 Paulo R. Protachevicz1,2
Paulo R. Protachevicz1,2 Fernando S. Borges3
Fernando S. Borges3 Kelly C. Iarosz1,4,5*
Kelly C. Iarosz1,4,5* Murilo S. Baptista6
Murilo S. Baptista6 Ewandson L. Lameu7
Ewandson L. Lameu7 Matheus Hansen2,8Iberê L. Caldas1
Matheus Hansen2,8Iberê L. Caldas1 José D. Szezech Jr.2,8
José D. Szezech Jr.2,8 Antonio M. Batista1,2,8
Antonio M. Batista1,2,8 Jürgen Kurths9,10,11
Jürgen Kurths9,10,11- 1Instituto de Física, Universidade de São Paulo, São Paulo, Brazil
- 2Graduate Program in Science–Physics, State University of Ponta Grossa, Ponta Grossa, Brazil
- 3Center for Mathematics, Computation, and Cognition, Federal University of ABC, São Paulo, Brazil
- 4Faculdade de Telêmaco Borba, FATEB, Telêmaco Borba, Brazil
- 5Graduate Program in Chemical Engineering, Federal Technological University of Paraná, Ponta Grossa, Brazil
- 6Institute for Complex Systems and Mathematical Biology, SUPA, University of Aberdeen, Aberdeen, United Kingdom
- 7Cell Biology and Anatomy Department, University of Calgary, Calgary, AB, Canada
- 8Department of Mathematics and Statistics, State University of Ponta Grossa, Ponta Grossa, Brazil
- 9Department of Physics, Humboldt University, Berlin, Germany
- 10Department Complexity Science, Potsdam Institute for Climate Impact Research, Potsdam, Germany
- 11Department of Human and Animal Physiology, Saratov State University, Saratov, Russia
In the brain, the excitation-inhibition balance prevents abnormal synchronous behavior. However, known synaptic conductance intensity can be insufficient to account for the undesired synchronization. Due to this fact, we consider time delay in excitatory and inhibitory conductances and study its effect on the neuronal synchronization. In this work, we build a neuronal network composed of adaptive integrate-and-fire neurons coupled by means of delayed conductances. We observe that the time delay in the excitatory and inhibitory conductivities can alter both the state of the collective behavior (synchronous or desynchronous) and its type (spike or burst). For the weak coupling regime, we find that synchronization appears associated with neurons behaving with extremes highest and lowest mean firing frequency, in contrast to when desynchronization is present when neurons do not exhibit extreme values for the firing frequency. Synchronization can also be characterized by neurons presenting either the highest or the lowest levels in the mean synaptic current. For the strong coupling, synchronous burst activities can occur for delays in the inhibitory conductivity. For approximately equal-length delays in the excitatory and inhibitory conductances, desynchronous spikes activities are identified for both weak and strong coupling regimes. Therefore, our results show that not only the conductance intensity, but also short delays in the inhibitory conductance are relevant to avoid abnormal neuronal synchronization.
1. Introduction
Network physiology reveals how organ systems dynamically interact (Bartsch et al., 2015). The human organism is a complex physiological and integrated system in which a fail in a specific component can produce a range of biological effects (Bashan et al., 2012). One of the biggest challenges is to understand how global behavior of the human organism emerges due to local causes (Ivanov et al., 2016). Brain-brain and brain-organ networks have been considered to study integrated physiological systems under neuronal control (Ivanov et al., 2009). Chen et al. (2006) investigated the relationship between the blood flow velocities in the cerebral arteries and beat-to-beat blood pressure. Liu et al. (2015) built a network of brain wave interactions. They found complex brain dynamics, such as desynchronous and synchronous activities (Xu et al., 2006) during quiet wake and deep sleep, respectively.
Time delay has been considered in several problems of biological interest (Glass et al., 1988), such as herbivore dynamics (Sun et al., 2015), polymerization processes (Mier-y-Terán-Romero et al., 2010), dynamics of tumor growth (Byrne, 1997; Borges et al., 2014), and dynamic behavior of coupled neurons (Esfahani et al., 2016). One of the brain's intrinsic properties is the delay in the transmission of information among separate brain regions (Deco et al., 2009). Stoelzel et al. (2017) investigated the relation between axonal conduction delays and visual information. They found that some conduction times in corticothalamic axons exceed 50 ms. Conduction latencies in mammalian brain about 100 ms are also reported by Aston-Jones et al. (1985).
Dynamic brain behavior can be mimicked by means of neuronal network models (Protachevicz et al., 2019), for instance, neuronal synchronous behavior (Borges et al., 2018). Neuronal synchronization is found in task conditions (Deco et al., 2011). Furthermore, many neurological disorders are also related to synchronous behavior in the brain (Uhlhaas and Singer, 2006). Network models have been used to study the effects of time delay in synchronized neuronal activities (Stepan, 2009). Dhamala et al. (2004) showed the enhancement of neuronal synchrony by time delay in a neuronal network. Wang et al. (2016) investigated synchronized stability in coupled neurons with distributed and discrete delays. Kim and Lim (2018a,b) studied synchronization in networks, where they considered plasticity (Borges et al., 2017a) and time delays between the pre-synaptic and post-synaptic spike times.
