 A. M. Pilotto
1
†
A. M. Pilotto
1
†
 E. Higueras-Liébana
2,3,4
†
E. Higueras-Liébana
2,3,4
†
 M. Ansaldo
1
M. Ansaldo
1
 I. Baltasar-Fernandez
2,3,4,5
I. Baltasar-Fernandez
2,3,4,5
 M. Neri
1
M. Neri
1
 L. Giusti
1
L. Giusti
1
 Á. Buendía-Romero
2,3,4
Á. Buendía-Romero
2,3,4
 P. L. Valenzuela
2,3,4
P. L. Valenzuela
2,3,4
 J. Alcazar
2,3,4
J. Alcazar
2,3,4
 F. Lauretani
6,7
F. Lauretani
6,7
 R. Re
8,9
R. Re
8,9
 A. Botter
10
A. Botter
10
 M. V. Franchi
11
M. V. Franchi
11
 I. Ara
2,3,4
I. Ara
2,3,4
 S. Porcelli
1,12*
S. Porcelli
1,12*- 1 Department of Molecular Medicine, University of Pavia, Pavia, Italy
- 2 GENUD Toledo Research Group, Faculty of Sport Sciences, University of Castilla-La Mancha, Toledo, Spain
- 3 Centro de Investigación Biomédica en Red Fragilidad y Envejecimiento Saludable (CIBERFES), Instituto de Salud Carlos III, Madrid, Spain
- 4 Department of Systems Biology, University of Alcalá, Madrid, Spain
- 5 Faculty of Health Sciences, University of Castilla-La Mancha, Talavera dela Reina, Spain
- 6 Geriatric Clinic Unit, Medical Geriatric Rehabilitative Department, University Hospital of Parma, Parma, Italy
- 7 Department of Medicine and Surgery, University of Parma, Parma, Italy
- 8 Department of Physics, Politecnico di Milano, Milano, Italy
- 9 Institute for Photonics and Nanotechnologies, National Research Council, Milano, Italy
- 10 Department of Electronics and Telecommunications, Laboratory for Engineering of the Neuromuscular System (LISiN), Politecnico di Torino, Torino, Italy
- 11 Department of Biomedical Sciences, University of Padova (UNIPD), Padova, Italy
- 12 Fondazione IRCCS Policlinico San Matteo, Pavia, Italy
Aging is associated with declines in cardiorespiratory fitness and endurance performance, but this association is usually confounded by age-related declines in physical activity levels. For this reason, world-class master athletes serve as exceptional models for elucidating the limits of endurance performance in old age.
Purpose: To examine the physiological responses to exercise and performance characteristics of an 81-year-old male runner who, in 2025, set a new world record in the 50-km race (4h47m39s, 10.5 km h-1) in the 80+ category.
Methods: Two weeks after the world record, maximal O2 uptake (
Results:
Conclusion: The exceptional endurance performance of this master athlete was attributed to his well-preserved
Introduction
Endurance performance declines with age (Hawkins and Wiswell, 2003; Heath et al., 1981; Kaminsky et al., 2015; Lepers and Stapley, 2016; Tanaka and Seals, 2008; Valenzuela et al., 2020). Specifically, there is a curvilinear pattern of decline in peak endurance performance, with a modest decrease from the age of 35 to 50–60 years and more notable declines after the age of 60 (Tanaka and Seals, 2008). Among others, reductions in the maximal O2 uptake (
In the last few decades, the endurance performance of the world-class master athletes has improved more rapidly than that of their younger counterparts (Lepers and Stapley, 2016), despite the inevitable age-related performance decline (Tanaka and Seals, 2008). This trend is attributed to advances in training strategies, together with an increase in the number of master athletes competing in endurance events, but it is still debated what are the physiological determinants of the preserved endurance performance in some aged adults (Lanza et al., 2025; Marcinek and Ferrucci, 2025). Master athletes, defined as individuals older than ∼35 years who train and compete in organized competitive events, provide a unique model for studying how regular training can mitigate or delay age-related physiological decline, accounting for the confounding effect of reduced physical activity levels (Mckendry et al., 2018; Valenzuela et al., 2020). Notably, master athletes with an average age of 67 years can exhibit
In this case study, we examined the training characteristics, physiological profile and performance of a male endurance athlete who set the world record in a 50-km race in the 80+ age category in the 2025 Master Championship in Malaga (Spain). Moreover, to identify the O2 cascade profile, we tested the limiting factors of
Methods
Participant
An 81-year-old Spanish master athlete (height: 1.57 m; body mass: 58.9 kg; body mass index (BMI): 23.9 kg m-2) participated in this case study. With no prior training experience, he initiated running at the age of 66 and started competing at 70 years old in distances ranging from 800 m to 100-km ultramarathons, with a predominant focus on long-distance races. The athlete’s training was monitored during the last 12 months through the continuous recording of heart rate, training distance, and exercise intensity by a GPS-enabled heart rate monitor (Fenix 3, Garmin ltd., United States of America), previously validated (Carrier et al., 2020). This athlete is the current male marathon world champion in the 80+ age category (3 h, 39 min and 10 s in the 2024 Bucharest Marathon).
The participant volunteered to take part in the study after being informed about the procedures and potential risks. Written informed consent was obtained for each assessment session. The present study conformed to the standards set by the Declaration of Helsinki. The athlete was part of a larger project (TRAJECTOR-AGE) for examining longitudinal aging physiological decline (Lauretani et al., 2025). The study is registered at http://clinicaltrials.gov (NCT06168591) and was approved by the AVEN Ethical Committee (Emilia Romagna region, Italy) on 5 July 2022 (protocol #28022; study ID 283/2022/SPER/UNIPR).
Assessment overview
On 3 May 2025, the athlete set the world record at the Spanish 50-km Master Championship in Malaga, Spain, and we analyzed his performance during this race. Two weeks after he had achieved the 50 km world record, he attended our laboratory on 4 different occasions interspersed by at least 48 h. During the first testing session, he was interviewed about his entire sport career and training habits, hemoglobin concentration ([Hb]) was obtained from venous blood, and his body composition was assessed through dual-energy X-ray absorptiometry (DXA). On the second and third testing sessions, he performed a treadmill graded exercise test (GXT) and maximal fat oxidation (MFO) determination, respectively. On the last occasion, he performed several repetitions of moderate-intensity constant work-rate exercises (CWR), with repeated femoral artery occlusions at the end, and a cardiopulmonary cycling test (CPET).
