 Patricia Parreira Perin1
Patricia Parreira Perin1 Caroline Leuchtenberger2,3,4
Caroline Leuchtenberger2,3,4 Mariana Malzoni Furtado2,5
Mariana Malzoni Furtado2,5 Livia de Almeida Rodrigues2,3
Livia de Almeida Rodrigues2,3 Grazielle Soresini2,3
Grazielle Soresini2,3 Nathalie Foerster2,3
Nathalie Foerster2,3 Guilherme Mourão6,7Wilson Junior Oliveira1Talita Oliveira Mendonça1Carmen Andrea Arias-Pacheco1Fabiana Alves Loureiro1
Guilherme Mourão6,7Wilson Junior Oliveira1Talita Oliveira Mendonça1Carmen Andrea Arias-Pacheco1Fabiana Alves Loureiro1 Estevam G. Lux Hoppe1*
Estevam G. Lux Hoppe1*- 1Parasitic Diseases and Zoonoses Laboratory (LabEPar), Department of Pathology, Reproduction and One Health (DPRSU), School of Agrarian and Veterinarian Sciences (FCAV), São Paulo State University (UNESP), Jaboticabal, Brazil
- 2Giant Otter Conservation Fund, Arroio do Meio, Brazil
- 3International Union for Conservation of Nature (IUCN) Species Survival Commission, Otter Specialist Group, Gland, Switzerland
- 4Federal Institute Farroupilha (IFFAR), Santa Maria, Brazil
- 5National Institute of Atlantic Forest, Santa Teresa, Brazil
- 6Federal University of Mato Grosso do Sul, Graduate Program in Ecology and Conservation, Institute of Biosciences, Campo Grande, Brazil
- 7Wildlife Laboratory, Embrapa Pantanal, Corumbá, Brazil
Introduction: Otters are top predators in aquatic ecosystems and serve as indicators of environmental health. However, knowledge of the parasitology of South American species remains limited. This study investigated the endoparasitic fauna of two otter species in Brazil, the giant otter (Pteronura brasiliensis) and the Neotropical otter (Lontra longicaudis), to better understand their parasite diversity and the ecological interactions between hosts and parasites.
Methods: Between February 2020 and November 2021, a total of 42 fecal samples and four carcasses (three P. brasiliensis and one L. longicaudis) were collected in the Pantanal Ecoregion. Coproparasitological analyses were performed to detect parasite eggs and oocysts, while necropsies allowed for the recovery and identification of helminth specimens. Molecular sequencing of 18S and 28S rDNA was conducted for selected taxa, with sequences deposited in GenBank.
Results: Coproparasitological analysis revealed the presence of parasites in 81.6% of P. brasiliensis fecal samples, with digeneans, strongylid-type, and ascarid eggs identified. Cystoisospora sp. oocysts and strongylid-type eggs were detected in the L. longicaudis fecal samples. Necropsies recovered specimens of digeneans and cestodes, including Alaria clathrata, Cryptocotyle thapari, and Spirometra sp. spargana, with the first 18S and 28S rDNA sequences for A. clathrata and C. thapari deposited in GenBank.
Discussion: These findings advance our understanding of otter–parasite dynamics in Neotropical wetlands and highlight the value of parasitological monitoring as part of conservation strategies for threatened wildlife.
1 Introduction
Otters are semiaquatic carnivores of the Mustelidae family that act as top predators in aquatic systems and play an important role in maintaining ecosystem balance (Groenendijk et al., 2023). In Brazil, the two species coexist in some areas: the giant otter (Pteronura brasiliensis) and the Neotropical otter (Lontra longicaudis).
Giant otters, among the largest carnivores in South America, are social animals globally classified as Endangered by the IUCN (Groenendijk et al., 2023), whereas the solitary and smaller Neotropical otter is considered Near Threatened (Rheingantz et al., 2017). The giant otter’s geographic distribution was significantly reduced in the past due to hunting driven by the pelt trade, and many populations have not recovered yet. In contrast, Neotropical otters are found widely across Brazil, inhabiting most regions of the country with suitable freshwater bodies. Currently, both species face threats from habitat loss and degradation, water contamination, human conflict, and climate change (Leuchtenberger et al., 2018; Rheingantz et al., 2022).
There are reports of infectious diseases affecting otters around the world, but this information is incomplete and fragmented, and the effects of these pathogens on otter populations are poorly understood. The presence of parasites in wildlife populations does not necessarily indicate disease or poor health, as some parasites can serve as indicators of biodiversity in healthy ecosystems (Hudson et al., 2006; Colwell et al., 2008; Lymbery et al., 2010). However, caution is needed when interpreting parasitological data from declining or threatened populations, such as giant otters. Information on the helminth fauna of Brazilian otters includes parasitism by digeneans (Alaria clathrata, Alaria pseudoclathrata, Baschkirovitrema incrassatum, Cryptocotyle thapari, Diplostomum alarioides, and Paragonimus rudis) and nematodes (Dirofilaria sp., Dirofilaria spectans, Galeiceps longispiculum, Molineus major, Subulura amazonica, and Subulura interrogans) in giant otters (Vieira et al., 2008; Muniz-Pereira et al., 2009). Additionally, there are records of Dioctophyme renale, Dioctophyme sp., Dirofilaria sp., Dirofilaria spectans, Dracunculus sp., oocysts of Eimeria spp., and eggs from Hymenolepis spp., Strongyloides spp., Ancylostomatidae, and Toxocara spp. in Neotropical otters (Vieira et al., 2008; Echenique et al., 2018; Uchôa et al., 2004).
Parasitological studies are valuable tools for addressing broader conservation threats, as parasites provide key indicators of host population dynamics, habitat quality, and anthropogenic pressures (Smith et al., 2009; Thompson et al., 2010; Gagne et al., 2022). Given the conservation challenges faced by both P. brasiliensis and L. longicaudis, understanding their parasitic fauna is essential not only for managing their health but also for gaining insight into their ecology and the environmental conditions they inhabit. Many of the parasites known to infect these animals have complex life cycles that depend on intermediate and/or paratenic hosts. In this study, we describe the parasites found in these two species through coproparasitological analysis and necropsy of otter carcasses found in nature, providing insights into their health status and ecological interactions.
2 Materials and methods
2.1 Study area
The field activities and sample collections were approved by the Instituto Chico Mendes de Conservação da Biodiversidade (ICMBio) (permit numbers 79173 and 85851). This study was conducted in the Pantanal Ecoregion in Brazil. Samples were collected from four locations: Aquidauana (-19,57845; -56,15308), Corumbá (-19,57964; -57,01819), and Miranda (-19,55746; -57,04869) in Mato Grosso do Sul state (MS), and Poconé (-17,43411; -56,82053), in Mato Grosso state (MT) (Figure 1).
