 Daniela Cesselli1
Daniela Cesselli1 Pietro Parisse2
Pietro Parisse2 Aneta Aleksova3
Aneta Aleksova3 Claudia Veneziano1Celeste Cervellin1
Claudia Veneziano1Celeste Cervellin1 Andrea Zanello1
Andrea Zanello1 Antonio Paolo Beltrami1*
Antonio Paolo Beltrami1*- 1Department of Medicine, University of Udine, Udine, Italy
- 2Elettra-Sincrotrone Trieste S.C.p.A., Trieste, Italy
- 3Cardiovascular Department, Azienda Sanitaria Universitaria Integrata di Trieste – University of Trieste, Trieste, Italy
Extracellular vesicles (EV) are at the center of an intense activity of investigation, both for their possible employment as biomarkers of ongoing pathologic processes and for their broad range of biological activities. EV can promote tissue repair in very different pathologic settings, including hindlimb and myocardial ischemia. Importantly, the exact mode of action of EV is still partly understood, since they may act by modulating growth factors and cytokines, signaling pathways, and by transferring non-coding RNAs to target cells. However, the term EV identifies cell derived, enveloped particles very heterogeneous in size, composition, and biogenesis. Therefore, part of the controversies on the biological effects exerted by EV is a consequence of differences in methods of separation that result in the enrichment of different entities. Since technical challenges still hamper the highly specific sorting of different EV subpopulations, up to now only few investigators have tried to verify differences in the biological effects of specific EV subtypes. This review summarizes the current state of the art on the comprehension of mechanisms involved in EV biogenesis and release, which is a prerequisite for understanding and investigating the impact that pathology and drug therapy may exert on the secretion and composition of EV. Finally, we described both the mechanism involved in the modulation of EV secretion by drugs commonly used in patients affected by heart failure, and how pathophysiological mechanisms involved in heart disease modify EV secretion.
Introduction
Extracellular vesicles (EV) are gaining momentum as potential biomarkers and for their broad range of biological activities. Indeed, a large body of literature, generated by independent laboratories, has shown, over the last few years, that EV can promote tissue repair in very different pathologic settings, ranging from cutaneous wound healing and bone regeneration to hindlimb and myocardial ischemia. Consistently, it is now generally accepted that stem cells and cell based therapies act, at least in part, through the release of EV (see Chen et al., 2017). However, the exact mode of action of EV is still object of intense investigation. Indeed, EV may act by modulating growth factors and cytokines (e.g., IGF, SDF1, NGF, and HGF), signaling pathways (e.g., Akt, ERK, Wnt/β-catenin, Hippo/YAP, and STAT3), and miRNA (e.g., miR-21, -23a, -125b, and -145) (Shabbir et al., 2015; Zhang et al., 2016; Fatima et al., 2017). Moreover, the coordinated action of these EV components has been shown to regulate stem cell proliferation and differentiation, stimulate angiogenesis, regulate epithelial to mesenchymal transition, reduce fibrosis and apoptosis (reviewed in Fatima et al., 2017).
Extracellular vesicles play a relevant role in the regulation of immunity, where they promote both immune stimulation and suppression (see Robbins and Morelli, 2014 for review), in a broad range of physiological and pathological conditions. With regard to immune stimulation, EV can both trigger T cell activation, acting directly as antigen-presenting vesicles, and elicit an indirect, dendritic cell (DC)-mediated, effect (Nolte-’t Hoen et al., 2009; Robbins and Morelli, 2014). Conversely, the mechanisms of the immunosuppressive function of EV, which have been experimentally exploited in the cardiac transplantation field to reduce cardiac allograft rejection, are still under scrutiny (Peche et al., 2003; Robbins and Morelli, 2014).
Given their complex mechanism of action and broad effects, EV are also involved in the pathogenesis of distinct pathological conditions, including inflammatory, autoimmune, and infectious diseases (Robbins and Morelli, 2014) as well as neurodegenerative or prion-related diseases, where they may enhance the spreading of toxic proteins or aggregates through the brain (Guo et al., 2016; Xiao T. et al., 2017).
Nonetheless, the term EV is a very general one that embraces cell derived, enveloped particles very heterogeneous in size, composition, and biogenesis. Therefore, part of the controversies on the biological effects exerted by EV is a consequence of differences in methods of separation that result in the enrichment of different entities. Specifically, EV have been divided into broad categories, mainly depending on their size and biogenesis (i.e., microvesicles (MVs), apoptotic bodies, and exosomes). While the first two bud from the plasma membrane, exosomes derive from multivesicular bodies (MVB) (Maas et al., 2017). Importantly, although both MVs and exosomes can transfer their cargo to recipient cells, influencing their behavior (Antonyak and Cerione, 2015), a large body of recent literature is focusing on exosomes, the smallest EV. However, only few investigators have tried to verify differences in the biological effects of these subtypes, possibly as a consequence of the difficulties in sorting with high specificity the different EV populations (Kanada et al., 2015).
Aim of this review is to discuss the impact that drugs and comorbidities exert on the biogenesis and function of EV, with particular reference to the cardiovascular system.
Exosomes and Microvesicles: What Are Them?
Although cell derived vesicles have been described in the mid 20th century, their biological function was initially only marginally understood, and they were considered to be mostly “cell dust” or a system of “trash disposal” (Laberge et al., 2018). It was only recently that the role played by EV as a primary mechanism of intercellular communication has emerged, mainly for their ability to transfer macromolecules, including proteins and RNA, among cells (Valadi et al., 2007; Al-Nedawi et al., 2009).
As anticipated, the term EV comprises different entities that differ in size, composition, and biogenesis. Specifically, three broad categories have been identified: exosomes, that are ≈30–100 nm in size and originate intracellularly from the MVB; MVs (also named ectosomes or microparticles), whose size ranges from ≈200 to 1,000 nm and arise via a direct budding of the plasma membrane; and apoptotic bodies, that are 1,000–5,000 nm in size and are generated as a consequence of programmed cell death. In recent years, it has become apparent that the picture is more complex than previously thought and that the overlap in size distribution among different EV types impairs our ability to separate them merely on physical basis (Tkach et al., 2018). Indeed, according to opinion leaders, the most popular methods of exosome purification (i.e., ultracentrifugation, 200 nm filtration, and precipitation) actually co-isolate different types of EV (Tkach and Thery, 2016).
Exosome Biogenesis and Sorting of Exosomal Cargo
Concerning the biogenesis of EV, it involves, for every subtype, the negative curvature of cellular membranes (i.e., bending the membrane away from the cytoplasm). While exosomes are first released within the MVB as intraluminal vesicles (ILV), larger EV are directly pinched off the plasma membrane. The negative curvature of the cellular membranes can be initiated by different mechanisms, that are divided into endosomal sorting complex required for transport (ESCRT)-dependent and ESCRT-independent.
Endosomal Sorting Complex Required for Transport, ESCRT
In the early 2000’s, a molecular machinery responsible for the negative bending of cell membranes (i.e., ESCRT) was identified. It comprises about 30 proteins, assembled in five functional units that are required for: cargo recognition (ESCRT-0), its sequestration into endosomal microdomains (ESCRT-I and II), the constriction of the neck of the nascent vesicle (ESCRT-III), and finally, membrane scission and ESCRT disassembly (promoted by the Vps4 complex) (Furthauer, 2018). A further support to membrane budding is provided by the interaction of syndecans and sintenin with Alix, an auxiliary component of the ESCRT complex (Baietti et al., 2012). In this pathway, that is particularly relevant for the regulation of the turnover of membrane proteins, ubiquitination of transmembrane proteins is required for their recognition and sorting by ESCRT-0, -I, and -II complexes (Urbanelli et al., 2013; Colombo et al., 2014).
However, other sorting mechanisms are in place too. Specifically, cytosolic proteins can be recruited to late endosomes by a microautophagy-like mechanism, which is distinct from chaperone-mediated autophagy, and requires hsc70, and ESCRT-I and -III (Sahu et al., 2011). Importantly, several molecules, including the pigment cell-specific type I integral membrane protein PMEL and CD63 in melanocytes (van Niel et al., 2011), and the class II major histocompatibility complex (MHC) in DCs (Buschow et al., 2009), can be inserted into ILV following ESCRT independent pathways.
Lipid Dependent Mechanisms
Ceramide production induced by neutral sphingomyelinase 2 (nSMase 2) is an important ESCRT-independent mechanism involved in ILV formation, possibly as a consequence of the ability of members of this family of lipids to cause membrane curvature (Trajkovic et al., 2008). However, the hypothesis that ceramide acts only by bending the membrane may be too simplistic (Kajimoto et al., 2013). Indeed, ceramide is at the center of the sphingolipid metabolism, where it acts as either a product or a precursor for several metabolites, including sphingosine-1-phosphate (S1P). Many of these metabolites have complex biological effects that are usually referred to as “sphingolipid rheostat” (see discussion later) (Newton et al., 2015). Specifically, it has been recently shown that the continuous activation of the inhibitory G protein (Gi) coupled S1P receptors on MVB is required for cargo sorting into ILV destined to the exosomal pathway. Conversely, inhibition of S1P production reduces CD63, CD81, and flotillin in ILV (Kajimoto et al., 2013). Intriguingly, a reciprocal role for ceramide and S1P on autophagy induction and mTOR pathway modulation has been described (Taniguchi et al., 2012). This finding is consistent with the concept that exosome secretion and the autophagy lysosome pathway are coordinated mechanisms and is reinforced by the observation that lysosome inhibition increases EV release (Baixauli et al., 2014; Miao et al., 2015). How these two different fates are regulated is still an area of investigation. Indeed, MVB are not a homogenous population, and it has been suggested that different subpopulations of MVB exist, with some (e.g., those rich in cholesterol) that are targeted for exocytosis.
Another lipid-dependent mechanism that is involved in the biogenesis of exosomes relies on the activity of phospholipase D (PLD). Upon stimulation with ionomycin, this enzyme is relocated from the plasma membrane to intracellular compartments and its activity correlates with the extent of exosome secretion (Laulagnier et al., 2004).
Furthermore, both PLD and phospholipase C (PLC) are involved in the production of diacylglycerol (DAG), which regulates several checkpoints of the vesicle secretory pathway (Alonso et al., 2011; Mazzeo et al., 2016). Diacylglycerol kinase (DGK) can convert DAG into phosphatidic acid, which can induce a negative membrane curvature and is a precursor of bis(diacylglycero)phosphate (BDP) (Kooijman et al., 2003). The latter or other substrates could be converted into Lysobisphosphatidic acid (LBPA) or bis(monoacylglycero)phosphate (BMP) by phospholipase A2 (PLA2) (Amidon et al., 1996). BMP, which increases its concentration in late endosomes/MVB (Subra et al., 2007), promotes the formation of ILV-like structures at low pH, interacting with Alix (Matsuo et al., 2004). Finally, BMP binds the chaperone Hsp70 (Kirkegaard et al., 2010) and seems to be critically involved in regulating the levels of cholesterol transported in endosomes (Chevallier et al., 2008).
Tetraspanins
An additional, ESCRT-independent mechanism of ILV formation is provided by proteins of the tetraspanin family (that include CD9, CD63, CD81, CD82, and CD151). An increasing amount of evidence supports this notion, as suggested by both: the ability of cells in which the ESCRT has been silenced to release CD63 positive EV, and the reduced ability of DCs of CD9 knockout mice to promote exosome secretion (Stuffers et al., 2009; Chairoungdua et al., 2010; Andreu and Yanez-Mo, 2014). Tetraspanins are palmytoilated proteins characterized by four transmembrane domains and the ability to organize membrane domains (named tetraspanins-enriched domains, TEM), clustering and interacting with a large number of distinct transmembrane and cytosolic proteins, cholesterol and gangliosides (Andreu and Yanez-Mo, 2014). They can regulate membrane shape (Grove, 2014), mostly promoting (or inhibiting, as in the case of CD82) membrane bending and regulating actin polymerization (Bari et al., 2011). Additionally, tetraspanins have been involved in sorting the intracellular cargo toward EV. Specifically, β-catenin, Wnt11, and PMEL, as well as matrix metalloproteases, such as CD10, and MHC-I and -II molecules have been shown to be sent to EV in a tetraspanin dependent fashion (Andreu and Yanez-Mo, 2014). Furthermore, by studying the interactome of EV pulled down employing antibodies against the C-terminal regions of both tetraspanins (CD9, CD81, and CD151) and tetraspanin-associated immunoglobulin superfamily receptors (ICAM-1, VCAM-1, and EWI-2), it was shown that ≈45% of the proteins annotated as exosomal indeed were part of the interactome of CD81 and/or EWI-2 (Perez-Hernandez et al., 2013).
