 Cláudia Gomes
Cláudia Gomes Lizeth Chicas
Lizeth Chicas Kelly A. Crotty
Kelly A. Crotty Ana Rodriguez
Ana Rodriguez- Department of Microbiology, New York University School of Medicine, New York, NY, United States
Cerebral malaria (CM) is a complication of infection with Plasmodium falciparum that can lead to cognitive sequelae and death. The diagnosis of CM is based on clinical criteria, which leads to frequent misdiagnosis as it is confused with other infections that induce coma in children. There is currently no possibility of early diagnosis of this complication, since CM is only identified after the presentation of neurological signs, which greatly decreases treatment success and also precludes the analysis of patient’s early samples for the identification of predictive/prognostic biomarkers. Here we have used the mouse model for CM (infection with Plasmodium berghei-ANKA) and compared it to a non-CM model (infection with P. berghei-NK65) to evaluate the early kinetics and specificity of two candidate biomarkers that are elevated in the plasma of patients with CM: Angiopoietin-2 and Angiopoietin-like 4. The mouse experimental CM (ECM) model allows for the study of the biomarker’s kinetics throughout infection, starting before neurological signs are evident, and for their specificity for ECM as compared to the non-cerebral model. Our results indicate that, similar to findings in P. falciparum malaria patients, Angiopoietin-2 and Angiopoietin-like-4 are significantly elevated in plasma during P. berghei infection in mice. In mice infected with P. berghei-NK65 there was a direct correlation with the levels of parasitemia, suggesting that this may be contributing to the increased levels of both candidate biomarkers during infection, however this was not observed in P. berghei-ANKA infected mice. In these mice, a high proportion developed ECM and showed elevated levels of Angiopoietin-like 4, which were not observed in mice with non-cerebral infections. Angiopoietin-like 4 levels were directly correlated with severity of ECM. This observation is similar to previous findings in human malaria patients and provide basis for the use of mice as a model to investigate early kinetics and specificity of potential biomarkers for human severe and cerebral malaria.
Introduction
Malaria is still the cause of approximately 600,000 death per year, mostly in children under the age of 5 (WHO, 2024). A high proportion of these deaths are caused by cerebral malaria (CM), a severe complication of Plasmodium falciparum infection that results in death in ~18% of the cases despite administration of anti-malarial medications (Guinovart et al., 2022), with 25% of survivors presenting important permanent neurological sequelae (John et al., 2008).
CM is characterized by the sequestration of P. falciparum-infected erythrocytes in the brain microvasculature, loss of blood-brain barrier integrity and brain swelling that may lead to death (Seydel et al., 2015). A major problem in the management of CM is that the diagnosis is frequently inaccurate, with both high rates of false positive (Taylor et al., 2004) and false negative (Berkley et al., 1999; Aipit et al., 2014) diagnosis.
In the absence of specific pharmacological therapy for CM, early diagnosis would be highly beneficial to improve the rate of success of general anti-malarial treatments. However, currently it is not possible to identify which children are at risk for developing CM among the very large numbers of children with uncomplicated malaria, since the initial presentation is frequently mild and non-specific (WHO, 2013). As a result, over 50% of children with severe malaria die at the community level without access to adequate healthcare and potentially lifesaving treatment (Rutherford et al., 2009). Therefore, there is an urgent need to identify biomarkers for the diagnosis of CM that can help clinicians identify this complication early and accurately in the course of infection. Several plasma molecules, with Angiopoietin-2 (ANG2) as the most solid candidate (Conroy et al., 2012), have been proposed as biomarkers for CM and have been used in clinical trials, but still do not meet the requirements for adequate sensitivity, specificity, and/or reproducibility between different cohorts (Muppidi et al., 2023).
In an attempt to identify biomarkers specific for CM that would not be triggered by the general inflammatory response to infections, but specifically in response to malaria, we previously performed a search for proteins secreted by human brain endothelial cells exposed to P. falciparum in vitro (Zuniga et al., 2022). This search identified a protein, Angiopoietin-like 4 (ANGPTL4), that is secreted by endothelial cells in response to P. falciparum and was found to be elevated in a cohort of African children with CM, compared to other malaria severe complications (Gomes et al., 2023). However, analysis of human CM samples provides limited information regarding the evolution of the biomarker levels over course of the disease, as samples are collected later in infection, when a small percentage of children with malaria start to show neurological signs.
Here, we have compared the experimental cerebral malaria (ECM) mouse model to a non-cerebral P. berghei infection (Ghazanfari et al., 2018) to perform longitudinal and specificity assessments of the two human CM candidate biomarkers, ANG2 and ANGPTL4, with the aim to characterize the kinetics and specificity of these biomarkers.
Material and methods
Mice
All procedures involving animals followed the Guide for the Care and Use of Laboratory Animals published by US National Institutes of Health (NIH Publication No. 85-23, revised 1985) and were performed with the approval of the Animal Care and Use Committee at New York University School of Medicine (Protocol IA16-00626).
Infections
C57BL/6J mice (4-6 weeks old) were obtained from Taconic Farms Inc. All animals were maintained under barrier conditions and had free access to water and normal laboratory diet. Mice were infected by i.p. inoculation of 3.5 x 105 P. berghei-ANKA (Franke-Fayard et al., 2004) or P. berghei-NK65 (Alger et al., 1971) infected erythrocytes. P. berghei lines were not cloned.