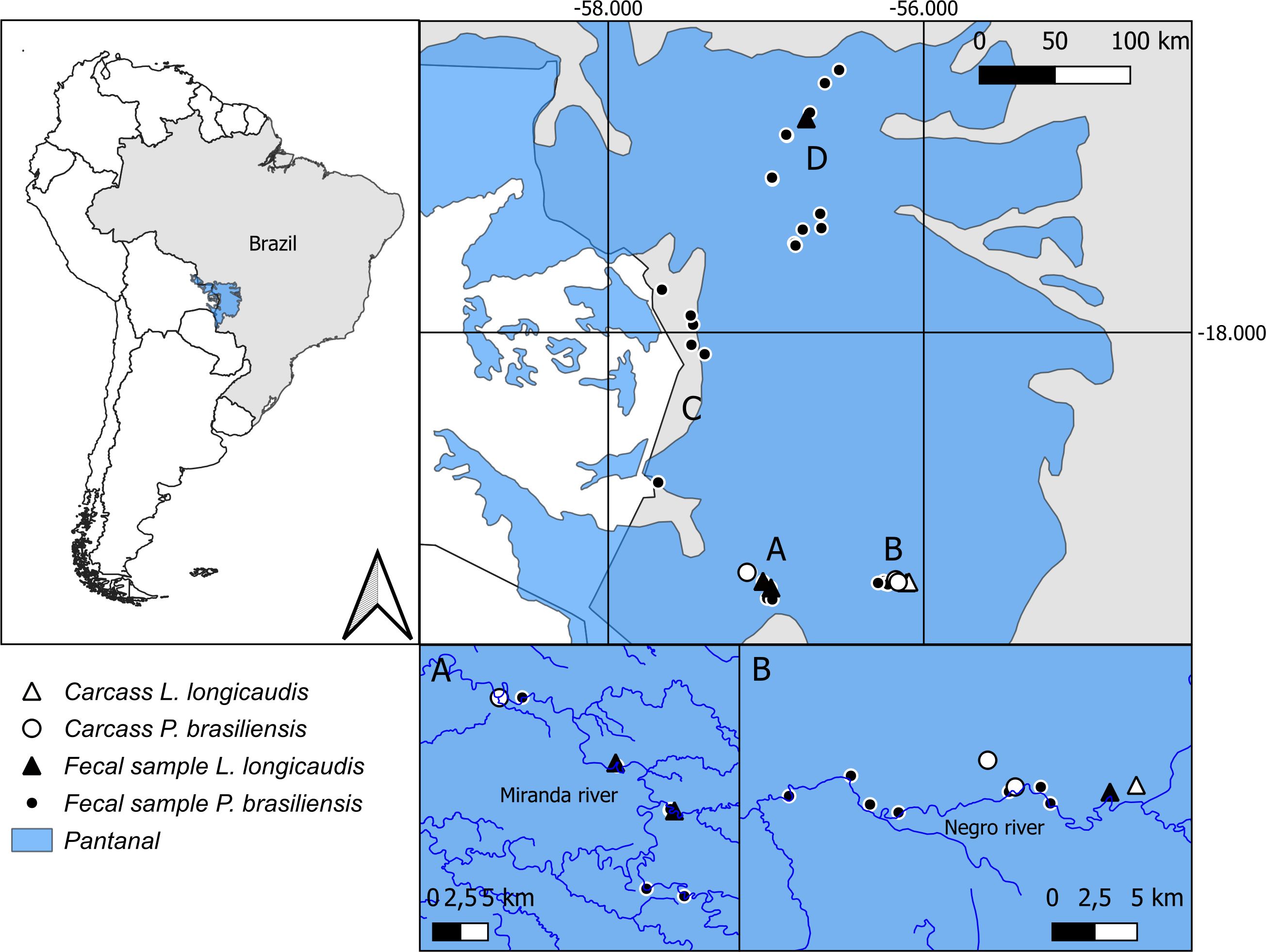
Figure 1. Study site, presenting the location of fecal samples and carcasses of Pteronura brasiliensis and Lontra longicaudis collected from February 2020 to November 2021 in the southern [(A) Aquidauana; (B) Miranda; (C) Corumbá] and northern [(D) Poconé] Brazilian Pantanal.
The Pantanal is the largest Neotropical wetland, covering 179,300 km², spanning Brazil (78%), Bolivia (18%), and Paraguay (4%) (Tomas et al., 2019). Although cattle ranching remains the primary economic activity in this wetland, ecotourism and agriculture are expanding, thereby transforming the landscape and altering human–wildlife dynamics. The Pantanal harbors significant populations of giant otters, while there are still data gaps for Neotropical otters, which seem to be widely distributed (Leuchtenberger et al., 2018; Rheingantz et al., 2022).
2.2 Parasitological assessment
A total of 38 fecal samples from 22 groups and one solitary giant otter (P. brasiliensis), as well as four fecal samples from L. longicaudis individuals, were collected in the Pantanal Ecoregion between February 2020 and November 2021. The feces were found during river-field expeditions, either through direct observation of otters using latrines or indirectly by detecting the characteristic odor of P. brasiliensis excreta. Only fresh samples were collected. Each was stored in labeled Falcon tubes containing 10% buffered formaldehyde solution (pH 7.4) and kept at room temperature until shipment to the laboratory. The presence of endoparasites was assessed using the Willis–Mollay flotation and Watanabe et al. sedimentation techniques (Zajac et al., 2021). Photomicrographs and measurements of oocysts and eggs were obtained using an Olympus BX-51 microscope equipped with an Olympus QColor3 digital camera (Olympus America Inc., Center Valley, PA, USA) and processed with Image-Pro Plus software (Media Cybernetics Inc., Bethesda, MD, USA). The measurements are noted in micrometers, expressed as arithmetic mean ± standard deviation. Representative images of the diagnosed oocysts and eggs were organized in a reference plate. Prevalence data were expressed as percentages, and 95% confidence intervals (CIs) for P. brasiliensis were calculated using the Wilson score method without continuity correction. For L. longicaudis, confidence intervals were not calculated due to the small sample size.
Carcasses of three giant otters, named PB1, PB2, and PB3, and one Neotropical otter, named LL1, were opportunistically collected during field monitoring activities in the Pantanal. Necropsies for PB1 and PB3 were performed on the same day they were found, while the carcasses of PB2 and LL1 were frozen at -25°C until processing. The organs were removed en bloc, dissected, and carefully inspected for the presence of macroparasites. The gastrointestinal tract was divided into anatomical segments (stomach, small intestine, and large intestine) and sectioned longitudinally. The contents and mucosa of the segments were washed with running water over 100 µm wire mesh sieves. The material retained in the sieves, along with helminths found during the macroscopic examination, was fixed and conserved in 70% ethanol in individual Falcon tubes. The contents of these tubes were carefully observed at a Leica EZ4 HD stereomicroscope (Leica Microsystems Inc., Buffalo Grove, IL, USA) for separation of parasites.
For taxonomic identification, 10 adult specimens of each digenean species were clarified in 80% acetic acid solution and then diaphanized in creosote. Since there were specimens at various degrees of development, we only included those with a uterus full of eggs in the analysis to avoid underdeveloped parasites. The specimens were mounted on temporary slides for observation under an Olympus BX-51 microscope equipped with an Olympus QColor3 digital camera (Olympus America Inc., Center Valley, PA, USA). Photomicrographs were captured and processed using Image-Pro Plus software (Media Cybernetics Inc., Bethesda, MD, USA). The total length of the metacestodes was measured macroscopically with a measuring tape. Measurements were recorded in millimeters, unless otherwise stated, expressed as arithmetic mean ± standard deviation. The identification of the helminths was based on the taxonomic keys proposed by Yamaguti (1958) and Travassos et al. (1969), with species redescriptions used when necessary (Gardner and Thew, 2006). All samples were processed in the Parasitic Diseases and Zoonoses Laboratory (LabEPar, FCAV, Unesp), and voucher specimens of each species were deposited in the LabEPar collection.
2.3 Molecular assessment
Two adult specimens of each digenean were preserved in absolute ethanol (Merk, Darmstadt, Germany) and stored at −20 °C until processing. The specimens selected for DNA extraction were individually transferred to microtubes and washed with sterile 1× PBS solution (pH 7.4). DNA extraction was performed using the DNeasy Blood & Tissue kit (Qiagen, Hilden, Germany) according to the manufacturer’s protocol. PCR was used to amplify the complete 18S rDNA and the D1–D3 region of 28S rDNA, using the primer sets and melting temperatures listed in Table 1. The preparation of the reaction mixes and the amplification cycles, conducted on a Nexus thermal cycler (Eppendorf, Hamburg, Germany), followed the methodology described by Perin et al. (2023).