RNA Sorting Mechanisms
A special mention deserves the mechanism that regulate RNA sorting into EV. Indeed, several studies confirmed that the RNA content of parental cells is significantly different from that of cell-derived vesicles, pointing to a selective sorting of target RNA species to the EV (Ratajczak et al., 2006; Skog et al., 2008; Nolte-’t Hoen et al., 2012). Deep sequencing experiments performed on DC derived EV have shown that the majority of extracellular RNA consists of small (<200 nucleotides) RNAs. Most of these are non-coding RNAs, distinct from microRNA and large intervening non-coding RNAs, and represent transcripts or cleavage products overlapping with protein coding regions, repeat sequences, or structural RNAs. It has been suggested that some of these could play a role in the specific sorting of regulatory RNAs into the EV (Nolte-’t Hoen et al., 2012). A broader picture was obtained by analyzing the extracellular RNA released by glioblastoma stem cells. In this setting, authors have shown that MV RNA content is more similar to that of the parent cell, while the exosomal one is more distinct (Wei et al., 2017). With regard to miRNA, bioinformatics and mutagenesis analyses have demonstrated that EXOmotifs regulate both their binding to hnRNPA2B1 and their loading into exosomes, upon sumoylation (Villarroya-Beltri et al., 2013). Additionally, Annexin-2, a protein that is found with high abundance in exosomes, binds miRNA in a Ca2+ dependent fashion. The role played by this protein in sorting miRNA into exosomes is provided by silencing experiments that showed a decrease in the amount of miRNA loaded into exosomes, without modifying their expression profile (Hagiwara et al., 2015). It has been additionally suggested that selective sorting of RNAs to EV could depend on their affinity to ceramide and sphingosine rich raft like structures present in MVB (Kosaka et al., 2010; Janas et al., 2012, 2015). This property may be either mediated by binding of target RNA to hnRNPA2B1 or consequent to hydrophobicity given directly by specific RNA sequences or by enzymatic modifications, such as miRNA methylation or tRNA isoprenylation (Janas et al., 2012, 2015).
Microvesicle Biogenesis
Somewhat less understood and investigated are the mechanisms that regulate plasma membrane budding and MV biogenesis. While, as for exosomes, the ESCRT complex is involved in the biogenesis of MV, MV production and release can also be induced by calcium signaling and ATP and lipid mediated mechanisms. Exosomes and MVs differ also in the mechanisms involved in protein and RNA sorting.
ESCRT
Although the biogenesis of exosomes and that of MVs are distinct processes, they both share part of the same molecular mechanisms. Indeed, one component of ESCRT-I (i.e., TSG101), that is hijacked by HIV to promote the release of viral particles, can be recruited to the plasma membrane by arrestin domain-containing protein 1 (ARRDC1) to promote, together with VPS4, MV release (Nabhan et al., 2012). Importantly, these ARRDC1-mediated MVs lack the endosomal markers CD63 and LAMP1 (Nabhan et al., 2012).
Additionally, it has been shown that ESCRT components can be found in the plasma membrane too, where they can form oligomeric complexes (Welsch et al., 2006). However, the ability of these molecules to promote the formation of MVs with the same mechanism that is observed in ILV biogenesis is uncertain (Yang and Gould, 2013).
Calcium Mediated Processes
Increased intracytoplasmic calcium levels, either employed as second messenger of different signal transduction pathways or occurring as a consequence of cell apoptosis and necrosis, can activate calcium sensitive proteins involved in MV biogenesis. Specifically, gelsolin, a ubiquitous actin binding protein that can remodel the actin cytoskeleton (Li et al., 2012) and calpain, a family of cysteine protease that modulate the function of their substrates by performing “limited proteolysis” (Ono et al., 2016), can act in synergy to promote cytoskeleton detachment and membrane blebbing (Miyoshi et al., 1996). Moreover, the activities of enzymes involved in regulating plasma membrane asymmetry, such as aminophospholipid translocase (that transports phosphatidylserine and phosphatidylethanolamine from the outer layer to the inner layer) and lipid scramblase (that promotes the movement of lipids across the cell membrane) are modulated in opposite fashions by calcium, resulting in loss of plasma membrane asymmetry and exposure of phosphatidylserine (Piccin et al., 2007). Furthermore, Annexin-2, a protein involved in calcium induced exocytosis has been also shown to promote MV release (Zhang W. et al., 2013).
ATP and Lipid Mediated Mechanisms
Upon stimulation of P2X7 purinergic receptors by ATP released in the extracellular space, the acid sphingomyelinase is rapidly activated and translocated to the outer layer of the plasma membrane, where it cleaves sphingomyelin, increasing the efflux of cholesterol and membrane fluidity, thus facilitating membrane shedding (Bianco et al., 2009).
Membrane Targeting and Protein Crowding
Protein sorting to nascent EV is thought to depend on the presence of anchors [i.e., protein myristoylation, phosphatidylinositol-4,5-bisphosphate (PIP2)-binding domains, phosphatidylinositol-(3,4,5)-trisphosphate-binding domains] that promote their association with the plasma membrane compartment (Shen et al., 2011). However, a second signal provided by the higher-order oligomerization of surface proteins seems to be required for their secretion within MV. Intriguingly, the presence of an endosomal targeting domain was not efficient in promoting protein secretion via EV, in this experimental model (Shen et al., 2011). A corollary to this observation is that membrane targeted proteins that form higher-order oligomeric complexes can be secreted into EV. Indeed, tetraspanins, that can be found as TEM also in the plasma membrane, can interact with the cytoskeleton via ezrin/radixin/moesin proteins, and can be thus released into MVs (Sala-Valdes et al., 2006; Yang and Gould, 2013; Andreu and Yanez-Mo, 2014).
Last, protein lateral confining and crowding mechanisms can bend membranes and may provide a biophysical explanation for EV formation (Stachowiak et al., 2012; Derganc and Copic, 2016).
RNA Sorting Mechanisms
As opposed to what was shown for exosomes, results obtained with next-generation sequencing suggest that the selection of the cargo may be, in this case, less stringent. Indeed, the RNA content of MV more closely resembles the parental cell one (Wei et al., 2017).
Secretion of Exosome and MV
A relevant question in EV biology is whether their secretion is constitutive or regulated. By understanding the mechanisms that regulate the secretion of EV, we could gain better insights into how ischemia and pharmacological therapies may impact on their release.
Regulation of Exosome Release
Once formed, ILV are released into the extracellular space, following the fusion of MVB or endosomal membranes with the plasma membrane, with a process that is regulated by the same mechanisms that control the trafficking of carrier vesicles. Specifically, transport, docking and fusion of vesicles to the plasma membrane are governed by the coordinated action of the cytoskeleton, of small GTPases of the Rab family, tethering factors, and soluble N-ethylmaleimide-sensitive factor attachment proteins -SNAP- and their receptors -SNARE-. More than 60 Rab proteins are encoded by the human genome. Each family member has a peculiar subcellular localization and is involved in distinct intracellular transport steps (Wandinger-Ness and Zerial, 2014). Of these, only Rab11, Rab27, Rab35, and possibly Rab22a, are validated regulators of exosome release (Colombo et al., 2014; Wang T. et al., 2014). Intriguingly, since Rab35, Rab11, and Rab27 are associated with early, recycling, and late endosomes, respectively, it has been hypothesized that they regulate the secretion of different subsets of exosomes. Differing in their origin, these subsets of exosomes are expected to express distinct markers, reflecting their sub cellular source (Colombo et al., 2014). Noteworthy, Rab proteins not directly involved in the exosome secretory pathways can be found within the vesicles, potentially reflecting a mechanism for the regulation of the correct intracellular levels and sub cellular localization of specific isoforms (Blanc and Vidal, 2017). With regard to the mechanisms regulating membrane fusion, SNAP and SNARE proteins, such as YKT6, VAMP7, VAMP3, and syntaxin-1a have been shown to be crucial for this process (Fader et al., 2009; Gross et al., 2012; Koles et al., 2012).
Exosomes can be released both in a constitutive and in a regulated fashion. With regard to upstream regulators of exosome secretion, it has been shown that the interaction between DCs and antigen specific CD4+ T lymphocytes can stimulate exosome release. Other triggers are represented by increased intracellular calcium levels, depolarization or neurotransmitter stimulation of neurons and glial cells (Colombo et al., 2014). Moreover, stressors able to induce p-53 can promote exosome secretion by upregulating the expression of its target gene TSAP6 (alias STEAP3), a multiple transmembrane protein involved both in iron homeostasis and Toll Like Receptor 4 mediated signaling (Lespagnol et al., 2008; Zhang et al., 2012).
Although considered to be a primary regulator of MV release, ADP ribosylation factor 6 (ARF6) promotes exosome secretion via its effector PLD2, an enzyme that regulates the cellular levels of phosphatidic acid, interacting with syntenin and Alix (Ghossoub et al., 2014). In line, DGK too, that metabolizes DAG to phosphatidic acid, promotes exosome secretion, in T cells (Alonso et al., 2011).
Other mechanisms of modulation of exosome secretion that have been described more recently are mediated by purinergic receptors, heparanase and ISGylation. The first ones (specifically P2X7 or P2Y12), are stimulated by extracellular ATP, and have been recently recognized as a relevant stimulus for exosome release (Drago et al., 2017). The second one is an enzyme that promotes angiogenesis and metastatic spread in many cancers, and can regulate the syntenin/Alix pathway by tailoring syndecans (Roucourt et al., 2015). ISGylation, instead, describes a mechanism of post-translational protein modification that leads to the covalent binding of target proteins to ISG15, an interferon induced ubiquitin-like protein. Recent data indicate that Interferon-I may reduce exosome secretion by promoting the ISGylation and degradation of TSG101 (Villarroya-Beltri et al., 2016).
Last, hypoxia is a prominent inducer of exosome release via HIF1α/HIF2α, both in the cancer and non-neoplastic settings (King et al., 2012; Kucharzewska et al., 2013; Zhang W. et al., 2017; Panigrahi et al., 2018).
Regulation of MV Release
While exosomes have been studied mainly in unstimulated cells, MV have been mostly analyzed following the application of specific stimuli (Colombo et al., 2014). However, recent data indicate that both mechanisms of constitutive and stimulated MV release exist, although the composition of MV released in these two different conditions can change (Drago et al., 2017).
Platelet derived MV (pMV) deserve a special mention, since they are the most abundant MV found in the circulation and exert procoagulant and proinflammatory functions (Badimon et al., 2016). Formation of pMV can be induced by a large number of stimuli, some of which shared with other cell types. Exercise training, heat stress, shear stress, cyclic strain, cardiac stress have all been shown to elicit pMV and MV release (Miyazaki et al., 1996; Augustine et al., 2014; Letsiou et al., 2015; Wilhelm et al., 2017). Moreover, different agonists have the ability to promote the release of MV. Among these, we should mention ATP or ADP stimulation, possibly in the presence of lipopolysaccharide priming, acting on either P2X7 or P2Y12 purinergic receptors, can lead to a raise in the intracellular Ca2+ levels, promoting cytoskeleton remodeling and MV release (Kahner et al., 2008; Bianco et al., 2009; Gulinelli et al., 2012; Colombo et al., 2014; Takenouchi et al., 2015; Drago et al., 2017). Furthermore, other proinflammatory (e.g., TNFα) and procoagulant factors (e.g., thrombin), together with reactive oxygen species can all induce MV release (Sapet et al., 2006; Szotowski et al., 2007). Last, hypoxia, via HIF1α- and HIF2α-induced Rab22a expression, has been associated with MV secretion (Wang T. et al., 2014). However, many variables can influence the response to the same stimulus, such as the cell type, the subject gender, age, and comorbidities.
Impact of Drugs and Diseases on Exosome Release
As anticipated, drugs and comorbidities can influence the mechanisms of biogenesis and release of exosomes and MV, thus influencing this mechanism of intercellular communication.
Cardiovascular Diseases and Comorbidities
Table 1 summarizes evidence of the association between the release of EV (both MVs and exosomes) and: cardiovascular risk factors (e.g., diabetes, hypertension, and sleep apnea), atherosclerosis and its complications. Furthermore, both the cell types involved in EV release and target cells are shown.
