 Julian C. Riano-Moreno1,2,3†
Julian C. Riano-Moreno1,2,3† Elizabeth Vargas-Castellanos4*†
Elizabeth Vargas-Castellanos4*† Amy Pedraza5
Amy Pedraza5 Laura Sofía Díaz-Quiñonez5
Laura Sofía Díaz-Quiñonez5 Víctor Saul Rangel-Ramos4*
Víctor Saul Rangel-Ramos4*- 1Medical Subdirection, National Institute of Cancerology, Bogotá, DC, Colombia
- 2Faculty of Medicine, Cooperative University of Colombia, Villavicencio, Colombia
- 3Bioethics Department, Universidad El Bosque, Bogotá, DC, Colombia
- 4Hospital Universitario Mayor-Méderi, Universidad del Rosario, Bogotá, DC, Colombia
- 5Universidad del Rosario, Bogotá, DC, Colombia
Preeclampsia (PE) is a multifactorial and multisystemic syndrome specific to human pregnancy, traditionally characterized by hypertension and proteinuria. Affecting 2%−10% of pregnancies, PE remains a leading cause of maternal and perinatal morbidity and mortality, particularly in low- and middle-income countries. This review traces the historical evolution of PE diagnosis, from early clinical observations to the incorporation of modern omics biomarkers. Early diagnostic criteria were based on observable clinical symptoms, but advancements in biomedical science have highlighted the significance of angiogenic and anti-angiogenic factors, such as sFlt-1 and PlGF, which are used as “Established Angiogenic Biomarkers.” Despite these advancements, clinical consensus on the use of these biomarkers remains elusive due to their variable sensitivity. Recent integrative approaches using omics technologies have provided deeper insights into the complex pathophysiology of PE, uncovering new pathways and “Potential Molecular Biomarkers” for early diagnosis and prediction. However, challenges remain in translating these findings into clinical practice, particularly due to the need for robust validation studies and the consideration of inter-individual and population variability. This review emphasizes the importance of continued research and validation of these biomarkers in diverse cohorts to develop effective predictive tools and improve maternal and fetal outcomes. By exploring the historical and modern perspectives on PE diagnosis, this review aims to provide a comprehensive understanding of the disease and highlight future directions in PE research.
1 Introduction
Preeclampsia (PE) is a multifactorial and complex multiorgan syndrome specific to human pregnancy and the puerperium. Traditionally, it has been characterized by the presence of hypertension with evidence of end-organ involvement, such as proteinuria, thrombocytopenia, renal insufficiency, pulmonary edema, or cerebral symptoms after 20 weeks' gestation (1, 2). Preeclampsia complicates 2%−10% of all pregnancies, and more than 99% of the annual 46,000 preeclampsia-related maternal deaths occur in low-income and middle-income countries (3). Although it is a well-recognized disease of pregnancy, it has historically been poorly defined due to changing diagnostic criteria over the years (4).
The evolving technoscientific and biomedical understanding of its etiology over time has contributed to clarifying the diagnosis and even predicting this condition. However, the etiology remains incompletely understood (4). Several biomedical advances have provided multiple diagnostic and predictive biomarkers for PE, but there is currently no consensus on their clinical use (5, 6).
To improve this understanding, we present a historical perspective on the diagnostic evolution of preeclampsia (PE), tracing the development from clinical and laboratory biomarkers to the latest advancements in omics biomarkers across different Eras: Ancient, Modern, Contemporary, and the Postgenomic era. By examining the progression of diagnostic criteria and the incorporation of potential novel biomarkers, we aim to elucidate how these changes have enhanced our ability to understand the underlying biological mechanisms of PE, while also opening new avenues for predicting and diagnosing the condition. This review highlights significant milestones (Figure 1) and future directions in the prediction and diagnosis of preeclampsia.
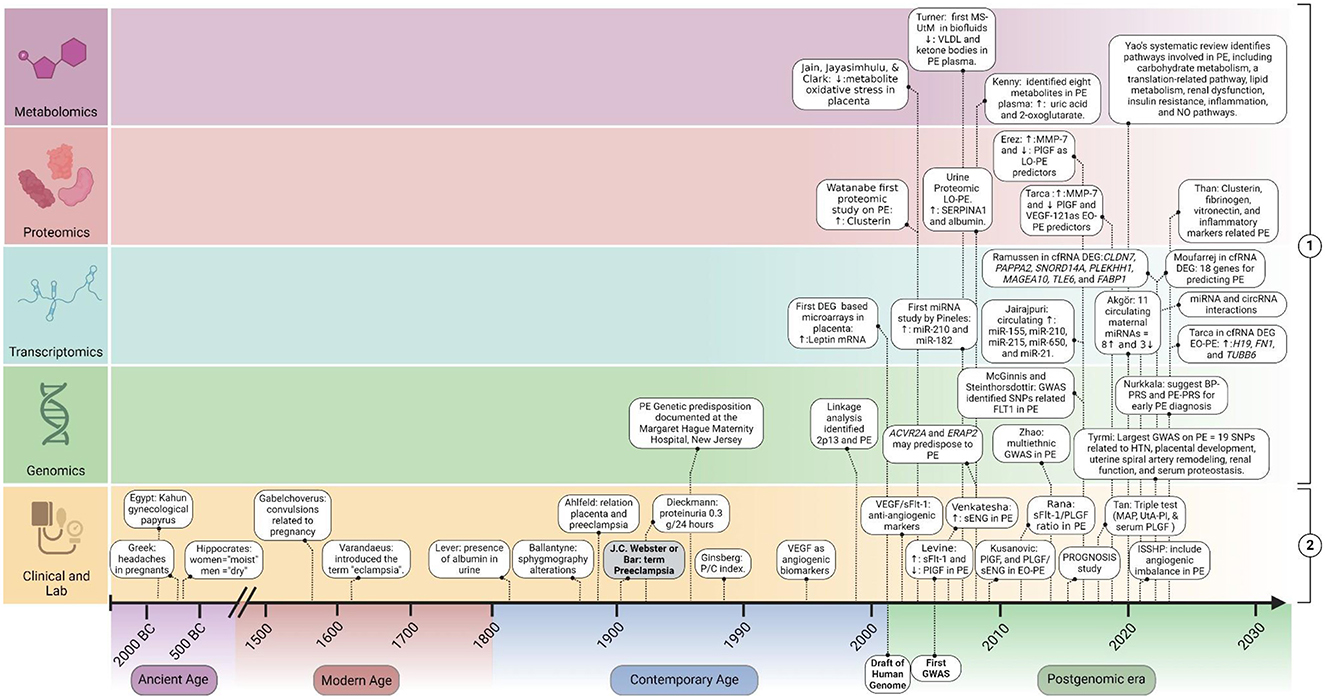
Figure 1. Timeline of milestones in biomarker discovery for prediction and diagnosis of preeclampsia. This figure illustrates the chronological progression of biomarker discovery in the prediction and diagnosis of PE. It includes two main categories: (1) potential Molecular Biomarkers: Omics studies findings to diagnose and predict PE; (2) clinical criteria and “Established Angiogenic Biomarkers” in PE. EO-PE, early-onset PE; LO-PE, late-onset PE; MAP, mean arterial pressure; UtA-PI, uterine artery pulsatility index; DEG, differential expressed genes studies; sENG, soluble endoglin; VEGF, vascular endothelial growth factor; PlGF, placental growth factor; sFlt-1, soluble fms-like tyrosine kinase-1. Created with BioRender.com.
2 Evolution of the diagnosis of hypertensive disorders in pregnancy
Hypertensive disorders of pregnancy (HDP) have been part of medical history since ancient times. The earliest related records date back to Ancient Age in Egypt, specifically the Kahun gynecological papyrus (1850–1700 BC), where treatments were described to prevent the pregnant woman from biting her tongue during childbirth (7). The ancient Greeks also mentioned associated symptoms, such as headaches in pregnant women, which are currently considered a precursor sign of some types of HDP (4, 8).
In the Modern Age, the clinical manifestations of HDP began to be described, starting with convulsions as the first concrete clinical sign associated with these conditions. In 1596, Gabelchoverus included convulsions as a specific group related to pregnancy or originating in the uterus in his work (8). However, it was Johannes Varandaeus who introduced the term “eclampsia” from the Greek “eklampsis,” meaning “a flash,” in his 1620 treatise “Tractatus de affectibus Renum et Vesicae,” which focused on convulsions associated with pregnancy (7, 9). Further, François Mauriceau in 1637 identified convulsions predominantly in primigravida women, proposing primigravidity as a risk factor for eclampsia (4). Additionally, François Boissier de Sauvages, a French physician and botanist, distinguished the convulsive episodes of eclampsia from those of epilepsy (7). This differentiation allowed eclampsia to be recognized in 1793 as a pathology distinct from epileptic processes, definitively integrating it into obstetric pathologies (4).
The laboratory manifestations of HDP were described in the Contemporary Age. In 1843, John C. W. Lever described the presence of albumin in the urine of women with puerperal convulsions, a finding confirmed by Sir James Young Simpson, a pioneer of modern obstetrics. Thus, for the first time, proteinuria was correlated with eclampsia. In 1940, William J. Dieckmann established a proteinuria cutoff of 0.3 g every 24 h for 3 days as abnormal in pregnancy (4). Further, in 1983, Ginsberg et al. de-emphasized proteinuria as the sole marker of eclampsia by introducing the proteinuria/creatininuria (P/C) index, considering 24-h proteinuria as a more suitable index for the diagnosis of eclampsia (10). This, along with studies establishing the relationship between proteinuria and maternal-fetal prognosis, relegated the routine measurement of 24-h proteinuria to a severity criterion (11), while the P/C index is increasingly considered a potential “Gold Standard” for diagnosis (10, 12).
On the other hand, a well-known sign previously considered pathognomonic of eclampsia and proteinuria, edema, described by Demanet in 1797 and supported by the studies of Friedman and Neff in 1977, has been dismissed for its non-specificity (13).
The first established relationship between neurohypertensive or vasospastic symptoms and eclampsia was in 1843 by Robert Johns, who identified these as warning symptoms of eclampsia (8). This relationship was confirmed in 1885 by John William Ballantyne, who observed that pregnant women with edema and albuminuria exhibited alterations in sphygmography reports (7). However, it was not until 1897 that Vasquez and Nobecourt established a clear correlation between eclampsia and gestational hypertension. Since then, arterial hypertension during pregnancy has become a diagnostic criterion for disorders such as PE.
In 1972, the American College of Obstetricians and Gynecologists (ACOG) established and specified week 20 as the cutoff point for the different hypertensive manifestations in pregnancy, primarily considering diastolic blood pressure and defining it as altered when values exceeded 90 mmHg (4, 8). Currently, there is controversy regarding the stages of blood pressure measurement, as the American Heart Association (AHA) has proposed stage cutoffs that focus not only on the second and third trimesters but also on the first trimester and prenatal stages (14, 15).
3 Evolution of the concept and clinical diagnosis of preeclampsia
Although eclampsia has traditionally been seen as the main feature of HDP, it is now recognized that hypertension and proteinuria precede convulsions. This preceding state is considered a different clinical condition within the HDP spectrum, called PE. While there is no consensus on who introduced the term, Leon Chesley attributed its introduction to John Clarence Webster in 1903 in the United States and to Bar in France (7). Since this condition is a precursor to eclampsia, most clinical efforts have focused on the diagnosis and prediction of PE to prevent the devastating impacts of eclampsia.
Currently, PE is recognized as a complex and multisystemic disease. Postmortem findings reveal macroscopic damage in the brain, kidneys, liver, hematopoietic system, and fetoplacental unit, and less frequently in the lungs, heart, pancreas, eyes, intestines, and endocrine and immune systems (7, 16). These damages have been associated with global maternal endothelial dysfunction (17). The state of maternal endothelial dysfunction and subsequent multisystem dysfunction has led to multiorgan damage being included as a diagnostic criterion for PE since 2000 (17, 18).
In addition to hypertension, PE is related to other hemodynamic changes associated with the maternal-fetal interaction, exhibiting diverse hemodynamic phenotypes. For instance, fetal growth restriction (FGR) without PE is more related to high peripheral vascular resistance (19). Conversely, when PE and FGR occur together, a distinct cardiovascular phenotype emerges, characterized by decreased cardiac output and increased vascular resistance. This suggests that fetal weight and gestational age are related to differences in the presentation of maternal endothelial dysfunction, highlighting the various ways in which PE can manifest (19, 20).
These variations in PE manifestation have led to the identification of two distinct forms: early-onset PE (EO-PE) and late-onset PE (LO-PE) (21, 22). This classification considers not only the gestational age at onset but also the underlying mechanisms of each form. EO-PE, which occurs before 34 weeks of gestation, is associated with abnormal placentation. This condition is often linked to severe placental insufficiency resulting from inadequate remodeling of the spiral arteries, leading to reduced placental perfusion. EO-PE has a stronger genetic component and is associated with a higher risk of recurrence within families, indicating a significant hereditary influence (23).
In contrast, LO-PE, presenting after 34 weeks, arises from the interaction between a normally developed placenta and a maternal predisposition to PE. This predisposition is often determined by pre-existing microvascular diseases such as hypertension, diabetes, or other metabolic imbalances. Consequently, LO-PE is less about placental abnormalities and more about the maternal body's inability to cope with the increased physiological demands imposed by the growing fetoplacental unit. This failure of the maternal cardiovascular system to adapt to pregnancy demands highlights the complex interplay between maternal health and fetal development (23).
Therefore, PE is not only a maternal multisystem disease but also a maternal-fetal disease, reflecting the intricate relationship between maternal health and fetal development (18). The historical fluctuation and clinical diversity of HDP have created significant difficulties in establishing specific diagnostic criteria for each of these conditions (24). This has led to discrepancies between scientific societies in defining diagnostic criteria for PE (Table 1). Consequently, it has become necessary to resort to additional strategies beyond traditional clinical and paraclinical evaluations to achieve an accurate diagnosis of PE. Although, the etiology of PE remains poorly understood, efforts to understand the underlying mechanisms have enabled the introduction of new methods to predict and diagnose PE.

Table 1. Diagnostic criteria for hypertensive pregnancy disorders from various gynecology and obstetrics societies.
4 Exploring the etiological hypotheses of preeclampsia
Several hypotheses about the etiology of PE have been proposed over time. In the Ancient times, Hippocrates described women as “moist” and men as “dry”, which he believed predisposed women to fluid absorption, leading to edema during pregnancy. Others considered the uterus to be a wandering organ that, in its displacement, damaged other organs (8). Additionally, an infectious hypothesis was proposed, suggesting that PE was caused by the infection of Bacillus eclampsiae, now known as Proteus vulgaris. François Mauriceau attributed PE to abnormalities in lochial flow or intrauterine fetal death (4, 8).
One of the most recognized hypotheses about the etiology of PE, originating from Modern times, the idea that toxins or poisons entering the maternal bloodstream could produce the symptoms. This hypothesis has given rise to widely used terms in the clinical context, such as “toxemia of pregnancy” or “gravidic toxemia”, which encompass not only PE but also eclampsia, hyperemesis gravidarum, acute yellow atrophy of the liver, gravidic pruritus, and ptyalism (7). However, although our understanding of the disease has advanced significantly from these early hypotheses, the exact cause of PE remains elusive.
In contrast to these earlier hypotheses, the uteroplacental dysfunction hypothesis has garnered the most support in contemporary research. The initial connection between the placenta and PE was proposed by Ahlfeld in 1894 (4). This hypothesis gained further validation in 1914 when James Young identified a relationship between placental infarctions and the development of eclampsia. Subsequent experimental studies demonstrated that guinea pigs administered placental extracts from PE women developed convulsions and other symptoms associated with eclampsia (7). Positioning placental dysfunction as a central factor in the pathogenesis of PE has paved the way for further research and study of PE.
During the second half of the 20th century, significant research efforts were directed toward understanding the relationship between the placenta and PE. One remarkable observation resulting from these studies was that trophoblast cells in PE failed to adequately invade the maternal spiral arteries and convert them from small muscular vessels into large, low-resistance vessels. This finding suggested an “uteroplacental antiangiogenic state” as the main pathophysiological mechanism of the disease (8), which precedes the maternal endothelial dysfunction associated with the multisystemic nature of PE. This concept was termed the “two-stage theory of PE.” Thus, the study and identification of uteroplacental antiangiogenic factors offer a rational starting point for exploring the pathophysiology of PE (25, 26). These insights were crucial for reevaluating diagnostic and predictive methods for PE.
Currently, it is recognized that neither blood pressure nor proteinuria is specific enough to define PE, and relying solely on these methods can lead to overdiagnosis of the disease (27). Therefore, the inclusion of measurements of factors related to the “uteroplacental antiangiogenic state” alongside traditional methods has become a cornerstone for a more accurate and reliable diagnosis of PE (26, 28). As a result, the International Society for the Study of Hypertension in Pregnancy (ISSHP) proposed a new definition of PE in 2021 that includes uteroplacental and target organ dysfunction, as well as the measurement of antiangiogenic biomarkers in maternal circulation as part of the diagnostic criteria for PE (1).
5 Angiogenic and anti-angiogenic factors: key discoveries in preeclampsia
Angiogenesis is regulated by a balance between pro-angiogenic and anti-angiogenic factors. Within the framework of the two-stage model, these alterations emerge initially as placental dysfunction (Stage 1) and later manifest as a systemic maternal response (Stage 2), with angiogenic imbalance as a central hallmark of the disease (Figures 2, 3). Vascular endothelial growth factor (VEGF) and placental growth factor (PlGF), produced by the placenta, are fundamental for vasculogenesis, angiogenesis, and placental development (29). One of the first angiogenic biomarkers studied in PE was VEGF in 1995 (30). VEGF, a specific factor for endothelial cells, induces endothelial proliferation and chemotaxis and stimulates the formation of new blood vessels. Additionally, VEGF increases microvascular permeability and promotes coagulation, distinctive characteristics of PE, making it a good candidate as an angiogenic biomarker in PE (29–31).