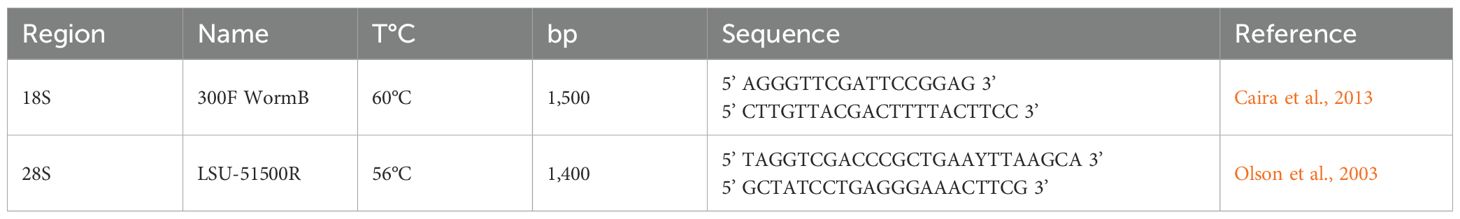
Table 1. Primer sets used in the PCR reactions.
The PCR products were run on 1% agarose gel electrophoresis to verify the amplification. Subsequently, these products were purified using the Wizard® SV Gel and PCR Clean-Up System kit (Promega, Madison, WI, USA). The purified PCR products were sequenced using the BigDye Terminator v3.1 kit (Applied Biosystems, Waltham, MA, USA). Sequencing was performed by capillary electrophoresis on an ABI3130 sequencer (Applied Biosystems), following the Sanger method (Sanger et al., 1977).
The electropherograms obtained from sequencing were processed using the Phred/Phrap/Consed software suite (Green, 1996; Ewing and Green, 1998; Gordon et al., 1998) to assess base quality and trim sequences, retaining only bases with a Phred score of 20 or higher. The resulting sequences were compared with those in the NCBI (National Center for Biotechnology Information) database using the BLAST tool (Altschul et al., 1990).
3 Results
In L. longicaudis, three parasite taxa were identified: Alaria clathrata, Cystoisospora sp., and strongylid-type eggs. In P. brasiliensis, six parasite taxa were recorded: three species identified from carcasses (A. clathrata, Cryptocotyle thapari, and Spirometra sp. spargana) and three morphotypes detected in feces (digenean eggs, strongylid-type eggs, and ascarid-type eggs).
Two of the four L. longicaudis fecal samples tested positive for parasitic infections: one for strongylid-type eggs and the other for Cystoisospora sp. In total, 31 of 38 fecal samples from P. brasiliensis (81.6%, 95% CI: 58.7 – 85.1%) tested positive for at least one parasite. The analysis of P. brasiliensis feces revealed the presence of digenean eggs in 28 out of 38 samples. Additionally, one P. brasiliensis sample showed co-infection with digeneans and strongylid-type eggs, another with digeneans and ascarids, and one sample was positive for digeneans, strongylid-type, and ascarids.
The digenean eggs (n=28) measured 34.5 ± 3.5 µm in length and 21.6 ± 1.1 µm in width. Ascarid eggs (n=8) were 60 ± 13 µm long and 46.2 ± 10 µm wide, and strongylid-type eggs (n=9) were 76.6 ± 20 µm long and 43.3 ± 10 µm wide. The Cystoisospora sp. oocysts (n=10) measured 21.8 ± 3.0 µm in length and 20.1 ± 2.2 µm in width. The oocysts and helminth eggs observed are illustrated in Figure 2.
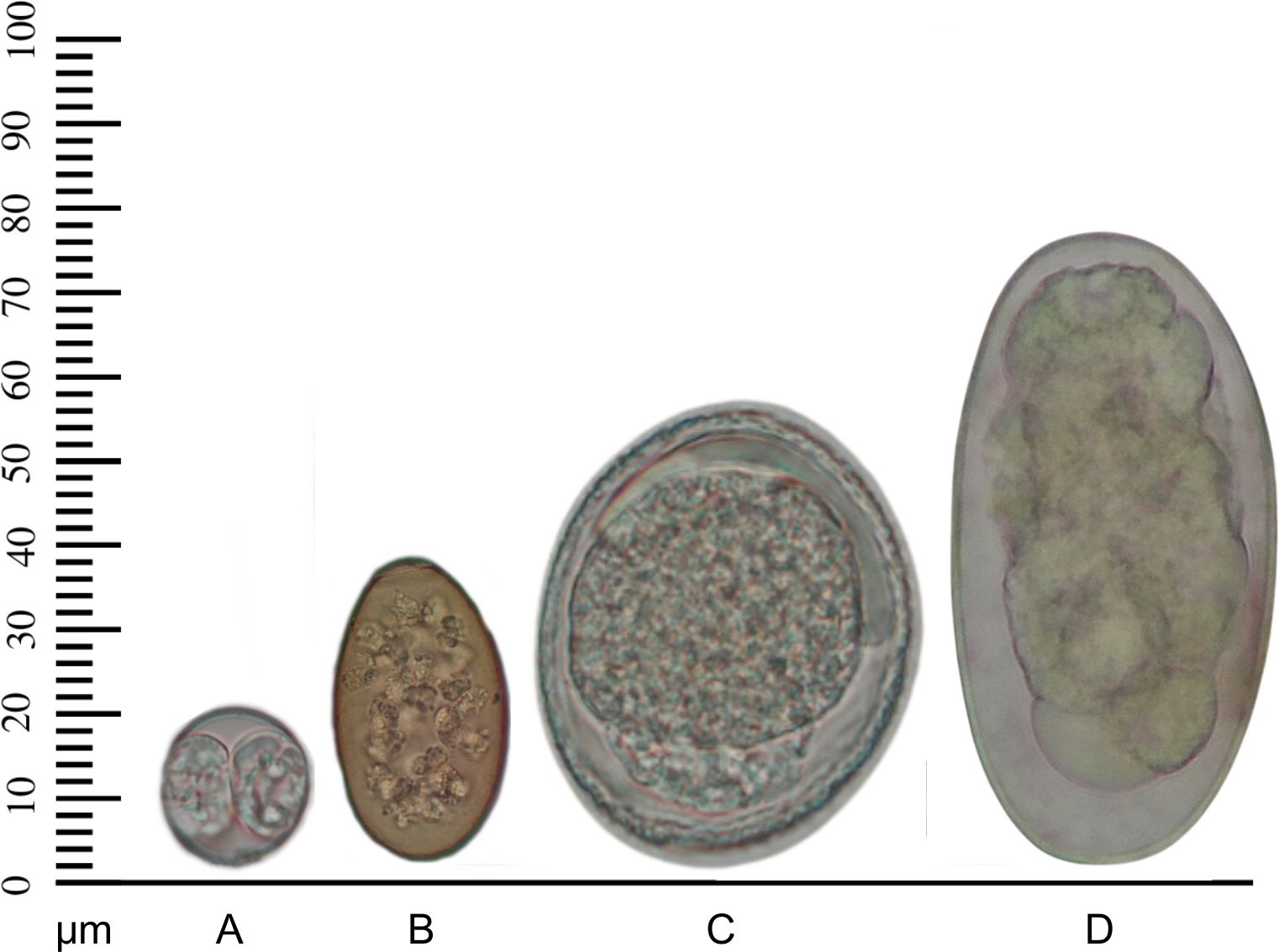
Figure 2. Oocyst and helminth egg morphotypes found in fecal samples from Brazilian semiaquatic mustelids (Pantanal, February 2020 – November 2021). (A) Cystoisospora sp. oocyst observed in Lontra longicaudis. (B) Digenean egg, (C) Ascarid egg, (D) Strongylid egg, all observed in Pteronura brasiliensis.
