 Christiane Lange-Küttner
Christiane Lange-Küttner Jörg Beringer
Jörg Beringer- 1Department of Psychology, University of Bremen, Bremen, Germany
- 2BeriSoft Inc., Redwood City, CA, United States
Introduction: This study investigates the long-standing Research Topic of the development of the space concept in children using a competitive, interactive, online, zero-sum game that required players, taking turns with a computer player, to use monochromatic electronic paint to complete coloring in places until an entire 10 × 10 grid was filled.
Methods: Children were 4- to 5-year-olds (n = 26), 6- to 7-year-olds (n = 32), 8- to 10-year-olds (n = 52), as well as adults (n = 48). The total sample consisted of 79 human and 79 machine players (N = 158). The human players colored in places on the grid in yellow, and the machine player in green. While human players could follow their own spatial heuristics, the machine system player was programmed to randomly color in anywhere in the grid. Neighboring places could visually merge into pathways or areas.
Results: Children explored the array somewhat less than adults, but both explored it less than the machine, which randomly colored in places across the entire array. Both children and adults as well as the machine players created one large area and many smaller ones. Their one large area left a large swath of the spatial array empty. This space was then filled in by the machine player at random places, which merged into a slightly smaller large area or many larger small areas.
Discussion: Hence, a similar outcome was achieved regardless of whether spatial concepts or random spatial explorations were devised. The use of spatial heuristics was demonstrated with correlational analyses, where long pathways and short-distance moves were significantly correlated with the largest area, but not in their machine opponents.
Introduction
This study is an experiment that contributes to the gamification of children's space concept (Long et al., 2023). An interactive grid game was created to investigate children's concept of aggregate space (Lange-Küttner, 2009), where an area is conceived of as an accumulation of places rather than a continuous expanse of Euclidean space. The concept of aggregated places is also the basis of the spatial layout of many board games like chess or Go. In the current study, children and adults played against a randomly behaving computer in an online coloring-in game. This game was zero-sum insofar as each player could not color in more than half of the places of the grid. It was interactive to the extent that the players were taking turns, each one coloring in places with their own color. Finally, it could be competitive because if a player did not just randomly color in individual places anywhere in the grid but was planning to build a cluster of places that would perceptually merge into a monochrome area, the other player could thwart that strategy by coloring places and patches within that region. Although the machine was programmed to only use a random strategy, the result of the current study was that the outcome of the game between the human and the machine players is structurally very similar insofar as both created one large major cluster of places distinct from many smaller clusters. To analyze the spatial heuristics of children and adults, we also measured the length of consecutively colored-in adjacent places creating spatial pathways, as well as the average distance between places and the average distance of the next move to a place anywhere in the grid.
Sequences and areas
Piagetian theory asserts that Eucledian space concepts develop from topological space concepts that are figurative, local, and practical (Piaget, 1977). For instance, children argue that the distance between A and B is reduced (and thus not conserved) when an object C is placed between them, taking away space (Liben, 1982). But there are also experiments on children's construction of sequences and spatial fields (Piaget and Inhelder, 1948; Piaget et al., 1948) that show that some wiggly spatial pathways develop early and that only straight spatial pathways take some time to develop (Flavell et al., 1991). Moreover, the spatial field also exerts an influence (Lange-Küttner, 2024). When Piaget asked children to connect two places A and B in a rectangular or circular array, at Stage 1, from 2 1/2 years of age onward, children would already sequence objects. The resulting lineup was not yet straight but meandered, nor did the lineup successfully connect A and B. Such sequences are called “threading” (Vinter et al., 2008).
At Stage 2, the connection between A and B is achieved, but a curious spatial field effect occurs where array edges or corners work as attractors (see also Hund and Plumert, 2005), or an orthogonal bias occurs which is also known from children's drawing of geometric angles (Bremner, 1984; Perner et al., 1984). Only at Stage 3 are A and B connected with a sequential lineup that forms a straight line, even if it is a diagonal.
Proximity when sequencing objects also plays an important role in the experiment on the conservation of areas (Piaget et al., 1948), see Figure 1. A lack of area conservation shows when, in their verbal response, children maintain that two areas are not the same size because there are differences in how they are occupied by figures in different configurations and numbers. The experiment requires children to answer the question which cow would have more greens to eat: the one whose farmer built terraced houses on array A or the one whose farmer built his houses in a random fashion dispersed across array A′? Children have no difficulty in saying that the green is identical (area conservation) as long as the houses are distributed in the same spatial pattern in both areas. However, if the houses are lined up, children up to 5 1/2–6 years of age notice the vast swathe of empty space and claim that the cow on this array has more to eat. In support of the children's view, one could say that the attached adjacent houses on the array A are of a different quality than the single houses on the array A′ as they form a new object, namely, a “terrace.” Moreover, in practical terms, in total, cows would need many shorter paths to surround the single houses than a row of terraced houses. In Piagetian theory, this would have been conceptualized as a practical grouping. At Stage 2, the notion of area equality of the two meadows is dependent on the number of houses as children's area conservation breaks down when numbers of houses increase and the more the perceptual appearance of the two meadows diverges from each other.

Figure 1. Conservation of areas: available space on two meadows with terraced and detached houses (illustration of the experiment by Piaget et al., 1948, chapter XI).
