 Sangami Pugazenthi1,2
Sangami Pugazenthi1,2 Shree S. Pari1
Shree S. Pari1 Ziyan Zhang1
Ziyan Zhang1 Julie Silverstein3,4
Julie Silverstein3,4 Albert H. Kim1,4,5
Albert H. Kim1,4,5 Bhuvic Patel1,4,5*
Bhuvic Patel1,4,5*- 1Taylor Family Department of Neurological Surgery, Washington University School of Medicine, St. Louis, MO, United States
- 2Department of Neurological Surgery, University of Pittsburgh Medical Center, Pittsburgh, PA, United States
- 3Division of Endocrinology, Metabolism and Lipid Research, Washington University School of Medicine, St. Louis, MO, United States
- 4WashU Medicine Pituitary Center, Washington University School of Medicine, St. Louis, MO, United States
- 5The Brain Tumor Center, Siteman Cancer Center, Washington University School of Medicine, St. Louis, MO, United States
Pituitary neuroendocrine tumors (PitNETs) are a heterogeneous group of intracranial neoplasms that vary in hormonal activity, histological features, and clinical behavior. The rise of high-throughput sequencing and molecular profiling technologies has enabled multiomic approaches—including genomics, transcriptomics, epigenomics, proteomics, and metabolomics—to deepen our understanding of PitNET pathogenesis. These studies have identified key mutations, transcriptional lineages, epigenetic modifications, and proteomic features that contribute to tumor subtype classification, invasiveness, and treatment response. Integrative multi-omic analyses have further revealed distinct molecular subtypes, complex regulatory networks, and molecular profiles that can predict recurrence and therapeutic efficacy. These approaches hold strong potential for advancing personalized medicine in PitNETs, supporting patient-specific diagnosis, prognostication, and therapeutic strategies. Future directions include the application of emerging -omic technologies and the development of robust computational tools to integrate and translate multi-layered data into clinically actionable insights.
Introduction
Pituitary tumors represent a diverse group of neoplasms that originate from the endocrine cells of the pituitary gland and account for approximately 17.8% of all intracranial tumors (1). Historically termed pituitary adenomas, these tumors have been considered largely benign and indolent. However, this perception has evolved significantly with advances in molecular pathology and clinical characterization. In 2022, the World Health Organization (WHO) officially reclassified these tumors as pituitary neuroendocrine tumors (PitNETs) to better reflect their neuroendocrine origin and biological spectrum (2). A pivotal aspect of the new WHO classification is the use of pituitary-specific transcription factors (TFs) to define tumor lineage more accurately than traditional hormonal immunostaining alone. The key TFs include pituitary-specific positive transcription factor 1 (PIT1), steroidogenic factor 1 (SF1), and T box transcription factor (TPIT), which correspond to the somatotroph/lactotroph/thyrotroph, gonadotroph, and corticotroph lineages, respectively (Figure 1). This molecular stratification helps distinguish morphologically similar but biologically distinct subtypes, thereby enhancing diagnostic precision and prognostic estimation (2). Despite these advances, the clinical management of PitNETs remains challenging due to a lack of robust biomarkers for tumor aggressiveness, treatment response, and recurrence risk. In this context, multiomic approaches—including genomic, transcriptomic, epigenomic, proteomic, and metabolomic profiling—offer powerful tools to dissect the complexity of PitNETs. Integrative multiomic analysis can provide a systems-level understanding of tumor biology, identify molecular subgroups, and uncover novel targets for therapy and early detection (3). This review summarizes the current landscape and emerging insights from multiomic studies in PitNETs, emphasizing their potential to revolutionize classification, prognosis, and individualized treatment strategies in pituitary tumor management.
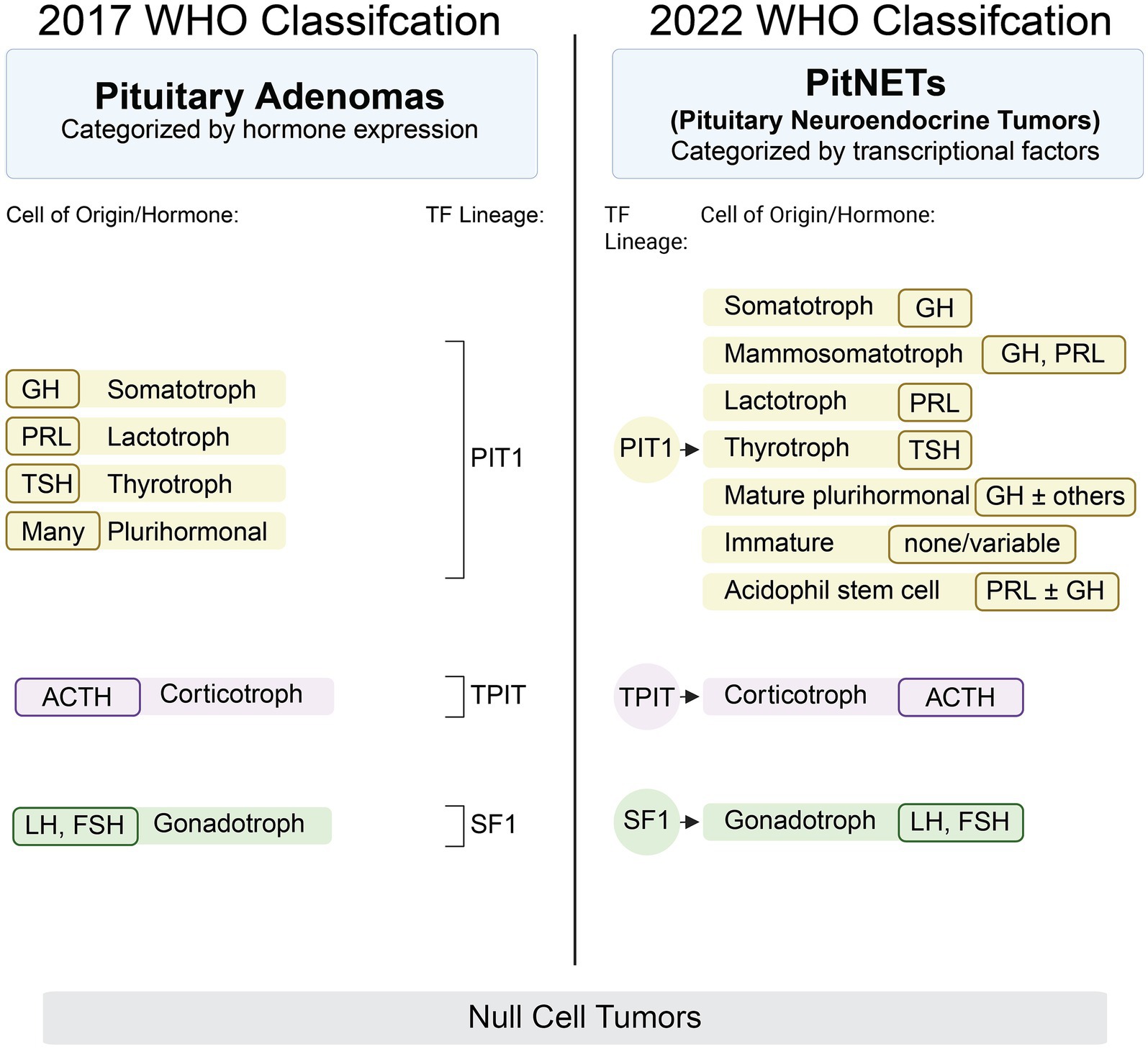
Figure 1. Comparison of 2017 and 2022 PitNET WHO classification schemes. PitNET, Pituitary neuroendocrine tumor; WHO, World Health Organization.
Genomic analysis
Genomic analyses have played a crucial role in uncovering the molecular underpinnings of PitNETs, shedding light on both sporadic and hereditary forms. Although PitNETs display a relatively low mutational burden compared to other solid tumors, several recurrent somatic and germline alterations have been identified that contribute to tumor initiation, hormonal dysregulation, and progression.
The most well-characterized somatic mutations in PitNETs are subtype specific. Guanine nucleotide-binding protein, alpha stimulating (GNAS) mutations are frequently found in somatotroph tumors, promoting cyclic adenosine monophosphate (cAMP) signaling and growth hormone (GH) overproduction. PitNETs with GNAS mutations have been associated with smaller size and decreased invasiveness (4). In corticotroph tumors causing Cushing’s Disease, ubiquitin carboxyl-terminal hydrolase 8 (USP8) mutations are present in up to 40% of cases and result in impaired degradation of epidermal growth factor receptor (EGFR), enhancing adrenocorticotropic hormone (ACTH) secretion and cellular proliferation (5, 6). Other mutations described in corticotroph PitNETs include ubiquitin specific peptidase 48 (USP48), B-Raf proto-oncogene, serine/threonine kinase (BRAF), and tumor protein p53 (TP53) (7, 8). Despite these discoveries, most PitNETs lack recurrent driver mutations, suggesting a significant role for epigenetic regulation, chromosomal instability, and post-transcriptional mechanisms in tumor biology. A subset of PitNETs arise in the context of hereditary tumor syndromes, most notably Multiple Endocrine Neoplasia type 1 (MEN1), caused by inactivating mutations in the MEN1 gene, which encodes the tumor suppressor menin. Other inherited mutations involve cyclin-dependent kinase inhibitor 1B (CDKN1B) (associated with MEN4), aryl hydrocarbon receptor interacting protein (AIP), and succinate dehydrogenase (SDHx) (9–12). Patients harboring AIP mutations most commonly present with somatotropinomas, often at a younger age, with larger tumors and more growth hormone (GH) secretion (13, 14). Succinate dehydrogenase complex iron sulfur subunit B/D (SDHB/D) mutations have been shown to be associated with combined paragangliomas, pheochromocytomas, and less frequently PitNETs, suggesting shared tumorigenesis pathways related to mitochondrial metabolism (12).
