 Huan Wang1*
Huan Wang1* Yuchao Yan2Delong Li3
Yuchao Yan2Delong Li3 Xiaojun Li4,5
Xiaojun Li4,5 Xiangzhuo Liu4,6
Xiangzhuo Liu4,6 Lei Fan7
Lei Fan7 Mengjia Wang8Ying Yao1
Mengjia Wang8Ying Yao1 Yiling Cai2Shijie Yan9Zanpin Xing10Yi Zheng11Yuqing Liu4
Yiling Cai2Shijie Yan9Zanpin Xing10Yi Zheng11Yuqing Liu4 Jean-Pierre Wigneron4*
Jean-Pierre Wigneron4*- 1College of Urban and Environmental Sciences, Institute of Carbon Neutrality, Sino-French Institute for Earth System Science, Peking University, Beijing, China
- 2Guangdong Key Laboratory for Urbanization and Geo-Simulation, School of Geography and Planning, Sun Yat-sen University, Guangzhou, China
- 3Institute of Geographical Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing, China
- 4INRAE, Bordeaux Sciences Agro, UMR 1391 ISPA, Villenave-d’Ornon, France
- 5Faculty of Geosciences and Engineering, Southwest Jiaotong University, Chengdu, China
- 6Géosciences Environnement Toulouse (GET), Université de Toulouse, Toulouse, France
- 7Chongqing Jinfo Mountain Karst Ecosystem National Observation and Research Station, School of Geographical Sciences, Southwest University, Chongqing, China
- 8School of Geo-Science and Technology, Zhengzhou University, Zhengzhou, China
- 9Aerospace Information Research Institute, Chinese Academy of Sciences, Beijing, China
- 10Center for the Pan-Third Pole Environment, Lanzhou University, Lanzhou, China
- 11School of Ecology, Hainan University, Haikou, China
Global climate change and accelerating human disturbance may trigger biosphere tipping points across a range of scales and push the terrestrial ecosystem undergoing irreversible critical transitions toward alternative ecosystems. The resilience of these systems—their capacity to resist and recover from perturbations and maintain structure and function—is being eroded by multiple drivers, including land-use change, altered disturbance regimes, and biogeochemical imbalances. These drivers interact in nonlinear ways, generating cascading effects across scales and amplifying the risk of state shifts. Increasing evidence based on remote sensing time series shows that many forests are losing resilience, suggesting an early warning signal for approaching tipping points. Once tipping points are crossed, recovery is highly uncertain or even impossible on human timescales, with profound implications for biodiversity, ecosystem services, and the global carbon cycle. Understanding the mechanisms of resilience loss and identifying early-warning signals of approaching thresholds are therefore central to predicting future ecosystem stability. Due to its ability to monitor key parameters related to vegetation dynamics, remote sensing has emerged as a key tool for monitoring vegetation resilience. This can be done over large areas and with high spatial (about 10 m) and temporal (week-month) resolutions. This review synthesizes current advances on the drivers, evidence, tipping dynamics of terrestrial ecosystems in transition, and advantages of remote sensing in resilience study. We further highlight urgent action to anticipate and manage critical risks, and mitigate climate change in the Anthropocene.
Introduction
Terrestrial ecosystems are fundamental to the functioning of the Earth system, regulating carbon and water cycles, maintaining biodiversity, and sustaining human societies. Yet, the Earth is increasingly exposed to unprecedented pressure from global warming, extreme droughts, floods, deforestation and fires. Mounting evidence suggests that these ecosystems are entering a period of unprecedented instability under the combined pressures of climate change and human disturbance (Barnosky et al., 2012). Such instability raises the risk of crossing tipping points, at which a small perturbation may cause an abrupt shift in the ecosystem state (Lenton et al., 2008), accelerated by land-atmosphere positive feedbacks (Flores and Staal, 2022), beyond which recovery becomes difficult or even impossible. Nine climate tipping points (Lenton et al., 2019) have been activated and teleconnections among tipping elements may have either destabilizing or stabilizing effects on other ecosystems (Liu et al., 2023), potentially leading to cascades of abrupt transitions. Such critical transitions represent a growing concern in global change science, as they may compromise the services on which human wellbeing depends and induce substantial carbon emissions. Thus, identifying the thresholds for such transitions has been a source of growing concern of policymakers and scientists.
A central challenge lies in understanding the drivers and mechanisms that propel ecosystems toward state shifts. Climate change, land-use intensification, and biogeochemical alterations interact across scales to destabilize ecological equilibria (Smith and Boers, 2023; Wang et al., 2024). As resilience declines, ecosystems become increasingly sensitive to disturbances, and early-warning signals of transition, such as critical slowing down (Dakos et al., 2008) and rising variability, begin to emerge (Scheffer et al., 2009). At the same time, accumulating empirical evidence from tropical forests (Boulton et al., 2022), high-latitude ecosystems, and drylands demonstrates that many ecosystems are approaching the tipping points (Forzieri et al., 2022; Smith et al., 2022). Identifying the climatic tipping points and other potential thresholds that delineate the boundaries of ecosystem stability has thus become a priority for predicting future trajectories and identifying current high-risk forests.
