 Liming Chen
Liming Chen Yao Chen
Yao Chen- Jingdezhen University, Jingdezhen, China
Introduction: Slope position is a crucial topographic factor influencing tea plant growth and element accumulation, yet its impact on nutrient uptake and metal distribution across organs remains unclear.
Methods: This study analyzed stems, mature leaves, litter, young leaves, and tea fruits collected from upper, middle, and lower slopes of low mountainous and hilly tea plantations. Concentrations of nitrogen (N), phosphorous (P), potassium (K), iron (Fe), manganese (Mn), copper (Cu), zinc (Zn), cadmium (Cd), lead (Pb), and chromium (Cr) were measured across plant organs with respect to slope position.
Results and discussion: The N, P, and K contents in organs from upper slopes were significantly higher (by 29.33%–52.25%, 14.59%–80.58%, and 16.20%–85.19%, respectively) than those from lower slopes, with the most pronounced enrichment occurring in fruits and young leaves. Fe and Mn accumulation was significantly elevated in stems, leaves, and tea fruits at the upper and middle slopes, respectively. Cu enrichment observed in mature leaves (upper slope) and young leaves (lower slope), while Zn accumulation was highest in young leaves and fruits on lower slopes. Cd and Pb were primarily concentrated in mature leaves at upper slopes, whereas Cr was significantly elevated across all organs, especially stems and fruits, at lower slopes. Health risk assessment revealed total hazard quotients (THQ) remained within safe limits (0.194–0.233), but total carcinogenic risks slightly exceeded 1×10–4 at both upper (1.05×10-4) and lower (1.15×10-4) slopes, primarily driven by Cd (93.9%) on upper slopes and Cr (70.4%) on lower slopes. These results advocate for slope-specific management strategies in low hilly tea plantations, entailing coordinated regulation of both fertilization practices and pollution sources to ensure product safety as well as mitigate toxic element accumulation.
1 Introduction
Topographic features, particularly slope position, play a pivotal role in modulating soil nutrient dynamics and pollutant migration in agroecosystems, thereby influencing crop productivity and safety (Liu et al., 2023; Zhang et al., 2016). In hilly regions, slope-driven variations in hydrological processes and soil erosion often lead to spatial heterogeneity in nutrient availability and heavy metal accumulation, posing challenges to sustainable tea production (Jiang et al., 2020; Xiang et al., 2021). While previous studies have explored the impacts of slope position on soil properties or individual nutrient elements (Amare et al., 2024; Yusra et al., 2020), a comprehensive understanding of how slope gradients regulate the organ-specific distribution of both essential nutrients and toxic metals in tea plants remains elusive.
Previous studies underscore the influence of slope position on soil nutrient retention and redistribution. Upper slopes often exhibit higher organic matter content due to reduced erosion, whereas lower slopes accumulate leached nutrients and pollutants (Yoo et al., 2006). For tea plants, nitrogen (N), phosphorus (P), and potassium (K) are vital for growth and secondary metabolite synthesis, yet their uptake efficiencies are highly dependent on soil moisture and pH (Huang et al., 2022; Zhou et al., 2024). Concurrently, heavy metals commonly defined as metallic elements with a density greater than 5 g/cm³, such as cadmium (Cd) and chromium (Cr) in tea leaves raise significant health concerns, as chronic exposure to these elements through tea consumption has been associated with carcinogenic risks (Peng et al., 2018; Tao et al., 2021). Studies have identified atmospheric deposition and soil-root interactions as primary pathways for metal uptake in tea plants (He et al., 2023; Uraguchi and Fujiwara, 2012). However, most studies have focused on flat terrains or single-slope systems, neglecting the interplay between slope-driven hydrological processes and organ-specific metal allocation (Wang et al., 2024; Fan et al., 2020). Furthermore, prior risk assessments often overlook the spatial variability of contaminants across slope positions, limiting the development of targeted mitigation strategies.
The agricultural utilization of mountainous soils represents a substantial element of China’s land use strategy, particularly evident in the southern hilly regions. In these areas, anthropogenic reclamation activities have led to the transformation of a considerable proportion of sloped terrain into tea plantations (Zhi et al., 2018). Notably, these land-use conversions induce substantial alterations in pedogenic processes, including the transformation dynamics of soil organic matter and the biogeochemical cycling of metallic elements (Lange et al., 2023). The spatial heterogeneity of slope positions (summit, backslope, and footslope) exerts profound influences on soil physicochemical properties through differential erosion-deposition patterns and microclimatic variations (Liu et al., 2019). This topographic modulation significantly affects the phytoavailability of essential nutrients (N, P, K) and the accumulation potential of both beneficial and toxic metallic elements in Camellia sinensis (Wen et al., 2020). Consequently, systematic investigation of slope-induced pedospheric modifications provides critical insights for optimizing fertilization strategies, mitigating heavy metal risks, and establishing sustainable management protocols in these agroecosystems.
This study systematically investigates the influence of slope position on the spatial distribution of macronutrients and heavy metals across various organs of tea plants grown in a low hilly region of Jiangxi Province, China. By integrating nutrient and metal accumulation patterns with environmental processes and health implications, the research aims to: 1) quantify slope-dependent nutrient distribution and enrichment mechanisms, 2) identify dominant pathways and driving factors of metal accumulation linked to slope-induced environmental variability, and 3) assess carcinogenic risks associated with tea consumption, thereby providing insights into slope-specific agricultural management and risk mitigation strategies.
2 Materials and methods
2.1 Sampling locations
Fuliang County (Jiangxi Province, China), a historically significant tea-producing region renowned for its premium tea varieties since the Tang Dynasty (618–907 AD), represents one of China’s most prominent production areas for high-quality teas. The study site was located in Hanxi Village (117°25’59’’E, 29°27’53’’N, 125 m asl) within the county’s predominant tea cultivation zone, characterized by low mountainous and hilly topography. This region experiences a typical subtropical monsoon climate, with a mean annual temperature of 16°C, annual precipitation exceeding 1500 mm, and an extended frost-free period, coupled with abundant sunshine duration. The tea plantations, predominantly established on red soils, feature terraced cultivation systems with approximately 1-meter-wide benches and risers, developed through manual reclamation practices. All sampled tea plants were collected from uniformly established Qimen Zhuyeshu tea seedlings (Camellia sinensis var. Qimen Zhuye Zhong) planted in the spring of 2017. These cultivation characteristics represent a prototypical tea garden ecosystem in the low mountainous and hilly areas of the Yangtze River Basin.
2.2 Sample collection and pretreatment
In November 2024, a comprehensive field investigation and systematic sampling were conducted in the selected tea plantation. Sampling sites were strategically chosen based on consistent slope aspects, uniform slope gradients, and homogeneous growth parameters including plant development status and vigor. According to topographical and altitude variations, the tea plantation was stratified into three distinct slope positions, namely the upper slope position (129.51 m asl), the middle slope position (91.13 m asl), and the lower slope position (56.42 m asl), for the collection of tea plant samples. The altitude information and soil basic physicochemical properties of the three slope positions are shown in Table 1. A total of ninety healthy and uniformly growing tea plants were sampled. Three replicated sampling plots were established at each slope position, with ten plants collected from each plot. Following field collection, plant samples were immediately transported to the laboratory and dissected into five morphological components: stems, mature leaves, litter, young leaves, and tea fruits. The organ selection was based on their distinct physiological and ecological functions: stems represent the transport pathway; mature and young leaves are the main sites of metal accumulation and the primary commercial product; litter (senesced leaves collected from the ground) was included to assess potential metal return to the soil via decomposition; and tea fruits were analyzed to evaluate the allocation of metals to reproductive structures.
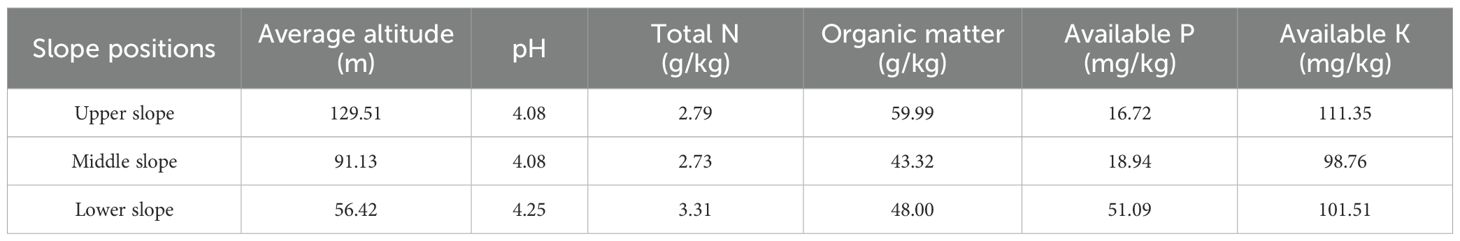
Table 1. Altitude information and basic physicochemical properties of soils at different slope positions.