With the development of next-generation sequencing (NGS), including whole-exome sequencing (WES) and whole-genome sequencing (WGS), studies utilizing these methods provided a broader landscape of mutational events in PitNETs. In 2016, Song et al. examined the somatic mutational landscape of 125 PitNETs, identifying low mutational burden, confirming the presence of previously described mutations such as GNAS, MEN1, and USP8, identifying novel mutations such as kinesin heavy chain isoform 5A (KIF5A) and growth factor receptor-bound protein 10 (GRB10), and determining that 18% of tumors harbor copy number alterations (CNAs). Gene ontology analysis revealed that plurihormonal, GH-, prolactin (PR)-, and ACTH-secreting PitNETs were enriched for somatic mutations in overlapping molecular pathways as were TSH- and LH/FSH-secreting PitNETs (15). Subsequently, Bi et al. identified that 29% of PitNETs have CNAs, but novel somatic alterations in genes were infrequent and often non-recurrent. They found that the tumors with more disrupted genomes (higher CNA burden) were more likely to be functional PitNETs or null cell tumors compared to PitNETs with less disrupted genomes, which were more likely nonfunctional (16). Large-scale sequencing efforts continue to uncover novel candidate genes and low-frequency variants that may contribute to tumor biology but integration of genomic data with transcriptomic and epigenomic profiles is essential to elucidate the mechanistic impact of these mutations, and inclusion of phenotypic data is critical for clinical relevance.
To facilitate clinical interpretation, Table 1 summarizes key PitNET biomarkers identified across multiomic studies, specifically highlighting their functional roles, prognostic value, and therapeutic relevance. Even though many markers remain investigational, this framework may inform future biomarker-guided therapy trials.
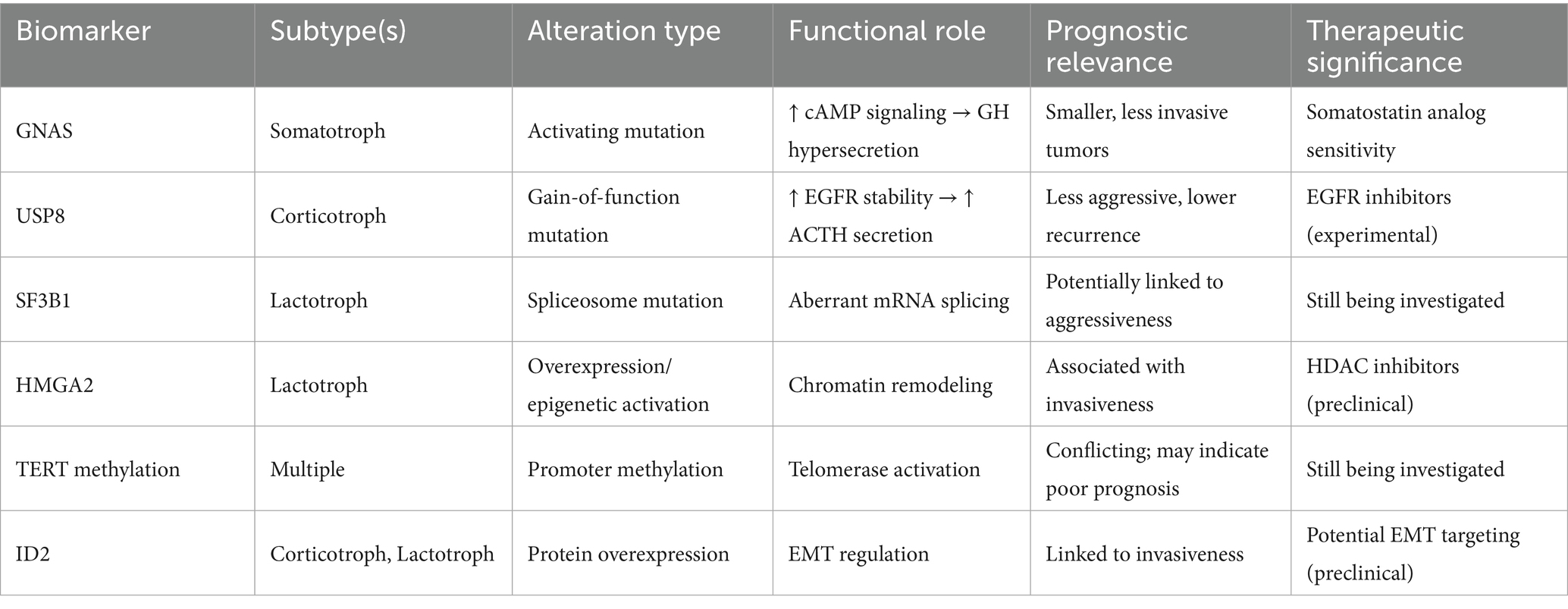
Table 1. Known biomarkers prognostic/therapeutic utility.
Transcriptional profiling
Transcriptomic profiling using techniques such as bulk and single-cell RNA sequencing has emerged as a powerful approach to characterize PitNETs beyond histology and hormonal output, offering insights into their functional identity, heterogeneity, and aggressiveness. Unlike genomic alterations, which are relatively infrequent in PitNETs, transcriptional changes are widespread and reflect both lineage commitment and tumor behavior.
Transcriptomic profiling has had a significant impact on the field of pituitary tumors as this method was used to discover the relevance of TFs in the classification of PitNETs highlighted in the 2022 WHO guidelines. The use of transcription factors has been shown to be more reliable than previous methods using histology, immunochemistry, in situ hybridization, and hormone expression to identify and classify these tumors (2). The biological role of PIT1, SF1 and TPIT in normal pituitary gland development and PitNET pathogenesis has also been investigated using bulk RNA sequencing (17, 18). In normal corticotroph development, TPIT along with paired like homeodomain 1 (PITX1) activate the proopiomelanocortin (POMC) gene (19, 20). On the other hand, suppression of TPIT causes pituitary neuroendocrine cells to differentiate into gonadotroph or thyrotroph cells (21). The PIT1 TF lineage is positively regulated by paired-like homeobox 1 (PROP1) and negatively regulated by HESX homeobox 1 (HESX1) (22, 23). Each hormonal subtype of PIT1 PitNETs have specific mechanisms through which PIT1 is involved in pathogenesis. Gonadotrophs are part of the SF1-lineage of PitNETs; SF1 transcription in part relies on the binding of estrogen-to-estrogen receptor alpha, which mediates chromatin remodeling of the SF1 locus (24).
Invasive PitNETs have significant differences in their transcriptional profiles compared to noninvasive tumors, including differentially expressed genes related to the Nuclear Factor-kappa B (NF-κB) and antitumoral immune response (25, 26). Invasive prolactinomas exhibited significantly different transcriptional profiles compared to noninvasive prolactinomas (27). Compared to noninvasive corticotrophs, invasive corticotroph tumors exhibit upregulation of cyclin D2 (CCND2) and zinc finger protein 676 (ZNF676) and downregulation of death-associated protein kinase 1 (DAPK1) and tissue inhibitor of metalloproteinase 2 (TIMP2) (28). Additionally, in corticotroph tumors, RNA-sequencing showed a decrease in RNA expression of secreted frizzled-related protein 2 (SFRP2), which may promote tumorigenesis by upregulating Wnt signaling (29). The transcriptional profile of lactotroph tumors showed activation of estrogen receptor signaling, oxidative phosphorylation signaling, and eukaryotic translation initiation factor (EIF) signaling. Network analysis of upstream regulators determined that potential pathogenic drivers may include early growth response 1 (EGR1), protein kinase cAMP-activated catalytic subunit Alpha (PRKACA), paired like homeodomain 2 (PITX2), cAMP responsive element binding protein 1 (CREB1), and Jun D (JUND) proto-oncogene, an AP-1 transcription factor subunit (30).
In addition to evaluating specific genes, pathways, and PitNET types, transcriptomic data has been used to cluster PitNETs based on molecular subtype. Consensus clustering of transcriptomic data from 117 PitNETs of all hormonal subtypes revealed three molecular subtypes of tumors defined by biological processes: Group I – signaling pathways, Group II – metabolic processes, and Group III – immune responses. Each group had different immune profiles, and Group III had the worst prognosis even though these tumors were smaller (31). Future investigation of the role of non-coding, long non-coding, micro, and circulating RNAs in PitNET biology represents a new frontier for transcriptional profiling of PitNETs (32).
Single cell RNA sequencing (scRNA-seq) has also been used to investigate biological pathways related to invasive PitNETs. Previous work has shown that silent corticotroph PitNETs have been associated with an invasive phenotype; scRNA-seq revealed that these tumors express epithelial to mesenchymal transition genes, which may be driving tumor invasion (20). scRNA-seq has also been utilized to more robustly identify the heterogeneous biology of PitNETs. For example, when analyzing tumor cells from PIT1-lineage tumors, expression of hormone-encoding genes represented the majority of variation between tumors. There were four major clusters of non-PIT-1 tumor cells, and of the three clusters with majority TPIT-lineage tumor cells, one had significantly elevated Granzyme K (GZMK) expression, suggesting a possible novel subtype of corticotroph tumor. The fourth cluster of non-PIT-1 tumor cells was predominantly composed of SF-1 lineage cells with overexpression of follicle stimulating hormone subunit beta (FSHB). Additionally, within the tumor microenvironment, two distinct tumor-associated macrophage (TAM) clusters were enriched in PitNETs, one with pro-inflammatory M1 features and the other with immunosuppressive M2 marker upregulation (SPP1, TREM2, and CX3CR1). This finding suggests that depletion of TAMs or macrophage repolarization may be therapeutically relevant in PitNET treatment. In addition, stress response pathways were upregulated in T cells, suggesting functional exhaustion. This finding suggests that certain PitNET subtypes may be responsive to immune checkpoint blockade and other relevant tumor microenvironment modulating therapies (33).
Through the integration of scRNA-seq and single cell genomic sequencing, transcriptional profiles of normal endocrine cells (gonadotrophs, somatotrophs, and lactotrophs) to cognate tumor cells revealed several tumor-related genes such as adhesion molecule with Ig like domain 2 (AMIGO2), zinc finger protein 36 (ZFP36), BTG anti-proliferation factor 1 (BTG1), and disks large MAGUK scaffold protein 5 (DLG5) (34). Although 62% of tumors harbored CNAs, there was no significant intratumoral CNA heterogeneity (34). Although single cell molecular analyses have been utilized extensively to reveal the underlying biology and microenvironment of several cancer types and central nervous system tumors, there are only a few robust studies analyzing PitNETs at a single cell resolution. Further work in this area will likely lead to a more sophisticated understanding of PitNET tumorigenesis, especially with regard to differences between hormonal subtypes, tumor microenvironment, the immune landscape, and molecular drivers.
Epigenetic profiling
While genomic mutations in PitNETs are relatively uncommon, epigenetic dysregulation influencing gene expression, hormonal activity, and tumor behavior has emerged as a critical mechanism of PitNET pathogenesis (35). Epigenetic changes—such as in DNA methylation, histone modifications, and chromatin remodeling—are key modulators of transcriptional activity and cellular identity in both normal pituitary cells and tumors (35). Indeed, the activity of lineage-specific transcription factors such as PIT1, SF1, and TPIT is modulated by epigenetic marks, and clustering of PitNETs profiled by methylation array separated tumors by TF lineage (36).