This review synthesizes current knowledge on the multiple dimensions of terrestrial ecosystem transitions. We first examine the primary drivers that erode resilience and accelerate nonlinear responses. We then evaluate progress in detecting early-warning signals, highlighting methodological advances and persistent uncertainties. Next, we synthesize empirical evidence of ecosystems already undergoing transitions across different biomes. Finally, we assess recent estimates of climatic thresholds and tipping points, emphasizing their implications for biodiversity conservation, carbon–climate feedbacks, and Earth system stability. Together, these perspectives aim to provide an integrated framework for understanding terrestrial ecosystems in transition and to inform strategies for anticipating and managing critical risks in the Anthropocene.
Early warning signals and empirical evidence for transitions
Detecting early-warning signals is a critical step toward anticipating ecosystem transitions before they become irreversible (Scheffer et al., 2009; Scheffer et al., 2001). Theoretical and empirical studies suggest that systems approaching a tipping point exhibit statistical signatures of declining resilience, often referred to as ‘critical slowing down’ (CSD). For example, experimental studies show that imminent extinction is signaled by CSD after environmental deterioration induced by experimental decline in environmental conditions and before the critical transition (Drake and Griffen, 2010). CSD includes slower recovery rates after perturbations, increasing variance in key ecological variables, and rising temporal autocorrelation in time series data. In space, complex ecosystems evade tipping through spatial pattern formation (Rietkerk et al., 2021), in which Turing patterns (Turing, 1990) has been shown as a prominent early warning signal. That is, the spontaneous formation of regular vegetation patterns driven by self-organization in ecosystems (Rietkerk et al., 2004). Other indicators in space, such as skewness, spatial lag-1 correlation variance have also been viewed as CSD-based spatial indicators for ecosystem transitions (Rietkerk et al., 2021; Majumder et al., 2019; Eby et al., 2017). Ecological manifestations of resilience loss include declines in species diversity (Srednick and Swearer, 2024) and slow shifts in functional group dominance to track climate change (Aguirre-Gutiérrez et al., 2025). With recent advances in remote sensing, machine learning, and network-based approaches, it has become possible to detect these early-warning signals at broader spatial and temporal scales, offering new opportunities for risk assessment. Nevertheless, challenges remain in distinguishing true warning signals from natural variability, scaling up from localized observations, and integrating diverse data sources into predictive frameworks.
Earth’s ecosystems have undergone regime shifts in the past at very long timescales, and they will happen in the future. Past critical transitions happen very quickly relative to their bracketing states, less than ∼5% of the lasted time of previous states. For example, at the Cambrian explosion, both large and mobile predators became part of the food chain for the first time (Barnosky et al., 2012). During the last transition from the glacial into the present interglacial condition, spending over millennia (Wim, 2008), megafaunal biomass changed from being dominated by many species to being dominated by Homo sapiens (Barnosky, 2008). The Greenland ice sheet is melting at an accelerating rate, and it may commit the Greenland ice sheet to raising sea level by about 7 m over millennia once the threshold is crossed (Lenton et al., 2019).
Accumulating evidence indicates that many terrestrial ecosystems are approaching transitions or even already undergoing them. For example, Figure 1 shows that at the global scale, most natural ecosystems are losing resilience, indicated by lag-1 month autocorrelation. In the tropics, recurrent severe droughts, increasing tree mortality, and escalating forest fragmentation point toward a potential shift from closed-canopy rainforest to open-canopy savanna-like states (Staver et al., 2011; Van Passel et al., 2024; Lovejoy and Nobre, 2018). High-latitude ecosystems provide additional evidence of ongoing transitions: continental steppe grasslands can expand to boreal forest where soil moisture declines further along the arid timberline (Hogg and Schwarz, 1997), while positive melt-elevation feedbacks drive the central-western Greenland Ice Sheet close to a critical transition, revealed by significant early warning signals (Boers and Rypdal, 2021). Collectively, these observations reveal that ecosystem transitions are not just theoretical projections but unfolding realities, with significant implications for biodiversity, ecosystem services, and human livelihoods.