All samples underwent standardized pretreatment procedures to ensure analytical reliability (GB/T8303-2013, 2013). The protocol involved initial enzyme deactivation through heat treatment at 105 °C for 30 minutes, followed by desiccation in a forced-air oven maintained at 80 °C until constant mass was achieved (typically 48–72 hours). Dry matter content was quantitatively determined gravimetrically. Subsequently, the dehydrated samples were homogenized using a stainless-steel grinder and passed through a 0.5-mm mesh sieve. The processed samples were subsequently subjected to quantitative analysis to determine essential macronutrients (N, P, and K) and trace metal elements, including iron (Fe), manganese (Mn), copper (Cu), zinc (Zn), cadmium (Cd), lead (Pb), and chromium (Cr), following established analytical protocols.
2.3 Element determination and analysis
The contents of total N, total P, and total K in various organs of the tea plant were determined using the Kjeldahl method, the molybdenum-antimony anti-colorimetric method, and flame photometry, respectively (NY/T2017-2011, 2011). To determine the concentrations of Fe, Mn, Cu, Zn, Cd, Pb, and Cr in tea plant samples, the samples were digested using the HNO3-H2O2 microwave digestion method (GB5009.268-2025, 2025). Briefly, 0.2000 g of pulverized tea plant sample was accurately weighed and placed into a microwave digestion vessel. Then, 8 mL of HNO3 and 2 mL of H2O2 were added, and the vessel was securely sealed. The sample was digested in a microwave digestion system (ETHOS UP, Milestone, Sorisole, Italy) following a predefined protocol. The concentrations of the target elements (Fe, Mn, Cu, Zn, Cd, Pb, and Cr) in various tea plant organs were determined by inductively coupled plasma mass spectrometry (iCAP-Q ICP-MS, Thermo Scientific, Waltham, MA, USA).
2.4 Human health risk assessment
The exposure risk associated with tea consumption was quantified using the health risk assessment model recommended by the United States Environmental Protection Agency (USEPA), with all assessment indicators derived from the average daily intake (ADI, mg·kg-1·day-1) (US EPA, 2015). In this study, the risk assessment calculations were performed using the mean concentrations of each element. The ADI is calculated as Equation 1 (Peng et al., 2018):
Where C is the concentration of metal elements in dried tea leaves (mg·kg-1). IR is the ingestion rate (1 L·d-1). EF is the exposure frequency (365 d·a-1). ED is the exposure duration (57 a). AT is the time period (d), which can be calculated as AT=ED×365. BW is the human body weight (70 kg for adults).
The non-carcinogenic risk associated with heavy metals in tea was assessed using the Hazard Quotient (HQ) methodology. The total non-carcinogenic risk, expressed as the Target Hazard Quotient (THQ), was defined as the arithmetic sum of individual HQ values for all detectable heavy metals, and were defined as Equations 2, 3 (Zhou et al., 2016; Ju et al., 2024):
Where RfDi is the oral intake amount of heavy metal (i) proposed by USEPA (mg·kg-1·d-1). RfDi values for Fe, Mn, Cu, Zn, Cd, Pb and Cr were 0.1800, 0.1400, 0.3000, 0.0400, 0.0010, 0.0035 and 0.0030 mg·kg-1·d-1. HQ and THQ< 1 indicates no human carcinogenic risk, while HQ and THQ > 1 indicates potential human carcinogenic risk, with the higher the value, the higher the risk. The carcinogenic risk can be calculated by the following Equations 4, 5 (Tao et al., 2021):
Where Riski is the carcinogenic risk of heavy metal (i). RiskTotal is the summation of each Riski value. SF is the slope factor of cancer. SF values for Cr, Cd, and Pb were 0.5, 15 and 0.008. There is no reference standard value for other elements. Risk< 10-6, Risk = 10-6-10–4 and Risk >10–4 indicates no significant risk, acceptable risk and unacceptable risk, respectively.
2.5 Statistical analysis
Statistical analyses were performed using one-way analysis of variance (ANOVA) implemented in SPSS statistical software (version 19.0; SPSS Inc., Chicago, IL, USA). Post hoc multiple comparisons between treatment groups were conducted using Fisher’s least significant difference (LSD) test, with statistical significance determined at P<0.05. The linear statistical model used for ANOVA included the replication number, Slope position (SP), Organ (O), and the interactions of SP×O.
3 Results
3.1 Nutrient element contents
A significant spatial heterogeneity was observed in the total N, P, and K contents across various organs of tea plants at different slope positions in low mountainous and hilly tea plantations. Overall, the total N content in the organs of tea plants at the upper slope position was significantly higher, ranging from 29.33% to 52.25%, compared to those at the lower slope position (Figure 1). Among all organs, tea fruits exhibited the highest total N content, with significantly greater levels at the upper slope position compared to the middle and lower slope positions. In contrast, litter displayed the lowest total N content, with significantly higher values at the upper slope position compared to the middle and lower slope positions. Furthermore, the total N content in stems, mature leaves, and young leaves also demonstrated a consistent trend, with higher levels at the upper slope position than at the middle and lower slope positions.

Figure 1. Content of total nitrogen in the same organ of the tea plant at different slope positions. Data are presented as the means ± SEM of three replicates. Different lowercase letters indicate significant differences among the slope positions (P< 0.05).
The total P content in the organs of tea plants located at the upper slope position was significantly higher, ranging from 14.59% to 80.58%, in comparison to those at the lower slope position (Figure 2). Tea fruits demonstrated the highest total P content, exhibiting significantly higher levels at the upper slope position than at the middle and lower slope positions. In contrast, litter displayed the lowest total P content, with significantly higher values at the upper slope position compared to the middle and lower slope positions. A similar trend was observed in stems, with mature leaves and young leaves displaying comparatively higher total P content at the upper slope position. However, in mature leaves, no significant difference in total P content was detected between the middle and lower slope positions.
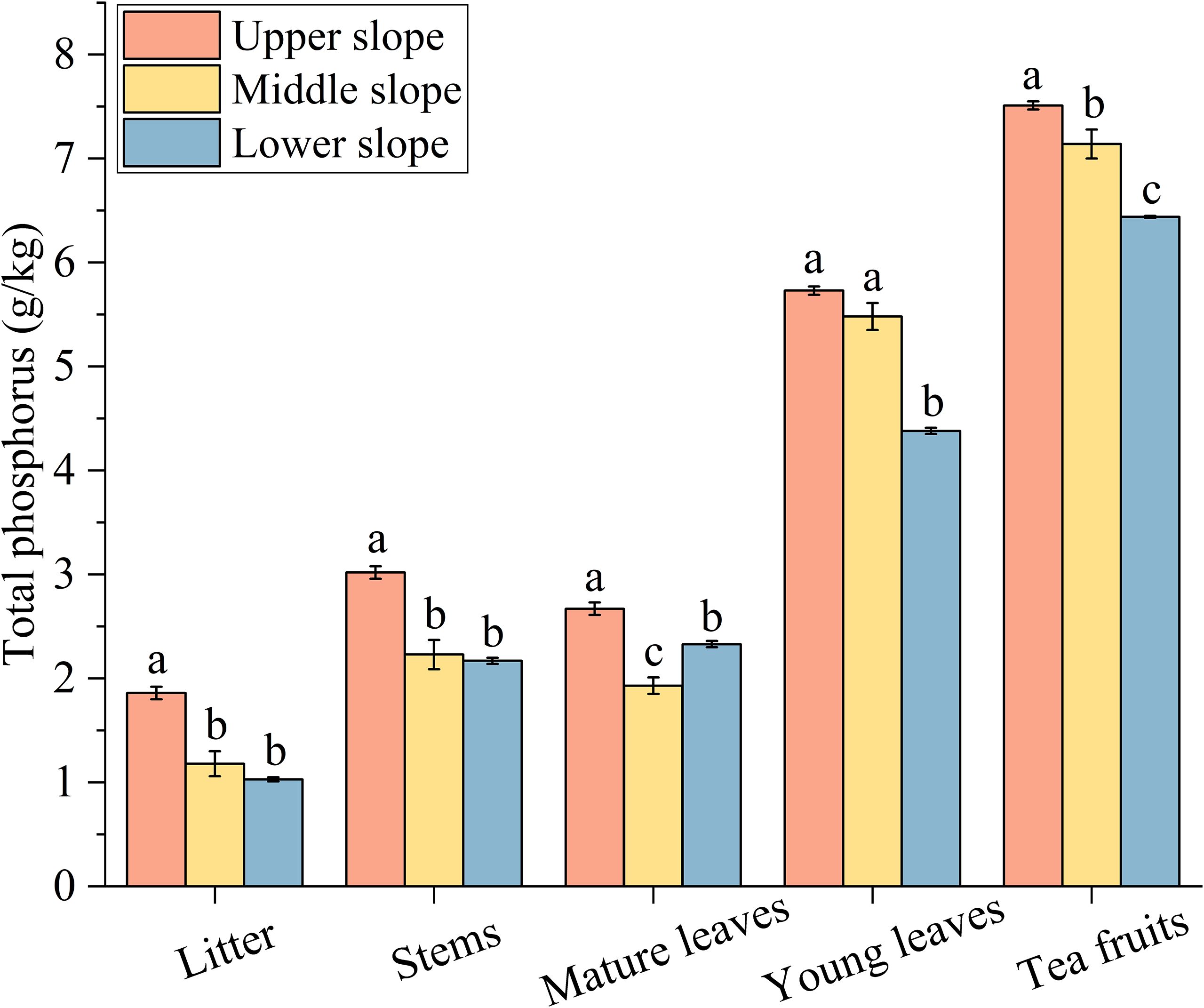
Figure 2. Content of total phosphorus in the same organ of the tea plant at different slope positions. Data are presented as the means ± SEM of three replicates. Different lowercase letters indicate significant differences among the slope positions (P< 0.05).