Body composition assessment
The body composition analysis was performed using a DXA device (QDR Discovery Wi; Hologic, Bedford, MA, United States of America). All DXA scans were analyzed using Physician’s Viewer, APEX System Software Version 3.1.2. (Bedford, MA). Body composition parameters included lean mass (LM), fat mass (FM), bone mineral content (BMC) and bone mineral density (BMD) in the whole body. Regional analysis was conducted to evaluate FM and LM in upper- and lower-limbs and trunk. Daily quality control and calibration were performed with a phantom according to the manufacturer’s guidelines. Assessments were performed with the participant in a supine position, wearing light clothing free of metal, and without shoes or jewelry.
Blood sampling
Blood samples were collected after an overnight fast of at least 12 h with the participant in a seated position and in a resting condition. To determine [Hb], blood sample was drawn from an antecubital vein in 3 tubes containing ethylenediaminetetraacetic acid (EDTA) (BD Vacutainer, Stockholm, Sweden). Blood sample underwent assessment for routine clinical chemistry measurements.
Treadmill graded exercise test
The GXT was conducted during the athlete’s usual training hours, using his habitual competition footwear, having maintained his usual diet in the preceding days, avoided physical exercise for 24 h, and consumed no food in the 3 h prior to testing.
The GXT was performed on a treadmill (HP Cosmos Pulsar; H Cosmos Sports and Medical GMBH, Nussdorf Traunstein, Germany) with a slope of 1.0% to match the energy cost of running outdoors (Jones and Doust, 1996). The athlete performed a 5-min warm-up at 8 km h-1. The GXT started at the same velocity with 0.1 km h-1 increments every 10 s until exhaustion (the participant was secured through a safety harness).
Pulmonary ventilation (
Maximal aerobic speed (MAS) was defined as the velocity associated with the
Maximal fat oxidation determination and running economy
The exercise testing protocol was adapted from validated protocols previously described (Jaén-Carrillo et al., 2025; Randell et al., 2017). The test was conducted on the same treadmill used for GXT, with a slope of 1.0% to replicate the energetic cost of outdoor running (Jones and Doust, 1996). The protocol began with a 6-min warm-up at an initial speed of 5.0 km h-1. From that point onward, speed was increased by 1 km h-1 every 4 min until a RER of 1.0 was reached.
Substrate oxidation was determined by means of average gas exchange measurements during the last 60 s of each 4-min stage of the incremental protocol (Amaro-Gahete et al., 2019a). Fat oxidation rate (g∙min−1) was calculated according to Frayn’s stoichiometric equations with the assumption that urinary nitrogen excretion was 0 g (Frayn, 1983). MFO rate and the speed at which MFO occurred (Fatmax) were identified. Fat oxidation values were expressed in absolute terms (g·min-1), relative to body mass (mg·min-1·kg-1) and relative to lean mass (mg·min-1·kg-1). The relationship between fat oxidation rate and relative exercise intensity (%) was determined using a second-order polynomial curve, based on the fat oxidation value obtained at rest and all completed stages of the test. The polynomial curve was inspected by an experienced evaluator to ensure the best goodness-of-fit (i.e., R 2 > 0.70).
Running economy (RE) was derived from measurements of
Muscle oxygen uptake recovery rate constant
The muscle oxygen uptake (m
Cycling cardiopulmonary exercise test
The CPET consisted of an incremental ramp test on the cycle ergometer. Power output was increased 20 W every minute starting from the initial unloading condition. The participant was instructed to maintain constant cadence at his preferred value (∼70 rpm). Intolerance was defined when the participant could no longer maintain his chosen pedaling frequency despite verbal encouragement.
Stroke volume (SV) was estimated beat-by-beat by means of transthoracic bioimpedance cardiography (PhysioFlow, Manatec Biomedical, France) and averaged every 10 beats. The accuracy of this device has been previously evaluated during incremental exercise in healthy subjects against the direct Fick method (Richard et al., 2001). A detailed description of the method has been provided elsewhere (Charloux et al., 2000). HR was obtained from the R-R interval determined on the ECG first lead. Cardiac output (Q̇) was then calculated by multiplying SV and HR.
Oxygenation changes in the vastus lateralis muscle were evaluated continuously during the tests by NIRS (Ferrari et al., 2004). As in previous studies (see, e.g., (DeLorey et al., 2003; Ferreira et al., 2007; Ferri et al., 2007; Grassi et al., 2003; Grassi et al., 2007; Kowalchuk et al., 2002; Lanfranconi et al., 2006; Porcelli et al., 2010)) [deoxy (Hb + Mb)] was taken as an estimate of skeletal muscle fractional O2 extraction, because this variable, unlike [oxy (Hb + Mb)], is relatively insensitive to changes in blood volume (Ferrari et al., 1997; Grassi and Quaresima, 2016). Because these data are expressed in arbitrary units, a prolonged arterial occlusion was used as physiological calibration and data obtained during the exercise protocol were expressed as a percentage of the values determined by the maximal deoxygenation of the muscle, i.e., Δ[deoxy (Hb + Mb)] plateau during ischemia. For more technical details of the measurement, see Porcelli et al. (2012).
Results
50-km race performance
The athlete completed the 50-km distance in 4 h, 47 min, and 39 s, corresponding to an average speed of 10.5 km h-1 (5:44 min km-1). With this performance, he broke the men’s 50-km world record in the 80+ age category, improving the previous record by 49 min and 2 s. The former record had been held by Josef Mathias Simon of Luxembourg since 2015, with a time of 5:36:41. The slowest kilometer was completed in 6 min and 21 s (corresponding to 9.4 km h-1), whereas the fastest kilometer was run at a pace of 5:18 min km-1, corresponding to an average speed of 11.3 km h-1 (Figure 1).

Figure 1. Pacing during the Spanish 50-km Master Championship. Grey circles represent the actual speed for each kilometer. Black line identifies the 50-km average speed.
Training characteristics
The athlete reported having performed regular endurance exercise for more than 10 years. His training program followed a linear periodization model. He ran between 65 km per week in general non-competitive phases, while this volume increased up to 120 km per week in specific phases (i.e., in the ∼2 months preceding the target competition). Weekly frequency ranged from 6 to 7 sessions. Annually, the subject covered over 3,500 km.