Scoring of CM
Animals were closely monitored to assess signs of CM following the score chart described by Waknine-Grinberg JH et al (Waknine-Grinberg et al., 2010) with modifications. All mice were scored daily after infection. Score of CM was based on appearance (Normal = 0; Coat ruffled = 1; Coat staring/panting = 2) and behavior (Normal = 0; Hunched or wobbly gait = 1; Partial paralysis or immobile = 2; Convulsions or coma = 3). Accumulated scores of 3 were considered severe cases and scores 4-5 critical cases of CM. Mice were sacrificed when reaching a cumulative score ≥ 4 or if they are immobile in response to stimulation or comatose.
Sample collection and determination of parasitemia
Blood samples of 12 μL were obtained by vein tail puncture at the indicated times after infection. Blood was collected with heparinized micro-hematocrit capillary tubes (Fisher Scientific). Parasitemias were determine by microscopic quantification of blood smears stained with Giemsa (Sigma-Aldrich).
ELISA determinations of ANG2 and ANGPTL4
Plasma concentrations of ANG2 and ANGPTL4 were measured (at dilutions of 1/40, and 1/100, respectively) using commercial ELISAs (R&D systems, Abcam, respectively) following manufacturer’s instructions. Plasma concentrations were determined using a standard curve for each of the recombinant proteins.
Statistical methods
Statistical analysis were performed with GraphPad Prism v10.4 software. The Normality Kolmogorov-Smirnov test was used to determine whether the analyzed variables follow a normal distribution. Unpaired samples t-tests were used to compare the levels of biomarkers in infected vs uninfected mice. Paired samples t-tests were carried out to compare the levels of each biomarker in each mouse between day 2 and days 4 and 6 after infection. One-way ANOVA was used to compare the levels of biomarkers between the uninfected, infected with P. berghei-NK65 and P. berghei-ANKA.
Results
The levels of the candidate human CM biomarkers ANG2 and ANGPTL4 were determined in the plasma of C57BL/6J mice to study the kinetics and specificity of each biomarker.
A first analysis was performed to determine whether the biomarkers could differentiate infected from uninfected mice, by comparing their levels in the plasma of control and infected mice (including both P. berghei-ANKA and P. berghei-NK65). We observed that the levels of ANGP2 and ANGPTL4 are significantly increased in infected mice (Figure 1), similarly to human malaria patients (Gomes et al., 2023).
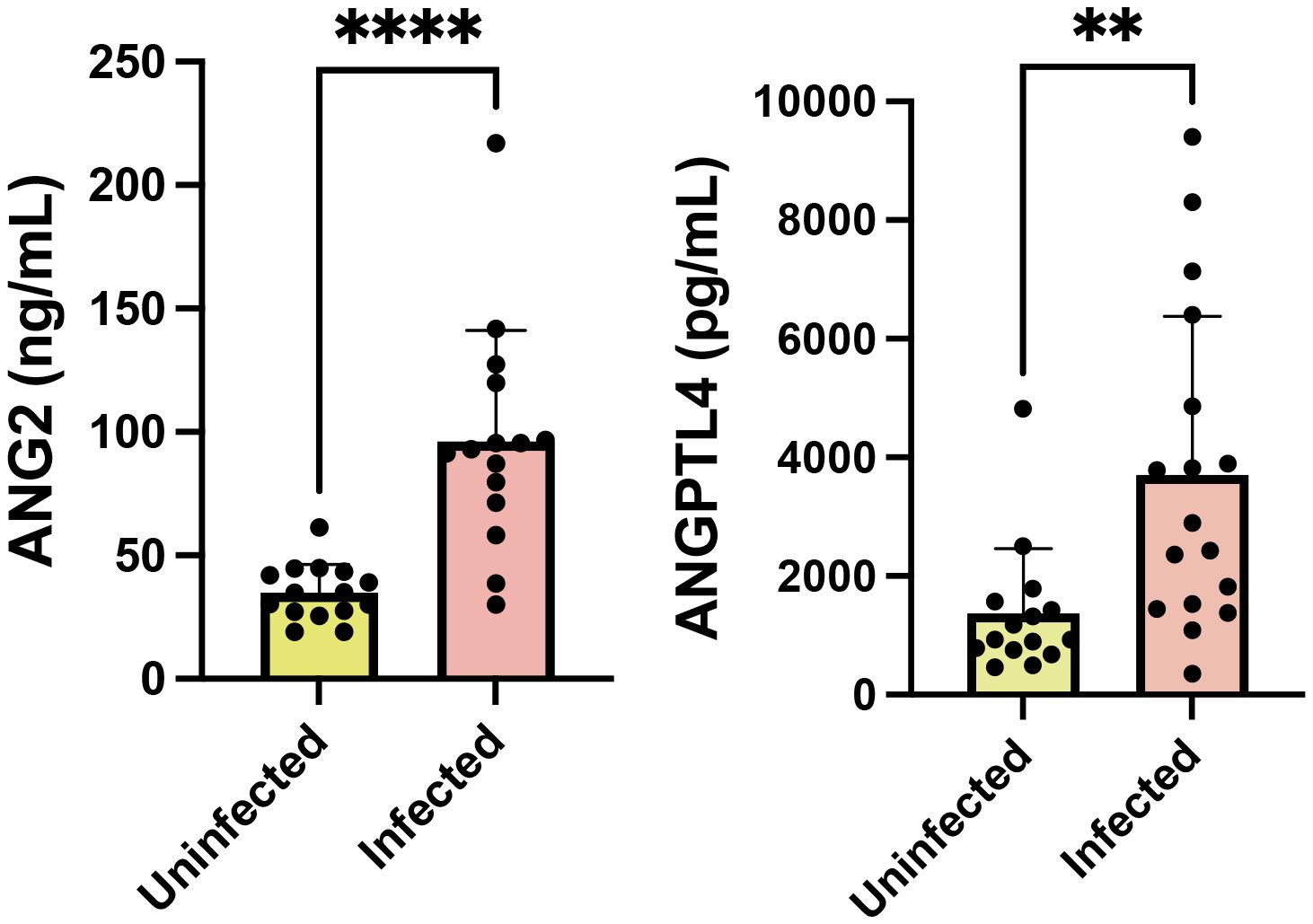
Figure 1. ANGP2 and ANGPTL4 are increased in mice infected with P. berghei. Levels of ANG2 and ANGPTL4 were determined in plasma of groups of C57/B6 mice uninfected (n=15) or infected with P. berghei at day 6 after infection (n=17). Bars show mean and standard deviation values for each group. p values determined by t-test. ****p<0.0001; **p<0.01.