Neurons can be modeled by differential equations. In 1907, Lapicque (Lapicque, 1907) used a linear differential equation (leaky integrate-and-fire) to simulate the neuron membrane potential. A system of non-linear differential equations was proposed by Hodgkin and Huxley (1952) to describe the action potential. The Hodgkin-Huxley model considers ion channels that open and close according to the voltage. Different connectivities among the neurons have been considered to form neuronal networks. The dynamics of coupled neurons was investigated in networks with random connections (Brunel, 2000), small-world (Tang et al., 2011), and scale-free (Batista et al., 2007, 2010) topologies were used to study neuronal synchronization.
We build here a network composed of adaptive exponential integrate-and-fire (AEIF) neurons. The AEIF model was introduced by Brette and Gerstner (2005). Depending on the parameter values, the AEIF neuron can exhibit different firing patterns (Naud et al., 2008). Synchronized firing patterns were observed in coupled AEIF neurons (Borges et al., 2017b). Pérez et al. (2011) studied the influence of conduction delays on spike synchronization in Hodgkin-Huxley neuronal networks. Previous works found that slow-rising inhibitory synaptic currents can induce synchrony (Abbott and van Vreeswijk, 1993; van Vreeswijk et al., 1994) and affect the stability of asynchronous state (e.g., splay state) (van Vreeswijk, 1996; Olmi et al., 2014). Chen et al. (2017) reported that the competition between coupling strength and synaptic time-constant leads to rich bifurcation in pulse-coupled neuronal networks with either excitatory or inhibitory synapses.
In this work, we study AEIF neurons randomly connected by means of excitatory and inhibitory conductivities. The neurons can exhibit not only spike but also burst activities (Santos et al., 2019). Our results show that the delayed conductance in both excitatory and inhibitory connections play an important role in the neuronal synchronization. Furthermore, we demonstrate that not only the values of the conductance intensity, but also small delays in inhibitory conductances are important to prevent abnormal synchronization.
The paper is organized as follows. In section 2, we introduce the neuronal network composed of AEIF neurons and delayed conductance. Section 3 shows our results about the effects of conduction delays in neuronal synchronization. We draw our conclusions in the last section.
2. Model and Methods
We construct a neuronal network with 100 AEIF neurons, where the connections are randomly chosen with probability equal to 0.5. The connection probability is defined as
where NT is the total connection number of the network and N·(N − 1) is the maximal possible number of connections for a network with N neurons without auto-connections. We consider that each neuron has at least one connection. The network has 80 and 20% of excitatory and inhibitory connections, respectively (Noback et al., 2005). The network dynamics is given by
where Vi, wi, and gi are the membrane potential, the adaptation current, and the conductance of the neuron i, respectively. We consider C = 200 pF (capacitance membrane), gL = 12 nS (leak conductance), EL = −70 mV (resting potential), Ii = 2·Irheo (constant input equal to two times the rheobase current Naud et al., 2008), ΔT = 2 mV (slope factor), VT = −50 mV (potential threshold), and τw = 300 ms (adaptation time constant). The level of subthreshold adaptation ai is randomly distributed in the interval [1.9, 2.1] nS. This set of parameters corresponds to the spike adaptation activity when neurons are uncoupled. In the model, the adaptation mechanism is able to generate burst activities when the neurons are connected by excitatory synapses (Fardet et al., 2018). For weak coupling, the neurons exhibit spike activities, while for strong, burst activities can occur for low inhibition (Protachevicz et al., 2019). The current input is calculated by the expression
where dj is the time delay in the conductance. We consider dj = dinh for inhibitory and dj = dexc for excitatory neurons. is the reversal potential (VREV = 0 mV for excitatory and VREV = −80 mV for inhibitory synapses). In the adjacency matrix (Aij), the element value is equal to 1 when the presynaptic neuron j and post-synaptic neuron i are connected, and 0 when they are not connected. gj has an exponential decay with the synaptic time constant τs = 2.728 ms. When the membrane potential of the neuron i is above a threshold (Vi > Vthres) (Naud et al., 2008), the state variables are updated according to the rules
where Vr = −58 mV is the reset potential and b = 70 pA is the triggered adaptation addition. The chemical conductance gs assumes gexc and ginh for excitatory and inhibitory neurons, respectively. We define a relative inhibitory conductance as g = ginh/gexc. Table 1 shows the standard parameter set that we use in our simulations.
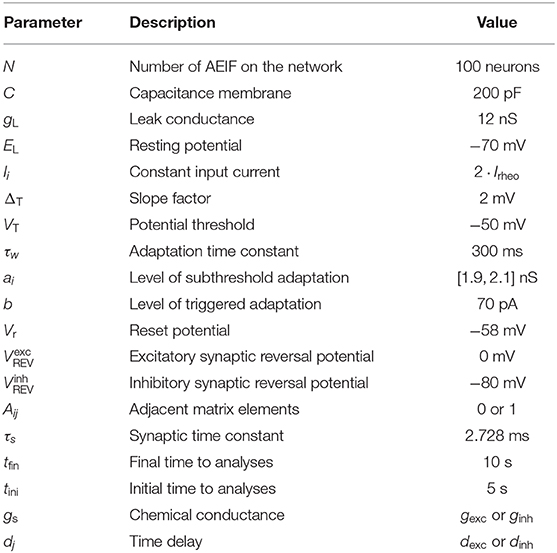
Table 1. Standard parameter set.
As a diagnostic tool to identify synchronization, we use the time average of the Kuramoto order parameter (Kuramoto, 1984; Batista et al., 2017)
where the final time in the simulation and initial time for analyses are tfin = 10 s and tini = 5 s, respectively. ranges from 0 to 1 and approaches 1 for synchronous behavior. The phase of each neuron j is calculated by Pikovsky et al. (1997)
where tj,m is the time at which neuron j suffers its m-th spike (m = 0, 1, 2, … ) and Φ is defined between two spikes in the interval [tj,m, tj,m+1].