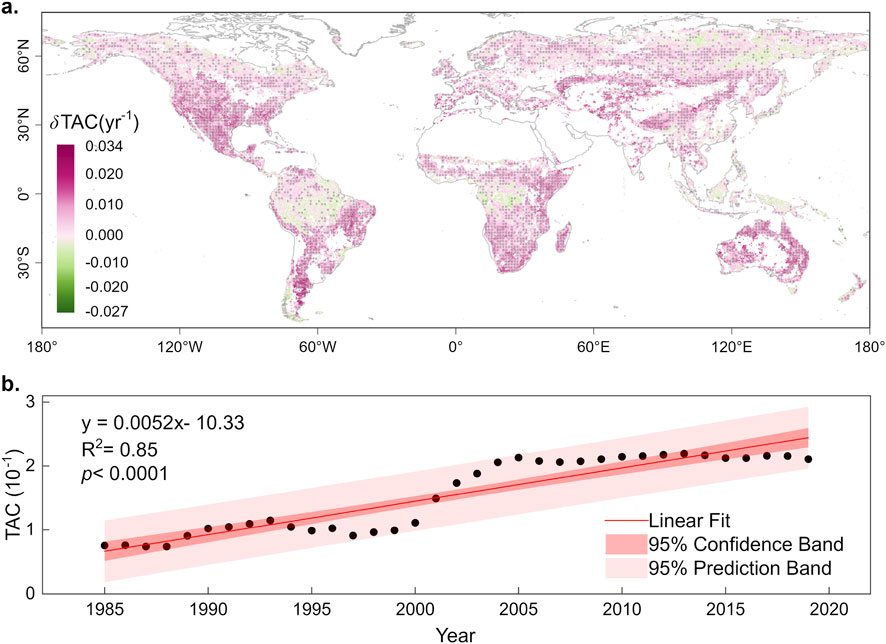
Figure 1. Temporal changes of lag-1 month autocorrelation (TAC) at global scale during 1985–2019 based on time series of remote sensing. (a) Spatial distribution of Theil–Sen slope of TAC. Positive δTAC signify declining resilience, and vice versa. (b). Temporal changes of TAC over global terrestrial ecosystems, except for croplands. Higher TAC indicates lower resilience, and vice versa. TAC is calculated from PKU GIMMS kNDVI (Li et al., 2023) based on 60-month smoothing window. Points in a suggest the temporal trends are statistically significant.
Drivers and mechanisms
Terrestrial ecosystems are increasingly impacted by multiple and interacting drivers that weaken their resilience and accelerate the likelihood of abrupt transitions. Humans now dominate Earth and change it in ways that destabilize the functions to sustain human-beings and other species (Curtis et al., 2018; Haberl et al., 2007). The shift from one state to another can be induced by a ‘threshold’, mainly driven from climate, and ‘sledgehammer’ effect, mainly driven by human land use change (Barnosky et al., 2012). The regime shift caused by threshold effect is usually difficult to simulate due to large uncertainties of Earth System Models and complex interaction between land and atmosphere. Threshold-caused regime shifts can result from ‘fold bifurcations’ and show hysteresis (Staal et al., 2020a). By contrast, a shift from a sledgehammer effect happens as no surprise, e.g., the clearing of a forest driven by agriculture. Sledgehammer effects represent abrupt, externally forced disturbances—such as deforestation, forest degradation, agricultural expansion, urbanization, and high-severity wildfires—that cause sudden structural collapse without passing through gradual climatic thresholds. Remote-sensing evidence supports their widespread occurrence: the Global Forest Change dataset (Hansen et al., 2013) reveals extensive forest loss across tropical regions; MODIS burned-area and fire-radiative-power products capture intensifying fire regimes; and global land-cover changes (Curtis et al., 2018) document rapid cropland and urban expansion. While these datasets provide robust spatial insights, uncertainties remain due to classification errors, mixed pixels, and temporal inconsistencies. Collectively, sledgehammer-type disturbances represent a dominant yet often underestimated pathway of structural ecosystem transformation, complementing the slower, climate-driven thresholds.
Climate change (Alley et al., 2003) plays a central role by rising temperatures, shifting precipitation patterns, and intensifying the frequency and severity of climatic stress from droughts (Van Passel et al., 2024; Wunderling et al., 2022), and wildfires. For example, severe drought and elevated temperatures significantly accelerated tree mortality by exacerbating hydraulic failure through increased water loss and, in Amazonian forests, by promoting high-intensity fires through drier fuels, higher fuel loads, and altered microclimatic conditions, despite some mitigating effects of elevated CO2 on carbon reserves (Duan et al., 2018; Brando et al., 2014). Rising atmospheric vapor pressure deficit (VPD), driven by long-term warming, has significantly increased tree mortality risk across tropical forests by pushing species closer to their upper hydraulic limits, even in the absence of strong drought trends in soil water deficit (Bauman et al., 2022). These changes are compounded by anthropogenic pressures (Curtis et al., 2018; Haberl et al., 2007; Sampaio et al., 2007), including deforestation, agricultural expansion, infrastructure development, and overgrazing, which disrupt natural disturbance regimes and fragment habitats. Biogeochemical perturbations, such as rising atmospheric CO2, enhanced nitrogen deposition, and altered nutrient cycles, further exacerbate stress on ecosystems by reshaping productivity and competitive interactions. Importantly, these drivers rarely act in isolation. Instead, they interact in nonlinear ways (Staal et al., 2020b), creating feedbacks across spatial and temporal scales that progressively erode ecological stability. Understanding the combined impacts of these drivers is therefore essential to explaining why terrestrial ecosystems are increasingly vulnerable to state shifts.