The total K content in various organs of tea plants at the upper slope position was significantly higher (16.20% to 85.19%) than those at the lower slope position (Figure 3). Amongst all organs, tea fruits demonstrated the highest total K content, with the upper slope position exhibiting significantly greater total K content in tea fruits than the middle and lower slope positions. In the case of litter, stems, and young leaves, the total K content at the upper slope position was significantly higher than that at the middle and lower slope positions. Conversely, for mature leaves, the total K content at both the upper and middle slope positions was significantly higher than that at the lower slope position. However, no significant difference was observed between the upper and middle slope positions.
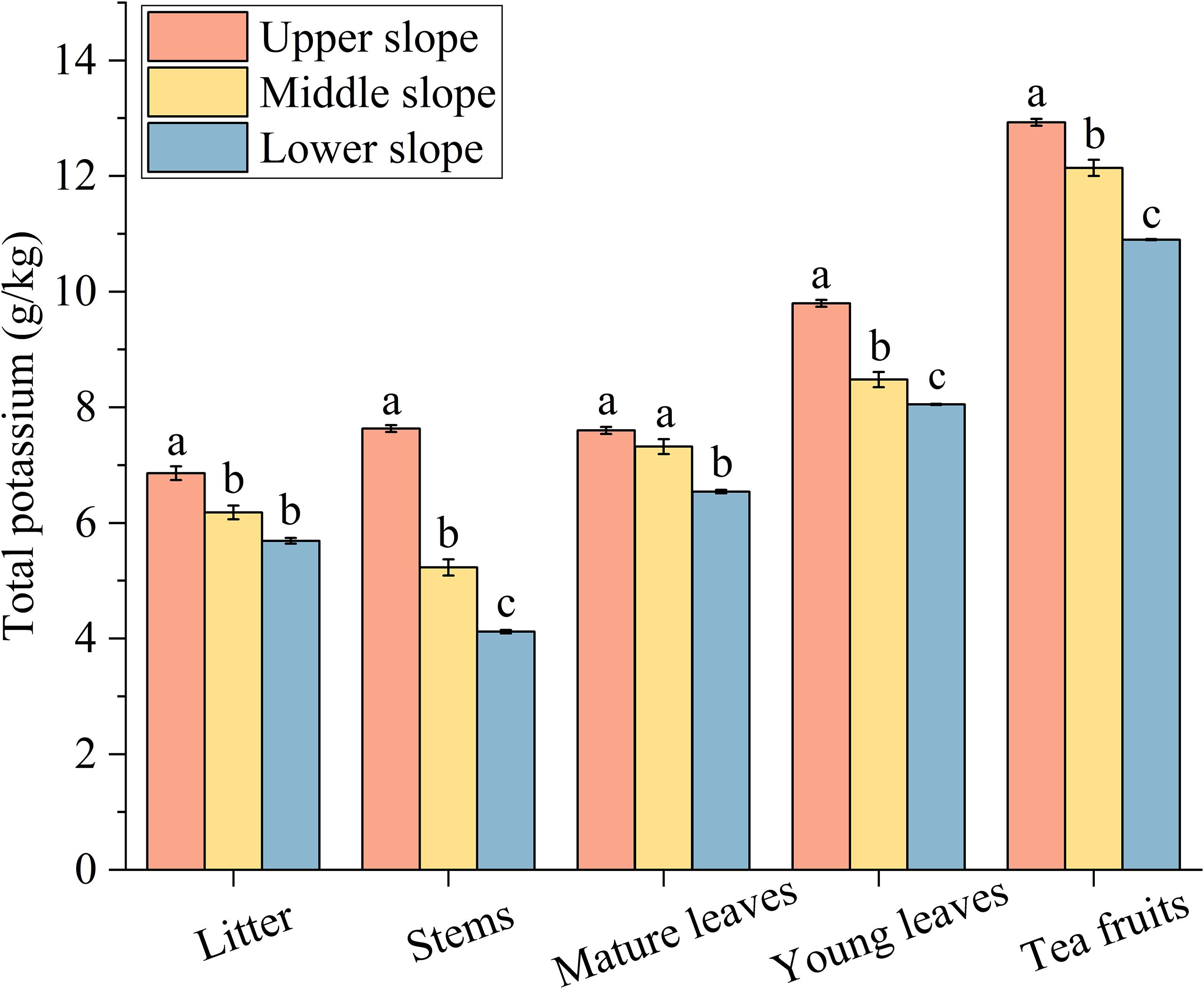
Figure 3. Content of total potassium in the same organ of the tea plant at different slope positions. Data are presented as the means ± SEM of three replicates. Different lowercase letters indicate significant differences among the slope positions (P< 0.05).
3.2 Content of Cu, Mn, Zn, and Fe
Slope position exerted a significant influence on the concentrations of various elements in different tea plant organs (Figure 4). This influence varied considerably depending on the specific element and organ type, exhibiting no consistent pattern across slope positions. Significant variations in Fe content were observed among slope positions for all organs. Litter Fe content differed substantially among slopes, with the lower slope exhibiting 14.5% and 22.7% higher concentrations than the middle and upper slopes, respectively (Figure 4A). Conversely, stems, mature leaves, young leaves, and tea fruits demonstrated significantly elevated Fe levels in the upper slope compared to both the middle and lower slopes. Specifically, upper slope stems contained 16.9% more Fe than lower slope stems and 112% more than middle slope stems, while upper slope tea fruits showed 15.6% and 38.5% higher Fe content than those from middle and lower slopes. Regarding Mn, litter Mn varied significantly across slopes, with lower slope concentrations exceeding upper and middle slope values by 39.1% and 102% (Figure 4B). Stems, mature leaves, and young leaves all exhibited peak Mn concentrations in the middle slope: stems showed 45.1% and 86.6% higher Mn than lower and upper slopes; mature leaves contained 22.9% and 67.2% more Mn than upper and lower slopes; young leaves displayed 28.9% and 82.2% greater Mn than upper and lower slopes. Tea fruits Mn showed no significant difference between upper and middle slopes, though both significantly exceeded lower slope values. For Cu, lower slope litter contained 36.6% and 83.7% more Cu than upper and middle slopes, while stems peaked in the middle slope (Figure 4C). Mature leaves showed significantly higher Cu in the upper slope, whereas young leaves peaked in the lower slope with 22.3% higher concentrations than the upper slope. Tea fruits Cu was significantly elevated in the lower slope compared to the upper slope. Zn content varied significantly across slopes for all organs, with litter, stems, and mature leaves all reaching maximum concentrations in the middle slope (Figure 4D). litter Zn was 12.8% and 37.0% higher than lower and upper slopes; stems Zn exceeded upper and lower slope values by 21.6% and 100%; mature leaves Zn was 33.3% and 60.8% greater than upper and lower slopes. Young leaves Zn peaked in the lower slope, surpassing middle and upper slope concentrations by 39.1% and 68.2%, while tea fruits Zn was significantly higher in the lower slope, exceeding upper and middle slope values by 18.0% and 20.7%.