The AEIF neuron can exhibit spike or burst activities. To identify these activities, we compute the coefficient of variation of the inter-spike interval (ISI)
where σISI and are the standard deviation and the mean value of ISI, respectively. We identify spike activities when and burst activities when (Protachevicz et al., 2018).
We calculate the mean firing frequency of the neuronal network by mean of the expression
We also compute the instantaneous Isyn(t) and the mean synaptic input (pA) of the network through
where is described by Equation (5). In all diagnostics, each point in the parameter space dinh × dexc is computed by means of the average of 10 different initial conditions. The initial conditions of Vi and wi are randomly distributed in the interval Vi = [−70, −50] mV and wi = [0, 80] nA, respectively. The initial conductance gi is equal to 0 for all neurons. To solve the delayed differential equations, we consider an initial profile of the network (for t ∈ [−dj, 0]) in which the neurons are not spiking.
3. Results
Neuronal conductances play a key role in network responses to stimuli (di Volo et al., 2019). Conduction delays were observed between the activities of the pre-synaptic and post-synaptic neurons (Ermentrout and Kopell, 1998). Figures 1A–D display , , , and , respectively, as a function of dexc for gexc = 0.2 nS, g = 6, and dinh = 5 ms. In Figures 1E–G (blue points, red points, black points), we show the raster plots for the parameters indicated by the respective filled colored circles. In Figures 1A–D, increasing dexc from 65 ms (blue) to 75 ms (red), the desynchronized spikes (Figure 1E) go to a synchronous behavior (Figure 1F), however, the spikes desynchronize when dexc is increased to 85 ms (Figure 1G). We find that a small change of the delayed conductance value can improve or suppress synchronous behavior.
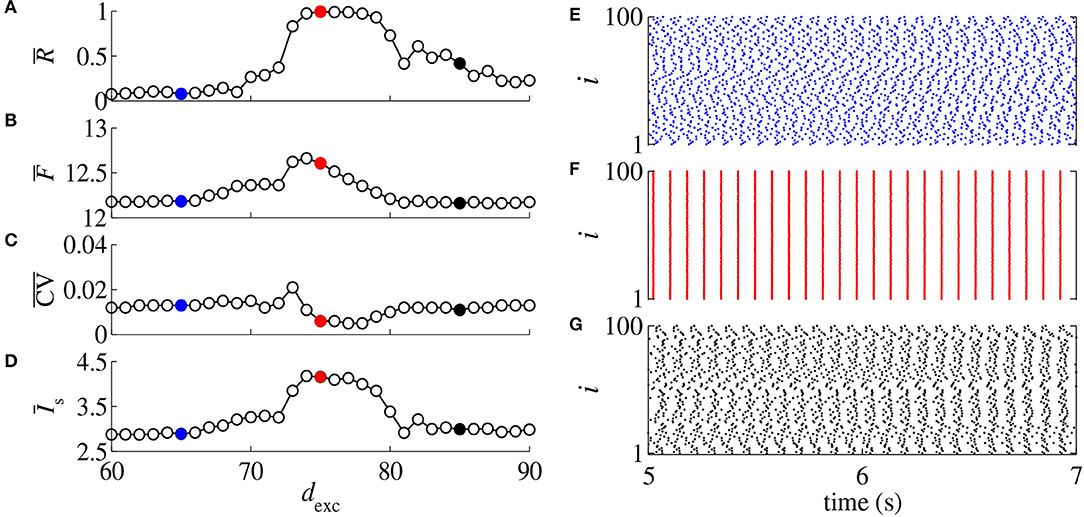
Figure 1. (A) Mean order parameter (), (B) mean firing frequency (), (C) mean coefficient of variation (), and (D) mean synaptic input () as a function of the excitatory delayed conduction dexc. Raster plots for dexc = 65 ms (E), dexc = 75 ms (F), and dexc = 85 ms (G) for gexc = 0.2 nS, g = 6, and dinh = 5 ms, and according to the colored circles.
Figures 2A,B display the parameter space dinh × dexc for gexc = 0.2 nS (weak coupling), where the color bar corresponds to the average order parameter . The parameter space exhibits synchronous (yellow region) and desynchronous (black region) spike patterns (). For g = 2 (Figure 2A), we verify vertical domains of synchronization that can be reached by maintaining dinh constant, and varying dexc. Increasing the relative inhibitory conductance for g = 6, separated domains with synchronized spikes appear, as shown in Figure 2B. For the considered parameter space, the highest and lowest values of (Figures 2C,D) and (Figures 2E,F) appear in the synchronized domain. In the domains with synchronized activities, we observe that the neuronal network achieves and maintains synchronized activities by means of changes in the mean firing frequency and synaptic current. In the region with a desynchronous pattern, the excitatory and inhibitory synaptic currents arrive in the neurons approximately at the same time.