This would be equivalent to a subjective grouping because idiosyncratic factors are the basis of an argument for why the two areas are different in size. However, after age 6, at Stage 3, children are simply counting the number of houses on each array and claim that the same space is left for both cows, showing their grasp of an abstract, geometric concept of area based on numerical computation.
Nevertheless, although proximity seems to play such a crucial role in young children's thinking, the Common Region Test (CRT) shows that young children also seem to have a strategy that isolates objects from each other. For instance, when asked to allocate a region around objects that are closer together or similar in appearance, up to 5 years of age, children would allocate individual places to a shape rather than a region; see Figure 2A. Only older children allocate a common region to pairs of shapes that are closer to one another than to others, and this spatial categorization strategy gradually increases with age (Lange-Küttner, 2006). Using a common region as a grouping principle (Palmer, 1992) greatly facilitates visual memory for places (Lange-Küttner, 2010a,b).
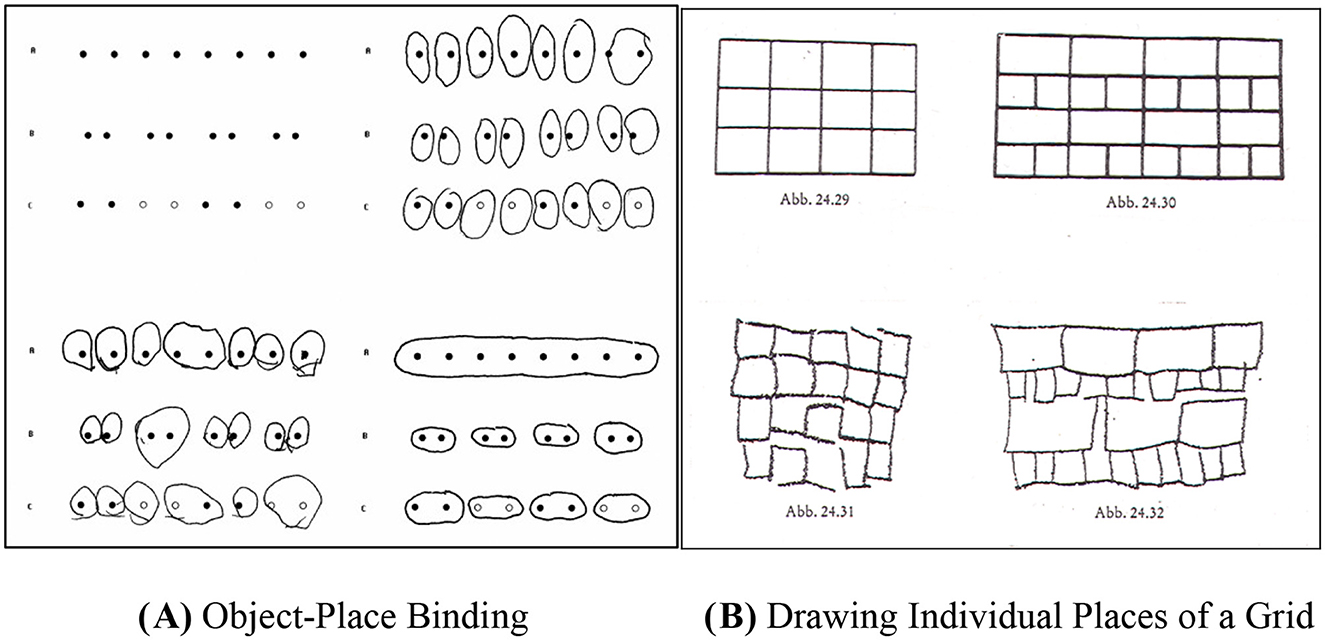
Figure 2. Two examples of aggregate space. (A) Common Region Test: Wertheimer array (upper left), object-place binding (upper right), object-region binding (lower right), and unsystematic coders (lower left; Lange-Küttner, 2006, with permission of the British Psychological Society). (B) Copying of a grid by preschoolers (Metzger, 1956, p. 541).
Another impressive example of developmental change in spatial grouping is children's changing understanding of the grid; see Figure 2B. Preschoolers would copy a grid as a wall that is built “brick by brick” without intersecting spatial axes (Metzger, 1956, 2006). Also other researchers found that young children up to the age of 5 prefer to segment spatially independent parts (Stiles and Tada, 1996), which has been termed aggregate space (Lange-Küttner, 2009). Children have no explanation for their object–place encoding. Against the backdrop of the Piagetian cow-in-the-meadow experiment on area conservation, one could assume that when drawing object–place bindings in the CRT, young children think action-based in terms of dynamic spatial pathways: Shapes are surrounded with a pencil like the cows would need to surround the randomly dispersed houses in the meadow when grazing. It is known that young children initially draw extremely action-based, hopping with a pencil like a rabbit over the page and only post-hoc interpret these traces (Freeman and Adi-Japha, 2008). Piaget (1977) always emphasized that logical thinking would develop from this kind of practical, action-based intelligence.
Interestingly, children who encode object–place units are better at remembering individual shapes, while children who allocate regions to matching objects are better at remembering locations (Lange-Küttner, 2010a,b, 2013). Spatial binding of matching shapes into common regions was especially important when children had to remember new memory sets with different spatial configurations. In contrast, when they remembered always the same objects in always the same places, spatial binding was not relevant, and memory performance improved just through repetition (Lange-Küttner and Küttner, 2015).
