 René Ortega-Monroy1†
René Ortega-Monroy1† Hugo A. Tomasini-Ovilla1†
Hugo A. Tomasini-Ovilla1† Frida Santillan-Valenzuela1
Frida Santillan-Valenzuela1 Alberto Montoya2
Alberto Montoya2 Lilia Gonzalez-Ceron1*
Lilia Gonzalez-Ceron1*- 1Regional Center of Research in Public Health, National Institute of Public Health, Ministry of Health, Tapachula, Mexico
- 2Parasitology Department, National Centre for Diagnosis Reference, Ministry of Health, Managua, Nicaragua
Background: Plasmodium vivax Duffy binding protein domain II (PvBDPII) is a promising vaccine candidate due to its crucial role in reticulocyte invasion. However, polymorphism is a concern as it may confer evasion of blocking antibodies. In this study, nucleotide and protein polymorphism were analyzed in parasites collected during a decline in malaria cases in Nicaragua.
Methods: Genomic DNA was extracted from P. vivax blood samples obtained from symptomatic patients in Nicaragua during 2012–2013. The PvdbpII gene was amplified and sequenced, and genetic structure, genealogical relationships, and amino acid polymorphism were analyzed. For comparison and to elucidate global variation of this gene, homologous sequences from other geographical regions were included.
Results: Sixty-three consensus sequences from Nicaragua were obtained, revealing 12 non-synonymous mutations and 6 haplotypes (Hd=0.704). Nucleotide diversity was lower (π=0.0044) than in other endemic regions. The Z test of selection (dN/dS) was positive (3.17; p < 0.001), similar to parasite populations worldwide (Latin America, Middle East, Asia, Southeast Asia, Africa, Papua New Guinea). Network analysis revealed that Nicaraguan pvdbpII haplotypes differed by 3–12 mutational steps. High-frequency haplotypes from six other Latin American countries were shared, showing signs of diversification and limited population structure. Five Nicaraguan haplotypes corresponded to the 10 most frequent globally. Haplotypes defined solely by amino acid changes at positions 417, 437, and 503 were of the Sal-I type (NWI), NWK, and KRK. These, along with KRI, were shared across most regions at varying frequencies. Based on these variations, two main divergent groups were identified.
Conclusions: The low diversity observed in pvdbpII suggests a population contraction aligning with the decline of malaria cases in Nicaragua during the sampling period. The PvDBPII haplotypes found may represent those best adapted in Nicaragua and in other endemic regions globally, encompassing both Sal-I-related and divergent types. It would be beneficial to assess the ability of the most frequent and persistent haplotypes to elicit phenotype-transcending immunity, which is critical for the development of a multicomponent vaccine and for monitoring its effectiveness.
Introduction
Malaria is caused by protozoan parasites of the genus Plasmodium and transmitted by female mosquitoes of the genus Anopheles. More than 85 countries are at risk of malaria transmission. In 2023, there were 263 million clinical cases, representing an increase of 11 million compared with the previous year (World Health Organization, 2024).
In the Region of the Americas, malaria cases declined by 65.4%, from 1.5 million to 0.6 million between 2000 and 2022, and in 2023 (World Health Organization, 2024). Plasmodium vivax accounted for approximately 73% of the 505,642 confirmed malaria cases, while about 27% were caused by Plasmodium falciparum (World Health Organization, 2024). The Central and South America have experienced significant changes in malaria transmission. In Nicaragua, malaria cases gradually decreased between 2000 and 2010, reaching more than 90% reduction (from 23,281 cases in 2000 to 896 in 2010 (World Health Organization, 2023). The number of cases fluctuated slightly between 2011 and 2014 (1,171–1,564 per year) and then drastically increased from 2,886 in 2015 to 23,323 and 16,158 in 2021 and 2022, respectively (World Health Organization, 2023). Although Nicaragua reported fewer cases in 2023 (6,716), it remained the country with the highest malaria burden in Central America (World Health Organization, 2024). Malaria elimination requires continuous assessment and improvement of control measures and strategies. Understanding parasite molecular diversity contributes to knowledge of disease epidemiology, delineates transmission patterns, and informs the development of vaccine candidates and antimalarial drug resistance monitoring. Studies of genetic and antigenic diversity can also help predict and monitor the effectiveness of interventions by detecting reductions in parasite diversity or the emergence of biologically important phenotypes (Arnott et al., 2012; Escalante and Pacheco, 2019).
P. vivax merozoites invade reticulocytes through complex interactions between parasite ligands and reticulocyte receptors. The Duffy binding protein (DBP) domain II (DBPII) participates in the formation of a tight, irreversible junction with the erythrocyte Duffy antigen receptor (DARC), which is essential for successful invasion (Singh et al., 2005). The pvdbp gene, located on chromosome six, encodes a 140-kDa type 1 integral membrane protein that is secreted into the microneme. Its structure contains seven domains: a signal peptide, two cysteine-rich domains (II and VI), three hydrophobic domains (III–V), and one transmembrane domain (VII) (Adams et al., 1992; Ranjan and Chitnis, 1999). The cysteine-rich DBPII consists of 170 amino acids spanning residues 198–522. This domain is highly immunogenic, and during infection it induces blocking antibodies that prevent P. vivax merozoite invasion of reticulocytes (Michon et al., 1998, 2000). These blocking antibodies are thought to be the mechanism by which a PvDBPII-based vaccine could confer protection, making this protein a promising target for vaccine development (Tsuboi et al., 1994). Both linear and conformational epitopes may be important in eliciting a protective immune response. Antibodies reactive to PvDBPII linear epitopes have been detected in residents of endemic areas in Papua New Guinea following repeated P. vivax exposure (Cole-Tobian et al., 2002; Xainli et al., 2003; Cole-Tobian et al., 2007).
Analysis of nucleotide diversity in pvdbpII provides information on circulating antigenic variants and the feasibility of a universal vaccine (Nobrega de Sousa et al., 2011). Domain II accounts for 93% of the polymorphism in the PvDBP molecule and may play a role in parasite immune evasion (Cole-Tobian et al., 2007; Chootong et al., 2014; Sousa et al., 2010; VanBuskirk et al., 2004). The 10 most frequent global amino acid variations in PvDBPII include R308S, K371Q, D384G, Q385K, K386N/E, R390H, N417K, L424I, W437R, and I503K (Sousa et al., 2011; Almeida-de-Oliveira et al., 2020)Residues N417K, W437R, and I503K have been implicated in immune evasion (VanBuskirk et al., 2004). More recently, residues D309–A326, Q401–N417, L315–K334, and Q401–W420 have been identified as participating in conformational epitopes (Urusova et al., 2019).
Central America appears to be the last region colonized by P. vivax, with parasite populations exhibiting low diversity and similarity to the Sal-I strain (Taylor et al., 2013). Previous genetic studies using markers such as pvama1 and pvmdr1 suggest that P. vivax in Nicaragua (Gonzalez-Ceron et al., 2017; 2023) may represent a unique population, possibly not exposed to pyrimethamine and distinct from South American populations (Gonzalez-Ceron et al., 2020). To characterize PvDBP diversity in Central America, this study analyzed nucleotide diversity and molecular polymorphism of P. vivax DBP domain II using isolates from Nicaragua collected in 2012–2013. Homologous sequences from Latin America and other geographical regions were included for comparative analysis and to assess global variation.
Materials and methods
The study was approved by the Ethics Committee of the National Institute of Public Health (CI1042) and the National Centre for Diagnosis and Reference, Ministry of Health, Nicaragua. The study followed bioethics guidelines (CITI program). All participants older than 18 years provided written informed consent, and minors aged 7–17 years gave assent accompanied by the written informed consent of one parent or guardian, in accordance with the Declaration of Helsinki. Patient personal information was encrypted, and only the municipality of residence was recorded.
P. vivax samples
P. vivax diagnosis was conducted by parasitological analysis of thick blood smears at the national sentinel laboratory network of the Ministry of Health. Blood samples were obtained from symptomatic patients living in affected areas of Nicaragua between 2012 and 2013 (Gonzalez Ceron et al., 2023; Gutierrez et al., 2016). P. vivax infected blood samples were spotted onto Whatman #2 filter paper, dried, and stored in the dark for later use. Sixty-five previously obtained samples from different municipalities were selected for this study.
Gene amplification and sequencing
DNA extraction was performed using the QIAamp Blood Minikit (Qiagen, Valencia, CA, USA) according to the manufacturer’s instructions. Three circles of 5 mm each were cut from the dried blood spots. A final elution volume of 50 µL containing extracted DNA was stored at −20°C for subsequent analysis. The Pvdbp domain II was amplified using nested PCR. The first PCR was performed in a 20 µl reaction mixture containing; 9.2µl H2O, 4µl 5x Buffer, 1.6µl 25mM MgCl2, 1µl 1.2mM dNTP, 2µl 10µM DBPF1 (5´ -GATAAAACTGGGGAGGAAAAAGAT- 3´) and DBPR1 (5´-CTTATCGGATTTGAATTGGTGGC- 3´) primers, 0.2µl GoTaq® DNA polymerase (Promega, Madison, WI, USA), and 2µl extracted DNA. The nested PCR was performed in a 50 µl reaction containing: 25.5 µl H2O, 10 µl 5x Buffer, 4 µl 25mM MgCl2, 2.5 µl 1.2mM dNTP, 5 µl 10µM DBPF3 (5´ -CCTCGAATGGTGGCAATCCT- 3’) and DBPR3 (5’ -CCTCTTCAACGGAACAAACGCA- 3’) primers, 0.5µl of GoTaq® Polymerase and 2.5µl of first PCR product to obtain a fragment of ∼984 bp.
Both PCR reactions were carried out under the following conditions: 95°C for 3 min; 35 cycles of 94°C for 40 s, 58°C for 40 s, and 72°C for 90 s; and a final extension at 72°C for 5 min. Reactions were performed in a T100 Thermal Cycler (Bio-Rad, Hercules, CA, USA).
Amplified fragments were resolved on 1% agarose gels stained with ethidium bromide and visualized under ultraviolet (UV) light using a digital photo documentation system (UVP Inc., Upland, CA, USA). PCR products were purified using the MiniElute PCR Purification Kit (Qiagen, Valencia, CA, USA) according to the manufacturer’s instructions and quantified using a NanoDrop ND2000 spectrophotometer (Thermo Scientific, USA). Samples with a concentration of at least 30ng/µl were sequenced by the Sanger method with DBPF3 and DBPR3 primers at the High-Throughput Genomics Unit, University of Washington, Seattle, USA. Electropherograms were reviewed in BioEdit 7.2.5 (Hall, 1999) using the Sal-I reference sequence (XM_001608337.1). All sequences showed single peaks, suggesting a single genotype per amplified product. Consensus sequences were deposited in GenBank (accession numbers PV581685–PV581747).