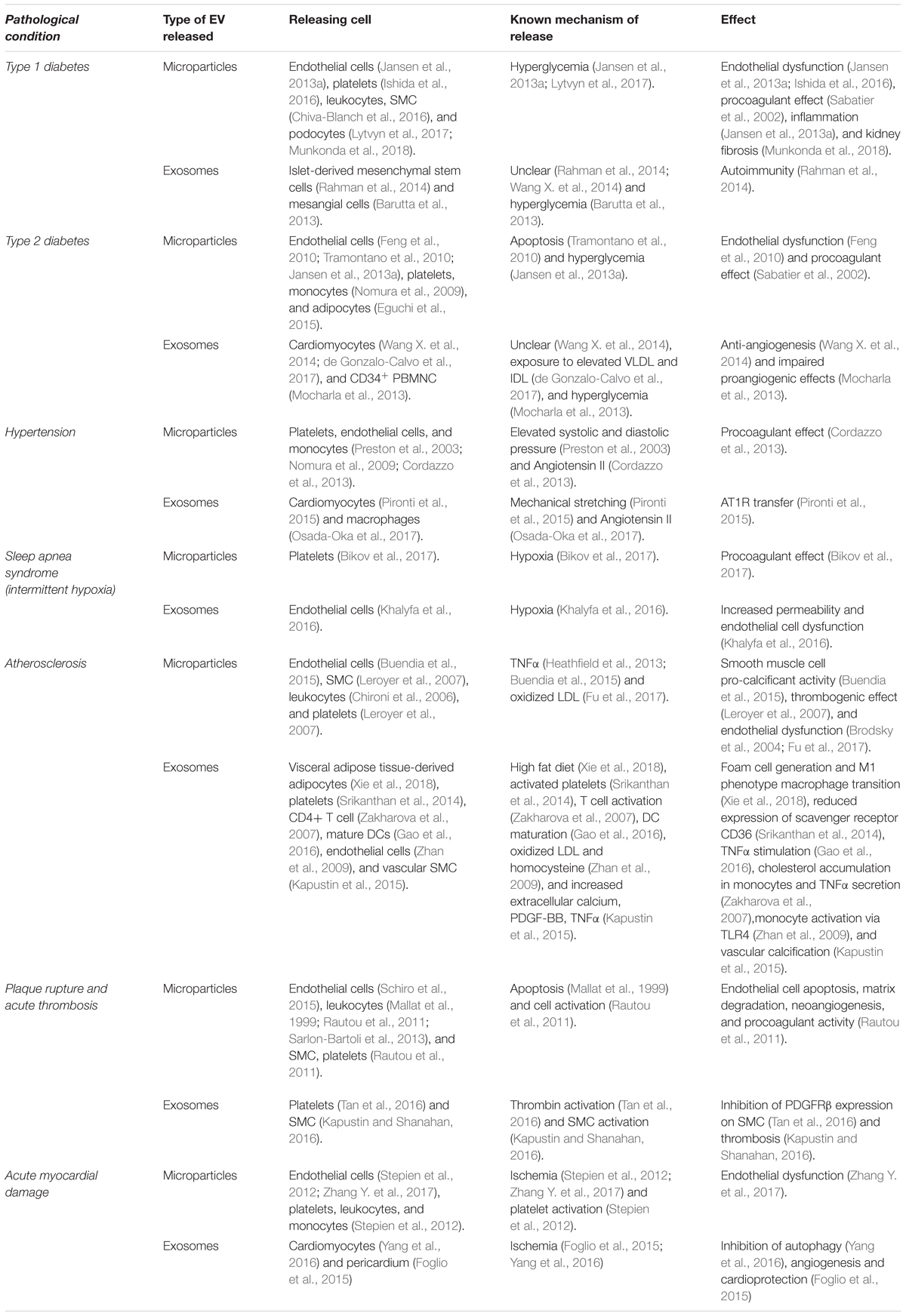
TABLE 1. Evidence of EV release in pathologic conditions.
Atherosclerosis
Concerning atherosclerosis, a large body of literature has shown the involvement of EV in crucial steps of its natural history. Endothelial cell dysfunction, the earliest event in the natural history of the disease can be promoted by endothelial derived microparticles. Indeed, early reports showed that MVs released by cultured endothelial cells reduce the acetylcholine induced vasorelaxation of isolated aortic rings, increasing the production of superoxide in target endothelial cells (Brodsky et al., 2004). More recently, microparticles were isolated both from the peripheral blood and atherosclerotic plaques of patients undergoing carotid endarterectomy. In both cases, most of the microparticles were of leukocyte origin. However, while a large fraction of circulating EV were of platelet origin, ≈13% of the vesicles isolated from the plaque were originated from SMC. Functionally, microparticles isolated from both sources expressed similar levels of tissue factor, but those of plaque origin generated more thrombin in vitro (Leroyer et al., 2007). Concerning atherosclerosis promoting factors, oxidized LDL and inflammatory mediators (e.g., TNFα) were shown to induce the release of microparticles from a broad range of cells, including endothelial cells, SMC, leukocytes, and platelets (Chironi et al., 2006; Leroyer et al., 2007; Heathfield et al., 2013; Buendia et al., 2015; Fu et al., 2017). Specifically, endothelial cells exposed to TNFα in vitro released microparticles that contained high amounts of BMP2 triggering vascular smooth muscle cell (SMC) calcification. Intriguingly, an identical procalcificant mechanism characterized MVs either released by senescent endothelial cells or isolated from patients affected by chronic kidney disease (Buendia et al., 2015). Similarly, stimulation of endothelial cells with oxidized LDL promoted the release of microparticles expressing ICAM-1 that could transfer this molecule to target endothelial cells, increasing their adhesion to monocytes (Fu et al., 2017).
Exosomes too have been implicated in the pathogenesis of atherosclerosis. Specifically, it has been shown that activated CD4+ T cells release exosomes that stimulated, in vitro, the accumulation of neutral lipids and free cholesterol, in THP-1 derived monocytes. Noteworthy, exosome-stimulated monocytes secreted TNFα as well (Zakharova et al., 2007). Furthermore, endothelial cells exposed to oxidized LDL or homocysteine released exosomes containing HSP70, which, in turn, increased the adhesion of monocytes to endothelial cells (Zhan et al., 2009). Exosomes derived from cultured, mature DCs may also represent an inflammatory stimulus for endothelial cells. Indeed, TNFα, exposed on the surface of these DC-derived particles, triggered, in target human umbilical vein endothelial cells, NFkB activation and expression of adhesion molecules (i.e., VCAM-1 and ICAM-1). Moreover, the in vivo administration of DC-derived particles to ApoE-/- mice worsened the atherosclerotic lesions of the animals (Gao et al., 2016). Cultured SMC too release exosomes in their supernatant, in a nSMase 2 dependent fashion. The release of SMC-derived exosomes was upregulated by calcifying conditions (i.e., in the presence of elevated levels of calcium and phosphate) and by PDGF-BB, while the proteomic analysis of these particles suggested their involvement in calcification processes. Consistently, SMC-derived exosomes could be detected in calcified arteries (Kapustin et al., 2015). Adipocyte derived exosomes too play a role in the atherogenic process. Specifically, visceral adipose tissue derived particles promoted cholesterol accumulation and enhanced macrophage foam cell formation by downregulating ATP-binding cassette transporter mediated cholesterol efflux (via ABCA1 and ABCG1) in RAW264.7 macrophages. Moreover, exosomes released by visceral adipocytes of animals fed on a high fat diet (VAT-HFD) induced the acquisition of a M1 phenotype by Raw264.7 macrophages. This process was a consequence of the activation of NFkB in macrophages and was paralleled by the increased secretion of proinflammatory cytokines IL6 and TNFα. In vivo administration of VAT-HFD derived exosomes too could exacerbate the atherosclerotic lesions of ApoE-/- mice (Xie et al., 2018). Last, exosomes may also have a protective effect, as in the case of those that are platelet derived and are produced during the atherothrombotic processes. Indeed, these particles promoted the ubiquitination of CD36, thus inhibiting both platelet activation and oxidized LDL binding to macrophages in vitro (Srikanthan et al., 2014).
Extracellular vesicles are involved in plaque rupture. Specifically, microparticles released by endothelial cells, leukocytes, SMC, and platelets can promote endothelial cell apoptosis, matrix degradation, inflammation, and intra plaque neoangiogenesis, thus destabilizing the plaque and increasing thrombus formation (Mallat et al., 1999; Rautou et al., 2011; Sarlon-Bartoli et al., 2013; Schiro et al., 2015). The clinical relevance of this phenomenon is supported by the observation that higher levels of circulating endothelial microparticles could be observed in the bloodstream of asymptomatic patients with unstable carotid artery plaques (Schiro et al., 2015). Moreover, the levels of circulating of CD11b+CD66b+ leukocyte derived microparticles proved to be independent predictors of carotid artery plaque instability (Sarlon-Bartoli et al., 2013).
Exosomes too could be produced by platelets and SMC in the context of a vulnerable plaque, contributing to the pathophysiology of the atherosclerotic plaque by favoring calcification and thrombosis (Kapustin and Shanahan, 2016; Tan et al., 2016).
Myocardial Infarction
With regard to ST-elevation myocardial infarction (STEMI), circulating microparticle levels increase acutely after the event and are associated with microvascular obstruction (Suades et al., 2016). Indeed, both ischemia and platelet activation trigger the release of MVs by endothelial cells, platelets, leukocytes, and monocytes (Stepien et al., 2012; Suades et al., 2016; Zhang Y. et al., 2017). Importantly, the levels of endothelial and platelet microparticles correlated with the dimension of the area at risk, in STEMI patients (Jung et al., 2012). Functionally, circulating microparticles isolated from infarcted patients were endowed with procoagulant activity, and altered endothelium-dependent vasorelaxation by impairing the endothelial nitric oxide pathway (Boulanger et al., 2001; Morel et al., 2004).
Concerning exosomes, their levels increase in the plasma of patients affected by both myocardial infarction and unstable angina, few hours after the acute event. Consistently, H9c2 myocytes released, in vitro, exosomes in their culture medium and the process was enhanced by hypoxia. Importantly, the HIF1α target microRNA30a that is released within exosomes, peaked after 4 h of hypoxia, was efficiently transferred to target myocytes, inhibited autophagy, and promoted apoptosis (Yang et al., 2016). Exosomes may also be found in the pericardial fluid (PF) and their concentration increases after acute myocardial infarction. These EV contain clusterin, that, in turn, promoted the epithelial to mesenchymal transition of epicardial cells, increasing the frequency of epicardial cells coexpressing smooth muscle actin and the stem cell marker c-Kit. Importantly, the in vivo administration of clusterin to infarcted mice increased angiogenesis and ameliorated myocardial function (Foglio et al., 2015). In line with these data, the direct proangiogenic effect of exosomes collected from the PF has been recently shown. Specifically, authors have shown the ability of PF derived exosomes to restore the angiogenic capacity of endothelial cells deprived of their microRNA content via Dicer silencing. Furthermore, PF exosomes improved post-ischemic blood flow recovery, in a mouse limb ischemia model (Beltrami et al., 2017).
Diabetes
In diabetes, different stimuli, such as hyperglycemia and apoptosis, promote the release of MVs from endothelial cells, platelets, leukocytes, SMCs, mesangial cells, and podocytes (Tramontano et al., 2010; Barutta et al., 2013; Jansen et al., 2013a; Chiva-Blanch et al., 2016; Ishida et al., 2016; Lytvyn et al., 2017; Munkonda et al., 2018). Functionally, microparticles isolated from diabetic rat plasma that originate mainly from platelets, were able to decrease acetylcholine-induced endothelium-dependent relaxation on carotid artery rings, ex vivo. These effects were associated with a decreased expression of eNOS, and with an incremented expression of caveolin-1 in target carotids (Ishida et al., 2016). Microparticles released by endothelial cells exposed to hyperglycemia were characterized by increased NADPH oxidase activity and reactive oxygen (ROS) levels. When human coronary endothelial cells were cultured in the presence of these pathologic microparticles, they increased intracellular ROS levels, activated p38 MAPK in a ROS dependent fashion, and expressed the adhesive proteins ICAM-1 and VCAM-1, thus incrementing monocyte adhesion (Jansen et al., 2013a). Microparticles released by endothelial cells and monocytes seem to link vascular injury with kidney disease. Indeed, addition of microparticles released by these two cell types, in response to TNFα stimulation, to differentiated podocytes upregulated the production of the proinflammatory mediators MCP-1 and interleukin-6, in vitro (Eyre et al., 2011). In turn, proximal tubule epithelial cells, cultured in the presence of podocyte derived microparticles, activated p38 MAPK, and TGFb signaling, promoting the secretion of fibronectin/collagen type IV, potentially contributing to renal fibrosis. This effect could be inhibited blocking the scavenger receptor CD36 (Munkonda et al., 2018). Last, Annexin V+ microparticles can be released by adipocytes during lipotoxic stress, in a Caspase-3 and Rho-associated kinase fashion, promoting migration of monocytes and macrophages to the adipose tissue (Eguchi et al., 2015).