Plausible transition trajectories and tipping points
Climate tipping points occur when tipping elements become self-perpetuating beyond a warming threshold. Tipping elements suggest large parts of climate system, changes in which become self-perpetuating beyond a warming threshold due to positive feedbacks, leading to considerable Earth system impacts. Notably, for slow-onset tipping elements, a threshold exceedance may not cause a change of system state if the overshoot time is shorter than the effective timescale of the tipping elements, such as, the collapse of the Atlantic Meridional Overturning Circulation (AMOC) (Ritchie et al., 2021). A potential collapse of the AMOC would similarly propagate terrestrial impacts through regional climate disruptions. Cooling over the North Atlantic and Europe, along with altered precipitation patterns in West Africa and South Asia, could shorten growing seasons, reduce soil moisture, and disrupt carbon storage in forests and grasslands. Heightened droughts and extreme rainfall events may induce vegetation shifts, for example, from grassland to semi-arid states, further increasing the likelihood of abrupt ecosystem transitions.
The recent identified tipping elements (Armstrong McKay et al., 2022) and their projected global warming tipping points are given in Figure 2. Triggering climate tipping points induce significant impacts, i.e., dieback of biodiverse Amazonia forests, permafrost thawing, or substantial sea level rise. These events could cause devastating impacts on human societies. Modeling studies suggest that exceeding 1.5 °C warming above pre-industrial could trigger multiple climate tipping points (Armstrong McKay et al., 2022). Notably, current 1.1 °C warming already lies within the lower end of five uncertainty ranges of climate tipping points. Models show that abrupt collapse in Arctic sea ice occurs beyond about 3 °C–4.5 °C warming (Drijfhout et al., 2015; Hezel et al., 2014; Notz and Community, 2020). The tipping point is ∼1.5 °C warming for collapse of West Antarctic ice sheet (Lenton et al., 2008). The best estimated threshold of Atlantic meridional overturning circulation is about 4 °C warming. Widespread Amazon dieback could occur at 3.5 °C warming (Armstrong McKay et al., 2022). Regime shifts may begin in boreal forests at ∼1.5 °C and become widespread by ∼3.5 °C (Gerten et al., 2013). By contrast, warming could result in abrupt increases in plants at 2 °C to 3.5 °C warming in Eastern Sahel in three Earth System models (Drijfhout et al., 2015). These thresholds are characterized by considerable spatial heterogeneity and uncertainty, but they consistently indicate that terrestrial ecosystems are approaching tipping points with potentially irreversible consequences. Quantifying these thresholds, while accounting for interacting drivers and feedbacks, is thus critical for predicting the future of Earth’s biosphere.
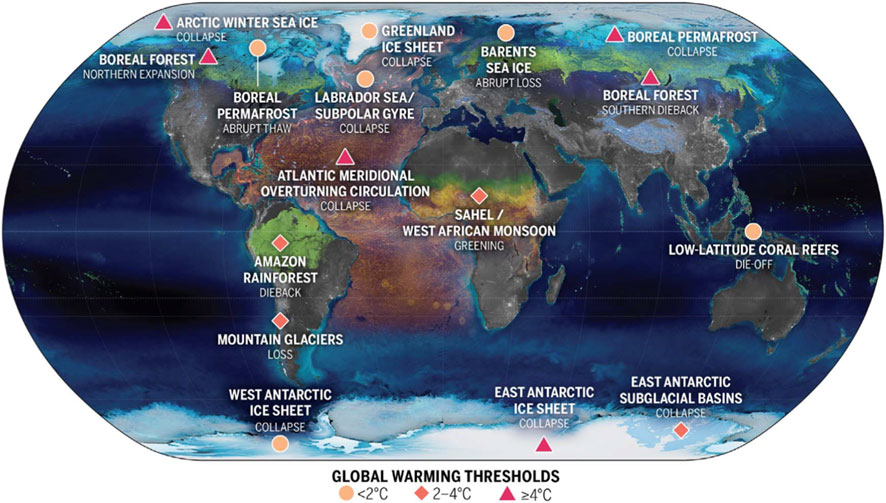
Figure 2. The location of climate tipping elements in the cryosphere, the biosphere, and ocean/atmosphere, and global warming levels at which their tipping points will likely be triggered, based on ref. (Armstrong McKay et al., 2022).
The capability of remote sensing time series on the quantification of critical slowing down
Previous research typically relies on the CSD theory and long-term observations of remote sensing data to assess changes in vegetation resilience (Forzieri et al., 2022; Smith et al., 2022). As forests approach a tipping point, CSD leads to an increase in the temporal autocorrelation coefficient and variance of the state variable. The mathematical principles are as follows:
Consider a system with periodic (
Here,
If
The autocorrelation coefficient,
When c is 0 and the mean value is 0, the variance is calculated as:
As the system approaches the critical point, the return to equilibrium slows down, with
In this study, based on the CSD theory, we utilized 60-month smoothing window to calculate the 1-lag time autocorrelation coefficients of the residual component of the kNDVI time series data from PKU GIMMS NDVI (Li et al., 2023), spanning from 1982 to 2022 with an 8 km resolution (Figure 1). The dataset’s accuracy surpasses that of its predecessor, GIMMS NDVI3g, effectively addressing issues related to orbital drift and sensor degradation in tropical regions. Data points influenced by snow, ice, or cloud cover, identified through the quality-control band and NDVI values < 0.1, were considered missing. Areas with over 75% missing data across all 16-day time steps were excluded from the analyses. In this study, bi-weekly data was aggregated into monthly values using Maximum Value Compositing to eliminate the contamination from cloud, atmosphere and changes in solar altitude angle. The original kNDVI time series are decomposed by the Seasonal and Trend decomposition using Loess (STL) method to three parts: trend, seasonal, and residual components.