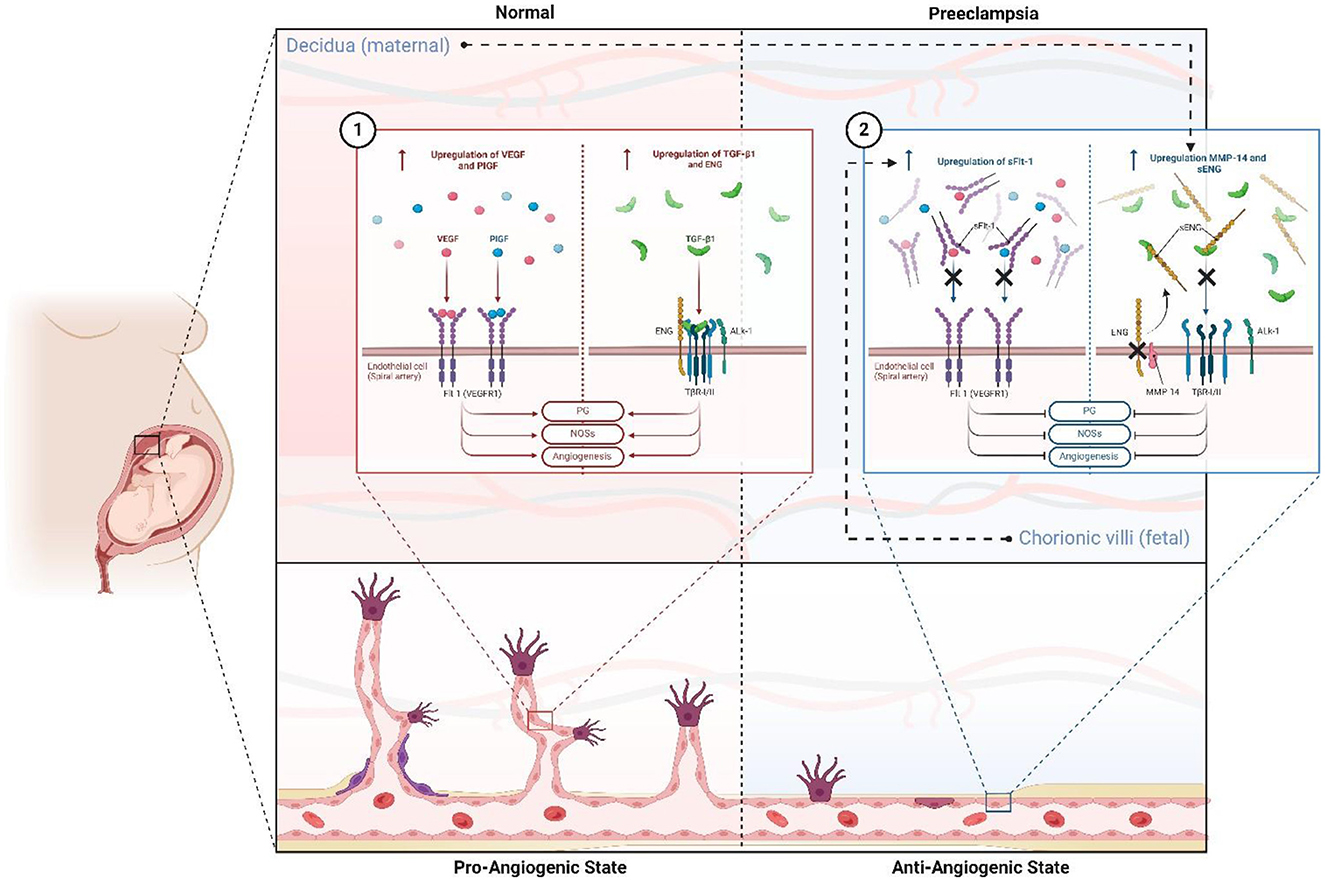
Figure 2. Antiangiogenic and proangiogenic state and factors in normal gestation and preeclampsia. In normal pregnancy (1), there is a pro-angiogenic state mediated by the upregulation and increase of Flt-1 ligands such as PlGF and VEGF, and TNF-β1 pathway activity supported by increased expression of the ENG coreceptor. This favors PG liberation, NOS activity, ON liberation, vasodilation, and angiogenesis. However, in PE, this state is reversed (2), leading to an antiangiogenic state primarily due to fetal-stimulated expression of sFlt-1, which competes for ligands PlGF and VEGF. Additionally, maternal upregulation of MMP-14 increases sENG, which antagonizes the TNF-β1 pathway. ENG, endoglin; sENG, soluble endoglin; Flt-1: fms-related receptor tyrosine kinase 1; VEGF, vascular endothelial growth factor; PlGF, placental growth factor; sFlt-1, soluble fms-like tyrosine kinase-1; NOSs, nitric oxide synthase; ON, nitric oxide. Created with BioRender.com.
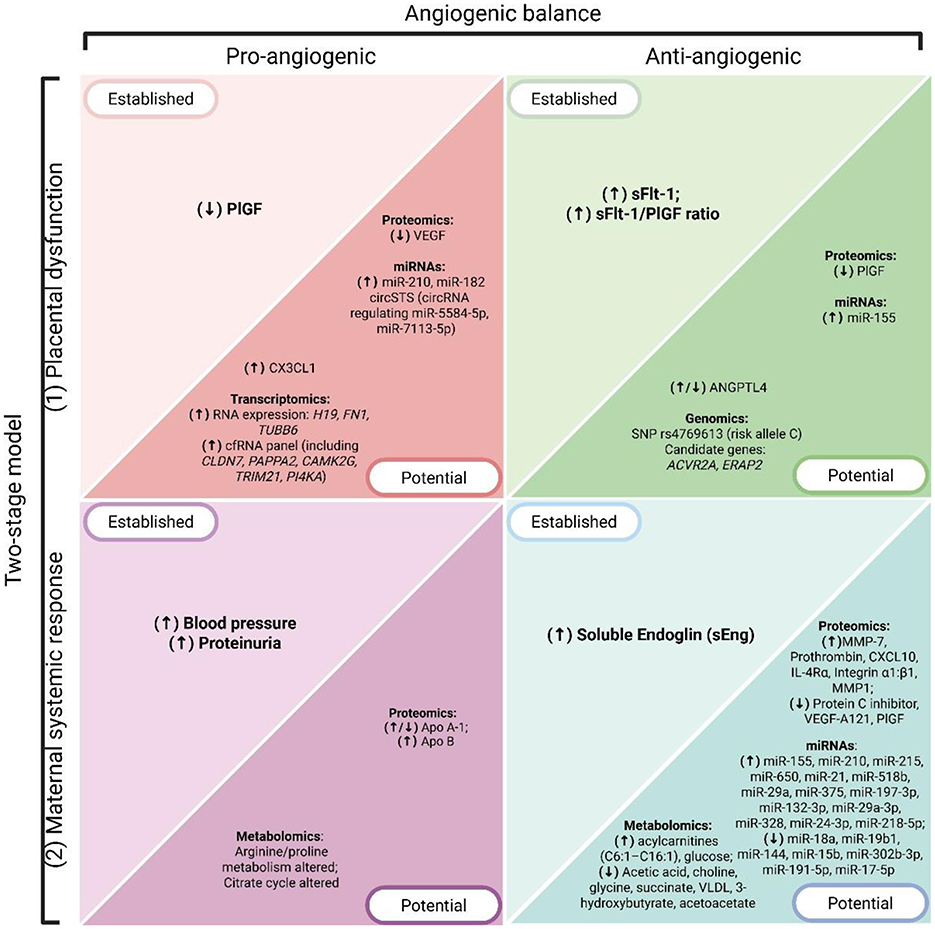
Figure 3. Overview of PE hallmarks and associated biomarkers, organized according to the two-stage model (Stage 1: placental dysfunction; Stage 2: maternal systemic response) and the angiogenic balance (pro-angiogenic vs anti-angiogenic factors). Biomarkers are grouped as established (currently used in clinical practice or guideline-endorsed) and potential (emerging candidates identified through omics technologies). Arrows indicate the direction of dysregulation in PE, where (↑) denotes overexpression or increased levels and (↓) denotes downregulation or decreased levels. Created with BioRender.com.
However, different studies have shown contradictory results regarding VEGF levels in women with PE. While several studies demonstrated that circulating concentrations of VEGF are elevated in these women, recent studies have shown that free VEGF levels in serum decrease in PE patients. These discrepancies are attributed to the limitations of conventional methods, such as sandwich-type Enzyme-Linked Immunosorbent Assay (ELISA), which struggle to measure VEGF protein complexes accurately. Consequently, VEGF has been deemed an unreliable biomarker for PE, prompting researchers to focus on other angiogenic and antiangiogenic factors and to explore new methods for effective measurement (31).
In 2003, two independent groups, Koga et al. (32) and Levine et al. (33), identified the importance of anti-angiogenic factors by evaluating the balance between VEGF and its natural antagonist, sVEGFR-1 (soluble vascular endothelial growth factor receptor-1) or sFlt-1 (soluble fms-like tyrosine kinase-1). They reported an increase in circulating levels of sFlt-1 in patients with PE compared to controls without PE (32). This increase was also associated with a reduction in the expression of PlGF and VEGF (33).
In 2004, in vitro experiments showed that normal villous explants induced endothelial cell migration and tube formation in vitro, both attenuated by preincubation with exogenous sFlt-1. The removal of sFlt-1 by immunoprecipitation significantly restored migration and tube formation to levels comparable to those induced by normal conditioned medium, demonstrating that elevated levels of sFlt-1 in PE are responsible for inhibiting angiogenesis due to the dysregulation of the VEGF/PlGF axis (34).
Between 2004 and 2005, Levine et al. identified that high levels of sFlt-1 and reduced PlGF are detectable even up to 5 weeks before the appearance of the classic clinical manifestations of PE, such as hypertension, proteinuria, and glomerular endotheliosis. This suggests that the relationship between sFlt-1 and PlGF could be used for the prediction or diagnosis of PE (33).
A year later, the relationship between sFlt-1 and PE was further strengthened through studies in murine models, demonstrating an imbalance in the production of anti-angiogenic and pro-angiogenic factors. The exogenous administration of sFlt-1 in mice induced a syndrome similar to human PE, including hypertension, proteinuria, and glomerular endotheliosis, directly linking sFlt-1 to the etiopathogenesis of PE (35). This discovery positioned sFlt-1 as a crucial factor and prompted multiple studies focused on its potential for the prediction and diagnosis of PE.
Deficient spiral artery remodeling, a hallmark of Stage 1 placental dysfunction, causes placental hypoxia and ischemia, increasing anti-angiogenic factors like sFlt-1. Hypoxia-inducible factor-1α (HIF-1α) upregulates sFlt-1, exacerbating endothelial damage, while also inducing angiopoietin-like protein-4 (ANGPTL4). Recently, ANGPTL4 has gained attention as a potential PE biomarker due to its proangiogenic effects, modulation of inflammation, and regulation of metabolism (36–39).
ANGPTL4 was first linked to PE by Liu et al. (40), who reported reduced placental and serum ANGPTL4 in early-onset severe PE, impairing trophoblast invasion and angiogenesis due to decreased PPARγ (Peroxisome Proliferator-Activated Receptor Gamma) activation. Conversely, Açikgözoglu et al. (41) observed elevated maternal serum ANGPTL4 in PE and gestational hypertension (GH), suggesting its utility as an early biomarker since levels increased before proteinuria onset. Also, Shu et al. (42) linked ANGPTL4 downregulation by lncRNA (Long Non-Coding RNA) HIF1A-AS2 (Hypoxia-Inducible Factor 1 Alpha Antisense RNA 2) to oxidative stress, apoptosis, and angiogenic dysfunction in PE (42).
Discrepancies in ANGPTL4 findings complicate its use as a biomarker. Elevated serum ANGPTL4 in early PE may indicate a compensatory response (41), while reduced placental ANGPTL4 in advanced stages may reflect hypoxia and reduced PPARγ activity (40). Similar ANGPTL4 levels in PE and GH further limit its specificity.
Similarly, fractalkine (CX3CL1) has emerged as a marker for early-onset PE and placental dysfunction (43, 44). It exists in membrane-bound and soluble forms, the latter produced by ADAM10 and ADAM17 and detectable in serum (45, 46). Szukiewicz et al. (45), were the first to demonstrate that hypoxia and inflammation stimulate CX3CL1 production and release into circulation. Later, Siwetz et al. (46), linked elevated serum fractalkine in severe early-onset PE to TNF-α (Tumor Necrosis Factor Alpha) activation via the NF-κB (Nuclear Transcription Factor Kappa Beta) pathway, highlighting its role in systemic inflammation. Pala et al. (43), found that while serum fractalkine levels showed no significant differences between controls and intrauterine growth restriction (IUGR) groups, amniotic fluid levels were markedly elevated in IUGR, reflecting localized placental hypoxia Variability in CX3CL1 expression between serum and amniotic fluid, along with small cohorts and inconsistent assays, limits its clinical application, as noted by Szewczyk et al. (47).
While recent biomarkers such as ANGPTL4 and CX3CL1 show potential, their findings remain inconsistent and insufficient for clinical implementation. These observations reinforce the concept that the interplay between pro- and anti-angiogenic factors is not only central to the two-stage model but also provides a framework for interpreting both established and emerging biomarkers (Figure 3). Larger, standardized studies are crucial to address these discrepancies. Meanwhile, well-established biomarkers like sFlt-1 continue to hold greater promise for clinical application due to their more robust evidence base and validated role in PE prediction and diagnosis.
5.1 The “antiangiogenic state” in preeclampsia
sFlt-1 (sVEGFR-1) is a soluble protein derived from an alternative splicing variant of the VEGFR-1 (Flt-1) receptor gene, and its upregulation is one of the best-characterized examples of the anti-angiogenic state that defines Stage 1 placental dysfunction and propagates into Stage 2 maternal systemic response (Figures 2, 3). This transmembrane receptor is crucial for the stabilization of endothelial cells in mature blood vessels, as well as the fenestrated and sinusoidal endothelium of the renal glomerulus, brain, and liver. As a circulating protein containing the same extracellular domain as Flt-1, sFlt-1 competes with Flt-1 for its ligands, particularly PlGF and VEGF, exhibiting strong antagonist activity (Figure 2). Additionally, it can form dominant negative complexes with the mitogenically competent VEGFR-2 (34).
The expression of sFlt-1 is associated with the angiotensin II pathway. The type 1 angiotensin II receptor is the main regulator of vascular tone at the systemic level in adulthood. Its activation triggers signaling pathways that activate the NF-κB, NADPH oxidase, and TNFα, increasing the production of antiangiogenic mediators such as soluble endoglin and sFlt-1. Overactivation of this pathway causes a vasopressor effect and increases peripheral vascular resistance (48).
During the early stages of gestation, there is generally a loss of sensitivity to angiotensin II (48). Levels of sFlt-1 begin to increase in the third trimester and gradually decrease toward the end of pregnancy, which is associated with a reduction in peripheral vascular resistance at the beginning and end of pregnancy. Various proangiogenic factors are expressed in trophoblastic cells and the placenta during gestation, with PlGF being one of the most characterized, predominantly in the syncytiotrophoblast. PlGF, a member of the VEGF family, promotes angiogenesis under conditions of ischemia, inflammation, and wound healing, acting as a VEGF enhancer through its binding to high-affinity Flt-1 receptors on endothelial cell membranes (35, 48). During gestation, the concentration of PlGF increases initially, peaks in mid-pregnancy, and then gradually decreases toward term.
In patients with PE, increased signaling of the type 1 angiotensin II receptor has been evidenced from the beginning of gestation (48), which prematurely and sustainedly elevates the concentrations of sFlt-1 secreted by the placenta throughout pregnancy. This causes an early decrease in circulating levels of PlGF, resulting in an antiangiogenic state that causes abnormal placentation due to insufficient remodeling of maternal spiral arteries and consequent placental ischemia (35). The increase in sFlt-1 in PE also antagonizes VEGF signaling, preventing its interaction with Flt-1. As a result, endothelial dysfunction occurs, decreasing the production of prostacyclins and nitric oxide and releasing procoagulant proteins, aggravating placental and maternal endothelial dysfunction, as well as the clinical signs of PE, such as hypertension and proteinuria (35).
While there is a substantial amount of evidence supporting the relationship between sFlt-1 and PE, the elevation of sFlt-1 is not evident in all cases of PE. Different studies have shown that women with severe LO-PE or term PE may present circulating sFlt-1 values within a normal range, suggesting that other antiangiogenic factors may be involved in the pathogenesis of PE (49).
Endoglin (ENG) is a transmembrane co-receptor of transforming growth factor β (TGF-β) with angiogenic effects, which is involved in various angiogenesis-dependent diseases and associated with endothelial dysfunction, such as PE (50). ENG is predominantly expressed in endothelial cells but also in tissues experiencing active angiogenesis, as well as in non-endothelial histotypes such as vascular smooth muscle cells, fibroblasts, hematopoietic progenitor cells, mesangial cells, and syncytiotrophoblast cells (51).
ENG expression is increased in the placentas of women with PE through mechanisms primarily induced by hypoxia, oxidative stress, and oxysterol-mediated liver X receptor (LXR) activation (50). Overexpression of endoglin results in an increase in its soluble form in the maternal circulation. This soluble isoform, known as soluble endoglin (sENG), has an antiangiogenic effect, unlike the transmembrane form, as described and associated with PE in 2006 by Venkatesha and colleagues (52).
sENG represents the extracellular domain of membrane endoglin, cleaved by the action of metalloproteinases, predominantly matrix metalloproteinase-14 (MMP-14) (50, 53). The antiangiogenic and prohypertensive activity of sENG is due to its ability to bind circulating TGF-β molecules (Figure 2). This way, it interferes with TGF-β1 and ALK1 (activin receptor-like kinase 1) receptor signaling pathways, inhibiting endothelial nitric oxide synthase activation, which disorganizes angiogenesis and promotes vasoconstriction (50). The concentration of sENG in maternal serum begins to increase between 8 and 10 weeks before the appearance of clinical manifestations of PE (54). It has even been observed that sENG concentrations can increase in the second trimester of pregnancy before clinical symptoms of PE (55). Due to these properties, sENG actively contributes to the poor placentation observed in PE, as well as to the pathogenesis and manifestation of its clinical signs and symptoms, especially hypertension and proteinuria.
sENG and sFlt-1 may exhibit synergistic effects in the development of PE (Figure 2). Each operates through separate mechanisms, but as demonstrated in studies with pregnant murine models, the endothelial dysfunction and severe PE phenotype caused by sENG are amplified by the co-administration of sFlt-1 (52, 56).
Thus, it is currently recognized that the combination of proangiogenic and antiangiogenic factors, particularly the balance among PlGF, sFLT1, and endoglin, characterizes the development of PE. Unfortunately, no single test has yet met the required specificity and sensitivity to allow its routine use as a diagnostic or predictive tool in clinical practice (57). Therefore, tests that evaluate the combined measure of these factors have shown promising sensitivity and specificity for the early prediction of PE (55, 58).
6 Established and potential molecular biomarkers in preeclampsia
The multifactorial nature of PE presents a significant challenge to biomarker discovery. Traditional biomarkers have been derived from known pathophysiological abnormalities, particularly those involving angiogenic imbalance as a central hallmark of the disease, consistent with the two-stage model (Figures 2, 3).
The NIH defines a biomarker as “a characteristic that is objectively measured and evaluated as an indicator of normal biological processes, pathogenic processes, or pharmacological responses to a therapeutic intervention” (59). An ideal biomarker possesses high specificity and sensitivity, is detectable through minimally invasive sampling procedures, and its concentration should indicate a disease state (60–62). Biomarkers can be categorized according to their clinical applications, with the most recognized being diagnostic, predictive, and prognostic biomarkers (26, 63, 64). Additionally, susceptibility/risk, monitoring, pharmacodynamic, and safety biomarkers have been described (64).