Concerning exosomes, it has been shown that they could play a relevant role in the pathogenesis of type 1 diabetes. Indeed, islet-derived mesenchymal stem cells (iMSC) released exosomes that were able to activate autoreactive B and T cells, primed in NOD mice. Immunization of mice with iMSC derived exosomes favored the expansion of diabetogenic T cells and accelerated the T cell mediated destruction of islets (Rahman et al., 2014). Although it has been shown that exosomes and specific microRNAs could be found in the urine during the evolution of diabetic nephropathy, few works have demonstrated the direct pathophysiological role of these small EV (Barutta et al., 2013). However, it was recently shown that rat proximal tubule cells exposed to advanced glycation end-products (AGE) increased the secretion of exosomes containing C-megalin. The latter is a multiligand endocytic scavenger receptor that binds to ligands such as AGE-modified bovine serum albumin, promoting dysfunction of the autophagy/lysosomal pathway and exosomal C-megalin excretion (De et al., 2017). Hyperglycemia exerts also a negative impact on CD34+CD14+ and CD34+CD14- circulating proangiogenic cells by inhibiting the secretion of the angiogenic microRNA126 within exosomes (Mocharla et al., 2013). Last, cardiomyocytes too, when exposed to hyperglycemia or VLDL/IDL, secrete exosomes (Wang X. et al., 2014; de Gonzalo-Calvo et al., 2017). These contain microRNA1 and microRNA133a and proved to be independent predictors of myocardial steatosis, in uncomplicated type-2 diabetes (de Gonzalo-Calvo et al., 2017). Functionally, exosomes released from cardiomyocytes isolated from Goto-Kakizaki rats (an animal model that spontaneously develops type 2 diabetes) inhibited the activities of proliferation, migration, and formation of tubule-like structures of cardiac endothelial cells. The effect was mediated by the transfer of the antiangiogenic microRNA320 to target cells (Wang X. et al., 2014).
Last, diabetes, may reduce the positive effect exerted by EV. Indeed, although endothelial microparticles can promote angiogenesis via microRNA126, diabetes reduces the microRNA126 content of these EV, thus impairing their proangiogenic capability (Jansen et al., 2013b). Similarly, while cardiomyocytes release exosomes that promote endothelial cell proliferation, migration, and capillary network formation in vitro, diabetes abrogated this protective effect (Wang X. et al., 2014).
Hypertension
With regard to other risk factors for cardiovascular disease, both elevated artery blood pressure and angiotensin II (Ang II) stimulate the release of microparticles from platelets, endothelial cells, and monocytes (Preston et al., 2003; Nomura et al., 2009; Cordazzo et al., 2013). Importantly, the levels of both platelet and endothelial microparticles correlated with: blood pressure, markers of endothelial and platelet activation (i.e., sVCAM-1 and sICAM-1), and vWF, a marker of endothelial injury/dysfunction (Preston et al., 2003). In a cohort of hypertensive patients with diabetes, systolic blood pressure, HDL cholesterol, sP-selectin, sE-selectin, sCD40L, RANTES, monocyte derived microparticle levels, and endothelial derived microparticle levels were independent predictors of the levels of platelet derived microparticles (Nomura et al., 2009). These results suggest that microparticles are at the crossroads between endothelial injury, coagulation, and inflammation. In line, exposure of mononuclear cells to Ang II, increased intracellular calcium levels and stimulated the release of procoagulant, tissue factor-bearing MVs (Cordazzo et al., 2013). Similarly, stimulation of endothelial cells with Ang II promoted superoxide generation, Rho kinase activity, and MV release, which, in turn, exerted prooxidative and proinflammatory effects on normal endothelial cells (Burger et al., 2011). Importantly, Ang II receptor blockers inhibited MV release.
Concerning exosomes, both mechanical stretching, and Ang II can promote their release from cardiomyocytes and macrophages (Pironti et al., 2015; Osada-Oka et al., 2017). Functionally, these vesicles could mediate the transfer of AT1R on cardiac and skeletal myocytes, and mesenteric vessels, conferring responsiveness to Ang II infusion in AT1R knockout mice (Pironti et al., 2015). Moreover, THP-1-derived macrophages, grown in the presence of Ang II, released exosomes that increased the expression of ICAM-1 and PAI-1 on target endothelial cells (Osada-Oka et al., 2017).
Sleep Apnea Syndrome and Hypoxic Conditions
In patients suffering from sleep apnea syndrome, episodes of intermittent hypoxia are associated with the release of circulating CD41+ and Annexin V+ microparticles of platelet origin, which show a diurnal fluctuation that reaches a significant peak at 5 pm. While the levels of CD41+ MVs positively correlated with the severity of obstructive sleep apnea, continuous positive airway pressure therapy reduced their diurnal peak (Bikov et al., 2017). Moreover, exposure of healthy volunteers to intermittent hypoxia led to the release of exosomes of endothelial cell, lymphocyte, monocyte and platelet origin into the bloodstream. Exosomes released after 4 days of exposure to intermittent hypoxia (4D IH) increased the expression of ICAM-1, decreased the expression of eNOS, and reduced the cell barrier function of cultured endothelial cells. Moreover, endothelial cells treated with 4D IH exosomes increased their adhesion to monocytes (Khalyfa et al., 2016). Indeed, hypoxia is a potent trigger for EV secretion, activating HIF1α and HIF2α (Wang T. et al., 2014). Furthermore, during ischemia reperfusion injury, stimuli such as ROS and activation of purinergic receptors can act as additional triggers of EV release (Szotowski et al., 2007; Kahner et al., 2008; Bianco et al., 2009; Drago et al., 2017).
Effect of Therapies on EV Release
Considering the biogenesis and the mechanisms of release of EV, it is conceivable how not only diseases but also drugs can impact EV production.
Since variations in intracellular calcium levels modulate the release of both exosomes and MV, we can anticipate that drugs interfering with calcium homeostasis modify the secretory profile of treated cells. In line, amiloride, a drug that inhibits both the Na+/H+ exchanger and the Na+/Ca2+ channel, has been shown to reduce both constitutive and stimulated exosome release in vitro (Savina et al., 2003). Similarly, nifedipine, a calcium channel blocker, was able to reduce the release of platelet derived microparticles (Lee et al., 1993). However, the effect of the latter drug on the release of EV by cardiac cells has not been assessed yet. Conversely, no effect was observed when the L-type calcium channel inhibitor verapamil was employed (Savina et al., 2003). Importantly, calcium channel blockers and β-blockers may also inhibit EV uptake, by reducing endothelial cell activation (Xiao X. et al., 2017).
An intriguing observation, conducted in the oncology field, is that a low environmental pH is associated with enhanced secretion and uptake of exosomes (Parolini et al., 2009). In line, omeprazole and other H+/K+ pump inhibitors, such as omeprazole, reduce both exosome release and their uptake (Chalmin et al., 2010). Although these observations have been conducted on tumors, it would be of outmost important to verify whether these drugs interfere with EV release during cardiac ischemic events, conditions that are coupled with the acidification of the interstitial space.
Consistently with the role played by cholesterol in exosome biogenesis, it has been shown that hydroxy-methylglutaryl-coenzyme A (HMG-CoA) inhibitors modulate the release of exosomes. Specifically, they increase exosome secretion from microglial cells, and promote the release of immunomodulatory exosomes from DCs (Tamboli et al., 2010; Li et al., 2016). Concerning other drugs interfering with lipid metabolism, it has been shown that amiloride could inhibit ceramide formation, possibly by indirectly inhibiting the acid sphingomyelinase, preventing the formation of an acidic microenvironment (Serrano et al., 2012). Similarly, GW4869, a nSMase 2 inhibitor, could blunt ceramide formation and exosome biogenesis (Panigrahi et al., 2018).
Given the described role played by ADP in promoting EV release, it is not surprising that thienopyridines, e.g., ticlopidine, clopidogrel, and prasugrel (P2Y12 inhibitors), can negatively modulate the release of EV (Badimon et al., 2017). Furthermore, other antiaggregant drugs, such as aspirin, lower the level of circulating microparticles both in vitro and in diabetic patients (Chiva-Blanch et al., 2016; Connor et al., 2016).
In line with the stimulatory effect of TNFα on the release of endothelial cell derived MVs (see Table 1), the use of an inhibitor of this cytokine (certolizumab) reduced EV secretion in vitro (Heathfield et al., 2013).
Moreover, while digoxin possibly reduces exosome release, acting as a non-specific HIF1α inhibitor, it has been shown to increase the release of endothelial cell and platelet derived microparticles, in patients affected by atrial fibrillation (Chirinos et al., 2005; Panigrahi et al., 2018).
Of particular relevance in oncology is that cisplatin, which is usually concentrated in the lysosomal compartment, could be secreted via the exosomal pathway, in drug resistant cells (Safaei et al., 2005). Similarly, doxorubicin can be secreted within exosomes, but this process could be inhibited in vitro by ketotifen, a stabilizer of the mastocyte membrane, thus reversing drug resistance (Khan et al., 2018).
Last, prompted by early reports, showing that the cardioprotective and proangiogenic activities of both mesenchymal stem cells and CD34+ progenitors could be found in the exosomal fraction of their secretome, several investigators have evaluated, in recent years, the cardioprotective role of EV secreted by different cell types (Lai et al., 2010; Sahoo et al., 2011). In line, microparticles released by apoptotic endothelial cells promote endothelial cell migration and proliferation, thus promoting reendothelialization in vivo, with a microRNA126-dependent mechanism (Jansen et al., 2013b). Cardiac progenitors (CPC) too release exosomes that are required to protect HL-1 myocytes from apoptosis and to promote angiogenesis in vitro. These exosomes, that contain proangiogenic and anti-apoptotic microRNAs, reduced the infarct size, decreasing cardiomyocyte apoptosis, and increasing angiogenesis in vivo (Barile et al., 2014). Intriguingly, the protective effect of CPC-derived exosomes mimicked the effect of cell therapy on acute myocardial infarction, possibly as a consequence of the modulation of the immune system. Specifically, exosomes reduced the accumulation of macrophages in the zone bordering the infarcted area and promoted their polarization toward an anti-inflammatory phenotype, both in vivo and in vitro. Exosome-mediated transfer of microRNA181b to macrophages may be the molecular mechanism responsible for the phenomenon (de Couto et al., 2017). The protein content of exosome too has been considered to be responsible for the beneficial effects of CPC-derived exosomes. Indeed, pregnancy-associated plasma protein-A (PAPP-A) is one of the most highly enriched proteins in the comparison between CPC-derived exosomes and bone marrow mononuclear cell derived exosomes. PAPP-A, whose active form was identified on the exosome surface, cleaves IGFBP-4 and increases the bioavailability of IGF-1, thus reducing cardiomyocyte apoptosis. Consistently, CPC-derived exosomes reduced infarct size and ameliorate ventricular function more efficiently than bone marrow derived ones (Barile et al., 2018).
Conclusion
In conclusion, the term EV encompasses a broad category of cell derived vesicles that differ in biogenesis and biological properties. As a consequence, their fractionation and characterization still represent a challenge. As consequence of this and of the fact that it is an emerging area of investigation, the current literature may be confounding. However, the conflicting results could just reflect heterogeneity in EV preparation and characterization and lack of knowledge or attention to confounding factors. Among the most prominent ones, comorbidities and ongoing pharmacological therapies may have a profound impact in either stimulating or inhibiting EV release or in modifying the characteristics of the secreted EV. In line, according to the literature, EV secreted by a specific cell type may exert protective effects, under normal conditions, while they could exert deleterious effects in pathological settings. Similarly, priming of a cell type with a specific drug may increase the immunomodulatory function of EV secreted by a given cell type.
Therefore, in future studies, the pathologic status, and the ongoing drug therapies must be considered as covariates, when analyzing for the biological effect of a specific EV type.
Methods
To identify the scientific literature regarding EV and cardiovascular diseases, we reviewed the literature in the PubMed database. We focused on publications written in English and published in the last ≈10 years. As search terms, we used “EV” or “MVs” or “exosomes” and either “cardiovascular disease” or “diabetes”. Due to the large number of papers, more than 2,000 scientific works, we were not able to cite all individual references and we summarized those that we consider the most important concepts and quoted representative works. We apologize to all authors whose important publications are not cited.
Author Contributions
AA, DC, and AB wrote and reviewed the manuscript. PP, CC, CV, and AZ reviewed the manuscript.