The current study investigates the binding of places into a common region. This is different from binding matching objects into common regions. It is known that objects and placeholders help young children remember places (Lange-Küttner et al., 2023). So the current task may appear simple because no figures or shapes are involved, but it tests whether there is a change in the way areas are produced from individual places.
The current study
Coloring in a grid in an interactive game paradigm has been used with adults before (Lange-Küttner and Beringer, 2023). The human player colors places in green, while the machine colors the places in yellow. If adjacent places were colored in, they merged into an area. The interactive experiment was programmed as a non-competitive game. Once a place in the grid was colored in, it could not be colored in by the computer system and vice versa, if the human had colored in a place, it became unavailable for the computer system. Thus, unlike in a game like Go, where regions can change hands, it was a zero-sum game where no territorial gains could be made, nor was there a race to be first. Instead, the grid game is an “alignment game,” where completing a configuration is the main aim (Gobet et al., 2004), similar to puzzles but more interactive.
It is known that the grid size is a crucial variable in grid games, even for artificial intelligence as, for instance, the larger and uniform 19 × 19 grid of Go is more difficult than an 8 × 8 chessboard (Silver et al., 2016). Hence, in a first pilot comparison, a 10 × 10 children's grid was smaller in size than the 20 × 20 grid of the adults in order to equate task difficulty in terms of cognitive load. Also, beginning players of Go often start with a smaller 9 × 9 or 13 × 13 grid (Matthews, 2003). The reason is that a larger grid size offers more possible moves1 that can and need to be considered. A larger grid offers more degrees of freedom and lower spatial constraints when selecting a place. A larger grid thus requires more computing power if all the possibilities of potential moves are explored, a cognitive process called “brute force” in comparison to a “heuristic,” which consists of a simpler, more selective strategy that is more economical in terms of computing power (Gobet et al., 2004).
In the current grid game, children and adults take turns with the computer system (henceforth called the “machine”) to color in each place in the grid in their own color. Coloring-in is a common activity for children, with ~12% of children's books sold in the United States being coloring and activity books, accounting for more than 104 million units and amounting to more than U.S. $230 million in annual revenue (Fitzpatrick and McPherson, 2010). Coloring-in is also supposed to relieve test anxiety in children (Carsley et al., 2015), most likely because it is a familiar activity that usually does not have achievement goals and is pre-structured with outlines and contours. In fact, coloring-in objectively reduces the stress hormone cortisol (Kline et al., 2020). Digitizing coloring-in improves the appeal of drawing for children (Tokuhisa and Kamiyama, 2010).
Strategic and random spatial strategies
Here we have made coloring-in somewhat more challenging. We test whether and how children and adults create areas by coloring in places in an online grid against a randomly behaving computer system player. The computer system responded to each move by a child or an adult by coloring in a random place in the array in a visible but unpredictable way for the participants.
That the coloring-in game was not competitive in the sense that a player could gain more ground than the other player—as each player could color in exactly 50% of the places—was not mentioned to the participants. It was also not mentioned that the machine, as the interactive game partner, would color in places randomly—although players could, of course, quickly realize that this was the case. As the aim of the game, it was only stated that the grid should be completely colored-in at the end, taking turns with the machine. However, should a player have a certain plan, the randomly responding machine could potentially get in the way. For instance, if human players had a plan to color in a green 3 × 3 square in the middle of the grid, the machine could randomly color in one of these nine places in yellow, ruining this square shape. In this sense, the game was competitive even if no territorial gain could be made. In a previous study with adults, coloring in places was sensitive to instructions (Lange-Küttner and Beringer, 2023). Compared to no explicit instruction, adults would color in faster if instructed to do so, resulting in smaller areas. They would also create larger areas if instructed to do so. Their areas were in between in size when instructed to create large areas fast. This showed that adults' playing behavior was flexible and adaptable.
A similar study was carried out by Falk (1981, cf. from Bar-Hillel and Wagenaar, 1991), which we did not know of at the time we created the task for the current study. Falk explicitly asked participants to randomly color in 50 places of a 10 × 10 grid in green vs. yellow. How many places bordered another place with a different color (alternation rate) was counted. The alternation rate in this coloring-in game and in card sequencing with 20 yellow and 20 green cards that were to be sorted at random, was a rate of 0.6 which is very close to selecting places and cards by a chance of 0.5, given that there were two colors. Interestingly, the longer a sequence of places, the less likely it was that adults created a randomly emerging sequence (Kubovy and Gilden, 1991). Another factor that may get in the way of adults' notion of randomness is that with rare exceptions, they were very good at remembering a random sequence that they created in a previous session, with correlations between 0.78 to 0.98 (see also Towse and Neil, 1998; Treisman and Faulkner, 1987). Turing (1950) states that in a game, including a random element in a learning machine would be wise as it could be more useful than searching for an optimal solution through a decision tree. There would be the additional advantage that a random strategy makes keeping track of values unnecessary. In fact, in chess, players appear to be bored by winning chess when having a supreme long-term memory for chess moves and have now embarked on a freestyle chess, where the pieces on the back row—king, queen, and so on—start in a randomized position (Agini, 2025). There is even an inaugural Freestyle Chess Grand Slam tour planned, with investors lined up.