It is important to note that higher values of the TAC indicate lower resilience, and vice versa. The trend in TAC variations is employed to characterize changes in resilience. A positive δTAC indicates a decreasing trend in resilience, while a negative δTAC suggests an increasing trend in resilience. The Theil-Sen slope analysis method is a non-parametric test commonly employed to estimate trends in time series data (Sen, 1968). This method is less susceptible to the influence of outliers and is considered more accurate compared to linear regression.
To derive the time series of kNDVI data, a kernel function was applied to the PKU GIMMS NDVI. kNDVI is a vegetation index derived from kernel method theory, fundamentally linearizing NDVI using the equation (Camps-Val et al., 2021):
In this equation, NIR represents the reflectance value in the near-infrared band, Red represents the reflectance value in the red band, and σ is the parameter controlling the distance between the near-infrared and red bands, simplified as σ = 0.5(NIR - Red). Thus, we have:
Discussions
We used TAC as a proxy for ecosystem resilience, based on the assumption that ecosystems near equilibrium exhibit linear recovery dynamics following perturbations. Higher TAC values indicate slower recovery and lower resilience, reflecting the phenomenon of critical slowing down as systems approach tipping points. However, the interpretation of TAC can be influenced by external drivers, non-stationary climate variability, observational noise, nonlinear dynamics, and the temporal or spatial scale of measurements. These factors can introduce potential artifacts, highlighting the need for careful consideration of method assumptions and appropriate sensitivity or robustness checks, such as detrending, or comparing across independent datasets.
The apparent tension between decreasing resilience (Forzieri et al., 2022; Smith et al., 2022) and vegetation greening (Piao et al., 2020; Zhang et al., 2025a; Zhang et al., 2025b) in some regions, e.g., arid/semi-arid, temperate, and boreal zones can be explained by multiple mechanisms. Elevated CO2, phenological shifts, and human management may drive sustained increases in vegetation productivity or leaf area, producing a greening signal, while the underlying system still recovers slowly from perturbations, leading to higher TAC. In other words, ecosystems can gain biomass while dynamical stability declines, because TAC captures intrinsic recovery near equilibrium, which can decrease even under persistent positive productivity trends. This suggests that long-term greening does not necessarily indicate enhanced resilience, and that combined analysis of TAC, productivity trends, and management factors is crucial. Future work should focus on disentangling the contributions of natural variability, anthropogenic inputs, and climate-driven changes to better predict vulnerability and guide ecosystem management under global change.
Concluding remarks
Looking forward, four priorities are essential to advance both science and policy. First, research must improve the integration of observations, models, and theory to capture the multi-scale feedbacks that govern transitions. This includes expanding long-term ecological monitoring, refining Earth system models, and developing methods that combine remote sensing, field data, and machine learning to better detect early-warning signals. Improving the ability of models to capture the known past regime shifts should increase the confidence to predict future critical transitions. Second, emphasis should be placed on quantifying thresholds and uncertainties, recognizing that ecological tipping points are context-dependent and shaped by socioeconomic drivers. Scenario-based approaches that couple ecological dynamics with human decision-making can help uncover cascading risks across systems. Third, management strategies should shift from reactive to anticipatory frameworks, focusing on resilience-building interventions, landscape connectivity, and adaptive governance to lower the risk of crossing undesirable thresholds. Fourth, satellite records are too short to provide early warnings in time (Lenton et al., 2024), yet their exceptional spatial coverage enables spatial indicators to detect resilience loss, while space-for-time substitution offers novel perspectives on potential transitions.
Looking ahead, advancing our understanding of ecosystem transitions requires moving beyond disciplinary silos toward a fully integrative framework. This entails linking empirical observations with mechanistic models and early-warning concepts to systematically identify vulnerable regions and critical thresholds, while building multi-scale monitoring networks and improving attribution of drivers and feedbacks. Embedding ecological tipping points within broader socioeconomic and policy pathways will be essential for assessing cascading risks and guiding effective responses. Ultimately, safeguarding terrestrial ecosystems in the Anthropocene will require bridging disciplinary boundaries and connecting ecological science with climate policy, conservation practice, and societal adaptation. By accelerating emissions reductions, strengthening resilience-building interventions, and enhancing our capacity to anticipate and manage critical transitions, we can lower the risk of crossing undesirable thresholds and ensure that ecosystems continue to regulate Earth’s climate, sustain biodiversity, and provide vital services for human wellbeing.