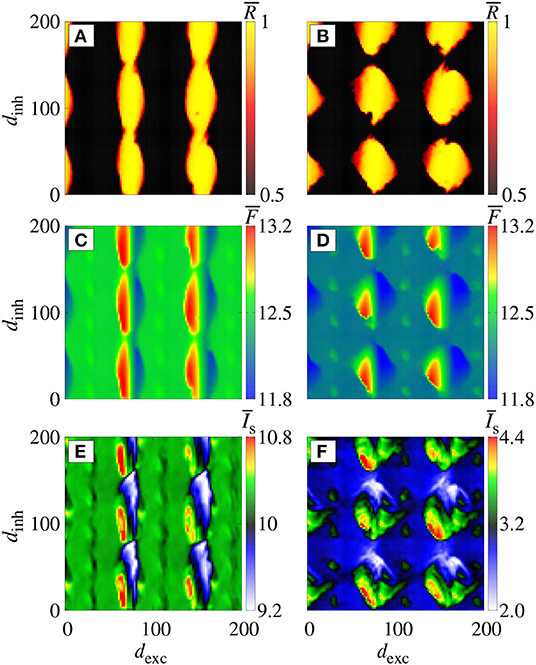
Figure 2. Colors represent , and on the parameter space dexc × dinh for gexc = 0.2 nS, where we consider g = 2 in (A,C,E), and g = 6 in (B,D,F).
Figure 3 displays magnifications of the parameter spaces shown in the right column of Figure 2 (40 ≤ dexc ≤ 110 ms and 0 ≤ dinh ≤ 70 ms). In the domain with a synchronous pattern, we observe that dexc and dinh have a significant influence on the mean firing frequency and mean synaptic current, respectively. The dynamics of neurons for some values of dexc, indicated in the vertical line (blue circles) in Figure 3 for dexc = 75 ms, are shown in Figure 4 by means of the temporal evolution of i (A,C,E) and Isyn (B,D,F), where we consider dinh = 70 ms (blue), dinh = 60 ms (red), and dinh = 10 ms (green).
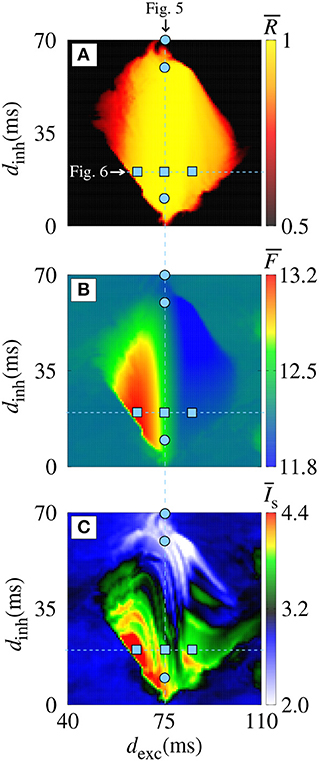
Figure 3. Magnifications of the parameter spaces shown in the right column of Figure 2. (A) , (B) , and (C) for gexc = 0.2 nS and g = 6 on the parameter space dexc × dinh.
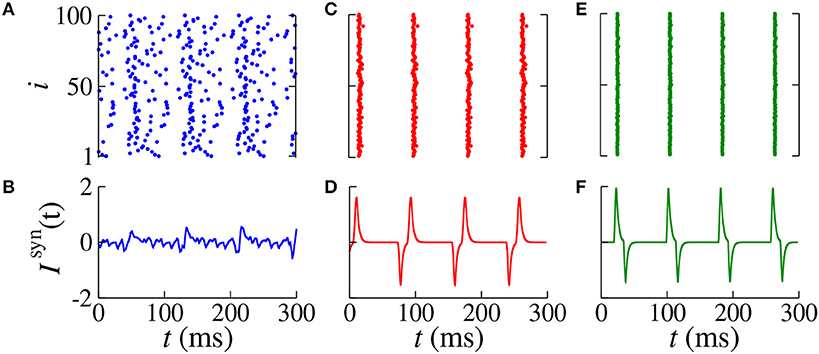
Figure 4. Raster plot (top) and Isyn(t) (bottom) for gexc = 0.2 nS, g = 6, and dexc = 75 ms for different values of dinh (blue circles in Figure 3). We consider (A,B) dinh = 70 ms (blue), (C,D) dinh = 60 ms (red), and (E,F) dinh = 10 ms (green).
In Figures 4A,B, we verify the existence of desynchronous spikes when excitatory and inhibitory inputs arrive in almost the same time (dexc ≈ dinh). Figure 5 shows raster plots (top) and Isyn(t) (bottom) for dinh = 30 ms (blue squares in Figure 3), where we consider dexc = 65 ms (blue), dexc = 75 ms (red), and dexc = 85 ms (green). The parameters correspond to the region where synchronization can occur. Furthermore, we observe that depending on the excitatory delay value, synchronization can be improved. We verify that the synchronization is improved for dexc = 75 ms, namely certain values of the delay can optimize the synchronization regime.
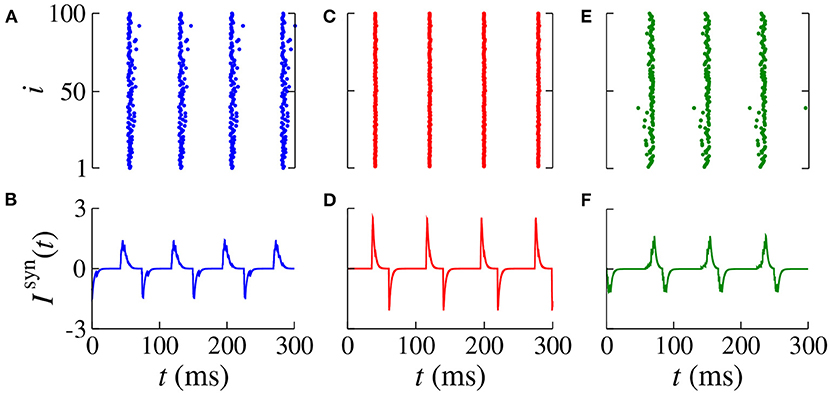
Figure 5. Raster plot (top) and Isyn(t) (bottom) for gexc = 0.2 nS, g = 6, and dinh = 30 ms for different values of dexc (blue squares in Figure 3). We consider (A,B) dexc = 65 ms (blue), (C,D) dexc = 75 ms (red), and (E,F) dexc = 85 ms (green).