The multifactorial nature of PE presents a significant challenge to biomarker discovery. Traditional biomarkers have been derived from known pathophysiological abnormalities associated with PE through a hypothesis-driven approach (57). Despite this, the discovery of new potential biomarkers remains crucial for a deeper understanding of the mechanisms underlying the pathophysiology of PE. Identifying new biomarkers not only offers insights into the disease mechanisms but also provides potential diagnostic tests that can enable effective therapy. This is particularly relevant for PE, where early detection and intervention can significantly impact maternal and fetal outcomes.
In the second part of this review, we will examine, following a historical approach, the current advances in the search for diagnostic and predictive biomarkers based on uteroplacental dysfunction, referred to here as “Established Angiogenic Biomarkers” that emerged in the Contemporary Age of Medicine. Additionally, we will review potential new biomarkers for the prediction and diagnosis of PE using omics technologies, designated as “Potential Molecular Biomarkers” that emerged in the Postgenomic Era. Figure 1 depicts the milestones, main studies, and findings in these two scenarios.
6.1 Techniques for established angiogenic biomarker detection
6.1.1 Conventional methods
The serum measurement of sFLT1, sVEGF, PlGF, and sENG, molecules related to the pathogenesis of PE, was initially performed using quantitative sandwich-type ELISA starting in 2003 (33, 35, 65). However, this method is inefficient for evaluating protein complexes and thus ineffective for assessing the total (free and bound) circulating levels of potential markers (31). Consequently, ELISA kits are restricted to research use only, and their cutoff values are not recommended for clinical use (35). Despite this limitation, many research groups continue to use ELISA measurements due to their cost-effectiveness. It is worth noting that ELISA measurements are recommended only for research purposes and for evaluating single biomarkers.
To address this limitation, clinical-use kits based on immunochemiluminescent assays, like the Elecsys types, have replaced ELISAs as the standard for measuring plasma levels of PE biomarkers. These assays show high correlation with ELISA determinations. Additionally, being automated, they can deliver results in ~18 min, significantly reducing diagnosis time (66). For clinical diagnosis and combined biomarker measurements, such as the sFlt-1/PlGF ratio, it is recommended to use Elecsys sFlt-1 and Elecsys PlGF assays, as their cutoff points are better standardized (66).
6.1.2 Emerging and point-of-care platforms
Beyond conventional ELISA and automated electrochemiluminescent immunoassays, several complementary and emerging techniques have been explored to broaden the methodological landscape. Multiplex immunoassays, such as bead-based Luminex platforms, have enabled the simultaneous measurement of panels including sFlt-1, PlGF, and soluble endoglin from minimal sample volumes, offering efficiency and the possibility of multimarker profiling (67, 68). Similarly, mass spectrometry–based proteomics, initially developed for discovery research, has been adapted for targeted validation of angiogenic proteins with high analytical specificity, thereby bridging experimental findings with potential clinical translation (69).
At the clinical interface, point-of-care strategies have begun to emerge, particularly lateral flow and fluorescence immunoassays for PlGF and sFlt-1/PlGF ratios (70). These rapid, portable assays offer immediate risk stratification and have already been piloted within national care pathways for suspected preterm preeclampsia, supporting timely triage in busy obstetric settings. In parallel, microfluidic “lab-on-a-chip” platforms and biosensors using electrochemical or nanomaterial-based detection are under development, showing clinically relevant sensitivity ranges for angiogenic biomarkers in proof-of-concept studies (71).
Despite their promise, most of these techniques remain costly, technically demanding, or still in early phases of validation, which limits their uptake compared with conventional ELISA and automated immunoassays. These barriers are particularly pronounced in low- and middle-income countries, where limited infrastructure and resource constraints hinder deployment and risk widening inequities in access to timely diagnosis and care (72, 73). The “last mile” conditions in rural clinics often degrade performance unless devices are ruggedized and simplified (74); indeed, many promising point-of-care innovations stall precisely because they are not engineered or validated under the realities of LMIC health systems (72). This underscores the need for harmonization, external validation in large and diverse cohorts, and pragmatic assessments of cost-effectiveness before these approaches can complement or replace conventional methods in routine preeclampsia diagnosis.
6.2 Predictive and diagnostic applications of pro and anti-angiogenic biomarkers in preeclampsia
One of the first studies to evaluate multiple serum biomarkers of angiogenesis was conducted by Kusanovic et al. (162). Within the framework of the two-stage model, these studies provided evidence that alterations in the angiogenic balance are detectable before the maternal systemic response becomes clinically apparent (Figures 2, 3). This cohort study included 1622 patients with singleton pregnancies, using the ELISA method to measure sFlt-1, PlGF, and sENG in maternal plasma to evaluate the independent and combined values of these biomarkers in early and late pregnancy stages. They observed that women with EO-PE had lower PlGF concentrations in the first trimester compared to women without PE. In the same study, it was identified that a lower PlGF/sENG ratio (lower circulating PlGF or higher sENG) is associated with PE, and this ratio has better predictive value, with a sensitivity of 100% and specificity of 98% for EO-PE prediction (37). However, other authors have shown that PlGF levels alone are sufficient as a predictor of EO-PE (75).
From these observations, the Fetal Medicine Foundation (FMF) developed a first-trimester screening model, endorsed by the International Federation of Gynecology and Obstetrics (FIGO), which estimates the individual risk of developing PE. The model combines maternal clinical factors, mean arterial pressure (MAP), uterine artery pulsatility index (UtA-PI), and serum measurement of PlGF through ELISA, also known as the triple test. This model provides a risk score ranging from 1 to 100, indicating lower to higher risk, with detection rates of 90%, 75%, and 41% for extremely preterm PE (before 32 weeks), preterm, and term PE, respectively (76). This model captures placental dysfunction in Stage 1 through the measurement of angiogenic factors, even before the onset of maternal systemic manifestations in Stage 2.
Subsequently, the measurement of circulating sFlt-1 was included in this model. This modified model was applied to 57 (0.6%) cases of EO-PE and 246 (2.6%) cases of LO-PE, finding a detection rate of 91.2% and 76.4%, respectively. Preventive interventions with acetylsalicylic acid reduce the risk of early PE by 62% using this model (77).
Other proposed combined markers, such as the sFlt-1/PlGF ratio, have been applied for diagnostic purposes. The sFlt-1/PlGF ratio has shown superior diagnostic power for both EO-PE and LO-PE compared to individual measurements of sFlt-1 or PlGF. For EO-PE, the sensitivity and specificity were even higher, reaching 100% and 95%, respectively (66). Other studies have established that an sFlt-1/PlGF ratio of 85 allows detection of PE before 34 0/7 weeks of gestation, with a sensitivity of 89% and specificity of 97%. It has also been identified that a value of 33 above 20 weeks and 110 at 34 weeks or more allows detection of PE with similar sensitivity of 88% and specificity of 99.5% (78).
The PROGNOSIS study, a prospective multicenter study that included 1,273 patients between 24 0/7 and 36 6/7 weeks of gestation, demonstrated that an sFlt-1/PLGF ratio of 38 or higher is indicative of the development of PE in the next 4 weeks, with a positive predictive value of 36.7%. Likewise, if the ratio is lower than this value, it allows the exclusion of PE within a week (79). These findings exemplify how quantifying the imbalance between pro-angiogenic and anti-angiogenic factors can guide clinical decision-making and risk stratification in PE (Figure 3). All this has led to the use of these biomarkers being part of routine obstetric clinical practice in some European countries (76).
The use of these combined biomarkers has been explored not only for the prediction and diagnosis of the disease but also for predicting adverse maternal and fetal outcomes, as well as determining the appropriate timing for ending the pregnancy in cases of PE. Rana et al. (80) was the first to evaluate the efficacy of the sFlt-1/PlGF ratio focused on predicting complications associated with PE. In 616 women with a high clinical suspicion of PE, they observed an inversely proportional relationship between the severity of the imbalance between pro-angiogenic and antiangiogenic factors through the sFlt-1/PlGF ratio and the remaining duration of the pregnancy (80).
The recognition of the role of angiogenic factors in the pathogenesis of PE and their value as biomarkers has led to the development of precision medicine strategies based on these biomarkers (Figure 3). Thadhani et al. (81) proposed different mechanisms to antagonize the effects of sFlt-1, such as saturating the system with its ligands VEGF and PlGF, administering anti-sFlt-1 antibodies, or using dextran sulfate cellulose columns for apheresis and extracorporeal removal of sFlt-1. In two studies that included patients with preterm PE, this latter procedure produced a reduction in sFlt-1 levels, improvement in proteinuria and blood pressure, without adverse effects for the mother and fetus (81, 82).
It is worth noting that the introduction of angiogenic biomarkers has mainly been valuable for the diagnosis and prediction of EO-PE, while evidence remains limited for LO-PE (83). Nonetheless, as both subtypes share pathogenic mechanisms linked to angiogenic imbalance within the two-stage framework, these targeted therapies may ultimately be relevant to both forms of the disease (Figures 2, 3).
6.3 Potential molecular biomarkers in preeclampsia
In the postgenomic era, high-throughput sequencing technologies enabled the detection of various molecular biomarkers for complex diseases like PE (Figure 3). When framed within the two-stage theory—placental dysfunction followed by maternal systemic response—these approaches map how molecular alterations across DNA, RNA, proteins, and metabolites converge on the angiogenic balance that underlies disease progression. These technologies have uncovered molecular signatures in DNA and expression biosignatures in various RNA species, which can serve as diagnostic or predictive biomarkers of PE and reveal underlying molecular mechanisms beyond antiangiogenic factors. Other omics technologies such as proteomics and metabolomics, involving the large-scale study of proteins and metabolites within a biological system, have identified proteins and metabolites that provide insight into PE pathophysiology and can potentially be used as predictive and diagnostic biomarkers of PE. Taken together, these omics approaches highlight a continuum—from genomic predisposition to transcriptional, proteomic, and metabolic dysregulation—that converges on angiogenic and anti-angiogenic imbalance. To maintain coherence with the two-stage model, the following subsections emphasize how omics data inform either Stage 1 placental dysfunction or Stage 2 maternal systemic response (Figure 3).
6.3.1 Genetic and genomic insights into preeclampsia
6.3.1.1 Stage 1: placental dysfunction
PE, as a multifactorial condition, presents severity determined by complex interactions between maternal and fetal genotypes as well as environmental factors (84). The genetic predisposition to PE was first documented in the 1960s when researchers at the Margaret Hague Maternity Hospital in New Jersey observed a higher incidence of PE and eclampsia among sisters, daughters, and granddaughters of affected women compared to daughters-in-law. Subsequently, through the Swedish Birth Register, heritability was estimated at ~55%. Additionally, it was identified that this is also dependent on fetal genetics, establishing that the contributions to PE heritability are of both maternal and fetal origin, with 35% and 20%, respectively (84). These findings have been recently supported by an approach based on Genome-based Restricted Maximum Likelihood (GREML), using chip genotypes from European and Central Asian subjects to estimate PE heritability from single nucleotide polymorphism (SNP). Results estimated that the heritability for maternal PE in Europe is 38.1% (95% CI: 29.3–46.8) and for fetal PE is 21.3% (95% CI: 7.4–35.3). In Central Asia, the heritability found was 54.4% (95% CI: 29.6–79.3) for maternal PE and 42.5% (95% CI: 17.3–67.7) for fetal PE (85), suggesting variability in the genetic component among different populations.
EO-PE, linked to severe placental insufficiency, is suggested to have a stronger genetic component. This form of PE is associated with a higher risk of recurrence within families, indicating significant heritability. In contrast, LO-PE is more related to the maternal body's inability to cope with the increased physiological demands imposed by the growing fetoplacental unit (23). These particularities in the heritability of PE have driven the search for the associated genetic background.
Early studies relied on candidate gene association models, which aim to identify genes by comparing allele frequencies and linkage between cases and controls. From this approach, at least eight different chromosomal regions associated with genes like ACVR2A and ERAP2 that may predispose to PE were identified (84). With the advent of next-generation sequencing (NGS) technologies, genome-wide association studies (GWAS) have overcome some of the inherent limitations of candidate gene studies (86). GWAS are based on the idea that many common diseases are complex and polygenic, with multiple variants, each with minimal but additive effects that can contribute to disease risk (87). These studies use SNPs as units of genetic variation in the genome (88, 89).
One of the pioneering studies and the largest GWAS on PE, which included 4,380 cases and 310,238 controls, was conducted by McGinnis and Steinthorsdottir and collaborators (90). They identified SNPs in linkage within the FLT1 gene on chromosome 13 (SNP rs4769613, risk allele C, p = 3.2 × 10−8). The C allele, present in 53% of the studied population, was estimated to confer a risk of PE (OR = 1.22, 95% CI (1.14–1.31) per allele), supporting the relationship of FLT1 with the development of PE. Additionally, two other SNPs of independent association with PE were identified: rs12050029 (A>G) (14% frequency) and rs149427560 (C>T) (6% frequency). SNPs rs4769613 (C>T) and rs12050029 (A>G) were located in placental enhancer regions near FLT1, suggesting that these variants may influence FLT1 expression by affecting gene transcription (90). This suggests that multiple causal variants in the fetal FLT1 locus contribute to disease risk. Interestingly, there was no difference in PE risk based on parental allele segregation, indicating that the increased risk is mediated by fetal gene expression, possibly in the placenta (90).
SNP rs4769613 C>T has been widely studied due to its high association with PE (84, 91). To understand whether there were effects between this SNP and PE subtype, its association in EO-PE and LO-PE, as well as with pregnancies with small-for-gestational-age (SGA) vs. non-SGA babies, was explored. From this approach, it was identified that SNP rs4769613[C] is more common in LO-PE pregnancies with non-SGA babies (90). These differential effects suggest that genetic variation in the FLT1 locus—specifically in a placental enhancer region near the gene— not only increases PE risk but also relates to the phenotypic heterogeneity of the disease. However, in other studies evaluating the relationship between fetal genotype rs4769613 (C>T) and FLT1 and sFlt1 levels in placentas of women with PE vs. control placentas, no association of genotype with protein level has been observed (91, 92).
6.3.1.2 Stage 2: maternal systemic response
In addition to placental pathways, maternal genetic factors play an important role in shaping the systemic response that defines the clinical phase of PE. Early candidate gene studies focused on variants within the renin–angiotensin system (RAS), coagulation factors, oxidative stress pathways, dyslipidemia, and immunoregulatory components, particularly within the HLA region (84). However, these approaches are limited as they start from a priori hypotheses of pathogenicity. They have often lacked replication due to issues such as inadequate sample sizes, inaccurate clinical phenotyping, lack of correction for multiple testing, poorly matched control groups, hidden ethnic bias, positive publication bias, and random error (84).
Zhao et al. (93) conducted a pioneering GWAS in the maternal genome, including 1,070 Afro-Caribbean mothers, 723 Hispanic mothers, and 1,257 European mothers from the HAPO study. Significant candidate SNPs were identified in different ethnic groups (Table 2). However, no individual SNP was significantly associated with PE after Bonferroni correction, indicating that findings should be interpreted with caution (94). Other studies, including SNP-based gene set enrichment analyses, have enabled the detection of some molecular pathways related to PE risk. A risk SNP, rs7579169 (T>C), is in the intergenic region near the inhibin beta B (INHBB) gene. Other SNPs associated with PE, such as rs12711941 (T>G), rs7576192 (A>G), and rs2681472 (A>G), were also identified near the inhibin gene (95). Inhibin is a glycoprotein that reduces levels of follicle-stimulating hormone (FSH). In pregnancies with PE, inhibin protein levels are much higher compared to normal pregnancies, although this relationship is not yet fully understood (94, 95).
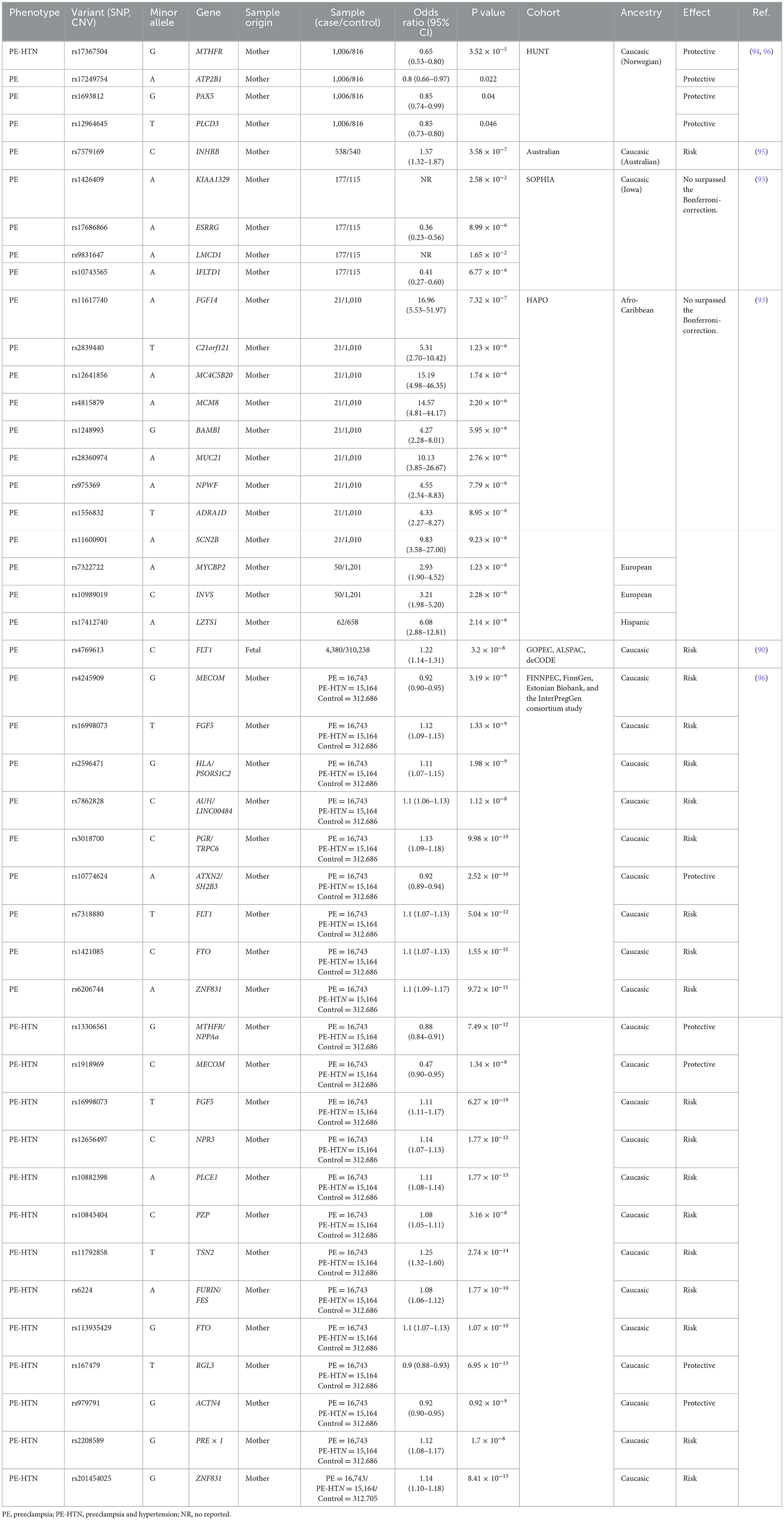
Table 2. GWAS-identified genes associated with preeclampsia.