In the current game, the computer randomly colors in places. In principle, we expected that because children understand aggregate space earlier than Euclidean space, the younger children would be able to solve this task as well as the older children. But we had no hypothesis on how they would react to the machine's stochastic behavior. In a pattern recognition task with stochastic feedback, children's accuracy was massively reduced (Hentschel et al., 2022, 2023; Lange-Küttner et al., 2012), and this lasted into adolescence, albeit not to the same degree (Lange-Küttner et al., 2021). Would children color in places in the same random way as the machine? The ability to randomly generate items such as numbers or letters is agreed to be an indicator of executive attention as it demands the inhibition of “natural” sequences, such as the number line or the alphabet, and increases in childhood (Towse and McLachlan, 1999) and decreases in dementia (Brugger et al., 1996). Thus, in light of this earlier research, it could be assumed that children and adults are unlikely to use the same random spatial strategy as the computer in the coloring-in game. We would rather assume that they use a heuristic to build a pathway out of a sequence of consecutive places and merge places that they have colored in with their color into monochrome areas. We thus measured (1) the size of clustered places, (2) the sequence lengths that participants built by lining up places, (3) the average distance between places, and (4) the average distance of the next move. We use the term cluster for places and area as the equivalent to clustered places of one color that produce a monochrome area.
Regarding the cluster size of places, we first predicted that young children would also be able to create large clusters as they are sensitive to practical, aggregate space while the machine could not have aims as it was programmed to randomly color in places. However, the younger age groups could also show an inclination toward coloring in isolated places; thus, their pathway creation was not for certain. Second, regarding the length of the place sequences, we predicted that some pathways would emerge in all human participants, while the machine could not form pathways because of the programmed random coloring-in. Third, while the average distance between places in the outcome of humans and machine may be similar, the average distance to the next move is expected be smaller in human players, because the machine, with its fixed random strategy, would color in places across the entire array. Thus, regarding intentionality and strategy, the fourth hypothesis is that the machine-generated areas would be the result of places that incidentally aggregate into areas, while the child- and adult-generated areas would be intentional and thus should significantly correlate with the pathways while they create a cognitive map of the grid during the experiment (Bostelmann et al., 2020; Herman, 1980).
Methods
Participants
A sample of N = 62 children from a lower middle-class area in North London, United Kingdom, completed the task. Preschool as well as primary school children were recruited as the task was doable for all age groups. There were three age groups, 4- to 5-year-olds, 6- to 7-year-olds, and 8- to 10-year-olds; see Table 1. The sample was recruited via a teacher at the school by contacting parents via email, which contained a link to the experiment. For seven children, the wrong date of birth was provided online (actual year instead of year of birth), which made allocating their data to an age group impossible. Thus, their data sets were omitted from analyses, resulting in a final sample size of 55. Children were described by their parents as Asian (12.7%), Black (10.9%), mixed (16.4%), and white (60%). There were 30 female (54.5%) and 25 male (45.5%) children. A sample of 22 mothers aged 21–46 years volunteered to take part using an adult 20 × 20 version of the grid. However, these data are not included in the current report because of the complexity of the statistical analysis. Instead, only the data from a second group of a more gender-balanced sample of mainly white adults (n = 24; 58.3% female) of about the same age range who solved the task using the same 10 × 10 grid was included in the current analyses. In the age group description in Table 1, the machine players were considered participants because they were the game partners for the participants. Thus, both the machine and the human player groups constituted the between-subjects factor.
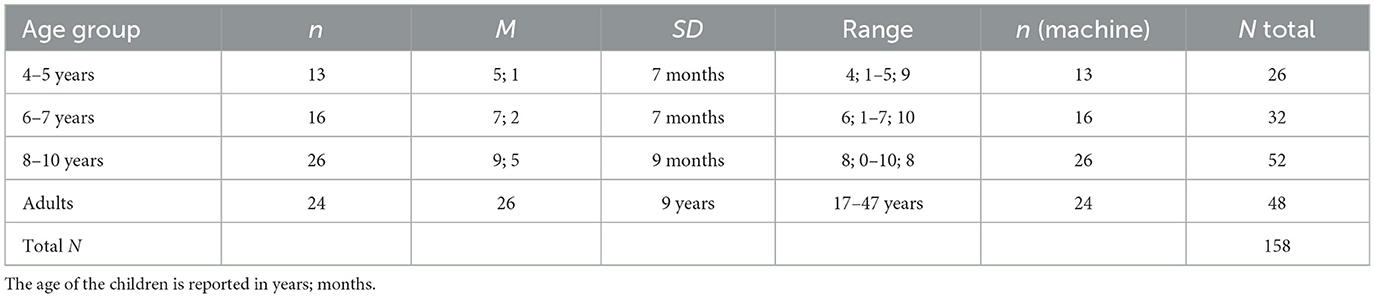
Table 1. Age group description.
Material and apparatus
The experimental code is deposited on OSF Depository (https://osf.io/tjgkn/). The coloring-in experiment could be run in a browser window on any computer system. The places in the grid could be clicked into with a mouse or tapped on with a finger on tablets with a touch screen to turn them colorful. The place color of the human participant was green, and the machine player, yellow; see Figure 3.

Figure 3. Coloring in the 10 × 10 grid. Green = human (dark gray), yellow = machine (light gray), and black = empty (not used).
