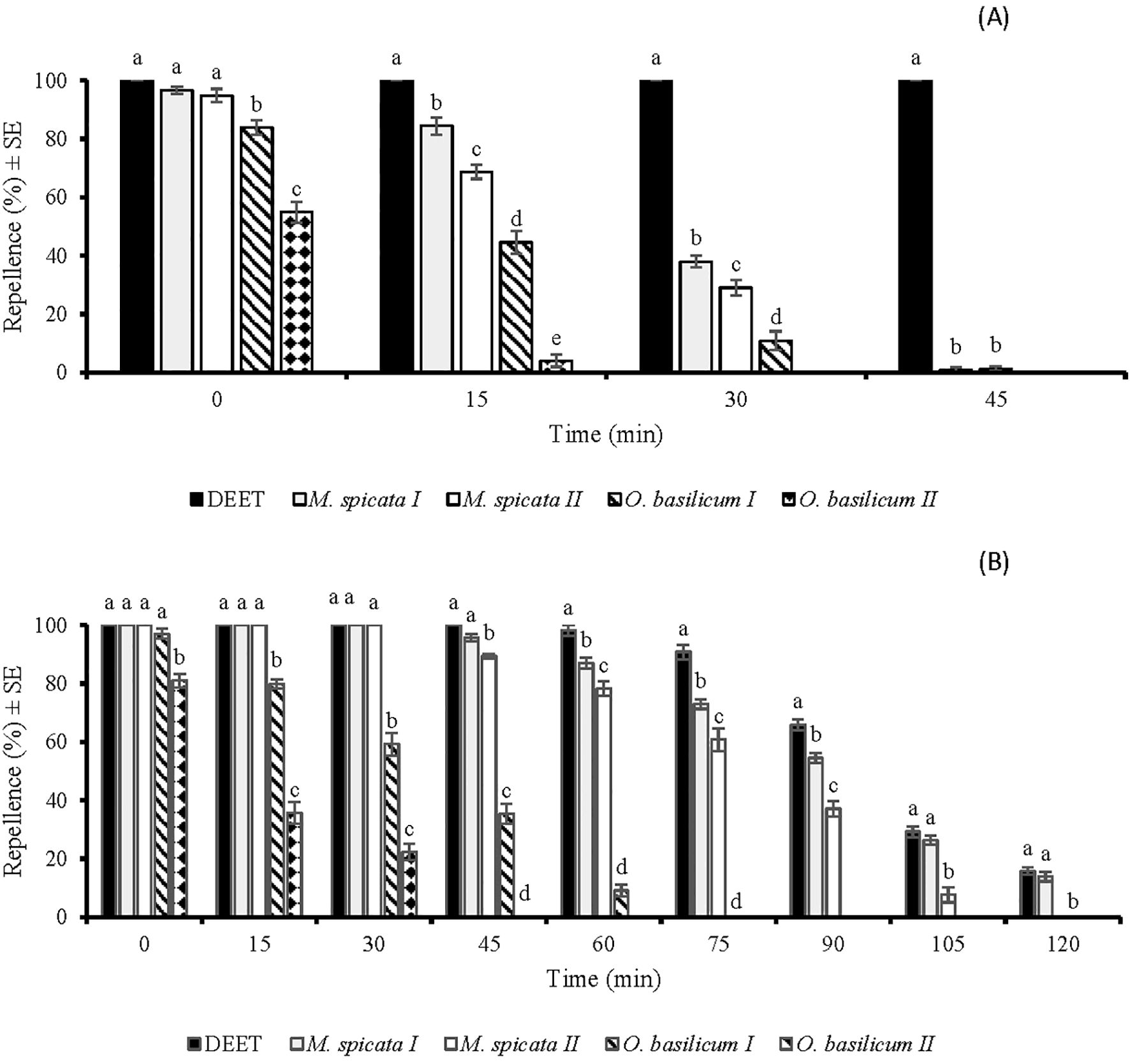
Figure 3. Time spans repellent effect of DEET and four EOs against Anopheles gambiae s. l. females at doses of 33.3 µg/cm2 (A) and 333 µg/cm2 (B). Different letters indicate significant differences (p < 0.05) in repellency between samples within each time interval according to the ANOVA post-hoc Tukey test. “SE” stands for standard error (n =5).
At a higher tested dose (333 µg/cm2), a significant difference (df = 4, F = 69, p < 0.001) was observed in the repellency of tested EOs against An. gambiae (Figure 3B). M. spicata I and M. spicata II showed complete protection similar to DEET against An. gambiae for up to 45 and 30 min, respectively. M. spicata I EO and positive control showed similar repellence against An. gambiae at 0, 15, 30, 45, 105, and 120 min post-treatment. O. basilicum I and O. basilicum II EOs exhibited an active time-span repellence against An. gambiae for up to 60 and 30 min, respectively (Figure 3B).
3.5 Time-span repellency of tested substances against Cx. quinquefasciatus
The repellency of all tested substances (EOs and DEET) was a significant difference (p < 0.05) at each tested time span except for the immediate post-treatment application of (p > 0.05). All the tested EOs showed 100% repellence against Cx. quinquefasciatus when tested immediately after application at a tested dose of 33.3 µg/cm2 (Figure 4A). The EOs of M. spicata I and M. spicata II exhibited complete protection (100%) against Cx. quinquefasciatus up to 60 and 35 min, respectively. Both these EOs showed repellence against Cx. quinquefasciatus up to 150 min and 135 min, respectively. O. basilicum I and O. basilicum II EOs showed 100% repellence against Cx. quinquefasciatus when observed immediately after application. However, at 30 min post-treatment, their repellency against Cx. quinquefasciatus decreased to 39% and 19%, and later, no repellency was observed (Figure 4A).
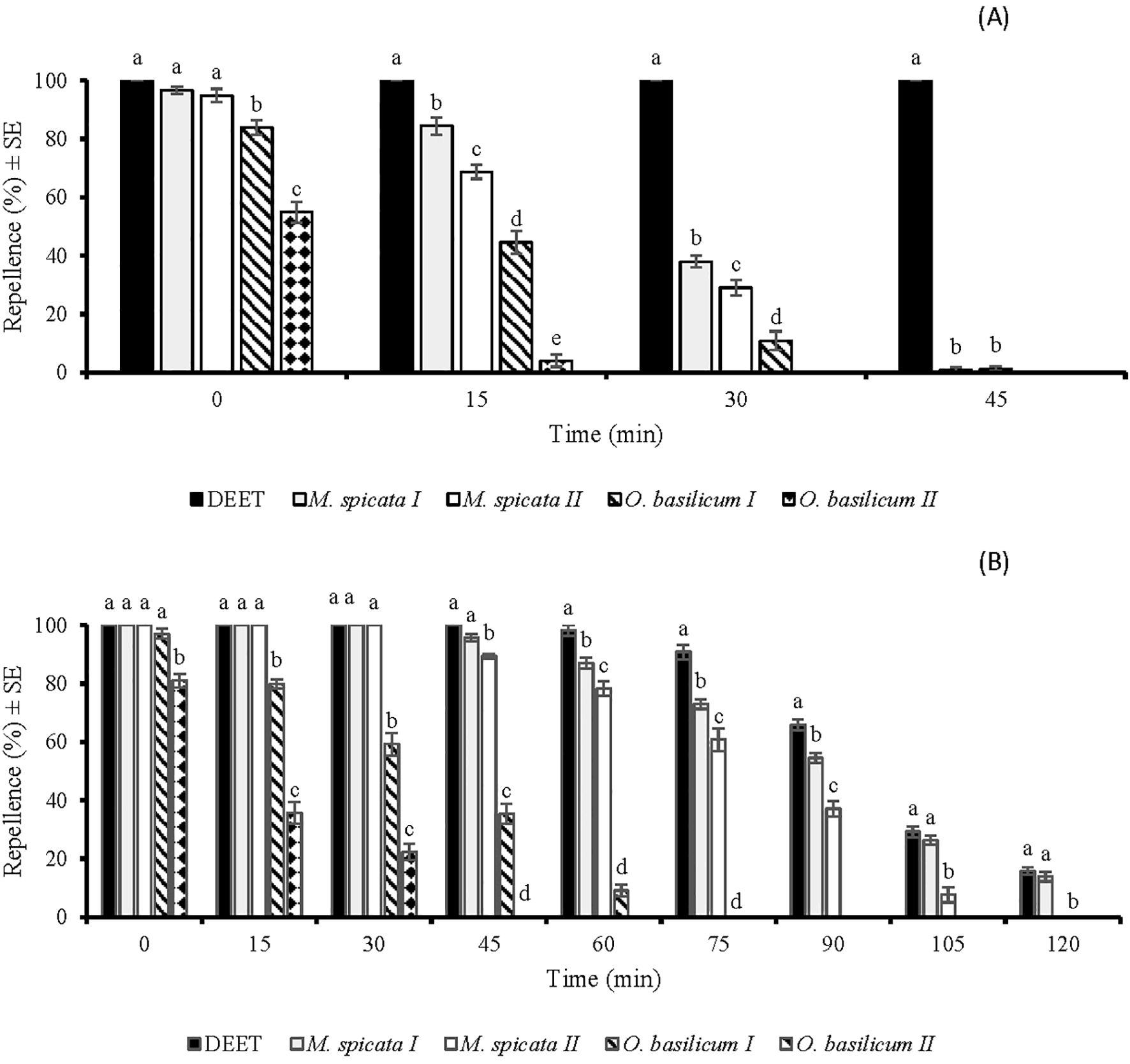
Figure 4. Time spans repellent effect of DEET and four EOs against Culex quinquefasciatus females at doses of 33.3 µg/cm2 (A) and 333 µg/cm2 (B). Different letters indicate significant differences (p < 0.05) in repellency between different substances within each time interval according to the ANOVA post-hoc Tukey test. “SE” stands for standard error (n =5).