Since the two biomarkers in this study were previously shown to differentiate CM from other forms of severe malaria in children (Conroy et al., 2009; Gomes et al., 2023), here we compared their levels in C57BL/6J mice infected with P. berghei-ANKA, which induces ECM with severe neurological signs upon 6-7 days after infection, to mice infected with P. berghei-NK65, where parasitemia increases steadily over time without neurological complications (Shaw et al., 2015).
Analysis of the levels of ANG2 collected over the course of infection showed increases in early infection in some of the mice infected with P. berghei-ANKA compared to P. berghei-NK65 (27% on days 4-5 and day 6) (Figures 2A, B). Similarly, analysis of plasma samples showed that the levels of ANGPTL4 progressively increased in the plasma of 50% of the mice after day 4 (Figures 2C, D). Twelve out of the total 14 mice presented a score ≥ 4 by days 6-8, which indicates severe signs of ECM and were euthanized. The two mice that developed less severe signs of ECM (score ≤ 3) and survived past day 10, presented low levels of ANGPTL4 over the course of infection, and lower levels of parasitemia (Figure 2E). Similar trends were observed for both male and female mice. In comparison, the levels of ANGPTL4 in mice infected with P. berghei-NK65, the non-cerebral strain, remained at lower levels throughout the course of infection. When analyzing the levels of ANGPTL4, we observed that all the CM strain-infected mice (P. berghei-ANKA) on day 6 post infection that presented higher levels of ANGPTL4 (over the NK65 max value of 3, 900) died within the next 2 days (Figures 2C, D). However, analysis of the parasitemia levels in all mice in the experiments revealed that the mice infected with P. berghei-NK65 mice had lower parasitemias compared to the mice infected with P. berghei-ANKA, even if both groups were infected with the same number of infected RBCs in the inoculum (Figures 2E, F).
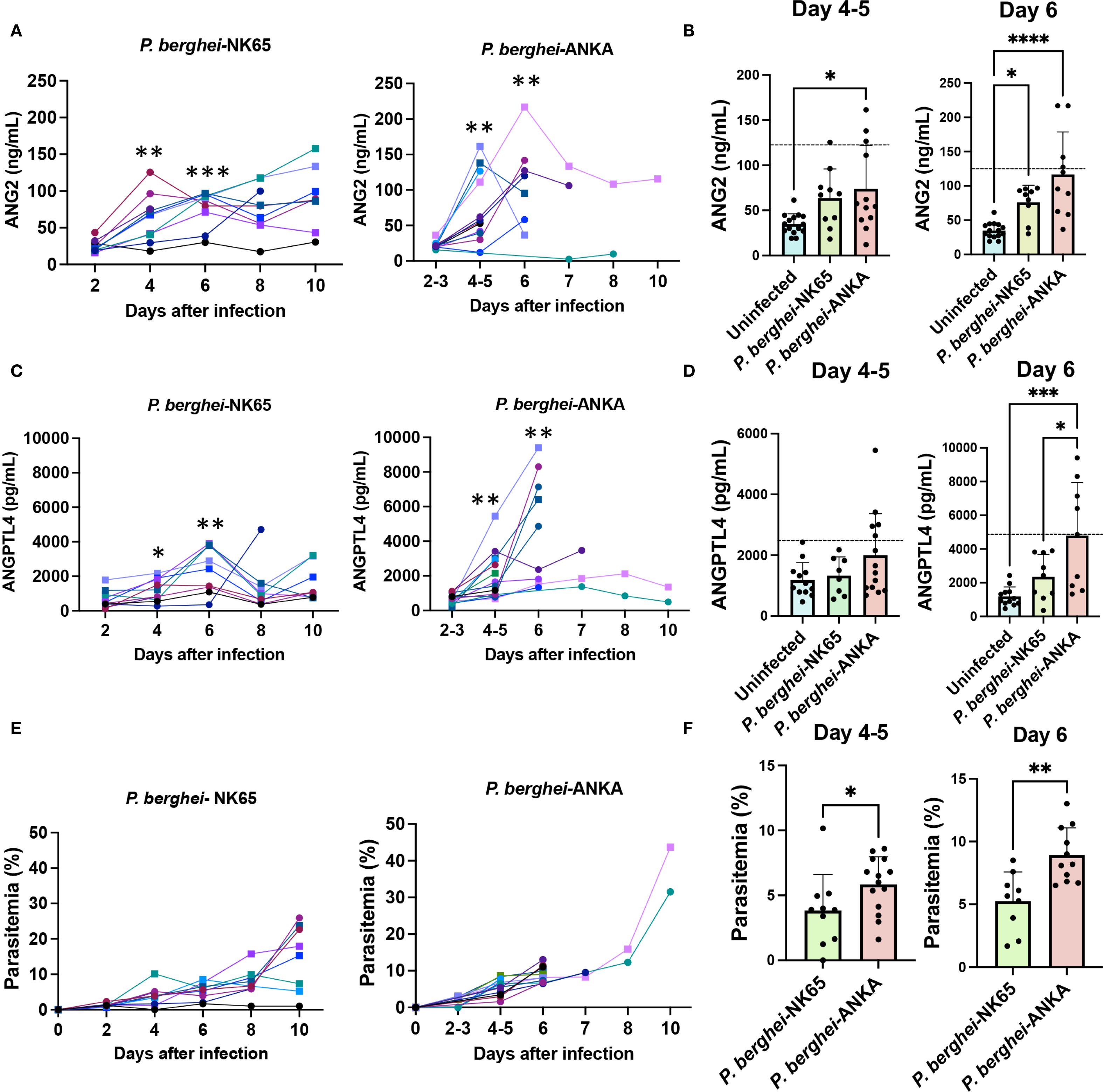