Procedure
The experiment's ethics proposal was approved by the Ethics Committee of the Psychology department of the London Metropolitan University, London, United Kingdom, where the first author worked at the time. The experiment was run in Cognition Lab, https://www.cognitionlab.com. Because of COVID-19 restrictions, parents were given an individual link to the child experiment by email through the school. If they felt like coloring in places themselves, they could take part in the adult 20 × 20 version of the experiment.
They had to agree to the consent form online before they could start with the experiment. If they declined, the experiment did not go ahead. If parents or children pressed the escape button during the experiment or closed the browser, the experiment was also aborted, and no data were saved. Parents supervised children to complete the coloring-in task. The instructions were “Welcome to the Grid Game! Please press the space bar to continue or tap on the continue button! This is a coloring-in game with the computer. YOU click into or tap on one place; the COMPUTER clicks into another place. Please always wait until it is your turn. Keep going until all the places are colored in.” Once a place was colored in, it could not be used again. Parents read aloud the instructions on the screen to the children and were advised to be present but not active. Data from the completed sessions were stored on an European Community server anonymously with a random number code and without an Internet Protocol address.
Data generation
The raw data were downloaded as one Excel output file from Cognition Lab and processed with R scripts accessible on the OSF Depository ‘Coloring-In Places Game (Children-Adults)' (https://osf.io/tjgkn/). Two PDF files per experimental session were generated; see Figures 4A, B.
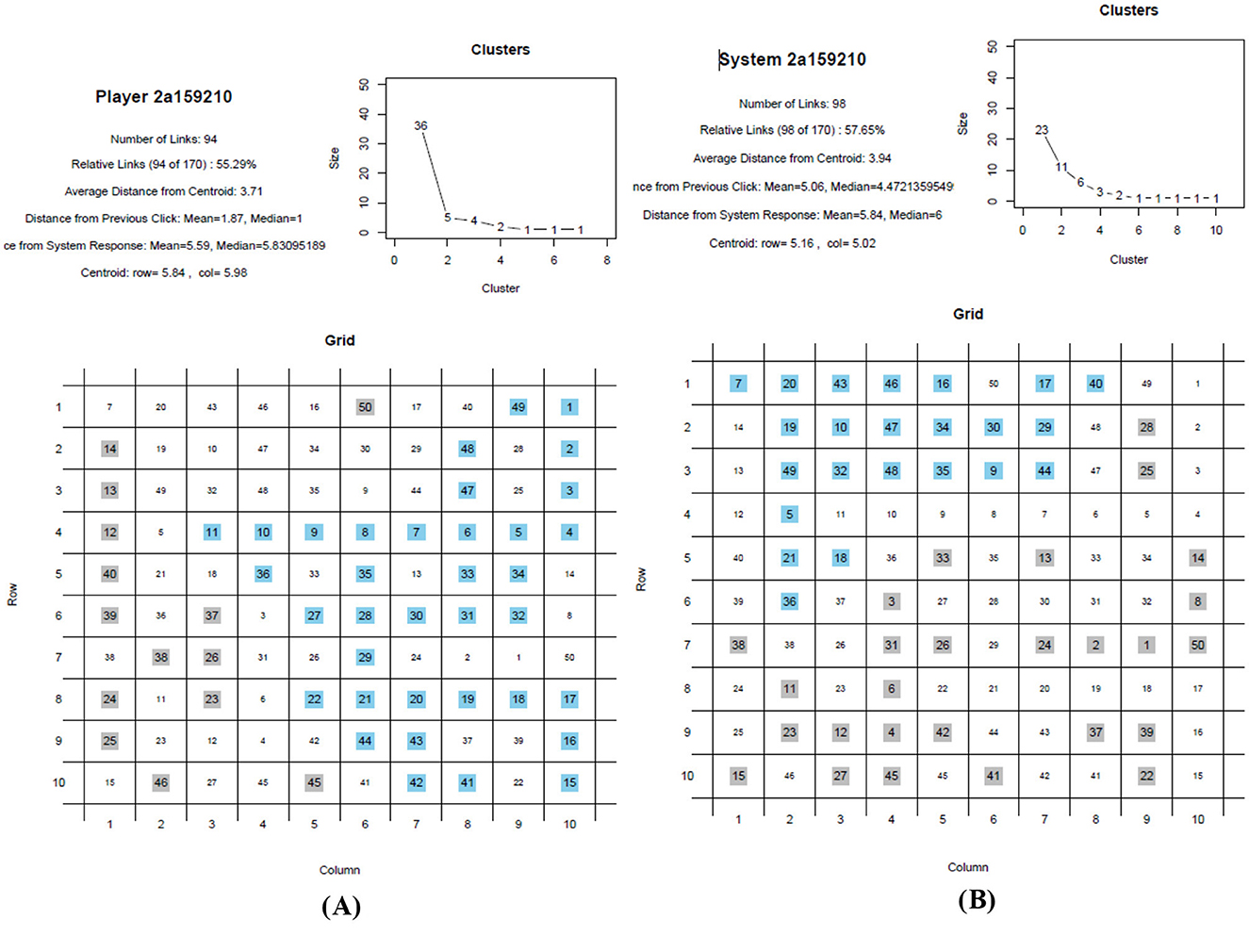
Figure 4. Graphic representation and data generation of coloring-in moves place by place. (A) Human player moves. (B) System player responses.