Helminths were recovered from all the carcasses (PB1, PB2, PB3, and LL1) and are described in Table 2. PB1 also had a cutaneous lesion measuring 8.5 × 6 cm in the frontoparietal region, with numerous open wounds at the edges, from which larvae of Cochliomyia hominivorax were recovered (Foerster et al., 2022). Three ticks were recovered from PB2, of which two were adults of Amblyomma sculptum and one nymph of Amblyomma spp (Soresini et al., 2023). Two ticks were recovered from PB3: a second-instar nymph of Ornithodoros rostratus (Argasidae) and a male of Amblyomma sculptum (Ixodidae) (Barros-Battesti et al., 2023).
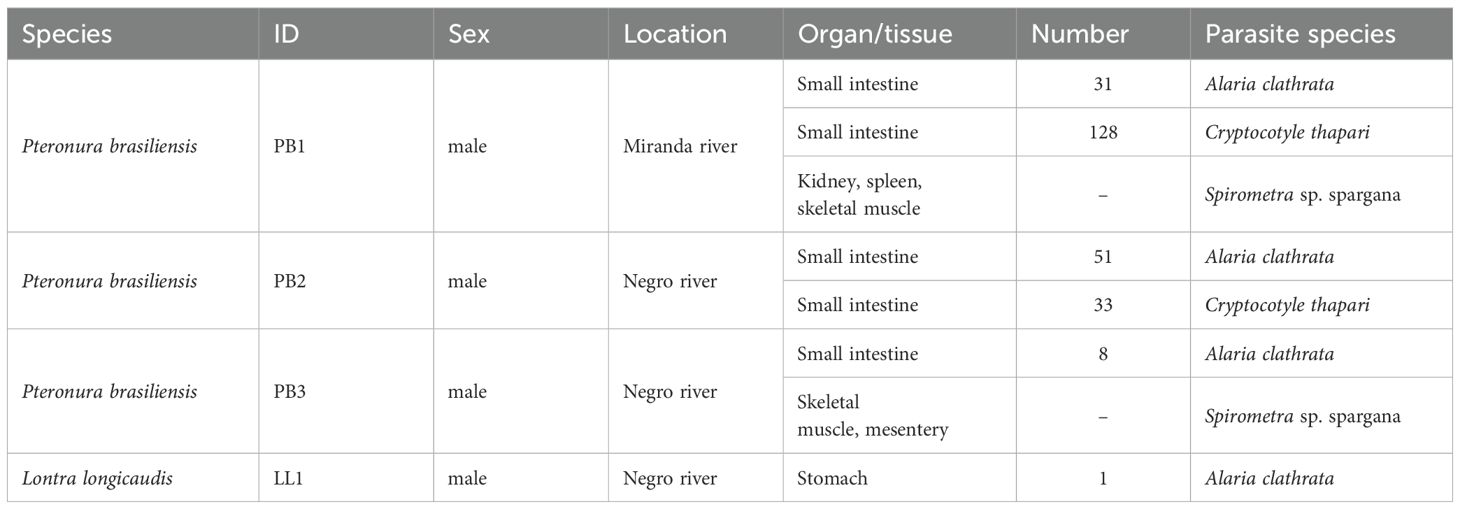
Table 2. Helminths recovered from Pteronura brasiliensis and Lontra longicaudis carcasses (Pantanal, February 2020- September 2022).
Alaria clathrata (Figure 3) has the body divided into two segments separated by a constriction, with the lateral edges of the cranial part slightly curved ventrally, partially covering the tribocytic organ. The (n=10) average body length is 2.89 ± 0.53, 0.72 ± 0.16 mm in width. Vitelaria is observed in the mid part of the body, especially around the tribocytic organ. The oral sucker is small and rounded, measuring 0.10 ± 0.05 in length and 0.13 ± 0.02 in width, sided by two pseudosuckers of variable shape and size. The pharynx measured 0.22 ± 0.03 mm in length and 0.18 ± 0.03 mm in width. The ventral sucker is covered by the tribocytic organ, measuring 0.11 ± 0,05 in length and 0.11 ± 0.06 in width. The testes and ovary are located after the body constraining, at the caudal segment. The testes and ovary are located beyond the body constriction, in the caudal segment. The anterior testes measured 0.22 ± 0.07 in length and 0.41 ± 0.09 in width, while the posterior testes measured 0.29 ± 0.06 in length and 0.46 ± 0.10 in width. The ovary is ellipsoid, smaller than the testes, measuring 0.15 ± 0.03 in length and 0.25 ± 0.07 in width. The eggs (n=10) are 99.30 ± 5.21 µm long and 67.97 ± 6.18 µm wide.
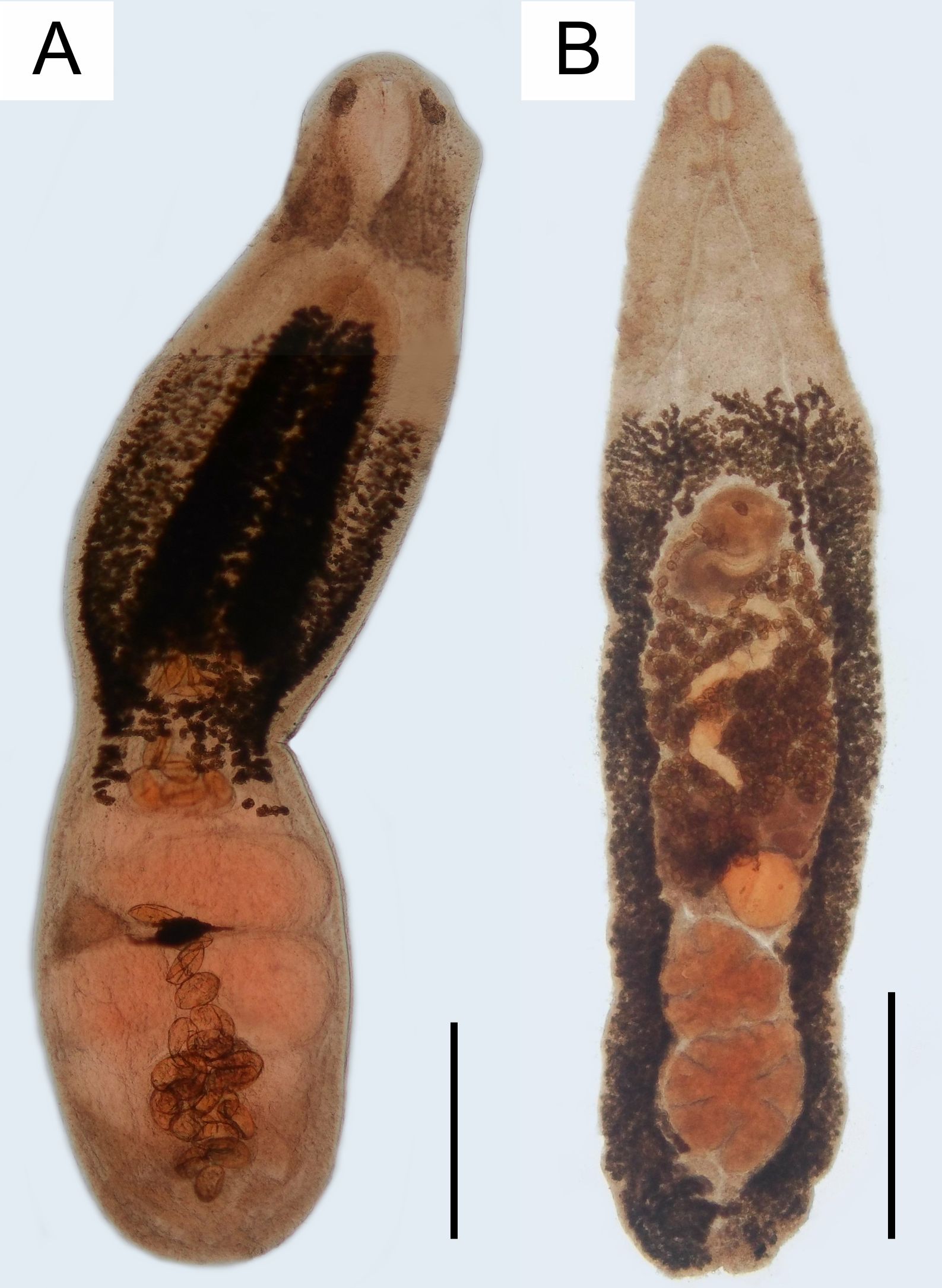
Figure 3. Alaria clathrata and Cryptocotyle thapari from Pteronura brasiliensis, specimen PB1 (Pantanal, February 2020 - September 2022). (A) Alaria clathrata adult worm, whole view, 3000 µm bar. (B) Cryptocotyle thapari adult worm, whole view, 3000 µm bar.