Funding
The authors acknowledge funding from the European Regional Development Fund Interreg V-A Italia–Austria 2014–2020 (EXOTHERA ITAT1036).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Al-Nedawi, K., Meehan, B., Kerbel, R. S., Allison, A. C., and Rak, J. (2009). Endothelial expression of autocrine VEGF upon the uptake of tumor-derived microvesicles containing oncogenic EGFR. Proc. Natl. Acad. Sci. U.S.A. 106, 3794–3799. doi: 10.1073/pnas.0804543106
Alonso, R., Mazzeo, C., Rodriguez, M. C., Marsh, M., Fraile-Ramos, A., Calvo, V., et al. (2011). Diacylglycerol kinase alpha regulates the formation and polarisation of mature multivesicular bodies involved in the secretion of Fas ligand-containing exosomes in T lymphocytes. Cell Death Differ. 18, 1161–1173. doi: 10.1038/cdd.2010.184
Amidon, B., Brown, A., and Waite, M. (1996). Transacylase and phospholipases in the synthesis of bis(monoacylglycero)phosphate. Biochemistry 35, 13995–14002. doi: 10.1021/bi961164o
Andreu, Z., and Yanez-Mo, M. (2014). Tetraspanins in extracellular vesicle formation and function. Front. Immunol. 5:442. doi: 10.3389/fimmu.2014.00442
Antonyak, M. A., and Cerione, R. A. (2015). Emerging picture of the distinct traits and functions of microvesicles and exosomes. Proc. Natl. Acad. Sci. U.S.A. 112, 3589–3590. doi: 10.1073/pnas.1502590112
Augustine, D., Ayers, L. V., Lima, E., Newton, L., Lewandowski, A. J., Davis, E. F., et al. (2014). Dynamic release and clearance of circulating microparticles during cardiac stress. Circ. Res. 114, 109–113. doi: 10.1161/CIRCRESAHA.114.301904
Badimon, L., Suades, R., Arderiu, G., Pena, E., Chiva-Blanch, G., and Padro, T. (2017). Microvesicles in atherosclerosis and angiogenesis: from bench to bedside and reverse. Front. Cardiovasc. Med. 4:77. doi: 10.3389/fcvm.2017.00077
Badimon, L., Suades, R., Fuentes, E., Palomo, I., and Padro, T. (2016). Role of platelet-derived microvesicles as crosstalk mediators in atherothrombosis and future pharmacology targets: a link between inflammation, atherosclerosis, and thrombosis. Front. Pharmacol. 7:293. doi: 10.3389/fphar.2016.00293
Baietti, M. F., Zhang, Z., Mortier, E., Melchior, A., Degeest, G., Geeraerts, A., et al. (2012). Syndecan-syntenin-ALIX regulates the biogenesis of exosomes. Nat. Cell Biol. 14, 677–685. doi: 10.1038/ncb2502
Baixauli, F., Lopez-Otin, C., and Mittelbrunn, M. (2014). Exosomes and autophagy: coordinated mechanisms for the maintenance of cellular fitness. Front. Immunol. 5:403. doi: 10.3389/fimmu.2014.00403
Bari, R., Guo, Q., Xia, B., Zhang, Y. H., Giesert, E. E., Levy, S., et al. (2011). Tetraspanins regulate the protrusive activities of cell membrane. Biochem. Biophys. Res. Commun. 415, 619–626. doi: 10.1016/j.bbrc.2011.10.121
Barile, L., Cervio, E., Lionetti, V., Milano, G., Ciullo, A., Biemmi, V., et al. (2018). Cardioprotection by cardiac progenitor cell-secreted exosomes: role of pregnancy-associated plasma protein-A. Cardiovasc. Res. 114, 992–1005. doi: 10.1093/cvr/cvy055
Barile, L., Lionetti, V., Cervio, E., Matteucci, M., Gherghiceanu, M., Popescu, L. M., et al. (2014). Extracellular vesicles from human cardiac progenitor cells inhibit cardiomyocyte apoptosis and improve cardiac function after myocardial infarction. Cardiovasc. Res. 103, 530–541. doi: 10.1093/cvr/cvu167
Barutta, F., Tricarico, M., Corbelli, A., Annaratone, L., Pinach, S., Grimaldi, S., et al. (2013). Urinary exosomal microRNAs in incipient diabetic nephropathy. PLoS One 8:e73798. doi: 10.1371/journal.pone.0073798
Beltrami, C., Besnier, M., Shantikumar, S., Shearn, A. I., Rajakaruna, C., Laftah, A., et al. (2017). Human pericardial fluid contains exosomes enriched with cardiovascular-expressed MicroRNAs and promotes therapeutic angiogenesis. Mol. Ther. 25, 679–693. doi: 10.1016/j.ymthe.2016.12.022
Bianco, F., Perrotta, C., Novellino, L., Francolini, M., Riganti, L., Menna, E., et al. (2009). Acid sphingomyelinase activity triggers microparticle release from glial cells. EMBO J. 28, 1043–1054. doi: 10.1038/emboj.2009.45
Bikov, A., Kunos, L., Pallinger, E., Lazar, Z., Kis, A., Horvath, G., et al. (2017). Diurnal variation of circulating microvesicles is associated with the severity of obstructive sleep apnoea. Sleep Breath 21, 595–600. doi: 10.1007/s11325-017-1464-y
Blanc, L., and Vidal, M. (2017). New insights into the function of Rab GTPases in the context of exosomal secretion. Small GTPases 9, 95–106. doi: 10.1080/21541248.2016.1264352
Boulanger, C. M., Scoazec, A., Ebrahimian, T., Henry, P., Mathieu, E., Tedgui, A., et al. (2001). Circulating microparticles from patients with myocardial infarction cause endothelial dysfunction. Circulation 104, 2649–2652. doi: 10.1161/hc4701.100516
Brodsky, S. V., Zhang, F., Nasjletti, A., and Goligorsky, M. S. (2004). Endothelium-derived microparticles impair endothelial function in vitro. Am. J. Physiol. Heart Circ. Physiol. 286, H1910–H1915. doi: 10.1152/ajpheart.01172.2003
Buendia, P., Montes de Oca, A., Madueno, J. A., Merino, A., Martin-Malo, A., Aljama, P., et al. (2015). Endothelial microparticles mediate inflammation-induced vascular calcification. FASEB J. 29, 173–181. doi: 10.1096/fj.14-249706
Burger, D., Montezano, A. C., Nishigaki, N., He, Y., Carter, A., and Touyz, R. M. (2011). Endothelial microparticle formation by angiotensin II is mediated via Ang II receptor type I/NADPH oxidase/ Rho kinase pathways targeted to lipid rafts. Arterioscler. Thromb. Vasc. Biol. 31, 1898–1907. doi: 10.1161/ATVBAHA.110.222703
Buschow, S. I., Nolte-’t Hoen, E. N., van Niel, G., Pols, M. S., ten Broeke, T., Lauwen, M., et al. (2009). MHC II in dendritic cells is targeted to lysosomes or T cell-induced exosomes via distinct multivesicular body pathways. Traffic 10, 1528–1542. doi: 10.1111/j.1600-0854.2009.00963.x
Chairoungdua, A., Smith, D. L., Pochard, P., Hull, M., and Caplan, M. J. (2010). Exosome release of beta-catenin: a novel mechanism that antagonizes Wnt signaling. J. Cell Biol. 190, 1079–1091. doi: 10.1083/jcb.201002049
Chalmin, F., Ladoire, S., Mignot, G., Vincent, J., Bruchard, M., Remy-Martin, J. P., et al. (2010). Membrane-associated Hsp72 from tumor-derived exosomes mediates STAT3-dependent immunosuppressive function of mouse and human myeloid-derived suppressor cells. J. Clin. Invest. 120, 457–471. doi: 10.1172/JCI40483
Chen, B., Li, Q., Zhao, B., and Wang, Y. (2017). Stem cell-derived extracellular vesicles as a novel potential therapeutic tool for tissue repair. Stem Cells Transl. Med. 6, 1753–1758. doi: 10.1002/sctm.16-0477
Chevallier, J., Chamoun, Z., Jiang, G., Prestwich, G., Sakai, N., Matile, S., et al. (2008). Lysobisphosphatidic acid controls endosomal cholesterol levels. J. Biol. Chem. 283, 27871–27880. doi: 10.1074/jbc.M801463200
Chirinos, J. A., Castrellon, A., Zambrano, J. P., Jimenez, J. J., Jy, W., Horstman, L. L., et al. (2005). Digoxin use is associated with increased platelet and endothelial cell activation in patients with nonvalvular atrial fibrillation. Heart Rhythm. 2, 525–529. doi: 10.1016/j.hrthm.2005.01.016
Chironi, G., Simon, A., Hugel, B., Del Pino, M., Gariepy, J., Freyssinet, J. M., et al. (2006). Circulating leukocyte-derived microparticles predict subclinical atherosclerosis burden in asymptomatic subjects. Arterioscler. Thromb. Vasc. Biol. 26, 2775–2780. doi: 10.1161/01.ATV.0000249639.36915.04
Chiva-Blanch, G., Suades, R., Padro, T., Vilahur, G., Pena, E., Ybarra, J., et al. (2016). Microparticle shedding by erythrocytes, monocytes and vascular smooth muscular cells is reduced by aspirin in diabetic patients. Rev. Esp. Cardiol. 69, 672–680. doi: 10.1016/j.rec.2015.12.033
Colombo, M., Raposo, G., and Thery, C. (2014). Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu. Rev. Cell Dev. Biol. 30, 255–289. doi: 10.1146/annurev-cellbio-101512-122326
Connor, D. E., Ly, K., Aslam, A., Boland, J., Low, J., Jarvis, S., et al. (2016). Effects of antiplatelet therapy on platelet extracellular vesicle release and procoagulant activity in health and in cardiovascular disease. Platelets 27, 805–811. doi: 10.1080/09537104.2016.1190008
Cordazzo, C., Neri, T., Petrini, S., Lombardi, S., Balia, C., Cianchetti, S., et al. (2013). Angiotensin II induces the generation of procoagulant microparticles by human mononuclear cells via an angiotensin type 2 receptor-mediated pathway. Thromb. Res. 131, e168–e174. doi: 10.1016/j.thromres.2013.01.019
De, S., Kuwahara, S., Hosojima, M., Ishikawa, T., Kaseda, R., Sarkar, P., et al. (2017). Exocytosis-Mediated urinary full-length megalin excretion is linked with the pathogenesis of diabetic nephropathy. Diabetes Metab. Res. Rev. 66, 1391–1404. doi: 10.2337/db16-1031
de Couto, G., Gallet, R., Cambier, L., Jaghatspanyan, E., Makkar, N., Dawkins, J. F., et al. (2017). Exosomal MicroRNA transfer into macrophages mediates cellular postconditioning. Circulation 136, 200–214. doi: 10.1161/CIRCULATIONAHA.116.024590
de Gonzalo-Calvo, D., van der Meer, R. W., Rijzewijk, L. J., Smit, J. W., Revuelta-Lopez, E., Nasarre, L., et al. (2017). Serum microRNA-1 and microRNA-133a levels reflect myocardial steatosis in uncomplicated type 2 diabetes. Sci. Rep. 7:47. doi: 10.1038/s41598-017-00070-6
Derganc, J., and Copic, A. (2016). Membrane bending by protein crowding is affected by protein lateral confinement. Biochim. Biophys. Acta 1858, 1152–1159. doi: 10.1016/j.bbamem.2016.03.009
Drago, F., Lombardi, M., Prada, I., Gabrielli, M., Joshi, P., Cojoc, D., et al. (2017). ATP modifies the proteome of extracellular vesicles released by microglia and influences their action on astrocytes. Front. Pharmacol. 8:910. doi: 10.3389/fphar.2017.00910
Eguchi, A., Mulya, A., Lazic, M., Radhakrishnan, D., Berk, M. P., Povero, D., et al. (2015). Microparticles release by adipocytes act as “find-me” signals to promote macrophage migration. PLoS One 10:e0123110. doi: 10.1371/journal.pone.0123110
Eyre, J., Burton, J. O., Saleem, M. A., Mathieson, P. W., Topham, P. S., and Brunskill, N. J. (2011). Monocyte- and endothelial-derived microparticles induce an inflammatory phenotype in human podocytes. Nephron Exp. Nephrol. 119, e58–e66. doi: 10.1159/000329575
Fader, C. M., Sanchez, D. G., Mestre, M. B., and Colombo, M. I. (2009). TI-VAMP/VAMP7 and VAMP3/cellubrevin: two v-SNARE proteins involved in specific steps of the autophagy/multivesicular body pathways. Biochim. Biophys. Acta 1793, 1901–1916. doi: 10.1016/j.bbamcr.2009.09.011
Fatima, F., Ekstrom, K., Nazarenko, I., Maugeri, M., Valadi, H., Hill, A. F., et al. (2017). Non-coding RNAs in mesenchymal stem cell-derived extracellular vesicles: deciphering regulatory roles in stem cell potency, inflammatory resolve, and tissue regeneration. Front. Genet. 8:161. doi: 10.3389/fgene.2017.00161
Feng, B., Chen, Y., Luo, Y., Chen, M., Li, X., and Ni, Y. (2010). Circulating level of microparticles and their correlation with arterial elasticity and endothelium-dependent dilation in patients with type 2 diabetes mellitus. Atherosclerosis 208, 264–269. doi: 10.1016/j.atherosclerosis.2009.06.037