Many studies have reported epigenetic changes in numerous genes associated with cell growth, cell signaling, and cell cycle signaling, including cyclin dependent kinase 1 (CDK1), cyclin dependent kinase inhibitor 1B (CDKN1B), cyclin dependent kinase inhibitor 2A (CDKN2A), cyclin dependent kinase inhibitor 2C (CDKN2C), growth arrest and DNA damage inducible gamma (GADD45G), Ras association domain family member 1 (RASSF1A), Ras association domain family member 3 (RASSF3), DAPK, pituitary tumor transforming gene 1 (PTTG1), maternally expressed 3 (MEG3), and fibroblast growth factor receptor 2 (FGFR2) (37–51). More aggressive PitNETs, defined by larger size and invasiveness, have been associated with the overexpression of DNA methyltransferases 1/3A (DNMT1/3A) and promoter hypermethylation of tumor suppressor genes (52). The first genome-wide methylation analysis of PitNETs in 2012 identified differentially methylated genes in nonfunctioning, GH-, and PRL-secreting PitNETs. Specifically, HHIP like 1 (HHIPL1) and transcription factor AP-2 epsilon (TFAP2E) were hypermethylated in nonfunctioning tumors (53). Multiple studies have shown that these nonfunctional tumors have global hypermethylation compared to hormonally active tumors (53–55). However, invasive nonfunctioning tumors have more hypomethylated cytosine-phosphate guanine (CpGs) sites compared to noninvasive nonfunctioning tumors (54), reminiscent of the global hypomethylation observed in many cancers (56). Biological pathways that were differentially methylated between invasive and noninvasive PitNETs included homophilic cell adhesion, cell–cell adhesion, and biological adhesion. The Polypeptide N-acetylgalactosaminyltransferase 9 (GALNT9) promoter was also found to be methylated with corresponding decreased RNA expression in invasive tumors, making GALNT9 expression a potential therapeutic target (55).
Although telomerase reverse transcriptase (TERT) promoter mutation is a marker of aggressiveness in numerous cancers and central nervous system tumors, the role of TERT promoter alterations such as methylation has been debated in PitNETs. In 2018, a study with 101 patients found no relationship between TERT promoter mutation or methylation and outcomes in patients with PitNETs (57). However, in a 2019 study analyzing 70 patients, TERT promoter methylation was associated with disease progression and shorter progression free survival (58, 59). Other common epigenetic biomarkers in brain tumors such as glioma include O6-methylguanine-DNA methyltransferase (MGMT) promoter methylation, which is related to response to temozolomide (TMZ) therapy. In contrast, in PitNETs the relationship between MGMT methylation status and prognosis or response to TMZ remains controversial (60–63).
Despite the ongoing debate surrounding prognostic epigenetic biomarkers like MGMT in PitNETs, the broader role of the epigenetic machinery itself presents a compelling target for therapeutic intervention. Importantly, DNA methyltransferase (DNMT) inhibitors and histone deacetylase (HDAC) inhibitors have demonstrated efficacy in other central nervous system tumors like glioblastoma and may be clinically relevant for the treatment of aggressive PitNETs (64, 65). While not yet clinically validated in PitNETs, DNMT and HDAC inhibitor therapies could be particularly beneficial when conventional therapies fail. Preclinical PitNET models will be essential in determinng whether modulation of the epigenetic landscape can suppress tumor proliferation, reduce hormonal hypersecretion, or enhance sensitivity to standard treatments such as temozolomide. As we further study PitNET epigenetics, targeted manipulation of regulators such as DNMTs and HDACs may emerge as a viable therapeutic strategy within a precision medicine framework.
Proteomic analysis
Proteomic analysis provides a direct readout of the functional state of cells by quantifying proteins and their post-translational modifications. In PitNETs, proteomic analyses offer unique insights into tumor activity, cellular heterogeneity, and treatment response.
Advanced mass spectrometry (MS)-based techniques, including tandem MS and data-independent acquisition (DIA), have enabled high-throughput profiling of PitNET proteomes and post-translational modifications. MS analysis reveals that nonfunctioning PitNETs have 2,000–6,000 differentially expressed proteins compared to normal pituitary glands (66, 67). Proteomic methods have also been used to identify the role of phosphorylation of proteins in nonfunctioning PitNETs. For example, phosphorylation of β-catenin at Serine552 is associated with aggressive disease characterized by invasion and recurrence (68). Meanwhile, comparison of nonfunctioning tumors to normal pituitary glands revealed 595 differentially phosphorylated proteins associated with biological pathways such as the spliceosome pathway, RNA transport pathway, and proteoglycans in cancer (69). Ubiquitination is another post-translational modification that has been investigated in PitNET biology. Ubiquitinated proteins in PitNETs were most involved in biological pathways such as the Phosphatidylinositol 3-kinase/protein kinase B (PI3K/AKT) pathway, Hippo (Hpo) pathway, ribosome signaling pathway, and nucleotide excision repair (70).
Alterations of specific protein abundances and functions have been investigated to identify their role in tumorigenesis in PitNETs. For example, hematopoietic cell signal transducer 1 (Hint1) is a protein marker that was found to have high expression in invasive PitNETs, especially those that expressed vascular endothelial growth factor (VEGF) and fetal liver kinase 1 (Flk1) (71). Invasive tumors were also found to have higher expression of cluster of differentiation 206 (CD206), a M2-macrophage marker, compared to noninvasive tumors based on immunohistochemical staining (72). Several protein components of the Notch pathway were altered in prolactinomas, in addition to increased expression of PIT1 and survival factor phosphoprotein associated with glycosphingolipid-enriched microdomains 1 (PAG1) and decreased expression of E-cadherin and N-cadherin (73).
Nitroproteomics is a subfield of proteomics that specifically studies nitropeptides and nitroproteins, which are often markers of oxidative damage and can be associated with tumorigenesis. In studies investigating nitroproteins in PitNETs, several nitroproteins and other proteins that interact with nitroproteins in nonfunctioning PitNETs were discovered using a nitrotyrosine affinity column (NTAC) (74, 75). Analysis of nitroproteins is important since identification of post-translational modifications such as nitrosylation may suggest potential new avenues for targeted therapy (76). Further work to identify the extent of the role of nitroproteomics in PitNET biology and tumorigenesis is warranted.
Metabolomics
Metabolomics—the comprehensive profiling of small-molecule metabolites in biological samples—provides a dynamic snapshot of cellular metabolism and its interaction with the tumor microenvironment. In PitNETs, metabolomic analysis has begun to uncover metabolic adaptations associated with hormone synthesis, tumor growth, and treatment resistance (77). Metabolomic methods such as matrix-assisted laser desorption/ionization (MALDI) mass spectrometry imaging have been used to confirm excess hormone production and classify PitNETs within 30 min (78). In patients with Cushing’s disease, biomarkers such as pyridoxate, deoxycholic acid, and 3-methyladipate were altered in plasma samples (79). Urine metabolites were analyzed using gas chromatography mass spectrometry system in prolactinoma patients, which showed an elevation of urinary 17-ketosteroids and all estrogen metabolite concentrations, as well as the ratios of delta 5/delta 4-steroids and 5 beta/5 alpha- hydrogensteroids (80). These findings have implications for understanding tumor biology, the systemic effect of disease, and identification of measurable biomarkers. For instance, PitNETs are defined by a distinct metabolic profile with higher succinic and lactic acid (72). These finding suggest possible mechanisms of disease development and progression as well as identification of biomarkers for diagnosis and targeted therapy. Although still an emerging field in pituitary tumor research, metabolomics holds significant promise for identifying biomarkers and therapeutic vulnerabilities, particularly in combination with other -omic methods.
Integrative Multiomic analysis
The advent of high-throughput -omics technologies has revolutionized our understanding of PitNETs, enabling comprehensive analyses at multiple molecular levels. These technologies each offer distinct advantages and limitations in terms of resolution, sensititivty, sample input, cost, and use-case. Table 2 provides a comparative overview of commonly used technologies across omics layers in an effort highlight pragmatic and methodological constraints across PitNET research. Integrative -omic analysis provides a holistic view of the molecular landscape of PitNETs, facilitates identification of biomarkers, elucidates complex regulatory networks, and uncovers potential therapeutic targets. Recent studies have demonstrated that such integrative analyses can reveal distinct molecular subtypes of PitNETs, improve correlations between molecular profiles and clinical outcomes, and provide insights into tumorigenesis and progression (Figure 2).
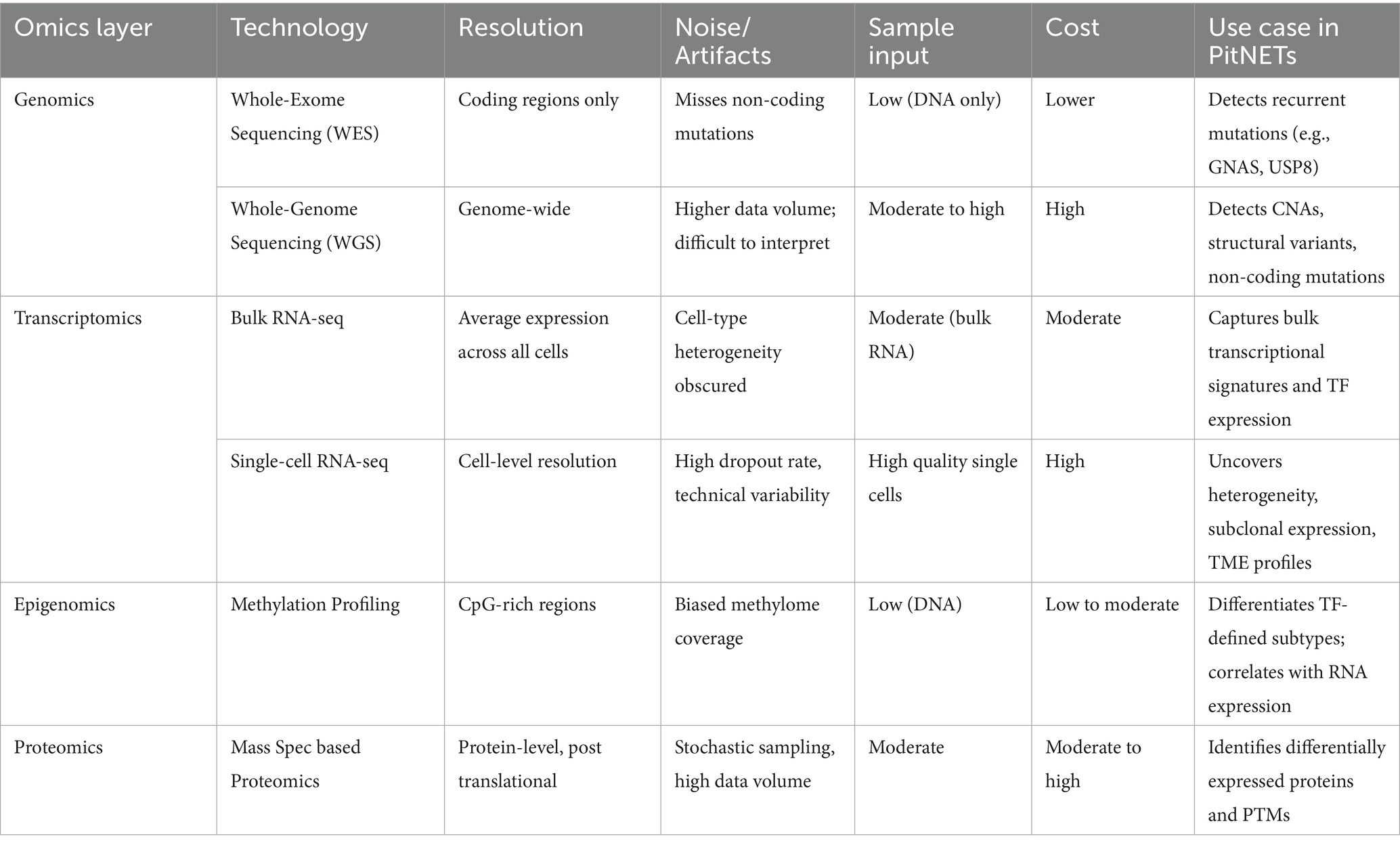
Table 2. Omics technology comparison table.