SNPs associated with protective effects against PE have also been identified, particularly in genes related to the calcium signaling pathway. This pathway includes genes such as ATP2B, ADRA1D, and PLCD3. Notably, SNP rs17367504 (G>A) has been found in the Norwegian population, and SNP rs13306561 (T>C) has been identified in other Caucasian cohorts (Table 2). These SNPs in the MTHFR gene have shown associations with protective effects against PE (94, 96).
Recently, Tyrmi and colleagues conducted one of the largest GWAS on PE, combining data from FINNPEC, the FinnGen project, the Estonian Biobank, and the previously published InterPregGen consortium GWAS. This study included a total of 16,743 women with a history of PE and 15,200 with PE or other maternal hypertension during pregnancy. From this analysis, 19 SNPs associated with PE were found, 13 of which were new. Seven of the novel loci harbor genes previously associated with hypertension traits such as NPPA, NPR3, PLCE1, TNS2, FURIN, RGL3, and PREX1. Additionally, risk loci were identified near genes involved in placental development (PGR, TRPC6, ACTN4, PZP), uterine spiral artery remodeling (NPPA, NPPB, NPR3, ACTN4), renal function (PLCE1, TNS2, ACTN4, TRPC6), and serum proteostasis maintenance during pregnancy (PZP) (96). This study revealed that genes related to hypertension traits are associated with PE, but many of these genes have additional pleiotropic effects on cardiometabolic, endothelial, and placental function. Moreover, several of the associated loci have no known connection to cardiovascular diseases but harbor genes that contribute to the maintenance of a successful pregnancy, whose dysfunctions can lead to PE-like symptoms.
6.3.2 Predictive utility of polygenic risk scores in preeclampsia
6.3.2.1 Stage 1: placental dysfunction
The diversity of loci associated with PE identified by GWAS allows for the consideration of incorporating polygenic risk scores (PRS) as a potential application of these findings in clinical practice. A PRS is typically derived from GWAS data and calculated as the weighted sum of the effect sizes estimated per SNP. PRS is an estimate of genetic susceptibility to the condition of interest, calculated from the effect size of the risk variants of a trait (97). These approaches have been applied in PE prediction and have shown significant potential for improving the identification of women at risk.
An initial study of PRS application in PE/eclampsia was based on a multi-ethnic meta-analysis that included 20,064 cases with PE and 703,117 controls, and gestational hypertension in 11,027 cases and 412,788 controls. This study identified 18 independent loci associated with these conditions, 12 of which are new. These loci highlight the role of natriuretic peptide signaling (NPPA, NPR3, FURIN), angiogenesis (FLT1), renal glomerular function (PLCE1, TNS2, TRPC6), trophoblast development (WNT3A), and immune dysfunction (MICA, SH2B3) (98). These findings provide new insights into the underlying mechanisms of PE beyond angiogenesis, weaving relationships with HTN mechanisms.
6.3.2.2 Stage 2: maternal systemic response
PRS derived from these loci effectively predicted PE and gestational hypertension in external cohorts, independently of other risk factors. A strong genetic correlation (rg) was observed between PE/eclampsia and gestational hypertension (rg = 0.71), and between systolic blood pressure (SBP) and gestational hypertension (rg = 0.73) (83). These results suggest that PRS provides an additional tool for PE risk stratification, improving the accuracy of risk prediction algorithms.
Under this approach, Nurkkala et al. (99) in a recent study, demonstrated the relevance of PRS in predicting PE and HDP. This study included 141,298 participants from the FinnGen study, followed from 1969 to 2021, to evaluate the association between Blood Pressure Polygenic Risk Scores (BP-PRS) and PE-specific PRS (PE-PRS) with gestational hypertension and PE. The genetic loci identified in this research include genes related to natriuretic peptide signaling, angiogenesis, renal glomerular function, trophoblast development, and immune regulation.
Additionally, results showed that BP-PRS is strongly associated with gestational hypertension (HR 1.38; 95% CI 1.35–1.41) and PE (HR 1.26; 95% CI 1.23–1.29), capturing the genetic architecture of PE better than PE-PRS alone (99). This way, it is suggested that the inclusion of BP-PRS not only improves the prediction of PE and gestational hypertension but also highlights the strong genetic correlation between hypertension predisposition and the occurrence of hypertensive disorders of pregnancy. This offers new perspectives for early diagnosis and efficient management of these conditions through interventions such as low-dose aspirin administration.
6.3.3 Transcriptomics and mRNA as potential biomarkers in preeclampsia
6.3.3.1 Stage 1: placental dysfunction
Transcriptomics applied to complex diseases such as PE has focused on measuring differential gene expression (DEG). This approach has identified gene sets and candidate pathways, as well as constructed gene networks and disease-specific pathways. Various transcriptomics studies in PE have concentrated on DEG in placental tissue using expression microarray techniques and RNA-seq (100). Except for leptin (LEP) and Flt1, most studies have reported inconsistent lists of differentially expressed genes in PE (101). This inconsistency is possibly due to small sample sizes, variability in matching, different maternal ethnicities, different types of chips and microarray platforms used, and indirect comparison designs (i.e., pooling mRNA from multiple individuals into a single sample) (100, 102). Moreover, many DEG-based studies often require invasive procedures, complicating their clinical application.
In a recent study, Tarca et al. (103) characterized cfRNA transcriptomic changes in maternal whole blood associated with early PE before and at the time of disease diagnosis, revealing a significant increase in H19, FN1, and TUBB6 gene expression in women with early PE at 11–17 weeks of gestation. Notably, both FN1 and H19 showed higher expression in placental tissue from women with PE. H19 was found to reduce cell viability while promoting invasion and autophagy in trophoblastic cells, along with the activation of PI3K/AKT/mTOR pathways. These findings provide direct evidence of transcriptomic alterations that contribute to impaired placentation and abnormal trophoblast function in the first stage of PE.
6.3.4 Stage 2: maternal systemic response
Advances in high-throughput sequencing have overcome some of the limitations of placental DEG studies by enabling the study of cell-free RNA (cfRNA), a fraction of RNA released into the blood from various tissues, through minimally invasive procedures. This has facilitated the application of transcriptomics in the clinical context, enabling the proposal of RNA-based biomarkers for complex diseases such as PE.
Rasmussen et al. (104) analyzed 2,539 plasma samples from 1,840 pregnant women from various cohorts with racial and ethnic diversity. They used cfRNA to track pregnancy progression and predict PE months before clinical presentation. Transcriptomic data were collected and analyzed using a machine learning model based on cfRNA profiles. Samples were collected on average 14.5 weeks before delivery, identifying seven key genes (CLDN7, PAPPA2, SNORD14A, PLEKHH1, MAGEA10, TLE6, FABP1) predictive of PE, with a sensitivity of 75% and a positive predictive value of 32.3%. This model identifies women at risk of developing PE from 16 to 27 weeks of gestation, providing a significant window for early interventions and improving obstetric management. These genes are associated with relevant biological processes such as placental angiogenesis, arterial morphogenesis, and embryonic placental development.
The applicability of cfRNA for predicting and diagnosing PE was supported by the work of Moufarrej et al. (105), who used cfRNA measurement in plasma in 404 blood samples from 199 pregnant women at different gestational stages. They identified transcriptomic changes associated with PE, dividing participants into discovery and validation cohorts. Samples collected 12 weeks before, between 13 and 20 weeks, from 23 weeks of gestation, and postpartum were analyzed. From these comparisons, it was demonstrated that changes in cfRNA expression between normotensive and preeclamptic mothers are marked and stable from early pregnancy stages. These changes are enriched in tissue-specific and neuromuscular, endothelial, and immune cell type-specific genes, reflecting key aspects of PE physiology. The identified set of 18 genes includes CAMK2G, DERA, FAM46A, KIAA1109, LRRC58, MYLIP, NDUFV3, NMRK1, PI4KA, PRTFDC1, PYGO2, RNF149, TFIP11, TRIM21, USB1, YWHAQP5, and Y_RNA. These genes are involved in molecular pathways related to muscle contraction, immune signaling, and endothelial function, highlighting the multifactorial nature of PE.
Based on these findings, the authors developed an 18-gene panel, which could predict PE with a sensitivity of 85% to 100% and a specificity of 85% to 97%. From these results, they suggest creating a liquid biopsy test based on evaluating the expression profiles of these 18 genes, which could predict the risk of PE between 5 and 16 weeks of gestation, well before clinical symptoms appear. Implementing such tests could facilitate the adoption of prophylactic interventions, such as low-dose aspirin use (105).
The integration of transcriptomic biomarkers, particularly cfRNA signatures, provides a bridge between placental dysfunction and the maternal systemic response. While DEG analyses of placental tissue highlight impaired trophoblast and angiogenic pathways consistent with stage 1 of PE, cfRNA studies in maternal circulation reveal systemic endothelial, immune, and neuromuscular alterations characteristic of stage 2. Together, these findings underscore the potential of transcriptomic approaches to capture the continuum of PE pathogenesis and support the development of minimally invasive tools for early prediction and diagnosis. These advancements in molecular biology could revolutionize the management of PE, enable early interventions and significantly improve maternal and neonatal outcomes, although rigorous research and validation studies remain essential to establish their clinical applicability and reliability.
6.3.5 MicroRNAs and RNA species: emerging biomarkers for preeclampsia
6.3.5.1 Stage 1: placental dysfunction
Transcriptomic studies have also enabled the identification of a wide variety of RNA species, such as microRNAs (miRNAs), long non-coding RNAs (lncRNAs), and circular RNAs (circRNAs). miRNAs are related to regulatory processes of the genome and are considered potential biomarkers for complex diseases like PE (Table 3). miRNAs are small RNA species, 22–24 nucleotides long, non-coding, endogenous, and highly conserved. They represent only 2%−3% of the genome but can regulate the expression of ~60% of genes (106). A single miRNA can regulate around 200 different transcripts, affecting various cellular pathways; similarly, the same mRNA can be regulated by multiple miRNAs (107). Different studies report that, during normal pregnancy, the miRNA expression profile changes according to the gestational stage (108, 109). Additionally, ~600 different types of miRNAs have been identified that are synthesized in the placenta (110). Different placental miRNAs have even been described as identified in maternal circulation. Due to their greater stability compared to mRNA, they have been considered possible candidates for biomarkers for pregnancy monitoring (111). The altered expression of some miRNAs in placental tissue and maternal circulation has been associated with various types of pregnancy complications, such as PE, preterm labor, and spontaneous abortion (112).
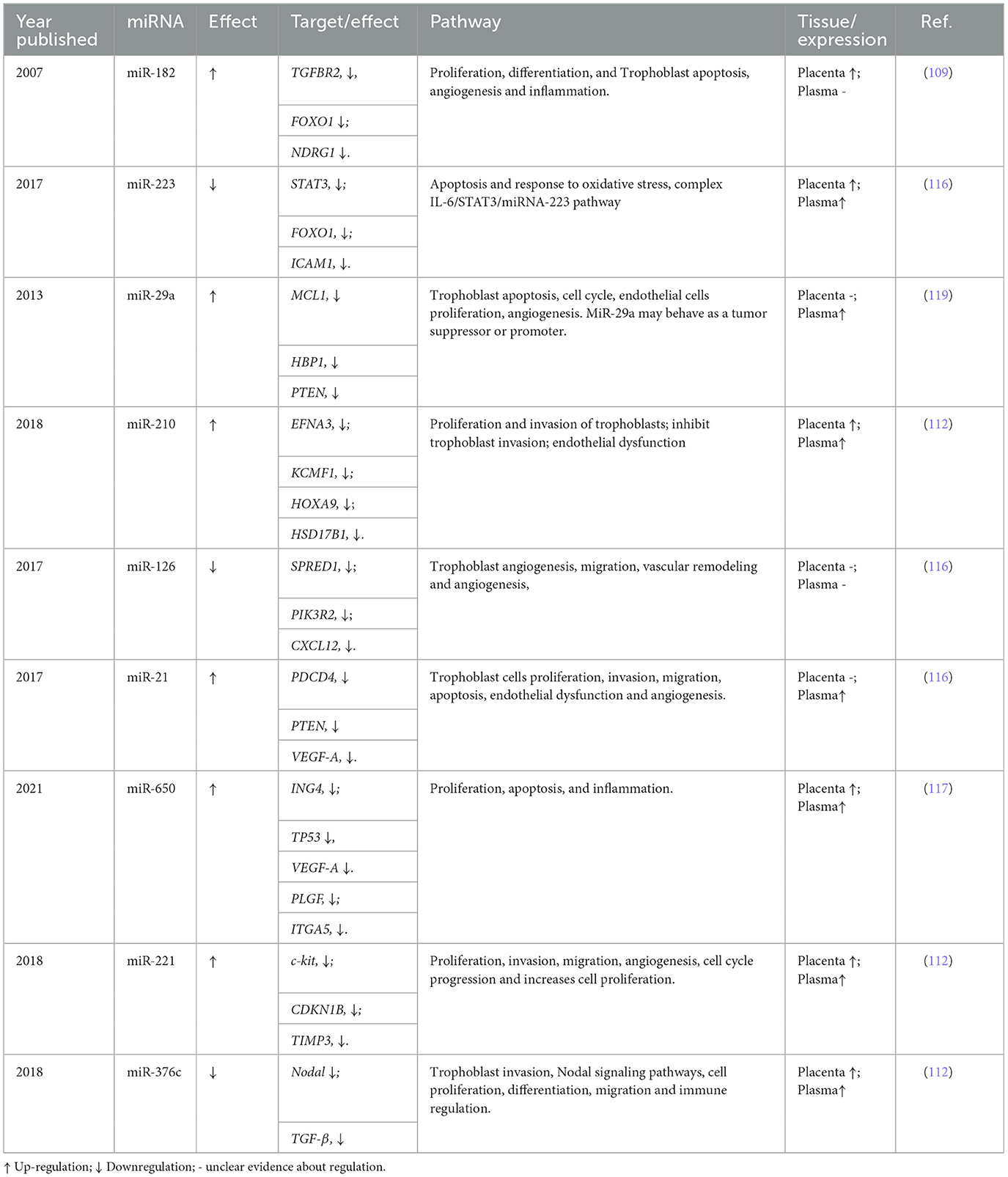
Table 3. Key miRNAs associated with the development of preeclampsia.
The first study that described miRNAs associated with PE was by Pineles et al. (109), in which the miRNAs miR-210 and miR-182 were highly expressed in preeclamptic placentas using RT-PCR. These miRNAs have been widely studied and are highlighted in immunological processes, being important regulators in inflammation, T cell differentiation and function, and NK cell modulation (113, 114). Subsequently, other miRNAs associated with PE were identified, such as miR-155, which is upregulated in the placentas of women with PE and can affect trophoblast migration and differentiation by inhibiting cell proliferation through cyclin D1 suppression (108, 115).
From these findings, different research groups, using various approaches, have identified numerous miRNA candidates and biological pathways related to PE, allowing PE to be recognized beyond the antiangiogenic hypothesis.
6.3.6 Stage 2: maternal systemic response
Among these studies, one by Jairajpuri et al. (116), using expression microarrays, evaluated the differential expression profile of circulating maternal miRNAs concerning disease severity. They identified an increase in the expression of miR-155, miR-210, miR-215, miR-650, and miR-21, and a decrease in miR-18a and miR-19b1. Additionally, they identified four new miRNAs: miR-518b and miR-29a, which were upregulated, and miR-144 and miR-15b, which were downregulated in severe PE compared to mild PE. These results confirm that miRNAs not only contribute to the pathogenesis of PE but can also predict its severity (116).
On the other hand, Akgör et al. (117) used RT-PCR to analyze circulating maternal miRNAs and identified 11 miRNAs related to PE, suggesting their use as potential biomarkers for the non-invasive prediction and diagnosis of the disease. Eight of these miRNAs (miR-210, miR-375, miR-197-3p, miR-132-3p, miR-29a-3p, miR-328, miR-24-3p, and miR-218-5p) were upregulated, while the remaining three (miR-302b-3p, miR-191-5p, and miR-17-5p) were downregulated (117). These results add to the evidence on miR-24 and miR-29a, which are involved in biological processes such as angiogenesis, inflammation, and hypoxia as potential markers for PE (118, 119).
In 2021, a pioneering study provided a comprehensive analysis of the human placental transcriptome using high-coverage RNA-Seq in rRNA-depleted placental samples (~100 million reads in 302 placental samples). Additionally, other RNA species, such as miRNA and circRNA, were characterized. The study found that the placenta contains multiple abundant and common circRNAs. circRNAs, functioning as miRNA sponges, can exert regulatory effects on them. In this study, a new circRNA (circSTS; chrX: 7,514,882–7,516,290) was identified that could modify the function of miR-5584-5p and miR-7113-5p. However, the functional consequences of the miRNA–circRNA interaction remain poorly understood, so these findings should be taken with caution (100).
Integrative analyses of miRNA, circRNA, and DEGs enabled the construction of gene networks and interaction pathways, revealing dysregulated gene sets in PE, particularly those involving the PSG (pregnancy-specific glycoprotein) family. Additionally, other key reproductive importance genes were identified, such as KIR immune receptor genes, which mediate the interaction between natural killer cells in the uterus and trophoblast cells. The study also found that 15% of women with PE and 8% with fetal growth restriction (FGR) had FSTL3 levels higher than 61.4 ng/ml. The study identifies overlapping transcriptomic alterations in PE and FGR, with FSTL3 emerging as a promising biomarker for adverse pregnancy outcomes (100).
Despite the abundant evidence and application in understanding PE through transcriptomic studies, the clinical application of cfRNA and other RNA species in early or mid-pregnancy detection is limited. Particularly, they tend to be disregarded in favor of usual detection methods based on biochemical, biophysical, and ultrasound markers, which may be more cost-effective and efficient (120). The study of the association between miRNAs and PE is relatively recent. However, it is essential to carry out more research that provides an understanding of the cellular miRNA mechanisms involved in developing PE. From the perspective of the two-stage model, altered miRNA and circRNA expression observed in the placenta reflects the first stage of defective placentation, while their detection in maternal blood highlights the second stage, where systemic responses become clinically evident. Together, these findings position non-coding RNA species as promising, though still emerging, biomarkers for both stages of the disease continuum.