Genetic analysis
Parameters of diversity and neutral evolution estimates were run in dnaSP v6.12 (Rozas et al., 2017), including the number of mutations (M), segregating sites (S), haplotypes (H), haplotype diversity (Hd), and synonymous and non-synonymous nucleotide changes (S, NS). Nucleotide diversity (π), defined as the average number of nucleotide differences per site between two or more sequences in a population, and genetic diversity (θ-w), defined as the variety in the DNA segment composition within a population, were also estimated.
Tajima’s D is based on the expectation that the estimates of S/a1 and k are unbiased estimates of θ-w under a neutral model (Tajima, 1989). Here, S is the number of segregating sites, n is the number of nucleotide sequences, and k is the average number of nucleotide differences between pairs of sequences. A negative Tajima’s D value indicates an excess of low-frequency polymorphism, which may suggest directional selection or population expansion, whereas a positive value suggests balancing selection or population contraction. D* and F* of Fu and Li evaluate whether the pattern of polymorphism is consistent with a neutral model. D* is based on the difference between ηs, the number of singletons (sites segregating at a frequency of 1/n or (n − 1)/n), and η, the total number of mutations, whereas F* is based on the difference between ηs and k (Fu and Li, 1993). Tests were run in DnaSP (Rozas et al., 2017).
The Z test of selection (dN/dS) was estimated using the Modified Nei–Gojobori model with Jukes–Cantor correction in MEGA v11 (Tamura et al., 2021). This test calculates the ratio of non-synonymous (dN) to synonymous (dS) substitutions per site across all sequence pairs, and the variance of the difference was computed using the bootstrap method with 1,000 replicates. To delineate genealogical relationships among pvdbpII haplotypes from Nicaragua and other regions, median-joining networks were generated using Population Analysis with Reticulate Trees in PopART v1.7 (Bandelt et al., 1999). Genetic structure differences in the distribution of variants were also analyzed to identify groups of sequences sharing similar patterns of variation, using a Bayesian clustering approach with Markov chain Monte Carlo (MCMC) estimation under an admixture model (Porras-Hurtado et al., 2013). Tests were run 20 times with a burn-in of 50,000 and 100,000 iterations for K values ranging from 2 to 5 in STRUCTURE v2.3.4 (Porras-Hurtado et al., 2013). The most probable number of clusters or subpopulations was estimated using the online tool Structure Harvester https://taylor0.biology.ucla.edu/structureHarvester/ (accessed 30 June 2023) (Earl and vonHoldt, 2012). Genetic differentiation between two or more populations was calculated using the FST index (Wright, 1951) in dnaSP v6.12.03 (Rozas et al., 2017). FST values range from 0 to 1, where 0 indicates genetically identical populations and 1 indicates completely unrelated populations.
To compare and gain a broader understanding of PvDBPII diversity, homologous sequences from various geographical sites were retrieved from NCBI. PvdbpII homologous sequences, accessible via NCBI (https://pubmed.ncbi.nlm.nih.gov) and PlasmoDB (http://plasmodb.org/plasmo/), were acquired and are listed as follows: Brazil (BRA, n = 122) EU812839.1 - EU812960.1 (Sousa et al., 2010); Mexico (MEX, n = 35) KP759780.1 - KP759814.1 (Gonzalez-Ceron et al., 2015), Ecuador (ECU, n = 51) Pv001-Pv091 (Nunez et al., 2023); Peru (PER, n = 11) Peru06-Peru10, Peru1008, Peru1021, Peru1022, Peru2025, Peru257-Peru262 (Hupalo et al., 2016); Colombia (COL, n = 38) U50590.1 - U50575.1 (Ampudia et al., 1996), and 30102100448 - 30102100440 (Hupalo et al., 2016); Iran (IRN; n = 12) KF791926.1 - KF791921.1 (Valizadeh et al., 2014a), KF318359.1 - KF318358.1 (Valizadeh et al., 2014b), and KF751810.1 - KF751807.1 (Nateghpour et al., 2017); Pakistan (PKN; n = 118) (Nguyễn et al., 2024); India (IND; n = 61) MN549534.1- MN549594.1 (unpublished), Sri Lanka (SLK; n = 100) GU143914.1 - GU144013.1 (Premaratne et al., 2011), China (CHN; n = 124) MZ765947.1 - MZ766070.1 (Shi et al., 2021); South Korea (SK; n = 13) JN989484.1 - JN989472.1 (Ju et al., 2013); Thailand (THL; n = 22) EF368159.1 - EF368180.1 (Gosi et al., 2008); Myanmar (MYN; n = 167) MN233573.1 - MN233489.1, MN233488.1 - MN233407.1 (Hu et al., 2019); Papua Nueva Guinea (PNG, n = 88) AF469515.1 - AF469602.1 (Cole-Tobian et al., 2002); Ethiopia (ETH; n = 75) MZ062409.1 - MZ062224.1 (Lo et al., 2021); Sudan (SUD; n = 97) MG805657.1 - MG805616.1 (Hoque et al., 2018), MZ062409.1 - MZ062224.3 (Lo et al., 2021); Uganda (UGA; n = 31) KX009560.1 - KX009537.1 (unpublished), JX174528.1 - JX174522.1 (unpublished); Botswana (BTW, n = 8) MZ062409.1 - MZ062224.2 (Lo et al., 2021).
Amino acid and haplotype polymorphism.
DNA sequences were translated into amino acid sequences, and the frequency of the most common amino acid substitutions and haplotypes found in Nicaragua was compared with those from other geographical regions. Additionally, the frequency of the eight amino acid combinations derived from polymorphic residues 417, 437, and 503 was compared regardless of other substitutions. These polymorphic residues are implicated in immune evasion (VanBuskirk et al., 2004).
P. vivax circulation in Nicaragua using pvdbpII versus pvdpbII - pvama1I-II
To test the sensitivity of pvdbpII haplotypes in tracking P. vivax circulation in Nicaragua, these haplotypes were mapped to the municipality level using ArcGIS v10.8.1. Furthermore, since pvama1I-II sequences were available for most isolates, their geographical distribution was compared with that of a concatenated sequence comprised of pvdbpII and pvama1I-II. Sixty-one isolates covering both pvdbpII (from this study) and pvama1I-II sequences (GenBank: ON730710.1–ON730730.1, ON730732.1–ON730736.1, ON730738.1–ON730746.1, ON730749.1–ON730774.1) (Gonzalez Ceron et al., 2023) were used. PvdbpII (1,806 bp) and pvama1I-II (915 bp) sequences were concatenated in MEGA v11, and haplotypes were resolved in DnaSP. Genetic relationships were investigated using a maximum likelihood phylogenetic tree in MEGA.
Results
Diversity parameters for pvdbpII, Nicaragua.
Sixty-three consensus pvdbpII sequences of 891 bp were obtained, corresponding to nucleotides 595–1485 of the Sal-I strain. Only nonsynonymous mutations were detected, and the mutation at codon 441 was a singleton (Supplementary Table 1). No mixed-genotype infections were detected. Nucleotide and genetic diversity were low (π = 0.006 and θ-w = 0.004, respectively). A sliding window analysis of nucleotide diversity showed higher values across nucleotides 900–1500 (Figure 1). Six haplotypes were detected, with haplotype diversity (Hd) of 0.704. Tajima’s D for pvdbpII was 1.531, and Fu and Li’s D* and F* statistics were 1.411 and 1.326, respectively; none were significant. The R² coefficient of linkage disequilibrium for the Nicaraguan parasites was 0.356.
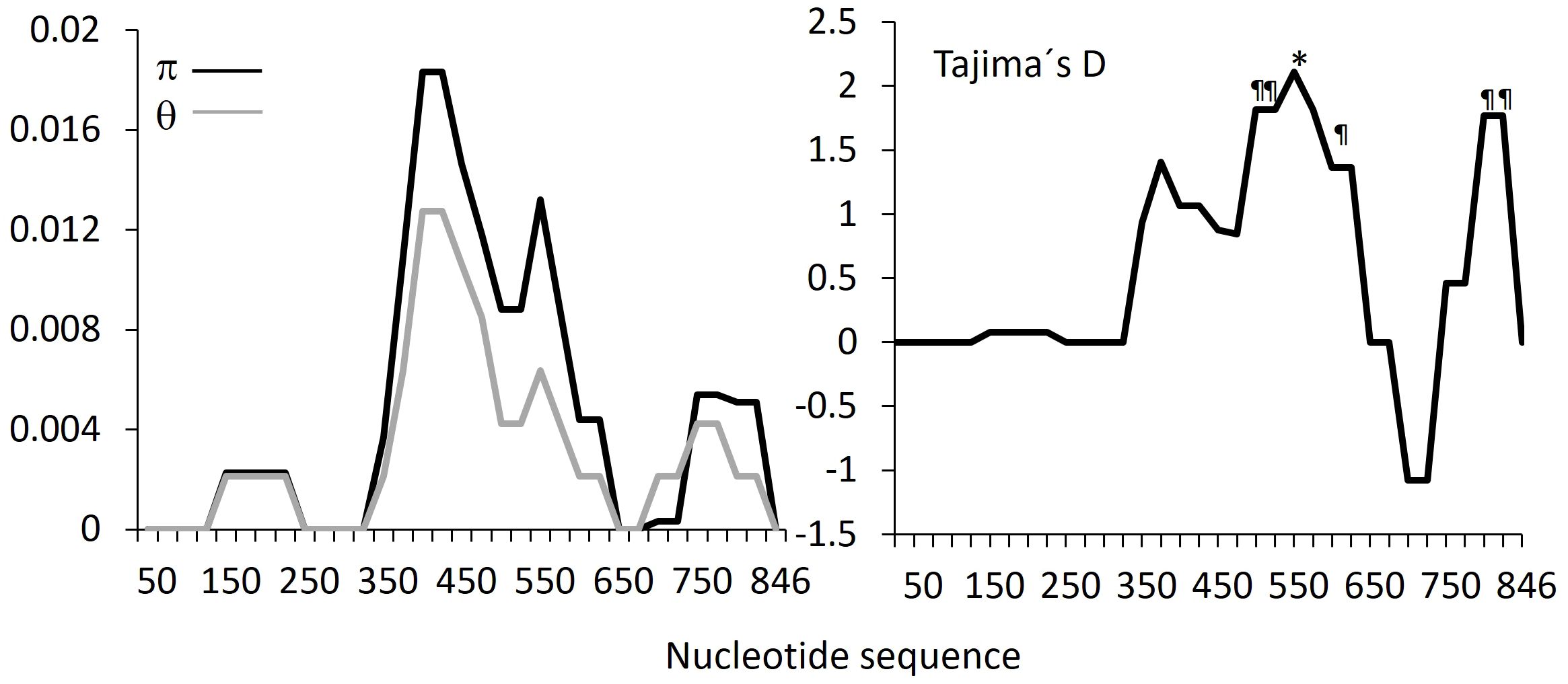
Figure 1. Sliding window analysis of pvdbpII gene fragments from Nicaraguan isolates. Midpoints were calculated for a window length of 100 bp with 25 overlapping nucleotides * P<0.05, ¶ P<0.10.