The Cryptocotyle thapari (Figure 3) specimens (n=10) have an elongated body, measuring 2.83 ± 0.26 in length and 0.72 ± 0.09 in width. Vitelaria is observed throughout the caudal two-thirds of the body length. The oral sucker measured 0.07 ± 0.02 in length and 0.08 ± 0.01 in width. The pharynx is small, 0.09 ± 0.01 in length and 0.06 ± 0.02 width. The ventral sucker is located in the mid part of the body, measuring 0.13 ± 0.05 in length and 0.16 ± 0.06 in width. The testes are irregularly lobated, located between the caudal segment of the ceca. The anterior testis is 0.31 ± 0.06 in length and 0.39 ± 0.06 in width, while the posterior measures 0.33 ± 0.05 in length and 0.40 ± 0.06 in width. The ovary is pretesticular, triangle-shaped, with irregular lobes, measuring 0.16 ± 0.03 in length and 0.19 ± 0.03 in width. The eggs were 34.34 ± 2.39 µm in length and 17.12 ± 1.24 µm in width.
The Spirometra sp. spargana (plerocercoids) (n=15) had an average body length of 106 ± 29. They were flat and thin, with an irregularly striated surface, yellowish before fixation, and often occurred in groups. The anterior end is slightly larger, and the posterior end bears a lip-shaped formation. The helminths described above can be observed in Figure 3.
Notably, no 18S rDNA or 28S rDNA sequences for A. clathrata or C. thapari had previously been available in GenBank. This study deposited the first sequences for these species (accession numbers PQ492324.1, PQ492325.1, PQ492326.1, and PQ492327.1). Detailed genetic identities can be observed in Table 3, below.
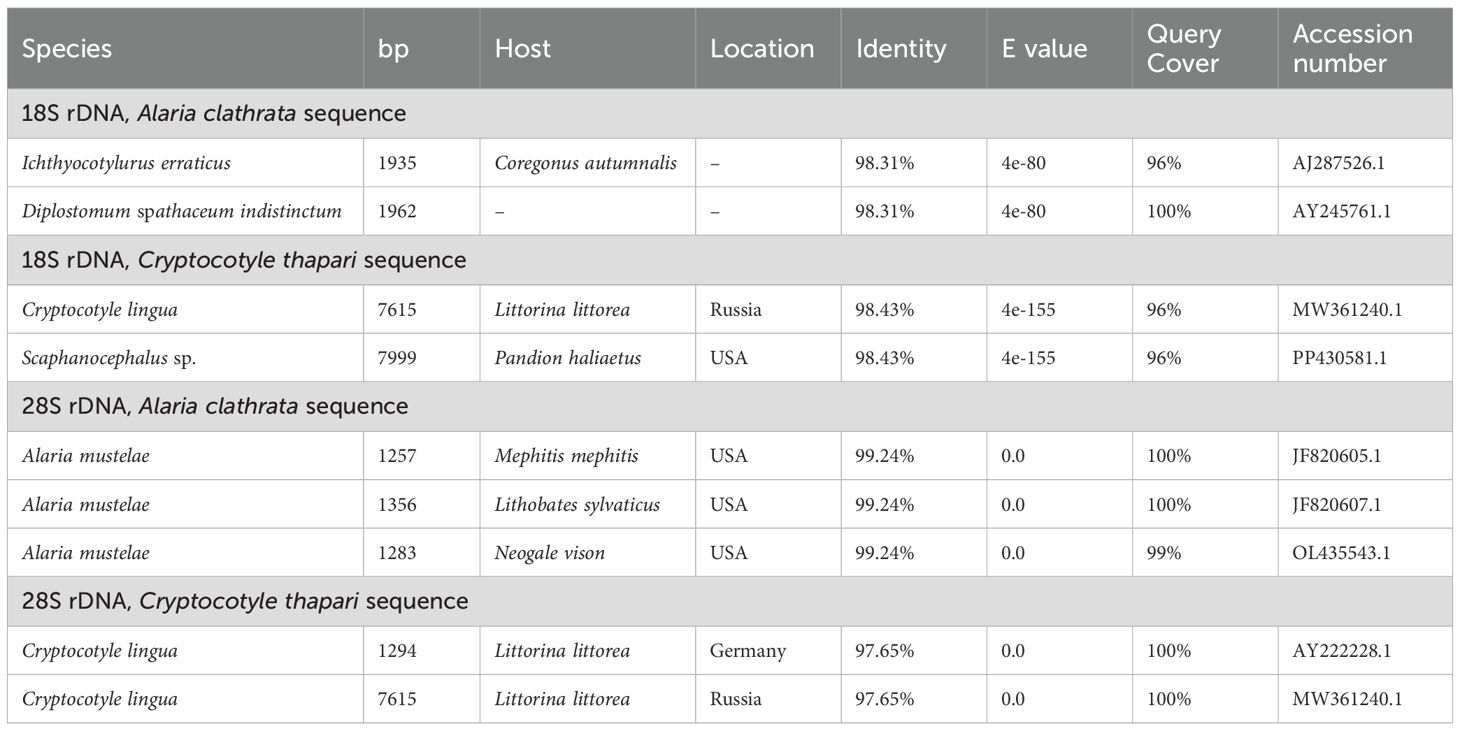
Table 3. Major genetic identities of 18S rDNA and 28S rDNA region sequences of digeneans from the gastrointestinal tract Pteronura brasiliensis specimen PB1 (Pantanal, February 2020- September 2022), deposited in the NCBI database.
4 Discussion
The results of this study revealed a high prevalence of parasites and identified at least three helminth species in samples from both Brazilian otter species, the giant otter and the neotropical otter. Nevertheless, parasite richness in these species remains poorly characterized. Despite their ecological significance as apex predators, these semiaquatic mustelids are understudied, largely due to their elusive behavior, low population densities, the logistical challenges of accessing remote habitats, and the difficulty of safely capturing individuals. These limitations have created significant knowledge gaps regarding their helminth fauna.
The detection of digeneans in both otter species suggests a complex ecological system within their habitats. These parasites typically have multi-stage life cycles involving mollusks as intermediate hosts, yet their specific cycles in freshwater environments remain poorly understood (Kudlai et al., 2015; Müller et al., 2015). Knowledge gaps include identifying mollusk hosts, understanding the role of paratenic hosts, and uncovering cercarial infection mechanisms. A comparable pattern in terms of developmental complexity is seen in species of the same genus, such as A. clathrata. Alaria alata has a wide array of paratenic hosts, including amphibians and mammals. Although human cases of A. alata infection have not been documented in Europe, its high prevalence in wild boars and the zoonotic potential of related species like A. americana highlight the risk of accidental transmission to mammals, including humans, through the food chain (Möhl et al., 2009; Guardone et al., 2022). Moreover, due to the poor knowledge of wild animal helminth fauna, new species are often described. A notable recent example is the identification of Dracunculus jaguape, a previously unknown parasite of L. longicaudis in Argentina (Natalini et al., 2023), whose life history remains undescribed but is presumed to involve multiple hosts, as is typical of the genus (Box et al., 2021). Similarly, Cryptocotyle dominicana, the only known congener of Cryptocotyle thapari in South America, has been recently identified in kelp gulls (Larus dominicanus), with its metacercariae recovered from Galaxias platei, a freshwater fish from Patagonia (Casalins et al., 2020).