Author contributions
HW: Conceptualization, Writing – original draft, Writing – review and editing, Data curation, Formal Analysis, Methodology. YuY: Methodology, Writing – review and editing. DL: Methodology, Writing – review and editing. XoL: Methodology, Writing – review and editing. XnL: Methodology, Writing – review and editing. LF: Methodology, Writing – review and editing. MW: Data curation, Writing – review and editing. YiY: Data curation, Writing – review and editing. YC: Methodology, Writing – review and editing. SY: Methodology, Writing – review and editing. ZX: Methodology, Writing – review and editing. YZ: Data curation, Writing – review and editing. YL: Methodology, Writing – review and editing. J-PW: Supervision, Writing – review and editing.
Funding
The authors declare that financial support was received for the research and/or publication of this article. This work was supported by the National Natural Science Foundation of China [grant number: 42401106], and by the Postdoctoral Innovation Talents Support Program (grant number: BX20230017).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
The handling editor YQ declared a past co-authorship with the authors ZX, JPW, XLi, LF.
Generative AI statement
The authors declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Aguirre-Gutiérrez, J., Díaz, S., Rifai, S. W., Corral-Rivas, J. J., Nava-Miranda, M. G., González-M, R., et al. (2025). Tropical forests in the americas are changing too slowly to track climate change. Science 387, eadl5414. doi:10.1126/science.adl5414
Alley, R. B., Marotzke, J., Nordhaus, W. D., Overpeck, J. T., Peteet, D. M., Pielke, R. A., et al. (2003). Abrupt climate change. Science 299, 2005–2010. doi:10.1126/science.1081056
Armstrong McKay, D. I., Staal, A., Abrams, J. F., Winkelmann, R., Sakschewski, B., Loriani, S., et al. (2022). Exceeding 1.5°C global warming could trigger multiple climate tipping points. Science 377, eabn7950. doi:10.1126/science.abn7950
Barnosky, A. D. (2008). Megafauna biomass tradeoff as a driver of Quaternary and future extinctions. Proc. Natl. Acad. Sci. 105, 11543–11548. doi:10.1073/pnas.0801918105
Barnosky, A. D., Hadly, E. A., Bascompte, J., Berlow, E. L., Brown, J. H., Fortelius, M., et al. (2012). Approaching a state shift in Earth’s biosphere. Nature 486, 52–58. doi:10.1038/nature11018
Bauman, D., Fortunel, C., Delhaye, G., Malhi, Y., Cernusak, L. A., Bentley, L. P., et al. (2022). Tropical tree mortality has increased with rising atmospheric water stress. Nature 608, 528–533. doi:10.1038/s41586-022-04737-7
Boers, N., and Rypdal, M. (2021). Critical slowing down suggests that the Western Greenland ice sheet is close to a tipping point. Proc. Natl. Acad. Sci. U. S. A. 118, e2024192118. doi:10.1073/pnas.2024192118
Boulton, C. A., Lenton, T. M., and Boers, N. (2022). Pronounced loss of amazon rainforest resilience since the early 2000s. Nat. Clim. Change 12, 271–278. doi:10.1038/s41558-022-01287-8
Brando, P. M., Balch, J. K., Nepstad, D. C., Morton, D. C., Putz, F. E., Coe, M. T., et al. (2014). Abrupt increases in Amazonian tree mortality due to drought-fire interactions. Proc. Natl. Acad. Sci. 111, 6347–6352. doi:10.1073/pnas.1305499111
Camps-Valls, G., Campos-Taberner, M., Moreno-Martínez, Á., Walther, S., Duveiller, G., Cescatti, A., et al. (2021). A unified vegetation index for quantifying the terrestrial biosphere. Sci. Adv. 7, eabc7447. doi:10.1126/sciadv.abc7447
Curtis, P. G., Slay, C. M., Harris, N. L., Tyukavina, A., and Hansen, M. C. (2018). Classifying drivers of global forest loss. Science 361, 1108–1111. doi:10.1126/science.aau3445
Dakos, V., Scheffer, M., van Nes, E. H., Brovkin, V., Petoukhov, V., and Held, H. (2008). Slowing down as an early warning signal for abrupt climate change. Proc. Natl. Acad. Sci. 105, 14308–14312. doi:10.1073/pnas.0802430105
Drake, J. M., and Griffen, B. D. (2010). Early warning signals of extinction in deteriorating environments. Nature 467, 456–459. doi:10.1038/nature09389