The application of tested EOs at higher dose (333 µg/cm2) extended the active time span of M. spicata I, M. spicata II¸ O. basilicum I, O. basilicum II. DEET, M. spicata I, and M. spicata II EOs showed 100% repellency for 75 min, 120 min, and 105 min, respectively. Afterwards, the efficiency of these substances started to decrease at different rates (p < 0.05). Importantly, EOs of M. spicata I and M. spicata II showed more repellence against Cx. quinquefasciatus after 75 min post-treatment as compared to other tested substances. Repellent effects of M. spicata I and M. spicata II against Cx. quinquefasciatus were observed up to 210 min, while repellent effects of O. basilicum I and O. basilicum II EOs were observed up to 120 and 105 min respectively (Figure 4B).
3.6 Larvicidal effects of EOs
All the tested EOs showed larvicidal effects against 2nd instar larvae of Ae. aegypti, An. gambiae, and Cx. quinquefasciatus (Tables 2, 3). Positive control (chlorpyriphos) exhibited higher larvicidal effects (p < 0.05) against all tested species of mosquitoes as compared to all tested EOs. Larvae of Ae. aegypti proved significantly more susceptible to M. spicata I and O. basilicum I compared to M. spicata II, O. basilicum II, and A. indicum EOs at 24 and 48 h of exposure (Table 2, 3). There was no significant difference (p > 0.05) in the larvicidal effects of M. spicata I and O. basilicum I EOs against Ae. Aegypti thus showed LC50 values of 15.7 mg/L and 18.5 mg/L respectively after 24 h exposure. In the case of An. gambiae, M. spicata I showed high toxic effects (LC50 = 52.1 mg/L (ppm) at 24 h exposure and LC50 = 42.9 mg/L at 48 h) against larvae of An. gambiae while A. indicum showed the least toxic effects compared to other tested EOs. In the case of Cx. quinquefasciatus, the LC50 value of M. spicata I was 18.2 mg/L at 24 h exposure, which decreased to 12.6 mg/L after 48 h of larvae exposure. Overall, M. spicata I showed higher while A. indicum showed the least larvicidal effects against Ae. aegypti, An. gambiae, and Cx. quinquefasciatus compared to M. spicata II, O. basilicum I, and O. basilicum II at both tested time (Tables 2, 3).
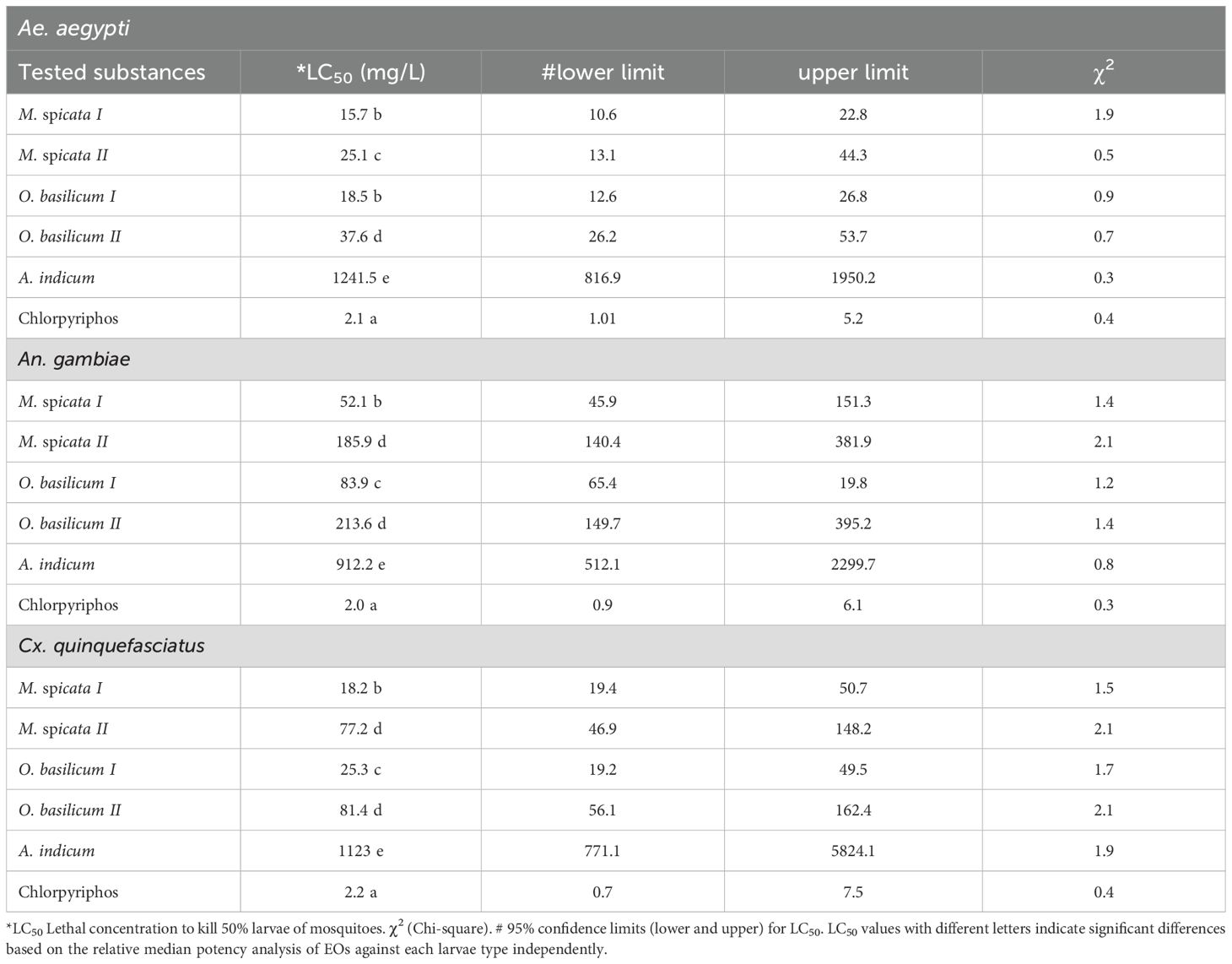
Table 2. Toxicity of tested substances against 2nd instar mosquito larvae after exposure of 24 h.