6.3.7 Proteomics in preeclampsia biomarker discovery
6.3.7.1 Placental dysfunction
Proteomics studies provide an integrated view of individual disease processes at the protein level through the simultaneous analysis of hundreds of proteins, enabling the comparison of protein patterns between healthy and diseased groups (121). These studies have been primarily conducted using techniques such as Mass Spectrometry (MS) for the qualitative and quantitative analysis of proteins and their post-translational modifications (122). MS-Proteomics techniques have contributed to the identification of new biomarkers of PE in a range of biological specimens, including placental tissue, placental trophoblast cells, and maternal biofluids such as blood serum, plasma, and urine (57). However, biofluids have been prioritized as noninvasive samples, making the discovery of differentially regulated proteins in these tissues more promising for clinical biomarker applications. Table 4 presents a summary of the main findings in proteomic studies of these tissues.
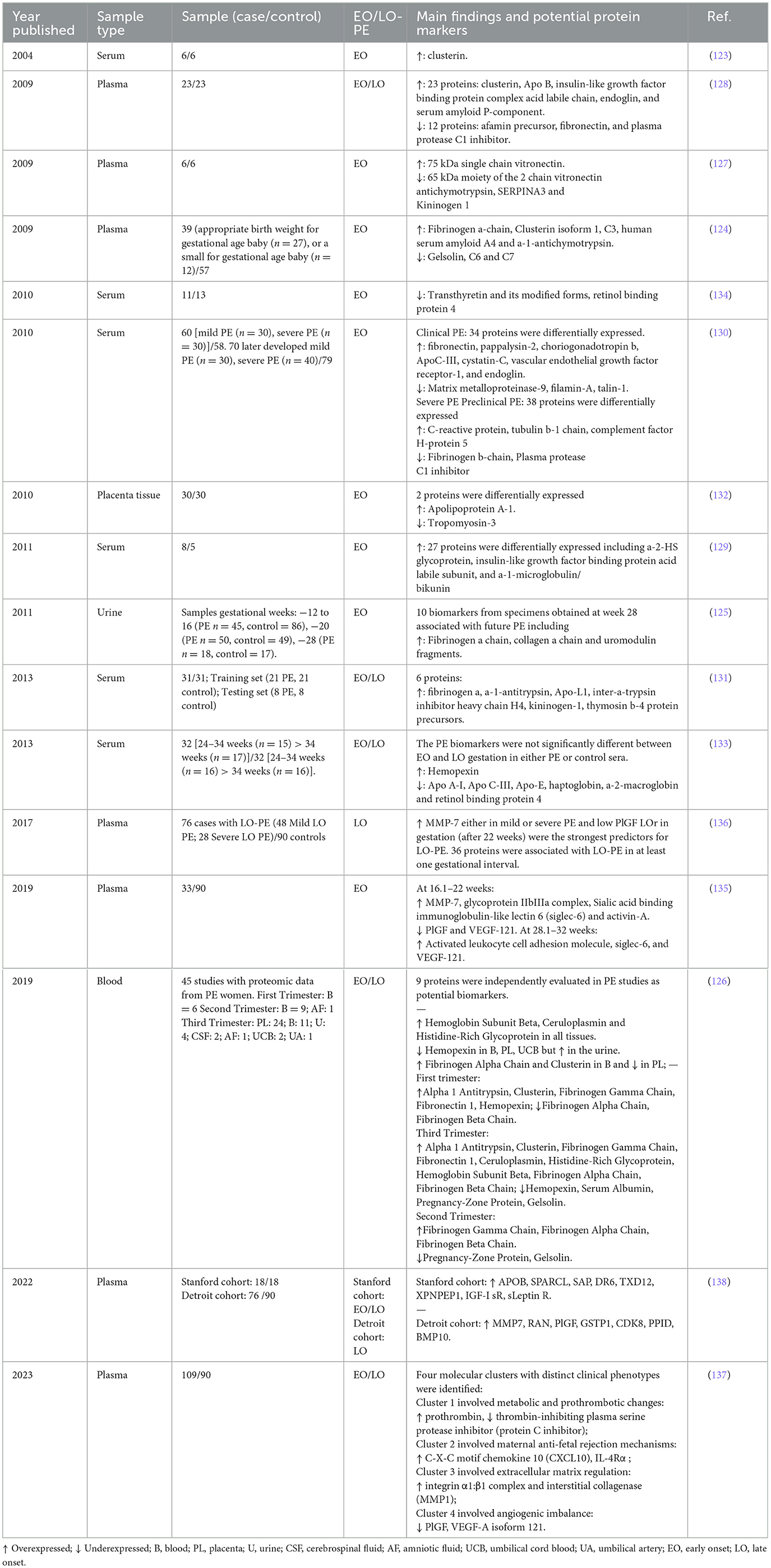
Table 4. Proteomic biomarkers linked to preeclampsia.
The first report of a proteomic study on PE was published in 2004 by Watanabe et al. They performed two-dimensional electrophoresis (2-DE) on sera from six patients with PE and six normal pregnant women, followed by MALDI-TOF-MS and peptide mass fingerprinting, identifying overexpressed Clusterin in the PE group. This finding was confirmed in sera from an additional 80 preeclamptic women and 80 normal pregnant women using immunoassays, which showed that Clusterin levels were significantly higher in preeclamptic women (123). Clusterin, a secreted heat shock protein or chaperone molecule in response to stress, potentially offers protection against cytotoxic agents (123). It has been proposed that Clusterin overexpression may result from endothelial dysfunction or renal issues related to PE (123). Additionally, Clusterin has been reported as upregulated in several proteomic studies, including both EO-PE and LO-PE (Table 4). These findings suggest that Clusterin could be a potential global biomarker for PE.
Most proteomic studies have focused on the identification of biomarkers in EO-PE, leading to the discovery of various potential biomarkers, primarily in serum and plasma. These biomarkers are associated with processes central to abnormal placental development, such as inflammation, coagulation, angiogenesis, and lipid metabolism. In addition to Clusterin, the most common findings in EO-PE include the overexpression of proteins such as multiple isoforms of fibrinogen, including the fibrinogen alpha chain (124–126), and vitronectin (127). Inflammatory and acute phase proteins such as serum amyloid P-component (128), alpha-2-HS glycoprotein (129), and C-reactive protein (130) have also been consistently overexpressed. Similarly, protease inhibitors like alpha-1-antitrypsin (131) have shown increased levels. Additionally, some studies have found overexpression of apolipoproteins such as Apo B (128) and Apo A-1 (132), although Apo A-1 has also been found to be underexpressed in other studies, along with Apo C-III and Apo-E (133).
Underexpressed proteins found in PE include inflammatory and acute phase proteins such as transthyretin and retinol-binding protein 4 (134), protease inhibitors such as plasma protease C1 inhibitor (128), antichymotrypsin, SERPINA3, and kininogen 1 (127). Additionally, proteins related to the innate immune system, such as gelsolin and complement proteins C6 and C7, have also been underexpressed (124). Proteomic studies have further supported the underexpression of angiogenic factors such as VEGF and PlGF (135), consistent with impaired placental angiogenesis.
6.3.8 Stage 2: maternal systemic response
Although fewer studies have focused specifically on LO-PE, proteomics has identified several biomarkers reflecting systemic vascular, immune, and metabolic responses. The pioneering study by Erez et al. (136) focused on LO-PE and identified the overexpression of MMP-7 in both mild and severe LO-PE, with low PlGF levels being the strongest predictors for LO-PE. Other overexpressed proteins identified in LO-PE include prothrombotic proteins such as prothrombin (137); inflammatory proteins such as C-X-C motif chemokine 10 (CXCL10) and Interleukin-4 Receptor Alpha (IL-4Rα) (137); and proteins related to extracellular matrix regulation, such as the integrin α1:β1 complex and interstitial collagenase (MMP1) (130). Additional proteins such as APOB, SPARCL, SAP, and DR6 have also been reported as overexpressed (138).
Several underexpressed proteins in LO-PE have also been described, including the thrombin-inhibiting plasma serine protease inhibitor (protein C inhibitor) (137). Additionally, underexpressed angiogenic proteins such as PlGF and VEGF-A isoform 121 have been observed in LO-PE, consistent with the antiangiogenic state that characterizes the disease (137). These patterns reflect the systemic endothelial dysfunction, prothrombotic changes, and inflammatory environment that dominate the second stage of PE.
Proteomic studies have contributed significantly by identifying hundreds of proteins that could serve as potential biomarkers in both early and late forms of PE. These studies have also highlighted the complexity of the mechanisms underlying the pathophysiology of PE. However, the biomarker discovery phase should be followed by biomarker verification and validation. Only a small number of the discovered markers have been further validated, such as Clusterin, Fibrinogen, Fibronectin, Angiotensinogen, Complement 4, Hemepexin, and Galectin-3, which have demonstrated clinical utility in predicting and diagnosing PE (57). Therefore, before these proteomic biomarkers can be included as clinical biomarkers, further studies and validations are necessary to ensure their reliability and clinical applicability in PE. From the perspective of the two-stage model, proteomic alterations in EO-PE reflect primary placental dysfunction, while those in LO-PE highlight maternal systemic inflammatory and vascular responses. Together, proteomics bridges both stages of PE pathogenesis and underscores the need for integrated multi-omics validation.
6.3.9 The role of metabolomics in preeclampsia biomarking
6.3.9.1 Stage 1: placental dysfunction
Another promising approach in biomarker discovery is metabolomics, which aims to identify metabolites correlated with diseases and environmental exposures. Metabolites are the substrates and products of metabolism that drive essential cellular functions such as energy production and storage, signal transduction, and apoptosis (139, 140). Metabolomic approaches focus on identifying metabolites and metabolic pathways associated with phenotypes and integrating this knowledge with functional and mechanistic biological studies (141). Given that metabolite biosignatures from human biofluids provide a link between genotype, environment, and phenotype (6), significant efforts have been directed toward identifying these biomarkers for the prediction and diagnosis of PE.
The main methodologies used for metabolite recovery and identification are untargeted (global) (UtM) and targeted mass spectrometry-based metabolomics (TM). UtM measures a broad range of metabolites in a sample without prior knowledge, while TM offers higher sensitivity and selectivity by analyzing metabolites based on prior information, typically used to validate and expand upon UtM results (141). Metabolomic studies aimed at discovering PE biomarkers have analyzed biological specimens like tissues (e.g., placenta) and biofluids. Various methods have been employed to measure potential biomarker molecules, including amino acids, fatty acids, organic acids, and acylcarnitines (142).
One of the first reported MS-UtM studies in PE was in 2004 (143). The study by Jain, Jayasimhulu, and Clark aimed to characterize phospholipids (PHLs) in the placenta of normotensive and PE women. The study found a notable deficiency in plasmenyl phosphatidylethanolamine (PPE), which was associated with oxidative stress as an additional biological factor linked to the pathogenesis of PE. Moreover, an increase in free fatty acids, particularly arachidonic acid, was observed in the PE placentas. This could result from heightened activity of placental phospholipase A2, leading to the production of potent vasoconstrictors such as thromboxanes and isoprostanes, which are elevated in PE.
6.3.9.2 Stage 2: maternal systemic response
In 2007, Turner et al. (144) reported the first MS-UtM study in biofluids, aiming to establish the metabolic profile in plasma of normal and preeclamptic women. The study included 11 normotensive pregnant women and 11 preeclamptic women. The results revealed lower concentrations of lipids such as very-low-density lipoprotein (VLDL) and lower levels of ketone bodies like 3-hydroxybutyrate and acetoacetate in preeclamptic women, suggesting that lipid and ketone body constituents contribute to the discrimination between the two groups (144). These findings support the oxidative stress mechanism as a significant underlying factor in the pathogenesis of PE. Additionally, other metabolites such as glycoprotein, proline, lactate, and various amino acids were present in lower concentrations in the PE group.
In 2008, Kenny et al. (145) conducted a study with an amplified sample that included plasma from 20 preeclamptic women and 20 matched controls. They identified eight metabolites overexpressed in PE: uric acid, 2-oxoglutarate, glutamate, alanine, 2-hydroxy-3-methyl-butanoic acid, 2-ethyl-3-hydroxypropionic acid, xylitol/ribitol, and creatinine, suggesting their potential as biomarkers for PE. Uric acid and 2-oxoglutarate are associated with ischemic conditions and may reflect the oxidative stress and metabolic disruptions characteristic of PE. The findings support the hypothesis that metabolic changes precede clinical symptoms of PE, allowing for early detection and targeted interventions.
In recent years, several metabolomic studies have identified various metabolites associated with PE, both in EO-PE and LO-PE. A recent systematic review by Yao et al. (140) summarized 41 human metabolomics studies conducted from 2000 to 2021, primarily focusing on biofluids to identify potential metabolic biomarkers and pathways linked to PE. The review highlighted frequently upregulated metabolites in EO-PE, including 3-hydroxyisovaleric acid, glucose, propylene glycol, hexanoylcarnitine, octenoylcarnitine, L-octanoylcarnitine, decenoylcarnitine, decanoylcarnitine, dodecenoylcarnitine, lauroylcarnitine, tetradecenoylcarnitine, and hexadecenoylcarnitine. Conversely, metabolites such as acetic acid, choline, formate, glycerol, glycine, L-phenylalanine, and succinate were found to be downregulated.
In LO-PE, commonly upregulated metabolites included glucose, octenoylcarnitine, L-octanoylcarnitine, decenoylcarnitine, decanoylcarnitine, and dodecenoylcarnitine, while downregulated metabolites included linoleylcarnitine, palmitoylcarnitine, oleylcarnitine, and stearoylcarnitine.
Pathway analysis demonstrated the significant involvement of multiple metabolic pathways. Among them, eight were related to amino acid metabolism, including arginine biosynthesis; alanine, aspartate, and glutamate metabolism; arginine and proline metabolism; glycine, serine, and threonine metabolism; valine, leucine, and isoleucine biosynthesis; phenylalanine metabolism; D-glutamine and D-glutamate metabolism; and histidine metabolism. Additionally, three pathways were linked to carbohydrate metabolism: butanoate metabolism; the citrate cycle; and glyoxylate and dicarboxylate metabolism (140). Other affected pathways included aminoacyl-tRNA biosynthesis, associated with translation, and sphingolipid metabolism, related to lipid metabolism (140). These findings indicate that these metabolites and pathways could serve as potential biomarkers for early diagnosis and provide valuable insights into the pathogenesis of PE.
Although metabolomics has highlighted different mechanisms underlying PE pathogenesis, utilizing metabolites as biomarkers presents several challenges. One of the largest challenges is overcoming interindividual metabolite variation, which arises from differences in genetic factors and environmental exposures (139, 140). Additionally, evaluating the biological roles of the identified metabolites is crucial for advancing from biomarkers to understanding disease mechanisms (141). The complexity of metabolic pathways in which each metabolite can be involved complicates the inference of its function and its differences in various contexts. Thus, translating these metabolomics findings into clinical practice must involve comprehensive validation studies, robust statistical analyses to account for individual variations, and integration with other omics data to fully elucidate the biological context and relevance of the identified potential biomarkers in PE (133, 141, 146). From the perspective of the two-stage model, early placental metabolic signatures such as oxidative stress and lipid alterations correspond to Stage 1 dysfunction, whereas systemic metabolic changes in maternal biofluids reflect Stage 2 responses. Together, these findings reinforce the value of metabolomics as a complementary approach to other omics in elucidating PE pathogenesis and developing predictive biomarkers.
7 Multi-omics approach in biomarkers research in preeclampsia
Omics technologies applied to biofluids have added additional layers to the biomarker discovery process in PE. Numerous biological markers based on genomics, transcriptomics, proteomics, and metabolomics have helped identify molecular signatures involved in various biological pathways, enhancing our understanding of the underlying pathogenesis of PE. Taken together, these findings converge with the two-stage theory of the disease: genomic and transcriptomic data highlight alterations in placental development, trophoblast invasion, and angiogenic signaling characteristic of the first stage, while proteomic and metabolomic studies often reflect the maternal systemic responses that dominate the second stage, including endothelial dysfunction, inflammation, oxidative stress, and metabolic disturbances. This integrated perspective shows how each omics layer contributes to different points in the disease continuum, supporting theories related to placental perfusion/vascular resistance, endocrine, renal, and immune dysregulation, as well as additional biological processes such as insulin resistance, lipid metabolism disorders, and activated inflammatory responses.
Despite these advances, using these molecular findings as biomarkers remains challenging in efforts to predict PE. Many potential biomarkers have demonstrated poor sensitivity and limited positive predictive values (6).
Consequently, no widely accepted molecular markers have yet proven to be reliable, reproducible, and cost-effective for predicting or diagnosing PE, which obstructs their application in clinical contexts. This underscores the need for continued research and rigorous validation to improve early detection.
Integrative-omics, or multi-omics, approaches that combine two or more omics datasets for data analysis, visualization, and interpretation (141, 146) have been proposed to overcome these limitations. By integrating omics data acquired from the same samples, these approaches can provide a more comprehensive understanding of the molecular landscape of PE and improve the accuracy of diagnosis and prognosis. Recently, such approaches have gained significant interest and are being adopted by research groups worldwide to accelerate progress in understanding complex diseases like PE and in the discovery of clinically relevant biomarkers (133, 147, 148).
8 Artificial intelligence in preeclampsia: promises and challenges
Artificial Intelligence (AI) methods, including machine learning (ML) and deep learning (DL), have demonstrated significant potential in maternal and perinatal care. These technologies are particularly valuable for handling complex data to predict and diagnose PE (149). Recently, numerous predictive and classification models have been developed, with most studies focusing on AI-based approaches to predict PE using clinical and paraclinical variables (146, 150, 151).
One of the first and largest clinical studies on PE prediction was conducted in 2019 by Jhee et al. (152). This study aimed to develop ML-based models for predicting late-onset PE using electronic medical records from 11,006 pregnant women in Seoul, South Korea. Maternal clinical and laboratory data from the early second trimester to 34 weeks of gestation were analyzed, including systolic blood pressure, serum creatinine, blood urea nitrogen (BUN), platelet count, urinary protein levels, white blood cell count, and electrolyte levels (e.g., calcium and magnesium). Among the predictive models tested, the stochastic gradient boosting model outperformed others, achieving a C-statistic of 0.924, an accuracy of 97.3%, and a false positive rate of 0.009. These findings demonstrated the potential of ML algorithms to identify meaningful temporal trends, improve prediction accuracy, and surpass traditional statistical approaches.
Later that year, Sandström et al. (153) explored an innovative approach to identify nulliparous women at risk of PE using multivariable predictive models. This study leveraged electronic health data from 62,562 pregnancies and compared three predictive methods: logistic regression with pre-specified variables, backward selection, and a Random Forest model. The models were developed based on early antenatal data, including clinical, social, and medical history, alongside physical measurements such as MAP, proteinuria, hemoglobin, and glucose levels. Among these, the pre-specified logistic regression model performed best, with an AUC of 0.68 for early-onset (< 34 weeks) and preterm PE, outperforming the NICE guidelines at a 10% false positive rate. However, when considering all three models, predictive performance remained modest, with AUCs ranging from 0.58 to 0.68, indicating that none of the approaches surpassed a moderate level of discrimination. Despite progress in personalized risk assessment, the findings underscored the need for external validation and the integration of novel biomarkers to enhance predictive accuracy.