Drijfhout, S., Bathiany, S., Beaulieu, C., Brovkin, V., Claussen, M., Huntingford, C., et al. (2015). Catalogue of abrupt shifts in intergovernmental panel on climate change climate models. Proc. Natl. Acad. Sci. U. S. A. 112, E5777–E5786. doi:10.1073/pnas.1511451112
Duan, H., Chaszar, B., Lewis, J. D., Smith, R. A., Huxman, T. E., and Tissue, D. T. (2018). CO2 and temperature effects on morphological and physiological traits affecting risk of drought-induced mortality. Tree Physiol. 38, 1138–1151. doi:10.1093/treephys/tpy037
Eby, S., Agrawal, A., Majumder, S., Dobson, A. P., and Guttal, V. (2017). Alternative stable states and spatial indicators of critical slowing down along a spatial gradient in a savanna ecosystem. Glob. Ecol. Biogeogr. 26, 638–649. doi:10.1111/geb.12570
Flores, B. M., and Staal, A. (2022). Feedback in tropical forests of the anthropocene. Glob. Chang. Biol. 28, 5041–5061. doi:10.1111/gcb.16293
Forzieri, G., Dakos, V., McDowell, N. G., Ramdane, A., and Cescatti, A. (2022). Emerging signals of declining forest resilience under climate change. Nature 608, 534–539. doi:10.1038/s41586-022-04959-9
Gerten, D., Lucht, W., Ostberg, S., Heinke, J., Kowarsch, M., Kreft, H., et al. (2013). Asynchronous exposure to global warming: freshwater resources and terrestrial ecosystems. Environ. Res. Lett. 8, 034032. doi:10.1088/1748-9326/8/3/034032
Haberl, H., Erb, K. H., Krausmann, F., Gaube, V., Bondeau, A., Plutzar, C., et al. (2007). Quantifying and mapping the human appropriation of net primary production in Earth's terrestrial ecosystems. Proc. Natl. Acad. Sci. 104, 12942–12947. doi:10.1073/pnas.0704243104
Hansen, M. C., Potapov, P. V., Moore, R., Hancher, M., Turubanova, S. A., Tyukavina, A., et al. (2013). High-resolution global maps of 21st-Century forest cover change. Science 342, 850–853. doi:10.1126/science.1244693
Hezel, P. J., Fichefet, T., and Massonnet, F. (2014). Modeled arctic sea ice evolution through 2300 in CMIP5 extended RCPs. Cryosphere 8, 1195–1204. doi:10.5194/tc-8-1195-2014
Hogg, E. H., and Schwarz, A. G. (1997). Regeneration of planted conifers across climatic moisture gradients on the Canadian prairies: implications for distribution and climate change. J. Biogeogr. 24, 527–534. doi:10.1111/j.1365-2699.1997.00138.x
Lenton, T. M., Held, H., Kriegler, E., Hall, J. W., Lucht, W., Rahmstorf, S., et al. (2008). Tipping elements in the earth's climate system. Proc. Natl. Acad. Sci. 105, 1786–1793. doi:10.1073/pnas.0705414105
Lenton, T. M., Rockström, J., Gaffney, O., Rahmstorf, S., Richardson, K., Steffen, W., et al. (2019). Climate tipping points - too risky to bet against. Nature 575, 592–595. doi:10.1038/d41586-019-03595-0
Lenton, T. M., Abrams, J. F., Bartsch, A., Bathiany, S., Boulton, C. A., Buxton, J. E., et al. (2024). Remotely sensing potential climate change tipping points across scales. Nat. Commun. 15, 343. doi:10.1038/s41467-023-44609-w
Li, M., Cao, S., Zhu, Z., Wang, Z., Myneni, R. B., and Piao, S. (2023). Spatiotemporally consistent global dataset of the GIMMS normalized difference vegetation index (PKU GIMMS NDVI) from 1982 to 2022. Earth Syst. Sci. Data 15, 4181–4203. doi:10.5194/essd-15-4181-2023
Liu, T., Chen, D., Yang, L., Meng, J., Wang, Z., Ludescher, J., et al. (2023). Teleconnections among tipping elements in the Earth system. Nat. Clim. Change 13, 67–74. doi:10.1038/s41558-022-01558-4
Lovejoy, T. E., and Nobre, C. (2018). Amazon tipping point. Sci. Adv. 4, eaat2340. doi:10.1126/sciadv.aat2340
Majumder, S., Tamma, K., Ramaswamy, S., and Guttal, V. (2019). Inferring critical thresholds of ecosystem transitions from spatial data. Ecology 100, e02722. doi:10.1002/ecy.2722
Notz, D., and Community, S. (2020). Arctic sea ice in CMIP6. Geophys. Res. Lett. 47, e2019GL086749. doi:10.1029/2019GL086749
Piao, S., Wang, X., Park, T., Chen, C., Lian, X., He, Y., et al. (2020). Characteristics, drivers and feedbacks of global greening. Nat. Rev. Earth and Environ. 1, 14–27. doi:10.1038/s43017-019-0001-x
Rietkerk, M., Dekker, S. C., de Ruiter, P. C., and van de Koppel, J. (2004). Self-organized patchiness and catastrophic shifts in ecosystems. Science 305, 1926–1929. doi:10.1126/science.1101867