Increasing gexc from 0.2 to 0.8 nS (strong coupling), in Figure 6, we observe in another range of the parameter space dinh × dexc (Figure 6A) where the region with synchronous behavior increases. Figure 6B displays the existence of regions with spike (blue) and burst (red) through the coefficient of variation value. Comparing Figures 6A,B, we verify that there are not only synchronized spikes, but also synchronized bursts. Moreover, desynchronous spike patterns are found for dinh ≈ dexc. Figures 6C,D show in color scale the values of and , respectively. We see that the synchronized spikes occur for the values of dinh and dexc in which and are low. The synchronized bursts can be found with different values of and . In addition, we also observe desynchronized activities for dinh ≈ dexc. Figure 7 displays the raster plot (top) and Isyn(t) (bottom) for gexc = 0.8 nS, g = 6, and different values of dexc and dinh, according to the parameters pointed by the symbols in Figure 6. Different delay values can generate desynchronized spikes (Figures 7A,B), synchronized bursts (Figures 7C,D,G,H), and synchronized spikes (Figures 7E,F).
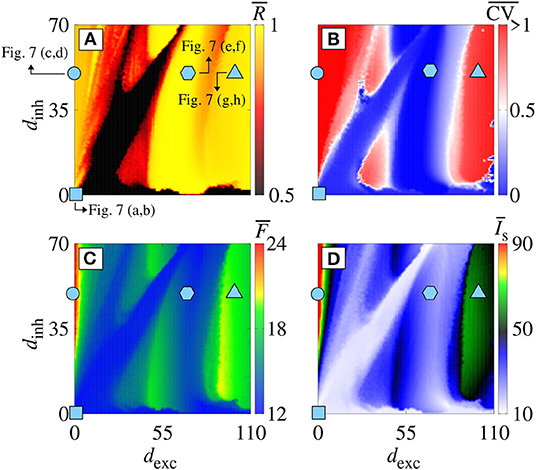
Figure 6. (A) , (B) , (C) , and (D) in the parameter space dexc × dinh for gexc = 0.8 nS and g = 6. Symbols in dexc × dinh correspond to dexc = dinh = 0 ms (cyan square), dexc = 0 ms and dinh = 50 ms (cyan circle), dexc = 70 ms and dinh = 50 ms (cyan hexagon), and dexc = 110 ms and dinh = 50 ms (cyan triangle).
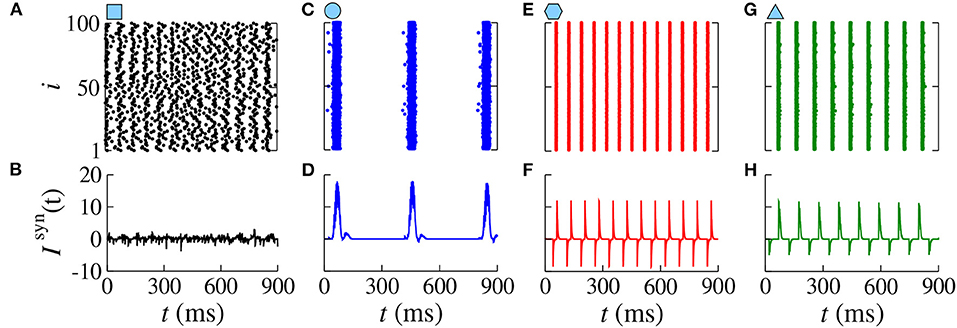
Figure 7. Raster plots (top) and Isyn(t) (bottom) for gexc = 0.8 nS, g = 6 for different values of dexc and dinh. Different delay values generate desynchronized spikes (A,B), synchronized bursts (C,D), synchronized spikes (E,F), and synchronized bursts (G,H).
4. Discussion and Conclusion
In this paper, we investigate the influence of delayed conductance on the neuronal synchronization. The study of neuronal synchronization is of great importance in neuroscience, due to the fact that it has been related to cognition, as well as to brain pathology. The conductance between the neurons plays a crucial role in the synchronous behavior. Many studies investigated the effects of the conductance on the neuronal activities (Bezanilla, 2008; Kispersky et al., 2012).
We construct a network composed of adaptive exponential integrate-and-fire (AEIF) neurons. The AEIF neuron has been used to mimic spike and burst patterns. In our network, we consider that the neurons are randomly connected by means of inhibitory and excitatory synapses. We find that for some network parameters, it is possible to observe spikes or bursts synchronization. We use the mean order parameter () and the mean coefficient of variation () as diagnostic tools to identify synchronization and spikes or bursts patterns, respectively. We also calculate and to analyse how they are related to synchronous behavior.