On each PDF, the human and machine players' selected places were visualized in figures that showed the numbered sequence of the moves as well as the cluster sizes. The next step was to average the trials per participant, human or machine, for import into SPSS with another R script. Two Excel sheets were generated with two further R scripts, one for the human players and one for the machine players; see the OSF Depository (https://osf.io/tjgkn/). We computed and analyzed (1) the eight largest area clusters because this was the cutoff point where data were available for all participants and (2) the longest sequence, as well as (3) the average distance of all places and (4) the average distance of a move to the next place anywhere. Places were counted as belonging to a cluster when they had at least one side in common with another place of the same color. Places of the same color touching at the corners were not counted.
Places were counted toward a sequence (pathway) when (1) they were colored in one after another and (2) they had one side in common with the next colored-in place. For sequence length, it did not matter whether the consecutive place was added to the side, upward, or downward in direction as long as one side of two consecutive places shared a boundary. Diagonal sequences were not considered. SPSS data are accessible on the OSF Depository (https://osf.io/tjgkn/).
Data analysis
We used analyses of variance using SPSS. We analyzed (a) the cluster size to test the size of the area that was built (human) or emerged (machine), (b) the length of sequences of consecutively colored in adjacent places to test the length of the pathways that were pursued, and (c) the average distance, and the distance of the next move to assess the exploration. When Mauchly's test of sphericity was significant, the degrees of freedom were adjusted according to the Greenhouse–Geisser correction. Pairwise comparisons within the model were corrected using SPSS according to the Bonferroni correction. Correlation heat maps were produced using JASP.
Results
First, the presence of sex differences was screened using t-tests for independent samples (two-sided). For this purpose, the children's age groups were collapsed. The cluster size was not significantly different for boys and girls for any of the eight clusters, ps > 0.459, nor was the average distance, t(53) = −0.24, p = 0.809, or the distance of the next move, t(53) = 0.66, p = 0.509. The same was true for the adult participants: The cluster sizes for any of the eight clusters were not significantly different, ps > 0.074, nor was the average distance, t(53) = 0.09, p = 0.926, or the distance of the next move, t(53) = 0.61, p = 0.545. Thus, sex differences were not further considered in the statistical analyses.
Cluster size (area)
The table with means, range, and SD can be found in Supplementary Table S1. An 8 (cluster) × 2 (human/machine) × 4 (age group) mixed analysis of variance (ANOVA) with repeated measures of cluster size was conducted. The effects of age group, F(3, 158) = 2.36, p = 0.073, and the interaction of age by machine, F(3, 158) = 0.40, p = 0.754, were not significant.
Cluster size was significant as a main effect, F(1.19, 158) = 691.27, p < 0.001, with a large effect size of η2 = 0.82. The cluster size varied from M = 30.88 to M = 6.87 to M = 3.79 to M = 2.26 to M = 1.60 to M = 1.16 to M = 0.87 to M = 0.67. Note that the last two clusters consist of isolated single places as the cluster size fell below 1. Pairwise comparisons within the model demonstrated that all differences were highly significant, ps ≤ 0.001 (see Appendix Table A1), showing not only that the largest cluster differed from the smaller ones but that the smaller cluster sizes also differed from each other. The decrease in cluster size is logical because as more places are colored in and integrated into self-contained monochrome areas, less space is left for the remaining clusters. This significant main effect of cluster size revealed the strong spatial constraints that were generated by building areas within the grid.
The eight cluster sizes had a two-way interaction with human vs. machine responses, F(1.19, 158) = 5.44, p = 0.016, η2 = 0.04 (see Figure 5).
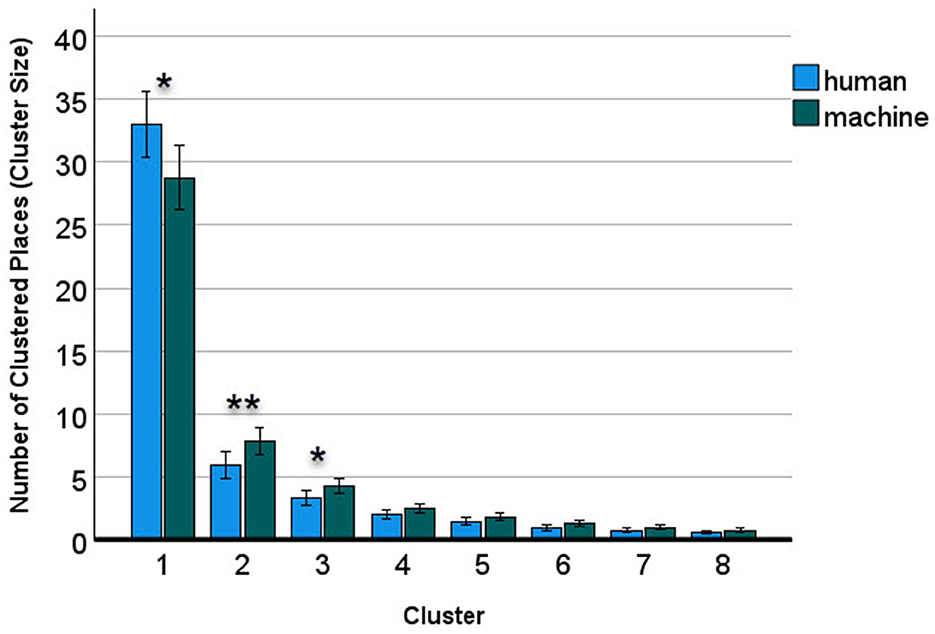
Figure 5. Human advantage in the largest cluster, machine advantage in the smaller clusters. The error bars denote the 95% confidence interval. * = p < 0.05; ** = p < 0.01.