Exercise sessions were divided into two main types: continuous training and interval training. During the general phases, the subject exclusively performed continuous training, typically at a pace ranging from 5:00 to 6:00 min km-1. The specific phase also included interval training, which initially started with 200-m intervals and progressively increased in distance throughout the training period (e.g., 400 m, 800 m, 1 km, 2 km, 4 km), reaching intervals of up to 8 km. These intervals were usually performed at a pace 5–10 s km-1 faster than the intended race pace.
Body composition
The subject’s anthropometric measurements were as follows: height 1.57 m, body mass 58.9 kg, and BMI 23.9 kg/m2. DXA analysis revealed a FM of 11.65 kg (19.5%), LM of 46.00 kg (76.8%), BMC of 2.24 kg, and BMD of 1.14 g/cm2 in the whole body. Regional analyses showed FM of 3.57 kg (18.9%) and LM of 14.45 kg in the lower limbs. In the upper limbs, FM totaled 1.34 kg (20.0%) while LM reached 5.01 kg. The trunk region contained 5.77 kg (19.1%) of FM and 23.86 kg of LM.
O2max and metabolic thresholds
From treadmill GXT, the participant showed a
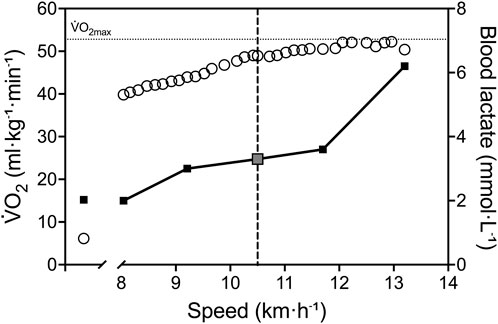
Figure 2. Physiological response during treadmill GXT. White circles represent
Maximal fat oxidation determination and running economy
His absolute fat oxidation rate at rest was 0.08 g min-1, equivalent to 1.36 mg min-1·kg-1 relative to body mass and 1.74 mg min-1·kg-1 relative to lean mass. His absolute MFO was 0.55 g min-1, corresponding to 9.34 mg min-1·kg-1 relative to body mass and 11.96 mg min-1·kg-1 relative to lean mass (Figure 3). MFO occurred at a velocity of 8 km h-1, equivalent to 61% of Vpeak and 77% of

Figure 3. Relationship between fat oxidation rate and relative exercise intensity (%) expressed as Vpeak panel (A) and
Muscle oxygen uptake recovery rate constant
Muscle oxidative capacity was characterized by a recovery rate constant k in HIGH range of 4.67 min-1 kLOW was 4.59 min-1, resulting in Δk value of 0.07.
CPET
The cycling CPET revealed a

Figure 4. Physiological response during maximal cycling CPET. White circles represent
Whole-body oxygen diffusion capacity (DO2) revealed a value of 75.3 mL min-1·mmHg-1, assuming mitochondrial partial pressure is very low pressure during maximal exercise and so could be neglected (Richardson et al., 2001).
Discussion
In this case study, we analyzed the performance and the physiological profile of an 81-year-old Spanish athlete who broke the 50-km world record for men over 80 years of age in May 2025, and currently holds first place in the marathon world championship in the 80+ age category (2024-2025). The superior endurance performance observed in this master athlete was primarily explained by a well-preserved
Although age-related performance decline is inevitable (Tanaka and Seals, 2008), largely due to reductions in
In this study we collected functional indexes of endurance performance in an octogenarian elite athlete. The incremental running test showed a very high cardiorespiratory fitness relative to his age, as indicated by a
To test the central limiting factors of
However, in the present athlete the reduction found in cardiac function was well compensated by a high value of Hb concentration, which allows for a large maximal oxygen delivery (Q̇aO2) (3.281 L min-1), higher than age-matched untrained individuals (∼2.900 mL L-1, (Capelli et al., 2025)).
Nevertheless, the unique endurance performance and the high values of
To better understand physiological adaptations in the O2 cascade from the lungs to the mitochondria, we utilized the collected parameters to reconstruct the Wagner diagram (Figure 5) through the Helsinki O2 Pathway Tool (Rissanen et al., 2024). This approach has been recently used by Goulding to demonstrate the role muscle diffusive capacity in response to sprint interval training from data collected by Mandic et al. (Goulding, 2024; Mandić et al., 2023). In Figure 5 it is possible to appreciate the unique features of our athlete in comparison to young healthy subjects. Although the
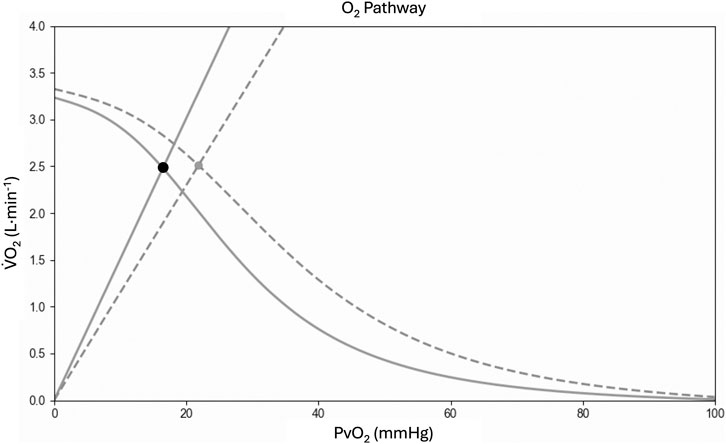
Figure 5. Wagner diagram based on cardiopulmonary cycling test data. In the graph, diagram from the present athlete (solid line) is compared with young healthy subjects from Mandic et al. (Goulding, 2024; Mandić et al., 2023) (dashed line). Fick’s principle (curved line) is conflated with Fick’s law of diffusion (straight line), with the point of intersection yielding the measured maximal oxygen uptake (
The exceptional physiological characteristics observed during exhaustion were also accompanied by unique submaximal features. The lactate threshold occurred at a velocity of 10.5 km h-1 (80% of Vpeak) and corresponded to 91% of
MFO corresponding to 0.55 g min-1 in this master athlete is comparable to normative data found in young athletic population (0.60 g min-1 for men) (Achten et al., 2003; Amaro-Gahete et al., 2019b). Furthermore, his Fatmax was found at 77% of
Among the physiological factors influencing running performance, we also evaluated O2 cost of exercise. The athlete’s running economy was lower than the typical values reported for elite younger males (39.9 mL kg-1·min-1 at 14 km h-1) and to those observed in younger recreational runners (36.7 mL kg-1·min-1 at 10 km·h-1) (Barnes and Kilding, 2015). This value contrasts the exemplary running economy value (179 mL kg-1·km-1 at 12 km h-1) demonstrated by a 70-year-old male marathon world record holder master athlete (Van Hooren and Lepers, 2023). We do not have a clear hypothesis for these differences, but it should be noted that our athlete is 10-year older than the marathon world record holder, and this gap may have affected tendon stiffness (Karamanidis and Arampatzis, 2006; Mademli and Arampatzis, 2008), resulting in a significant negative impact on running economy. Moreover, our athlete reported an average running distance of 65 km wk-1, with a maximum of 120 km wk-1 in the specific phases. In comparison, the 70-year-old male master marathon world record holder routinely ran 135–140 km wk-1 (Van Hooren and Lepers, 2023). Thus, the lower weekly distance could have led to smaller adaptations that enhanced running economy (Morgan et al., 1995). It is also important to note that the O2 cost of running was measured in fresh condition while it would have been of interest to have information about the running economy along the different segments of the 50-km race to better describe the unique performance (Zanini et al., 2024; Scheer et al., 2018). However, the race monitoring data revealed a 10% decrease in running speed, which is more pronounced than the 5% decrease observed in 40-year-old ultradistance running athletes following a 60-km ultramarathon (Schena et al., 2014).