Given their indirect and complex life cycles, these parasites are highly susceptible to ecological changes, serving as sensitive indicators of ecosystem health (Sures et al., 2017; Schwelm et al., 2021). Environmental disturbances, including but not limited to droughts, wildfires, habitat loss, and fragmentation, may have a direct impact on their presence and persistence. Understanding these dynamics is crucial for informing conservation strategies, as the health of host-parasite systems can reflect broader environmental changes. Moraes et al. (2024) suggest that seasonal floods and droughts in the Pantanal may influence infections of Spirometra spp. in wild animals. Notably, the carcasses of giant otters infected with plerocercoids of these cestodes were found during extreme drought conditions, which have become more frequent in recent years (Marengo et al., 2021). These fluctuations in water levels likely alter the dynamics of parasite transmission, as droughts may concentrate hosts near the water bodies and increase the likelihood of encounters with infected intermediate hosts, while floods could disperse the parasites over a wider area. The changing climate patterns in the Pantanal could thus exacerbate the spread of such infections, including other parasites that were not found in this study, posing new challenges to the persistence of giant otter populations in the region.
Unsporulated coccidian oocysts, and digeneans, strongylid-type, and cestodes eggs were found in the feces of giant otters from the same region in the Pantanal using flotation and sedimentation tests (Borges et al., 2022; Soresini et al., 2016). Although the presence of Cystoisospora oocysts could indicate an undescribed parasite, a possible spillover from domestic animals may explain their occurrence, with potential impacts on both wildlife and public health. Meanwhile, Spirometra sp. spargana, known to cause sparganosis in humans and animals, underscores the need for monitoring zoonotic diseases in areas of close human-wildlife interaction.
This study represents the first molecular analysis of the digeneans Alaria clathrata and Cryptocotyle thapari, with newly acquired sequences now deposited in GenBank, to support future research. Specifically, we report four novel DNA sequences, two from A. clathrata and two from C. thapari, targeting the 18S rDNA and 28S rDNA regions. The challenges posed by the conservation status of P. brasiliensis and L. longicaudis, as well as difficulties in managing these species in captivity, limit the feasibility of conducting experimental infections. Non-invasive molecular approaches, such as eDNA sampling targeting larval forms and potential intermediate or paratenic hosts (Reza Varzandi et al., 2024), therefore provide a viable alternative. However, these methods remain limited by the scarcity of reference genetic data, a widespread issue in wildlife helminth research (Poulin et al., 2019).
Despite the important role that parasites play in ecosystem functioning, food web stability, and host population regulation (Dougherty et al., 2016; Carlson et al., 2020; Milotic et al., 2020), there are still many knowledge gaps regarding parasite diversity, especially in freshwater systems. This study provides novel insights into the relationship between parasites and Brazilian otters, highlighting the importance of parasite surveillance in wildlife, particularly in aquatic ecosystems. From a conservation perspective, documenting new host–parasite interactions is highly valuable, particularly when assessing potential threats to declining populations and endangered species. Moreover, a comprehensive understanding of natural host-parasite relationships may be useful to prevent the introduction of novel parasites into naïve environments during translocation activities for conservation or rehabilitation purposes (Warne and Chaber, 2023) or through domestic animals carrying generalist pathogens (Soresini et al., 2023). Given the ongoing transformation of natural habitats, the escalating anthropogenic pressure, and the accelerating impacts of climate change, the surveillance of parasitic infections in wild populations can serve as an early warning system for emerging threats. Even the absence of parasites, particularly those with complex ecologies that were previously observed at high abundance or prevalence, may signal environmental change. This is particularly critical for declining populations and threatened species, such as the giant otter. The dynamics of parasite infection may influence health, survival, and long-term conservation outcomes (Gonchoroski et al., 2025). Additionally, considering the ecological role that otters play in their ecosystems, this group can also be used as a proxy for assessing environmental health and may be a valuable tool for public health assessments.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: NCBI GenBank, accession, PQ492324.1, PQ492325.1, PQ492326.1, PQ492327.1.
Ethics statement
The animal study was approved by Instituto Chico Mendes de Conservação da Biodiversidade (ICMBio). The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
PP: Methodology, Supervision, Project administration, Writing – review & editing, Data curation, Conceptualization, Validation, Investigation, Writing – original draft, Formal analysis, Funding acquisition, Software, Visualization, Resources. CL: Data curation, Methodology, Validation, Conceptualization, Investigation, Supervision, Project administration, Resources, Funding acquisition, Writing – review & editing, Formal analysis, Software, Writing – original draft, Visualization. MF: Methodology, Validation, Conceptualization, Data curation, Supervision, Software, Investigation, Resources, Formal analysis, Writing – review & editing, Funding acquisition, Project administration, Writing – original draft, Visualization. LR: Validation, Data curation, Visualization, Resources, Methodology, Formal analysis, Project administration, Investigation, Conceptualization, Writing – review & editing, Software, Funding acquisition, Writing – original draft, Supervision. GS: Formal analysis, Software, Methodology, Data curation, Resources, Writing – original draft, Visualization, Project administration, Writing – review & editing, Validation, Conceptualization, Supervision, Investigation, Funding acquisition. NF: Writing – review & editing, Funding acquisition, Resources, Formal analysis, Writing – original draft, Project administration, Conceptualization, Visualization, Methodology, Software, Validation, Data curation, Investigation, Supervision. GM: Investigation, Supervision, Conceptualization, Validation, Funding acquisition, Software, Writing – review & editing, Project administration, Writing – original draft, Formal analysis, Methodology, Resources, Data curation, Visualization. WO: Writing – original draft, Visualization, Software, Data curation, Formal analysis, Resources, Funding acquisition, Conceptualization, Methodology, Project administration, Investigation, Supervision, Validation, Writing – review & editing. TM: Investigation, Supervision, Data curation, Writing – original draft, Methodology, Software, Conceptualization, Resources, Writing – review & editing, Project administration, Visualization, Validation, Funding acquisition, Formal analysis. CA-P: Resources, Software, Funding acquisition, Project administration, Writing – original draft, Conceptualization, Writing – review & editing, Supervision, Methodology, Data curation, Visualization, Formal analysis, Investigation, Validation. FL: Methodology, Conceptualization, Software, Writing – original draft, Investigation, Supervision, Data curation, Visualization, Validation, Funding acquisition, Resources, Formal analysis, Project administration, Writing – review & editing. EL: Conceptualization, Investigation, Writing – review & editing, Writing – original draft, Supervision, Funding acquisition, Software, Data curation, Visualization, Validation, Resources, Formal analysis, Methodology, Project administration.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. Field activities for biological sample collection were made possible through funding provided by the Houston Zoo and Zoo Miami. EL is granted a CNPq fellowship (311063/2022-5).