Haplotype networks
A median-joining network using 63 sequences from Nicaraguan parasites showed six haplotypes separated by 1–12 mutational steps. Haplotype Nh1 was the most frequent and was separated by three mutational steps from Nh2 and Nh6, and by four steps from Nh5. Nh4 and Nh3 were separated by one and three mutational steps from Nh5, respectively. Nh2 and Nh6 shared two nonsynonymous mutations (R384G and R390H), while Nh3, Nh4, and Nh5 (divergent haplotypes) shared four nonsynonymous mutations (N417K, L424I, W437R, and I503K) (Figure 2).
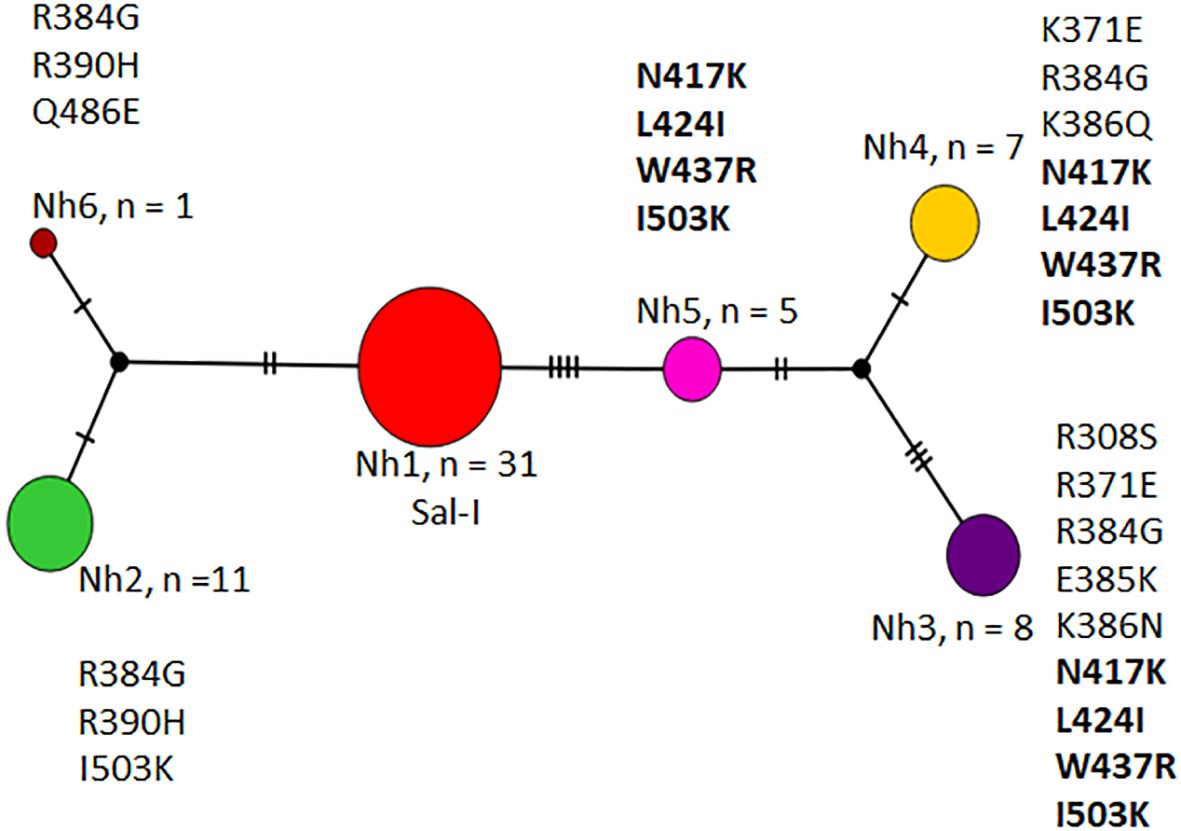
Figure 2. Haplotype network of P. vivax dbpII from Nicaragua, 2012–2013. Haplotypes are represented by different colors, with each circle corresponding to a unique haplotype. The circle size is proportional to the number of sequences sharing that haplotype. As only nonsynonymous mutations were detected across the 63 sequences, the amino acid polymorphism defining each haplotype is indicated. The four common mutations present in haplotypes Nh3, Nh4, and Nh5 are shown in bold.
The haplotype network of global isolates was highly complex, with multiple genealogical groups and no discernible geographic structure, as reported previously (Nguyễn et al., 2024). Figure 3A shows a network of Latin American pvdbpII haplotypes present in at least two individuals. Intra-haplotype mutational steps ranged from 1 to 30. Four of the six haplotypes from Nicaragua represented the most frequent haplotypes (Nh1, Nh2, Nh3, and Nh5), albeit at different frequencies (Figure 3A). A higher number of unique haplotypes were detected in Ecuador > Brazil > Colombia compared with other Latin American sites.
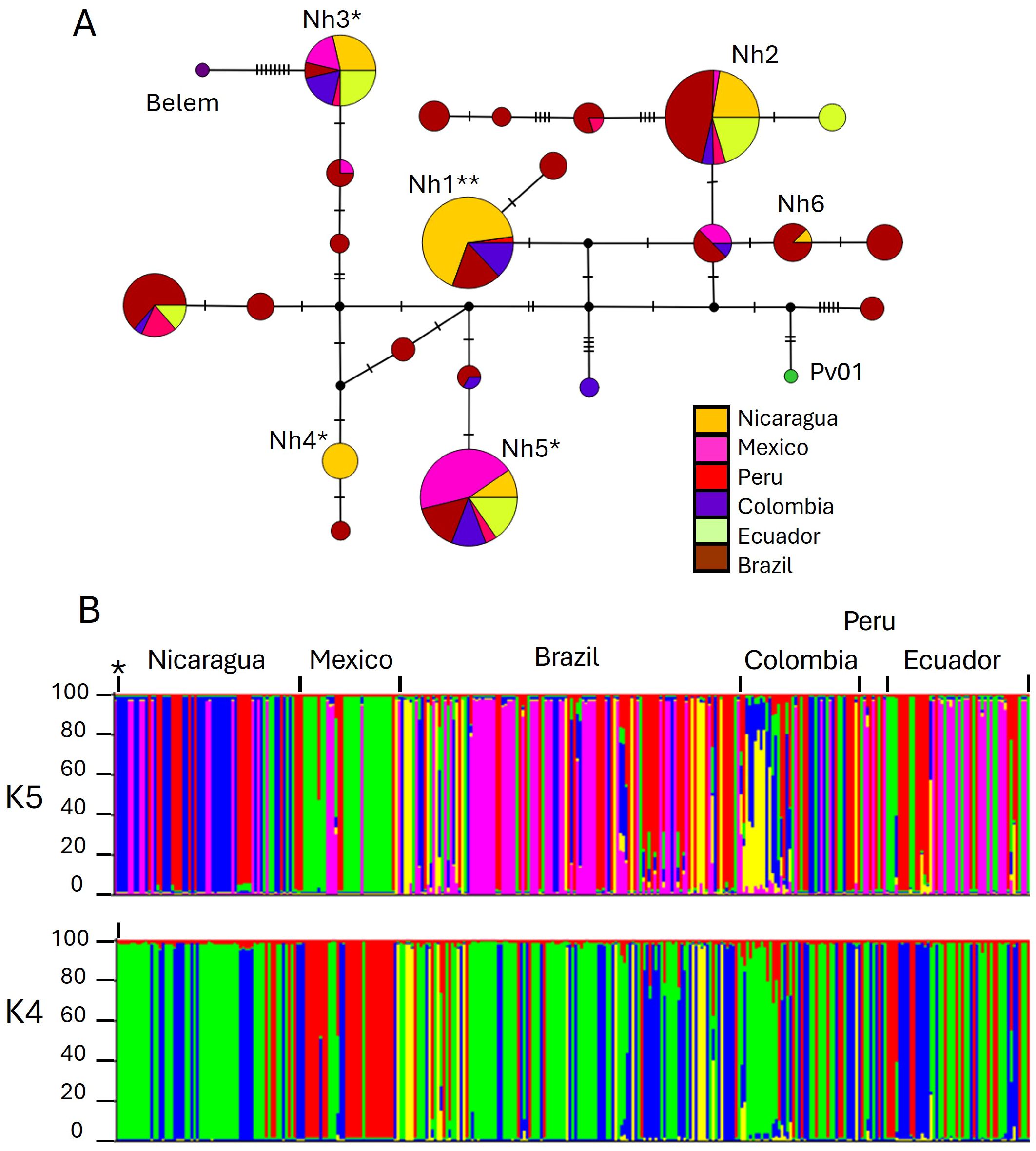
Figure 3. Haplotype network and genetic structure of P. vivax dbpII from Latin America. (A) Median-joining haplotype network of pvdbpII haplotypes. Each circle represents a unique haplotype, and its size is proportional to haplotype frequency (based on 269 sequences, including only haplotypes with two or more sequences). Haplotypes are connected by lines, with short vertical stripes indicating the number of mutational steps between them. Nh1–Nh6 denotes Nicaraguan haplotypes. Reference strains such as Sal-I (**), Pv01, and Belem were included. *Haplotypes showing four simultaneous amino acid changes at residues N417K, L424I, W437R, and I503K are marked. (B) Genetic structure of 319 P. vivax isolates. Each color represents one subpopulation. The Y-axis indicates the percentage of shared ancestry among individuals. The blue (K = 5) or green (K = 4) subpopulation includes the *Sal-I strain. Individuals displaying more than one color indicate mixed ancestry.
Structure analysis
The Evanno test for structure analysis based on pvdbpII suggested five distinct subpopulations in Latin America; however, the Delta K value was low (0.549). Population structure at K = 5 and K = 4 did not differ significantly (Figure 3B). Most admixture was observed in Brazil and Colombia. Nicaraguan subpopulations were shared with South American parasites, with the greatest difference observed against Mexico.
Genetic parameters of pvdbpII in Nicaragua, Latin America and globally
This analysis included 1,234 sequences covering 224 codons (739–1410 nucleotides) from 19 affected countries: 318 sequences from six regions in Latin America (LA; 25.8%), including Nicaragua; 137 from the Middle East (11.1%); 155 from South Asia (12.6%); 189 from Southeast Asia (15.3%); 135 from East Asia (10.9%); 210 from Africa (17%); and 88 from Papua New Guinea (PNG; 7.1%). A total of 133 polymorphic sites and 144 mutations were identified, of which 60 were singletons. Of the 244 haplotypes detected, 70 were present in two or more individuals, representing 85.9% of all sequences.