Limitations
This study captures an informative snapshot of the athlete’s physiology close to the 50-km record. More specifically, the distance from the establishment of the new 50-km record and the tests was 2 weeks. This is the optimal timeframe for evaluating physiological characteristics in proximity to performance, while simultaneously avoiding the inclusion of detrimental acute effects associated with long-distance running. Thus, our results seem to support an outstanding physiological profile of our athlete.
Nevertheless, it should be considered that we did not have longitudinal data that allowed us to trace the developmental trajectory or identify factors that shaped the physiological adaptations underlying this performance. Moreover, in the present study we investigated the physiological determinants of
Additionally, the comparisons of physiological determinants between our athlete and data from the literature was used to help the readers in better understanding the excellence of this case report. This approach was strengthen using comparable experimental approach between the present study and previous literature (Cerezuela-Espejo et al., 2018; Jaén-Carrillo et al., 2025; Randell et al., 2017), as well as high reproducible and valid testing. However, there are differences between exercising in the laboratory setting and performing a race on the field. For example, running economy was evaluated on a 1% uphill treadmill slope to more accurately reflect the energy cost of outdoor running (Jones and Doust, 1996) but there is likely a significant difference in running economy when running on a trail vs. running the same speed on a treadmill (Sabater et al., 2023). Thus, future study should try to explore physiological limitations of performance by also collecting data during actual athletic performance.
Finally, the approach used in the present study to identify limiting factors of
Conclusion
This case report on the world-record holder for the 50-km running distance in the male 80+ age category revealed the highest
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The studies involving humans were approved by AVEN Ethical Committee (Emilia Romagna region, Italy) on 5 July 2022 (protocol #28022; study ID 283/2022/SPER/UNIPR). The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study. Written informed consent was obtained from the individual(s) for the publication of any potentially identifiable images or data included in this article.
Author contributions
AP: Methodology, Validation, Data curation, Visualization, Project administration, Supervision, Conceptualization, Funding acquisition, Resources, Investigation, Software, Writing – original draft, Formal Analysis, Writing – review and editing. EH-L: Funding acquisition, Conceptualization, Resources, Validation, Investigation, Project administration, Visualization, Supervision, Formal Analysis, Writing – review and editing, Writing – original draft, Methodology, Data curation, Software. MA: Writing – original draft, Writing – review and editing. IB-F: Formal Analysis, Project administration, Writing – original draft, Conceptualization, Supervision, Software, Data curation, Writing – review and editing, Investigation, Methodology, Visualization. MN: Methodology, Formal Analysis, Writing – original draft, Investigation, Writing – review and editing. LG: Methodology, Investigation, Writing – review and editing, Writing – original draft, Formal Analysis. ÁB-R: Formal Analysis, Methodology, Writing – original draft, Software, Writing – review and editing, Data curation, Visualization, Investigation. PV: Software, Supervision, Investigation, Writing – review and editing, Writing – original draft, Visualization. JA: Resources, Software, Writing – original draft, Project administration, Data curation, Visualization, Writing – review and editing, Conceptualization, Supervision, Investigation, Methodology. FL: Resources, Supervision, Writing – original draft, Writing – review and editing, Conceptualization, Funding acquisition. RR: Supervision, Resources, Funding acquisition, Writing – review and editing, Writing – original draft. AB: Funding acquisition, Resources, Writing – original draft, Writing – review and editing, Software, Project administration, Conceptualization, Methodology. MF: Supervision, Writing – original draft, Funding acquisition, Writing – review and editing, Investigation, Resources, Project administration, Conceptualization, Methodology. IA: Project administration, Writing – review and editing, Methodology, Supervision, Investigation, Writing – original draft. SP: Conceptualization, Project administration, Data curation, Supervision, Writing – review and editing, Methodology, Writing – original draft, Visualization, Investigation, Formal Analysis, Software.
Funding
The author(s) declared that financial support was received for this work and/or its publication. AP, FL, RR, AB, MF, SP, are supported by the Italian Ministry of University and Research (PRIN 2020477RW5) TRAJECTORAGE. EH-L, IB-F, ÁB-R, PV, IA and JA are supported by CIBERFES (CB16/10/00477), Plan Propio de Investigación of the University of Castilla-La Mancha and FEDER funds from the European Union (2025-GRIN-38408), by the Instituto de Investigación Sanitaria de Castilla-La Mancha-IDISCAM (TEC 2022-007), and by the Government of Castilla-La Mancha (SBPLY/19/180501/000312). ÁB-R is supported by a postdoctoral contract granted by Spanish Ministry of Science and Innovation (JDC 2023-052593-I, funded by MCIU/AEI/10.13039/501100011033). PLV was supported by a postdoctoral contract granted by Agencia Estatal de Investigación (RYC2024-048275-I) and University of Castilla-La Mancha and Fondo Social Europeo Plus (FSE+) (2024-UNIVERS-12850).