In order to explore the effects of different delayed conductances on the neuronal synchronization, in the section 3, we consider delay in both inhibitory and excitatory conductances. When all neurons are spiking (weak coupling), the delays induce synchronization domains in the parameter space dinh × dexc. Inside the parameter domains with synchronized neurons, we observe separated parameter subdomains representing neurons with higher and lower values of the mean firing frequency (Hz), as well as different values of the mean synaptic input (pA). For the neuronal network with strong coupling, we do not find domains with behavior similar to weak coupling in the parameter space dinh × dexc. However, we see synchronous and desynchronous activities with either spike and burst activities. We also observe a range of high values of and when only inhibitory delayed conductance is increased (dexc ≈ 0), responsible for turning desynchronous spikes into synchronous burst patterns. For dexc ≈ dinh and strong coupling, we also observed desynchronous spike activities. Desynchronous spike activities can be associated with lower mean firing frequency and synaptic currents for strong coupling.
For weak coupling, the size of the region with synchronized behavior in dinh × dexc decreases when the number of connections is decreased. In this situation, we observe that the size of the small regions can be increased by increasing gexc. In addition, for strong coupling and decreasing the number of connections, there is no burst activity and we verify the existence of synchronized and desynchronized spiking patterns, as shown for weak coupling and no sparse connectivity. Therefore, the connectivity and the synaptic conductance play an important role in the synchronization.
In conclusion, we verify that the delay in the conductances plays a crucial role in the behavior of the neurons in the neuronal network. For weak coupling, we uncover that not only the synchronous behavior, but also the mean firing frequency and the mean synaptic input depend on the delayed inhibitory and excitatory conductances. We identify which range of synaptic current allow the neuronal network to achieve and maintain synchronous activities. In the region with desynchronized activities, excitatory and inhibitory currents arrive in different times, consequently, high synchronization does not appear. For strong coupling, we see that also spike and burst patterns depend on the delayed conductances. The domain with synchronous pattern is characterized by having different delays in the inhibitory and excitatory conductances. Considering dexc ≈ dinh, we observe desynchronous spikes activities for both weak and strong coupling. In addition, our results demonstrate that not only intensity of synaptic conductance, but also a short delay in the inhibitory conductance are relevant to avoid abnormal neuronal synchronization.
Our results can be useful to clarify how synchronous and desynchronous activities are reached in a context of neuronal population with delayed conductance. In future works, we plan to analyse the influence of the connection probability between excitatory and inhibitory neurons in the neuronal synchronization, as well as the appearance of clusters synchronization.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author Contributions
PP, FB, KI, EL, and MH designed the work, developed the theory, and performed the numerical simulations. AB wrote the manuscript with support from MB, IC, JS, and JK. The authors revised the manuscript several times and gave promising suggestions. All authors also contributed to manuscript revision, read, and approved the submitted version.
Funding
This study was possible by partial financial support from the following Brazilian government agencies: Fundação Araucária, National Council for Scientific and Technological Development (CNPq) (302665/2017-0, 310124/2017-4, 428388/2018-3, 150153/2019-8), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior-Brasil (CAPES) (Finance Code No. 001), and São Paulo Research Foundation (FAPESP) (2015/07311-7, 2015/50122-0, 2016/23398-8, 2017/18977-1, 2018/03211-6, 2020/04624-2). We also wish to thank the Newton Fund and IRTG 1740/TRP 2015/50122-0, funded by the DFG/FAPESP and the project RF Goverment Grant 075-15-2019-1885.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Abbott, L. F., and van Vreeswijk, C. (1993). Asynchronous states in networks of pulse-coupled oscillators. Phys. Rev. E 48:1483. doi: 10.1103/PhysRevE.48.1483
Aston-Jones, G., Foote, S. L., and Segal, M. (1985). Impulse conduction properties of noradrenergic locus coeruleus axons projecting to monkey cerebrocortex. Neuroscience 15, 765–777. doi: 10.1016/0306-4522(85)90077-6
Bartsch, R. P., Liu, K. K. L., Bashan, A., and Ivanov, P. Ch. (2015). Network Physiology: how organ systems dynamically interact. PLoS ONE 10:e0142143. doi: 10.1371/journal.pone.0142143
Bashan, A., Bartsch, R. P., Kantelhardt, J. W., Havlin, S., and Ivanov, P. Ch. (2012). Network physiology reveals relations between network topology and physiologic function. Nat. Commun. 3, 1–9. doi: 10.1038/ncomms1705
Batista, C. A. S., Lopes, S. R., Viana, R. L., and Batista, A. M. (2010). Delayed feedback control of bursting synchronization in a scale-free neuronal network. Neural Netw. 23, 114–124. doi: 10.1016/j.neunet.2009.08.005