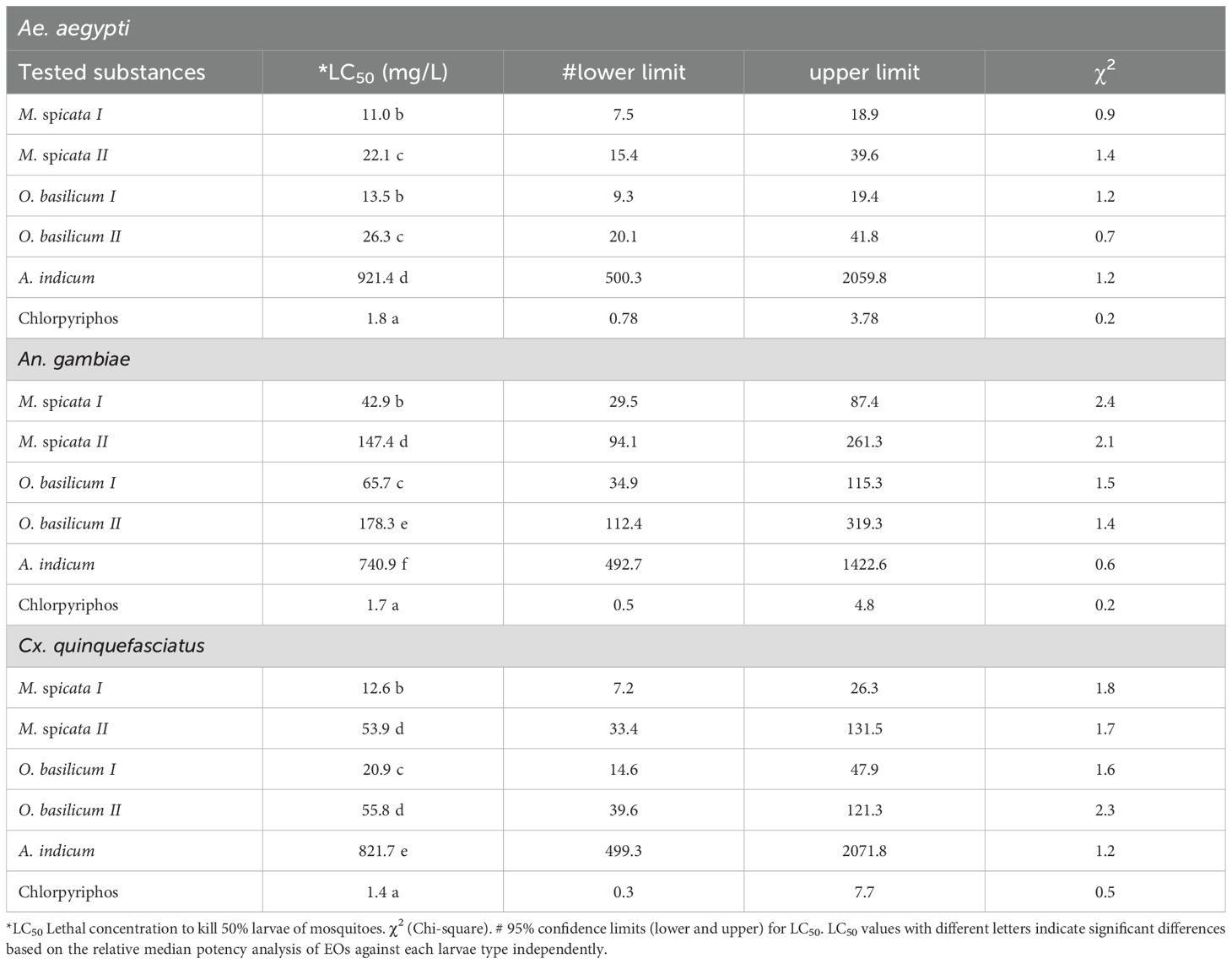
Table 3. Toxicity of tested substances against 2nd instar mosquito larvae after exposure of 48 h.
3.7 Oviposition deterrence
In the oviposition deterrence bioassay, a significant difference was observed between EOs against Ae. aegypti (df = 4, F = 69.5, p < 0.001), An. gambiae (df = 4, F = 20.9, p < 0.001), and Cx. quinquefasciatus (df = 4, F = 44.6, p < 0.001) at lower tested concentration of 60µg/cm2. At this concentration, higher oviposition deterrence (p < 0.05) was observed in the case of M. spicata I against all tested species of mosquitoes compared to other tested EOs. M. spicata I and A. indicum showed similar patterns of deterrence against Ae. aegypti, An. gambiae, and Cx. quinquefasciatus (Figure 5A). There was a significant difference in the oviposition deterrent effects of the tested EOs against Ae. aegypti (df = 4, F = 96.7, p < 0.001), An. gambiae (df = 4, F =69, p < 0.001), and Cx. quinquefasciatus (df = 4, F = 176, p < 0.001) at a tested concentration of 600 µg/cm2. EOs of M. spicata I and M. spicata II showed higher oviposition deterrence toward Ae. aegypti, An. gambiae, and Cx. quinquefasciatus as compared to the oviposition deterrent potential of O. basilicum I, O. basilicum II, and A. indicum EOs. Overall, greater oviposition deterrence of tested EOs was observed against Cx. quinquefasciatus as compared to Ae. aegypti and An. gambiae (Figure 5B).
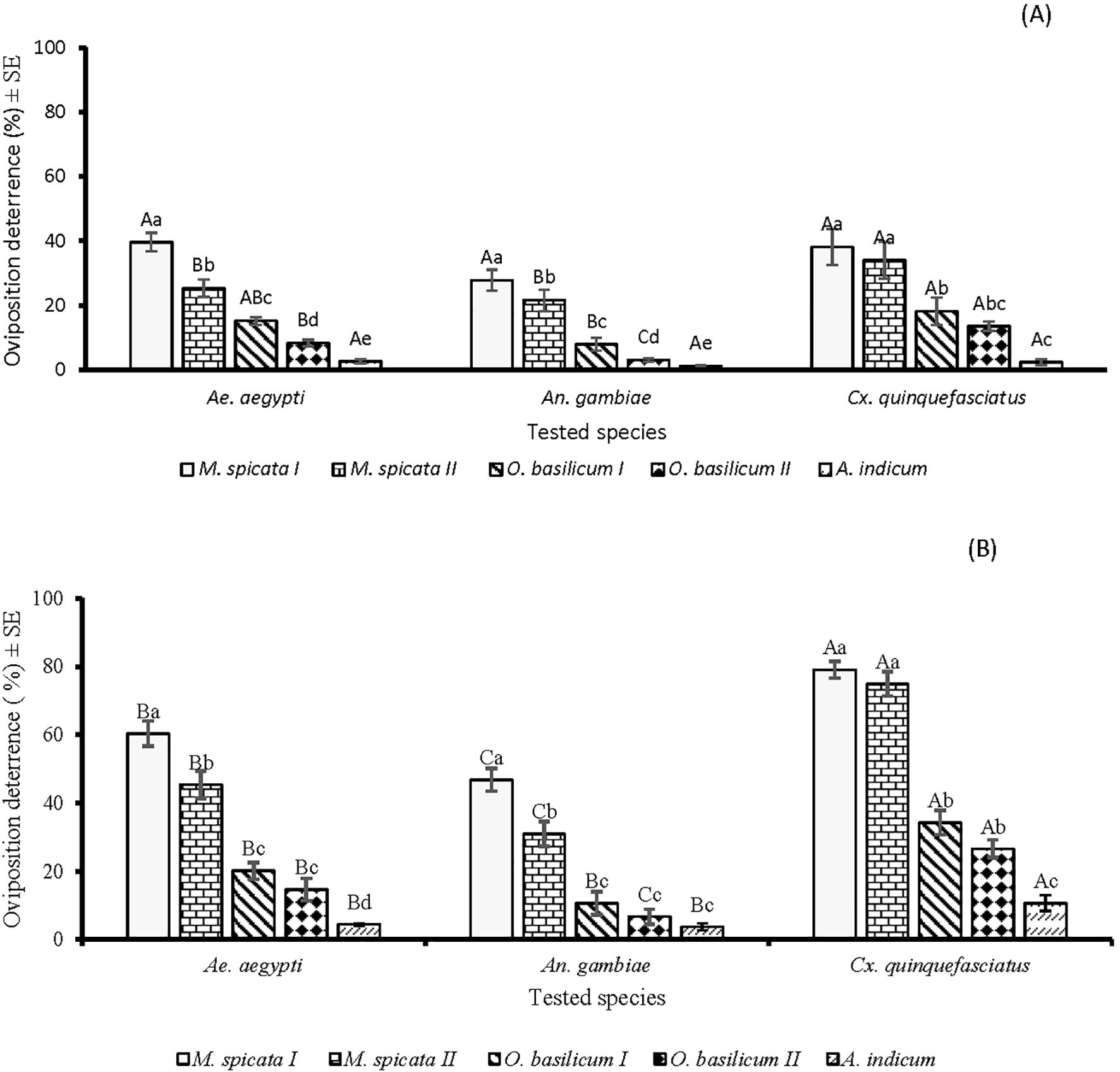
Figure 5. Oviposition behaviour modifying effectof five EOs at the tested doses of 60 µg/cm2 (A) and 600 µg/cm2 (B) against Aedes aegypti, Anopheles gambiae s.l., and Culex quinquefasciatus mosquitos. Small letters above the columns present the significant difference (p < 0.05) among tested EOs against mosquitoes, while capital letters present the significant difference (p < 0.05) among mosquitoes towards particular EO, according to ANOVA post-hoc Tukey test for each mosquito species and each EO separately. Error bars represent the standard error (n =5).