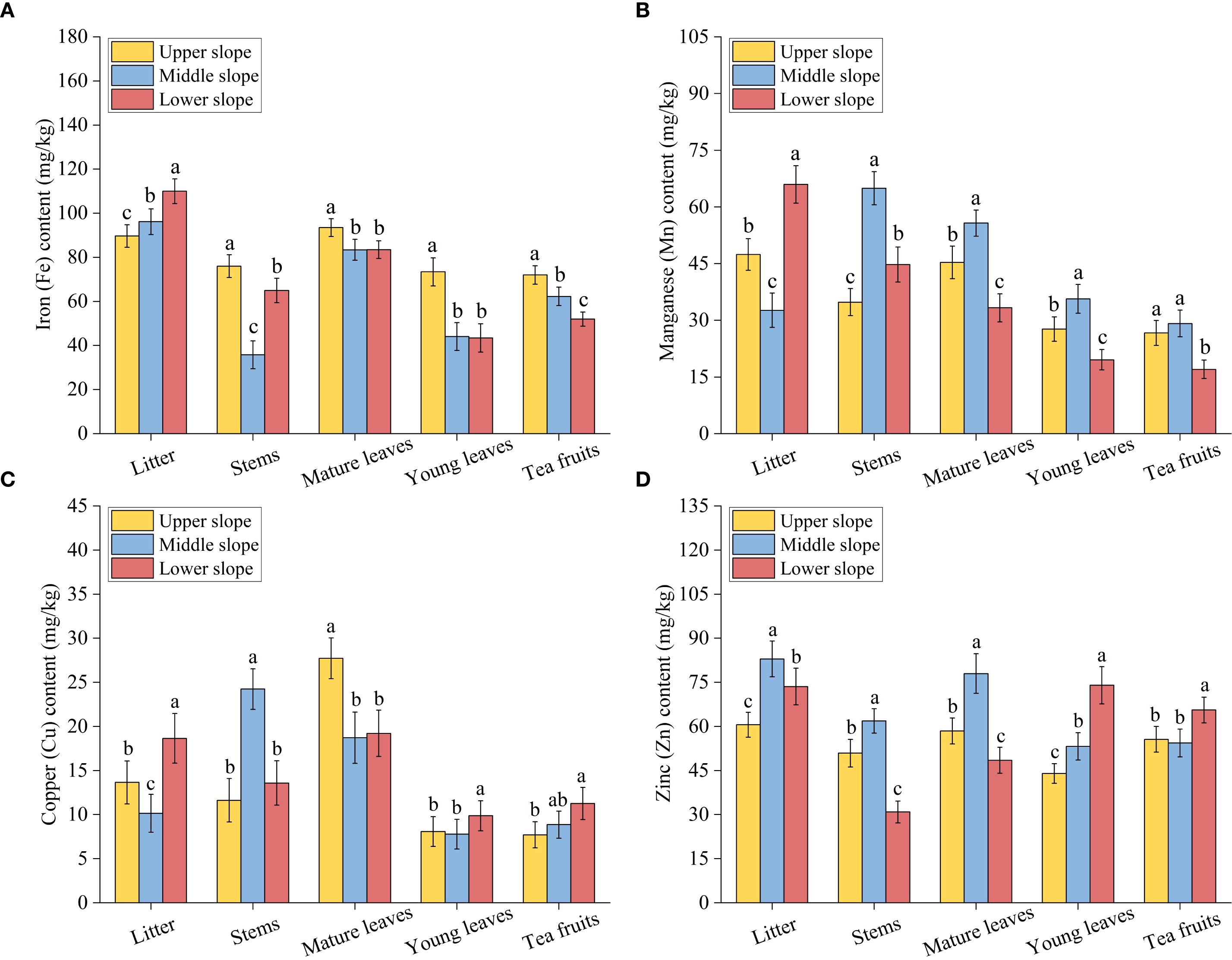
Figure 4. Contents of Fe (A), Mn (B), Cu (C), and Zn (D) in various organs of tea plants at different slope positions. Data are presented as the means ± SEM of three replicates. Different lowercase letters indicate significant differences among the slope positions (P< 0.05).
3.3 Content of Cd, Pb, and Cr
Significant variations in Cd content were observed across slope positions (Figure 5A). Litter Cd was significantly higher in the middle and lower slopes than in the upper slope, reaching 1.72 times the upper slope concentration. Stem Cd content was 45.5% higher in the middle slope than in the lower slope and 71.4% higher than in the upper slope. Conversely, mature leaves exhibited significantly higher Cd in the upper slope, exceeding lower and middle slope values by 29.6% and 84.2% respectively. Young leaves showed substantially elevated Cd in the upper slope, containing 2.31 and 2.85 times the concentrations found in the middle and lower slopes. Tea fruit Cd peaked in the middle slope at 2.33 times the upper slope levels and 2.24 times the lower slope levels. For Pb, litter, stems, mature leaves, and young leaves all showed significant inter-slope differences, with upper slope concentrations exceeding middle and lower slopes by 43.7% and 58.7% respectively (Figure 5B). Middle slope stem Pb was 9.2% higher than the upper slope and 36.3% higher than the lower slope values. Upper slope mature leaf Pb exceeded lower and middle slope concentrations by 50.7% and 66.4%, while middle slope young leaf Pb was 58.1% higher than lower slope and 65.9% higher than upper slope. Tea fruit Pb reached 2.06 and 2.68 times the upper and lower slope levels in the middle slope. Cr content varied significantly across all three slope positions for all organs (Figure 5C). Lower slope litter Cr exceeded upper and middle slope concentrations by 3.2% and 407% respectively. Lower slope stems contained 229% and 863% more Cr than the upper and middle slopes. Lower slope mature leaves showed 86.8% and 484% higher Cr than middle and upper slopes, while lower slope young leaves exhibited 529% and 1273% greater Cr than middle and upper slopes. Middle slope tea fruit Cr exceeded lower and upper slope values by 14.5% and 839%.

Figure 5. Contents of Cd (A), Pb (B), and Cr (C) in various organs of tea plants at different slope positions. Data are presented as the means ± SEM of three replicates. Different lowercase letters indicate significant differences among the slope positions (P< 0.05).
3.4 Effects of slope position, tea plant organs and interactions on element distribution
Multivariate analysis of variance revealed statistically significant effects (P< 0.01) of slope position, plant organ, and their interaction on the concentrations of Fe, Mn, Zn, Cd, Pb, and Cr (Table 2). While Cu accumulation exhibited no statistically significant variation across slope positions, both organ-specific distribution patterns (P< 0.01) and slope-organ interaction effects (P< 0.01) demonstrated high significance for this element.
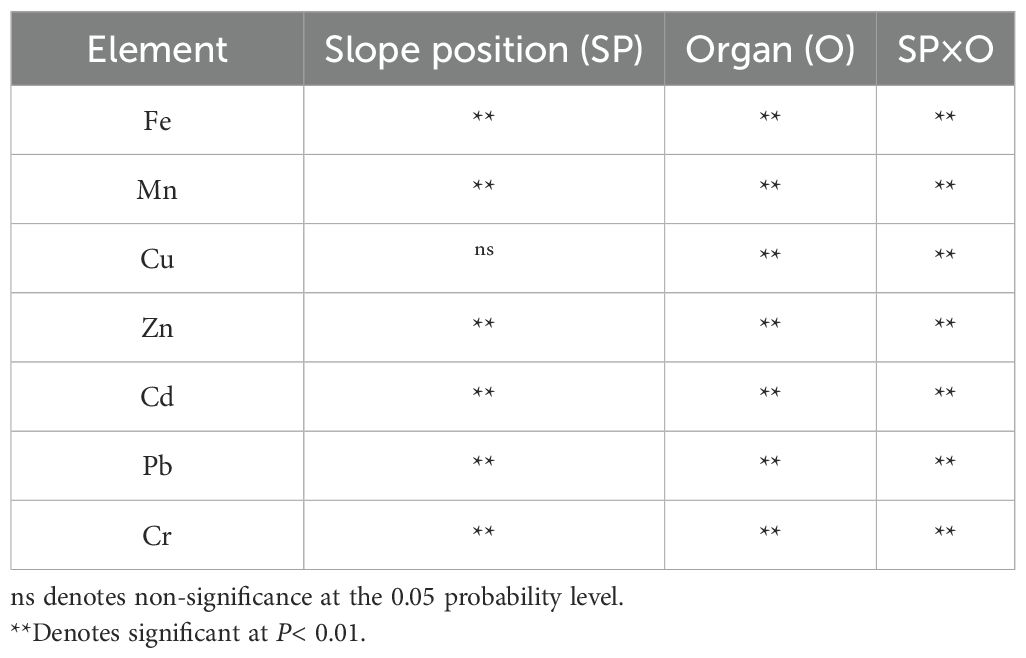
Table 2. Analysis of variance (ANOVA) of heavy metals in tea plant organs at different slope positions.
3.5 Health risk assessment
The ADI values of Fe, Mn, Cu, and Zn ranged from 7.734×10-3 - 1.314×10-2, 3.432×10-3 - 6.473×10-3, 1.358×10-3 - 1.803×10-3, and 7.607×10-3 - 1.351×10–2 mg·kg-1·day-1, respectively, in all slopes, which did not show a significant spatial difference (Table 3). The ADI magnitudes of Cd, Pb and Cr were lower (10-6~10-4), but the ADI of Cr on Lower slope (1.621×10–4 mg·kg-1·day-1) was significantly higher than that of other slope positions. The mean single-factor HQ values for all elements were below 1 (0.002-0.073), and the total HQ values ranged from 0.194 to 0.233. The mean HQ values for each slope are shown in descending order as follows: lower slope (0.230) > upper slope (0.198) > middle slope (0.195). The element exhibiting the highest mean value of carcinogenic risk was Cd across all the different slopes, with Risk values approaching the upper acceptable limit (1×10-4) for Cd (9.870×10-5) in the upper slope and Cr (8.103×10-5) in the lower slope. The total Risk of the three slopes is calculated to be 1.050×10-4 (upper slope), 5.610×10-5 (middle slope) and 1.150×10-4 (lower slope), where the total Risk of the upper slope and lower slope is slightly more than 1×10-4.

Table 3. Average daily intake (ADI) and non-carcinogenic risk (HQ) and carcinogenic risk (Risk) of heavy metals.
The contribution rate of Fe exhibited a progressive decrease from upper to lower slope positions (36.90% →22.30% →18.70%), while Mn reached its maximum contribution (23.40%) at the middle slope position (Figure 6). Comparatively modest variations were observed in the contribution rates of Cu and Zn across different slope positions, ranging from 17.80% to 19.10% and 13.10% to 19.10%, respectively. Cd and Pb demonstrated generally lower contribution rates, both displaying descending trends with decreasing slope elevation (Cd: 3.50% → 0.90%; Pb: 14.20% → 7.80%). Notably, Cr exhibited a marked elevation in contribution rate at the lower slope position (23.50%), substantially exceeding values recorded at upper (2.0%) and middle (4.60%) slope positions.