Figure 2. ANG2 and ANGPTL4 are elevated in mice with ECM. Levels of ANG2 and ANGPTL4 in plasma were determined in groups of C57/BL6 mice infected with P. berghei-ANKA (n = 14) or P. berghei-NK65 (n=10). (A, C) Levels of each biomarker in each individual mouse over the course of infection. Males are represented by squares and females by circles. T-test was used to calculate the significance of paired values for each mouse in days 4 and 6 were compared to day 2. (B, D) Comparison of biomarker levels between groups of uninfected and infected mice. Dotted line represents the average of non-CM mice (P. berghei-NK65) plus two times the standard deviation. (E, F) Levels of parasitemia in each individual mice over the course of infection (E) and comparison of parasitemias by t-test. Each symbol represents the same mouse in (A, C, E). p values determined by ANOVA (A-D) or t-test (F). ****p<0.0001; ***p<0.001; **p<0.01; *p<0.05.
These results are comparable to data collected from human patients, which showed significantly increased levels of ANGPTL4 in children with CM versus children with severe (non-cerebral) malaria (Gomes et al., 2023). Since only the mice infected with P. berghei-ANKA died during infection, the results obtained comparing the levels of ANGPTL4 in day 6 also indicate that this biomarker is elevated in a large proportion of mice before death.
We then analyzed the statistical relation of ANG2 and ANGPTL4 with CM (measured as standard scores for each mouse) and with parasitemia over the different days of the assay. No significant correlations with ANG2 were found. A significant direct correlation between the levels of ANGPTL4 and CM scores (ρ 0.41, p < 0.05) was observed, but no correlation was found for each of them with levels of parasitemia (p > 0.05 for both biomarkers) (Figure 3). Although there was no significant correlation between the severity of CM (scores) and the levels of parasitemia in mice infected with P. berghei-ANKA, we observed that these mice had overall higher levels of parasitemia compared to mice infected with P. berghei-NK65 (Figures 2E, F). Therefore, we analyzed the relation between parasitemia levels and biomarkers in mice infected with P. berghei-NK65, observing a direct correlation for both candidate biomarkers (ANG2 ρ 0.54, p < 0.05; ANGPTL4 ρ 0.51, p < 0.05). These results suggest that parasitemia may be contributing to the increase in the levels of the candidate biomarkers in these mice.