Nicaraguan parasites showed the second-lowest nucleotide diversity (π) in LA and the third-lowest globally. PvdbpII haplotype diversity in Nicaragua was lower than in Peru, Brazil, and Colombia but higher than in Mexico. Overall, genetic diversity in South America was higher than that in Mesoamerica. Globally, the highest nucleotide and haplotype diversity were observed in Thailand and South Korea, respectively (Supplementary Table 2).
Tajima’s D values varied widely across regions, ranging from −1.562 in PNG to 1.92 in India. Similarly, Fu and Li’s D* and F* values ranged from −5.57 to 1.74 and from −4.74 to 1.91 in PNG and Brazil, respectively. The Z test of selection also varied among parasite populations and was positive and significant in parasites from Nicaragua, Mexico, and Colombia, as well as in populations from the Middle East, Asia, Africa, and PNG (Supplementary Table 2).
The fixation index (FST) was estimated between parasite populations from different geographical regions. FST values between Nicaraguan and other LA parasites were low (0.032–0.114), except with Mexican parasites (0.247). Low FST values were also observed between parasites from Nicaragua and those from India (0.043) and Iran (0.064) (Supplementary Table 3).
Frequency of PvDBPII amino acid substitutions and haplotypes in Nicaragua, Latin America and globally
All mutations detected in Nicaraguan parasites were shared with other geographical regions (Table 1). The most frequent substitution in Nicaraguan parasites was I503K (n = 63; 49.2%), followed by D384G (42.9%). The simultaneous occurrence of four changes at N417K, L424I, W437R, and I503K was observed in 31.8% of isolates from Nicaragua. These were present in all Latin American (LA) regions and at frequencies ranging from 26% to 100% globally. In Nicaragua, mutations at K386N and K386Q occurred at similar frequencies (12.7% and 11.1%, respectively), but K386Q was infrequent globally (Table 1).
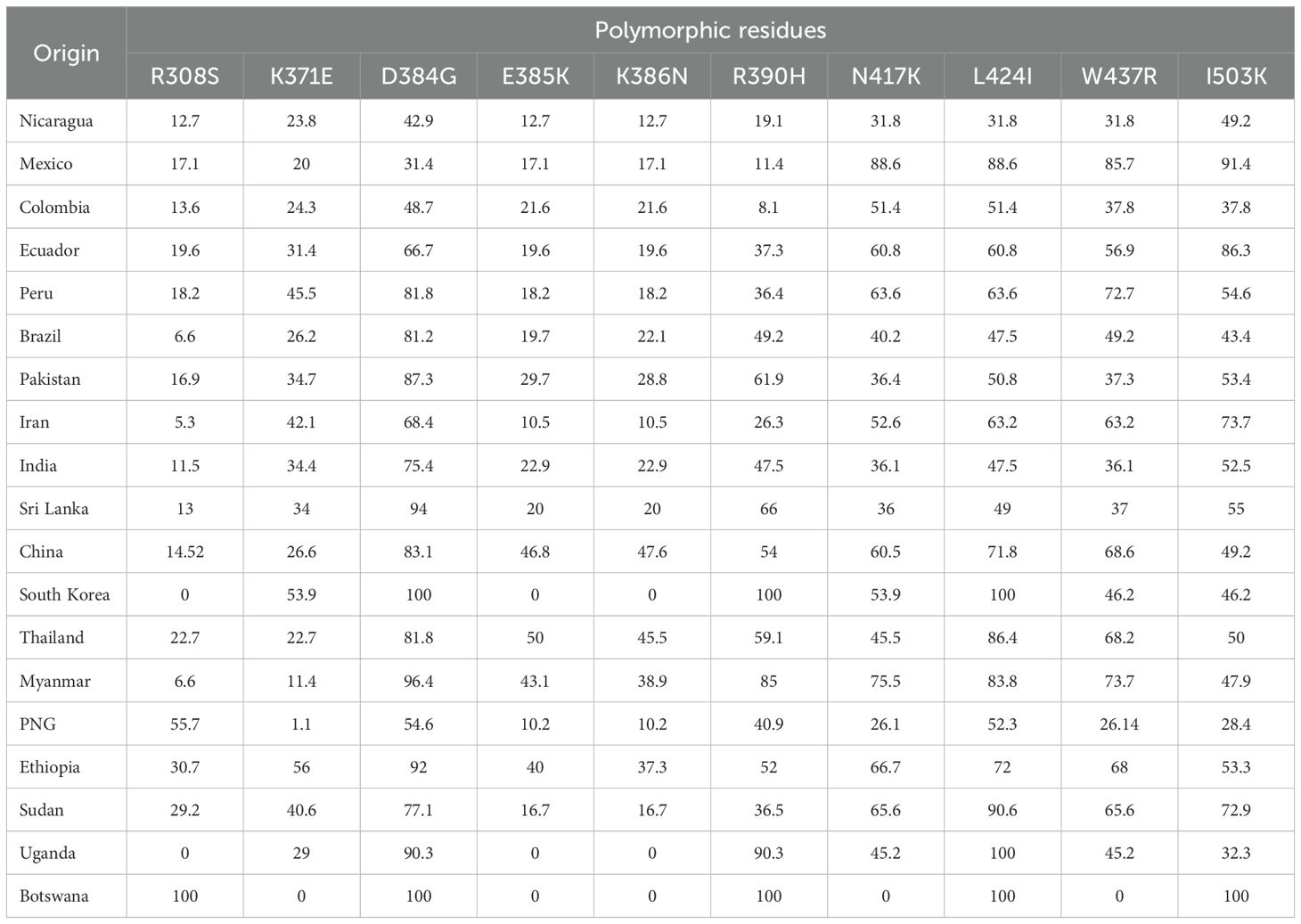
Table 1. Frequency (%) of polymorphisms in PvBDPII from Nicaragua compared with other geographical sites.
No exclusive PvDBPII haplotypes defined by 14 residues (R308S, L333F, K371E, N375D, D384G, E385K, K386N/Q, R390H, N417K, I419M, L424I, W437R, S447K, I503K) were detected in Nicaraguan parasites. Five of the six haplotypes corresponded to the 10 most frequent haplotypes estimated in our study (gh1–gh10), four of which occurred at >5% frequency (Table 2). Gh2 (Sal-I/Nh1), gh3 (Nh5), and gh5 (Nh3) were present in all LA regions, whereas gh7, gh8, gh9, and gh10 had the least distribution. Gh10 (Nh4) from Nicaragua was also observed in the Middle East and Africa. In LA, both Sal-I–related and divergent haplotypes were widely distributed (Table 2). Supplementary Table 4 shows the frequency of the most frequent haplotypes by country.
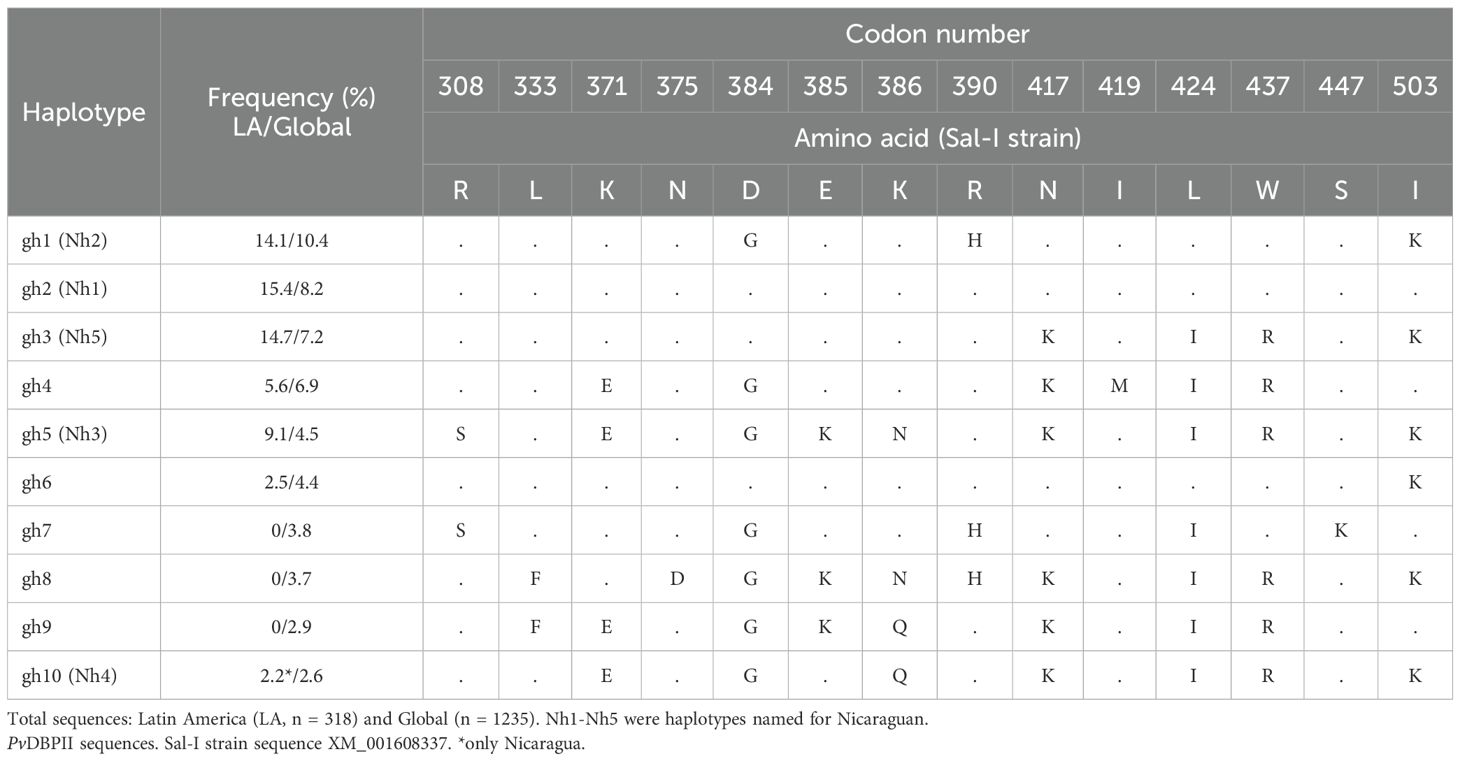
Table 2. P. vivax DBPII: most frequent global haplotypes.
The frequencies of PvDBPII phenotypes defined by combinations at residues N417K, W437R, and I503K—known to participate in antibody evasion (VanBuskirk et al., 2004) —were also estimated, regardless of other substitutions. Phenotypes corresponding to the Sal-I sequence (NWI) and NWK were present in all regions at varying frequencies. The divergent phenotype KRK was detected in 31.7% of Nicaraguan parasites and showed wide variation in LA and outside the continent (Table 3). Haplotypes with the NWI and KRK phenotypes were found in most regions, followed by NWK and KRI. While Nicaragua had the top three phenotypes globally, parasite populations from Myanmar and Sudan exhibited seven of the eight phenotypes (Table 3).