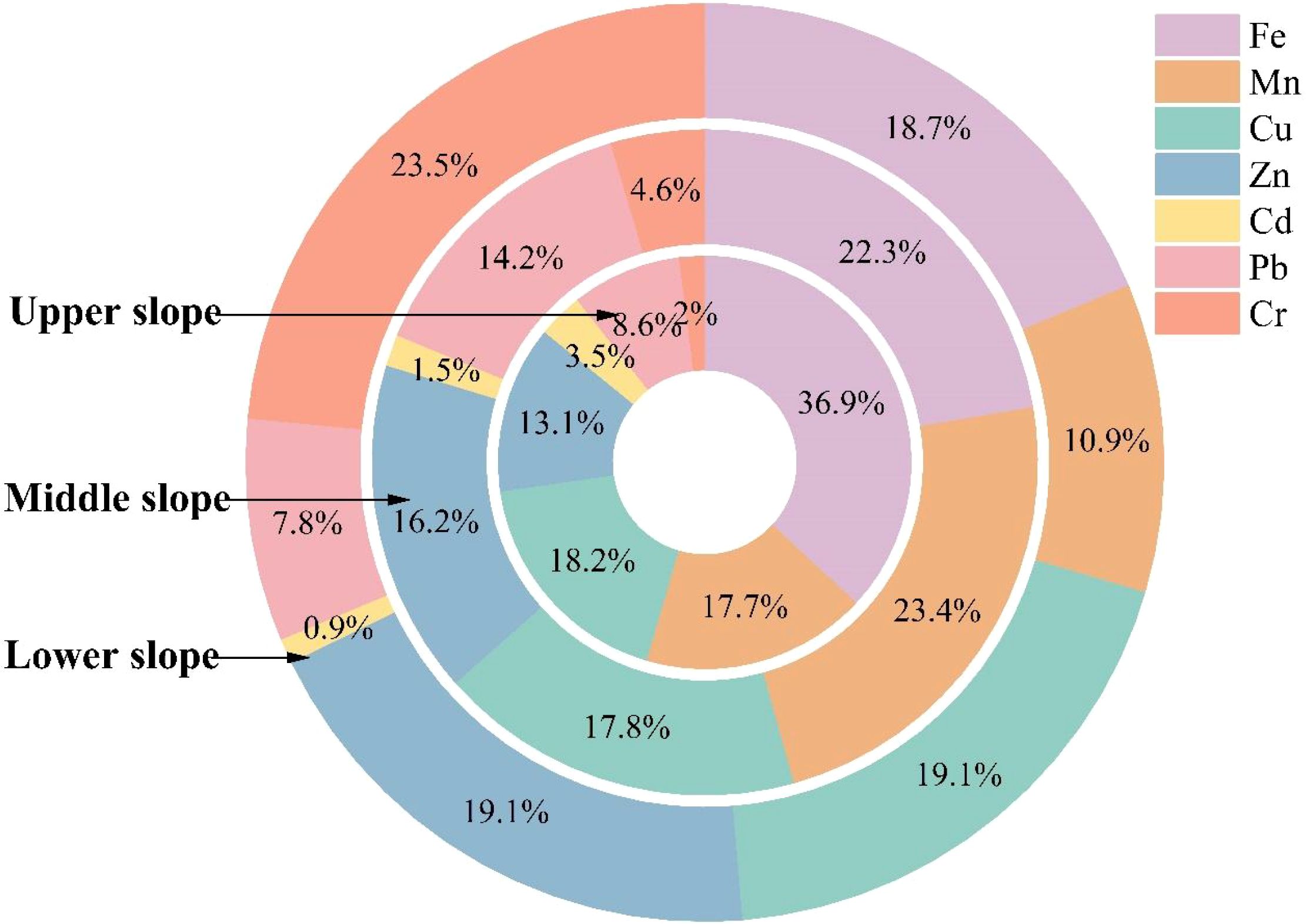
Figure 6. The percentage contribution of Target Hazard Quotient (THQ) of heavy metals.
With regard to the percentage contribution to RiskTotal, Cd dominated RiskTotal contributions in the upper slope (93.90%), which progressively decreasing to 75.60% in the middle slope and sharply declining to 29.20% in the lower slope (Figure 7). Conversely, Cr exhibited an inverse spatial pattern, with its contribution increasing from 5.60% (upper slope) to 23.00% (middle slope) and peaking at 70.40% in the lower slope. Pb maintained consistently minimal contributions across all slope positions (0.50–1.50%), demonstrating negligible influence on RiskTotal.
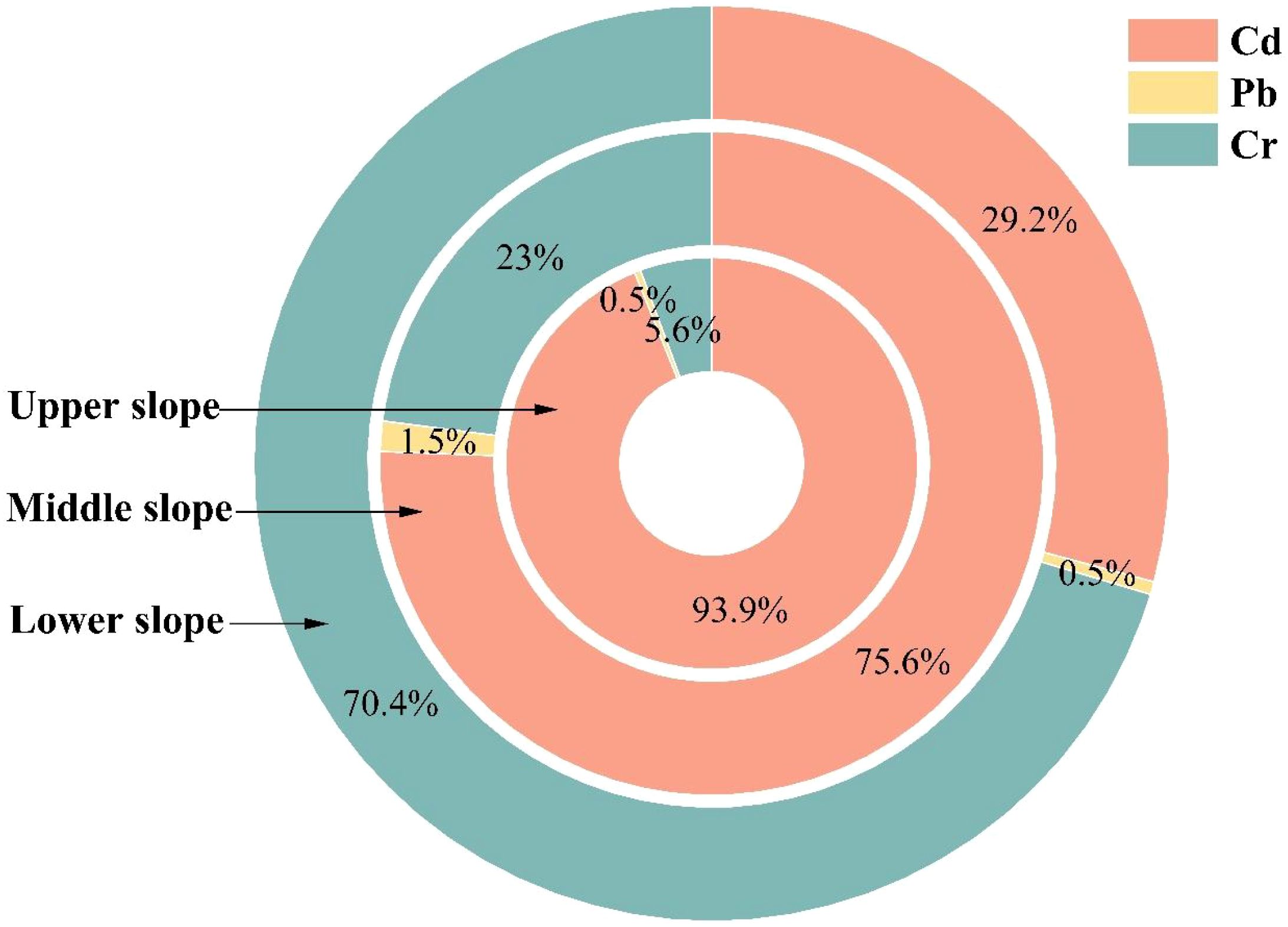
Figure 7. The percentage contribution of RiskTotal of Cd, Pb, and Cr.