3.8 Chemical profile of EOs
The most abundant compounds in M. spicata I EO were piperitenone oxide (38.8%), piperitone oxide (25.6%), β-caryophyllene (6.3%), and limonene (4.1%). M. spicata II EO contained 35.4% piperitone oxide, 22.6% piperitenone oxide, and 7.6% limonene, accounting for approximately 64.92% of its composition (Table 4). Estragole (55.3%), eucalyptol (10.3%), linalool (10.2), and trans-α-bergamotene (9%), were the most abundant compounds in O. basilicum I, while linalool (43.8%), estragole (14.60%), and trans-α-bergamotene (9%) were the major compounds in O. basilicum II (Table 4).
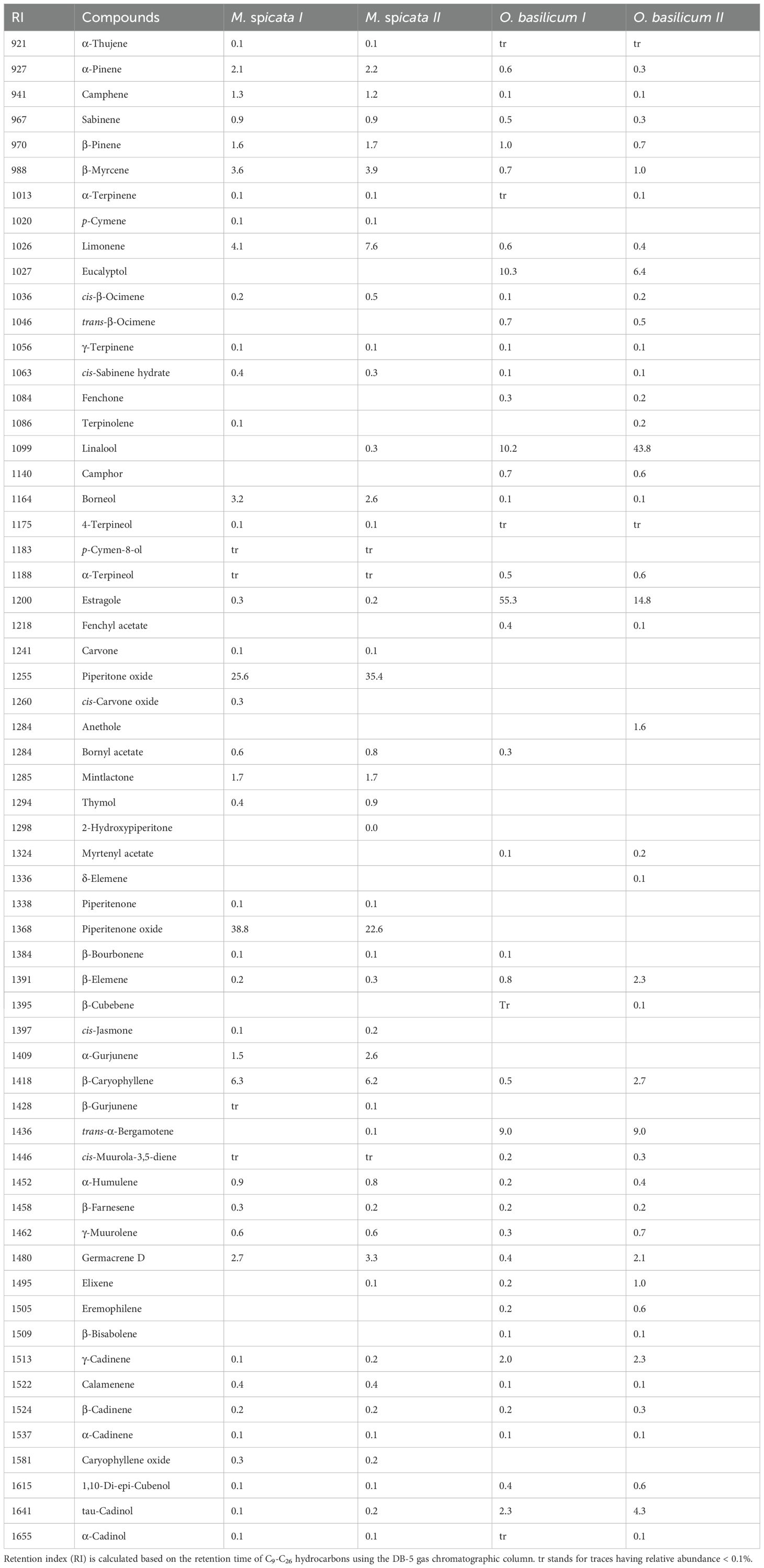
Table 4. Chemical profile of the EOs.
4 Discussion
Plant-based products like EOs are receiving more attention due to their potential in controlling blood sucking insects, mosquitoes and ticks etc. Here, we evaluated the repellent, larvicidal, and oviposition deterrent activities of five EO samples derived from two populations of M. spicata and O. basilicum as well as a population of A. indicum against Ae. aegypti, An. gambiae, and Cx. quinquefasciatus mosquitoes. Repellent results of the current study revealed that the tested EOs showed higher repellence against Cx. quinquefasciatus as compared to Ae. aegypti and An. gambiae. The difference in the behavioural response of different mosquito species towards the same test substance could be explained based on the presence of mismatched types of chemoreceptors in mosquito species (42). Our results are aligned with the previous studies where different levels of repellence of EOs were observed against other species of mosquitoes. For example, in a comparative study, Cx. quinquefasciatus was significantly more repelled compared to Ae. aegypti when exposed to the same EOs (42). In another study, menthol propylene glycol carbonate and DEET were significantly more repellent against Cx. quinquefasciatus as compared to An. gambiae (43). This could be due to differences in the sensitivity of olfactory receptors of different species to the chemical constituents present in the tested EOs (44).
Furthermore, the repellent activity of tested EOs against Ae. aegypti, An. gambiae, and Cx. quinquefasciatus was observed in a dose-dependent manner. At the higher tested dose 333 μg/cm2, all tested EOs showed greater and long-lasting repellence against Ae. aegypti, An. gambiae, and Cx. quinquefasciatus compared to the lower tested dose 33.3 μg/cm2. Our study results are aligned with the previous studies where the repellence of EOs has been documented as dose/concentration-dependent (35, 38, 45–47).
M. spicata I and M. spicata II EOs exhibited higher repellent effects against Ae. aegypti, Cx. quinquefasciatus, and An. gambiae as compared to the EOs of O. basilicum I, O. basilicum II, and A. indicum. However, in time-based bioassays, different repellent longevity of M. spicata I and M. spicata II was observed against Ae. aegypti, An. gambiae, and Cx. quinquefasciatus. The difference in repellence longevity of M. spicata I and M. spicata II might be due to their chemical compounds, particularly different proportions of piperitenone oxide and piperitone oxide. Previously, piperitenone oxide had displayed repellent efficacy against An. stephensi and Ae. albopictus (28, 30). In the study of Giatropoulos et al. (28) piperitenone oxide exhibited 95% repellency against Ae. albopictus at a tested dose of 40 μg/cm2. When the dose was increased to at 80 μg/cm2 and 200 μg/cm2 the repellency was 100%. Tripathi et al. demonstrated 100% repellency of piperitenone oxide against An. stephensi at the tested dose of 10.0 mg/mL (30). In our previous study, 45.5% piperitone oxide and 30% piperitenone oxide were the major components of the M. longifolia EO that showed 100% repellence against Ae. aegypti as compared to other tested EOs. Our current study and the previous studies showed that EOs containing a higher proportion of piperitenone oxide along with other compounds like piperitone oxide exhibited higher repellency compared to one where the relative proportion of piperitenone oxide was lower than piperitone oxide. Moreover, pure piperitenone oxide also exhibited lower repellency compared to the combined effect (28). Like in a study, though, the repellency of piperitone oxide against Ae. albopictus was moderate, its combined effect was significant in the case of EO, which contained 23% piperitone oxide and 41% piperitenone oxide (28). A study from India reported the presence of 32.4% piperitone oxide and 41.5% piperitenone oxide in Plectranthus incanus EO, which showed excellent repellency against An. stephensi and Cx. fatigans (48). Thus, the synergetic effects of different components of M. spicata I and M. spicata II EOs make it a potent repellent for Ae. aegypti, An. gambiae, and Cx. quinquefasciatus.