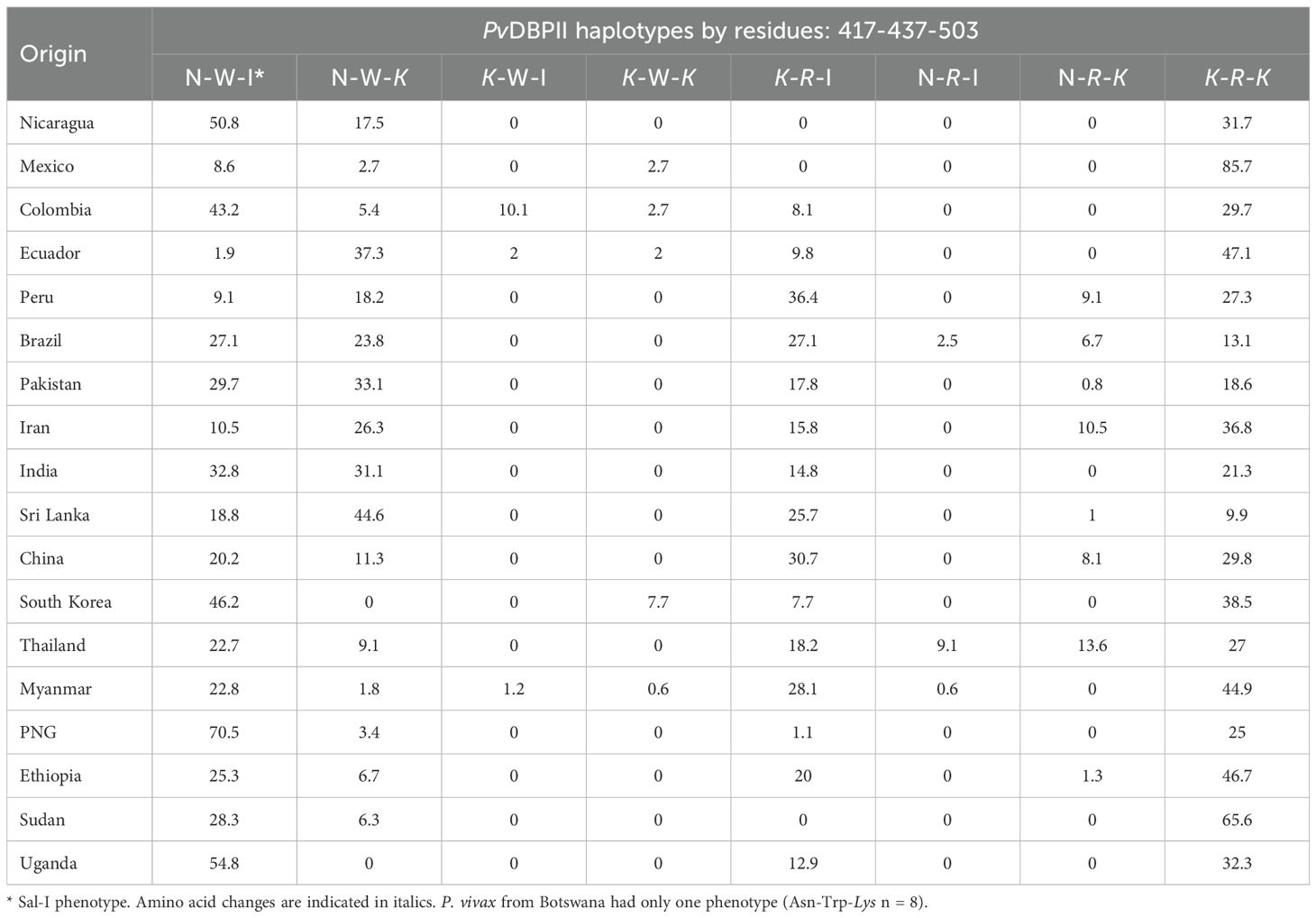
Table 3. Comparison of the frequencies (%) of P. vivax BDPII phenotypes in parasites from Nicaragua (2012-2013) and other geographical sites.
Geographic distribution of P. vivax haplotypes in Nicaragua municipalities
Most pvdbpII haplotypes were widely distributed in Nicaragua. Haplotype Nh1 (Sal-I) was common in RACCN and found in one isolate from the Pacific coast (Figure 4A). Haplotype Nh2 was the second most frequent (17.4%) in RACCN. Haplotypes Nh3–Nh5 were restricted to RACCN municipalities, while Nh4 predominated on the Pacific coast (Figure 4A). Of the six pvdbpII haplotypes, only two were exclusive to a single municipality.
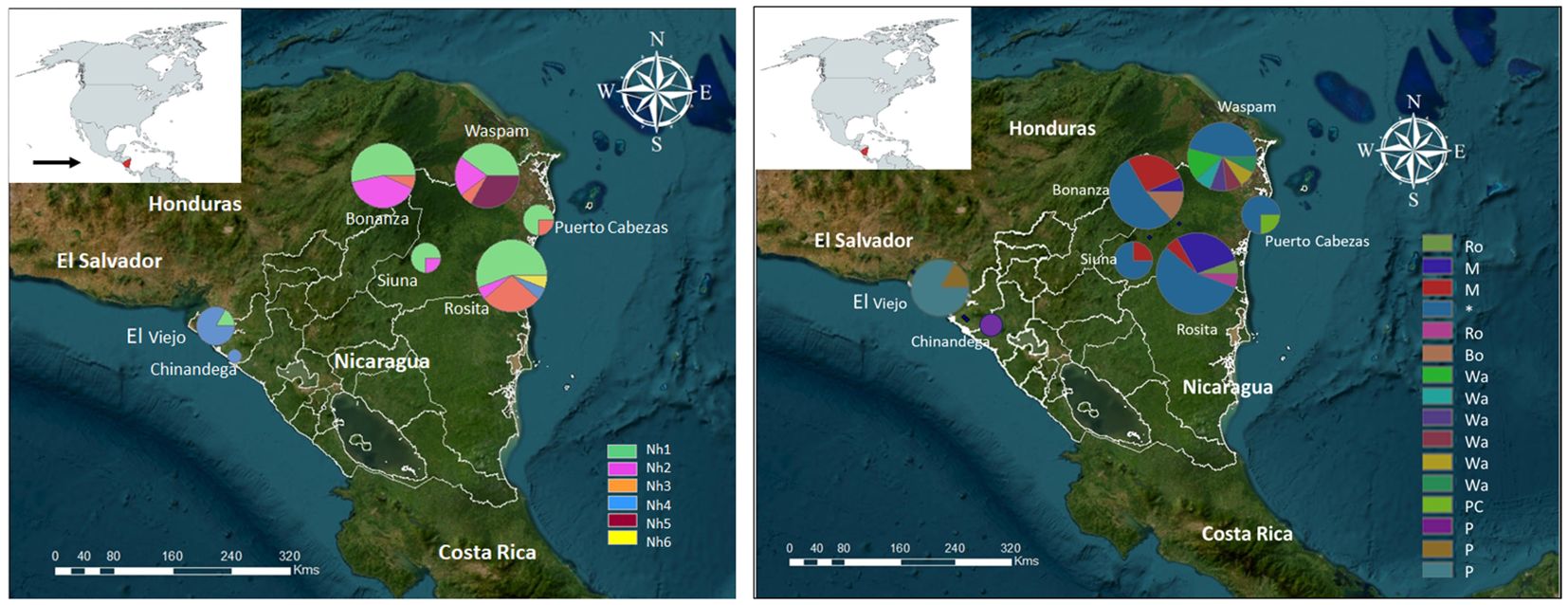
Figure 4. Geographic distribution of P. vivax haplotypes in Nicaragua, 2012–2013. (A) dbpII and (B) dbpII–ama1I-II haplotypes (H) were mapped using ArcGIS v10.8.1. The Sal-I haplotype*, and haplotypes exclusive to the Mining region (M), Pacific coast (P), RACCN (Ro, Rosita; Bo, Bonanza; Wa, Waspam; and PC, Puerto cabezas).
The concatenated sequence (pvdbpII–pvama1I-II) significantly increased resolution, yielding 16 haplotypes and identifying exclusive haplotypes for each region or municipality (Figure 4B). Notably, 30 of 31 parasites with pvdbpII Sal-I also exhibited pvama1 I-II Sal-I. The Mining region harbored two frequent and exclusive haplotypes, while Bonanza had one and Rosita had two. Waspam and Puerto Cabezas displayed six and one exclusive haplotypes, respectively. All three haplotypes detected on the Pacific coast were exclusive (Figure 4B).
A phylogenetic tree revealed several genetic groups that exhibited a degree of homogeneity when analyzed with either pvdbpII alone or the concatenated sequence (Figure 5).