4 Discussion
4.1 Impact of slope position on nutrient element uptake in tea plants
The present study revealed that the concentrations of N, P and K in various tea plant organs within hilly tea plantations exhibited a consistent trend of upper slope > middle slope > lower slope, with tea fruits and young leaves demonstrating the highest nutrient concentrations. This phenomenon may be closely associated with slope position-mediated regulation of soil nutrient distribution and hydrological transport. Previous research has indicated that upper slope soils typically exhibit higher organic matter accumulation and mineral nutrient retention due to reduced water infiltration caused by gravitational effects and weaker soil erosion (Zhang et al., 2023; Buraka et al., 2022). Conversely, middle and lower slopes are prone to surface runoff-induced nutrient leaching losses, leading to diminished availability of soil nutrients. The observed nutrient enrichment in tea fruits and young leaves is likely to reflect their substantial nutrient demands as organs characterized by elevated metabolic activity. Specifically, tea fruits, as reproductive organs, may prioritize nutrient acquisition through hormonal signaling pathways, particularly through the allocation mechanisms of nutrients mediated by cytokines (Liu et al., 2021). Furthermore, leaf litter exhibited the lowest nutrient levels, with further reductions observed in lower slope positions, which may be attributable to slope-dependent variations in decomposition rates and nutrient recycling processes (Hosseini et al., 2022). This phenomenon is primarily driven by nutrient loss during decomposition: soluble nutrients (e.g., K+, NO3-) are rapidly leached by rainwater, while labile carbon compounds are mineralized by microbes, leaving recalcitrant compounds (e.g., lignin) with low nutrient density. Lower slope positions likely accelerate these processes due to higher soil moisture and prolonged litter submersion during runoff events, which further depletes nutrients (Chen et al., 2023).
Notably, mature and young leaves exhibited significantly higher N, P, and K concentrations in upper slope positions compared to middle and lower slopes (Figure 2), a phenomenon potentially attributed to active nutrient demands driven by photosynthetic requirements. Specifically, leaf N predominantly contributes to chlorophyll biosynthesis and Rubisco enzyme production, while K facilitates stomatal regulation (Ruan et al., 2019). The superior light availability in upper slopes may enhance photosynthetic carbon assimilation, thereby indirectly stimulating root nutrient uptake through carbohydrate-mediated signaling (Chen et al., 2016). However, no significant difference in P concentration was observed between middle and lower slopes for mature leaves, likely reflecting P’s limited mobility in soil systems. In acidic red soils, phosphate ions are readily immobilized through adsorption to iron/aluminum oxides, with their bioavailability largely dependent on root-exuded organic acids (e.g., citrate) for activation (Lei et al., 2024). Furthermore, elevated soil moisture content in middle and lower slope positions may exacerbate P leaching losses (Kochian et al., 2015). While leaching transports soluble nutrients downslope, our results for N, P, and K consistently showed higher concentrations in upper slope plant organs, contrary to a potential expectation of enrichment in lower slopes. This indicates that plant uptake and soil retention processes at the upper slope (e.g., reduced erosion, higher organic matter) are more dominant factors for these macronutrients than the translocation via leaching for the tea plants in this system. This highlights the importance of implementing slope-specific nutrient management strategies in hilly tea plantations. Specifically, upper slopes should prioritize reduced N inputs to minimize fertilizer redundancy, given their inherent nutrient retention capacity. In contrast, middle and lower slopes require targeted supplementation of P and K fertilizers to counteract leaching-induced deficiencies and optimize nutrient stoichiometry.
4.2 Impact of slope position on metal element accumulation in tea plants
The absorption and allocation of heavy metals in tea plants (Camellia sinensis) within low hilly plantations demonstrated marked organ-specific patterns and slope-position interactions. The pronounced enrichment of Cu or Fe in stems, leaves, and tea fruits from upper slope positions can likely be attributed to enhanced metal bioavailability under acidic soil conditions (Jia et al., 2024). Table 1 shows corroborating evidence of lower pH values in upper slope soils compared to lower slopes. Under such acidic conditions, the dissolution of Cu2+ and Fe3+ is facilitated (Alloway, 2013). In response, tea plants may upregulate metal transporters (e.g., YSL family proteins) to improve Fe chelation and uptake (Curie et al., 2009). Meanwhile, increased accumulation of Cu in mature leaves, which serves as a cofactor for key oxidases like polyphenol oxidase, may indicate strengthened antioxidant metabolism in plants growing upper slope positions (Maksymiec, 2007). For instance, the Fe concentration in upper slope mature leaves (93.487 mg/kg) resides within the agronomically optimal range for tea plants, but the Mn content in middle-slope young leaves (19.574 mg/kg) approaches the upper limit of typical phytotoxicity thresholds (Alloway, 2013), indicating a potential risk of stress or warranting monitoring. This discrepancy highlights the role of soil redox dynamics in modulating Mn bioavailability at middle slopes. Notably, abnormal Mn enrichment in mature and young leaves at middle slopes may be associated with dynamic fluctuations in soil redox potential at this slope. Seasonal waterlogging in hilly tea plantations facilitates the reductive dissolution of Mn oxides (e.g., MnO2) into soluble Mn2+ (Millaleo et al., 2010), while tea leaves actively absorb Mn through plasma membrane-localized transporters (NRAMP family) to support photosynthetic function, particularly in the oxygen-evolving complex of photosystem II (Pittman, 2005). The Mn levels in middle-slope litter (32.646 mg/kg) are comparable to those reported in other horticultural crops under similar redox conditions (Rotter et al., 2017), but the significantly higher Mn in upper slope litter (47.396 mg/kg) underscores the role of acidification in mobilizing this element. This finding aligns with field studies showing that pH< 5.5 dramatically increases Mn solubility (Mayanna et al., 2015).
The accumulation patterns of Cd, Pb, and Cr further elucidate slope-mediated regulation of pollutant migration and transformation. The pronounced Cd enrichment observed in upper-slope mature leaves (0.95-fold higher than in young leaves) may predominantly arise from stomatal uptake pathways. Elevated wind velocities at upper slopes enhance the deposition of Cd-containing atmospheric particulates onto leaf surfaces, facilitating Cd entry into the symplast via stomatal infiltration (Uraguchi et al., 2009; Ouyang et al., 2023). The Cd concentration in upper-slope mature leaves (0.035 mg/kg) is within the range reported for tea plants in non-polluted regions (Hu et al., 2023), but the elevated Cd in middle-slope stems (0.048 mg/kg) exceeds the maximum level (0.02 mg/kg) recommended for processed tea leaves by FAO/WHO (2021), suggesting a potential risk of Cd transfer into consumable tea products and associated health concerns. This discrepancy warrants further investigation into whether slope-specific hydrological processes (e.g., leaching) amplify Cd bioavailability in transitional zones. The elevated Cd accumulation in middle-slope stems (1.55-fold higher than in litter) likely results from vertical leaching of soil Cd. The position of this transitional slope, characterized by active hydrological transport, facilitates the downward migration of dissolved Cd with soil water. This process is then intercepted by stem xylem flow (Lux et al., 2011), thereby inducing a sequestration mechanism analogous to Cd compartmentalization in rice nodes (Uraguchi and Fujiwara, 2012). The Pb concentration in upper-slope mature leaves (1.594 mg·kg-1) significantly exceeds the typical range for cultivated C. sinensis in Asian humid subtropical regions (Zhang et al., 2020), a deviation attributable to localized atmospheric deposition. Conversely, Cr levels in lower slope stems (1.04 mg·kg-1) correspond to pedogenic background concentrations in agricultural soils (Ju et al., 2024), reflecting geogenic sourcing. The heightened Pb concentration in upper slope mature leaves (4.86-fold greater than in young leaves) may be correlated with atmospheric particulate deposition through stomatal uptake (Shahid et al., 2017). Additional retention mechanisms, including adsorption by the leaf cuticular wax layer and cell wall sequestration, likely restrict the translocation of Pb to young leaves (Rahman et al., 2024). However, the Pb in upper-slope litter (1.525 mg/kg) remains within safe thresholds for soil ecosystems (Alloway, 2013), suggesting minimal long-term environmental risks. This contrast between foliar and litter Pb concentrations underscores the importance of organ-specific partitioning in mitigating metal toxicity. Cr primarily accumulates in lower-slope stems and fruits, potentially driven by depositional processes related to topography. Chronic water flow transports Cr to lower-slope soils, where subsequent root uptake facilitates its systemic distribution (Becquer et al., 2003). The Cr concentration in lower-slope tea fruits (0.861 mg/kg) is consistent with reports for fruiting crops in unpolluted areas (Hu et al., 2023), but the elevated Cr in lower-slope stems (1.04 mg/kg) warrants further investigation, including speciation analysis, to assess both phytotoxicity and broader ecological risks, given the significantly higher toxicity of Cr(VI) compared to Cr(III) (Shahid et al., 2017). However, as this study provides point-in-time measurements, the precise rates of downslope transport and leaching fluxes for elements like Cr remain unquantified. Future studies incorporating longitudinal sampling and detailed monitoring of soil pore water dynamics are essential to directly quantify these rates and model the kinetics of element movement between slope positions. Overall, the metal accumulation patterns reveal a key distinction: Cr conformed to the expected downslope leaching and accumulation behavior, acting as a hydrological sink. In contrast, the distributions of Cd and Pb were primarily governed by atmospheric deposition and subsequent plant uptake processes at the upper slope, overriding leaching effects.