Conflict of interest
The author(s) declared that this work was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declared that generative AI was not used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Achten J., Venables M. C., Jeukendrup A. E. (2003). Fat oxidation rates are higher during running compared with cycling over a wide range of intensities. Metabolism 52 (6), 747–752. doi:10.1016/S0026-0495(03)00068-4
Adami A., Rossiter H. B. (2018). Principles, insights, and potential pitfalls of the noninvasive determination of muscle oxidative capacity by near-infrared spectroscopy. J. Appl. Physiology 124 (1), 245–248. doi:10.1152/japplphysiol.00445.2017
Adami A., Cao R., Porszasz J., Casaburi R., Rossiter H. B. (2017). Reproducibility of NIRS assessment of muscle oxidative capacity in smokers with and without COPD. Respir. Physiology and Neurobiol. 235, 18–26. doi:10.1016/j.resp.2016.09.008
Amaro-Gahete F. J., Sanchez-Delgado G., Alcantara J. M. A., Martinez-Tellez B., Acosta F. M., Helge J. W., et al. (2019a). Impact of data analysis methods for maximal fat oxidation estimation during exercise in sedentary adults. Eur. J. Sport Sci. 19 (9), 1230–1239. doi:10.1080/17461391.2019.1595160
Amaro-Gahete F. J., Sanchez-Delgado G., Jurado-Fasoli L., De-la-O A., Castillo M. J., Helge J. W., et al. (2019b). Assessment of maximal fat oxidation during exercise: a systematic review. Scand. J. Med. and Sci. Sports 29 (7), 910–921. doi:10.1111/sms.13424
Barnes K. R., Kilding A. E. (2015). Running economy: measurement, norms, and determining factors. Sports Med. - Open 1 (1), 8. doi:10.1186/s40798-015-0007-y
Beaudry R., Haykowsky M. J., Baggish A., La Gerche A. (2016). A modern definition of the athlete’s Heart—for research and the clinic. Cardiol. Clin. 34 (4), 507–514. doi:10.1016/j.ccl.2016.06.001
Beever A. T., Tripp T. R., Zhang J., MacInnis M. J. (2020). NIRS-derived skeletal muscle oxidative capacity is correlated with aerobic fitness and independent of sex. J. Appl. Physiology 129 (3), 558–568. doi:10.1152/japplphysiol.00017.2020
Bishop D. J., Granata C., Eynon N. (2014). Can we optimise the exercise training prescription to maximise improvements in mitochondria function and content? Biochimica Biophysica Acta 1840 (4), 1266–1275. doi:10.1016/J.BBAGEN.2013.10.012
Borg G. (1982). Ratings of perceived exertion and heart rates during short-term cycle exercise and their use in a new cycling strength test. Int. J. Sports Med. 3 (3), 153–158. doi:10.1055/s-2008-1026080
Bouckaert J., Vrijens J., Pannier J. L. (1990). Effect of specific test procedures on plasma lactate concentration and peak oxygen uptake in endurance athletes. J. Sports Med. Phys. Fit. 30 (1), 13–18.
Brizendine J. T., Ryan T. E., Larson R. D., McCully K. K. (2013). Skeletal muscle metabolism in endurance athletes with near-infrared spectroscopy. Med. and Sci. Sports and Exerc. 45 (5), 869–875. doi:10.1249/MSS.0b013e31827e0eb6
Capelli C., Ferretti G., di Prampero P. E., Tam E. (2025). Cardiovascular and peripheral factors affecting the decay of maximal oxygen uptake across the spectrum of age in humans. Eur. J. Appl. Physiology. doi:10.1007/s00421-025-06031-6
Carrick-Ranson G., Howden E. J., Brazile T. L., Levine B. D., Reading S. A. (2023). Effects of aging and endurance exercise training on cardiorespiratory fitness and cardiac structure and function in healthy midlife and older women. J. Appl. Physiology 135 (6), 1215–1235. doi:10.1152/japplphysiol.00798.2022
Carrier B., Creer A., Williams L. R., Holmes T. M., Jolley B. D., Dahl S., et al. (2020). Validation of garmin fenix 3 HR fitness tracker biomechanics and metabolics (VO2max). J. Meas. Phys. Behav. 3 (4), 331–337. doi:10.1123/jmpb.2019-0066
Cerezuela-Espejo V., Courel-Ibáñez J., Morán-Navarro R., Martínez-Cava A., Pallarés J. G. (2018). The relationship between lactate and ventilatory thresholds in runners: validity and reliability of exercise test performance parameters. Front. Physiology 9, 1320. doi:10.3389/fphys.2018.01320
Charloux A., Lonsdorfer-Wolf E., Richard R., Lampert E., Oswald-Mammosser M., Mettauer B., et al. (2000). A new impedance cardiograph device for the non-invasive evaluation of cardiac output at rest and during exercise: comparison with the “direct” fick method. Eur. J. Appl. Physiology 82 (4), 313–320. doi:10.1007/S004210000226
Coyle E. F., Hemmert M. K., Coggan A. R. (1986). Effects of detraining on cardiovascular responses to exercise: role of blood volume. J. Appl. Physiology (Bethesda, Md, 1985) 60 (1), 95–99. doi:10.1152/JAPPL.1986.60.1.95
da Mota Moreira I., Willigenburg T. R., Kregting W. J. W., Van Steijn E. R. C., Floor-Westerdijk M. J., Colier W. N. J. M. (2023). “Assessing stability and accuracy of a novel commercial wearable near-infrared spectroscopy device,” in Biophotonics in exercise science, sports medicine, health monitoring technologies, and wearables IV. Editors B. Shadgan, and A. H. Gandjbakhche (SPIE), 13. doi:10.1117/12.2657434
Dandanell S., Meinild-Lundby A., Andersen A. B., Lang P. F., Oberholzer L., Keiser S., et al. (2018). Determinants of maximal whole-body fat oxidation in elite cross-country skiers: role of skeletal muscle mitochondria. Scand. J. Med. and Sci. Sports 28 (12), 2494–2504. doi:10.1111/sms.13298
DeLorey D. S., Kowalchuk J. M., Paterson D. H. (2003). Relationship between pulmonary O2 uptake kinetics and muscle deoxygenation during moderate-intensity exercise. J. Appl. Physiology 95 (1), 113–120. doi:10.1152/japplphysiol.00956.2002
Fernhall B., Kohrt W. (1990). The effect of training specificity on maximal and submaximal physiological responses to treadmill and cycle ergometry. J. Sports Med. Phys. FitnessThe J. Sports Med. Phys. Fit. 30 (3), 268–275.