In 2020, Marić et al. (154) developed predictive models for early-onset PE using statistical learning methods. This retrospective cohort study analyzed data from 5,245 pregnancies, of which 561 cases (10.7%) involved PE. Two statistical learning algorithms—elastic net and gradient boosting—were used to construct models based on 67 variables, including maternal characteristics, medical history, vital signs, laboratory results, and medication use. The model for all PE achieved an AUC of 0.79 (95% CI: 0.75–0.83), with a sensitivity of 45.2% and a false-positive rate of 8.1%. For early-onset PE, performance was notably higher, with an AUC of 0.89 (95% CI: 0.84–0.95), a sensitivity of 72.3%, and a false-positive rate of 8.8%. Elastic net proved particularly effective for feature selection, identifying key predictors such as chronic hypertension, a history of preeclampsia, systolic and diastolic blood pressure, and laboratory findings like proteinuria and glucose levels. These results highlighted the utility of statistical learning methods for accurate early prediction of PE.
A recent longitudinal study conducted in China (155), followed 4,644 pregnant women from early pregnancy to develop predictive models for both term (≥37 weeks) and preterm (< 37 weeks) PE. Clinical variables such as maternal age, pre-pregnancy weight, parity, and medical history (e.g., chronic hypertension, diabetes, and systemic lupus erythematosus) were recorded. Biophysical markers, including MAP and UtA-PI, and biochemical markers, such as pregnancy-associated plasma protein A (PAPP-A) and PlGF, were collected during the first trimester. The Voting Classifier achieved the highest performance for predicting preterm PE (AUC: 0.884; sensitivity: 86%; specificity: 83.4%), while Logistic Regression reached an AUC of 0.826 for overall PE prediction, with MAP contributing 44.3% to model accuracy. The inclusion of PlGF and UtA-PI enhanced predictive accuracy for preterm PE. While ML algorithms demonstrated robust performance, their results were comparable to traditional models such as the competing risk model by the Fetal Medicine Foundation (AUC = 0.831 vs. 0.797) (155).
A systematic review by Ranjbar et al. (150), evaluated 128 studies focusing on ML models for PE prediction, integrating maternal characteristics, clinical history, and paraclinical findings. The predictors analyzed included maternal age, body mass index (BMI), serum creatinine, uric acid, platelet count, and ultrasound findings such as uterine artery pulsatility index. The review highlighted nine ML models, with Stochastic Gradient Boosting achieving the highest AUC (0.973) and a false positive rate of 0.009 for late-onset PE prediction. Extreme Gradient Boosting showed strong performance for early-onset PE (AUC: 0.955), while Random Forest demonstrated robust general prediction (AUC: 0.860) (150). These findings emphasize the ability of ML methods to achieve high accuracy with minimal false positives.
Despite their promising results, versatility, and the availability of multimodal and multivariable modeling techniques, these models face significant challenges in clinical generalization. Population variability, inconsistent feature definitions, and limited external validation continue to hinder their broader application. Nevertheless, AI's ability to process extensive and complex datasets offers a valuable opportunity to integrate multi-omics data with clinical and paraclinical variables. This integration presents a promising pathway to address the complexity of PE and improve maternal and fetal care (149).
8.1 Artificial intelligence and multi-omics integration
Multi-omics approaches present significant challenges in data processing and interpretation. Consequently, bioinformatics and computational tools are indispensable for minimizing redundancy in these intricate datasets and facilitating the identification of the most relevant metabolites. The rapid expansion of the omics field and digital resources, combined with the vast data and complexity inherent in multi-omics, surpass traditional computational workflows, underscoring the need for innovation. Automated computational workflows markedly expedite data upload and mining processes, offering novel methods for the identification and biological interpretation of data. In this context, AI methods are becoming potent resources for handling multi-omics data (146, 150, 151).
Multi-omics and AI produce complex datasets that necessitate computational tools to identify and correlate omics data across samples, examining their interconnectivity in metabolic pathways relative to the phenotype or pathological processes—patterns that are undetectable by traditional methods. Although AI methods have been applied to PE, particularly utilizing clinical and paraclinical data to efficiently predict PE risk (152, 156–158), the potential of integrating AI methods and multi-omics approaches in PE has not been extensively explored. To our knowledge, the only comprehensive multi-omics study was conducted by Marić et al. (154) in 2022. This prospective study included 33 women in the discovery cohort (17 preeclamptic, 16 normotensive) and 16 women in the validation cohort (12 preeclamptic, 4 normotensive), incorporating transcriptomics, proteomics, metabolomics, lipidomic, and microbiome data from longitudinally collected samples from pregnant women. The study also integrated immune system mass cytometry data for a subset of women and combined it with clinical and demographic data.
For early prediction of preeclampsia within the first 16 weeks of gestation, machine learning models, specifically Elastic Net (EN) regression, were employed to extract the most predictive features from high-dimensional datasets. The urine metabolome model achieved the highest performance (AUC = 0.88), identifying predictive metabolites such as adenine and carnitines, while the proteome model followed closely (AUC = 0.87), with proteins including endothelial growth factor A (VEGFA), leptin (LEP), vascular, L-selectin (SELL), E-selectin (SELE), kinase transmembrane receptor (ROR1), and C-X-C motif chemokine ligand 10 (CXCL10), showing strong predictive value. Validation of the urine metabolite model in an independent cohort yielded an AUC of 0.83. Importantly, integration of multi-omics datasets further improved prediction accuracy (AUC ≈ 0.91). Pathway analyses highlighted tryptophan, caffeine, and arachidonic acid metabolism, while integration with immune cytometry data revealed novel associations between immune and proteomic dynamics (148).
The integration of multi-omics and AI methods holds promise in unraveling the biological mechanisms underlying PE, offering opportunities for biomarker discovery and validation. However, significant challenges remain, the largest being Biomedical Data Inequality (159–161). Recent statistics indicate that over 80% of data from clinical omics studies are derived from individuals of European ancestry, who constitute < 20% of the global population (160). Most PE studies predominantly feature cohorts from Caucasian populations (Table 2). Although AI is revolutionizing biomedical research and healthcare, it also has the potential to exacerbate data inequality, leading to negative impacts (160). Therefore, it is imperative to incentivize omics research with a multiethnic focus and strengthen research efforts in developing countries. This will enable the inclusion of representative cohorts, facilitating the validation and emergence of biomarkers for PE that are truly applicable in clinical contexts worldwide.
9 Final remarks and conclusions
Preeclampsia has long been described as a “babel of schemata,” reflecting the persistent difficulty in defining its diagnostic boundaries. The evolution from early clinical observations of hypertension and proteinuria to the use of conventional biomarkers such as proteinuria and blood pressure measurement, and later to angiogenic and anti-angiogenic markers like sFlt-1, PlGF, and soluble endoglin, illustrates how the field has moved from descriptive criteria toward a more mechanistic understanding. Yet, these biomarkers still show limited sensitivity and predictive value, underscoring the need for combined approaches.
The integration of multi-omics approaches with artificial intelligence represents a promising frontier for PE prediction and diagnosis. AI can process complex, multidimensional datasets and enhance risk stratification, paving the way toward precision medicine. However, challenges remain regarding external validation, population variability, and limited generalizability, which currently restrict the clinical adoption of AI-based models.
Beyond these methodological hurdles, the translation of omics technologies themselves into clinical practice remains limited. They demand specialized equipment, trained personnel, and high costs that often confine their use to research settings. These barriers are most evident in low- and middle-income countries, where routine application is impractical and risks widening inequities in maternal health. Simplified essays, cost reduction, and context-specific validation will be essential for equitable implementation in PE.
Therefore, clinical translation remains the greatest challenge. Large, multiethnic validation studies and standardized strategies are urgently required to ensure reproducibility and applicability across diverse populations. Bridging discovery with implementation will be critical to transform biomarker and AI research into effective clinical practice and improve maternal and perinatal outcomes.
Author contributions
JR-M: Conceptualization, Formal analysis, Writing – original draft, Writing – review & editing. EV-C: Conceptualization, Formal analysis, Writing – original draft, Writing – review & editing. AP: Writing – original draft, Writing – review & editing. LD-Q: Writing – original draft, Writing – review & editing. VR-R: Conceptualization, Formal analysis, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This review was funded by Hospital Universitario Mayor-Méderi (Project No. C-011-2023).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abbreviations
ACOG, American College of Obstetricians and Gynecologists; AHA, American Heart Association; AI, artificial intelligence; ALK1, activin receptor-like kinase 1; ANGPTL4, angiopoietin-like protein 4; APO, apolipoprotein; BP, blood pressure; BP-PRS, blood pressure polygenic risk scores; cfRNA, cell-free RNA; circRNA, circular RNA; CX3CL1, fractalkine; DEG, differentially expressed genes; DL, deep learning; EO-PE, early-onset preeclampsia; ENG, endoglin; FIGO, International Federation of Gynecology and Obstetrics; FGR, fetal growth restriction; FLT1, fms-related tyrosine kinase 1; FMF, Fetal Medicine Foundation; GWAS, genome-wide association study; HDP, hypertensive disorders of pregnancy; HIF-1α, hypoxia-inducible factor 1 alpha; HLA, human leukocyte antigen; IL, interleukin; ISSHP, international society for the study of hypertension in pregnancy; IUGR, intrauterine growth restriction; LO-PE, late-onset preeclampsia; LXR, liver X receptor; MAP, mean arterial pressure; miRNA, MicroRNA; ML, machine learning; MMP, matrix metalloproteinase; MS, mass spectrometry; NGS, next-generation sequencing; PE, preeclampsia; PE-PRS, preeclampsia polygenic risk scores; PlGF, placental growth factor; PRS, polygenic risk score; RAS, renin-angiotensin system; RCOG, Royal College of Obstetricians and Gynecologists; RNA-seq, RNA sequencing; SOGC, Society of Obstetricians and Gynecologists of Canada; SNP, single nucleotide polymorphism; SOMANZ, Society of Obstetric Medicine of Australia and New Zealand; sENG, soluble endoglin; sFlt-1, soluble fms-like tyrosine kinase-1; sVEGFR-1, soluble vascular endothelial growth factor receptor-1; TGF-β, transforming growth factor beta; TM, targeted metabolomics; UtA-PI, uterine artery pulsatility index; UtM, untargeted metabolomics; VEGF, vascular endothelial growth factor.
References
1. Magee LA, Brown MA, Hall DR, Gupte S, Hennessy A, Karumanchi SA, et al. The 2021 international society for the study of hypertension in pregnancy classification, diagnosis & management recommendations for international practice. Pregnancy Hypertens. (2022) 27:148–69. doi: 10.1016/j.preghy.2021.09.008
2. Magee LA, Nicolaides KH, von Dadelszen P. Preeclampsia. N Engl J Med. (2022) 386:1817–32. doi: 10.1056/NEJMra2109523
3. Kassebaum N, Steiner C, Murray CJL, Lopez AD. Global, regional, and national levels of maternal mortality, 1990–2015: a systematic analysis for the Global Burden of Disease Study 2015. Lancet. (2016) 388:1775–812. doi: 10.1016/S0140-6736(16)31470-2
4. Tanner MS, Davey MA, Mol BW, Rolnik DL. The evolution of the diagnostic criteria of preeclampsia- eclampsia. Am J Obstet Gynecol. (2022) 226:S835–43. doi: 10.1016/j.ajog.2021.11.1371
5. Polsani S, Phipps E, Jim B. Emerging new biomarkers of preeclampsia. Adv Chronic Kidney Dis. (2013) 20:271–9. doi: 10.1053/j.ackd.2013.01.001
6. Qiu S, Cai Y, Yao H, Lin C, Xie Y, Tang S, et al. Small molecule metabolites: discovery of biomarkers and therapeutic targets. Signal Transduct Target Ther. (2023) 8:132. doi: 10.1038/s41392-023-01399-3
7. Erez O, Romero R, Jung E, Chaemsaithong P, Bosco M, Suksai M, et al. Preeclampsia/eclampsia: the conceptual evolution of a syndrome - PMC. Am J Obstet Gynecol. (2023) 226:1–42. doi: 10.1016/j.ajog.2021.12.001
8. Bell MJ. A historical overview of preeclampsia-eclampsia. JOGNN J Obstet Gynecol Neonatal Nurs. (2010) 39:510–8. doi: 10.1111/j.1552-6909.2010.01172.x
9. Lees C, Ferrazzi E. Relevance of haemodynamics in treating pre-eclampsia. Curr Hypertens Rep. (2017) 19:3–7. doi: 10.1007/s11906-017-0766-6
10. Rubin DB, Drab EA, Ward WF, Bauer KD, Nov N. Cell cycle progression in irradiated endothelial cells cultured from bovine aorta. JSTOR. (2016) 116:364–71. doi: 10.2307/3577473
11. Yildiz GA, Yilmaz EPT. The association between protein levels in 24-hour urine samples and maternal and neonatal outcomes of pregnant women with preeclampsia. J Turkish Ger Gynecol Assoc. (2022) 23:190–8. doi: 10.4274/jtgga.galenos.2022.2022-4-3
13. Friedman EA, Neff RK. Pregnancy Hypertension: A Systematic Evaluation of Clinical Diagnostic Criteria. Littleton, CO: PSG Publishing Company (1977).