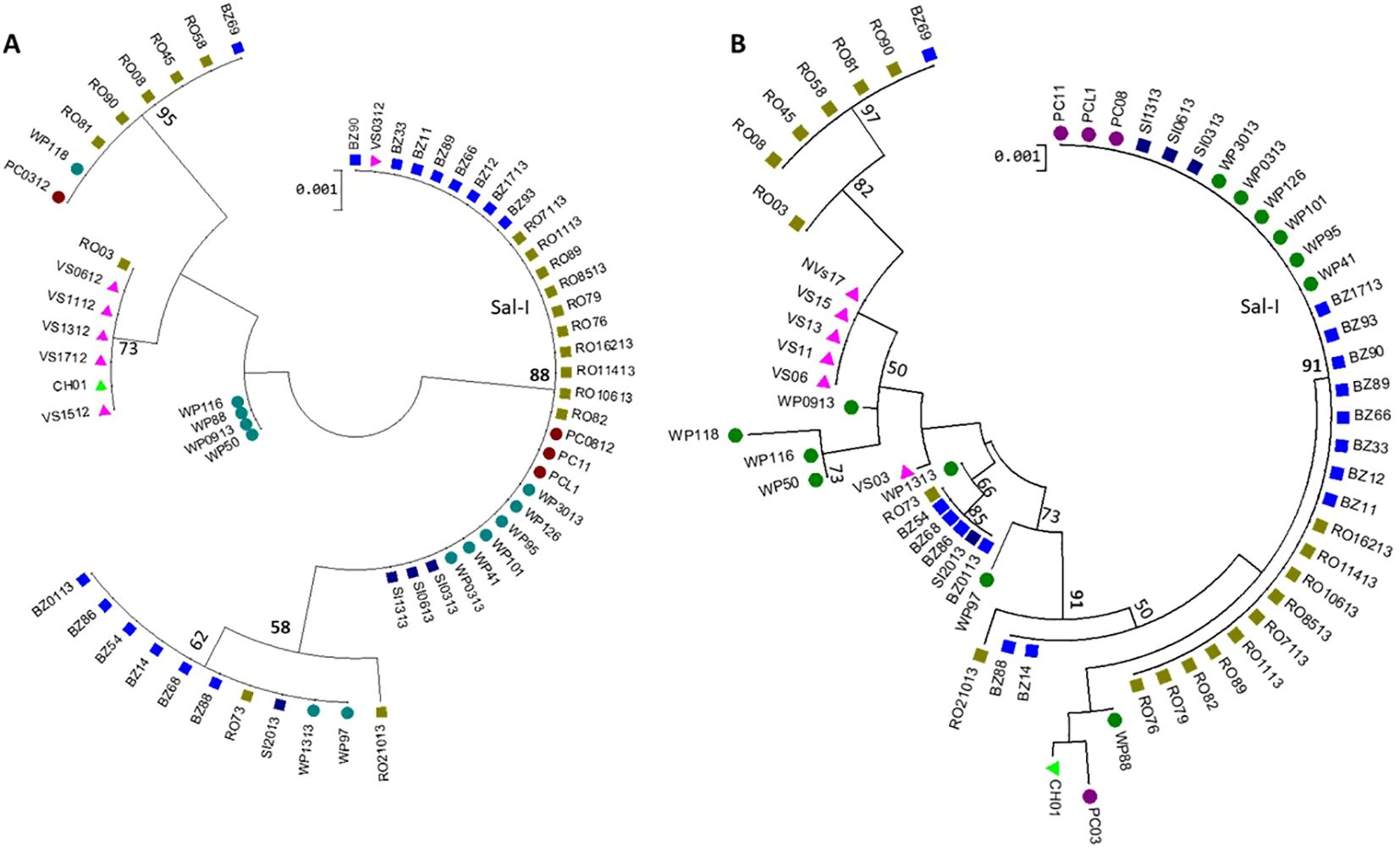
Figure 5. Maximum likelihood (ML) phylogenetic trees of 61 P. vivax isolates from Nicaragua, 2012-2013. (A) PvdbpII and (B) PvdbpII - pvama1I-II concatenated haplotypes (H) were analyzed in MEGA v11 using the HKY model and bootstrap method with 1,000 replications. Thirty of 31 pvdbpII Sal-I haplotypes also carried pvama1I-II Sal-I.
Discussion
The low P. vivax dbpII diversity found in Nicaragua aligns with a continuous decline in the number of P. vivax cases since 2001, including a significant drop between 2007 and 2012. This decline was likely intensified operational activities for malaria control in the country (Pan American Health Organization, 2020; World Health Organization, 2021). Low genetic diversity was also observed in pvama1I-II (Gonzalez Ceron et al., 2023), which is consistent with mitochondrial DNA analysis of Central American isolates compared with those from South America or outside the continent (Taylor et al., 2013).
The invasion process of P. vivax requires the interaction of the Duffy binding protein domain II (DBPII) and the DARC receptor on reticulocytes (Singh et al., 2005). This molecule shows polymorphism that might hinder its effectiveness as a vaccine candidate (VanBuskirk et al., 2004). PvdbpII from Nicaragua exhibited mutation and haplotype patterns similar to those found in Latin America and elsewhere (Sousa et al., 2011; Almeida-de-Oliveira et al., 2020). Haplotype network analysis, STRUCTURE analysis, and the FST index based on pvdbpII suggest limited genetic differentiation between Nicaraguan and other Latin American parasites, a finding echoed in recent studies of global parasites (Nguyễn et al., 2024). In contrast, analysis of pvmsp142, pvama1I-II, and pvmdr1 gene markers has shown a differentiated population in Nicaragua (Gutierrez et al., 2016; Gonzalez-Ceron et al., 2017; 2020). This is supported by the finding that mutations associated with pyrimethamine resistance in PvDHFR, which are common in South America, were not present in parasites from Nicaragua or Mexico collected the same period (Gonzalez-Ceron et al., 2020). The presence of similar mutations in pvdbpII globally might indicate convergent evolution, possibly similar to genes involved in drug resistance or other genes under selective pressure (Abera et al., 2021; Flannery et al., 2015).
The exclusive presence of nonsynonymous mutations and a positive Z test of selection in Nicaraguan parasites may indicate that this protein is under antibody pressure, consistent with findings from other geographic regions (Almeida-de-Oliveira et al., 2020; Cole-Tobian and King, 2003; Chen et al., 2016; Michon et al., 2000). Varying intensities of antibody selective pressure and/or genetic drift (VanBuskirk et al., 2004), might explain the variation in haplotype frequencies observed between Nicaragua and other geographical regions. Although PvDBPII is less diverse compared with other molecules more exposed to the immune system (Gonzalez-Ceron et al., 2015), a proportion of infected individuals develop variable antibody titers, which appear to increase with exposure (Zakeri et al., 2011; Nicolete et al., 2016). These antibodies inhibited the binding of P. vivax to human erythrocytes (Grimberg et al., 2007) and may be phenotype-transcending and long-lasting (King et al., 2008; Nicolete et al., 2016; Thawornpan et al., 2022).
In Plasmodium, amino acid substitutions in molecules involved in parasite development and in vaccine candidates frequently modify the protein’s antigenic profile, potentially allowing parasites to escape heterologous antibody responses. Experimental studies have shown that antibodies targeting PvDBPII Sal-I (N417-W437-I503) inhibit erythrocyte adhesion to COS cells expressing the homologous phenotype. However, naturally occurring mutants showed varying levels of inhibition, with NRK and KRK variants showing the strongest inhibition (VanBuskirk et al., 2004). Similar results were observed in P. vivax ex vivo short-term assays using heterologous monoclonal antibodies (Rawlinson et al., 2019). Remarkably, heterologous haplotypes carrying NWI/NWK versus KRK, NRK, and KRI were the most prevalent globally, including in Nicaragua.
Further experiments using a mouse model demonstrated the induction of cross-reactive inhibitory antibodies by different PvDBPII phenotypes prevalent in Thailand and the humoral responses were either phenotype-transcending or phenotype-specific (Thawornpan et al., 2022). Haplotypes with variations similar to global haplotypes gh8 and gh9 showed >50% inhibition of adhesion in vitro against all polymorphic haplotypes except the Sal-I phenotype. In contrast, one haplotype similar to the highly frequent gh1 induced strain-specific antibodies. Sera from P. vivax-infected individuals showed variable inhibition, blocking adhesion of between two and five PvDBPII phenotypes (Thawornpan et al., 2022). Conserved continuous or conformational B-cell epitopes may contribute to phenotype-transcending immunity (VanBuskirk et al., 2004; Sousa et al., 2011; Ntumngia et al., 2012; Chootong et al., 2014; Valizadeh et al., 2014a; Urusova et al., 2019; Rawlinson et al., 2019; Almeida-de-Oliveira et al., 2020; Valizadeh et al., 2014c).
Previous findings suggest that the overall pattern of polymorphism is more relevant than individual amino acid substitutions in generating phenotype-transcending humoral responses. However, the quality and quantity of antibody responses vary among individuals. In Nicaragua, two main PvDBPII phenotype groups were detected, representing highly frequent and widespread haplotypes globally: gh1, gh2, and gh6, which are related to Sal-I, and divergent haplotypes (gh3, gh4, and gh5) exhibiting variant phenotypes N417K-W437R-I503K or N417K-W437R. Both groups occurred at similar frequencies, with haplotype gh3 (Nh5) being the most widespread globally.
In Nicaragua, over half a million people are at risk of malaria, with P. vivax being the predominant species. In RACCN, where malaria transmission is most persistent, indigenous groups such as the Miskito population (Ministerio de salud, 2014), are particularly vulnerable due to natural disasters, climate change, economic instability, and limited access to health services (Ministerio de salud, 2014; Larrañaga et al., 2013; Soto et al., 2022). Notably, the RACCN accounted for 67.6% of all malaria cases in Nicaragua between 2013 and 2018. During this period, reported P. vivax cases in the region increased from 950 in 2013 to 14,501 in 2018, alongside a high rate of presumed relapse episodes (Soto et al., 2022). Puerto Cabezas was the most affected municipality. Significant population movement from areas such as Waspam to Puerto Cabezas may have contributed to the dispersal of P. vivax haplotypes, a factor that warrants further evaluation. The pvdbpII marker alone is insufficient for parasite surveillance, but it could be informative when used in combination with other markers.
Because sampling in Nicaragua was conducted after transmission had declined, the prevalent pvdbpII haplotypes in Nicaragua might correspond to those that were best adapted. These haplotypes would persist, but their frequencies could fluctuate—a phenomenon previously observed using temporal networks for pvama1I-II (Flores-Alanis et al., 2017) and pvmsp142 (Flores-Alanis et al., 2022) in southern Mexico.
More integrated and temporal studies could help explain how haplotypes (both Sal-I–related and divergent) interact and fluctuate in a population over time, and explore evidence of immune evasion in natural infections. In addition, studying histocompatibility leukocyte antigen (HLA) associations with homologous and heterologous antibody responses would be beneficial. For instance, in the Brazilian Amazon, antibody responses against the Sal-I haplotype were associated with specific HLA antigens (Kano et al., 2016). Moreover, some Nicaraguan haplotypes were among the most frequent globally, providing potential candidates for mono- or polyvalent vaccines beyond just the Sal-I haplotype.
Conclusions
P. vivax dbpII in Nicaragua showed low genetic diversity, and while its mutations and haplotypes were shared with other geographic areas both within Latin America and globally, their frequencies varied. Non-synonymous mutations predominated and were located at sites involved in B-cell epitopes under immunological selective pressure, consistent with findings in other regions. This information contributes to the development of PvBDPII-based vaccines, suggesting that the extent and type of polymorphism relevant for vaccine design may already have been identified. However, ongoing parasitological surveillance remains crucial to detect new variants that could potentially lead to immune evasion.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://www.ncbi.nlm.nih.gov/genbank/, accession numbers: ON730710-ON730774.
Ethics statement
The studies involving humans were approved by Ethic Committees of the National Center for Diagnosis and Reference (CNDR) of the Health Ministry in Nicaragua, and the National Institute of Public Health, Mexico (CI1042). The studies were conducted in accordance with the local legislation and institutional requirements. The human samples used in this study were acquired from primarily isolated as part of your previous study for which ethical approval was obtained. Written informed consent for participation was not required from the participants or the participants’ legal guardians/next of kin in accordance with the national legislation and institutional requirements.
Author contributions
RO-M: Methodology, Formal Analysis, Writing – original draft. HT-O: Methodology, Writing – original draft. FS-V: Methodology, Data curation, Formal Analysis, Writing – original draft. AM: Conceptualization, Writing – original draft, Funding acquisition, Resources, Methodology. LG-C: Writing – review & editing, Supervision, Writing – original draft, Formal Analysis, Investigation, Funding acquisition, Project administration, Conceptualization, Validation.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This work was supported by AMI/RAVEDRA-PAHO/WHO-Nicaragua and Global Fund-Malaria Component-Nicaragua, and CONACyT-Mexico: project CB-2009-01-131247.