Previously, it has been proved that the bio-efficacy of EOs mainly depends on the type of chemical compounds present in them (49–51). In previous studies, EOs derived from M. spicata have possessed different levels of repellency against various species of mosquitoes. For example, EO of M. spicata possessed 73% repellence against Ochlerotatus caspius at a tested concentration of 20% (52). Likewise, Giatropoulos et al. from Greece reported 90% repellence of M. spicata EO against Ae. albopictus at a tested dose of 40 μg/cm2 (28). In the current study, even at a lower tested dose of 33.3 μg/cm2, greater repellence of M. spicata EO was observed against Ae. aegypti, An. gambiae and Cx. quinquefasciatus. The difference in the repellent effects of M. spicata EOs evaluated in the current and previous studies might be due to differences in the chemical composition of EOs or due to the presence of different chemoreceptors in different mosquitoes. Notably, the EOs derived from M. spicata I and M. spicata II provided long-lasting repellency at a higher tested dose 333 μg/cm2 against Cx. quinquefasciatus even longer than the positive control, i.e., DEET. The presence of less volatile compounds like piperitenone oxide, piperitone oxide and borneol in the EO of M. spicata might have been attributed towards the long-lasting effects of M. spicata I and M. spicata II EOs against the tested species of mosquito.
The EOs distilled from O. basilicum I, and O. basilicum II showed varying levels of repellency against Ae. aegypti, Cx. quinquefasciatus, and An. gambiae. A similar study was conducted by Baba et al., (53), where different levels of repellency of O. basilicum EO were observed against other species of mosquitoes (53). O. basilicum EO provided more repellence for a longer period (303 min) against An. gambiae as compared to Cx. quinquefasciatus (180 min) (53). In the current study, O. basilicum I EO showed up to 60 min repellence against An. gambiae while 105 min against Cx. quinquefasciatus at a tested dose of 333 μg/cm2. The difference in the repellency time of EO tested in the current study and Baba et al., (53) could be due to the different doses used, here we used 10% EO solution (333 μg/cm2), whereas Baba et al., (53) used pure EO having ten times higher concentration (53). Data showed the applied concentration/dose poses a significant effect on bioactivity. In the current study, the most abundant compounds of O. basilicum EO were estragole, eucalyptol, and linalool. These compounds, along with other compounds present in the EOs of O. basilicum I and O. basilicum II, might have contributed to the repellent activity of O. basilicum EOs against Ae. aegypti, An. gambiae, and Cx. quinquefasciatus.
In larvicidal bioassays, EOs showed varying levels of toxicity toward 2nd instar larvae of Ae. aegypti, Cx. quinquefasciatus, and An. gambiae. M. spicata I exhibited the highest larvicidal activity compared to all other tested EOs. The variation in the larvicidal effects of M. spicata I and M. spicata II might be due to the difference in the proportion of the same chemical compounds as piperitenone oxide and piperitone oxide. Previously, piperitenone oxide, which was an abundant compound (35.7%) in M. spicata EO showed prominent larvicidal effects against larvae of Cx. pipiens (LC50 9.95 ppm) (54). In another study, piperitenone oxide showed toxic effects against larvae of Ae. albopictus with an LC50 value of 162.2 ppm (28). In our previous study, EO having piperitone oxide (45.5%), piperitenone oxide (30.1%), and limonene (4.6%) as the most abundant compounds provided toxic effects against Ae. aegypti larvae (2nd instar) with an LC50 of 39.3 ppm (35). Limonene also has been shown toxic effects against An. stephensi, Ae. aegypti, and Cx. quinquefasciatus with the LC50 values of 8.83, 12.01, and 14.07 ppm, respectively (29). So, we can say that the toxic effects of M. spicata I and M. spicata II against tested species of mosquitoes might be due to the presence of these compounds like piperitenone oxide, piperitone oxide, and limonene, however, the contribution of other minor compounds can also be involved in the toxic effects of M. spicata I and M. spicata II.
O. basilicum I and O. basilicum II showed strong toxic effects against 2nd instar larvae of Ae. aegypti, An. gambiae, and Cx. quinquefasciatus. Our results are aligned with the previous studies where the EO of O. basilicum had been proven as a toxicant against various disease-carrying mosquitoes like Ae. aegypti (55), Cx. quinquefasciatus (56, 57), Cx. tritaeniorhynchus, An. subpictus, Ae. albopictus (37), An. stephensi (58), An. culicifacies (59), and An. stephensi (58). However, the toxic effect of O. basilicum EOs varied within the study as well as with previous studies. For example, in the current study, O. basilicum I and O. basilicum II showed different toxicity levels against Cx. quinquefasciatus with the LC50 value of 21 ppm and 61 ppm, respectively. Likewise in a previous study, EO of O. basilicum showed toxicity to Cx. quinquefasciatus larvae having an LC50 of 92 ppm (60). In another study, the EO of O. basilicum showed larvicidal activity against Cx. quinquefasciatus with the LC50 value of 68 ppm (61). The difference in larvicidal activity of O. basilicum EOs within the study and previous studies might be due to changes in the proportions of the same chemicals or different chemical compositions of O. basilicum EOs used in the current and previous studies. However, it is difficult to assess or compare results with other published data because of differences in plant sources, variation in chemical composition, percentage of secondary metabolites, extraction methods, collection times in different seasons and species of different mosquito genera (62–64). Larvicidal activity of O. basilicum EOs against Ae. aegypti, An. gambiae, and Cx. quinquefasciatus might be due to the presence of linalool and eucalyptol, which were present abundantly in the O. basilicum EO. Previously, both compounds have been proven to have toxic effects on the larvae of mosquitoes. For example, eucalyptol possessed 100% mortality in larvae of Ae. aegypti at a tested dose of 100 ppm (65). In another study, eucalyptol had toxic effects with an LC50 of > 200 ppm (28). Likewise, linalool displayed larvicidal activity against Ae. aegypti with an LC50 of 50 ppm (66). So, here we can conclude that chemical compounds present in EOs significantly effects the bioactivity of EO.
In current study, 2nd instar larvae were used to evaluate the larvicidal activity of the tested EOs over a 48 h exposure period. This approach was chosen to avoid pupation, which could occur with 3rd instar larvae during prolonged exposure. It is well-documented that younger larvae (2nd instar) are more susceptible to insecticides compared to older larvae (3rd or 4th instar) due to their thinner cuticle, smaller size, and underdeveloped detoxification mechanisms (67–69). For instance, Rajkumar et al. (67) demonstrated that 2nd instar larvae of Ae. aegypti exhibited significantly lower LC50 values when exposed to plant-derived compounds compared to 3rd and 4th instar larvae (67). Similarly, Chellappandian et al. (69) demonstrated that 2nd instar larvae of Ae. aegypti exhibited significantly lower LC50 values when exposed to chlorpyrifos, a standard insecticide, compared to 3rd and 4th instar larvae (69). While this higher susceptibility and the extended exposure period may limit direct comparability with studies using 3rd or 4th instar larvae and a 24-hour exposure (as per WHO protocols), our findings provide valuable insights into the efficacy of the tested oils against early larval stages, which are critical targets for mosquito control programs. Future studies could include parallel experiments with 3rd instar larvae and standardized exposure times to facilitate broader comparisons and further validate these results.
In oviposition deterrence bioassays, M. spicata I and M. spicata II EOs provided high oviposition deterrent activity against Ae. aegypti, An. gambiae, and Cx. quinquefasciatus as compared to the EOs of O. basilicum I, O. basilicum II, and A. indicum. Oviposition deterrent activity of M. spicata EO has been documented in previous studies against mosquitoes (30). Previously, M. spicata EO showed 47%, and 97% oviposition deterrence against An. stephensi at a tested dose of 30 µg/mL and 60 µg/mL, respectively (30). Chemical compounds of the M. spicata EO might be responsible for the oviposition deterrent activity against mosquitoes. Previously, piperitenone oxide, the most abundant compound of M. spicata EO, had displayed 77% and 100% oviposition deterrence against An. stephensi at 30 µg/mL and 60 µg/mL, respectively (30). Thus, the oviposition deterrent activity of the M. spicata EO against tested species of mosquitoes might be due to the presence of major and minor compounds in it.
EO of O. basilicum showed significantly different levels of oviposition deterrent activity against Cx. quinquefasciatus (30%), An. gambiae (20%), and Ae. aegypti (10%). Our results are aligned with the previous studies where different levels of oviposition deterrence were observed against various mosquitoes. For example, O. basilicum EO showed 95% oviposition deterrent at varying doses against An. stephensi (146 μg/mL), Ae. aegypti (211 μg/mL), and Cx. quinquefasciatus (215 μg/mL) (41). In another study, O. basilicum EOs displayed 95% oviposition deterrence against Ae. aegypti at the tested concentration of 10% (70). The difference in the oviposition deterrent activity of O. basilicum EO within the study and with respect to previous studies can be due to a change in the chemical composition of EOs.