4.3 Health risk assessment
Health risk assessment revealed that although the acceptable daily intake (ADI) of Fe, Mn, Cu, and Zn remained below threshold levels, the ADI of Cr at lower slopes (1.621×10-4 mg·kg-1·day-1) showed significant elevation. This elevation is likely associated with Cr enrichment caused by poor soil drainage in these positions (Antoniadis et al., 2017). Under reducing conditions, Cr predominantly exists as Cr(III), but rhizospheric oxidation may generate toxic Cr(VI), which enters plants via sulfate transporters (Riaz et al., 2024). As hydrological sinks, lower slopes accumulate pollutants transported from upper slopes through surface runoff, exacerbating Cr accumulation risks (Li et al., 2011). Although the total hazard quotient (HQTotal: 0.194-0.233) remained below safety thresholds, the maximum HQ at lower slopes (0.230) suggests potential concerns regarding low-dose cumulative exposure from chronic tea consumption. Carcinogenic risk (RiskTotal) marginally exceeded 1×10-4 at upper and lower slopes, with Cd (93.9% contribution at upper slopes) and Cr (70.4% contribution at lower slopes) identified as primary risk drivers (Figure 7). The elevated Cd risk correlates with its strong bioaccumulation capacity in tea leaves and low toxicity thresholds (Ju et al., 2024), while lower soil pH at upper slopes likely enhances Cd bioavailability (Hao et al., 2024). Conversely, the spatial differentiation in Cr risk (70.4% contribution at lower slopes) may reflect localized anthropogenic inputs (e.g., pesticide application or industrial contamination), necessitating further quantification through source apportionment models (Zhou et al., 2016).
The primary risk drivers are starkly contrasted across the slope, with Cd (atmospheric) dominating at the top and Cr (leaching or runoff) at the base, a pattern that directly reflects their divergent transport and accumulation mechanisms. This finding underscores the critical need for slope-specific management strategies and provides a solid foundation for developing tailored mitigation measures. To mitigate the aforementioned risks, the following management strategies are recommended: 1) restrict the application of chromium-based agrochemicals and inputs of industrial pollutants on lower slopes; 2) employ rationalized fertilization practices to ensure a balanced nutrient supply; 3) utilize organic management strategies, such as biochar or lime amendment, to modulate soil pH and lower the bioavailability of heavy metals. Furthermore, optimizing the spatial arrangement of tea plantations in low hilly regions and prioritizing the cultivation of high-value tea varieties on upper slopes can help reconcile nutrient use efficiency with tea product safety.
5 Conclusions
This study revealed distinct slope-dependent variations in nutrient and heavy metal accumulation in tea plants within low mountainous and hilly plantations. Upper slopes showed higher concentrations of N, P, and K in tea organs, especially in tea fruits and young leaves, whereas lower slopes showed reduced nutrient availability. Fe was elevated across multiple organs on upper slopes, while Mn peaked in stems and leaves at the middle slope. Cu accumulation was slope-dependent, highest in mature leaves (upper) and young leaves (lower). Zn showed distinct organ-specificity, concentrating in young leaves and fruits at lower slopes. Cd and Pb were mainly concentrated in mature leaves on upper slopes, and Cr was notably elevated in all plant organs, particularly stems and fruits, on lower slopes. Health risk assessments confirmed that total hazard quotients (THQs) below safety thresholds, whereas total carcinogenic risks slightly exceeded 1×10-4 on both upper and lower slopes, primarily driven by Cd on upper slopes and Cr on lower slopes. Slope-specific management strategies, including optimized fertilization, restricted use of agrochemicals, and soil amendments, are recommended to mitigate risks and support sustainable tea production.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Author contributions
LC: Conceptualization, Data curation, Funding acquisition, Investigation, Project administration, Writing – review & editing. YC: Data curation, Investigation, Writing – original draft. JA: Investigation, Validation, Writing – original draft.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This research was funded by the Science and Technology Research Project of Jiangxi Provincial Department of Education (Grant No. GJJ2402305), the Jingdezhen University 2024 University-Level Scientific Research Project (Incubation Category) (Grant No. 2024xjkt-01), and the National-Level College Student Innovation and Entrepreneurship Training Program Project for 2025 (Grant No. 202510894003).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Alloway B. J. (2013). Heavy metals in Soils: Trace metals and metalloids in soils and their bioavailability. 3rd (Dordrecht, The Netherlands: Springer).
Amare H., Admase H., and Ewunetu T. (2024). Influence of land-use types and topographic slopes on the physico-chemical characteristics of soils in Northwestern Ethiopia. Front. Soil Sci. 4, 1463315. doi: 10.3389/fsoil.2024.1463315
Antoniadis V., Levizou E., Shaheen S. M., Ok Y. S., Sebastian A., Baum C., et al. (2017). Trace elements in the soil-plant interface: phytoavailability, translocation, and phytoremediation—a review. Earth-Sci. Rev. 171, 621–645. doi: 10.1016/j.earscirev.2017.06.005
Becquer T., Quantin C., Sicot M., and Boudot J. P. (2003). Chromium availability in ultramafic soils from new Caledonia. Sci. Total Environ. 301, 251–261. doi: 10.1016/S0048-9697(02)00298-X
Buraka T., Elias E., and Lago A. L. (2022). Soil organic carbon and its stock potential in different land-use types along slope position in coka watershed, southern Ethiopia. Heliyon. 8, e10261. doi: 10.1016/j.heliyon.2022.e10261
Chen K. Q., Pan Y. F., Li Y. Q., Cheng J. Y., Lin H. H., Zhuo W. H., et al. (2023). Slope position- mediated soil environmental filtering drives plant community assembly processes in hilly shrublands of Guilin, China. Front. Plant Sci. 13, 1074191. doi: 10.3389/fpls.2022.1074191
Chen X., Yao Q., Gao X., Jiang C., and Fu X. (2016). Shoot-to-root mobile transcription factor HY5 coordinates plant carbon and nitrogen acquisition. Curr. Biol. 26, 640–646. doi: 10.1016/j.cub.2015.12.066
Curie C., Cassin G., Couch D., Divol F., Higuchi K., Le Jean M., et al. (2009). Metal movement within the plant: contribution of nicotianamine and yellow stripe 1-like transporters. Ann. Bot. 103, 1–11. doi: 10.1093/aob/mcn207
FAO/WHO (2021). The state of food security and nutrition in the world 2021. (Food and Agriculture Organization of the United Nations).
Fan B., Tao W., Qin G., Hopkins I., and Guo L. (2020). Soil micro-climate variation in relation to slope aspect, position, and curvature in a forested catchment. Agric. For. Meteorol. 290, 107999. doi: 10.1016/j.agrformet.2020.107999
GB/T 8303-2013 (2013). Tea - Preparation of ground sample and determination of dry matter content (Beijing: Standards Press of China).
GB 5009.268-2025 (2025). National food safety standard—Determination of multiple elements in food (Beijing: Standards Press of China).
Hao X., Xiahou L., Zhao H., Liu J. T., Guo F., Wang P., et al. (2024). CsABCG11.2 mediates theanine uptake to alleviate cadmium toxicity in tea plants (Camellia sinensis). Hortic. Adv. 2, 19. doi: 10.1007/s44281-024-00036-5
He L., Wang S., Liu M., Chen Z., Xu J., and Dong Y. (2023). Transport and transformation of atmospheric metals in ecosystems: a review. J. Hazard. Mater. Adv. 9, 100218. doi: 10.1016/j.hazadv.2022.100218
Hosseini Bai S., Gallart M., Singh K., Hannet G., Komolong B., Yin Il D., et al. (2022). Leaf litter species affects decomposition rate and nutrient release in a cocoa plantation. Agric. Ecosyst. Environ. 324, 107705. doi: 10.1016/j.agee.2021.107705
Hu C., Zhang X., Zhan N., and Liu Y. (2023). Current status and health risk assessment of heavy metals contamination in tea across China. Toxics. 11, 662. doi: 10.3390/toxics11080662
Huang W., Lin M., Liao J., Li A., Tsewang W., Chen X., et al. (2022). Effects of potassium deficiency on the growth of tea (Camellia sinensis) and strategies for optimizing potassium levels in soil: a critical review. Horticulturae. 8, 660. doi: 10.3390/horticulturae8070660
Jia M., Wang Y., Zhang Q., Lin S., Zhang Q., Chen Y., et al. (2024). Effect of soil pH on the uptake of essential elements by tea plant and subsequent impact on growth and leaf quality. Agronomy. 14, 1338. doi: 10.3390/agronomy14061338
Jiang Z. Y., Wang X. D., Zhang S. Y., He B., and Zeng Y. C. (2020). Response of soil water dynamics to rainfall on a collapsing gully slope: based on continuous multi-depth measurements. Water. 12, 2272. doi: 10.3390/w12082272
Ju Y., Luo Z., Bi J., Liu C., and Liu X. (2024). Transfer of heavy metals from soil to tea and the potential human health risk in a regional high geochemical background area in southwest China. Sci. Total Environ. 908, 168122. doi: 10.1016/j.scitotenv.2023.168122
Kochian L. V., Piñeros M. A., Liu J., and Magalhaes J. V. (2015). Plant adaptation to acid soils: the molecular basis for crop aluminum resistance. Annu. Rev. Plant Biol. 66, 571–598. doi: 10.1146/annurev-arplant-043014-114822
Lange M., Eisenhauer N., Chen H., and Gleixner G. (2023). Increased soil carbon storage through plant diversity strengthens with time and extends into the subsoil. Glob. Change Biol. 29, 2627–2639. doi: 10.1111/gcb.16641
Lei J., Yin J., Chen S., Fenton O., Liu R., Chen Q., et al. (2024). Understanding phosphorus mobilization mechanisms in acidic soil: roles of chemical and biological processes. Sci. Total Environ. 916, 170294. doi: 10.1016/j.scitotenv.2024.170294
Li X., Zhang Z., Yang J., Zhang G., and Wang B. (2011). Effects of bahia grass cover and mulch on runoff and sediment yield of sloping red soil in southern China. Pedosphere. 21, 238–243. doi: 10.1016/S1002-0160(11)60123-9
Liu J., Liu M., Fang H., Zhang Q., and Ruan J. Y. (2021). Accumulation of amino acids and flavonoids in young tea shoots is highly correlated with carbon and nitrogen metabolism in roots and mature leaves. Front. Plant Sci. 12, 756433. doi: 10.3389/fpls.2021.756433
Liu X., Tian J., Liu G., and Sun L. (2023). Multi-omics analysis reveals mechanisms of strong phosphorus adaptation in tea plant roots. Int. J. Mol. Sci. 24, 12431. doi: 10.3390/ijms241512431
Liu Y., Zhang L., Lu J., Chen W., and Lin Y. (2019). Topography affects the soil conditions and bacterial communities along a restoration gradient on loess-plateau. Appl. Soil Ecol. 150, 103471. doi: 10.1016/j.apsoil.2019.103471
Lux A., Martinka M., Vaculík M., and White P. J. (2011). Root Responses to cadmium in the rhizosphere: a review. J. Exp. Bot. 62, 21–37. doi: 10.1093/jxb/erq281
Maksymiec W. (2007). Signaling Responses in plants to heavy metal stress. Acta Physiol. Plant 29, 177–187. doi: 10.1007/s11738-007-0036-3
Mayanna S., Peacock C. L., Schäffner F., Grawunder A., Merten D., and Kothe E. (2015). Biogenic precipitation of manganese oxides and enrichment of heavy metals at acidic soil pH. Chem. Geol. 402, 6–17. doi: 10.1016/j.chemgeo.2015.02.029
Millaleo R., Reyes-Díaz M., Ivanov A. G., Mora M. L., and Alberdi M. (2010). Manganese as essential and toxic element for plants: transport, accumulation and resistance mechanisms. J. Soil Sci. Plant Nutr. 10, 476–494. doi: 10.4067/S0718-95162010000200008
NY/T 2017-2011 (2011). Determination of nitrogen,phosphorus and potassium in plants (Beijing: Standards Press of China).