Ferrari M., Binzoni T., Quaresima V. (1997). Oxidative metabolism in muscle. Philosophical Trans. R. Soc. Lond. Ser. B Biol. Sci. 352 (1354), 677–683. doi:10.1098/rstb.1997.0049
Ferrari M., Mottola L., Quaresima V. (2004). Principles, techniques, and limitations of near infrared spectroscopy. Can. J. Appl. Physiology 29 (4), 463–487. doi:10.1139/h04-031
Ferreira L. F., Koga S., Barstow T. J. (2007). Dynamics of noninvasively estimated microvascular O2 extraction during ramp exercise. J. Appl. Physiology 103 (6), 1999–2004. doi:10.1152/japplphysiol.01414.2006
Ferri A., Adamo S., Longaretti M., Marzorati M., Lanfranconi F., Marchi A., et al. (2007). Insights into central and peripheral factors affecting the “oxidative performance” of skeletal muscle in aging. Eur. J. Appl. Physiology 100 (5), 571–579. doi:10.1007/s00421-006-0371-x
Fleg J. L., Lakatta E. G. (1988). Role of muscle loss in the age-associated reduction in VO2 max. J. Appl. Physiology 65 (3), 1147–1151. doi:10.1152/jappl.1988.65.3.1147
Foulkes S. J., Haykowsky M. J., Kistler P. M., McConell G. K., Trappe S., Hargreaves M., et al. (2024). Lifelong physiology of a former marathon world-record holder: the pros and cons of extreme cardiac remodeling. J. Appl. Physiology (Bethesda, Md, 1985) 137 (3), 461–472. doi:10.1152/japplphysiol.00070.2024
Frayn K. N. (1983). Calculation of substrate oxidation rates in vivo from gaseous exchange. J. Appl. Physiology 55 (2), 628–634. doi:10.1152/jappl.1983.55.2.628
Goulding R. P. (2024). Re-evaluating central versus peripheral contributions to maximal oxygen uptake: the role of muscle diffusive capacity. J. Physiology 602 (20), 5391–5393. doi:10.1113/JP287378
Grassi B., Quaresima V. (2016). Near-infrared spectroscopy and skeletal muscle oxidative function in vivo in health and disease: a review from an exercise physiology perspective. J. Biomed. Opt. 21 (9), 091313. doi:10.1117/1.JBO.21.9.091313
Grassi B., Pogliaghi S., Rampichini S., Quaresima V., Ferrari M., Marconi C., et al. (2003). Muscle oxygenation and pulmonary gas exchange kinetics during cycling exercise on-transitions in humans. J. Appl. Physiology 95 (1), 149–158. doi:10.1152/japplphysiol.00695.2002
Grassi B., Marzorati M., Lanfranconi F., Ferri A., Longaretti M., Stucchi A., et al. (2007). Impaired oxygen extraction in metabolic myopathies: detection and quantification by near-infrared spectroscopy. Muscle and Nerve 35 (4), 510–520. doi:10.1002/mus.20708
Grevendonk L., Connell N. J., McCrum C., Fealy C. E., Bilet L., Bruls Y. M. H., et al. (2021). Impact of aging and exercise on skeletal muscle mitochondrial capacity, energy metabolism, and physical function. Nat. Commun. 12 (1), 4773. doi:10.1038/s41467-021-24956-2
Haseler L. J., Lin A. P., Richardson R. S. (2004). Skeletal muscle oxidative metabolism in sedentary humans: 31P-MRS assessment of O2 supply and demand limitations. J. Appl. Physiology 97 (3), 1077–1081. doi:10.1152/japplphysiol.01321.2003
Hawkins S. A., Wiswell R. A. (2003). Rate and mechanism of maximal oxygen consumption decline with aging. Sports Med. 33 (12), 877–888. doi:10.2165/00007256-200333120-00002
Heath G. W., Hagberg J. M., Ehsani A. A., Holloszy J. O. (1981). A physiological comparison of young and older endurance athletes. J. Appl. Physiology 51 (3), 634–640. doi:10.1152/jappl.1981.51.3.634
Hood D. A., Memme J. M., Oliveira A. N., Triolo M. (2019). Maintenance of skeletal muscle mitochondria in health, exercise, and aging. Annu. Rev. Physiology 81 (1), 19–41. doi:10.1146/annurev-physiol-020518-114310
Jaén-Carrillo D., García-Pinillos F., Plaza-Florido A., Riquelme-Sebastián L., Fernández-Navarrete I., Ruiz-Alias S. A. (2025). Maximal fat oxidation rate in healthy young adults. Influence of cardiorespiratory fitness level and sex. Am. J. Hum. Biol. 37 (1), e24212. doi:10.1002/ajhb.24212
Jones A. M., Doust J. H. (1996). A 1% treadmill grade most accurately reflects the energetic cost of outdoor running. J. Sports Sci. 14 (4), 321–327. doi:10.1080/02640419608727717
Kaminsky L. A., Arena R., Myers J. (2015). Reference standards for cardiorespiratory fitness measured with cardiopulmonary exercise testing. Mayo Clin. Proc. 90 (11), 1515–1523. doi:10.1016/j.mayocp.2015.07.026
Karamanidis K., Arampatzis A. (2006). Mechanical and morphological properties of human quadriceps femoris and triceps surae muscle–tendon unit in relation to aging and running. J. Biomechanics 39 (3), 406–417. doi:10.1016/j.jbiomech.2004.12.017
Karlsen T., Leinan I. M., Bækkerud F. H., Lundgren K. M., Tari A., Steinshamn S. L., et al. (2015). How to be 80 year old and have a VO 2max of a 35 year old. Case Rep. Med. 2015, 1–6. doi:10.1155/2015/909561
Kokkinos P., Faselis C., Samuel I. B. H., Pittaras A., Doumas M., Murphy R., et al. (2022). Cardiorespiratory fitness and mortality risk across the spectra of age, race, and sex. J. Am. Coll. Cardiol. 80 (6), 598–609. doi:10.1016/j.jacc.2022.05.031
Kowalchuk J. M., Rossiter H. B., Ward S. A., Whipp B. J. (2002). The effect of resistive breathing on leg muscle oxygenation using near-infrared spectroscopy during exercise in men. Exp. Physiol. 87 (5), 601–611. doi:10.1113/eph8702456
Lacour J. R., Padilla-Magunacelaya S., Chatard J. C., Arsac L., Barthlmy J. C. (1991). Assessment of running velocity at maximal oxygen uptake. Eur. J. Appl. Physiology Occup. Physiology 62 (2), 77–82. doi:10.1007/BF00626760
Lakatta E. G., Levy D. (2003). Arterial and cardiac aging: major shareholders in cardiovascular disease enterprises. Circulation 107 (2), 346–354. doi:10.1161/01.CIR.0000048893.62841.F7
Lanfranconi F., Borrelli M., Ferri A., Porcelli S., Maccherini M., Chiavarelli M., et al. (2006). Noninvasive evaluation of skeletal muscle oxidative metabolism after heart transplant. Med. and Sci. Sports and Exerc. 38 (8), 1374–1383. doi:10.1249/01.mss.0000228943.62776.69
Lanza I. R., Sundberg C. W., Kent J. A. (2025). Reduced oxidative capacity of skeletal muscle IS NOT an inevitable consequence of adult ageing. J. Physiology 603 (1), 21–24. doi:10.1113/JP285042
Lauretani F., Maggio M., Pilotto A. M., Ansaldo M., Brusco C. M., Carbonaro M., et al. (2025). The trajectories of neuromuscular aging (TRAJECTOR-AGE clinical trial): study rationale and methodological protocol. J. Am. Geriatrics Soc. 73 (9), 2819–2827. doi:10.1111/jgs.70005
Lepers R., Stapley P. J. (2016). Master athletes are extending the limits of human endurance. Front. Physiology 7, 613. doi:10.3389/fphys.2016.00613
Liguori G., Feito Y., Fountaine C., Roy B. A. (2022). ACSM’s guidelines for exercise testing and prescription. Am. Coll. Sports Med.