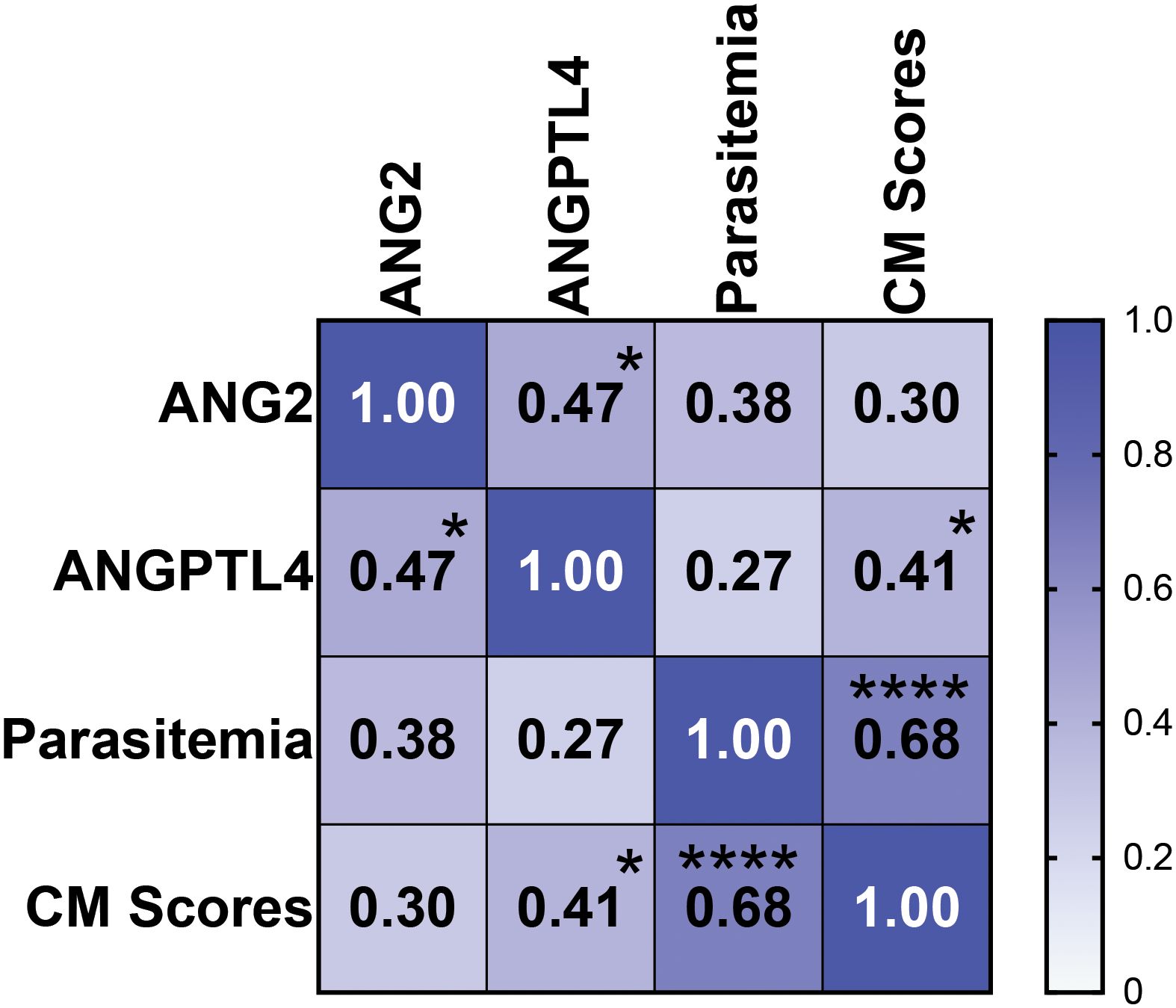
Figure 3. Correlation of study parameters in P. berghei-ANKA infected mice. Spearman coefficients were calculated for each pair of variables. Numbers indicate the Spearman’s ρ between the two indicated variables. The p values for each Spearman’s ρ are shown by asterisks (*p < 0.05; ****p < 0.0001). No inverse correlations were observed.
These results complement previous findings in human patients (Gomes et al., 2023) and suggest that levels of ANGPTL4 may be directly related to CM pathology in mice. ANG2 relation with CM scores (ρ 0.30, p = 0.14) did not reach significance. As expected, a high correlation between the levels of parasitemia and CM scores was observed (Figure 3).
The prognostic value of the biomarkers was assessed by determining whether their levels are elevated before the symptoms (CM scores) appear. Considering the maximum level of each biomarker in P. berghei-NK65-infected mice each day, this analysis revealed that 3 mice for ANG2 and 5 mice for ANGPTL4 had increased levels at least one day before CM scores were increased (Score ≥ 1) in a total of 13 mice analyzed. The calculated predictive value is 23% for ANG2 and 38% for ANGPTL4.
Discussion
The diagnosis of CM is performed following WHO criteria that requires a Blantyre coma score ≤2 and the presence of P. falciparum parasites in peripheral blood (WHO, 2014). However, this diagnosis is frequently inaccurate, as was evidenced by an autopsy study where 23% of children that had been diagnosed with CM following WHO criteria, lacked sequestration of P. falciparum infected erythrocytes in brain vessels and an alternative cause of death was identified for each one (Taylor et al., 2004). The high inaccuracy of the CM diagnosis is caused by the unspecificity of the clinical features of malaria coma (convulsions, anemia, respiratory distress and abnormal posturing) compared to other causes of coma and by the high proportion of children in malaria endemic areas with detectable circulating P. falciparum parasitemia in the absence of any clinical symptoms [as high as 50% (O’Meara et al., 2008)]. Moreover, CM diagnosis is frequently confounded by other lethal infectious diseases like bacterial meningitis or is accompanied by other invasive infections, with an important increase in the case fatality rate of those patients (Berkley et al., 1999; Aipit et al., 2014).
Under these circumstances, the identification of specific biomarkers for CM could help increase the accuracy of the diagnosis, resulting in improved clinical care. Previous studies in children with CM have identified a limited number of potential biomarkers, however variability between different studies and lack of sensitivity and specificity of the candidate biomarkers for CM have precluded the establishment of a reliable biomarker for CM (Foko et al., 2022).