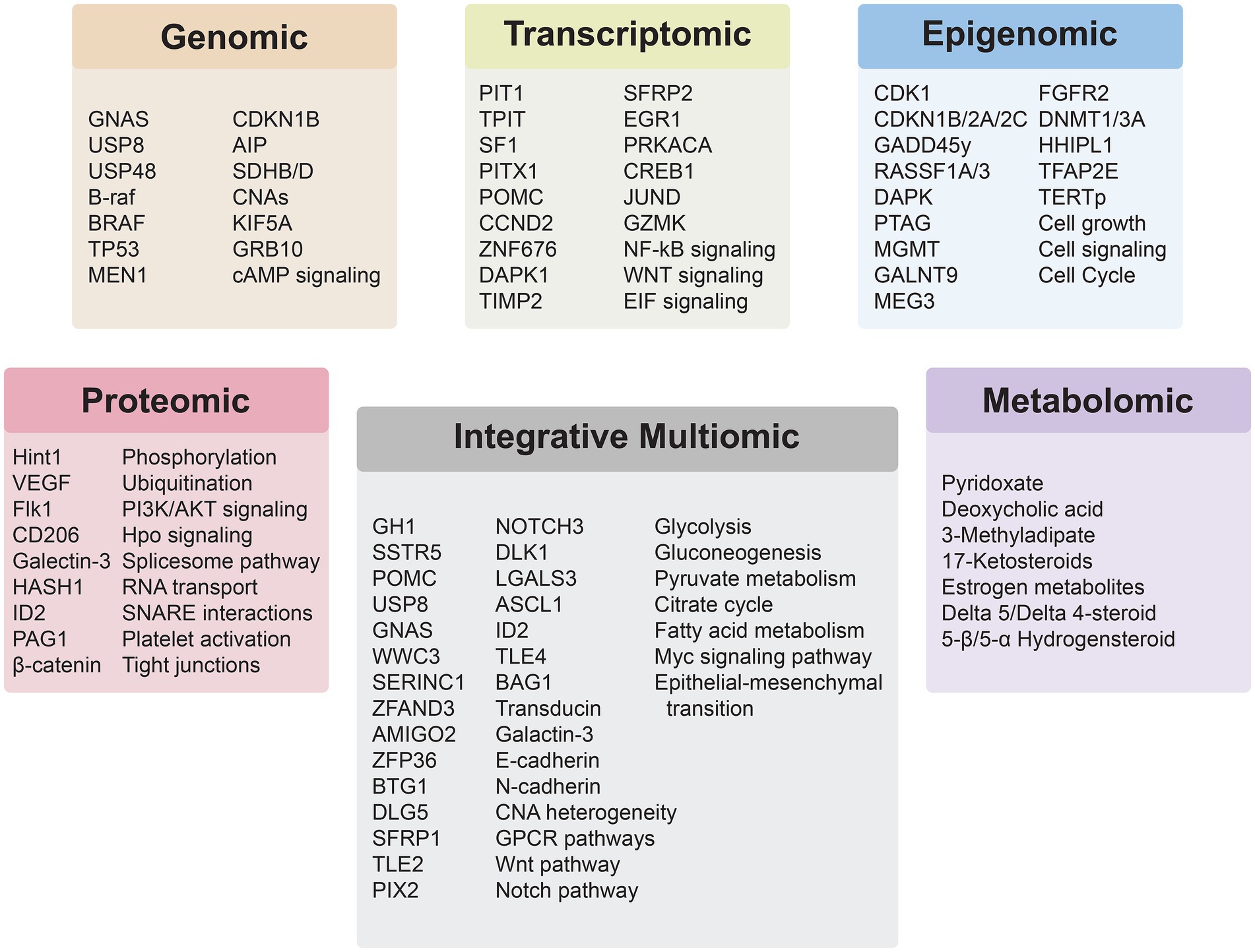
Figure 2. Insights derived from the application of individual and integrative multiomics analyses for PitNETs.
As Table 3 summarizes, each PitNET subtype is characterized by distinct molecular features across genomic, transcriptomic, epigenomic, proteomic, and metabolomic layers. Despite their differences, these multiomic signatures converge on shared biological pathways across subtype. For instance, somatotroph tumors exhibit GNAS mutations, PIT1-driven transcription, and enrichment of proteins in PI3K/AKT signaling, which collectively support growth hormone hypersecretion via cAMP signaling and metabolic reprogramming (81). Corticotroph tumors exhibit USP8 mutations, upregulation of proopiomelanocortin (POMC), transcriptomic changes in Wnt regulators like SFRP2, and proteomic changes in Galectin-3 and ID2, linking chromatin remodeling and epithelial-to-mesenchymal (EMT) transition with sustained ACTH hypersecretion (82–84). Finally, lactotroph tumors with FIPA or SF3B1 mutations and estrogen receptor activation display epigenetic change (HMGA regulation via chromatin architecture) and proteomic shifts in Galectin-3, HADH1, and ID2, linking genetic mutations and estrogen signaling to altered tumor epigenetics and protein expression patterns that drive tumor aggressiveness and treatment resistance (85, 86). These convergences evidently highlight shared mechanisms such as hormone hypersecretion, chromatin remodeling, biological pathway activation, and metabolic rewiring across tumor types, underscoring the translational value of integrative multiomic analysis in PitNET research. Additional molecular studies across different subtypes remain necessary, as certain subtypes such as Gonadotroph PitNETs lack any published molecular data (87).
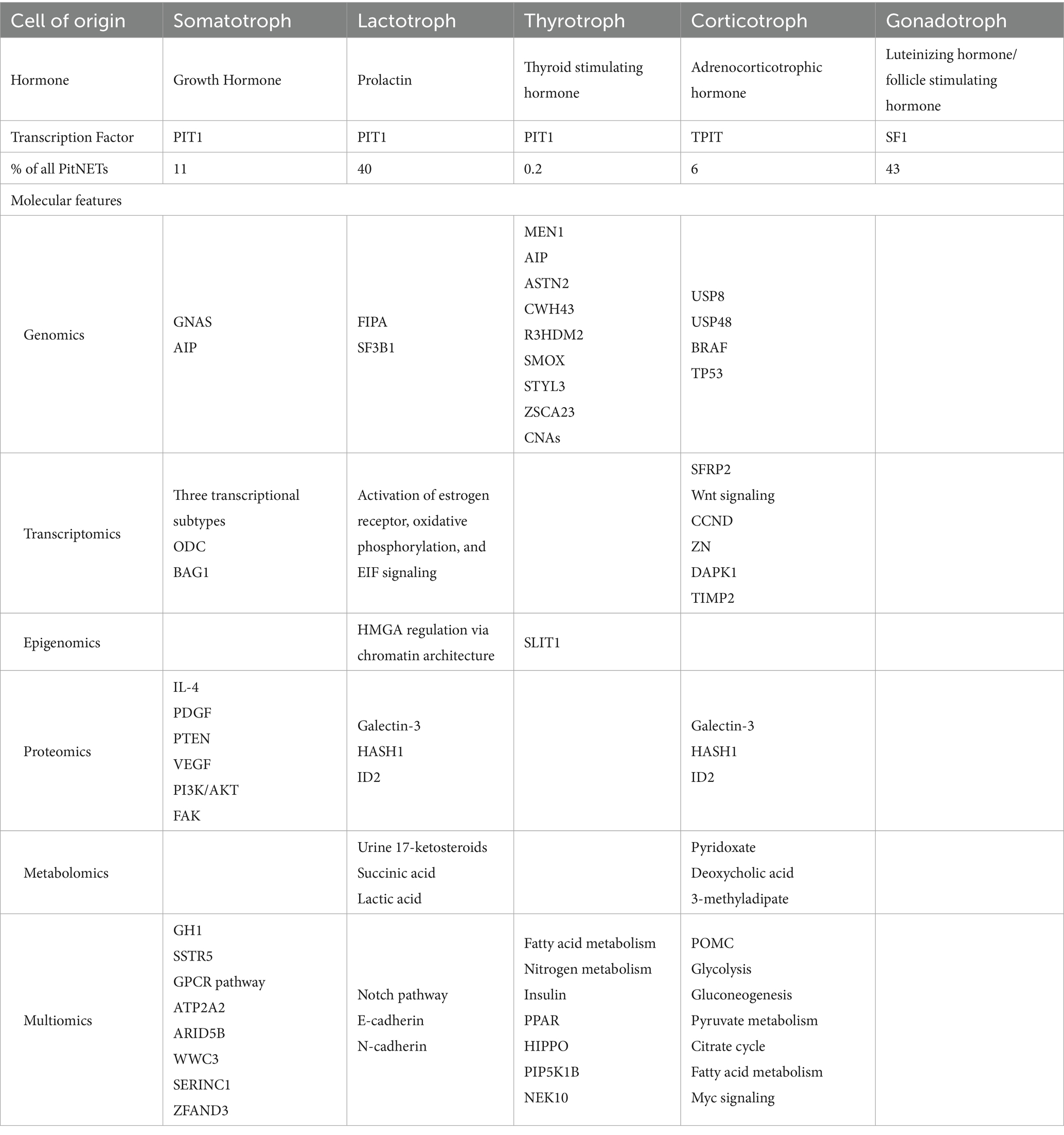
Table 3. Molecular features of PitNETs by hormonal expression.