Liu Y., Christensen P. M., Hellsten Y., Gliemann L. (2022). Effects of exercise training intensity and duration on skeletal muscle capillarization in healthy subjects: a meta-analysis. Med. Sci. Sports Exerc. 54 (10), 1714–1728. doi:10.1249/MSS.0000000000002955
Mademli L., Arampatzis A. (2008). Mechanical and morphological properties of the triceps surae muscle–tendon unit in old and young adults and their interaction with a submaximal fatiguing contraction. J. Electromyogr. Kinesiol. 18 (1), 89–98. doi:10.1016/j.jelekin.2006.09.008
Mandić M., Eriksson L. M. J., Melin M., Skott V., Sundblad P., Gustafsson T., et al. (2023). Increased maximal oxygen uptake after sprint-interval training is mediated by central haemodynamic factors as determined by right heart catheterization. J. Physiology 601 (12), 2359–2370. doi:10.1113/JP283807
Marcinek D. J., Ferrucci L. (2025). Reduced oxidative capacity of skeletal muscle mitochondria IS a fundamental consequence of adult ageing. J. Physiology 603 (1), 17–20. doi:10.1113/JP285040
Mckendry J., Breen L., Shad B. J., Greig C. A. (2018). Muscle morphology and performance in master athletes: a systematic review and meta-analyses. Ageing Res. Rev. 45, 62–82. doi:10.1016/j.arr.2018.04.007
Montero D., Lundby C. (2018). “Regulation of red blood cell volume with exercise training,” in Comprehensive physiology (Wiley), 149–164. doi:10.1002/cphy.c180004
Moreira-da-Costa M., Russo A. K., Pićarro I. C., Barros Neto T. L., Silva A. C., Tarasantchi J. (1989). Oxygen consumption and ventilation during constant-load exercise in runners and cyclists. J. Sports Med. Phys. Fit. 29 (1), 36–44.
Morgan D. W., Bransford D. R., Costill D. L., Daniels J. T., Howley E. T., Krahenbuhl G. S. (1995). Variation in the aerobic demand of running among trained and untrained subjects. Med. Sci. Sports Exerc. 27 (3), 404–409. doi:10.1249/00005768-199503000-00017
Noakes T. D., Prins P. J., Volek J. S., D’Agostino D. P., Koutnik A. P. (2023). Low carbohydrate high fat ketogenic diets on the exercise crossover point and glucose homeostasis. Front. Physiology 14, 1150265. doi:10.3389/fphys.2023.1150265
Petot H., Meilland R., Le Moyec L., Mille-Hamard L., Billat V. L. (2012). A new incremental test for VO2max accurate measurement by increasing VO2max plateau duration, allowing the investigation of its limiting factors. Eur. J. Appl. Physiology 112 (6), 2267–2276. doi:10.1007/s00421-011-2196-5
Pilotto A. M., Adami A., Mazzolari R., Brocca L., Crea E., Zuccarelli L., et al. (2022). Near-infrared spectroscopy estimation of combined skeletal muscle oxidative capacity and O 2 diffusion capacity in humans. J. Physiology 600 (18), 4153–4168. doi:10.1113/JP283267
Pollock M. L., Mengelkoch L. J., Graves J. E., Lowenthal D. T., Limacher M. C., Foster C., et al. (1997). Twenty-year follow-up of aerobic power and body composition of older track athletes. J. Appl. Physiology 82 (5), 1508–1516. doi:10.1152/jappl.1997.82.5.1508
Porcelli S., Marzorati M., Lanfranconi F., Vago P., Pisot R., Grassi B. (2010). Role of skeletal muscles impairment and brain oxygenation in limiting oxidative metabolism during exercise after bed rest. J. Appl. Physiology (Bethesda, Md, 1985) 109 (1), 101–111. doi:10.1152/japplphysiol.00782.2009
Porcelli S., Marzorati M., Pugliese L., Adamo S., Gondin J., Bottinelli R., et al. (2012). Lack of functional effects of neuromuscular electrical stimulation on skeletal muscle oxidative metabolism in healthy humans. J. Appl. Physiology 113 (7), 1101–1109. doi:10.1152/japplphysiol.01627.2011
Psatha A., Al-Mahayri Z. N., Mitropoulou C., Patrinos G. P. (2024). Meta-analysis of genomic variants in power and endurance sports to decode the impact of genomics on athletic performance and success. Hum. Genomics 18 (1), 47. doi:10.1186/s40246-024-00621-9
Randell R. K., Rollo I., Roberts T. J., Dalrymple K. J., Jeukendrup A. E., Carter J. M. (2017). Maximal fat oxidation rates in an athletic population. Med. and Sci. Sports and Exerc. 49 (1), 133–140. doi:10.1249/MSS.0000000000001084
Richard R., Lonsdorfer-Wolf E., Charloux A., Doutreleau S., Buchheit M., Oswald-Mammosser M., et al. (2001). Non-invasive cardiac output evaluation during a maximal progressive exercise test, using a new impedance cardiograph device. Eur. J. Appl. Physiology 85 (3–4), 202–207. doi:10.1007/S004210100458
Richardson R. S., Newcomer S. C., Noyszewski E. A. (2001). Skeletal muscle intracellular PO(2) assessed by myoglobin desaturation: response to graded exercise. J. Appl. Physiology 91 (6), 2679–2685. doi:10.1152/jappl.2001.91.6.2679