Foglio, E., Puddighinu, G., Fasanaro, P., D’Arcangelo, D., Perrone, G. A., Mocini, D., et al. (2015). Exosomal clusterin, identified in the pericardial fluid, improves myocardial performance following MI through epicardial activation, enhanced arteriogenesis and reduced apoptosis. Int. J. Cardiol. 197, 333–347. doi: 10.1016/j.ijcard.2015.06.008
Fu, Z., Zhou, E., Wang, X., Tian, M., Kong, J., Li, J., et al. (2017). Oxidized low-density lipoprotein-induced microparticles promote endothelial monocyte adhesion via intercellular adhesion molecule 1. Am. J. Physiol. Cell Physiol. 313, C567–C574. doi: 10.1152/ajpcell.00158.2016
Furthauer, M. (2018). The ESCRT machinery: when function follows form. Semin. Cell Dev. Biol. 74, 1–3. doi: 10.1016/j.semcdb.2017.11.003
Gao, W., Liu, H., Yuan, J., Wu, C., Huang, D., Ma, Y., et al. (2016). Exosomes derived from mature dendritic cells increase endothelial inflammation and atherosclerosis via membrane TNF-alpha mediated NF-kappaB pathway. J. Cell Mol. Med. 20, 2318–2327. doi: 10.1111/jcmm.12923
Ghossoub, R., Lembo, F., Rubio, A., Gaillard, C. B., Bouchet, J., Vitale, N., et al. (2014). Syntenin-ALIX exosome biogenesis and budding into multivesicular bodies are controlled by ARF6 and PLD2. Nat. Commun. 5:3477. doi: 10.1038/ncomms4477
Gross, J. C., Chaudhary, V., Bartscherer, K., and Boutros, M. (2012). Active Wnt proteins are secreted on exosomes. Nat. Cell Biol. 14, 1036–1045. doi: 10.1038/ncb2574
Grove, J. (2014). Super-resolution microscopy: a virus’ eye view of the cell. Viruses 6, 1365–1378. doi: 10.3390/v6031365
Gulinelli, S., Salaro, E., Vuerich, M., Bozzato, D., Pizzirani, C., Bolognesi, G., et al. (2012). IL-18 associates to microvesicles shed from human macrophages by a LPS/TLR-4 independent mechanism in response to P2X receptor stimulation. Eur. J. Immunol. 42, 3334–3345. doi: 10.1002/eji.201142268
Guo, B. B., Bellingham, S. A., and Hill, A. F. (2016). Stimulating the release of exosomes increases the intercellular transfer of prions. J. Biol. Chem. 291, 5128–5137. doi: 10.1074/jbc.M115.684258
Hagiwara, K., Katsuda, T., Gailhouste, L., Kosaka, N., and Ochiya, T. (2015). Commitment of Annexin A2 in recruitment of microRNAs into extracellular vesicles. FEBS Lett. 589(24 Pt B), 4071–4078. doi: 10.1016/j.febslet.2015.11.036
Heathfield, S. K., Parker, B., Zeef, L. A., Bruce, I. N., and Alexander, M. Y. (2013). Certolizumab pegol attenuates the pro-inflammatory state in endothelial cells in a manner that is atheroprotective. Clin. Exp. Rheumatol. 31, 225–233.
Ishida, K., Taguchi, K., Hida, M., Watanabe, S., Kawano, K., Matsumoto, T., et al. (2016). Circulating microparticles from diabetic rats impair endothelial function and regulate endothelial protein expression. Acta Physiol. 216, 211–220. doi: 10.1111/apha.12561
Janas, T., Janas, M. M., Sapon, K., and Janas, T. (2015). Mechanisms of RNA loading into exosomes. FEBS Lett. 589, 1391–1398. doi: 10.1016/j.febslet.2015.04.036
Janas, T., Janas, T., and Yarus, M. (2012). Human tRNA(Sec) associates with HeLa membranes, cell lipid liposomes, and synthetic lipid bilayers. RNA 18, 2260–2268. doi: 10.1261/rna.035352.112
Jansen, F., Yang, X., Franklin, B. S., Hoelscher, M., Schmitz, T., Bedorf, J., et al. (2013a). High glucose condition increases NADPH oxidase activity in endothelial microparticles that promote vascular inflammation. Cardiovasc. Res. 98, 94–106. doi: 10.1093/cvr/cvt013
Jansen, F., Yang, X., Hoelscher, M., Cattelan, A., Schmitz, T., Proebsting, S., et al. (2013b). Endothelial microparticle-mediated transfer of MicroRNA-126 promotes vascular endothelial cell repair via SPRED1 and is abrogated in glucose-damaged endothelial microparticles. Circulation 128, 2026–2038. doi: 10.1161/CIRCULATIONAHA.113.001720
Jung, C., Sorensson, P., Saleh, N., Arheden, H., Ryden, L., and Pernow, J. (2012). Circulating endothelial and platelet derived microparticles reflect the size of myocardium at risk in patients with ST-elevation myocardial infarction. Atherosclerosis 221, 226–231. doi: 10.1016/j.atherosclerosis.2011.12.025
Kahner, B. N., Dorsam, R. T., and Kunapuli, S. P. (2008). Role of P2Y receptor subtypes in platelet-derived microparticle generation. Front. Biosci. 13, 433–439. doi: 10.2741/2690
Kajimoto, T., Okada, T., Miya, S., Zhang, L., and Nakamura, S. (2013). Ongoing activation of sphingosine 1-phosphate receptors mediates maturation of exosomal multivesicular endosomes. Nat. Commun. 4:2712. doi: 10.1038/ncomms3712
Kanada, M., Bachmann, M. H., Hardy, J. W., Frimannson, D. O., Bronsart, L., Wang, A., et al. (2015). Differential fates of biomolecules delivered to target cells via extracellular vesicles. Proc. Natl. Acad. Sci. U.S.A. 112, E1433–E1442. doi: 10.1073/pnas.1418401112
Kapustin, A. N., Chatrou, M. L., Drozdov, I., Zheng, Y., Davidson, S. M., Soong, D., et al. (2015). Vascular smooth muscle cell calcification is mediated by regulated exosome secretion. Circ. Res. 116, 1312–1323. doi: 10.1161/CIRCRESAHA.116.305012
Kapustin, A. N., and Shanahan, C. M. (2016). Emerging roles for vascular smooth muscle cell exosomes in calcification and coagulation. J. Physiol. 594, 2905–2914. doi: 10.1113/JP271340
Khalyfa, A., Zhang, C., Khalyfa, A. A., Foster, G. E., Beaudin, A. E., Andrade, J., et al. (2016). Effect on intermittent hypoxia on plasma exosomal micro RNA signature and endothelial function in healthy adults. Sleep 39, 2077–2090. doi: 10.5665/sleep.6302
Khan, F. M., Saleh, E., Alawadhi, H., Harati, R., Zimmermann, W. H., and El-Awady, R. (2018). Inhibition of exosome release by ketotifen enhances sensitivity of cancer cells to doxorubicin. Cancer Biol. Ther. 19, 25–33. doi: 10.1080/15384047.2017.1394544
King, H. W., Michael, M. Z., and Gleadle, J. M. (2012). Hypoxic enhancement of exosome release by breast cancer cells. BMC Cancer 12:421. doi: 10.1186/1471-2407-12-421
Kirkegaard, T., Roth, A. G., Petersen, N. H., Mahalka, A. K., Olsen, O. D., Moilanen, I., et al. (2010). Hsp70 stabilizes lysosomes and reverts Niemann-Pick disease-associated lysosomal pathology. Nature 463, 549–553. doi: 10.1038/nature08710
Koles, K., Nunnari, J., Korkut, C., Barria, R., Brewer, C., Li, Y., et al. (2012). Mechanism of evenness interrupted (Evi)-exosome release at synaptic boutons. J. Biol. Chem. 287, 16820–16834. doi: 10.1074/jbc.M112.342667
Kooijman, E. E., Chupin, V., de Kruijff, B., and Burger, K. N. (2003). Modulation of membrane curvature by phosphatidic acid and lysophosphatidic acid. Traffic 4, 162–174. doi: 10.1034/j.1600-0854.2003.00086.x
Kosaka, N., Iguchi, H., Yoshioka, Y., Takeshita, F., Matsuki, Y., and Ochiya, T. (2010). Secretory mechanisms and intercellular transfer of microRNAs in living cells. J. Biol. Chem. 285, 17442–17452. doi: 10.1074/jbc.M110.107821
Kucharzewska, P., Christianson, H. C., Welch, J. E., Svensson, K. J., Fredlund, E., Ringner, M., et al. (2013). Exosomes reflect the hypoxic status of glioma cells and mediate hypoxia-dependent activation of vascular cells during tumor development. Proc. Natl. Acad. Sci. U.S.A. 110, 7312–7317. doi: 10.1073/pnas.1220998110
Laberge, A., Arif, S., and Moulin, V. J. (2018). Microvesicles: intercellular messengers in cutaneous wound healing. J. Cell Physiol. 233, 5550–5563. doi: 10.1002/jcp.26426
Lai, R. C., Arslan, F., Lee, M. M., Sze, N. S., Choo, A., Chen, T. S., et al. (2010). Exosome secreted by MSC reduces myocardial ischemia/reperfusion injury. Stem Cell Res. 4, 214–222. doi: 10.1016/j.scr.2009.12.003
Laulagnier, K., Grand, D., Dujardin, A., Hamdi, S., Vincent-Schneider, H., Lankar, D., et al. (2004). PLD2 is enriched on exosomes and its activity is correlated to the release of exosomes. FEBS Lett. 572, 11–14. doi: 10.1016/j.febslet.2004.06.082
Lee, Y. J., Jy, W., Horstman, L. L., Janania, J., Reyes, Y., Kelley, R. E., et al. (1993). Elevated platelet microparticles in transient ischemic attacks, lacunar infarcts, and multiinfarct dementias. Thromb. Res. 72, 295–304. doi: 10.1016/0049-3848(93)90138-E
Leroyer, A. S., Isobe, H., Leseche, G., Castier, Y., Wassef, M., Mallat, Z., et al. (2007). Cellular origins and thrombogenic activity of microparticles isolated from human atherosclerotic plaques. J. Am. Coll. Cardiol. 49, 772–777. doi: 10.1016/j.jacc.2006.10.053
Lespagnol, A., Duflaut, D., Beekman, C., Blanc, L., Fiucci, G., Marine, J. C., et al. (2008). Exosome secretion, including the DNA damage-induced p53-dependent secretory pathway, is severely compromised in TSAP6/Steap3-null mice. Cell Death Differ. 15, 1723–1733. doi: 10.1038/cdd.2008.104
Letsiou, E., Sammani, S., Zhang, W., Zhou, T., Quijada, H., Moreno-Vinasco, L., et al. (2015). Pathologic mechanical stress and endotoxin exposure increases lung endothelial microparticle shedding. Am. J. Respir. Cell Mol. Biol. 52, 193–204. doi: 10.1165/rcmb.2013-0347OC
Li, G. H., Arora, P. D., Chen, Y., McCulloch, C. A., and Liu, P. (2012). Multifunctional roles of gelsolin in health and diseases. Med. Res. Rev. 32, 999–1025. doi: 10.1002/med.20231
Li, X. L., Li, H., Zhang, M., Xu, H., Yue, L. T., Zhang, X. X., et al. (2016). Exosomes derived from atorvastatin-modified bone marrow dendritic cells ameliorate experimental autoimmune myasthenia gravis by up-regulated levels of IDO/Treg and partly dependent on FasL/Fas pathway. J. Neuroinflammation 13:8. doi: 10.1186/s12974-016-0475-0
Lytvyn, Y., Xiao, F., Kennedy, C. R., Perkins, B. A., Reich, H. N., Scholey, J. W., et al. (2017). Assessment of urinary microparticles in normotensive patients with type 1 diabetes. Diabetologia 60, 581–584. doi: 10.1007/s00125-016-4190-2
Maas, S. L. N., Breakefield, X. O., and Weaver, A. M. (2017). Extracellular vesicles: unique intercellular delivery vehicles. Trends Cell Biol. 27, 172–188. doi: 10.1016/j.tcb.2016.11.003
Mallat, Z., Hugel, B., Ohan, J., Leseche, G., Freyssinet, J. M., and Tedgui, A. (1999). Shed membrane microparticles with procoagulant potential in human atherosclerotic plaques: a role for apoptosis in plaque thrombogenicity. Circulation 99, 348–353. doi: 10.1161/01.CIR.99.3.348
Matsuo, H., Chevallier, J., Mayran, N., Le Blanc, I., Ferguson, C., Faure, J., et al. (2004). Role of LBPA and Alix in multivesicular liposome formation and endosome organization. Science 303, 531–534. doi: 10.1126/science.1092425
Mazzeo, C., Calvo, V., Alonso, R., Merida, I., and Izquierdo, M. (2016). Protein kinase D1/2 is involved in the maturation of multivesicular bodies and secretion of exosomes in T and B lymphocytes. Cell Death Differ. 23, 99–109. doi: 10.1038/cdd.2015.72
Miao, Y., Li, G., Zhang, X., Xu, H., and Abraham, S. N. (2015). A TRP channel senses lysosome neutralization by pathogens to trigger their expulsion. Cell 161, 1306–1319. doi: 10.1016/j.cell.2015.05.009
Miyazaki, Y., Nomura, S., Miyake, T., Kagawa, H., Kitada, C., Taniguchi, H., et al. (1996). High shear stress can initiate both platelet aggregation and shedding of procoagulant containing microparticles. Blood 88, 3456–3464.