An additional obstacle that difficulties adequate management of CM is the lack of a reliable early diagnostic tool. Since most malaria cases are either asymptomatic or uncomplicated, and only a small percentage of cases develops into severe disease, it would be highly beneficial to predict which children are at risk of developing severe complications. However, the identification of prognostic biomarkers for CM would require the collection and analysis of thousands of patient samples at the early/uncomplicated stage to be able to identify and analyze sufficient number of samples from the early stages of CM, when prognostic biomarkers may be apparent.
Considering these difficulties, we have analyzed the onset of human CM biomarker candidates in mice, where we can easily obtain early samples of individual mice that later develop ECM. Our approach confirmed that two previously identified biomarkers of CM in humans (Gomes et al., 2023), ANG2 and ANGPTL4, are elevated in a high proportion of the mice that develop CM compared to mice with uncomplicated infections. These results indicate that the mouse model can reflect the changes observed in humans and opening the possibility to use mice as a model to study CM biomarkers.
We also observed a sustained increase of biomarker levels and clinical signs (CM scores) in plasma throughout the course of infection. Detailed analysis to identify whether the increase in biomarker levels preceded clinical signs indicated a moderate predictive value of the biomarkers (lower than 50%), which suggests they may be useful for the early identification of CM in combination with other biomarkers. A broader kinetic screen of human candidate CM biomarkers in the mouse model could help identify specific candidates that increase their levels early in cerebral infections and may be useful as prognostic biomarkers.
ANG2 is a broad biomarker for diseases with a disfunctional endothelial component, including cancer (Wu et al., 2017) and various infections (Akwii et al., 2019). ANG2 is readily released in the circulation upon endothelial activation induced by inflammatory stimuli, which is a common characteristic of multiple infections (Fiedler and Augustin, 2006).
ANGPTL4, on the other hand, is upregulated in response to hypoxia, promoting vascular permeability through the activation of HIF-1 (Xin et al., 2013). It has been identified as a candidate biomarker for infections which have a major endothelial component, such as sepsis (Xu et al., 2015), severe COVID-19 (Bhatraju et al., 2023) and Acute Respiratory Distress Syndrome (ARDS) (Hu et al., 2021).
ANGPTL4, which is elevated in the plasma of patients with CM (Gomes et al., 2023), was secreted by endothelial cells in response to P. falciparum a 2D in vitro model (Marsh et al., 1995; Gomes et al., 2023), but was not in an 3D model including pericytes (Long et al., 2025). Since pericytes regulate endothelial cells in the BBB (Armulik et al., 2010), these results may indicate a negative regulation of ANGPTL4 expression by these cells. In a mouse model of sepsis, ANGPTL4 was also identified as a key regulatory component of the inflammatory response (Sun et al., 2024).
ANG2 and ANGPTL4 were found to be elevated in human severe malaria when compared to uncomplicated, but also in severe, non-cerebral malaria compared to CM (Conroy et al., 2009; Gomes et al., 2023). However, some patients with CM present also with other severe complications, such as ARDS, which was found to range from 10% to 20% of all CM cases (Marsh et al., 1995; Gomes et al., 2023). It should be noted that mice infected with P. berghei-ANKA can also develop overlapping lung pathology (Lovegrove et al., 2008) that could contribute to the elevation of the candidate biomarkers in this study. Additionally, ANGPTL4 was found to induce pulmonary tissue leakiness in a model of pneumonia, suggesting that this factor could be directly involved in lung pathology (Li et al., 2015).
A detailed analysis of the relation of these biomarkers with CM scores in the mice, showed that levels of ANGPTL4 are directly correlated with CM scores. It is likely that the increase of plasma ANGPTL4 in CM is caused by hypoxia, which is characteristic of CM and is also known to trigger the secretion of ANGPTL4 (Xin et al., 2013). ANGPTL4 levels did not correlate with parasitemia in mice infected with P. berghei-ANKA (Figure 3), which would indicate that this biomarker is related to CM pathology, rather than levels of infection in the mice. A limitation of this study is that a more significant comparison of mice infected with P. berghei-ANKA that developed or not CM pathology was not possible because of the low percentage of mice that do not develop the pathology (14%). Since there is a positive correlation between the levels of parasitemia in P. berghei-NK65 mice and both candidate biomarkers, it is likely that parasitemia is also a driver for the increased levels of both candidate biomarkers. Similar results were found in human patients where higher levels of ANGPTL4 were found in children with CM compared with other forms of severe malaria, and were positively correlated with the levels of HRP2 (a surrogate for parasitemia) (Conroy et al., 2009; Gomes et al., 2023).
ANG2 association with CM scores was not statistically significant. It is likely that differences with human CM studies where ANG2 correlates strongly with CM (Conroy et al., 2012), are due to the low number of mice samples compared to human patients.