The most abundant compounds in M. spicata I EO were piperitenone oxide (38.8%), piperitone oxide (25.6%), β-caryophyllene (6.3%), and limonene (4.1%). M. spicata II EO contained 35.4% piperitone oxide, 22.6% piperitenone oxide, and 7.6% limonene. In our previous published data, the major compounds of M. spicata EO were piperitenone oxide (47.0%), eucalyptol (12.0%), and borneol (9.5%) (24). A study by Koliopoulos et al. identified the major constituents in M. spicata as piperitenone oxide (35.7%) and 1,8-cineole (14.5%) (54). Several factors can influence the difference in the chemical composition of M. spicata EOs within the study and previous studies, including climate, altitude, soil type, growth conditions, agricultural methods and practices, plant part extracted, developmental stage, and harvesting time (71).
In the current study estragole (55.3%), eucalyptol (10.3%), linalool (10.2), and trans-α-bergamotene (9%) were the most abundant compounds in O. basilicum I, while linalool (43.8%), estragole (14.6%), and trans-α-bergamotene (9.0%) were the major compounds in O. basilicum II EO. Previously, linalool (56.7–60.6%) was the main constituent of O. basilicum EO, followed by epi-α-cadinol (8.6–11.4%), α-bergamotene (7.4–9.2%), γ-cadinene (3.3–5.4%) (72). Purkayastha et al. reported camphor, limonene and β-selinene as the major compounds of O. basilicum EO (73). The observed difference in the chemical composition of O. basilicum EOs within the current study and with respect to the previous studies might be due to the presence of different chemotypes, soil types and availability of nutrients to plants. Previously, O. basilicum EO exhibited a wide and varying array of chemical compounds, depending on variations in chemotypes, leaf and flower colours, aroma and origin of the plants (72, 74, 75).
The industrial application of essential oil-based formulations as mosquito repellents holds promise due to their natural origin and lower environmental impact. However, challenges such as formulation stability, volatility, and cost-effective large-scale production must be addressed. While essential oils offer a sustainable alternative to DEET, their commercial viability depends on optimizing extraction processes, enhancing longevity through encapsulation techniques, and ensuring affordability for widespread consumer adoption.
Conclusions
The use of EOs in controlling mosquitoes is expected to reduce the cost and harmful environmental effects of synthetic mosquito control measures. M. spicata I EO proved effective as a repellent, larvicidal, and oviposition deterrent against Ae. aegypti, An. gambiae, and Cx. quinquefasciatus. In the time span bioassay, M. spicata I EO showed significantly higher repellence than the “golden repellent standard” DDET against Ae. aegypti and Cx. quinquefasciatus; therefore, this EO can be considered a potential candidate for controlling mosquitoes. The limited repellent, larvicidal, and oviposition deterrent efficacy of A. indicum EO reveals no potential of this formulation for mosquito management. Further research should focus on optimizing the formulation of M. spicata I EO for field applications, including microencapsulation or emulsification techniques to enhance stability and longevity.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The studies involving humans were approved by Bahauddin Zakariya University, Multan. The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study. The manuscript presents research on animals that do not require ethical approval for their study.
Author contributions
MAb: Formal analysis, Methodology, Writing – original draft. MB: Conceptualization, Funding acquisition, Supervision, Writing – review & editing. MAz: Conceptualization, Funding acquisition, Supervision, Writing – review & editing, Formal analysis, Investigation, Methodology, Project administration, Visualization, Writing – original draft. SM: Formal analysis, Investigation, Writing – review & editing. ZS: Formal analysis, Investigation, Writing – review & editing, Methodology. AN: Formal analysis, Investigation, Methodology, Writing – review & editing. MS: Investigation, Methodology, Writing – review & editing. AP: Investigation, Methodology, Writing – review & editing. RM: Methodology, Writing – review & editing, Formal analysis, Funding acquisition, Visualization.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was funded by the International Foundation for Science (IFS), Sweden (Grant No. I-1-F-6041-1) to MA and by the Lithuanian state grant through State Scientific Research Institute Nature Research Centre, program 2 Climate and Eco-systems, Vilnius, Lithuania, available to RM.
Acknowledgments
We are very thankful to Abdullah Haris, Muhammad Sajjad, and Muhammad Umar Bashir for helping carry out the experiment. Ken Keefover-Ring, UW-Madison, USA, is highly acknowledged for proofreading this manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Benelli G and Duggan MF. Management of arthropod vector data - Social and ecological dynamics facing the One Health perspective. Acta tropica. (2018) 182:80–91. doi: 10.1016/j.actatropica.2018.02.015
2. Krauer F, Riesen M, Reveiz L, Oladapo OT, Martínez-Vega R, Porgo TV, et al. Zika virus infection as a cause of congenital brain abnormalities and guillain-barré Syndrome: systematic review. PloS medicine. (2017) 14:e1002203. doi: 10.1371/journal.pmed.1002203
3. Becker N, Petrić D, Zgomba M, Boase C, Madon M, Dahl C, et al. Subfamily anophelinae. In: Mosquitoes & Their control. New York, USA: Springer (2010). p. 163–86.
4. Terenius O, Marinotti O, Sieglaff D, and James AA. Molecular genetic manipulation of vector mosquitoes. Cell Host Micr. (2008) 4:417–23. doi: 10.1016/j.chom.2008.09.002
5. WHO. Dengue and severe dengue. Geneva, Switzerland: WHO (2020). Available at: https://www.who.int/news-room/fact-sheets/detail/dengue-and-severe-dengue (Accessed November 13, 2022).
6. Al Zahrani MR, Gharsan FN, Al-Ghamd KM, Mahyoub JA, and Alghamdi TS. Toxicity of two groups of pesticides against the mosquito Aedes aegypti. GSC Biol Pharmacol Sci. (2020) 13:148–55. doi: 10.30574/gscbps.2020.13.1.0334
7. Junkum A, Intirach J, Chansang A, Champakaew D, Chaithong U, Jitpakdi A, et al. Enhancement of temephos and deltamethrin toxicity by petroselinum crispum oil and its main constituents against Aedes aegypti (diptera: culicidae). Med Entomol. (2021) 58:1298–315. doi: 10.1093/jme/tjab008
8. Devine GJ, Vazquez-Prokopec GM, Bibiano-Marín W, Pavia-Ruz N, Che-Mendoza A, Medina-Barreiro A, et al. The entomological impact of passive metofluthrin emanators against indoor Aedes aegypti: A randomized field trial. PloS Neg Tropic Dis. (2021) 15:e0009036. doi: 10.1371/journal.pntd.0009036
9. Singhi M, Joshi V, Sharma R, and Sharma K. Ovipositioning behaviour of Aedes aegypti in different concentrations of latex of Calotropis procera: studies on refractory behaviour and its sustenance across gonotrophic cycles. Dengue Bull. (2004) 28:184–8.
10. Uwaifo F and John-Ohimai F. Dangers of organophosphate pesticide exposure to human health. Mat Sci Med. (2020) 4:27. doi: 10.4103/MTSM.MTSM_2_20
11. Pratiwi MAM and Purwati. The repellent activity test of rosemary leaf (Rosmarinus officinalis l) essential oil gel preparations influence on Aedes aegypti mosquito. J. Phys.: Conf. Ser. (2021) 1788, 012016. doi: 10.1088/1742-6596/1788/1/012016
12. Briassoulis G, Narlioglou M, and Hatzis T. Toxic encephalopathy associated with use of DEET insect repellents: a case analysis of its toxicity in children. Hum Exp Toxicol. (2001) 20:8–14. doi: 10.1191/096032701676731093
13. Fradin MS and Day JF. Comparative efficacy of insect repellents against mosquito bites. New Engl J Med. (2002) 347:13–8. doi: 10.1056/NEJMoa011699
14. Santhanam A, Miller MA, and Kasting GB. Absorption and evaporation of N, N-diethyl-m-toluamide from human skin in vitro. Toxicol app Pharmacol. (2005) 204:81–90. doi: 10.1016/j.taap.2004.08.016
15. Sukumar K, Perich MJ, and Boobar L. Botanical derivatives in mosquito control: a review. J Am Mosquito Cont Ass. (1991) 7:210–37.
16. Prabakar K and Jebanesan A. Larvicidal efficacy of some Cucurbitacious plant leaf extracts against Culex quinquefasciatus (Say). Biores Technol. (2004) 95:113–4. doi: 10.1016/j.biortech.2003.05.001
17. Hillary VE, Ceasar SA, and Ignacimuthu S. Efficacy of plant products in controlling disease vector mosquitoes, a review. Entomol Exp App. (2024) 172:195–214. doi: 10.1111/eea.v172.3
18. Arnason JT, Sims SR, and Scott IM. Natural products from plants as insecticides. Encyclopedia Life Support Syst (EOLSS). (2012) 1:1–8.