Miyoshi, H., Umeshita, K., Sakon, M., Imajoh-Ohmi, S., Fujitani, K., Gotoh, M., et al. (1996). Calpain activation in plasma membrane bleb formation during tert-butyl hydroperoxide-induced rat hepatocyte injury. Gastroenterology 110, 1897–1904. doi: 10.1053/gast.1996.v110.pm8964416
Mocharla, P., Briand, S., Giannotti, G., Dorries, C., Jakob, P., Paneni, F., et al. (2013). AngiomiR-126 expression and secretion from circulating CD34(+) and CD14(+) PBMCs: role for proangiogenic effects and alterations in type 2 diabetics. Blood 121, 226–236. doi: 10.1182/blood-2012-01-407106
Morel, O., Hugel, B., Jesel, L., Lanza, F., Douchet, M. P., Zupan, M., et al. (2004). Sustained elevated amounts of circulating procoagulant membrane microparticles and soluble GPV after acute myocardial infarction in diabetes mellitus. Thromb. Haemost. 91, 345–353. doi: 10.1160/TH03-05-0294
Munkonda, M. N., Akbari, S., Landry, C., Sun, S., Xiao, F., Turner, M., et al. (2018). Podocyte-derived microparticles promote proximal tubule fibrotic signaling via p38 MAPK and CD36. J. Extracell Vesicles 7:1432206. doi: 10.1080/20013078.2018.1432206
Nabhan, J. F., Hu, R., Oh, R. S., Cohen, S. N., and Lu, Q. (2012). Formation and release of arrestin domain-containing protein 1-mediated microvesicles (ARMMs) at plasma membrane by recruitment of TSG101 protein. Proc. Natl. Acad. Sci. U.S.A. 109, 4146–4151. doi: 10.1073/pnas.1200448109
Newton, J., Lima, S., Maceyka, M., and Spiegel, S. (2015). Revisiting the sphingolipid rheostat: evolving concepts in cancer therapy. Exp. Cell Res. 333, 195–200. doi: 10.1016/j.yexcr.2015.02.025
Nolte-’t Hoen, E. N., Buermans, H. P., Waasdorp, M., Stoorvogel, W., Wauben, M. H., and t Hoen, P. A. (2012). Deep sequencing of RNA from immune cell-derived vesicles uncovers the selective incorporation of small non-coding RNA biotypes with potential regulatory functions. Nucleic Acids Res. 40, 9272–9285. doi: 10.1093/nar/gks658
Nolte-’t Hoen, E. N., Buschow, S. I., Anderton, S. M., Stoorvogel, W., and Wauben, M. H. (2009). Activated T cells recruit exosomes secreted by dendritic cells via LFA-1. Blood 113, 1977–1981. doi: 10.1182/blood-2008-08-174094
Nomura, S., Inami, N., Shouzu, A., Urase, F., and Maeda, Y. (2009). Correlation and association between plasma platelet-, monocyte- and endothelial cell-derived microparticles in hypertensive patients with type 2 diabetes mellitus. Platelets 20, 406–414. doi: 10.1080/09537100903114545
Ono, Y., Saido, T. C., and Sorimachi, H. (2016). Calpain research for drug discovery: challenges and potential. Nat. Rev. Drug Discov. 15, 854–876. doi: 10.1038/nrd.2016.212
Osada-Oka, M., Shiota, M., Izumi, Y., Nishiyama, M., Tanaka, M., Yamaguchi, T., et al. (2017). Macrophage-derived exosomes induce inflammatory factors in endothelial cells under hypertensive conditions. Hypertens. Res. 40, 353–360. doi: 10.1038/hr.2016.163
Panigrahi, G. K., Praharaj, P. P., Peak, T. C., Long, J., Singh, R., Rhim, J. S., et al. (2018). Hypoxia-induced exosome secretion promotes survival of African-American and Caucasian prostate cancer cells. Sci. Rep. 8:3853. doi: 10.1038/s41598-018-22068-4
Parolini, I., Federici, C., Raggi, C., Lugini, L., Palleschi, S., De Milito, A., et al. (2009). Microenvironmental pH is a key factor for exosome traffic in tumor cells. J. Biol. Chem. 284, 34211–34222. doi: 10.1074/jbc.M109.041152
Peche, H., Heslan, M., Usal, C., Amigorena, S., and Cuturi, M. C. (2003). Presentation of donor major histocompatibility complex antigens by bone marrow dendritic cell-derived exosomes modulates allograft rejection. Transplantation 76, 1503–1510. doi: 10.1097/01.TP.0000092494.75313.38
Perez-Hernandez, D., Gutierrez-Vazquez, C., Jorge, I., Lopez-Martin, S., Ursa, A., Sanchez-Madrid, F., et al. (2013). The intracellular interactome of tetraspanin-enriched microdomains reveals their function as sorting machineries toward exosomes. J. Biol. Chem. 288, 11649–11661. doi: 10.1074/jbc.M112.445304
Piccin, A., Murphy, W. G., and Smith, O. P. (2007). Circulating microparticles: pathophysiology and clinical implications. Blood Rev. 21, 157–171. doi: 10.1016/j.blre.2006.09.001
Pironti, G., Strachan, R. T., Abraham, D., Mon-Wei Yu, S., Chen, M., Chen, W., et al. (2015). Circulating exosomes induced by cardiac pressure overload contain functional angiotensin II Type 1 receptors. Circulation 131, 2120–2130. doi: 10.1161/CIRCULATIONAHA.115.015687
Preston, R. A., Jy, W., Jimenez, J. J., Mauro, L. M., Horstman, L. L., Valle, M., et al. (2003). Effects of severe hypertension on endothelial and platelet microparticles. Hypertension 41, 211–217. doi: 10.1161/01.HYP.0000049760.15764.2D
Rahman, M. J., Regn, D., Bashratyan, R., and Dai, Y. D. (2014). Exosomes released by islet-derived mesenchymal stem cells trigger autoimmune responses in NOD mice. Diabetes Metab. Res. Rev. 63, 1008–1020. doi: 10.2337/db13-0859
Ratajczak, J., Miekus, K., Kucia, M., Zhang, J., Reca, R., Dvorak, P., et al. (2006). Embryonic stem cell-derived microvesicles reprogram hematopoietic progenitors: evidence for horizontal transfer of mRNA and protein delivery. Leukemia 20, 847–856. doi: 10.1038/sj.leu.2404132
Rautou, P. E., Vion, A. C., Amabile, N., Chironi, G., Simon, A., Tedgui, A., et al. (2011). Microparticles, vascular function, and atherothrombosis. Circ. Res. 109, 593–606. doi: 10.1161/CIRCRESAHA.110.233163
Robbins, P. D., and Morelli, A. E. (2014). Regulation of immune responses by extracellular vesicles. Nat. Rev. Immunol. 14, 195–208. doi: 10.1038/nri3622
Roucourt, B., Meeussen, S., Bao, J., Zimmermann, P., and David, G. (2015). Heparanase activates the syndecan-syntenin-ALIX exosome pathway. Cell Res. 25, 412–428. doi: 10.1038/cr.2015.29
Sabatier, F., Darmon, P., Hugel, B., Combes, V., Sanmarco, M., Velut, J. G., et al. (2002). Type 1 and type 2 diabetic patients display different patterns of cellular microparticles. Diabetes Metab. Res. Rev. 51, 2840–2845.