Although genomic profiling suggests infrequent rates of somatic mutations in PitNETs, CNAs are common among all TF-lineage subtypes. Integrating analysis of methylation and transcriptional data suggests that hypomethylation of promoter regions is associated with increased RNA expression of GH1 and Somatostatin Receptor subtype 5 (SSTR5) in GH-secreting PitNETs and POMC in ACTH-secreting PitNETs (88). In a 2020 multi-omic study, three molecular classes of PitNETs were identified by integrating somatic mutations, chromosomal alterations, and profiling of the miRNAome, methylome, and transcriptome (89). This classification scheme clustered PitNETs similar to the classification based on TF lineage. Prognostic analysis identified that USP8 wildtype (WT) compared to USP8 mutant corticotroph PitNETs were more aggressive with invasive properties (89). The transcriptome of these invasive corticotrophs was enriched for genes associated with epithelial-mesenchymal-transition, consistent with their invasive clinical behavior (89). Gene ontology analysis in a transcriptomic and proteomic integrated analysis of GNAS mutant vs. wildtype somatotrophs suggested that GNAS mutations may impact endocrine features through induction of G protein-coupled receptor (GPCR) pathways. Higher protein expression of WW and C2 domain-containing protein-3 (WWC3), serine incorporator 1 (SERINC1), and zinc finger AN1-type containing 3 (ZFAND3) was correlated with increased tumor volume after somatostatin analog treatment (90). Recurrence as a clinical marker of aggressive disease has also been investigated utilizing multiomic methodologies. A robust longitudinal study of primary and recurrent PitNETs from the same patient determined primary and recurrent PitNETs to have similar genomic profiles but divergent transcriptomic profiles (91). Interestingly, several metabolic pathways that were differentially expressed among primary and recurrent tumors based on transcriptional data did not seem to be regulated by methylation, raising the possibility of alternative regulatory mechanisms that warrant further investigation (91).
Multiomic analyses have also incorporated both proteomic and transcriptomic data to further understand PitNET biology. For example, nonfunctioning PitNETs had almost 300 differentially expressed genes and 50 differentially expressed proteins compared to controls including secreted frizzled-related protein 1 (SFRP1), transducin like enhancer of split 2 (TLE2), PITX2, Notch receptor 3 (NOTCH3), and delta like non-canonical Notch ligand 1 (DLK1) (92). These findings suggest potential critical molecular pathways implicated in this tumor type such as the Wnt and Notch pathways. Integrative proteomic and transcriptomic analysis has also been used to analyze metastatic PitNETs, which led to the identification of almost 5,000 differentially expressed genes, and the downregulation of beta-galactoside binding protein galactin-3. Other genes that may play important roles in metastatic PitNETs include lectin, galactoside-binding, soluble, 3 (LGALS3), achaete-scute family bHLH transcription factor 1 (ASCL1), ID2, and transducin like enhancer of split 4 (TLE4) (93). Lastly, transcriptomic and proteomic analysis of prolactinomas compared to normal pituitary glands identified a unique transcriptomic and proteomic profile. Notably, several components of the Notch pathway were altered in prolactinomas, along with increased expression of survival factor BCL2 associated athanogene 1 (BAG1) and decreased expression of E-cadherin and N-cadherin (73).
Metabolomics has been used alongside other -omic methods such as proteomics and lipidomics to delve further into the mechanisms of PitNET pathogenesis. In ACTH-secreting PitNETs, integrated analysis identified that these tumors were significantly enriched in protein-metabolite joint pathways such as glycolysis/gluconeogenesis, pyruvate metabolism, citrate cycle, and fatty acid metabolism (94). The Myc signaling pathway was also identified to have a significant role in the metabolic changes and tumorigenesis of these tumors (94). A broader study using desorption electrospray ionization (DESI-MS) derived phospholipid signals that differed between gray matter, white matter, gliomas, meningiomas and pituitary tumors. Principal component analysis of lipid and metabolite profiles from this analysis were able to separate different tumor types: gliomas, meningiomas, and pituitary tumors (95).
However, while these studies underscore the value of integrative multiomics, they also highlight the significant computational hurdles in merging heterogenous omic datasets. Despite the growing number of multi-omic studies in PitNETs, integration and standardization across datasets remain computationally challenging, as omics data is inherently heterogenous. Several bioinformatic tools have been developed to address these issues. Multi-omics factor analysis uses unsupervised latent factor modeling to identify hidden sources of variation across omics layers (96). Similarity network fusion constructs networks of samples and merges these networks effectively to discover subtypes (97). By contrast, iClusterPlus applies joint latent variable modeling to integrate multiple subtypes of genomic data for subtype identification (98). Unfortunately, these distinct data fusion techniques differ in scalability, handling of missing data, and interpretability. Moreover, these methods are rarely tailored to PitNET-specific datasets, which tend to be small and sparse.
Standardization of data in PitNET omics research faces similar issues. Batch effects, inconsistent normalization strategies, and variable bioinformatics pipelines undermine reproducibility of data. Transcriptomic analysis heavily relies on normalization and batch correction tools like ComBat or Harmony (99, 100). Proteomic and epigenomic analyses use quantile normalization and reference-based scaling to address technical variability (101). Collectively, these techniques’ inconsistencies can complicate downstream integration efforts. Hence, adhering to data frameworks such as the NIH’s Findable, Accessible, Interoperable, Reusable (FAIR) principles, standardizing pipelines, and reporting metadata in PitNET research would allow for increased reproducibility and comparability of data, facilitating the development of robust PitNET-specific computational pipeline that provide clinically meaningful data.
In parallel with efforts to integrate and standardize multiomic workflows, artificial intelligence (AI) and machine learning (ML) have emerged as powerful tools for analyzing complex multi-omic datasets. Although still in the nascent stages of adoption in PitNET research, these methods are beginning to prove extremely useful. Several studies have already utilized AI and ML to create robust PitNET classifiers for risk stratification and diagnosis. Wang et al. used LASSO regression and Support Vector Machine Recursive Feature Elimination to develop a Programmed Cell Death-associated index (PCDI) classifier that outperforms traditional prognostic models in identifying invasive PitNETs with a high degree of accuracy (102). In another study, Li et al. used radiomic features derived from T2-weighted MRI to construct a Gaussian process model capable of preoperatively predicting histological subtypes of PitNETs, such as prolactinoma (103). Despite these promising results, the translational potential of these approaches is limited by the paucity of PitNET datasets. Collaborative future modeling efforts may allow for more robust and accurate model construction and generalization.
Integrative multi-omics analyses have significantly advanced our understanding of PitNETs by revealing multiple molecular subtypes and the complex regulatory networks that underlie tumor behavior. Building upon these approaches, spatial omics technologies are emerging as vital tools for resolving tumor heterogeneity in its native context. Spatial transcriptomics and proteomics offer significant resolution advancement for characterizing intratumoral heterogeneity and tumor microenvironment architecture in PitNETs. For instance, spatial transcriptomics could distinguish between non-invasive and invasive PitNET phenotypes by localizing EMT markers. Similarly, spatial proteomic analysis could enable the visualization of PTMs throughout the invasive PitNET front. These tools have the potential to refine the current understanding of PitNET pathophysiology and support the development of spatially-informed, precision medicine strategies.
Translational gaps
While multiomics PitNET research has yielded invaluable biological insights, a significant gap remains between academic discovery and clinical translation. Cost and infrastructure requirements for generating and analyzing multi-layered omics data remains prohibitive, especially outside of academic centers. Governmental regulatory pathways for clinical grade omics assays are still evolving, with no PitNET omics-based biomarker panels still having received FDA clearance. Clinical trials for multiomic biomarker validation also remain rare and underpowered.
Conclusion
In conclusion, the integration of multi-omics technologies has profoundly advanced our understanding of PitNETs, offering a comprehensive view of their molecular landscape. By combining data from genomics, transcriptomics, proteomics, epigenomics, and metabolomics, researchers have identified distinct molecular subtypes, unveiled regulatory networks, and discovered novel biomarkers, thereby enhancing diagnostic precision and informing therapeutic strategies. Clinically, these integrative approaches hold promise for the development of personalized medicine in PitNET management, which is a critical need, in particular for recurrent tumors and tumors not cured by the current standard of care. The ability to correlate multiomic profiles with clinical outcomes facilitates more accurate prognostication and the potential for tailored treatment regimens. Looking forward, the continued evolution of computational tools and machine learning algorithms will be critical in managing the complexity of multiomic data, enabling real-time integration and interpretation in clinical settings. Advancements in single-cell and spatial omics technologies are expected to further define tumor heterogeneity and microenvironmental interactions, providing deeper insights into PitNET pathogenesis. Collectively, these developments herald a new era in PitNET management, where multiomic integration becomes central to patient-specific diagnosis, prognosis, and therapy.
Author contributions
SP: Conceptualization, Methodology, Visualization, Writing – original draft, Writing – review & editing. SSP: Methodology, Visualization, Writing – original draft, Writing – review & editing. ZZ: Methodology, Validation, Writing – original draft, Writing – review & editing. JS: Writing – original draft, Writing – review & editing, Project administration. AK: Conceptualization, Supervision, Writing – original draft, Writing – review & editing, Funding acquisition. BP: Conceptualization, Supervision, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. AK: Alvin J. Siteman Cancer Research Fund (GF0010215), the Christopher Davidson and Knight Family Fund, and the Duesenberg Research Fund.