19. Sharifi-Rad J, Sureda A, Tenore GC, Daglia M, Sharifi-Rad M, Valussi M, et al. Biological activities of essential oils: From plant chemoecology to traditional healing systems. Molecules. (2017) 22:70. doi: 10.3390/molecules22010070
20. Tripathi AK, Shikha U, and Mantu B. A review on prospects of essential oils as biopesticide in insect-pest management. J Pharm Phytotherapy. (2009) 1:052–63.
21. Soleimani-Ahmadi M, Sanei-Dehkordi A, Turki H, Madani A, Abadi YS, Paksa A, et al. Phytochemical properties and insecticidal potential of volatile oils from Tanacetum persicum and Achillea kellalensis against two medically important mosquitoes. J Essl Oil Bearing Plants. (2017) 20:1254–65. doi: 10.1080/0972060X.2017.1388752
22. Sharma R, Rao R, Kumar S, Mahant S, and Khatkar S. Therapeutic potential of citronella essential oil: a review. Cur Drug Dis Technol. (2019) 16:330–9. doi: 10.2174/1570163815666180718095041
23. Carroll SP and Loye J. PMD, a registered botanical mosquito repellent with deet-like efficacy. J Am Mosquito Cont Ass. (2006) 22:507–14. doi: 10.2987/8756-971X(2006)22[507:PARBMR]2.0.CO;2
24. Azeem M, Zaman T, Tahir M, Haris A, Iqbal Z, Binyameen M, et al. Chemical composition and repellent activity of native plants essential oils against dengue mosquito, Aedes aegypti. Ind Crops Prod. (2019) 140:111609. doi: 10.1016/j.indcrop.2019.111609
25. Hussain AI, Anwar F, Shahid M, Ashraf M, and Przybylski R. Chemical composition, and antioxidant and antimicrobial activities of essential oil of spearmint (Mentha spicata L.) from Pakistan. J Essential Oil Res. (2010) 22:78–84. doi: 10.1080/10412905.2010.9700269
26. Chowdhury JU, Nandi NC, Uddin M, and Rahman M. Chemical constituents of essential oils from two types of spearmint (Mentha spicata L. and M. cardiaca L.) introduced in Bangladesh. Bangladesh J Sci Industrial Res. (2007) 42:79–82. doi: 10.3329/bjsir.v42i1.359
27. Saba I and Anwar F. Effect of harvesting regions on physico-chemical and biological attributes of supercritical fluid-extracted spearmint (Mentha spicata L.) leaves essential oil. J Essential Oil Bearing Plants. (2018) 21:400–19. doi: 10.1080/0972060X.2018.1458658
28. Giatropoulos A, Kimbaris A, Michaelakis A, Papachristos DP, Polissiou MG, and Emmanouel N. Chemical composition and assessment of larvicidal and repellent capacity of 14 Lamiaceae essential oils against Aedes albopictus. Parasitol Res. (2018) 117:1953–64. doi: 10.1007/s00436-018-5892-9
29. Govindarajan M, Sivakumar R, Rajeswari M, and Yogalakshmi K. Chemical composition and larvicidal activity of essential oil from Mentha spicata (Linn.) against three mosquito species. Parasitol Res. (2012) 110:2023–32. doi: 10.1007/s00436-011-2731-7
30. Tripathi AK, Prajapati V, Ahmad A, Aggarwal KK, and Khanuja SP. Piperitenone oxide as toxic, repellent, and reproduction retardant toward malarial vector Anopheles stephensi (Diptera: Anophelinae). J Med Entomol. (2004) 41:691–8. doi: 10.1603/0022-2585-41.4.691
31. Kovendan K, Murugan K, Panneerselvam C, Kumar PM, Amerasan D, Subramaniam J, et al. Laboratory and field evaluation of medicinal plant extracts against filarial vector, Culex quinquefasciatus Say (Diptera: Culicidae). Parasitol Res. (2012) 110:2105–15. doi: 10.1007/s00436-011-2736-2
32. Rahuman AA, Gopalakrishnan G, Venkatesan P, and Geetha K. Isolation and identification of mosquito larvicidal compound from Abutilon indicum (Linn.) Sweet. Parasitol Res. (2008) 102:981–8. doi: 10.1007/s00436-007-0864-5
33. Tariq RM, Naqvi SNH, Choudhary MI, and Abbas A. Importance and implementation of essential oil of Pakistanian Acorus calamus linn., as a biopesticide. Pak J Botany. (2010) 42:2043–50.
34. Sullivan JR and Nazaire M. Specimen collection and preparation for a changing flora. Rhodora. (2022) 123:50–66, 17. doi: 10.3119/20-32
35. Abbas MG, Haris A, Binyameen M, Nazir A, Mozūratis R, and Azeem M. Chemical composition, larvicidal and repellent activities of wild plant essential oils against Aedes aegypti. Biol. (2023) 12:8. doi: 10.3390/biology12010008
37. Govindarajan M, Sivakumar R, Rajeswary M, and Yogalakshmi K. Chemical composition and larvicidal activity of essential oil from Ocimum basilicum (L.) against Culex tritaeniorhynchus, Aedes albopictus and Anopheles subpictus (Diptera: Culicidae). Exp parasitol. (2013) 134:7–11. doi: 10.1016/j.exppara.2013.01.018
38. Parveen A, Abbas MG, Keefover-Ring K, Binyameen M, Mozūraitis R, and Azeem M. Chemical composition of essential oils from natural populations of Artemisia scoparia collected at different altitudes: antibacterial, mosquito repellent, and larvicidal effects. Molecules. (2024) 29:1359–67. doi: 10.3390/molecules29061359
39. Haris A, Azeem M, Abbas MG, Mumtaz M, Mozūratis R, and Binyameen M. Prolonged repellent activity of plant essential oils against dengue vector, Aedes aegypti. Molecules. (2023) 28:1351–3. doi: 10.3390/molecules28031351
40. Ali A, Wang Y-H, and Khan IA. Larvicidal and biting deterrent activity of essential oils of Curcuma longa, ar-turmerone, and curcuminoids against Aedes aegypti and Anopheles quadrimaculatus (Culicidae: Diptera). J Med Entomol. (2015) 52:979–86. doi: 10.1093/jme/tjv072
41. Prajapati V, Tripathi A, Aggarwal K, and Khanuja S. Insecticidal, repellent and oviposition-deterrent activity of selected essential oils against Anopheles stephensi, Aedes aegypti and Culex quinquefasciatus. Biores Technol. (2005) 96:1749–57. doi: 10.1016/j.biortech.2005.01.007
42. Amer A and Mehlhorn H. Repellency effect of forty-one essential oils against Aedes, Anopheles, and Culex mosquitoes. Parasitol Res. (2006) 99:478–90. doi: 10.1007/s00436-006-0184-1
43. Kweka EJ, Munga S, Mahande AM, Msangi S, Mazigo HD, Adrias AQ, et al. Protective efficacy of menthol propylene glycol carbonate compared to N, N-diethyl-methylbenzamide against mosquito bites in Northern Tanzania. Parasitol Vect. (2012) 5:1–10. doi: 10.1186/1756-3305-5-189
44. Afify A and Potter CJ. Insect repellents mediate species-specific olfactory behaviours in mosquitoes. Malaria J. (2020) 19:1–10. doi: 10.1186/s12936-020-03206-8
45. Azeem M, Zaman T, Abbasi AM, Abid M, Mozūratis R, Alwahibi MS, et al. Pesticidal potential of some wild plant essential oils against grain pests Tribolium castaneum (Herbst, 1797) and Aspergillus flavus (Link, 1809). Arabian J Chem. (2022) 15:103–9. doi: 10.1016/j.arabjc.2021.103482
46. Farina P and Conti B. Liabilities of essential oils as insect repellents. Curr Opin Environ Sci Health. (2024) 1:100–564. doi: 10.1016/j.coesh.2024.100564
47. Luker HA. A critical review of current laboratory methods used to evaluate mosquito repellents. Front Insect Sci. (2024) 4:132–8. doi: 10.3389/finsc.2024.1320138
48. Pal M, Kumar A, and Tewari KS. Chemical composition and mosquito repellent activity of the essential oil of Plectranthus incanus link. Facta universitatis-series: Phys Chem Technol. (2011) 9:57–64. doi: 10.2298/FUPCT1101057P
49. Auysawasdi N, Chuntranuluck S, Phasomkusolsil S, and Keeratinijakal V. Improving the effectiveness of three essential oils against Aedes aegypti (Linn.) and Anopheles dirus (Peyton and Harrison). Parasitol Res. (2016) 115:99–106. doi: 10.1007/s00436-015-4725-3
50. Herman A, Tambor K, and Herman A. Linalool affects the antimicrobial efficacy of essential oils. Cur Microbiol. (2016) 72:165–72. doi: 10.1007/s00284-015-0933-4
51. Angioni A, Barra A, Coroneo V, Dessi S, and Cabras P. Chemical composition, seasonal variability, and antifungal activity of Lavandula stoechas L. ssp. stoechas essential oils from stem/leaves and flowers. J Agric Food Chem. (2006) 54:4364–70. doi: 10.1021/jf0603329
52. Koc S, Oz E, and Cetin H. Repellent activities of some Labiatae plant essential oils against the saltmarsh mosquito Ochlerotatus caspius (Pallas, 1771)(Diptera: Culicidae). Parasitol Res. (2012) 110:2205–9. doi: 10.1007/s00436-011-2750-4
53. Baba G, Lawal A, and Shariff HB. Mosquito repellent activity and phytochemical characterization of essential oils from Striga hermonthica, Hyptis spicigera and Ocimum basilicum leaf extracts. Br J Pharmacol Toxicol. (2012) 3:43–8.