Post-hoc tests for independent samples of humans and machine (two-tailed) showed that humans (M = 32.6) were at an advantage against the system player (M = 28.4) with respect to the largest cluster, t(156) = 2.35, p = 0.020, but it was the reverse for the second, t(156) = −2.92, p = 0.004, and third smaller clusters t(156) = −2.39, p = 0.018 (see Appendix Table A2). This result showed that the human participants were able to build a larger area in the grid under low spatial constraints. However, under high spatial constraints caused by a large cluster already occupying a large area, a human advantage did not materialize. On the contrary, the system player built reliably larger small areas than humans in two clusters out of the seven remaining ones. There was no significant interaction of age with the human/machine factor, F(3.56, 158) = 1.64, p = 0.173, nor was the three-way interaction with cluster significant, p > 0.360.
Length of sequences of consecutively colored-in adjacent places
The length of sequences of adjacent consecutive places, one after another, side by side, was assessed to test for a coloring-in strategy. A univariate 2 (human/machine) × 4 (age group) mixed ANOVA was run with the longest place sequence length as a dependent variable. There was a significant difference between human and machine, F(1, 158) = 74.44, p < 0.001, = 0.33. The length of spatial pathways was, on average, more than twice as long (M = 5.34) than those of the machine (M = 1.90). The age group effect was not significant, neither as a main effect, nor in interaction with the machine, ps > 0.366. This showed that there was no difference in pathway length between children and adults.
Average distance and distance of the next move
Average distance
The average distance between all places of one player and the distance of the next move were indicative of spatial exploration of the grid. One could expect that because the machine was coloring in randomly, the use of the spatial grid would be more extensive than in humans. The univariate 2 (human/machine) × 4 (age group) mixed ANOVA with average distance between places as dependent variable showed a main effect of machine, F(1, 158) = 13.25, p < 0.001, = 0.08. On average, the distance between colored places was M = 3.84 for the machine, and M = 3.67 for the humans showing a small but significant average distance effect for the machine compared to the humans. There was no significant effect of age or interaction of age with the human/machine factor, ps > 0.135 (see Figure 6A).
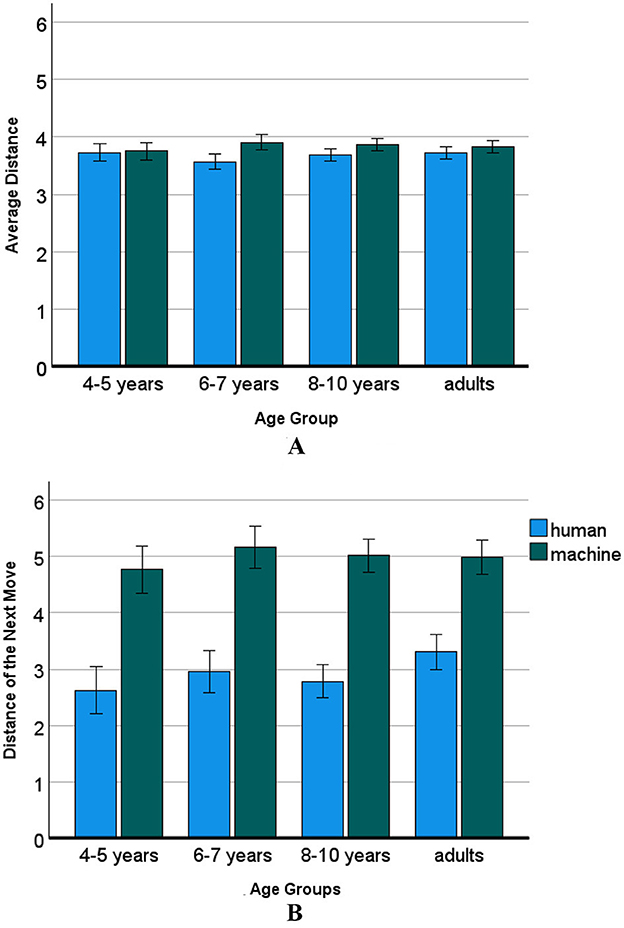
Figure 6. Machine advantage in average distance of next move (grid exploration). (A) Average distance. (B) Distance of the next move. The bars denote the confidence interval. The distance of the next move is significantly farther away for the machine than for humans. Age effects were not significant.
Average distance of the next move
While the average distance between places was a general measure of the distance between colored-in places of a player, the distance of the next consecutive move to anywhere in the array was more indicative of reaching out within the spatial array. The univariate 2 (human/machine) × 4 (age group) mixed ANOVA with distance of the next move places as dependent variable showed a main effect of machine, F(1, 158) = 268.26, p < 0.001, with a medium effect size of = 0.64. On average, the machine made moves that were approximately five places away from the previous move, M = 4.98, showing more exploration of the grid than the humans who colored in places approximately three places away from the previous one, M = 2.92. The effect of age group did not reach significance, F(3, 158) = 2.27, p = 0.083, =0.04, nor was the spatial exploration of the human/machine factor interacting with the age of the human players, p = 0.251 (see Figure 6B).