Conflict of interest
AK is a consultant for Monteris Medical and has received nonrelated research grants from Stryker for study of dural substitute.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Price, M, Ballard, C, Benedetti, J, Neff, C, Cioffi, G, Waite, KA, et al. Cbtrus statistical report: primary brain and other central nervous system tumors diagnosed in the United States in 2017–2021. Neuro-Oncology. (2024) 26:vi1–vi85. doi: 10.1093/neuonc/noae145
2. Asa, SL, Mete, O, Perry, A, and Osamura, RY. Overview of the 2022 who classification of pituitary tumors. Endocr Pathol. (2022) 33:6–26. doi: 10.1007/s12022-022-09703-7
3. Mouchtouris, N, Smit, RD, Piper, K, Prashant, G, Evans, JJ, and Karsy, M. A review of multiomics platforms in pituitary adenoma pathogenesis. Front Biosci. (2022) 27:77. doi: 10.31083/j.fbl2703077
4. Tang, C, Zhong, C, Zhu, J, Yuan, F, Yang, J, Xu, Y, et al. Gnas mutations suppress cell invasion by activating Meg3 in growth hormone-secreting pituitary adenoma. Oncol Res. (2024) 32:1079–91. doi: 10.32604/or.2024.046007
5. Reincke, M, Sbiera, S, Hayakawa, A, Theodoropoulou, M, Osswald, A, Beuschlein, F, et al. Mutations in the deubiquitinase gene Usp8 cause Cushing's disease. Nat Genet. (2015) 47:31–8. doi: 10.1038/ng.3166
6. Perez-Rivas, LG, Theodoropoulou, M, Ferraù, F, Nusser, C, Kawaguchi, K, Stratakis, CA, et al. The gene of the ubiquitin-specific protease 8 is frequently mutated in adenomas causing Cushing's disease. J Clin Endocrinol Metab. (2015) 100:E997–E1004. doi: 10.1210/jc.2015-1453
7. Perez-Rivas, LG, Simon, J, Albani, A, Tang, S, Roeber, S, Assié, G, et al. Tp53 mutations in functional Corticotroph tumors are linked to invasion and worse clinical outcome. Acta Neuropathol Commun. (2022) 10:139. doi: 10.1186/s40478-022-01437-1
8. Chen, J, Jian, X, Deng, S, Ma, Z, Shou, X, Shen, Y, et al. Identification of recurrent Usp48 and Braf mutations in Cushing's disease. Nat Commun. (2018) 9:3171. doi: 10.1038/s41467-018-05275-5
9. Dénes, J, and Korbonits, M. The clinical aspects of pituitary tumour genetics. Endocrine. (2021) 71:663–74. doi: 10.1007/s12020-021-02633-0
10. Chin, SO, Chik, C, and Tateno, T. Pituitary neuroendocrine tumors in multiple endocrine neoplasia. Endocrinol Metab (Seoul). (2025) 40:39–46. doi: 10.3803/EnM.2024.2074
11. Stratakis, CA, Tichomirowa, MA, Boikos, S, Azevedo, MF, Lodish, M, Martari, M, et al. The role of germline Aip, Men1, Prkar1a, Cdkn1b and Cdkn2c mutations in causing pituitary adenomas in a large cohort of children, adolescents, and patients with genetic syndromes. Clin Genet. (2010) 78:457–63. doi: 10.1111/j.1399-0004.2010.01406.x
12. Xekouki, P, and Stratakis, CA. Succinate dehydrogenase (Sdhx) mutations in pituitary tumors: could this be a new role for mitochondrial complex II and/or Krebs cycle defects? Endocr Relat Cancer. (2012) 19:C33–40. doi: 10.1530/erc-12-0118
13. Daly, AF, Tichomirowa, MA, Petrossians, P, Heliövaara, E, Jaffrain-Rea, ML, Barlier, A, et al. Clinical characteristics and therapeutic responses in patients with germ-line Aip mutations and pituitary adenomas: an international collaborative study. J Clin Endocrinol Metab. (2010) 95:E373–83. doi: 10.1210/jc.2009-2556
14. Hernández-Ramírez, LC, Gabrovska, P, Dénes, J, Stals, K, Trivellin, G, Tilley, D, et al. Landscape of familial isolated and young-onset pituitary adenomas: prospective diagnosis in Aip mutation carriers. J Clin Endocrinol Metab. (2015) 100:E1242–54. doi: 10.1210/jc.2015-1869
15. Song, Z-J, Reitman, ZJ, Ma, Z-Y, Chen, J-H, Zhang, Q-L, Shou, X-F, et al. The genome-wide mutational landscape of pituitary adenomas. Cell Res. (2016) 26:1255–9. doi: 10.1038/cr.2016.114
16. Bi, WL, Horowitz, P, Greenwald, NF, Abedalthagafi, M, Agarwalla, PK, Gibson, WJ, et al. Landscape of genomic alterations in pituitary adenomas. Clin Cancer Res. (2017) 23:1841–51. doi: 10.1158/1078-0432.Ccr-16-0790
17. Oh, JY, Osorio, RC, Jung, J, Carrete, L, Choudhary, N, Lad, M, et al. Transcriptomic profiles of normal pituitary cells and pituitary neuroendocrine tumor cells. Cancers (Basel). (2022) 15:110. doi: 10.3390/cancers15010110
18. Tebani, A, Jotanovic, J, Hekmati, N, Sivertsson, Å, Gudjonsson, O, Edén Engström, B, et al. Annotation of pituitary neuroendocrine tumors with genome-wide expression analysis. Acta Neuropathol Commun. (2021) 9:181. doi: 10.1186/s40478-021-01284-6
19. Lamolet, B, Pulichino, AM, Lamonerie, T, Gauthier, Y, Brue, T, Enjalbert, A, et al. A pituitary cell-restricted T box factor, Tpit, activates Pomc transcription in cooperation with Pitx Homeoproteins. Cell. (2001) 104:849–59. doi: 10.1016/s0092-8674(01)00282-3
20. Zhang, D, Hugo, W, Bergsneider, M, Wang, MB, Kim, W, Vinters, HV, et al. Single-cell Rna sequencing in silent Corticotroph tumors confirms impaired Pomc processing and provides new insights into their invasive behavior. Eur J Endocrinol. (2022) 187:49–64. doi: 10.1530/eje-21-1183
21. Pulichino, AM, Vallette-Kasic, S, Tsai, JP, Couture, C, Gauthier, Y, and Drouin, J. Tpit determines alternate fates during pituitary cell differentiation. Genes Dev. (2003) 17:738–47. doi: 10.1101/gad.1065703
22. Zhu, X, Gleiberman, AS, and Rosenfeld, MG. Molecular physiology of pituitary development: signaling and transcriptional networks. Physiol Rev. (2007) 87:933–63. doi: 10.1152/physrev.00006.2006
23. Toda, M, Tamura, R, and Toda, M. Recent progress in stem cell research of the pituitary gland and pituitary adenoma. Endocrine. (2020) 1:49–57. doi: 10.3390/endocrines1010006
24. Zhao, L, Bakke, M, Krimkevich, Y, Cushman, LJ, Parlow, AF, Camper, SA, et al. Steroidogenic factor 1 (Sf1) is essential for pituitary Gonadotrope function. Development. (2001) 128:147–54. doi: 10.1242/dev.128.2.147
25. Jotanovic, J, Tebani, A, Hekmati, N, Sivertsson, Å, Lindskog, C, Uhlèn, M, et al. Transcriptome analysis reveals distinct patterns between the invasive and noninvasive pituitary neuroendocrine tumors. J Endocr Soc. (2024) 8:bvae040. doi: 10.1210/jendso/bvae040
26. Wierinckx, A, Auger, C, Devauchelle, P, Reynaud, A, Chevallier, P, Jan, M, et al. A diagnostic marker set for invasion, proliferation, and aggressiveness of prolactin pituitary tumors. Endocr Relat Cancer. (2007) 14:887–900. doi: 10.1677/erc-07-0062
27. Galland, F, Lacroix, L, Saulnier, P, Dessen, P, Meduri, G, Bernier, M, et al. Differential gene expression profiles of invasive and non-invasive non-functioning pituitary adenomas based on microarray analysis. Endocr Relat Cancer. (2010) 17:361–71. doi: 10.1677/erc-10-0018
28. de Araújo, LJ, Lerario, AM, de Castro, M, Martins, CS, Bronstein, MD, Machado, MC, et al. Transcriptome analysis showed a differential signature between invasive and non-invasive corticotrophinomas. Front Endocrinol (Lausanne). (2017) 8:55. doi: 10.3389/fendo.2017.00055
29. Wu, Q, Yin, X, Zhao, W, Xu, W, and Chen, L. Downregulation of Sfrp2 facilitates Cancer Stemness and Radioresistance of glioma cells via activating Wnt/β-catenin signaling. PLoS One. (2021) 16:e0260864. doi: 10.1371/journal.pone.0260864
30. Hattori, Y, Tahara, S, Ozawa, H, Morita, A, and Ishii, H. Transcriptomic profiling of Lactotroph pituitary neuroendocrine tumors via Rna sequencing and ingenuity pathway analysis. Neuroendocrinology. (2024) 114:670–80. doi: 10.1159/000539017
31. Peng, J, Yuan, L, Kang, P, Jin, S, Ma, S, Zhou, W, et al. Comprehensive transcriptomic analysis identifies three distinct subtypes of pituitary adenomas: insights into tumor behavior, prognosis, and stem cell characteristics. J Transl Med. (2024) 22:892. doi: 10.1186/s12967-024-05702-w
32. Butz, H. Circulating noncoding Rnas in pituitary neuroendocrine tumors-two sides of the same coin. Int J Mol Sci. (2022) 23:5122. doi: 10.3390/ijms23095122
33. Yan, N, Xie, W, Wang, D, Fang, Q, Guo, J, Chen, Y, et al. Single-cell transcriptomic analysis reveals tumor cell heterogeneity and immune microenvironment features of pituitary neuroendocrine tumors. Genome Med. (2024) 16:2. doi: 10.1186/s13073-023-01267-3
34. Cui, Y, Li, C, Jiang, Z, Zhang, S, Li, Q, Liu, X, et al. Single-cell transcriptome and genome analyses of pituitary neuroendocrine tumors. Neuro-Oncology. (2021) 23:1859–71. doi: 10.1093/neuonc/noab102
35. Hauser, BM, Lau, A, Gupta, S, Bi, WL, and Dunn, IF. The Epigenomics of pituitary adenoma. Front Endocrinol (Lausanne). (2019) 10:290. doi: 10.3389/fendo.2019.00290
36. Mosella, MS, Sabedot, TS, Silva, TC, Malta, TM, Dezem, FS, Asmaro, KP, et al. DNA methylation-based signatures classify sporadic pituitary tumors according to Clinicopathological features. Neuro-Oncology. (2021) 23:1292–303. doi: 10.1093/neuonc/noab044
37. Lidhar, K, Korbonits, M, Jordan, S, Khalimova, Z, Kaltsas, G, Lu, X, et al. Low expression of the cell cycle inhibitor P27kip1 in normal corticotroph cells, corticotroph tumors, and malignant pituitary tumors. J Clin Endocrinol Metab. (1999) 84:3823–30. doi: 10.1210/jcem.84.10.6066
38. Zhou, Y, Zhang, X, and Klibanski, A. Genetic and epigenetic mutations of tumor suppressive genes in sporadic pituitary adenoma. Mol Cell Endocrinol. (2014) 386:16–33. doi: 10.1016/j.mce.2013.09.006
39. Kirsch, M, Mörz, M, Pinzer, T, Schackert, HK, and Schackert, G. Frequent loss of the Cdkn2c (P18ink4c) gene product in pituitary adenomas. Genes Chromosomes Cancer. (2009) 48:143–54. doi: 10.1002/gcc.20621
40. Hossain, MG, Iwata, T, Mizusawa, N, Qian, ZR, Shima, SW, Okutsu, T, et al. Expression of P18(Ink4c) is down-regulated in human pituitary adenomas. Endocr Pathol. (2009) 20:114–21. doi: 10.1007/s12022-009-9076-0
41. Simpson, DJ, Hibberts, NA, McNicol, AM, Clayton, RN, and Farrell, WE. Loss of Prb expression in pituitary adenomas is associated with methylation of the Rb1 Cpg island. Cancer Res. (2000) 60:1211–6.