14. Reddy M, Rolnik DL, Harris K, Li W, Mol BW, Da Silva Costa F, et al. Challenging the definition of hypertension in pregnancy: a retrospective cohort study. Am J Obstet Gynecol. (2020) 222:606.e1–21. doi: 10.1016/j.ajog.2019.12.272
15. Porcelli BA, Diveley E, Meyenburg K, Woolfolk C, Rosenbloom JI, Raghuraman N, et al. A new definition of gestational hypertension? New-onset blood pressures of 130 to 139/80 to 89 mm Hg after 20 weeks of gestation. Am J Obstet Gynecol. (2020) 223:442.e1–7. doi: 10.1016/j.ajog.2020.06.019
16. Hecht JL, Ordi J, Carrilho C, Ismail MR, Zsengeller ZK, Karumanchi SA, et al. The pathology of eclampsia: an autopsy series. Hypertens Pregnancy. (2017) 36:259–68. doi: 10.1080/10641955.2017.1329430
17. Goodlin RC. Severe pre-eclampsia: another great imitator. Am J Obstet Gynecol. (1976) 125:747–52. doi: 10.1016/0002-9378(76)90841-3
18. Brown MA, Hague WM, Higgins J, Lowe S, McCowan L, Oats J, et al. The detection, investigation and management of hypertension in pregnancy: executive summary. Aust New Zeal J Obstet Gynaecol. (2000) 40:133–8. doi: 10.1111/j.1479-828X.2000.tb01136.x
19. Tay J, Foo L, Masini G, Bennett PR, McEniery CM, Wilkinson IB, et al. Early and late preeclampsia are characterized by high cardiac output, but in the presence of fetal growth restriction, cardiac output is low: insights from a prospective study. Am J Obstet Gynecol. (2018) 218:517.e1–12. doi: 10.1016/j.ajog.2018.02.007
20. Masini G, Foo LF, Tay J, Wilkinson IB, Valensise H, Gyselaers W, et al. Preeclampsia has two phenotypes which require different treatment strategies. Am J Obstet Gynecol. (2022) 226:S1006–18. doi: 10.1016/j.ajog.2020.10.052
21. Roberts JM, Rich-Edwards JW, McElrath TF, Garmire L, Myatt L, GPC. Subtypes of preeclampsia: recognition and determining clinical usefulness. Nature. (2008) 456:814–8. doi: 10.1161/HYPERTENSIONAHA.120.14781
22. Robillard PY, Dekker G, Scioscia M, Saito S. Progress in the understanding of the pathophysiology of immunologic maladaptation related to early-onset preeclampsia and metabolic syndrome related to late- onset preeclampsia. Am J Obstet Gynecol. (2022) 226:S867–75. doi: 10.1016/j.ajog.2021.11.019
23. Oudejans CBM, van Dijk M, Oosterkamp M, Lachmeijer A, Blankenstein MA. Genetics of preeclampsia: paradigm shifts. Hum Genet. (2007) 120:607–12. doi: 10.1007/s00439-006-0259-1
24. Fishel Bartal M, Lindheimer MD, Sibai BM. Proteinuria during pregnancy: definition, pathophysiology, methodology, and clinical significance. Am J Obstet Gynecol. (2022) 226:S819–34. doi: 10.1016/j.ajog.2020.08.108
25. Redman CW, Sargent IL. Latest advances in understanding preeclampsia. Science. (2005) 308:1592–4. doi: 10.1126/science.1111726
26. MacDonald TM, Walker SP, Hannan NJ, Tong S, Kaitu'u-Lino TJ. Clinical tools and biomarkers to predict preeclampsia. eBioMedicine. (2022) 75:103780. doi: 10.1016/j.ebiom.2021.103780
27. Zhang J, Klebanoff MA, Roberts JM. Prediction of adverse outcomes by common definitions of hypertension in pregnancy. Obstet Gynecol. (2001) 97:261–7. doi: 10.1097/00006250-200102000-00018
28. Khan N, Andrade W, De Castro H, Wright A, Wright D, Nicolaides KH. Impact of new definitions of pre- eclampsia on incidence and performance of first-trimester screening. Ultrasound Obstet Gynecol. (2020) 55:50–7. doi: 10.1002/uog.21867
29. Bujold E, Romero R, Chaiworapongsa T, Kim YM, Kim GJ, Kim MR, et al. Evidence supporting that the excess of the sVEGFR-1 concentration in maternal plasma in preeclampsia has a uterine origin. J Matern neonatal Med Off J Eur Assoc Perinat Med Fed Asia Ocean Perinat Soc Int Soc Perinat Obstet. (2005) 18:9–16. doi: 10.1080/14767050500202493
30. Baker PN, Krasnow J, Roberts JM, Yeo KT. Elevated serum levels of vascular endothelial growth factor in patients with preeclampsia. Obstet Gynecol. (1995) 86:815–21. doi: 10.1016/0029-7844(95)00259-T
31. Lee ES, Oh M-J, Jung JW, Lim J-E, Seol H-J, Lee K-J, et al. The levels of circulating vascular endothelial growth factor and soluble Flt-1 in pregnancies complicated by preeclampsia. J Korean Med Sci. (2007) 22:94–8. doi: 10.3346/jkms.2007.22.1.94
32. Koga K, Osuga Y, Yoshino O, Hirota Y, Ruimeng X, Hirata T, et al. Elevated serum soluble vascular endothelial growth factor receptor 1 (sVEGFR-1) levels in women with preeclampsia. J Clin Endocrinol Metab. (2003) 88:2348–51. doi: 10.1210/jc.2002-021942
33. Levine RJ, Maynard SE, Qian C, Lim KH, England LJ Yu KF, et al. Circulating angiogenic factors and the risk of preeclampsia. Surv Anesthesiol. (2005) 49:14–5. doi: 10.1056/NEJMoa031884
34. Ahmad S, Ahmed A. Elevated placental soluble vascular endothelial growth factor receptor-1 inhibits angiogenesis in preeclampsia. Circ Res. (2004) 95:884–91. doi: 10.1161/01.RES.0000147365.86159.f5
35. Maynard SE, Min J-Y, Merchan J, Lim K-H, Li J, Mondal S, et al. Excess placental soluble fms-like tyrosine kinase 1 (sFlt1) may contribute to endothelial dysfunction, hypertension, and proteinuria in preeclampsia. J Clin Invest. (2003) 111:649–58. doi: 10.1172/JCI200317189
36. Nirupama R, Divyashree S, Janhavi P, Muthukumar SP, Ravindra PV. Preeclampsia: pathophysiology and management. J Gynecol Obstet Hum Reprod. (2021) 50:101975. doi: 10.1016/j.jogoh.2020.101975
37. Caniggia I, Mostachfi H, Winter J, Gassmann M, Lye SJ, Kuliszewski M, et al. Hypoxia-inducible factor-1 mediates the biological effects of oxygen on human trophoblast differentiation through TGFβ3. J Clin Invest. (2000) 105:577–87. doi: 10.1172/JCI8316
38. Le Jan S, Amy C, Cazes A, Monnot C, Lamandé N, Favier J, et al. Angiopoietin-like 4 is a proangiogenic factor produced during ischemia and in conventional renal cell carcinoma. Am J Pathol. (2003) 162:1521–8. doi: 10.1016/S0002-9440(10)64285-X
39. Karumanchi SA, Bdolah Y. Hypoxia and sFlt-1 in preeclampsia: the “chicken-and-egg” question. Endocrinology. (2004) 145:4835–7. doi: 10.1210/en.2004-1028
40. Liu L, Zhuang X, Jiang M, Guan F, Fu Q, Lin J. Angptl4 mediates the protective role of pparγ activators in the pathogenesis of preeclampsia. Cell Death Dis. (2017) 8:1–12. doi: 10.1038/cddis.2017.419
41. Açikgözoglu MK, Pala S, Atilgan R, Ilhan N, Ilhan N. High serum angiopoietin-like protein-4 levels are associated with gestational hypertension and preeclampsia: a case-control study. Turkish J Biochem. (2024) 49:344–8. doi: 10.1515/tjb-2023-0087
42. Shu L, Wang C, Ding Z, Tang J, Zhu Y, Wu L, et al. A novel regulated network mediated by downregulation HIF1A-AS2 lncRNA impairs placental angiogenesis by promoting ANGPTL4 expression in preeclampsia. Front Cell Dev Biol. (2022) 10:1–13. doi: 10.3389/fcell.2022.837000
43. Pala S, Atilgan R, Ilhan N. High amniotic fluid fractalkine and MIP-1β levels are associated with intrauterine growth restriction: a prospective cohort study. Turkish J Med Sci. (2024) 54:280–90. doi: 10.55730/1300-0144.5789
44. Kahouadji S, Giguère Y, Lambert S, Forest JC, Bernard N, Blanchon L, et al. CX3CL1/Fractalkine as a biomarker for early pregnancy prediction of preterm premature rupture of membranes. Clin Chem Lab Med. (2024) 62:1101–8. doi: 10.1515/cclm-2023-1202
45. Szukiewicz D, Kochanowski J, Mittal TK, Pyzlak M, Szewczyk G, Cendrowski K. CX3CL1 (fractalkine) and TNFα production by perfused human placental lobules under normoxic and hypoxic conditions in vitro: the importance of CX3CR1 signaling. Inflamm Res. (2014) 63:179–89. doi: 10.1007/s00011-013-0687-z
46. Siwetz M, Dieber-Rotheneder M, Cervar-Zivkovic M, Kummer D, Kremshofer J, Weiss G, et al. Placental fractalkine is up-regulated in severe early-onset preeclampsia. Am J Pathol. (2015) 1851334–43. doi: 10.1016/j.ajpath.2015.01.019
47. Szewczyk G, Pyzlak M, Pankiewicz K, Szczerba E, Stangret A, Szukiewicz D, et al. The potential association between a new angiogenic marker fractalkine and a placental vascularization in preeclampsia. Arch Gynecol Obstet. (2021) 304:365–76. doi: 10.1007/s00404-021-05966-3
48. Imanishi T, Hano T, Nishio I. Angiotensin II potentiates vascular endothelial growth factor-induced proliferation and network formation of endothelial progenitor cells. Hypertens Res. (2004) 27:101–8. doi: 10.1291/hypres.27.101
49. Hirashima C, Ohkuchi A, Matsubara S, Suzuki H, Takahashi K, Usui R, et al. Alteration of serum soluble endoglin levels after the onset of preeclampsia is more pronounced in women with early-onset. Hypertens Res. (2008) 31:1541–8. doi: 10.1291/hypres.31.1541
50. Margioula-Siarkou G, Margioula-Siarkou C, Petousis S, Margaritis K, Vavoulidis E, Gullo G, et al. The role of endoglin and its soluble form in pathogenesis of preeclampsia. Mol Cell Biochem. (2022) 477:479–91. doi: 10.1007/s11010-021-04294-z
51. Fonsatti E, Maio M. Highlights on endoglin (CD105): from basic findings towards clinical applications in human cancer. J Transl Med. (2004) 2:18. doi: 10.1186/1479-5876-2-18
52. Venkatesha S, Toporsian M, Lam C, Hanai J, Mammoto T, Kim YM, et al. Soluble endoglin contributes to the pathogenesis of preeclampsia. Nat Med. (2006) 12:642–9. doi: 10.1038/nm1429
53. Hawinkels LJAC, Kuiper P, Wiercinska E, Verspaget HW, Liu Z, Pardali E, et al. Matrix metalloproteinase- 14 (MT1-MMP)-mediated endoglin shedding inhibits tumor angiogenesis. Cancer Res. (2010) 70:4141–50. doi: 10.1158/0008-5472.CAN-09-4466
54. Gregory AL, Xu G, Sotov V, Letarte M. Review: the enigmatic role of endoglin in the placenta. Placenta. (2014) 35:S93–9. doi: 10.1016/j.placenta.2013.10.020
55. Robinson CJ, Johnson DD. Soluble endoglin as a second-trimester marker for preeclampsia. Am J Obstet Gynecol. (2007) 197:174.e1–5. doi: 10.1016/j.ajog.2007.03.058
56. Levine RJ, Lam C, Qian C, Yu KF, Maynard SE, Sachs BP, et al. Soluble endoglin and other circulating antiangiogenic factors in preeclampsia: commentary. Obstet Gynecol Surv. (2007) 62:82–3. doi: 10.1097/01.ogx.0000253489.32189.4d
57. Dahabiyeh LA. The discovery of protein biomarkers in pre-eclampsia: the promising role of mass spectrometry. Biomarkers Biochem Indic Expo response, susceptibility to Chem. (2018) 23:609–21. doi: 10.1080/1354750X.2018.1474257
58. Verlohren S, Galindo A, Schlembach D, Zeisler H, Herraiz I, Moertl MG, et al. An automated method for the determination of the sFlt-1/PIGF ratio in the assessment of preeclampsia. Am J Obstet Gynecol. (2010) 202:161.e1–11. doi: 10.1016/j.ajog.2009.09.016
59. Biomarkers Definitions Working Group. Biomarkers and surrogate endpoints: preferred definitions and conceptual framework. Clin Pharmacol Ther. (2001) 69:89–95. doi: 10.1067/mcp.2001.113989
60. Correia CN, Nalpas NC, McLoughlin KE, Browne JA, Gordon S V, MacHugh DE, et al. Circulating microRNAs as potential biomarkers of infectious disease. Front Immunol. (2017) 8:118. doi: 10.3389/fimmu.2017.00118
61. Bates SE. Clinical applications of serum tumor markers. Ann Intern Med. (1991) 115:623–38. doi: 10.7326/0003-4819-115-8-623
62. Passaro A, Al Bakir M, Hamilton EG, Diehn M, André F, Roy-Chowdhuri S, et al. Cancer biomarkers: emerging trends and clinical implications for personalized treatment. Cell. (2024) 187:1617–35. doi: 10.1016/j.cell.2024.02.041
63. Nalejska E, Maczyńska E, Lewandowska MA. Prognostic and predictive biomarkers: tools in personalized oncology. Mol Diagn Ther. (2014) 18:273–84. doi: 10.1007/s40291-013-0077-9
64. Bernhardt AM, Tiedt S, Teupser D, Dichgans M, Meyer B, Gempt J, et al. A unified classification approach rating clinical utility of protein biomarkers across neurologic diseases. EBioMedicine. (2023) 89:104456. doi: 10.1016/j.ebiom.2023.104456
65. Hirashima C, Ohkuchi A, Arai F, Takahashi K, Suzuki H, Watanabe T, et al. Establishing reference values for both total soluble Fms-like tyrosine kinase 1 and free placental growth factor in pregnant women. Hypertens Res. (2005) 28:727–32. doi: 10.1291/hypres.28.727
66. Ohkuchi A, Hirashima C, Suzuki H, Takahashi K, Yoshida M, Matsubara S, et al. Evaluation of a new and automated electrochemiluminescence immunoassay for plasma sFlt-1 and PlGF levels in women with preeclampsia. Hypertens Res. (2010) 33:422–7. doi: 10.1038/hr.2010.15
67. Cui L, Shu C, Liu Z, Tong W, Cui M, Wei C, et al. The expression of serum sEGFR, sFlt-1, sEndoglin and PLGF in preeclampsia. Pregnancy Hypertens. (2018) 13:127–32. doi: 10.1016/j.preghy.2018.05.011
68. Stepan H, Herraiz I, Schlembach D, Verlohren S, Brennecke S, Chantraine F, et al. Implementation of the sFlt-1/PlGF ratio for prediction and diagnosis of pre-eclampsia in singleton pregnancy: implications for clinical practice. Ultrasound Obstet Gynecol. (2015) 45:241–6. doi: 10.1002/uog.14799
69. Lim JH, Lim JM, Lee HM, Lee HJ, Kwak DW, Han YJ, et al. Systematic proteome profiling of maternal plasma for development of preeclampsia biomarkers. Mol Cell Proteomics. (2024) 23:100826. doi: 10.1016/j.mcpro.2024.100826
70. Creswell L, O'Gorman N, Palmer KR, da Silva Costa F, Rolnik DL. Perspectives on the use of placental growth factor (plgf) in the prediction and diagnosis of pre-eclampsia: recent insights and future steps. Int J Womens Health. (2023) 15:255–71. doi: 10.2147/IJWH.S368454
71. Cao S, Chen R, Yang Q, He X, Chiavaioli F, Ran Y, et al. Point-of-care diagnosis of pre-eclampsia based on microfiber Bragg grating biosensor. Biosens Bioelectron. (2024) 249:116014. doi: 10.1016/j.bios.2024.116014
72. Ng KW, Chaturvedi N, Coté GL, Fisher SA, Mabbott S. Biomarkers and point of care screening approaches for the management of preeclampsia. Commun Med. (2024) 4:208. doi: 10.1038/s43856-024-00642-4
73. Soundararajan R, Khan T, von Dadelszen P. Pre-eclampsia challenges and care in low and middle-income countries: understanding diagnosis, management, and health impacts in remote and developing regions. Best Pract Res Clin Obstet Gynaecol. (2024) 96:102525. doi: 10.1016/j.bpobgyn.2024.102525
74. Kumar MM, Naik G. Maternal near miss: reaching the last mile. J Obstet Gynaecol J Inst Obstet Gynaecol. (2021) 41:675–83. doi: 10.1080/01443615.2020.1820467
75. Lubis MP, Hariman H, Lumbanraja SN, Bachtiar A. The role of placental growth factor, soluble endoglin, and uterine artery diastolic notch to predict the early onset of preeclampsia. Open access Maced J Med Sci. (2019) 7:1153–9. doi: 10.3889/oamjms.2019.154
76. Chaemsaithong P, Sahota DS, Poon LC. First trimester preeclampsia screening and prediction. Am J Obstet Gynecol. (2022) 226:S1071–97.e2. doi: 10.1016/j.ajog.2020.07.020
77. O'Gorman N, Wright D, Poon LC, Rolnik DL, Syngelaki A, de Alvarado M, et al. Multicenter screening for pre-eclampsia by maternal factors and biomarkers at 11–13 weeks' gestation: comparison with NICE guidelines and ACOG recommendations. Ultrasound Obstet Gynecol. (2017) 49:756–60. doi: 10.1002/uog.17455
78. Verlohren S, Herraiz I, Lapaire O, Schlembach D, Zeisler H, Calda P, et al. New gestational phase-specific cutoff values for the use of the soluble fms-like tyrosine kinase-1/placental growth factor ratio as a diagnostic test for preeclampsia. Hypertens (Dallas, Tex 1979). (2014) 63:346–52. doi: 10.1161/HYPERTENSIONAHA.113.01787
79. Hughes RCE, Phillips I, Florkowski CM, Gullam J. The predictive value of the sFlt-1/PlGF ratio in suspected preeclampsia in a New Zealand population: a prospective cohort study. Aust New Zeal J Obstet Gynaecol. (2023) 63:34–41. doi: 10.1111/ajo.13549
80. Rana S, Hacker MR, Modest AM, Salahuddin S, Lim KH, Verlohren S, et al. Angiogenic factors and the risk of adverse outcomes in women with suspected preeclampsia. Circulation. (2012) 125:911–9. doi: 10.1161/CIRCULATIONAHA.111.054361
81. Thadhani R, Hagmann H, Schaarschmidt W, Roth B, Cingoez T, Karumanchi SA, et al. Removal of soluble fms-like tyrosine kinase-1 by dextran sulfate apheresis in preeclampsia. J Am Soc Nephrol. (2016) 27:903–13. doi: 10.1681/ASN.2015020157
82. Rana S, Burke SD, Karumanchi SA. Imbalances in circulating angiogenic factors in the pathophysiology of preeclampsia and related disorders. Am J Obstet Gynecol. (2022) 226:S1019–34. doi: 10.1016/j.ajog.2020.10.022
83. Chaiworapongsa T, Romero R, Gomez-Lopez N, Suksai M, Gallo DM, Jung E, et al. Preeclampsia at term: evidence of disease heterogeneity based on the profile of circulating cytokines and angiogenic factors. Am J Obstet Gynecol. (2024) 230:450.e1–e18. doi: 10.1016/j.ajog.2023.10.002
84. Gray KJ, Saxena R, Karumanchi SA. Genetic predisposition to preeclampsia is conferred by fetal DNA variants near FLT1, a gene involved in the regulation of angiogenesis. Am J Obstet Gynecol. (2018) 218:211–8. doi: 10.1016/j.ajog.2017.11.562
85. Steinthorsdottir V, McGinnis R, Williams NO, Stefansdottir L, Thorleifsson G, Shooter S, et al. Genetic predisposition to hypertension is associated with preeclampsia in European and Central Asian women. Nat Commun. (2020) 11:5976. doi: 10.1038/s41467-020-19733-6
86. Siu H, Zhu Y, Jin L, Xiong M. Implication of next-generation sequencing on association studies. BMC Genomics. (2011) 12:322. doi: 10.1186/1471-2164-12-322
87. Chang CQ, Yesupriya A, Rowell JL, Pimentel CB, Clyne M, Gwinn M, et al. A systematic review of cancer GWAS and candidate gene meta-analyses reveals limited overlap but similar effect sizes. Eur J Hum Genet. (2014) 22:402–8. doi: 10.1038/ejhg.2013.161
88. Schork AJ, Thompson WK, Pham P, Torkamani A, Roddey JC, Sullivan PF, et al. All SNPs are not created equal: genome-wide association studies reveal a consistent pattern of enrichment among functionally annotated SNPs. PLoS Genet. (2013) 9:e1003449. doi: 10.1371/journal.pgen.1003449
89. Yang J, Manolio TA, Pasquale LR, Boerwinkle E, Caporaso N, Cunningham JM, et al. Genome partitioning of genetic variation for complex traits using common SNPs. Nat Genet. (2011) 43:519–25. doi: 10.1038/ng.823