Safaei, R., Larson, B. J., Cheng, T. C., Gibson, M. A., Otani, S., Naerdemann, W., et al. (2005). Abnormal lysosomal trafficking and enhanced exosomal export of cisplatin in drug-resistant human ovarian carcinoma cells. Mol. Cancer Ther. 4, 1595–1604. doi: 10.1158/1535-7163.MCT-05-0102
Sahoo, S., Klychko, E., Thorne, T., Misener, S., Schultz, K. M., Millay, M., et al. (2011). Exosomes from human CD34+ stem cells mediate their proangiogenic paracrine activity. Circ. Res. 109, 724–728. doi: 10.1161/CIRCRESAHA.111.253286
Sahu, R., Kaushik, S., Clement, C. C., Cannizzo, E. S., Scharf, B., Follenzi, A., et al. (2011). Microautophagy of cytosolic proteins by late endosomes. Dev. Cell 20, 131–139. doi: 10.1016/j.devcel.2010.12.003
Sala-Valdes, M., Ursa, A., Charrin, S., Rubinstein, E., Hemler, M. E., Sanchez-Madrid, F., et al. (2006). EWI-2 and EWI-F link the tetraspanin web to the actin cytoskeleton through their direct association with ezrin-radixin-moesin proteins. J. Biol. Chem. 281, 19665–19675. doi: 10.1074/jbc.M602116200
Sapet, C., Simoncini, S., Loriod, B., Puthier, D., Sampol, J., Nguyen, C., et al. (2006). Thrombin-induced endothelial microparticle generation: identification of a novel pathway involving ROCK-II activation by caspase-2. Blood 108, 1868–1876. doi: 10.1182/blood-2006-04-014175
Sarlon-Bartoli, G., Bennis, Y., Lacroix, R., Piercecchi-Marti, M. D., Bartoli, M. A., Arnaud, L., et al. (2013). Plasmatic level of leukocyte-derived microparticles is associated with unstable plaque in asymptomatic patients with high-grade carotid stenosis. J. Am. Coll. Cardiol. 62, 1436–1441. doi: 10.1016/j.jacc.2013.03.078
Savina, A., Furlan, M., Vidal, M., and Colombo, M. I. (2003). Exosome release is regulated by a calcium-dependent mechanism in K562 cells. J. Biol. Chem. 278, 20083–20090. doi: 10.1074/jbc.M301642200
Schiro, A., Wilkinson, F. L., Weston, R., Smyth, J. V., Serracino-Inglott, F., and Alexander, M. Y. (2015). Elevated levels of endothelial-derived microparticles, and serum CXCL9 and SCGF-beta are associated with unstable asymptomatic carotid plaques. Sci. Rep. 5:16658. doi: 10.1038/srep16658
Serrano, D., Bhowmick, T., Chadha, R., Garnacho, C., and Muro, S. (2012). Intercellular adhesion molecule 1 engagement modulates sphingomyelinase and ceramide, supporting uptake of drug carriers by the vascular endothelium. Arterioscler. Thromb. Vasc. Biol. 32, 1178–1185. doi: 10.1161/ATVBAHA.111.244186
Shabbir, A., Cox, A., Rodriguez-Menocal, L., Salgado, M., and Van Badiavas, E. (2015). Mesenchymal stem cell exosomes induce proliferation and migration of normal and chronic wound fibroblasts, and enhance angiogenesis In Vitro. Stem Cells Dev. 24, 1635–1647. doi: 10.1089/scd.2014.0316
Shen, B., Wu, N., Yang, J. M., and Gould, S. J. (2011). Protein targeting to exosomes/microvesicles by plasma membrane anchors. J. Biol. Chem. 286, 14383–14395. doi: 10.1074/jbc.M110.208660
Skog, J., Wurdinger, T., van Rijn, S., Meijer, D. H., Gainche, L., Sena-Esteves, M., et al. (2008). Glioblastoma microvesicles transport RNA and proteins that promote tumour growth and provide diagnostic biomarkers. Nat. Cell Biol. 10, 1470–1476. doi: 10.1038/ncb1800
Srikanthan, S., Li, W., Silverstein, R. L., and McIntyre, T. M. (2014). Exosome poly-ubiquitin inhibits platelet activation, downregulates CD36 and inhibits pro-atherothombotic cellular functions. J. Thromb. Haemost. 12, 1906–1917. doi: 10.1111/jth.12712
Stachowiak, J. C., Schmid, E. M., Ryan, C. J., Ann, H. S., Sasaki, D. Y., Sherman, M. B., et al. (2012). Membrane bending by protein-protein crowding. Nat. Cell Biol. 14, 944–949. doi: 10.1038/ncb2561
Stepien, E., Stankiewicz, E., Zalewski, J., Godlewski, J., Zmudka, K., and Wybranska, I. (2012). Number of microparticles generated during acute myocardial infarction and stable angina correlates with platelet activation. Arch. Med. Res. 43, 31–35. doi: 10.1016/j.arcmed.2012.01.006
Stuffers, S., Sem Wegner, C., Stenmark, H., and Brech, A. (2009). Multivesicular endosome biogenesis in the absence of ESCRTs. Traffic 10, 925–937. doi: 10.1111/j.1600-0854.2009.00920.x
Suades, R., Padro, T., Crespo, J., Ramaiola, I., Martin-Yuste, V., Sabate, M., et al. (2016). Circulating microparticle signature in coronary and peripheral blood of ST elevation myocardial infarction patients in relation to pain-to-PCI elapsed time. Int. J. Cardiol. 202, 378–387. doi: 10.1016/j.ijcard.2015.09.011
Subra, C., Laulagnier, K., Perret, B., and Record, M. (2007). Exosome lipidomics unravels lipid sorting at the level of multivesicular bodies. Biochimie 89, 205–212. doi: 10.1016/j.biochi.2006.10.014
Szotowski, B., Antoniak, S., Goldin-Lang, P., Tran, Q. V., Pels, K., Rosenthal, P., et al. (2007). Antioxidative treatment inhibits the release of thrombogenic tissue factor from irradiation- and cytokine-induced endothelial cells. Cardiovasc. Res. 73, 806–812. doi: 10.1016/j.cardiores.2006.12.018
Takenouchi, T., Tsukimoto, M., Iwamaru, Y., Sugama, S., Sekiyama, K., Sato, M., et al. (2015). Extracellular ATP induces unconventional release of glyceraldehyde-3-phosphate dehydrogenase from microglial cells. Immunol. Lett. 167, 116–124. doi: 10.1016/j.imlet.2015.08.002
Tamboli, I. Y., Barth, E., Christian, L., Siepmann, M., Kumar, S., Singh, S., et al. (2010). Statins promote the degradation of extracellular amyloid {beta}-peptide by microglia via stimulation of exosome-associated insulin-degrading enzyme (IDE) secretion. J. Biol. Chem. 285, 37405–37414. doi: 10.1074/jbc.M110.149468
Tan, M., Yan, H. B., Li, J. N., Li, W. K., Fu, Y. Y., Chen, W., et al. (2016). Thrombin stimulated platelet-derived exosomes inhibit platelet-derived growth factor receptor-beta expression in vascular smooth muscle cells. Cell Physiol. Biochem. 38, 2348–2365. doi: 10.1159/000445588
Taniguchi, M., Kitatani, K., Kondo, T., Hashimoto-Nishimura, M., Asano, S., Hayashi, A., et al. (2012). Regulation of autophagy and its associated cell death by “sphingolipid rheostat”: reciprocal role of ceramide and sphingosine 1-phosphate in the mammalian target of rapamycin pathway. J. Biol. Chem. 287, 39898–39910. doi: 10.1074/jbc.M112.416552
Tkach, M., Kowal, J., and Thery, C. (2018). Why the need and how to approach the functional diversity of extracellular vesicles. Philos. Trans. R. Soc. Lond. B Biol. Sci. 373:20160479. doi: 10.1098/rstb.2016.0479
Tkach, M., and Thery, C. (2016). Communication by extracellular vesicles: where we are and where we need to go. Cell 164, 1226–1232. doi: 10.1016/j.cell.2016.01.043
Trajkovic, K., Hsu, C., Chiantia, S., Rajendran, L., Wenzel, D., Wieland, F., et al. (2008). Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science 319, 1244–1247. doi: 10.1126/science.1153124
Tramontano, A. F., Lyubarova, R., Tsiakos, J., Palaia, T., Deleon, J. R., and Ragolia, L. (2010). Circulating endothelial microparticles in diabetes mellitus. Mediators Inflamm. 2010:250476. doi: 10.1155/2010/250476
Urbanelli, L., Magini, A., Buratta, S., Brozzi, A., Sagini, K., Polchi, A., et al. (2013). Signaling pathways in exosomes biogenesis, secretion and fate. Genes 4, 152–170. doi: 10.3390/genes4020152
Valadi, H., Ekstrom, K., Bossios, A., Sjostrand, M., Lee, J. J., and Lotvall, J. O. (2007). Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 9, 654–659. doi: 10.1038/ncb1596
van Niel, G., Charrin, S., Simoes, S., Romao, M., Rochin, L., Saftig, P., et al. (2011). The tetraspanin CD63 regulates ESCRT-independent and -dependent endosomal sorting during melanogenesis. Dev. Cell 21, 708–721. doi: 10.1016/j.devcel.2011.08.019
Villarroya-Beltri, C., Baixauli, F., Mittelbrunn, M., Fernandez-Delgado, I., Torralba, D., Moreno-Gonzalo, O., et al. (2016). ISGylation controls exosome secretion by promoting lysosomal degradation of MVB proteins. Nat. Commun. 7:13588. doi: 10.1038/ncomms13588
Villarroya-Beltri, C., Gutierrez-Vazquez, C., Sanchez-Cabo, F., Perez-Hernandez, D., Vazquez, J., Martin-Cofreces, N., et al. (2013). Sumoylated hnRNPA2B1 controls the sorting of miRNAs into exosomes through binding to specific motifs. Nat. Commun. 4:2980. doi: 10.1038/ncomms3980
Wandinger-Ness, A., and Zerial, M. (2014). Rab proteins and the compartmentalization of the endosomal system. Cold Spring Harb. Perspect. Biol. 6:a022616. doi: 10.1101/cshperspect.a022616
Wang, T., Gilkes, D. M., Takano, N., Xiang, L., Luo, W., Bishop, C. J., et al. (2014). Hypoxia-inducible factors and RAB22A mediate formation of microvesicles that stimulate breast cancer invasion and metastasis. Proc. Natl. Acad. Sci. U.S.A. 111, E3234–E3242. doi: 10.1073/pnas.1410041111
Wang, X., Huang, W., Liu, G., Cai, W., Millard, R. W., Wang, Y., et al. (2014). Cardiomyocytes mediate anti-angiogenesis in type 2 diabetic rats through the exosomal transfer of miR-320 into endothelial cells. J. Mol. Cell Cardiol. 74, 139–150. doi: 10.1016/j.yjmcc.2014.05.001
Wei, Z., Batagov, A. O., Schinelli, S., Wang, J., Wang, Y., El Fatimy, R., et al. (2017). Coding and noncoding landscape of extracellular RNA released by human glioma stem cells. Nat. Commun. 8:1145. doi: 10.1038/s41467-017-01196-x
Welsch, S., Habermann, A., Jager, S., Muller, B., Krijnse-Locker, J., and Krausslich, H. G. (2006). Ultrastructural analysis of ESCRT proteins suggests a role for endosome-associated tubular-vesicular membranes in ESCRT function. Traffic 7, 1551–1566. doi: 10.1111/j.1600-0854.2006.00489.x
Wilhelm, E. N., Gonzalez-Alonso, J., Chiesa, S. T., Trangmar, S. J., Kalsi, K. K., and Rakobowchuk, M. (2017). Whole-body heat stress and exercise stimulate the appearance of platelet microvesicles in plasma with limited influence of vascular shear stress. Physiol. Rep. 5:e13496. doi: 10.14814/phy2.13496
Xiao, T., Zhang, W., Jiao, B., Pan, C. Z., Liu, X., and Shen, L. (2017). The role of exosomes in the pathogenesis of Alzheimer’ disease. Transl. Neurodegener. 6:3. doi: 10.1186/s40035-017-0072-x
Xiao, X., Xiao, F., Zhao, M., Tong, M., Wise, M. R., Stone, P. R., et al. (2017). Treating normal early gestation placentae with preeclamptic sera produces extracellular micro and nano vesicles that activate endothelial cells. J. Reprod. Immunol. 120, 34–41. doi: 10.1016/j.jri.2017.04.004
Xie, Z., Wang, X., Liu, X., Du, H., Sun, C., Shao, X., et al. (2018). Adipose-Derived exosomes exert proatherogenic effects by regulating macrophage foam cell formation and polarization. J. Am. Heart Assoc. 7:e007442. doi: 10.1161/JAHA.117.007442
Yang, J. M., and Gould, S. J. (2013). The cis-acting signals that target proteins to exosomes and microvesicles. Biochem. Soc. Trans. 41, 277–282. doi: 10.1042/BST20120275
Yang, Y., Li, Y., Chen, X., Cheng, X., Liao, Y., and Yu, X. (2016). Exosomal transfer of miR-30a between cardiomyocytes regulates autophagy after hypoxia. J. Mol. Med. 94, 711–724. doi: 10.1007/s00109-016-1387-2
Zakharova, L., Svetlova, M., and Fomina, A. F. (2007). T cell exosomes induce cholesterol accumulation in human monocytes via phosphatidylserine receptor. J. Cell. Physiol. 212, 174–181. doi: 10.1002/jcp.21013
Zhan, R., Leng, X., Liu, X., Wang, X., Gong, J., Yan, L., et al. (2009). Heat shock protein 70 is secreted from endothelial cells by a non-classical pathway involving exosomes. Biochem. Biophys. Res. Commun. 387, 229–233. doi: 10.1016/j.bbrc.2009.06.095
Zhang, B., Shi, Y., Gong, A., Pan, Z., Shi, H., Yang, H., et al. (2016). HucMSC Exosome-delivered 14-3-3zeta orchestrates self-control of the Wnt response via modulation of YAP during cutaneous regeneration. Stem Cells 34, 2485–2500. doi: 10.1002/stem.2432
Zhang, F., Tao, Y., Zhang, Z., Guo, X., An, P., Shen, Y., et al. (2012). Metalloreductase Steap3 coordinates the regulation of iron homeostasis and inflammatory responses. Haematologica 97, 1826–1835. doi: 10.3324/haematol.2012.063974
Zhang, W., Zhao, P., Xu, X. L., Cai, L., Song, Z. S., Cao, D. Y., et al. (2013). Annexin A2 promotes the migration and invasion of human hepatocellular carcinoma cells in vitro by regulating the shedding of CD147-harboring microvesicles from tumor cells. PLoS One 8:e67268. doi: 10.1371/journal.pone.0067268
Zhang, W., Zhou, X., Yao, Q., Liu, Y., Zhang, H., and Dong, Z. (2017). HIF-1-mediated production of exosomes during hypoxia is protective in renal tubular cells. Am. J. Physiol. Renal Physiol. 313, F906–F913. doi: 10.1152/ajprenal.00178.2017
Keywords: extracellular vesicles, exosomes, non-coding RNA, miRNA, heart failure, pharmacological therapy
Citation: Cesselli D, Parisse P, Aleksova A, Veneziano C, Cervellin C, Zanello A and Beltrami AP (2018) Extracellular Vesicles: How Drug and Pathology Interfere With Their Biogenesis and Function. Front. Physiol. 9:1394. doi: 10.3389/fphys.2018.01394
Received: 05 May 2018; Accepted: 13 September 2018;
Published: 01 October 2018.
Edited by:
Vincenzo Lionetti, Scuola Sant’Anna di Studi Avanzati, ItalyReviewed by:
Lucio Barile, Cardiocentro Ticino, SwitzerlandMihaela Gherghiceanu, Carol Davila University of Medicine and Pharmacy, Romania
Khaled Qanud, The Feinstein Institute for Medical Research, United States
Copyright © 2018 Cesselli, Parisse, Aleksova, Veneziano, Cervellin, Zanello and Beltrami. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Antonio Paolo Beltrami, YW50b25pby5iZWx0cmFtaUB1bml1ZC5pdA==