42. Tateno, T, Nakano-Tateno, T, Ezzat, S, and Asa, SL. Ng2 targets tumorigenic Rb inactivation in Pit1-lineage pituitary cells. Endocr Relat Cancer. (2016) 23:445–56. doi: 10.1530/erc-16-0013
43. Nakayama, K, Ishida, N, Shirane, M, Inomata, A, Inoue, T, Shishido, N, et al. Mice lacking P27(Kip1) display increased body size, multiple organ hyperplasia, retinal dysplasia, and pituitary tumors. Cell. (1996) 85:707–20. doi: 10.1016/s0092-8674(00)81237-4
44. Zhang, X, Sun, H, Danila, DC, Johnson, SR, Zhou, Y, Swearingen, B, et al. Loss of expression of Gadd45 gamma, a growth inhibitory gene, in human pituitary adenomas: implications for tumorigenesis. J Clin Endocrinol Metab. (2002) 87:1262–7. doi: 10.1210/jcem.87.3.8315
45. Carrier, F, Georgel, PT, Pourquier, P, Blake, M, Kontny, HU, Antinore, MJ, et al. Gadd45, a P53-responsive stress protein, modifies DNA accessibility on damaged chromatin. Mol Cell Biol. (1999) 19:1673–85. doi: 10.1128/mcb.19.3.1673
46. Bahar, A, Bicknell, JE, Simpson, DJ, Clayton, RN, and Farrell, WE. Loss of expression of the growth inhibitory gene Gadd45gamma, in human pituitary adenomas, is associated with Cpg Island methylation. Oncogene. (2004) 23:936–44. doi: 10.1038/sj.onc.1207193
47. Qian, ZR, Sano, T, Yoshimoto, K, Yamada, S, Ishizuka, A, Mizusawa, N, et al. Inactivation of Rassf1a tumor suppressor gene by aberrant promoter Hypermethylation in human pituitary adenomas. Lab Investig. (2005) 85:464–73. doi: 10.1038/labinvest.3700248
48. Simpson, DJ, Clayton, RN, and Farrell, WE. Preferential loss of death associated protein kinase expression in invasive pituitary Tumours is associated with either Cpg Island methylation or homozygous deletion. Oncogene. (2002) 21:1217–24. doi: 10.1038/sj.onc.1205195
49. Zhang, X, Zhou, Y, and Klibanski, A. Isolation and characterization of novel pituitary tumor related genes: a Cdna representational difference approach. Mol Cell Endocrinol. (2010) 326:40–7. doi: 10.1016/j.mce.2010.02.040
50. Zhao, J, Dahle, D, Zhou, Y, Zhang, X, and Klibanski, A. Hypermethylation of the promoter region is associated with the loss of Meg3 gene expression in human pituitary tumors. J Clin Endocrinol Metab. (2005) 90:2179–86. doi: 10.1210/jc.2004-1848
51. Zhu, X, Lee, K, Asa, SL, and Ezzat, S. Epigenetic silencing through DNA and histone methylation of fibroblast growth factor receptor 2 in neoplastic pituitary cells. Am J Pathol. (2007) 170:1618–28. doi: 10.2353/ajpath.2007.061111
52. Ma, HS, Wang, EL, Xu, WF, Yamada, S, Yoshimoto, K, Qian, ZR, et al. Overexpression of DNA (Cytosine-5)-methyltransferase 1 (Dnmt1) and DNA (Cytosine-5)-methyltransferase 3a (Dnmt3a) is associated with aggressive behavior and hypermethylation of tumor suppressor genes in human pituitary adenomas. Med Sci Monit. (2018) 24:4841–50. doi: 10.12659/msm.910608
53. Duong, CV, Emes, RD, Wessely, F, Yacqub-Usman, K, Clayton, RN, and Farrell, WE. Quantitative, genome-wide analysis of the DNA Methylome in sporadic pituitary adenomas. Endocr Relat Cancer. (2012) 19:805–16. doi: 10.1530/erc-12-0251
54. Ling, C, Pease, M, Shi, L, Punj, V, Shiroishi, MS, Commins, D, et al. A pilot genome-scale profiling of DNA methylation in sporadic pituitary macroadenomas: association with tumor invasion and histopathological subtype. PLoS One. (2014) 9:e96178. doi: 10.1371/journal.pone.0096178
55. Gu, Y, Zhou, X, Hu, F, Yu, Y, Xie, T, Huang, Y, et al. Differential DNA Methylome profiling of nonfunctioning pituitary adenomas suggesting tumour invasion is correlated with cell adhesion. J Neuro-Oncol. (2016) 129:23–31. doi: 10.1007/s11060-016-2139-4
56. Toraño, EG, Petrus, S, Fernandez, AF, and Fraga, MF. Global DNA Hypomethylation in Cancer: review of validated methods and clinical significance. Clin Chem Lab Med. (2012) 50:1733–42. doi: 10.1515/cclm-2011-0902
57. Boresowicz, J, Kober, P, Rusetska, N, Maksymowicz, M, Goryca, K, Kunicki, J, et al. Telomere length and tert abnormalities in pituitary adenomas. Neuro Endocrinol Lett. (2018) 39:49–55.
58. Miyake, Y, Adachi, JI, Suzuki, T, Mishima, K, Araki, R, Mizuno, R, et al. Tert promoter methylation is significantly associated with Tert upregulation and disease progression in pituitary adenomas. J Neuro-Oncol. (2019) 141:131–8. doi: 10.1007/s11060-018-03016-8
59. Alzoubi, H, Minasi, S, Gianno, F, Antonelli, M, Belardinilli, F, Giangaspero, F, et al. Alternative lengthening of telomeres (alt) and telomerase reverse transcriptase promoter methylation in recurrent adult and primary pediatric pituitary neuroendocrine tumors. Endocr Pathol. (2022) 33:494–505. doi: 10.1007/s12022-021-09702-0
60. Kovacs, K, Scheithauer, BW, Lombardero, M, McLendon, RE, Syro, LV, Uribe, H, et al. Mgmt Immunoexpression predicts responsiveness of pituitary tumors to Temozolomide therapy. Acta Neuropathol. (2008) 115:261–2. doi: 10.1007/s00401-007-0279-5
61. Widhalm, G, Wolfsberger, S, Preusser, M, Woehrer, A, Kotter, MR, Czech, T, et al. O(6)-Methylguanine DNA methyltransferase Immunoexpression in nonfunctioning pituitary adenomas: are progressive tumors potential candidates for Temozolomide treatment? Cancer. (2009) 115:1070–80. doi: 10.1002/cncr.24053
62. Hirohata, T, Asano, K, Ogawa, Y, Takano, S, Amano, K, Isozaki, O, et al. DNA mismatch repair protein (Msh6) correlated with the responses of atypical pituitary adenomas and pituitary carcinomas to Temozolomide: the National Cooperative Study by the Japan Society for Hypothalamic and Pituitary Tumors. J Clin Endocrinol Metab. (2013) 98:1130–6. doi: 10.1210/jc.2012-2924
63. McCormack, AI, McDonald, KL, Gill, AJ, Clark, SJ, Burt, MG, Campbell, KA, et al. Low O6-methylguanine-DNA methyltransferase (Mgmt) expression and response to temozolomide in aggressive pituitary tumours. Clin Endocrinol. (2009) 71:226–33. doi: 10.1111/j.1365-2265.2008.03487.x
64. Kushihara, Y, Tanaka, S, Kobayashi, Y, Nagaoka, K, Kikuchi, M, Nejo, T, et al. Glioblastoma with high O6-methyl-guanine DNA methyltransferase expression are more immunologically active than tumors with low Mgmt expression. Front Immunol. (2024) 15:1328375. doi: 10.3389/fimmu.2024.1328375
65. Chen, R, Zhang, M, Zhou, Y, Guo, W, Yi, M, Zhang, Z, et al. The application of histone deacetylases inhibitors in glioblastoma. J Exp Clin Cancer Res. (2020) 39:138. doi: 10.1186/s13046-020-01643-6
66. Candy, NG, Ramezanpour, M, Bouras, G, Chegeni, N, Chataway, T, Ormsby, RJ, et al. Differential proteomic expression in non-functional pituitary neuroendocrine Tumours and pituitary glands. Rhinology. (2024) 62:750–8. doi: 10.4193/Rhin24.216
67. Cheng, T, Wang, Y, Lu, M, Zhan, X, Zhou, T, Li, B, et al. Quantitative analysis of proteome in non-functional pituitary adenomas: clinical relevance and potential benefits for the patients. Front Endocrinol (Lausanne). (2019) 10:854. doi: 10.3389/fendo.2019.00854
68. Rai, A, Yelamanchi, SD, Radotra, BD, Gupta, SK, Mukherjee, KK, Tripathi, M, et al. Phosphorylation of β-catenin at Serine552 correlates with invasion and recurrence of non-functioning pituitary neuroendocrine Tumours. Acta Neuropathol Commun. (2022) 10:138. doi: 10.1186/s40478-022-01441-5
69. Li, J, Wen, S, Li, B, Li, N, and Zhan, X. Phosphorylation-mediated molecular pathway changes in human pituitary neuroendocrine tumors identified by quantitative Phosphoproteomics. Cells. (2021) 10:2225. doi: 10.3390/cells10092225
70. Qian, S, Zhan, X, Lu, M, Li, N, Long, Y, Li, X, et al. Quantitative analysis of Ubiquitinated proteins in human pituitary and pituitary adenoma tissues. Front Endocrinol (Lausanne). (2019) 10:328. doi: 10.3389/fendo.2019.00328
71. Carrillo-Najar, C, Rembao-Bojórquez, D, Tena-Suck, ML, Zavala-Vega, S, Gelista-Herrera, N, Ramos-Peek, MA, et al. Comparative proteomic study shows the expression of Hint-1 in pituitary adenomas. Diagnostics. (2021) 11:330. doi: 10.3390/diagnostics11020330