Acknowledgments
The authors are grateful to Projeto Ariranhas, Barranco Alto Farm, Panthera Brasil, the Jaguar Identification Project, Embrapa Pantanal, the Federal University of Mato Grosso do Sul, and Instituto Homem Pantaneiro for their logistical support and collaboration. We also thank the staff of the Pantanal Study Base and the Zoological Reference Collection of the Federal University of Mato Grosso do Sul for their invaluable assistance during the necropsies.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Altschul S. F., Gish W., Miller W., Myers E. W., and Lipman D. J. (1990). Basic local alignment search tool. J. Mol. Biol. 215, 403–410. doi: 10.1016/S0022-2836(05)80360-2, PMID: 2231712
Barros-Battesti D. M., André M. R., Hoppe E. G. L., Seron G., Bassini-Silva R., Calchi A. C., et al. (2023). Noteworthy records of the ticks Ornithodoros rostratus and Amblyomma sculptum parasitizing Pteronura brasiliensis in the central-western region of Brazil, with pathogen investigation notes. Rev. Bras. Parasitol. Vet. 33, e014523. doi: 10.1590/S1984-29612024003, PMID: 38126574
Borges J. C. G., Lima V. F., Da Silva E. M., dos Santos Lima D., Marmontel M., Carvalho V. L., et al. (2022). Use of the FLOTAC technique as a new coproparasitological diagnostic method in aquatic mammals and comparison with traditional methods. Anais. Acad. Bras. Ci. 94, e20201184. doi: 10.1590/0001-3765202220201184, PMID: 35239771
Box E. K., Yabsley M. J., Garrett K. B., Thompson A. T., Wyckoff S. T., and Cleveland C. A. (2021). Susceptibility of anurans, lizards, and fish to infection with Dracunculus species larvae and implications for their roles as paratenic hosts. Sci. Rep. 11, 11802. doi: 10.1038/s41598-021-91122-5, PMID: 34083638
Caira J. N., Marques F. P., Jensen K., Kuchta R., and Ivanov V. (2013). Phylogenetic analysis and reconfiguration of genera in the cestode order Diphyllidea. Int. J. Parasitol. 43, 621–639. doi: 10.1016/j.ijpara.2013.03.001, PMID: 23603519
Carlson C. J., Hopkins S., Bell K. C., Doña J., Godfrey S. S., Kwak M. L., et al. (2020). A global parasite conservation plan. Biol. Conserv. 250, 108596. doi: 10.1016/j.biocon.2020.108596
Casalins L. M., Arbetman M. P., Viozzi G. P., and Flores V. R. (2020). A new species of Cryptocotyle (Digenea: Heterophyidae) infecting kelp gull and a galaxiid fish in Patagonian freshwater environments: morphological and molecular analyses. J. Parasitol. 106, 203–210. doi: 10.1645/18-44, PMID: 32164027
Colwell D. D., Otranto D., and Stevens J. R. (2008). Oestrid flies: eradication and extinction versus biodiversity. Trends Parasitol. 25, 500–504. doi: 10.1016/j.pt.2009.07.011, PMID: 19762281
Dougherty E. R., Carlson C. J., Bueno V. M., Burgio K. R., Cizauskas C. A., Clements C. F., et al. (2016). Paradigms for parasite conservation. Conserv. Biol. 30, 724–733. doi: 10.1111/cobi.12634, PMID: 26400623
Echenique J. V., Soares M. P., Mascarenhas C. S., Bandarra P. M., Quadros P., Driemeier D., et al. (2018). Lontra longicaudis infected with canine parvovirus and parasitized by Dioctophyma renale. Pesq. Vet. Bras. 38, 1844–1848. doi: 10.1590/1678-5150-PVB-5744
Ewing B. and Green P. (1998). Base calling of automated sequencer traces using PHRED. II. Error probabilities. Genome Res. 8, 186–194. doi: 10.1101/gr.8.3.175, PMID: 9521922
Foerster N., Soresini G., Paiva F., Silva F. A., Leuchtenberger C., and Mourão G. (2022). First report of myiasis caused by Cochliomyia hominivorax in free-ranging giant otter (Pteronura brasiliensis). Rev. Bras. Parasitol. Vet. 31, e009522. doi: 10.1590/S1984-29612022058, PMID: 36417628
Gagne R. B., Crooks K. R., Craft M. E., Chiu E. S., Fountain-Jones N. M., Malmberg J. L., et al. (2022). Parasites as conservation tools. Conserv. Biol. 36, e13719. doi: 10.1111/cobi.13719, PMID: 33586245
Gardner S. L. and Thew P. T. (2006). Redescription of cryptocotyle thapari mcIntosh 1953 (Trematoda: Heterophyidae), in the river otter Lutra longicaudis from Bolivia. Comp. Parasitol. 73, 20–23. doi: 10.1654/0001.1
Gonchoroski G. Z., Martin A., Raphael G., Rodrigues L. A., Furtado M. M., Mourão G. M., et al. (2025). Intergroup conflict and myiasis-induced mortality in a giant otter from the Brazilian pantanal: implications for population conservation. IUCN. Otter. Spec. Group. Bull. 42, 63–70.
Gordon D., Abajian C., and Green P. (1998). Consed: A graphical tool for sequence finishing. Genome Res. 8, 195–202. doi: 10.1101/gr.8.3.195, PMID: 9521923
Green P. (1996). Phrad documentation. Available online at: http://bozeman.mbt.washington.edu/phrap.docs/phrap.html (Accessed July 7, 2025).
Groenendijk J., Leuchtenberger C., Marmontel M., Van Damme P. A., Wallace R., and Schenck C. (2023). “Pteronura brasiliensis (amended version of 2022 assessment),” in The IUCN red list of threatened Species, e.T18711A244867206. Gland, Switzerland: International Union for Conservation of Nature (IUCN). doi: 10.2305/IUCN.UK.2023-1.RLTS.T18711A244867206.en
Guardone L., Armani A., Mancianti F., and Ferroglio E. (2022). A review on Alaria alata, Toxoplasma gondii and Sarcocystis spp. in mammalian game meat consumed in Europe: epidemiology, risk management and future directions. Animals 12, 263. doi: 10.3390/ani12030263, PMID: 35158587
Hudson P. J., Dobson A. P., and Lafferty K. D. (2006). Is a healthy ecosystem one that is rich in parasites? Trends Ecol. Evol. 21, 381–385. doi: 10.1016/j.tree.2006.04.007, PMID: 16713014
Kudlai O., Stunžėnas V., and Tkach V. (2015). The taxonomic identity and phylogenetic relationships of Cercaria pugnax and C. helvetica XII (Digenea: Lecithodendriidae) based on morphological and molecular data. Int. J. Parasitol. 45, 881–890. doi: 10.14411/fp.2015.003, PMID: 25960547
Leuchtenberger C., Barocas A., Thoisy B., Ward C., Evangelista E., Michalski F., et al. (2018). “Giant otter,” in Global Otter Conservation Strategy. Eds. N. Duplaix N. and Savage M. (IUCN Otter Specialist Group, Four Corners Institute, Salem, Oregon), 74–81.