Batista, C. A. S., Szezech, J. D. Jr., Batista, A. M., Macau, E. E. N., and Viana, R. L. (2017). Synchronization of phase oscillators with coupling mediated by a diffusing substance. Phys. A 470, 236–248. doi: 10.1016/j.physa.2016.11.140
Batista, C. S., Batista, A. M., de Pontes, J. A., Viana, R. L., and Lopes, S. R. (2007). Chaotic phase synchronization in scale-free networks of bursting neurons. Phys. Rev. E 76:016218. doi: 10.1103/PhysRevE.76.016218
Bezanilla, F. (2008). Ion channels: from conductance to structure. Neuron 60, 456–468. doi: 10.1016/j.neuron.2008.10.035
Borges, F. S., Iarosz, K. C., Ren, H. P., Batista, A. M., Baptista, M. S., Viana, R. L., et al. (2014). Model of tumour growth with treatment by continuous and pulsed chemotherapy. Biosystems 116, 43–48. doi: 10.1016/j.biosystems.2013.12.001
Borges, F. S., Lameu, E. L., Iarosz, K. C., Protachevicz, P. R., Caldas, I. L., Viana, R. L., et al. (2018). Inference of topology and the nature of synapses, and the flow of information in neuronal networks. Phys. Rev. E 97:022303. doi: 10.1103/PhysRevE.97.022303
Borges, F. S., Protachevicz, P. R., Lameu, E. L., Bonetti, R. C., Iarosz, K. C., Caldas, I. L., et al. (2017b). Synchronised firing patterns in a random network of adaptive exponential integrate-and-fire neuron model. Neural Netw. 90, 1–7. doi: 10.1016/j.neunet.2017.03.005
Borges, R. R., Borges, F. S., Lameu, E. L., Batista, A. M., Iarosz, K. C., Caldas, I. L., et al. (2017a). Spike timing-dependent plasticity induces non-trivial topology in the brain. Neural Netw. 88, 58–64. doi: 10.1016/j.neunet.2017.01.010
Brette, R., and Gerstner, W. (2005). Adaptive exponential integrate-and-fire model as an effective description of neuronal activity. J. Neurophysiol. 94, 3637–3642. doi: 10.1152/jn.00686.2005
Brunel, N. (2000). Dynamics of networks of randomly connected excitatory and inhibitory spiking neurons. J. Physiol. 94, 445–463. doi: 10.1016/S0928-4257(00)01084-6
Byrne, H. M. (1997). The effect of time delays on the dynamics of avascular tumor growth. Math. Biosci. 144, 83–117. doi: 10.1016/S0025-5564(97)00023-0
Chen, B., Engelbrecht, J. R., and Mirollo, R. (2017). Cluster synchronization in networks of identical oscillators with α-function pulse coupling. Phys. Rev. E 95:022207. doi: 10.1103/PhysRevE.95.022207
Chen, Z., Hu, K., Stanley, H. E., Novak, V., and Ivanov, P. Ch. (2006). Cross-Correlation of instantaneous phase increments in pressure-flow fluctuations: applications to cerebral autoregulation. Phys. Rev. E 73:031915. doi: 10.1103/PhysRevE.73.031915
Deco, G., Buehlmann, A., Masquelier, T., and Hugues, E. (2011). The role of rhythmic neural synchronization in rest and task conditions. Front. Hum. Neurosci. 5:4. doi: 10.3389/fnhum.2011.00004
Deco, G., Jirsa, V., Mcintosh, A. R., Sporns, O., and Kötter, R. (2009). Key role of coupling, delay, and noise in resting brain fluctuations. Proc. Natl. Acad. Ssi. U.S.A. 106, 10302–10307. doi: 10.1073/pnas.0901831106
Dhamala, M., Jirsa, V. K., and Ding, M. (2004). Enhancement of neural synchrony by time delay. Phys. Rev. Lett. 92:074104. doi: 10.1103/PhysRevLett.92.074104
di Volo, M., Romagnoni, A., Capone, C., and Destexhe, A. (2019). Biologically realistic mean-field models of conductance-based networks of spiking neurons with adaptation. Neural Comput. 31, 653–680. doi: 10.1162/neco_a_01173
Ermentrout, G. B., and Kopell, N. (1998). Fine structure of neural spiking and synchronization in the presence of conduction delays. Proc. Natl. Acad. Ssi. U.S.A. 95, 1259–1264. doi: 10.1073/pnas.95.3.1259
Esfahani, Z. G., Gollo, L. L., and Valizadeh, A. (2016). Stimulus-dependent synchronization in delayed-coupled neuronal networks. Sci. Rep. 6:23471. doi: 10.1038/srep23471
Fardet, T., Ballandras, M., Bottani, S., Métens, S., and Monceau, P. (2018). Understanding the generation of network bursts by adaptive oscillatory neurons. Front. Neurosci. 2:41. doi: 10.3389/fnins.2018.00041
Glass, L., Beuter, A., and Larocque, D. (1988). Time delays, oscillations and chaos in physiological control systems. Math. Biosci. 90, 111–125. doi: 10.1016/0025-5564(88)90060-0
Hodgkin, A. L., and Huxley, A. F. (1952). A quantitative description of membrane curent and its application to conduction and excitation in nerve. J. Physiol. 117, 500–544. doi: 10.1113/jphysiol.1952.sp004764
Ivanov, P. C., Liu, K. K. L., and Bartsch, R. (2016). Focus on the emerging new fields of network physiology and network medicine. New J. Phys. 18:100201. doi: 10.1088/1367-2630/18/10/100201
Ivanov, P. C., Ma, Q. D. Y., and Bartsch, R. P. (2009). Maternal-fetal heartbeat phase-synchronization. Proc. Natl. Acad. Sci. U.S.A. 106, 13641–13642. doi: 10.1073/pnas.0906987106
Kim, S.-Y., and Lim, W. (2018a). Stochastic spike synchronization in a small-world neural network with spike-timing-dependent plasticity. Neural Netw. 97, 92–106. doi: 10.1016/j.neunet.2017.09.016
Kim, S.-Y., and Lim, W. (2018b). Effect of inhibitory spike-timing-dependent plasticity on fast sparsely synchronized rhythms in a small-world neuronal network. Neural Netw. 106, 50–66. doi: 10.1016/j.neunet.2018.06.013
Kispersky, T. J., Caplan, J. S., and Marder, E. (2012). Increase in sodium conductance decreases firing rate and gain in model neurons. J. Neurosci. 32, 10995–11004. doi: 10.1523/JNEUROSCI.2045-12.2012
Lapicque, L. (1907). Recherches quantitatives sur l'excitation électrique des nerfs traitée comme une polarisation. J. Physiol. Pathol. Gen. 9, 620–635.