Rietkerk, M., Bastiaansen, R., Banerjee, S., van de Koppel, J., Baudena, M., and Doelman, A. (2021). Evasion of tipping in complex systems through spatial pattern formation. Science 374, eabj0359. doi:10.1126/science.abj0359
Ritchie, P. D. L., Clarke, J. J., Cox, P. M., and Huntingford, C. (2021). Overshooting tipping point thresholds in a changing climate. Nature 592, 517–523. doi:10.1038/s41586-021-03263-2
Sampaio, G., Nobre, C., Costa, M. H., Satyamurty, P., Soares-Filho, B. S., and Cardoso, M. (2007). Regional climate change over eastern Amazonia caused by pasture and soybean cropland expansion. Geophys. Res. Lett. 34. doi:10.1029/2007GL030612
Scheffer, M., Carpenter, S., Foley, J. A., Folke, C., and Walker, B. (2001). Catastrophic shifts in ecosystems. Nature 413, 591–596. doi:10.1038/35098000
Scheffer, M., Bascompte, J., Brock, W. A., Brovkin, V., Carpenter, S. R., Dakos, V., et al. (2009). Early-warning signals for critical transitions. Nature 461, 53–59. doi:10.1038/nature08227
Smith, T., Traxl, D., and Boers, N. (2022). Empirical evidence for recent global shifts in vegetation resilience. Nat. Clim. Change 12, 477–484. doi:10.1038/s41558-022-01352-2
Smith, T., and Boers, N. (2023). Global vegetation resilience linked to water availability and variability. Nat. Commun. 14, 498. doi:10.1038/s41467-023-36207-7
Srednick, G., and Swearer, S. E. (2024). Understanding diversity–synchrony–stability relationships in multitrophic communities. Nat. Ecol. and Evol. 8, 1259–1269. doi:10.1038/s41559-024-02419-3
Staal, A., Fetzer, I., Wang-Erlandsson, L., Bosmans, J. H. C., Dekker, S. C., van Nes, E. H., et al. (2020a). Hysteresis of tropical forests in the 21st century. Nat. Commun. 11, 4978. doi:10.1038/s41467-020-18728-7
Staal, A., Flores, B. M., Aguiar, A. P. D., Bosmans, J. H. C., Fetzer, I., and Tuinenburg, O. A. (2020b). Feedback between drought and deforestation in the amazon. Environ. Res. Lett. 15, 044024. doi:10.1088/1748-9326/ab738e
Staver, A. C., Archibald, S., and Levin, S. A. (2011). The global extent and determinants of savanna and forest as alternative biome states. Science 334, 230–232. doi:10.1126/science.1210465
Turing, A. M. (1990). The chemical basis of morphogenesis. Bull. Math. Biol. 52, 153–197. doi:10.1016/s0092-8240(05)80008-4
Van Passel, J., Bernardino, P. N., Lhermitte, S., Rius, B. F., Hirota, M., Conradi, T., et al. (2024). Critical slowing down of the amazon forest after increased drought occurrence. Proc. Natl. Acad. Sci. U. S. A. 121, e2316924121. doi:10.1073/pnas.2316924121
Wang, H., Ciais, P., Sitch, S., Green, J. K., Tao, S., Fu, Z., et al. (2024). Anthropogenic disturbance exacerbates resilience loss in the amazon rainforests. Glob. Change Biol. 30, e17006. doi:10.1111/gcb.17006
Wim, Z. H. (2008). The last glacial-interglacial transition. Int. Union Geol. Sci. 31, 226–229. doi:10.18814/epiiugs/2008/v31i2/007
Wunderling, N., Staal, A., Sakschewski, B., Hirota, M., Tuinenburg, O. A., Donges, J. F., et al. (2022). Recurrent droughts increase risk of cascading tipping events by outpacing adaptive capacities in the amazon rainforest. Proc. Natl. Acad. Sci. U. S. A. 119, e2120777119. doi:10.1073/pnas.2120777119
Zhang, Y., Mao, J., Sun, G., Guo, Q., Atkins, J., Li, W., et al. (2025a). Earth's record-high greenness and its attributions in 2020. Remote Sens. Environ. 316, 114494. doi:10.1016/j.rse.2024.114494
Keywords: resilience loss, critical transition, tipping points, remote sensing, terrestrial ecosystem
Citation: Wang H, Yan Y, Li D, Li X, Liu X, Fan L, Wang M, Yao Y, Cai Y, Yan S, Xing Z, Zheng Y, Liu Y and Wigneron J-P (2025) Terrestrial ecosystems are in transition. Front. Remote Sens. 6:1705386. doi: 10.3389/frsen.2025.1705386
Received: 15 September 2025; Accepted: 10 November 2025;
Published: 26 November 2025.
Edited by:
Yuanwei Qin, Hohai University, ChinaReviewed by:
Yulong Zhang, Duke University, United StatesCopyright © 2025 Wang, Yan, Li, Li, Liu, Fan, Wang, Yao, Cai, Yan, Xing, Zheng, Liu and Wigneron. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Huan Wang, c3VtbWVyX3NpeEAxNjMuY29t; Jean-Pierre Wigneron, amVhbi1waWVycmUud2lnbmVyb25AaW5yYWUuZnI=