Acknowledgments
To the personnel of the sentinel sites in Nicaragua for their commitment in sample collection, identification and storage.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmala.2025.1620450/full#supplementary-material
References
Abera D., Kibet C. K., Degefa T., Amenga-Etego L., Bargul J. L., and Golassa L. (2021). Genomic analysis reveals independent evolution of Plasmodium falciparum populations in Ethiopia. Malar. J. 20, 129. doi: 10.1186/s12936-021-03660-y
Adams J. H., Sim B. K., Dolan S. A., Fang X., Kaslow D. C., Miller L. H., et al. (1992). A family of erythrocyte binding proteins of malaria parasites. Proc. Natl. Acad. Sci. U. S. A. 89, 7085–7089. doi: 10.1073/pnas.89.15.7085
Almeida-de-Oliveira N. K., Lima-Cury L., de Abreu-Fernandes R., de Rosa Lavigne A., de Pina-Costa A., de Souza Perce-da-Silva D., et al. (2020). Extensive genetic diversity of Plasmodium vivax dbp-II in Rio de Janeiro Atlantic Forest and Brazilian Amazon Basin: evidence of positive selection. Malar. J. 19, 81. doi: 10.1186/s12936-020-03159-y
Ampudia E., Patarroyo M. A., Patarroyo M. E., and Murillo L. A. (1996). Genetic polymorphism of the Duffy receptor binding domain of Plasmodium vivax in Colombian wild isolates. Mol. Biochem. Parasitol. 78, 269–272. doi: 10.1016/s0166-6851(96)02611-4
Arnott A., Barry A. E., and Reeder J. C. (2012). Understanding the population genetics of Plasmodium vivax is essential for malaria control and elimination. Malar J. 11, 14. doi: 10.1186/1475-2875-11-14
Bandelt H. J., Forster P., and Rohl A. (1999). Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 16, 37–48. doi: 10.1093/oxfordjournals.molbev.a026036
Chen E., Salinas N. D., Huang Y., Ntumngia F., Plasencia M. D., Gross M. L., et al. (2016). Broadly neutralizing epitopes in the Plasmodium vivax vaccine candidate Duffy Binding Protein. Proc. Natl. Acad. Sci. U S A. 113 (22), 6277–6282. doi: 10.1073/pnas.1600488113
Chootong P., McHenry A. M., Ntumngia F. B., Sattabongkot J., and Adams J. H. (2014). The association of Duffy binding protein region II polymorphisms and its antigenicity in Plasmodium vivax isolates from Thailand. Parasitol. Int. 63, 858–864. doi: 10.1016/j.parint.2014.07.014
Cole-Tobian J. L., Cortés A., Baisor M., Kastens W., Xainli J., Bockarie M., et al. (2002). Age-acquired immunity to a plasmodium vivax invasion ligand, the duffy binding protein. J. Infect. Dis. 186, 531–539. doi: 10.1086/341776
Cole-Tobian J. and King C. L. (2003). Diversity and natural selection in Plasmodium vivax Duffy binding protein gene. Mol. Biochem. Parasitol. 127, 121–132. doi: 10.1016/s0166-6851(02)00327-4
Cole-Tobian J. L., Michon P., Dabod E., Mueller I., and King C. L. (2007). Dynamics of asymptomatic Plasmodium vivax infections and Duffy binding protein polymorphisms in relation to parasitemia levels in Papua New Guinean children. Am. J. Trop. Med. Hyg 77, 955–962. doi: 10.4269/ajtmh.2007.77.955
Earl D. and vonHoldt B. (2012). STRUCTURE HARVESTER: a website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 4, 359–361. doi: 10.1007/s12686-011-9548-7
Escalante A. A. and Pacheco M. A. (2019). Malaria molecular epidemiology: an evolutionary genetics perspective. Microbiol. Spectr. 7, 1–17. doi: 10.1128/microbiolspec.ame-0010-2019
Flannery E. L., Wang T., Akbari A., Corey V. C., Gunawan F., Bright A. T., et al. (2015). Next-generation sequencing of plasmodium vivax patient samples shows evidence of direct evolution in drug-resistance genes. ACS Infect. Dis. 1, 367–379. doi: 10.1021/acsinfecdis.5b00049
Flores-Alanis A., González-Cerón L., Santillán-Valenzuela F., Ximenez C., Sandoval-Bautista M. A., and Cerritos R. (2017). Temporal genetic changes in Plasmodium vivax apical membrane antigen 1 over 19 years of transmission in southern Mexico. Parasitol. Vectors 10, 217. doi: 10.1186/s13071-017-2156-y
Flores-Alanis A., Gonzalez-Ceron L., Santillan-Valenzuela F., Ximenez C., Sandoval-Bautista M. A., and Cerritos R. (2022). Spatiotemporal Changes in Plasmodium vivax msp142 Haplotypes in Southern Mexico: From the Control to the Pre-Elimination Phase. Microorganisms 10. doi: 10.3390/microorganisms10010186
Fu Y. X. and Li W. H. (1993). Statistical tests of neutrality of mutations. Genetics 133, 693–709. doi: 10.1093/genetics/133.3.693
Gonzalez-Ceron L., Cerritos R., Corzo-Mancilla J., and Santillan F. (2015). Diversity and evolutionary genetics of the three major Plasmodium vivax merozoite genes participating in reticulocyte invasion in southern Mexico. Parasitol. Vectors 8, 651. doi: 10.1186/s13071-015-1266-7
Gonzalez-Ceron L., Montoya A., Corzo-Gomez J. C., Cerritos R., Santillan F., and Sandoval M. A. (2017). Genetic diversity and natural selection of Plasmodium vivax multi-drug resistant gene (pvmdr1) in Mesoamerica. Malar. J. 16, 261. doi: 10.1186/s12936-017-1905-x
Gonzalez Ceron L., Piedra-Arevalo F. O., Casanova-Hernandez D., Santillan-Valenzuela F., and Montoya Perez A. (2023). Plasmodium vivax apical membrane antigen 1 I-II from Nicaragua, (2012-2013): genetic and antigenic polymorphism. Salud Publica Mex. 65, 127–135. doi: 10.21149/14039
Gonzalez-Ceron L., Rodriguez M. H., Montoya A., Santillan-Valenzuela F., and Corzo-Gomez J. C. (2020). Molecular variation of Plasmodium vivax dehydrofolate reductase in Mexico and Nicaragua contrasts with that occurring in South America. Salud Publica Mex. 62, 364–371. doi: 10.21149/10129
Gosi P., Khusmith S., Khalambaheti T., Lanar D. E., Schaecher K. E., Fukuda M. M., et al. (2008). Polymorphism patterns in Duffy-binding protein among Thai Plasmodium vivax isolates. Malar. J. 7, 112. doi: 10.1186/1475-2875-7-112
Grimberg B. T., Udomsangpetch R., Xainli J., McHenry A., Panichakul T., Sattabongkot J., et al. (2007). Plasmodium vivax invasion of human erythrocytes inhibited by antibodies directed against the Duffy binding protein. PloS Med. 4, e337. doi: 10.1371/journal.pmed.0040337
Gutierrez S., Gonzalez-Ceron L., Montoya A., Sandoval M. A., Torres M. E., and Cerritos R. (2016). Genetic structure of Plasmodium vivax in Nicaragua, a country in the control phase, based on the carboxyl terminal region of the merozoite surface protein-1. Infect. Genet. Evol. 40, 324–330. doi: 10.1016/j.meegid.2015.08.040
Hall T. A. (1999). BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT Nucl. Acids Symp. Ser. 41, 95–98.
Hoque M. R., Elfaki M. M. A., Ahmed M. A., Lee S.-K., Muh F., Ali Albsheer M. M., et al. (2018). Diversity pattern of Duffy binding protein sequence among Duffy-negatives and Duffy-positives in Sudan. Malar. J. 17, 1–10. doi: 10.1186/s12936-018-2425-z
Hu Y., Wang L., Mbenda H. G. N., Soe M. T., Yu C., Feng H., et al. (2019). Genetic diversity, natural selection and haplotype grouping of Plasmodium vivax Duffy-binding protein genes from eastern and western Myanmar borders. Parasitol. Vectors 12, 546. doi: 10.1186/s13071-019-3803-2
Hupalo D. N., Luo Z., Melnikov A., Sutton P. L., Rogov P., Escalante A., et al. (2016). Population genomics studies identify signatures of global dispersal and drug resistance in. Plasmodium vivax. Nat. Genet. 48, 953–958. doi: 10.1038/ng.3588
Ju H. L., Kang J. M., Moon S. U., Bahk Y. Y., Cho P. Y., Sohn W. M., et al. (2013). Genetic diversity and natural selection of Duffy binding protein of Plasmodium vivax Korean isolates. Acta Trop. 125, 67–74. doi: 10.1016/j.actatropica.2012.09.016
Kano F. S., Souza-Silva F. A., Torres L. M., Lima B. A., Sousa T. N., Alves J. R., et al. (2016). The presence, persistence and funcional properties of plasmodium vivax duffy binding protein II antibodies are influenced by HLA class II allelic variants. PloS Negl. Trop. Dis. 10, e0005177. doi: 10.1371/journal.pntd.0005177
King C. L., Michon P., Shakri A. R., Marcotty A., Stanisic D., Zimmerman P. A., et al. (2008). Naturally acquired Duffy-binding protein-specific binding inhibitory antibodies confer protection from blood-stage Plasmodium vivax infection. Proc. Natl. Acad. Sci. U.S.A. 105, 8363–8368. doi: 10.1073/pnas.0800371105
Larrañaga N., Mejía R. E., Hormaza J. I., Montoya A., Soto A., and Fontecha G. A. (2013). Genetic structure of Plasmodium falciparum populations across the Honduras-Nicaragua border. Malar. J. 12, 354. doi: 10.1186/1475-2875-12-354
Lo E., Russo G., Pestana K., Kepple D., Abagero B. R., Dongho G. B. D., et al. (2021). Contrasting epidemiology and genetic variation of Plasmodium vivax infecting Duffy-negative individuals across Africa. Int. J. Infect. Dis. 108, 63–71. doi: 10.1016/j.ijid.2021.05.009
Michon P. A., Arevalo-Herrera M., Fraser T., Herrera S., and Adams J. H. (1998). Serologic responses to recombinant Plasmodium vivax Duffy binding protein in a Colombian village. Am. J. Trop. Med. Hyg. 59, 597–599. doi: 10.4269/ajtmh.1998.59.597
Michon P. A. ,. P., Fraser T., and Adams J. H. (2000). Naturally acquired and vaccine-elicited antibodies block erythrocyte cytoadherence of the Plasmodium vivax Duffy binding protein. Infect. Immun. 68, 3164–3171. doi: 10.1128/IAI.68.6.3164-3171.2000
Ministerio de Salud. Esquema Nacional de Tratamiento de Malaria (2014). Available online at: https://www.paho.org/es/documentos/esquema-nacional-tratamiento-malaria-ministerio-salud-Nicaragua-2014 (Accessed 20 april 2025).