Liu, K. K. L., Bartsch, R. P., Lin, A., Mantegna, R. N., and Ivanov, P. Ch. (2015). Plasticity of brain wave network interactions and evolution across physiologic states. Front. Neural Circuits 9:62. doi: 10.3389/fncir.2015.00062
Mier-y-Terán-Romero, L., Silber, M., and Hatzimanikatis, V. (2010). The origins of time-delay in template biopolymerization processes. PLoS Comput. Biol. 6:e1000726. doi: 10.1371/journal.pcbi.1000726
Naud, R., Marcille, N., Clopath, C., and Gerstner, W. (2008). Firing patterns in the adaptive exponential integrate-and-fire model. Biol. Cybernet. 99, 335–347. doi: 10.1007/s00422-008-0264-7
Noback, C. R., Strominger, N. L., Demarest, R. J., and Ruggiero, D. A. (2005). The Human Nervous System: Structure and Function. Totowa, NJ: Humana Press.
Olmi, S., Torcini, A., and Politi, A. (2014). Linear stability in networks of pulsed-coupled neurons. Front. Comput. Neurosci. 8:8. doi: 10.3389/fncom.2014.00008
Pérez, T., Garcia, G. C., Eguíluz, V. M., Vicente, R., Pipa, G., and Mirasso, C. (2011). Effect of the topology and delayed interactions in neuronal networks synchronization. PLoS ONE 6:e19900. doi: 10.1371/journal.pone.0019900
Pikovsky, A. S., Rosenblum, M. G., Osipov, G. V., and Kurths, J. (1997). Phase synchronization of chaotic oscillators by external driving. Phys. D 104, 219–238. doi: 10.1016/S0167-2789(96)00301-6
Protachevicz, P. R., Borges, F. S., Lameu, E. L., Ji, P., Iarosz, K. C., Kihara, A. H., et al. (2019). Bistable firing pattern in a neural network model. Front. Comput. Neurosci. 13:19. doi: 10.3389/fncom.2019.00019
Protachevicz, P. R., Borges, R. R., Reis, A. S., Borges, F. S., Iarosz, K. C., Caldas, I. L., et al. (2018). Synchronous behaviour in network model based on human cortico-cortical connections. Physiol. Meas. 39:074006. doi: 10.1088/1361-6579/aace91
Santos, M. S., Protachevicz, P. R., Iarosz, K. C., Caldas, I. L., Viana, R. L., Borges, F. S., et al. (2019). Spike-burst chimera states in an adaptive exponential integrate-and-fire neuronal network. Chaos 29:043106. doi: 10.1063/1.5087129
Stepan, G. (2009). Delay effects in brain dynamics. Philos. Trans. R. Soc. A 367, 1059–1062. doi: 10.1098/rsta.2008.0279
Stoelzel, C. R., Bereshpolova, Y., Alonso, J.-M., and Swadlow, H. A. (2017). Axonal conduction delays, brain state, and corticogenuculate communication. J. Neurosci. 37, 6342–6358. doi: 10.1523/JNEUROSCI.0444-17.2017
Sun, G.-Q., Wang, S.-L., Ren, Q., Jin, Z., and Wu, Y.-P. (2015). Effects of time delay and space on herbivere dynamics: linking inducible defenses of plants to herbivore outbreak. Sci. Rep. 5:11246. doi: 10.1038/srep11246
Tang, J., Ma, J., Yi, M., Xia, H., and Yang, X. (2011). Delay and diversity-induced synchronization transitions in a small-world neuronal network. Phys. Rev. E 83:046207. doi: 10.1103/PhysRevE.83.046207
Uhlhaas, P. J., and Singer, W. (2006). Neural synchrony in brain disorders: relevance for cognitive dysfunctions and pathophysiology. Neuron 52, 155–168. doi: 10.1016/j.neuron.2006.09.020
van Vreeswijk, C. (1996). Partial synchronization in populations of pulse-coupled oscillators. Phys. Rev. E 54:5522. doi: 10.1103/PhysRevE.54.5522
van Vreeswijk, C., Abbott, L. F., and Ermentrout, G. B. (1994). When not excitation synchronizes neural firing. J. Comput. Neurosci. 1, 313–321. doi: 10.1007/BF00961879
Wang, L., Zhao, H., and Cao, J. (2016). Synchronized bifurcation and stability in a ring of diffusively coupled neurons with time delay. Neural Netw. 75, 32–46. doi: 10.1016/j.neunet.2015.11.012
Keywords: synchronization, integrate-and-fire, neuronal network, time delay, conductance
Citation: Protachevicz PR, Borges FS, Iarosz KC, Baptista MS, Lameu EL, Hansen M, Caldas IL, Szezech JD Jr, Batista AM and Kurths J (2020) Influence of Delayed Conductance on Neuronal Synchronization. Front. Physiol. 11:1053. doi: 10.3389/fphys.2020.01053
Received: 14 April 2020; Accepted: 31 July 2020;
Published: 03 September 2020.
Edited by:
Plamen Ch. Ivanov, Boston University, United StatesReviewed by:
Bolun Chen, Brandeis University, United StatesGrigory Osipov, Lobachevsky State University of Nizhny Novgorod, Russia
Copyright © 2020 Protachevicz, Borges, Iarosz, Baptista, Lameu, Hansen, Caldas, Szezech, Batista and Kurths. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kelly C. Iarosz, a2lhcm9zekBnbWFpbC5jb20=