In summary, our results in mice reflect previous findings in humans for candidate biomarkers ANGPTL4 and ANG2 and provide novel evidence that mouse models may help in the identification and selection of prognostic and/or diagnostic biomarkers for CM.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was approved by Institutional Animal Care and Use Committee of New York University School of Medicine. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
CG: Conceptualization, Data curation, Investigation, Writing – review & editing, Funding acquisition. LC: Investigation, Writing – review & editing. KC: Investigation, Writing – review & editing. IS: Writing – review & editing, Investigation. DT: Writing – review & editing, Investigation. AR: Conceptualization, Formal analysis, Funding acquisition, Project administration, Supervision, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This work was supported by NIH grants R01AI181219, R01HL150145 and R01NS105910 to AR and a Research Grant 2023 of the European Society of Clinical Microbiology and Infectious Diseases (ESCMID) to CG.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Aipit J., Laman M., Hwaiwhanje I., Bona C., Pomat N., Siba P., et al. (2014). Accuracy of initial clinical diagnosis of acute bacterial meningitis in children from a malaria-endemic area of Papua New Guinea. Trans. R Soc. Trop. Med. Hyg 108, 444–448. doi: 10.1093/trstmh/tru067
Akwii R. G., Sajib M. S., Zahra F. T., and Mikelis C. M. (2019). Role of angiopoietin-2 in vascular physiology and pathophysiology. Cells 8 (5), 471. doi: 10.3390/cells8050471
Alger N. E., Branton M., Harant J., and Silverman P. H. (1971). Plasmodium berghei NK65 in the inbred A-J mouse: variations in virulence of P. berghei demes. . J. Protozool 18, 598–601. doi: 10.1111/j.1550-7408.1971.tb03382.x
Armulik A., Genove G., Mae M., Nisancioglu M. H., Wallgard E., Niaudet C., et al. (2010). et al: Pericytes regulate the blood-brain barrier. Nature 468, 557–561. doi: 10.1038/nature09522
Berkley J. A., Mwangi I., Mellington F., Mwarumba S., and Marsh K. (1999). Cerebral malaria versus bacterial meningitis in children with impaired consciousness. QJM 92, 151–157. doi: 10.1093/qjmed/92.3.151
Bhatraju P. K., Morrell E. D., Stanaway I. B., Sathe N. A., Srivastava A., Postelnicu R., et al. (2023). et al: angiopoietin-like4 is a novel marker of COVID-19 severity. Crit. Care Explor. 5, e0827. doi: 10.1097/CCE.0000000000000827
Conroy A. L., Glover S. J., Hawkes M., Erdman L. K., Seydel K. B., Taylor T. E., et al. (2012). Angiopoietin-2 levels are associated with retinopathy and predict mortality in Malawian children with cerebral malaria: a retrospective case-control study*. Crit. Care Med. 40, 952–959. doi: 10.1097/CCM.0b013e3182373157
Conroy A. L., Lafferty E. I., Lovegrove F. E., Krudsood S., Tangpukdee N., Liles W. C., et al. (2009). Whole blood angiopoietin-1 and -2 levels discriminate cerebral and severe (non-cerebral) malaria from uncomplicated malaria. Malar J. 8, 295. doi: 10.1186/1475-2875-8-295
Fiedler U. and Augustin H. G. (2006). Angiopoietins: a link between angiogenesis and inflammation. Trends Immunol. 27, 552–558. doi: 10.1016/j.it.2006.10.004
Foko L. P. K., Narang G., Tamang S., Hawadak J., Jakhan J., Sharma A., et al. (2022). The spectrum of clinical biomarkers in severe malaria and new avenues for exploration. Virulence 13, 634–653. doi: 10.1080/21505594.2022.2056966
Franke-Fayard B., Trueman H., Ramesar J., Mendoza J., van der Keur M., van der Linden R., et al. (2004). A Plasmodium berghei reference line that constitutively expresses GFP at a high level throughout the complete life cycle. Mol. Biochem. Parasitol. 137, 23–33. doi: 10.1016/j.molbiopara.2004.04.007
Ghazanfari N., Mueller S. N., and Heath W. R. (2018). Cerebral malaria in mouse and man. Front. Immunol. 9, 2016. doi: 10.3389/fimmu.2018.02016
Gomes C., Varo R., Duran-Frigola M., Sitoe A., Bila R., Machevo S., et al. (2023). Endothelial transcriptomic analysis identifies biomarkers of severe and cerebral malaria. JCI Insight, 8 (22), e172845. doi: 10.1172/jci.insight.172845
Guinovart C., Sigauque B., Bassat Q., Loscertales M. P., Nhampossa T., Acacio S., et al. (2022). et al: The epidemiology of severe malaria at Manhica District Hospital, Mozambique: a retrospective analysis of 20 years of malaria admissions surveillance data. Lancet Glob Health 10, e873–e881. doi: 10.1016/S2214-109X(22)00125-5
Hu J., Liu L., Zeng X., Wang K., Wang H., Zeng Z., et al. (2021). Prognostic value of angiopoietin-like 4 in patients with acute respiratory distress syndrome. Shock 56, 403–411. doi: 10.1097/SHK.0000000000001734
John C. C., Bangirana P., Byarugaba J., Opoka R. O., Idro R., Jurek A. M., et al. (2008). Cerebral malaria in children is associated with long-term cognitive impairment. Pediatrics 122, e92–e99. doi: 10.1542/peds.2007-3709