90. McGinnis R, Steinthorsdottir V, Williams NO, Thorleifsson G, Shooter S, Hjartardottir S, et al. Variants in the fetal genome near FLT1 are associated with risk of preeclampsia. Nat Genet. (2017) 49:1255–60. doi: 10.1038/ng.3895
91. Kikas T, Inno R, Ratnik K, Rull K, Laan M. C-allele of rs4769613 Near FLT1 represents a high- confidence placental risk factor for preeclampsia. Hypertens (Dallas, Tex 1979). (2020) 76:884–91. doi: 10.1161/HYPERTENSIONAHA.120.15346
92. Ohwaki A, Nishizawa H, Kato A, Kato T, Miyazaki J, Yoshizawa H, et al. Placental genetic variants in the upstream region of the FLT1 gene in pre-eclampsia. J Reprod Infertil. (2020) 21:240–6. doi: 10.18502/jri.v21i4.4328
93. Zhao L, Bracken MB, DeWan AT. Genome-wide association study of pre-eclampsia detects novel maternal single nucleotide polymorphisms and copy-number variants in subsets of the Hyperglycemia and Adverse Pregnancy Outcome (HAPO) study cohort. Ann Hum Genet. (2013) 77:277–87. doi: 10.1111/ahg.12021
94. Benny PA, Alakwaa FM, Schlueter RJ, Lassiter CB, Garmire LX. A review of omics approaches to study preeclampsia. Placenta. (2020) 92:17–27. doi: 10.1016/j.placenta.2020.01.008
95. Johnson MP, Brennecke SP, East CE, Göring HHH, Kent JWJ, Dyer TD, et al. Genome-wide association scan identifies a risk locus for preeclampsia on 2q14, near the inhibin, beta B gene. PLoS ONE. (2012) 7:e33666. doi: 10.1371/journal.pone.0033666
96. Tyrmi JS, Kaartokallio T, Lokki AI, Jääskeläinen T, Kortelainen E, Ruotsalainen S, et al. Genetic risk factors associated with preeclampsia and hypertensive disorders of pregnancy. JAMA Cardiol. (2023) 8:674–83. doi: 10.1001/jamacardio.2023.1312
97. Abu-El-Haija A, Reddi H V, Wand H, Rose NC, Mori M, Qian E, et al. The clinical application of polygenic risk scores: a points to consider statement of the American College of Medical Genetics and Genomics (ACMG). Genet Med Off J Am Coll Med Genet. (2023) 25:100803. doi: 10.1016/j.gim.2023.100803
98. Honigberg MC, Truong B, Khan RR, Xiao B, Bhatta L, Vy HMT, et al. Polygenic prediction of preeclampsia and gestational hypertension. Nat Med. (2023) 29:1540–9. doi: 10.1038/s41591-023-02374-9
99. Nurkkala J, Kauko A, FinnGen, Laivuori H, Saarela T, Tyrmi JS, et al. Associations of polygenic risk scores for preeclampsia and blood pressure with hypertensive disorders of pregnancy. J Hypertens. (2023) 41:380–7. doi: 10.1097/HJH.0000000000003336
100. Gong S, Gaccioli F, Dopierala J, Sovio U, Cook E, Volders P-J, et al. The RNA landscape of the human placenta in health and disease. Nat Commun. (2021) 12:2639. doi: 10.1038/s41467-021-22695-y
101. Enquobahrie DA, Meller M, Rice K, Psaty BM, Siscovick DS, Williams MA. Differential placental gene expression in preeclampsia. Am J Obstet Gynecol. (2008) 199:566.e1–11. doi: 10.1016/j.ajog.2008.04.020
102. Sitras V, Paulssen RH, Grønaas H, Leirvik J, Hanssen TA, Vårtun A, et al. Differential placental gene expression in severe preeclampsia. Placenta. (2009) 30:424–33. doi: 10.1016/j.placenta.2009.01.012
103. Tarca AL, Romero R, Erez O, Gudicha DW, Than NG, Benshalom-Tirosh N, et al. Maternal whole blood mRNA signatures identify women at risk of early preeclampsia: a longitudinal study. J Matern neonatal Med Off J Eur Assoc Perinat Med Fed Asia Ocean Perinat Soc Int Soc Perinat Obstet. (2021) 34:3463–74. doi: 10.1080/14767058.2019.1685964
104. Rasmussen M, Reddy M, Nolan R, Camunas-Soler J, Khodursky A, Scheller NM, et al. RNA profiles reveal signatures of future health and disease in pregnancy. Nature. (2022) 601:422–7. doi: 10.1038/s41586-021-04249-w
105. Moufarrej MN, Vorperian SK, Wong RJ, Campos AA, Quaintance CC, Sit RV, et al. Early prediction of preeclampsia in pregnancy with cell-free RNA. Nature. (2022) 602:689–94. doi: 10.1038/s41586-022-04410-z
106. Cho WCS. MicroRNAs as therapeutic targets and their potential applications in cancer therapy. Expert Opin Ther Targets. (2012) 16:747–59. doi: 10.1517/14728222.2012.696102
107. Krek A, Grün D, Poy MN, Wolf R, Rosenberg L, Epstein EJ, et al. Combinatorial microRNA target predictions. Nat Genet. (2005) 37:495–500. doi: 10.1038/ng1536
108. Fu G, Brkić J, Hayder H, Peng C. MicroRNAs in human placental development and pregnancy complications. Int J Mol Sci. (2013) 14:5519–44. doi: 10.3390/ijms14035519
109. Pineles BL, Romero R, Montenegro D, Tarca AL, Han YM, Kim YM, et al. Distinct subsets of microRNAs are expressed differentially in the human placentas of patients with preeclampsia. Am J Obstet Gynecol. (2007) 196:261.e1–e6. doi: 10.1016/j.ajog.2007.01.008
110. Miura K, Miura S, Yamasaki K, Higashijima A, Kinoshita A, Yoshiura KI, et al. Identification of pregnancy-associated microRNAs in maternal plasma. Clin Chem. (2010) 56:1767–71. doi: 10.1373/clinchem.2010.147660
111. Chim SSC, Shing TKF, Hung ECW, Leung TY, Lau TK, Chiu RWK, et al. Detection and characterization of placental microRNAs in maternal plasma. Clin Chem. (2008) 54:482–90. doi: 10.1373/clinchem.2007.097972
112. Skalis G, Katsi V, Miliou A, Georgiopoulos G, Papazachou O, Vamvakou G, et al. MicroRNAs in preeclampsia. MicroRNA. (2018) 8:28–35. doi: 10.2174/2211536607666180813123303
113. Stittrich A-B, Haftmann C, Sgouroudis E, Kühl AA, Hegazy AN, Panse I, et al. The microRNA miR-182 is induced by IL-2 and promotes clonal expansion of activated helper T lymphocytes. Nat Immunol. (2010) 11:1057–62. doi: 10.1038/ni.1945
114. Kim YM, Romero R, Oh SY, Kim CJ, Kilburn BA, Armant DR, et al. Toll-like receptor 4: a potential link between “danger signals,” the innate immune system, and preeclampsia? Am J Obstet Gynecol. (2005) 193:921–7. doi: 10.1016/j.ajog.2005.07.076
115. Dai Y, Qiu Z, Diao Z, Shen L, Xue P, Sun H, et al. MicroRNA-155 inhibits proliferation and migration of human extravillous trophoblast derived HTR-8/SVneo cells via down-regulating cyclin D1. Placenta. (2012) 33:824–9. doi: 10.1016/j.placenta.2012.07.012
116. Jairajpuri DS, Malalla ZH, Mahmood N, Almawi WY. Circulating microRNA expression as predictor of preeclampsia and its severity. Gene. (2017) 627:543–8. doi: 10.1016/j.gene.2017.07.010
117. Akgör U, Ayaz L, Çayan F. Expression levels of maternal plasma microRNAs in preeclamptic pregnancies. J Obstet Gynaecol. (2021) 41:910–4. doi: 10.1080/01443615.2020.1820465
118. Wu L, Zhou H, Lin H, Qi J, Zhu C, Gao Z, et al. Circulating microRNAs are elevated in plasma from severe preeclamptic pregnancies. Reproduction. (2012) 143:389–97. doi: 10.1530/REP-11-0304
119. Li P, Guo W, Du L, Zhao J, Wang Y, Liu L, et al. MicroRNA-29b contributes to pre-eclampsia through its effects on apoptosis, invasion and angiogenesis of trophoblast cells. Clin Sci. (2013) 124:27–40. doi: 10.1042/CS20120121
120. Carbone IF, Conforti A, Picarelli S, Morano D, Alviggi C, Farina A. Circulating nucleic acids in maternal plasma and serum in pregnancy complications: are they really useful in clinical practice? A Systematic Review. Mol Diagnosis Ther. (2020) 24:409–31. doi: 10.1007/s40291-020-00468-5
121. Starodubtseva N, Poluektova A, Tokareva A, Kukaev E, Avdeeva A, Rimskaya E, et al. Proteome-based maternal plasma and serum biomarkers for preeclampsia: a systematic review and meta-analysis. Life. (2025) 15:776. doi: 10.3390/life15050776
122. Navajas R, Corrales F, Paradela A. Quantitative proteomics-based analyses performed on pre-eclampsia samples in the 2004–2020 period: a systematic review. Clin Proteomics. (2021) 18:6. doi: 10.1186/s12014-021-09313-1
123. Watanabe H, Hamada H, Yamada N, Sohda S, Yamakawa-Kobayashi K, Yoshikawa H, et al. Proteome analysis reveals elevated serum levels of clusterin in patients with preeclampsia. Proteomics. (2004) 4:537–43. doi: 10.1002/pmic.200300565
124. Blumenstein M, McMaster MT, Black MA, Wu S, Prakash R, Cooney J, et al. A proteomic approach identifies early pregnancy biomarkers for preeclampsia: novel linkages between a predisposition to preeclampsia and cardiovascular disease. Proteomics. (2009) 9:2929–45. doi: 10.1002/pmic.200800625
125. Carty DM, Siwy J, Brennand JE, Zürbig P, Mullen W, Franke J, et al. Urinary proteomics for prediction of preeclampsia. Hypertens (Dallas, Tex 1979). (2011) 57:561–9. doi: 10.1161/HYPERTENSIONAHA.110.164285
126. Nguyen TPH, Patrick CJ, Parry LJ, Familari M. Using proteomics to advance the search for potential biomarkers for preeclampsia: a systematic review and meta-analysis. PLoS ONE. (2019) 14:e0214671. doi: 10.1371/journal.pone.0214671
127. Blumenstein M, Prakash R, Cooper GJS, North RA. Aberrant processing of plasma vitronectin and high- molecular-weight kininogen precedes the onset of preeclampsia. Reprod Sci. (2009) 16:1144–52. doi: 10.1177/1933719109342756
128. Blankley RT, Gaskell SJ, Whetton AD, Dive C, Baker PN, Myers JE, et al. Proof-of-principle gel-free proteomics strategy for the identification of predictive biomarkers for the onset of pre-eclampsia. BJOG. (2009) 116:1473–80. doi: 10.1111/j.1471-0528.2009.02283.x
129. Park J, Cha DH, Lee SJ, Kim YN, Kim YH, Kim KP. Discovery of the serum biomarker proteins in severe preeclampsia by proteomic analysis. Exp Mol Med. (2011) 43:427–35. doi: 10.3858/emm.2011.43.7.047
130. Rasanen J, Girsen A, Lu X, Lapidus JA, Standley M, Reddy A, et al. Comprehensive maternal serum proteomic profiles of preclinical and clinical preeclampsia. J Proteome Res. (2010) 9:4274–81. doi: 10.1021/pr100198m
131. Wen Q, Liu LY, Yang T, Alev C, Wu S, Stevenson DK, et al. Peptidomic identification of serum peptides diagnosing preeclampsia. PLoS ONE. (2013) 8:e65571. doi: 10.1371/journal.pone.0065571
132. Centlow M, Hansson SR, Welinder C. Differential proteome analysis of the preeclamptic placenta using optimized protein extraction. J Biomed Biotechnol. (2010) 2010:458748. doi: 10.1155/2010/458748
133. Liu LY, Yang T, Ji J, Wen Q, Morgan AA, Jin B, et al. Integrating multiple “omics” analyses identifies serological protein biomarkers for preeclampsia. BMC Med. (2013) 11:236. doi: 10.1186/1741-7015-11-236
134. Pecks U, Schütt A, Röwer C, Reimer T, Schmidt M, Preschany S, et al. A mass spectrometric multicenter study supports classification of preeclampsia as heterogeneous disorder. Hypertens Pregnancy. (2012) 31:278–91. doi: 10.3109/10641955.2011.640375
135. Tarca AL, Romero R, Benshalom-Tirosh N, Than NG, Gudicha DW, Done B, et al. The prediction of early preeclampsia: results from a longitudinal proteomics study. PLoS ONE. (2019) 14:e0217273. doi: 10.1371/journal.pone.0217273
136. Erez O, Romero R, Maymon E, Chaemsaithong P, Done B, Pacora P, et al. The prediction of late-onset preeclampsia: results from a longitudinal proteomics study. PLoS ONE. (2017) 12:e0181468. doi: 10.1371/journal.pone.0181468
137. Than NG, Romero R, Györffy D, Posta M, Bhatti G, Done B, et al. Molecular subclasses of preeclampsia characterized by a longitudinal maternal proteomics study: distinct biomarkers, disease pathways and options for prevention. J Perinat Med. (2023) 51:51–68. doi: 10.1515/jpm-2022-0433
138. Ghaemi MS, Tarca AL, Romero R, Stanley N, Fallahzadeh R, Tanada A, et al. Proteomic signatures predict preeclampsia in individual cohorts but not across cohorts - implications for clinical biomarker studies. J Matern Fetal Neonatal Med Off J Eur Assoc Perinat Med Fed Asia Ocean Perinat Soc Int Soc Perinat Obstet. (2022) 35:5621–8. doi: 10.1080/14767058.2021.1888915
139. Gh BFNM. Application of metabolomics to preeclampsia diagnosis. Syst Biol Reprod Med. (2018) 64:324–39. doi: 10.1080/19396368.2018.1482968
140. Yao M, Xiao Y, Yang Z, Ge W, Liang F, Teng H, et al. Identification of biomarkers for preeclampsia based on metabolomics. Clin Epidemiol. (2022) 14:337–60. doi: 10.2147/CLEP.S353019
141. Johnson CH, Ivanisevic J, Siuzdak G. Metabolomics: beyond biomarkers and towards mechanisms. Nat Rev Mol Cell Biol. (2016) 17:451–9. doi: 10.1038/nrm.2016.25
142. Lee SM, Kang Y, Lee EM, Jung YM, Hong S, Park SJ, et al. Metabolomic biomarkers in midtrimester maternal plasma can accurately predict the development of preeclampsia. Sci Rep. (2020) 10:16142. doi: 10.1038/s41598-020-72852-4
143. Jain S, Jayasimhulu K, Clark JF. Metabolomic analysis of molecular species of phospholipids from normotensive and preeclamptic human placenta electrospray ionization mass spectrometry. Front Biosci. (2004) 9:3167–75. doi: 10.2741/1470
144. Turner E, Brewster JA, Simpson NAB, Walker JJ, Fisher J. Plasma from women with preeclampsia has a low lipid and ketone body content–a nuclear magnetic resonance study. Hypertens Pregnancy. (2007) 26:329–42. doi: 10.1080/10641950701436073
145. Kenny LC, Broadhurst D, Brown M, Dunn WB, Redman CWG, Kell DB, et al. Detection and identification of novel metabolomic biomarkers in preeclampsia. Reprod Sci. (2008) 15:591–7. doi: 10.1177/1933719108316908
146. Kharb S, Joshi A. Multi-omics and machine learning for the prevention and management of female reproductive health. Front Endocrinol. (2023) 14:1081667. doi: 10.3389/fendo.2023.1081667
147. Zhang L, Bi S, Liang Y, Huang L, Li Y, Huang M, et al. Integrated metabolomic and lipidomic analysis in the placenta of preeclampsia. Front Physiol. (2022) 13:807583. doi: 10.3389/fphys.2022.807583
148. Marić I, Tsur A, Aghaeepour N, Montanari A, Stevenson DK, Shaw GM, et al. Early prediction of preeclampsia via machine learning. Am J Obstet Gynecol MFM. (2020) 2:1–17. doi: 10.1016/j.ajogmf.2020.100100
149. Hackelöer M, Schmidt L, Verlohren S. New advances in prediction and surveillance of preeclampsia: role of machine learning approaches and remote monitoring. Arch Gynecol Obstet. (2023) 308:1663–77. doi: 10.1007/s00404-022-06864-y
150. Ranjbar A, Montazeri F, Ghamsari SR, Mehrnoush V, Roozbeh N, Darsareh F. Machine learning models for predicting preeclampsia: a systematic review. BMC Pregnancy Childbirth. (2024) 24:6. doi: 10.1186/s12884-023-06220-1
151. Espinosa C, Becker M, Marić I, Wong RJ, Shaw GM, Gaudilliere B, et al. Data-driven modeling of pregnancy-related complications. Trends Mol Med. (2021) 27:762–76. doi: 10.1016/j.molmed.2021.01.007
152. Jhee JH, Lee S, Park Y, Lee SE, Kim YA, Kang S-W, et al. Prediction model development of late-onset preeclampsia using machine learning-based methods. PLoS ONE. (2019) 14:e0221202. doi: 10.1371/journal.pone.0221202
153. Sandström A, Snowden JM, Höijer J, Bottai M, Wikström AK. Clinical risk assessment in early pregnancy for preeclampsia in nulliparous women: a population based cohort study. PLoS ONE. (2019) 14:1–16. doi: 10.1371/journal.pone.0225716
154. Marić I, Contrepois K, Moufarrej MN, Stelzer IA, Feyaerts D, Han X, et al. Early prediction and longitudinal modeling of preeclampsia from multiomics. Patterns. (2022) 3:100655. doi: 10.1016/j.patter.2022.100655
155. Li T, Xu M, Wang Y, Wang Y, Tang H, Duan H, et al. Prediction model of preeclampsia using machine learning based methods: a population based cohort study in China. Front Endocrinol. (2024) 15:1–14. doi: 10.3389/fendo.2024.1345573
156. Kovacheva VP, Eberhard BW, Cohen RY, Maher M, Saxena R, Gray KJ. Preeclampsia prediction using machine learning and polygenic risk scores from clinical and genetic risk factors in early and late pregnancies. Hypertens (Dallas, Tex 1979). (2024) 81:264–72. doi: 10.1161/HYPERTENSIONAHA.123.21053
157. Gil MM, Cuenca-Gómez D, Rolle V, Pertegal M, Díaz C, Revello R, et al. Validation of machine-learning model for first-trimester prediction of pre-eclampsia using cohort from PREVAL study. Ultrasound Obstet Gynecol. (2024) 63:68–74. doi: 10.1002/uog.27478
158. Montgomery-Csobán T, Kavanagh K, Murray P, Robertson C, Barry SJE, Vivian Ukah U, et al. Machine learning-enabled maternal risk assessment for women with pre-eclampsia (the PIERS-ML model): a modelling study. Lancet Digit Heal. (2024) 6:e238–50. doi: 10.1016/S2589-7500(23)00267-4
159. Ali MR, Lawson CA, Wood AM, Khunti K. Addressing ethnic and global health inequalities in the era of artificial intelligence healthcare models: a call for responsible implementation. J R Soc Med. (2023) 116:260–2. doi: 10.1177/01410768231187734
160. Gao Y, Sharma T, Cui Y. Addressing the challenge of biomedical data inequality: an artificial intelligence perspective. Annu Rev Biomed data Sci. (2023) 6:153–71. doi: 10.1146/annurev-biodatasci-020722-020704
161. Kumar M, Nguyen TPN, Kaur J, Singh TG, Soni D, Singh R, et al. Opportunities and challenges in application of artificial intelligence in pharmacology. Pharmacol Rep. (2023) 75:3–18. doi: 10.1007/s43440-022-00445-1
162. Kusanovic JP, Romero R, Chaiworapongsa T, Erez O, Mittal P, Vaisbuch E, et al. A prospective cohort study of the value of maternal plasma concentrations of angiogenic and anti-angiogenic factors in early pregnancy and midtrimester in the identification of patients destined to develop preeclampsia. J Matern Fetal Neonatal Med. (2009) 22:1021–38. doi: 10.3109/14767050902994754
Keywords: preeclampsia, biomarkers, omics technologies, hypertensive disorders, history, artificial intelligence
Citation: Riano-Moreno JC, Vargas-Castellanos E, Pedraza A, Díaz-Quiñonez LS and Rangel-Ramos VS (2025) Preeclampsia prediction and diagnosis: a comprehensive historical review from clinical insights to omics perspectives. Front. Med. 12:1689745. doi: 10.3389/fmed.2025.1689745
Received: 21 August 2025; Accepted: 30 September 2025;
Published: 23 October 2025.
Edited by:
Andrea Cruz, International Iberian Nanotechnology Laboratory (INL), PortugalReviewed by:
Iane Oliveira Pires Porto, University of Rio Verde, BrazilLokanatha Oruganti, Tulane University, United States
Copyright © 2025 Riano-Moreno, Vargas-Castellanos, Pedraza, Díaz-Quiñonez and Rangel-Ramos. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Elizabeth Vargas-Castellanos, ZWxpemFiZXRoLnZhcmdhc0BtZWRlcmkuY29tLmNv; Víctor Saul Rangel-Ramos, dmljdG9yLnJhbmdlbEBtZWRlcmkuY29tLmNv
†These authors have contributed equally to this work