54. Koliopoulos G, Pitarokili D, Kioulos E, Michaelakis A, and Tzakou O. Chemical composition and larvicidal evaluation of Mentha, Salvia, and Melissa essential oils against the West Nile virus mosquito Culex pipiens. Parasitology Res. (2010) 107:327–35. doi: 10.1007/s00436-010-1865-3
55. Murugan K, Murugan P, and Noortheen A. Larvicidal and repellent potential of Albizzia amara Boivin and Ocimum basilicum Linn against dengue vector, Aedes aegypti (Insecta: Diptera: Culicidae). Biores Technol. (2007) 98:198–201. doi: 10.1016/j.biortech.2005.12.009
56. Rajamma A, Dubey S, Sateesha S, Tiwari S, and Ghosh S. Comparative larvicidal activity of different species of Ocimum against Culex quinquefasciatus. Natural prod Res. (2011) 25:1916–22. doi: 10.1080/14786419.2010.551755
57. Pushpanathan T, Jebanesan A, and Govindarajan M. Larvicidal, ovicidal and repellent activities of Cymbopogan citratus Stapf (Graminae) essential oil against the filarial mosquito Culex quinquefasciatus (Say)(Diptera: Culicidae). Trop Biomed. (2006) 23:208–12.
58. Murugan K, Aarthi N, Kovendan K, Panneerselvam C, Chandramohan B, Kumar PM, et al. Mosquitocidal and antiplasmodial activity of Senna occidentalis (Cassiae) and Ocimum basilicum (Lamiaceae) from Maruthamalai hills against Anopheles stephensi and Plasmodium falciparum. Parasitol Res. (2015) 114:3657–64. doi: 10.1007/s00436-015-4593-x
59. Pandey A, Pandey U, and Shrivastava P. Studies on mosquito larvicidal efficacy of indigenous plant extracts. J Ecophysiol Occup Health. (2010) 1:223–7.
60. Manzoor F, Samreen K, and Parveen Z. Larvicidal activity of essential oils against Aedes aegypti and Culex quinquefasciatus larvae (Diptera: Culicidae). J Anim Plant Sci. (2013) 23:420–4.
61. Benelli G, Pavela R, Giordani C, Casettari L, Curzi G, Cappellacci L, et al. Acute and sub-lethal toxicity of eight essential oils of commercial interest against the filariasis mosquito Culex quinquefasciatus and the housefly Musca domestica. Ind Crops prod. (2018) 112:668–80. doi: 10.1016/j.indcrop.2017.12.062
62. Knio K, Usta J, Dagher S, Zournajian H, and Kreydiyyeh S. Larvicidal activity of essential oils extracted from commonly used herbs in Lebanon against the seaside mosquito, Ochlerotatus caspius. Biores Technol. (2008) 99:763–8. doi: 10.1016/j.biortech.2007.01.026
63. Okumu FO, Knols BG, and Fillinger U. Larvicidal effects of a neem (Azadirachta indica) oil formulation on the malaria vector Anopheles Gambiae. Malaria J. (2007) 6:1–8. doi: 10.1186/1475-2875-6-63
64. Lima MA, de Oliveira FFM, Gomes GA, Lavor PL, Nagao-Dias AT, Lemos TL, et al. Evaluation of larvicidal activity of the essential oils of plants species from Brazil against Aedes aegypti (Diptera: Culicidae). Afr J Biotechnol. (2011) 10:11716–20.
65. Araújo EC, Silveira ER, Lima MAS, Neto MA, de Andrade IL, Lima MAA, et al. Insecticidal activity and chemical composition of volatile oils from Hyptis martiusii Benth. J Agric Food Chem. (2003) 51:3760–2. doi: 10.1021/jf021074s
66. Cheng S-S, Liu J-Y, Tsai K-H, Chen W-J, and Chang S-T. Chemical composition and mosquito larvicidal activity of essential oils from leaves of different Cinnamomum osmophloeum provenances. J Agric Food Chem. (2004) 52:4395–400. doi: 10.1021/jf0497152
67. Rajkumar S, Jebanesan A, and Nagarajan R. Effect of leaf essential oil of Coccinia indica on egg hatchability and different larval instars of malarial mosquito Anopheles stephensi. Asian Pacific J Trop Med. (2011) 4:948–51. doi: 10.1016/S1995-7645(11)60224-1
68. World Health Organization. Guidelines for laboratory and field testing of mosquito larvicides. Geneva: World Health Organization (2005).
69. Chellappandian M, Vasantha-Srinivasan P, Senthil-Nathan S, Karthi S, Thanigaivel A, Ponsankar A, et al. Botanical essential oils and uses as mosquitocides and repellents against dengue. Environ Int. (2018) 113:214–30. doi: 10.1016/j.envint.2017.12.038
70. Warikoo R, Wahab N, and Kumar S. Oviposition-altering and ovicidal potentials of five essential oils against female adults of the dengue vector, Aedes aegypti L. Parasitol Res. (2011) 109:1125–31. doi: 10.1007/s00436-011-2355-y
71. Moghaddam M and Mehdizadeh L. Chemistry of essential oils and factors influencing their constituents. Soft Chem Food Ferment. (2017) p:379–419. doi: 10.1016/B978-0-12-811412-4.00013-8
72. Hussain AI, Anwar F, Sherazi STH, and Przybylski R. Chemical composition, antioxidant and antimicrobial activities of basil (Ocimum basilicum) essential oils depends on seasonal variations. Food Chem. (2008) 108:986–95. doi: 10.1016/j.foodchem.2007.12.010
73. Purkayastha J and Nath SC. Composition of the camphor-rich essential oil of Ocimum basilicum L. native to Northeast India. J Ess Oil Res. (2006) 18:332–4. doi: 10.1080/10412905.2006.9699104
74. Sajjadi SE. Analysis of the essential oils of two cultivated basil (Ocimum basilicum L.) from Iran. DARU J. Pharm. Sci. (2006) 14(3):128–30.
Keywords: repellence, chemical constituents, mentha spicata, control strategies, eco-friendly, gas chromatography-mass spectrometry, bioactive compounds
Citation: Abbas MG, Binyameen M, Azeem M, Majeed S, Sarwar ZM, Nazir A, Sharif MMI, Parveen A and Mozūratis R (2025) Chemical analysis, repellent, larvicidal, and oviposition deterrent activities of plant essential oils against Aedes aegypti, Anopheles gambiae, and Culex quinquefasciatus. Front. Insect Sci. 5:1582669. doi: 10.3389/finsc.2025.1582669
Received: 24 February 2025; Accepted: 22 April 2025;
Published: 15 May 2025.
Edited by:
Maristella Mastore, University of Insubria, ItalyReviewed by:
Raimundo Nonato Souto, Universidade Federal do Amapá, BrazilPathalam Ganesan, St. Xaviers College, India
Pablo Francisco Martina, Institute of Subtropical Biology - CONICET, Argentina
Copyright © 2025 Abbas, Binyameen, Azeem, Majeed, Sarwar, Nazir, Sharif, Parveen and Mozūratis. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Muhammad Azeem, bXVoYXplZW1AY3VpYXRkLmVkdS5waw==; Raimondas Mozūratis, cmFpbW9uZGFzLm1venVyYWl0aXNAc3Uuc2U=
†These authors have contributed equally to this work