Li L., Chong H. C., Ng S. Y., Kwok K. W., Teo Z., Tan E. H. P., et al. (2015). et al: Angiopoietin-like 4 Increases Pulmonary Tissue Leakiness and Damage during Influenza Pneumonia. Cell Rep. 10, 654–663. doi: 10.1016/j.celrep.2015.01.011
Long R. K. M. K., Ronchi P., Fleckenstein P., Schorb M., Mirza W., Mallorquí M., et al. (2025). Plasmodium falciparum impairs Ang-1 secretion by pericytes in a 3D brain microvessel model. Nat. Commun. 16, 7262. doi: 10.1038/S41467-025-62514-2
Lovegrove F. E., Gharib S. A., Pena-Castillo L., Patel S. N., Ruzinski J. T., Hughes T. R., et al. (2008). Parasite burden and CD36-mediated sequestration are determinants of acute lung injury in an experimental malaria model. PloS Pathog. 4, e1000068. doi: 10.1371/journal.ppat.1000068
Marsh K., Forster D., Waruiru C., Mwangi I., Winstanley M., Marsh V., et al. (1995). Indicators of life-threatening malaria in African children. N Engl. J. Med. 332, 1399–1404. doi: 10.1056/NEJM199505253322102
Muppidi P., Wright E., Wassmer S. C., and Gupta H. (2023). Diagnosis of cerebral malaria: Tools to reduce Plasmodium falciparum associated mortality. Front. Cell Infect. Microbiol. 13, 1090013. doi: 10.3389/fcimb.2023.1090013
O’Meara W. P., Bejon P., Mwangi T. W., Okiro E. A., Peshu N., Snow R. W., et al. (2008). Effect of a fall in malaria transmission on morbidity and mortality in Kilifi, Kenya. Lancet 372, 1555–1562. doi: 10.1016/S0140-6736(08)61655-4
Rutherford M. E., Dockerty J. D., Jasseh M., Howie S. R., Herbison P., Jeffries D. J., et al. (2009). Access to health care and mortality of children under 5 years of age in the Gambia: a case-control study. Bull. World Health Organ 87, 216–224. doi: 10.2471/BLT.08.052175
Seydel K. B., Kampondeni S. D., Valim C., Potchen M. J., Milner D. A., Muwalo F. W., et al. (2015). et al: Brain swelling and death in children with cerebral malaria. N Engl. J. Med. 372, 1126–1137. doi: 10.1056/NEJMoa1400116
Shaw T. N., Stewart-Hutchinson P. J., Strangward P., Dandamudi D. B., Coles J. A., Villegas-Mendez A., et al. (2015). et al: perivascular arrest of CD8+ T cells is a signature of experimental cerebral malaria. PloS Pathog. 11, e1005210. doi: 10.1371/journal.ppat.1005210
Sun B., Bai L., Li Q., Sun Y., Li M., Wang J., et al. (2024). Knockdown of angiopoietin-like 4 suppresses sepsis-induced acute lung injury by blocking the NF-kappaB pathway activation and hindering macrophage M1 polarization and pyroptosis. Toxicol. In Vitro 94, 105709. doi: 10.1016/j.tiv.2023.105709
Taylor T. E., Fu W. J., Carr R. A., Whitten R. O., Mueller J. S., Fosiko N. G., et al. (2004). Differentiating the pathologies of cerebral malaria by postmortem parasite counts. Nat. Med. 10, 143–145. doi: 10.1038/nm986
Waknine-Grinberg J. H., Hunt N., Bentura-Marciano A., McQuillan J. A., Chan H. W., Chan W. C., et al. (2010). Artemisone effective against murine cerebral malaria. Malar J. 9, 227. doi: 10.1186/1475-2875-9-227
WHO (2013). Programme WGM: WHO informal consultation on fever management in peripheral health care settings: A global review of evidence and practice.
Wu X., Giobbie-Hurder A., Liao X., Connelly C., Connolly E. M., Li J., et al. (2017). et al: angiopoietin-2 as a biomarker and target for immune checkpoint therapy. Cancer Immunol. Res. 5, 17–28. doi: 10.1158/2326-6066.CIR-16-0206
Xin X., Rodrigues M., Umapathi M., Kashiwabuchi F., Ma T., Babapoor-Farrokhran S., et al. (2013). et al: Hypoxic retinal Muller cells promote vascular permeability by HIF-1-dependent up-regulation of angiopoietin-like 4. Proc. Natl. Acad. Sci. U.S.A. 110, E3425–E3434. doi: 10.1073/pnas.1217091110
Xu X., Shi Z., Hu J., Yuan B., Huang H., Fang H., et al. (2015). Identification of differentially expressed genes associated with burn sepsis using microarray. Int. J. Mol. Med. 36, 1623–1629. doi: 10.3892/ijmm.2015.2374
Keywords: cerebral malaria (CM), Plasmodium berghei (ANKA strain), Plasmodium berghei (NK65 strain), biomarkers, mice, experimental cerebral malaria (ECM)
Citation: Gomes C, Chicas L, Crotty KA, Salzano I, Tawfiles D and Rodriguez A (2025) Specificity and kinetics of human candidate cerebral malaria biomarkers in mice. Front. Malar. 3:1553466. doi: 10.3389/fmala.2025.1553466
Received: 30 December 2024; Accepted: 04 September 2025;
Published: 17 September 2025.
Edited by:
Sam C Wassmer, University of London, United KingdomReviewed by:
Laurent Rénia, Nanyang Technological University, SingaporeAmma Larbi, Kwame Nkrumah University of Science and Technology, Ghana
Copyright © 2025 Gomes, Chicas, Crotty, Salzano, Tawfiles and Rodriguez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ana Rodriguez, YW5hLnJvZHJpZ3VlekBueXVtYy5vcmc=