72. Lin, K, Zhang, J, Lin, Y, Pei, Z, and Wang, S. Metabolic characteristics and M2 macrophage infiltrates in invasive nonfunctioning pituitary adenomas. Front Endocrinol (Lausanne). (2022) 13:901884. doi: 10.3389/fendo.2022.901884
73. Evans, CO, Moreno, CS, Zhan, X, McCabe, MT, Vertino, PM, Desiderio, DM, et al. Molecular pathogenesis of human prolactinomas identified by gene expression profiling, Rt-Qpcr, and proteomic analyses. Pituitary. (2008) 11:231–45. doi: 10.1007/s11102-007-0082-2
74. Zhan, X, Wang, X, and Desiderio, DM. Pituitary adenoma Nitroproteomics: current status and perspectives. Oxidative Med Cell Longev. (2013) 2013:580710. doi: 10.1155/2013/580710
75. Zhan, X, and Desiderio, DM. Nitroproteins from a human pituitary adenoma tissue discovered with a Nitrotyrosine affinity column and tandem mass spectrometry. Anal Biochem. (2006) 354:279–89. doi: 10.1016/j.ab.2006.05.024
76. Bhat, FA, Mangalaparthi, KK, Ding, H, Jain, A, Hsu, JS, Peterson, JA, et al. Exploration of Nitrotyrosine-containing proteins and peptides by antibody-based enrichment strategies. Mol Cell Proteomics. (2024) 23:100733. doi: 10.1016/j.mcpro.2024.100733
77. Pînzariu, O, Georgescu, B, and Georgescu, CE. Metabolomics-a promising approach to pituitary adenomas. Front Endocrinol (Lausanne). (2018) 9:814. doi: 10.3389/fendo.2018.00814
78. Calligaris, D, Feldman, DR, Norton, I, Olubiyi, O, Changelian, AN, Machaidze, R, et al. Maldi mass spectrometry imaging analysis of pituitary adenomas for near-real-time tumor delineation. Proc Natl Acad Sci USA. (2015) 112:9978–83. doi: 10.1073/pnas.1423101112
79. Oklu, R, Deipolyi, AR, Wicky, S, Ergul, E, Deik, AA, Chen, JW, et al. Identification of small compound biomarkers of pituitary adenoma: a bilateral inferior petrosal sinus sampling study. J Neurointerv Surg. (2014) 6:541–6. doi: 10.1136/neurintsurg-2013-010821
80. Lee, SH, Nam, SY, and Chung, BC. Altered profile of endogenous steroids in the urine of patients with Prolactinoma. Clin Biochem. (1998) 31:529–35. doi: 10.1016/s0009-9120(98)00063-0
81. Yang, Y, Yao, Y, Deng, K, Xing, B, Lian, W, You, H, et al. Somatic Gnas mutations in acromegaly: prevalence, clinical features and gender differences. Endocr Connect. (2025) 14:266. doi: 10.1530/EC-24-0266
82. Huang, C, Shi, Y, and Zhao, Y. Usp8 mutation in Cushing's disease. Oncotarget. (2015) 6:18240–1. doi: 10.18632/oncotarget.4856
83. Paes, T, Hofland, LJ, Iyer, AM, and Feelders, RA. Epigenetic implications in the pathogenesis of Corticotroph tumors. Pituitary. (2025) 28:51. doi: 10.1007/s11102-025-01522-3
84. Sumal, AKS, Zhang, D, and Heaney, AP. Refractory Corticotroph adenomas. Pituitary. (2023) 26:269–72. doi: 10.1007/s11102-023-01308-5
85. Biagetti, B, and Simo, R. Molecular pathways in Prolactinomas: translational and therapeutic implications. Int J Mol Sci. (2021) 22:22 (20). doi: 10.3390/ijms222011247
86. Paes, T, Buelvas Mebarak, J, Magnotto, JC, Stamatiades, GA, Kuang, Y, Paweletz, CP, et al. Somatic activating Esr1 mutation in an aggressive Prolactinoma. J Clin Endocrinol Metab. (2025) 110:1166–76. doi: 10.1210/clinem/dgae615
87. Ilie, MD, Vasiljevic, A, Louvet, C, Jouanneau, E, and Raverot, G. Gonadotroph tumors show subtype differences that might have implications for therapy. Cancers (Basel). (2020) 12:e1012. doi: 10.3390/cancers12041012
88. Salomon, MP, Wang, X, Marzese, DM, Hsu, SC, Nelson, N, Zhang, X, et al. The epigenomic landscape of pituitary adenomas reveals specific alterations and differentiates among acromegaly, Cushing's disease and endocrine-inactive subtypes. Clin Cancer Res. (2018) 24:4126–36. doi: 10.1158/1078-0432.Ccr-17-2206
89. Neou, M, Villa, C, Armignacco, R, Jouinot, A, Raffin-Sanson, M-L, Septier, A, et al. Pangenomic classification of pituitary neuroendocrine tumors. Cancer Cell. (2020) 37:123–34.e5. doi: 10.1016/j.ccell.2019.11.002
90. Yamato, A, Nagano, H, Gao, Y, Matsuda, T, Hashimoto, N, Nakayama, A, et al. Proteogenomic landscape and clinical characterization of Gh-producing pituitary adenomas/Somatotroph pituitary neuroendocrine tumors. Commun Biol. (2022) 5:1304. doi: 10.1038/s42003-022-04272-1
91. Taniguchi-Ponciano, K, Hinojosa-Alvarez, S, Hernandez-Perez, J, Chavez-Santoscoy, RA, Remba-Shapiro, I, Guinto, G, et al. Longitudinal multiomics analysis of aggressive pituitary neuroendocrine tumors: comparing primary and recurrent tumors from the same patient, reveals genomic stability and heterogeneous transcriptomic profiles with alterations in metabolic pathways. Acta Neuropathol Commun. (2024) 12:142. doi: 10.1186/s40478-024-01796-x
92. Moreno, CS, Evans, CO, Zhan, X, Okor, M, Desiderio, DM, and Oyesiku, NM. Novel molecular signaling and classification of human clinically nonfunctional pituitary adenomas identified by gene expression profiling and proteomic analyses. Cancer Res. (2005) 65:10214–22. doi: 10.1158/0008-5472.Can-05-0884
93. Ruebel, KH, Leontovich, AA, Jin, L, Stilling, GA, Zhang, H, Qian, X, et al. Patterns of gene expression in pituitary carcinomas and adenomas analyzed by high-density oligonucleotide arrays, reverse transcriptase-quantitative Pcr, and protein expression. Endocrine. (2006) 29:435–44. doi: 10.1385/endo:29:3:435
94. Feng, J, Zhang, Q, Zhou, Y, Yu, S, Hong, L, Zhao, S, et al. Integration of proteomics and metabolomics revealed metabolite-protein networks in Acth-secreting pituitary adenoma. Front Endocrinol (Lausanne). (2018) 9:678. doi: 10.3389/fendo.2018.00678
95. Jarmusch, AK, Pirro, V, Baird, Z, Hattab, EM, Cohen-Gadol, AA, and Cooks, RG. Lipid and metabolite profiles of human brain tumors by desorption electrospray ionization-Ms. Proc Natl Acad Sci USA. (2016) 113:1486–91. doi: 10.1073/pnas.1523306113
96. Argelaguet, R, Velten, B, Arnol, D, Dietrich, S, Zenz, T, Marioni, JC, et al. Multi-omics factor analysis-a framework for unsupervised integration of multi-omics data sets. Mol Syst Biol. (2018) 14:e8124. doi: 10.15252/msb.20178124
97. Wang, B, Mezlini, AM, Demir, F, Fiume, M, Tu, Z, Brudno, M, et al. Similarity network fusion for aggregating data types on a genomic scale. Nat Methods. (2014) 11:333–7. doi: 10.1038/nmeth.2810
98. Shen, R, Olshen, AB, and Ladanyi, M. Integrative clustering of multiple genomic data types using a joint latent variable model with application to breast and lung cancer subtype analysis. Bioinformatics. (2009) 25:2906–12. doi: 10.1093/bioinformatics/btp543
99. Arevalo, J, Su, E, Ewald, JD, van Dijk, R, Carpenter, AE, and Singh, S. Evaluating batch correction methods for image-based cell profiling. Nat Commun. (2024) 15:6516. doi: 10.1038/s41467-024-50613-5
100. Korsunsky, I, Millard, N, Fan, J, Slowikowski, K, Zhang, F, Wei, K, et al. Fast, sensitive and accurate integration of single-cell data with harmony. Nat Methods. (2019) 16:1289–96. doi: 10.1038/s41592-019-0619-0
101. Zhao, Y, Wong, L, and Goh, WWB. How to do quantile normalization correctly for gene expression data analyses. Sci Rep. (2020) 10:15534. doi: 10.1038/s41598-020-72664-6
102. Wang, G, Yan, S, Zhang, L, Lin, L, Liu, R, Han, Y, et al. Machine learning-driven Pcdi classifier for invasive Pitnets. Curr Gene Ther. (2025) 25:831. doi: 10.2174/0115665232399193250529074831
Keywords: pituitary, PitNET, multiomics, molecular sequencing, transcriptomics, genomics, epigenomics, proteomics
Citation: Pugazenthi S, Pari SS, Zhang Z, Silverstein J, Kim AH and Patel B (2025) The evolution and application of multi-omic analysis for pituitary neuroendocrine tumors. Front. Med. 12:1629621. doi: 10.3389/fmed.2025.1629621
Edited by:
HaiHui Huang, Shaoguan University, ChinaReviewed by:
Dola Sundeep, Indian Institute of Information Technology Design and Manufacturing, IndiaQiang Zhang, University of Texas MD Anderson Cancer Center, United States
Copyright © 2025 Pugazenthi, Pari, Zhang, Silverstein, Kim and Patel. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bhuvic Patel, Ymh1dmljLnBhdGVsQHd1c3RsLmVkdQ==