Rissanen A.-P. E., Mikkola T., Gagnon D. D., Lehtonen E., Lukkarinen S., Peltonen J. E. (2024). Wagner diagram for modeling O2 pathway—calculation and graphical display by the helsinki O2 pathway tool. Physiol. Meas. 45 (5), 055028. doi:10.1088/1361-6579/ad4c36
Robergs R. A., Dwyer D., Astorino T. (2010). Recommendations for improved data processing from expired gas analysis indirect calorimetry. Sports Med. 40 (2), 95–111. doi:10.2165/11319670-000000000-00000
Robinson A. T., Watso J. C., Babcock M. C., Joyner M. J., Farquhar W. B. (2019). Record-breaking performance in a 70-Year-Old marathoner. N. Engl. J. Med. 380 (15), 1485–1486. doi:10.1056/NEJMc1900771
Sabater P. F., Besson T., Berthet M., Varesco G., Kennouche D., Dandrieux P. E., et al. (2023). Elite road vs. trail runners: comparing economy, biomechanics, strength, and power. J. Strength Cond. Res. 37 (1), 181–186. doi:10.1519/JSC.0000000000004226
Scheer V., Vieluf S., Cramer L., Jakobsmeyer R., Heitkamp H.-C. (2018). Changes in running economy during a 65-km ultramarathon. Front. Physiology 9, 1809. doi:10.3389/fphys.2018.01809
Schena F., Pellegrini B., Tarperi C., Calabria E., Luca Salvagno G., Capelli C. (2014). Running economy during a simulated 60-km trial. Int. J. Sports Physiology Perform. 9 (4), 604–609. doi:10.1123/ijspp.2013-0302
Seals D. R., Hagberg J. M., Spina R. J., Rogers M. A., Schechtman K. B., Ehsani A. A. (1994). Enhanced left ventricular performance in endurance trained older men. Circulation 89 (1), 198–205. doi:10.1161/01.CIR.89.1.198
Takeshima N., Kobayashi F., Watanabe T., Tanaka K., Tomita M., Pollock M. L. (1996). Cardiorespiratory responses to cycling exercise in trained and untrained healthy elderly: with special reference to the lactate threshold. Appl. Hum. Sci. J. Physiological Anthropol. 15 (6), 267–273. doi:10.2114/jpa.15.267
Tanaka H., Seals D. R. (2008). Endurance exercise performance in masters athletes: age-associated changes and underlying physiological mechanisms. J. Physiology 586 (1), 55–63. doi:10.1113/jphysiol.2007.141879
Tanaka H., Monahan K. D., Seals D. R. (2001). Age-predicted maximal heart rate revisited. J. Am. Coll. Cardiol. 37 (1), 153–156. doi:10.1016/S0735-1097(00)01054-8
Trappe S., Hayes E., Galpin A., Kaminsky L., Jemiolo B., Fink W., et al. (2013). New records in aerobic power among octogenarian lifelong endurance athletes. J. Appl. Physiology (Bethesda, Md. 1985) 114 (1), 3–10. doi:10.1152/japplphysiol.01107.2012
Valenzuela P. L., Maffiuletti N. A., Joyner M. J., Lucia A., Lepers R. (2020). Lifelong endurance exercise as a countermeasure against age-related VO2max decline: physiological overview and insights from masters athletes. Sports Med. 50 (4), 703–716. doi:10.1007/s40279-019-01252-0
Van Hooren B., Lepers R. (2023). A physiological comparison of the new—over 70 years of age—marathon record holder and his predecessor: a case report. Front. Physiology 14, 1122315. doi:10.3389/fphys.2023.1122315
Villanova S., Pastorio E., Pilotto A. M., Marciano A., Quaresima V., Adami A., et al. (2025). Oxidative and O2 diffusive function in triceps brachii of recreational to world class swimmers. Exp. Physiol. 110, 1721–1731. doi:10.1113/EP092299
Wiswell R. A., Jaque S. V., Marcell T. J., Hawkins S. A., Tarpenning K. M., Constantino N., et al. (2000). Maximal aerobic power, lactate threshold, and running performance in master athletes. Med. and Sci. Sports and Exerc. 32 (6), 1165–1170. doi:10.1097/00005768-200006000-00021
Zanini M., Folland J. P., Blagrove R. C. (2024). Durability of running economy: differences between quantification methods and performance status in Male runners. Med. Sci. Sports Exerc 56 (11), 2230–2240. doi:10.1249/MSS.0000000000003499
Keywords: aging, maximal oxygen consumption, NIRS, running, wagner diagram
Citation: Pilotto AM, Higueras-Liébana E, Ansaldo M, Baltasar-Fernandez I, Neri M, Giusti L, Buendía-Romero Á, Valenzuela PL, Alcazar J, Lauretani F, Re R, Botter A, Franchi MV, Ara I and Porcelli S (2026) Exploring the physiological limits of aging: a case study of the male 50-km world record in the 80+ age category. Front. Physiol. 16:1735019. doi: 10.3389/fphys.2025.1735019
Received: 29 October 2025; Accepted: 12 December 2025;
Published: 12 January 2026.
Edited by:
Anna Pedrinolla, University of Trento, ItalyReviewed by:
Enrico Tam, University of Verona, ItalyDaniel Craighead, University of Minnesota Twin Cities, United States
Copyright © 2026 Pilotto, Higueras-Liébana, Ansaldo, Baltasar-Fernandez, Neri, Giusti, Buendía-Romero, Valenzuela, Alcazar, Lauretani, Re, Botter, Franchi, Ara and Porcelli. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: S. Porcelli, c2ltb25lLnBvcmNlbGxpQHVuaXB2Lml0
†These authors have contributed equally to this work