Nateghpour M., Haghi A. M., Naderi B., and Sepehrizadeh Z. (2017). Genetic polymorphism of Plasmodium vivax Duffy Binding Protein in malarious areas in southeastern of Iran. J. Parasitol. Dis. 41, 1132–1138. doi: 10.1007/s12639-017-0946-y
Nguyễn Đ. T. D., Võ T. C., Nguyễn K. O., Lê H. G., Kang J. M., and Nguyễn T. H. (2024). Genetic polymorphism of Duffy binding protein in Pakistan Plasmodium vivax isolates. Acta Trop. 260, 107421. doi: 10.1016/j.actatropica.2024.107421
Nicolete V. C., Frischmann S., Barbosa S., King C. L., and Ferreira M. U. (2016). Naturally acquired binding-inhibitory antibodies to plasmodium vivax duffy binding protein and clinical immunity to malaria in rural amazonians. J. Infect. Dis. 214, 1539–1546. doi: 10.1093/infdis/jiw407
Ntumngia F. B., Schloegel J., Barnes S. J., McHenry A. M., Singh S., King C. L., et al. (2012). Conserved and variant epitopes of Plasmodium vivax Duffy binding protein as targets of inhibitory monoclonal antibodies. Infect. Immun. 80 (3), 1203–8. doi: 10.1128/IAI.05924-11
Nunez A., Ntumngia F. B., Guerra Y., Adams J. H., and Saenz F. E. (2023). Genetic diversity and natural selection of Plasmodium vivax reticulocyte invasion genes in Ecuador. Malar. J. 22, 225. doi: 10.1186/s12936-023-04640-0
Pan American Health Organization (2020). Actualización Epidemiológica: Situación de la malaria en las Américas en el contexto de la pandemia de COVID-19. Available online at: https://iris.paho.org/handle/10665.2/52288 (Accessed 22 april 2025).
Porras-Hurtado L., Ruiz Y., Santos C., Phillips C., Carracedo A., and Lareu M. V. (2013). An overview of STRUCTURE: applications, parameter settings, and supporting software. Front. Genet. 4. doi: 10.3389/fgene.2013.00098
Premaratne P. H., Aravinda B. R., Escalante A. A., and Udagama P. V. (2011). Genetic diversity of Plasmodium vivax Duffy Binding Protein II (PvDBPII) under unstable transmission and low intensity malaria in Sri Lanka. Infect. Genet. Evol. 11, 1327–1339. doi: 10.1016/j.meegid.2011.04.023
Ranjan A. and Chitnis C. E. (1999). Mapping regions containing binding residues within functional domains of Plasmodium vivax and Plasmodium knowlesi erythrocyte-binding proteins. Proc. Natl. Acad. Sci. U.S.A. 96, 14067–14072. doi: 10.1073/pnas.96.24.14067
Rawlinson T. A., Barber N. M., Mohring F., Cho J. S., Kosaisavee V., Gérard S. F., et al. (2019). Structural basis for inhibition of Plasmodium vivax invasion by a broadly neutralizing vaccine-induced human antibody. Nat. Microbiol. 4, 1497–1507. doi: 10.1038/s41564-019-0462-1
Rozas J., Ferrer-Mata A., Sanchez-DelBarrio J. C., Guirao-Rico S., Librado P., Ramos-Onsins S. E., et al. (2017). DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 34, 3299–3302. doi: 10.1093/molbev/msx248
Shi T. Q., Shen H. M., Chen S. B., Kassegne K., Cui Y. B., Xu B., et al. (2021). Genetic diversity and natural selection of plasmodium vivax duffy binding protein-II from China-Myanmar border of yunnan province, China. Front. Microbiol. 12. doi: 10.3389/fmicb.2021.758061
Singh A. P., Ozwara H., Kocken C. H., Puri S. K., Thomas A. W., and Chitnis C. E. (2005). Targeted deletion of Plasmodium knowlesi Duffy binding protein confirms its role in junction formation during invasion. Mol. Microbiol. 55, 1925–1934. doi: 10.1111/j.1365-2958.2005.04523.x
Soto A. M., González-Cerón L., Santillán-Valenzuela F., Parrales M. E., and Montoya A. (2022). Recurrent plasmodium vivax cases of both short and long latency increased with transmission intensity and were distributed year-round in the most affected municipalities of the RACCN, Nicaragua 2013-2018. Int. J. Environ. Res. Public. Health 19, 6195. doi: 10.3390/ijerph19106195
Sousa T. N., Carvalho L. H., and Alves de Brito C. F. (2011). Worldwide genetic variability of the Duffy binding protein: insights into Plasmodium vivax vaccine development. PloS One 6, e22944. doi: 10.1371/journal.pone.0022944
Sousa T. N., Tarazona-Santos E. M., Wilson D. J., Madureira A. P., Falcao P. R., Fontes C. J., et al. (2010). Genetic variability and natural selection at the ligand domain of the Duffy binding protein in Brazilian Plasmodium vivax populations. Malar. J. 9, 334. doi: 10.1186/1475-2875-9-334
Tajima F. (1989). Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 123, 585–595. doi: 10.1093/genetics/123.3.585
Tamura K., Stecher G., and Kumar S. (2021). MEGA 11: molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 38, 3022–3027. doi: 10.1093/molbev/msab120
Taylor J. E., Pacheco M. A., Bacon D. J., Beg M. A., Dantas MaChado R. L., Fairhurst R. M., et al. (2013). The evolutionary history of Plasmodium vivax as inferred from mitochondrial genomes: parasite genetic diversity in the Americas. Mol. Biol. Evol. 30, 2050–2064. doi: 10.1093/molbev/mst104
Thawornpan P., Changrob S., Kochayoo P., Wangriatisak K., Ntumngia F. B., De S. L., et al. (2022). Cross-reactive inhibitory antibody and memory B cell responses to variant strains of Duffy binding protein II at post-Plasmodium vivax infection. PloS One 17, e0276335. doi: 10.1371/journal.pone.0276335
Tsuboi T., Kappe S. H., al-Yaman F., Prickett M. D., Alpers M., and Adams J. H. (1994). Natural variation within the principal adhesion domain of the Plasmodium vivax duffy binding protein. Infect. Immun. 62, 5581–5586. doi: 10.1128/iai.62.12.5581-5586.1994
Urusova D., Carias L., Huang Y., Nicolete V. C., Popovici J., and Roesch C. (2019). Structural basis for neutralization of Plasmodium vivax by naturally acquired human antibodies that target DBP. Nat. Microbiol. 4, 2024. doi: 10.1038/s41564-019-0587-2
Valizadeh V., Zakeri S., Mehrizi A. A., and Djadid N. D. (2014a). Population genetics and natural selection in the gene encoding the Duffy binding protein II in Iranian Plasmodium vivax wild isolates. Infect. Genet. Evol. 21, 424–435. doi: 10.1016/j.meegid.2013.12.012
Valizadeh V., Zakeri S., Mehrizi A. A., and Djadid N. D. (2014b). Non-allele specific antibody responses to genetically distinct variant forms of Plasmodium vivax Duffy binding protein (PvDBP-II) in Iranians exposed to seasonal malaria transmission. Acta Trop. 136, 89–100. doi: 10.1016/j.actatropica.2014.03.019
Valizadeh V., Zakeri S., Mehrizi A. A., and Djadid N. D. (2014c). Is the single variant form of Plasmodium vivax Duffy binding protein-II (PvDBP-II) adequate for inclusion in a PvDBP-II-based vaccine? Mal. J. 13, 95. doi: 10.1186/1475-2875-13-s1-p95
VanBuskirk K. M., Cole-Tobian J. L., Baisor M., Sevova E. S., Bockarie M., King C.L., et al (2004). Antigenic drift in the ligand domain of Plasmodium vivax duffy binding protein confers resistance to inhibitory antibodies. J. Infect. Dis. 190, 1556–1562. doi: 10.1086/424852
World Health Organization (2021). World malaria report. Available online at: https://www.who.int/publications/i/item/9789240040496 (Accessed April 22, 2025).
World Health Organization (2023). World malaria report. Available online at: https://www.who.int/publications/i/item/9789240086173 (Accessed April 22, 2025).
World Health Organization (2024). World malaria report. Available online at: https://www.who.int/publications/i/item/9789240104440 (Accessed April 22, 2025).
Wright S. (1951). The genetical structure of populations. Ann. Eugenics 15, 323–354. doi: 10.1111/j.1469-1809.1949.tb02451.x
Xainli J., Cole-Tobian J. L., Baisor M., Kastens W., Bockarie M., Yazdani S. S., et al. (2003). Epitope-specific humoral immunity to Plasmodium vivax Duffy binding protein. Infect. Immun. 71, 2508–2515. doi: 10.1128/IAI.71.5.2508-2515.2003
Zakeri S., Babaeekhou L., Mehrizi A. A., Abbasi M., and Djadid N. D. (2011). Antibody responses and avidity of naturally acquired anti-Plasmodium vivax Duffy binding protein (PvDBP) antibodies in individuals from an area with unstable malaria transmission. Am. J. Trop. Med. Hyg. 84, 944–950. doi: 10.4269/ajtmh.2011.11-0001
Keywords: Plasmodium vivax, Nicaragua, PvDBPII, polymorphism, nucleotide diversity
Citation: Ortega-Monroy R, Tomasini-Ovilla HA, Santillan-Valenzuela F, Montoya A and Gonzalez-Ceron L (2025) Molecular polymorphism of Plasmodium vivax Duffy binding protein domain II from Nicaragua, and global diversity patterns. Front. Malar. 3:1620450. doi: 10.3389/fmala.2025.1620450
Received: 29 April 2025; Accepted: 13 August 2025;
Published: 16 September 2025.
Edited by:
Alena Pance, University of Hertfordshire, United KingdomReviewed by:
Fauzi Muh, Diponegoro University, IndonesiaJin-Hee Han, Kangwon National University College of Medicine, Republic of Korea
Sonalika Kar, National Institute of Malaria Research (ICMR), India
Copyright © 2025 Ortega-Monroy, Tomasini-Ovilla, Santillan-Valenzuela, Montoya and Gonzalez-Ceron. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lilia Gonzalez-Ceron, bGdvbnphbEBpbnNwLm14
†Present addresses: René Ortega-Monroy, Departamento de Epidemiología. Hospital General de Zona 67, IMSS. Ciudad Apodaca, Mexico
Hugo A Tomasini-Ovilla, Laboratorio Clínico, Hospital de pediatría de CMN Siglo XXI, Ciudad de México, Mexico