Ouyang X., Ma J., Feng H., Wang X., Zhang X., and Weng L. (2023). Foliar cadmium uptake, transfer, and redistribution in Chili: a comparison of foliar and root uptake, metabolomic, and contribution. J. Hazard. Mater. 456, 131421. doi: 10.1016/j.jhazmat.2023.131421
Peng C. Y., Zhu X. H., Hou R. Y., Ge G. F., Hua R. M., Wan X. C., et al. (2018). Aluminum and heavy metal accumulation in tea leaves: an interplay of environmental and plant factors and an assessment of exposure risks to consumers. J. Food Sci. 83, 1165–1172. doi: 10.1111/1750-3841.14093
Pittman J. K. (2005). Managing the manganese: molecular mechanisms of manganese transport and homeostasis. New Phytol. 167, 733–742. doi: 10.1111/j.1469-8137.2005.01453.x
Rahman S. U., Qin A., Zain M., Mushtaq Z., Mehmood F., Riaz L., et al. (2024). Pb uptake, accumulation, and translocation in plants: plant physiological, biochemical, and molecular response: a review. Heliyon. 10, e27724. doi: 10.1016/j.heliyon.2024.e27724
Riaz A., Qin Y., Zheng Q., Chen X., Jiang W., Riaz B., et al. (2024). Cr(VI) behaves differently than Cr(III) in the uptake, translocation and detoxification in rice roots. Sci. Total Environ. 948, 174736. doi: 10.1016/j.scitotenv.2024.174736
Rotter P., Kuta J., Vácha R., and Sáňka M. (2017). The role of mn and fe oxides in risk elements retention in soils under different forest types. Plant Soil Environ. 63, 213–219. doi: 10.17221/116/2017-PSE
Ruan J., Wu X., and Hrdter R. (2019). Effects of potassium and magnesium nutrition on the quality components of different types of tea. J. Sci. Food Agric. 79, 47–52. doi: 10.1002/(SICI)1097-0010(199901)79:1<47::AID-JSFA172>3.0.CO;2-A
Shahid M., Dumat C., Khalid S., Schreck E., Xiong T., and Niazi N. K. (2017). Foliar heavy metal uptake, toxicity and detoxification in plants: a comparison of foliar and root metal uptake. Environ. Sci. pollut. Res. 325, 36–58. doi: 10.1016/j.jhazmat.2016.11.063
Tao C., Song Y., Chen Z., Zhao W., and Frost R. L. (2021). Geological load and health risk of heavy metals uptake by tea from soil: what are the significant influencing factors? Catena. 204, 105419. doi: 10.1016/j.catena.2021.105419
Uraguchi S. and Fujiwara T. (2012). Cadmium transport and tolerance in rice: perspectives for reducing grain cadmium accumulation. Rice. 5, 1–8. doi: 10.1186/1939-8433-5-5
Uraguchi S., Mori S., Kuramata M., Kawasaki A., Arao T., and Ishikawa S. (2009). Root-to-shoot Cd translocation via the xylem is the major process determining shoot and grain cadmium accumulation in rice. J. Exp. Bot. 60, 2677–2688. doi: 10.1093/jxb/erp119
US EPA (2015). Integrated risk information system; environmental protection agency (Washington. DC, USA: U.S. Environmental Protection Agency), 2015.
Wang S., Yan Y., Zhao Y. J., Fu Z. Y., and Chen H. S. (2024). Co-evolution among soil thickness, epikarst weathering degree, and runoff characteristics on a subtropical karst hillslope. J. Hydrol. 628, 130499. doi: 10.1016/j.jhydrol.2023.130499
Wen B., Ren S., Zhang Y., Duan Y., Shen J., Zhu X., et al. (2020). Effects of geographic locations and topographical factors on secondary metabolites distribution in green tea at a regional scale. Food Control. 110, 106979. doi: 10.1016/j.foodcont.2019.106979
Xiang M., Li Y., Yang J., Lei K., and Cao Y. (2021). Heavy metal contamination risk assessment and correlation analysis of heavy metal contents in soil and crops. Environ. pollut. 278, 116911. doi: 10.1016/j.envpol.2021.116911
Yoo K., Amundson R., Heimsath A. M., and Dietrich W. E. (2006). Spatial patterns of soil organic carbon on hillslopes: Integrating geomorphic processes and the biological C cycle. Geoderma. 135, 97–109. doi: 10.1016/j.geoderma.2005.01.008
Yusra Y., Khusrizal K., and Diannastiti F. A. (2020). “Soil chemical characteristics at three slope positions in the smallholder’s Piper nigrum L. @ in Lhokseumawe City, Aceh Province. IOP Conf. Ser. Earth Environ. Sci. 497, 012040. doi: 10.1088/1755-1315/497/1/012040
Zhang S., Jiang L., Liu X., Zhang X., Fu S., and Dai L. (2016). Soil nutrient variance by slope position in a Mollisol farmland area of Northeast China. Chin. Geogr. Sci. 26, 508–517. doi: 10.1007/s11769-015-0737-2
Zhang J., Yang R. D., Li Y. C., Peng Y. S., Wen. X. F., and Ni X. R. (2020). Distribution, accumulation, and potential risks of heavy metals in soil and tea leaves from geologically different plantations. Ecotoxicol. Environ. Saf. 195, 110475. doi: 10.1016/j.ecoenv.2020.110475
Zhang Q., Zhou Z. Z., Zhao W. W., Huang G. H., Liu G. F., Li X. F., et al. (2023). Effect of slope position on leaf and fine root C, N and P stoichiometry and rhizosphere soil properties in tectona grandis plantations. J. For. Res. 34, 1997–2009. doi: 10.1007/s11676-022-01582-2
Zhi X., Huang X., Zheng Z., and Hong Y. (2018). Spatio-temporal variation and the driving forces of tea production in China over the last 30 years. J. Geogr. Sci. 28, 275–290. doi: 10.1007/s11442-018-1472-2
Zhou G., Bruijnzeel L. A., Dai A., Wang F., Gentine P., Zhang G., et al. (2024). Rising rainfall intensity induces spatially divergent hydrological changes within a large river basin. Nat. Commun. 15, 823. doi: 10.1038/s41467-023-44562-8
Keywords: Camellia sinensis, slope position, nutrient absorption, heavy metals, health risk assessment
Citation: Chen L, Chen Y and Ao J (2025) Tea plantation slope position modulates nutrient and heavy metal allocation in tea plant organs: implications for health risks and sustainable plantation management. Front. Agron. 7:1670025. doi: 10.3389/fagro.2025.1670025
Received: 21 July 2025; Accepted: 08 September 2025;
Published: 24 September 2025.
Edited by:
Davey Jones, Bangor University, United KingdomReviewed by:
Nguyen Cong-Hau, Nguyen Tat Thanh university, VietnamHaizhong Wu, Henan Agricultural University, China
Copyright © 2025 Chen, Chen and Ao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Liming Chen, MjMwNjdAamR6dS5lZHUuY24=