Lymbery A. J., Hassan M., Morgan D. L., Beatty S. J., and Doupé R. G. (2010). Parasites of native and exotic freshwater fishes in south-western Australia. J. Fish. Biol. 76, 1770–1785. doi: 10.1111/j.1095-8649.2010.02615.x, PMID: 20557630
Marengo J. A., Cunha A. P., Cuartas L. A., Deusdará Leal K. R., Broedel E., Seluchi M. E., et al. (2021). Extreme drought in the Brazilian Pantanal in 2019 – 2020: Characterization, causes, and impacts. Front. Water 3. doi: 10.3389/frwa.2021.639204
Milotic M., Lymbery A., Thompson A., Doherty J. F., and Godfrey S. (2020). Parasites are endangered by the conservation of their hosts: Meta-analyses of the effect of host captivity on the odds of parasite infection. Biol. Conserv. 248, 108702. doi: 10.1016/j.biocon.2020.108702
Möhl K., Große K., Hamedy A., Wüste T., Kabelitz P., and Lücker E. (2009). Biology of Alaria spp. and human exposition risk to Alaria mesocercariae—a review. Parasitol. Res. 105, 1–15. doi: 10.1007/s00436-009-1444-7, PMID: 19430816
Moraes I. D. S., Silva V. L. D. B., Andrade-Silva B. E. D., Gomes A. P. N., Urzedo N. F. D., Abolis V. B., et al. (2024). Gastrointestinal Helminths in Wild Felids in the Cerrado and Pantanal: zoonotic bioindicators in important Brazilian Biomes. Animals 14, 1622. doi: 10.3390/ani14111622, PMID: 38891670
Müller T., Czarnoleski M., Labecka A. M., Cichy A., Zając K., and Dragosz-Kluska D. (2015). Factors affecting trematode infection rates in freshwater mussels. Hydrobiologia 742, 59–70. doi: 10.1007/s10750-014-1965-7
Muniz-Pereira L. C., Vieira F. M., and Luque J. L. (2009). Checklist of helminth parasites of threatened vertebrate species from Brazil. Zootaxa 2123, 1–45. doi: 10.11646/zootaxa.2123.1.1
Natalini M. B., Notarnicola J., Monje L. D., Vallejos A. B., Gavier F. S., and Kowalewski M. M. (2023). A new Dracunculus species (Nematoda: Dracunculoidea) in Neotropical otters (Lontra longicaudis) from Argentina: Morphological and molecular characterization. Int. J. Parasitol. Parasites Wildl. 22, 234–242. doi: 10.1016/j.ijppaw.2023.11.002, PMID: 38045874
Olson P. D., Cribb T. H., Tkach V. V., Bray R. A., and Littlewood D. T. J. (2003). Phylogeny and classification of the digenea (Platyhelminthes: trematoda). Int. J. Parasitol. 33, 733–755. doi: 10.1016/S0020-7519(03)00049-3, PMID: 12814653
Perin P. P., Lapera I. M., Arias-Pacheco C. A., Mendonça T. O., Oliveira W. J., de Souza Pollo A., et al. (2023). Epidemiology and integrative taxonomy of helminths of invasive wild boars, Brazil. Pathogens 12, 175. doi: 10.3390/pathogens12020175, PMID: 36839447
Poulin R., Hay E., and Jorge F. (2019). Taxonomic and geographic bias in the genetic study of helminth parasites. Int. J. Parasitol. 49, 429–435. doi: 10.1016/j.ijpara.2018.12.005, PMID: 30797772
Reza Varzandi A., Zanet S., Rubele E., Occhibove F., Vada R., Benatti F., et al. (2024). Development of a qPCR Duplex Assay for simultaneous detection of Fascioloides magna and Galba truncatula in eDNA samples: Monitoring beyond boundaries. Sci. Total. Environ. 916, 170338. doi: 10.1016/j.scitotenv.2024.170338, PMID: 38266734
Rheingantz M. L., Rosas-Ribeiro P., Gallo-Reynoso J., Fonseca da Silva V. C., Wallace R., Utreras V., et al. (2022). “Lontra longicaudis (amended version of 2021 assessment),” in The IUCN Red List of Threatened Species, e.T12304A219373698 Gland, Switzerland: International Union for Conservation of Nature (IUCN). doi: 10.2305/IUCN.UK.2022-2.RLTS.T12304A219373698.en
Rheingantz M. L., Santiago-Plata V. M., and Trinca C. S. (2017). The Neotropical otter Lontra longicaudis: A comprehensive update on the current knowledge and conservation status of this semiaquatic carnivore. Mamm. Rev. 47, 291–305. doi: 10.1111/mam.12098
Sanger F., Nicklen S., and Coulson A. R. (1977). DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. U.S.A. 74, 5463–5467. doi: 10.1073/pnas.74.12.5463, PMID: 271968
Schwelm J., Selbach C., Kremers J., and Sures B. (2021). Rare inventory of trematode diversity in a protected natural reserve. Sci. Rep. 11, 22066. doi: 10.1038/s41598-021-01457-2, PMID: 34764355
Smith K. F., Acevedo-Whitehouse K., and Pedersen A. B. (2009). The role of infectious diseases in biological conservation. Anim. Conserv. 12, 1–12. doi: 10.1111/j.1469-1795.2008.00228.x
Soresini G., Foerster N., Paiva F., Mourão G., and Leuchtenberger C. (2023). Amblyomma sculptum ticks on a giant otter from the Brazilian Pantanal. Rev. Bras. Parasitol. Vet. 32, e010923. doi: 10.1590/S1984-29612023053, PMID: 37878901
Soresini G. C. G., Teixeira V. N., Aguiar F., and Mourão G. M. (2016). Occurrence of gastrointestinal parasites in free-ranging giant otter (Pteronura brasiliensis) in the Southern Pantanal, Brazil. 13th International Otter Congress,15. Abstract retrieved from Programme Booklet. Singapore: IUCN-SSC Otter Specialist Group (OSG).
Sures B., Nachev M., Selbach C., and Marcogliese D. J. (2017). Parasite responses to pollution: what we know and where we go in ‘Environmental Parasitology’. Parasitol. Vectors 10, 65. doi: 10.1186/s13071-017-2001-3, PMID: 28166838
Thompson R. C. A., Lymbery A. J., and Smith A. (2010). Parasites, emerging disease and wildlife conservation. Int. J. Parasitol. 40, 1163–1170. doi: 10.1016/j.ijpara.2010.04.009, PMID: 20452354
Tomas W. M., de Oliveira Roque F., Morato R. G., Medici P. E., Chiaravalloti R. M., Tortato F. R., et al. (2019). Sustainability agenda for the Pantanal wetland: perspectives on a collaborative interface for science, policy, and decision-making. Trop. Conserv. Sci. 12, 1940082919872634. doi: 10.1177/194008291987263
Travassos L., Teixeira De Freitas J. F., and Kohn A. (1969). Trematódeos do Brazil. Mem. Inst. Oswaldo. Cruz. 67, 1–886.
Uchôa T., Vidolin G. P., Fernandes T. M., Velastin G. O., and Mangini P. R. (2004). Aspectos ecológicos e sanitários da lontra (Lontra longicaudis Olfers 1818) na reserva natural salto Morato, Guaraqueçaba, Paraná, Brasil. Cad. Biodivers. 4, 19–28.
Vieira F. M., Luque J. L., and Muniz-Pereira L. C. (2008). Checklist of helminth parasites in wild carnivore mammals from Brazil. Zootaxa 1721, 1–23. doi: 10.5281/zenodo.181136vieira
Warne R. K. and Chaber A. L. (2023). Assessing disease risks in wildlife translocation projects: a comprehensive review of disease incidents. Animals 13, 3379. doi: 10.3390/ani13213379, PMID: 37958133
Yamaguti S. (1958). Systema Helminthum. Vol. I. The Digenetic Trematodes of Vertebrates - Part II (New York: Interscience Publishers).
Keywords: neotropical otter, giant otter, Pantanal, parasites, helminths, wildlife health, fecal sampling
Citation: Perin PP, Leuchtenberger C, Furtado MM, Rodrigues LdA, Soresini G, Foerster N, Mourão G, Oliveira WJ, Mendonça TO, Arias-Pacheco CA, Loureiro FA and Lux Hoppe EG (2025) Parasitological evaluation of the neotropical otter Lontra longicaudis and the giant otter Pteronura brasiliensis: swimming in little-known waters before it is too late. Front. Mamm. Sci. 4:1678138. doi: 10.3389/fmamm.2025.1678138
Received: 01 August 2025; Accepted: 21 August 2025;
Published: 17 September 2025.
Edited by:
Pedro Manoel Galetti Jr, Federal University of São Carlos, BrazilReviewed by:
Manuel Uribe, Justus Liebig University Giessen, GermanyLuciano Dos Anjos, São Paulo State University, Brazil
Copyright © 2025 Perin, Leuchtenberger, Furtado, Rodrigues, Soresini, Foerster, Mourão, Oliveira, Mendonça, Arias-Pacheco, Loureiro and Lux Hoppe. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Estevam G. Lux Hoppe, bHV4LmhvcHBlQHVuZXNwLmJy