Are the strategies of place sequencing and area building related?
To be sure that in humans, but not in the machine, the strategy of coloring-in consecutive places into a sequence contributed to area building, correlations were separately computed for machine and human responses. The Supplementary Table S2 (children) and Supplementary Table S3 (child machine opponent), as well as Supplementary Table S4 (adults) and Supplementary Table S5 (adult machine opponent) show the Pearson correlations, p-values, and visualization of the distributions (please view at 200%). Children's age groups were collapsed because there were no significant age group differences for either area or sequence length. The longer pathway and the shorter next move were correlated in children, r = −0.68, but not in the child machine opponent, r = −0.09.
In Figure 7, the correlations of the cluster with pathway length (max. sequence) and array exploration (next move) are visualized numerically with the strength of the p-value in color. For children, the most striking difference to be found in the comparison of Figure 7A and Figure 7B is that pathway and next move show significant correlations with the clusters, but not in the child machine opponent. The shorter pathway length of the machine has close to zero correlations with the clusters, and the correlations of the next move are clearly smaller in size compared to the children. In particular, the shorter distance of the next move in human children is highly correlated with the largest cluster, r = −0.60, but not as much in the child machine opponent, r = −0.15.
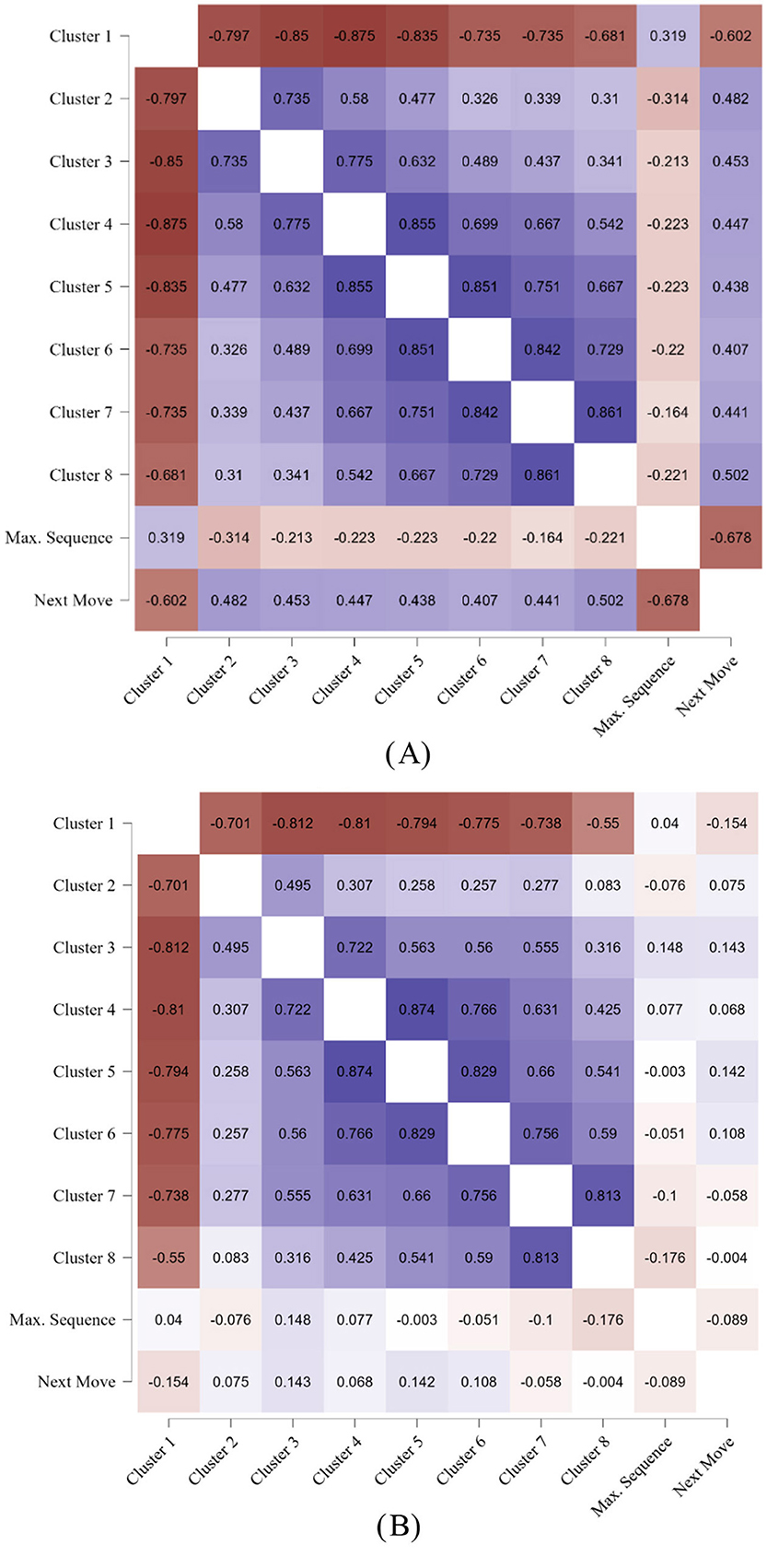
Figure 7. Children's correlations between clusters, pathway length, and distance of the next move. (A) Children. (B) Child machine opponent.
However, it is noticeable that there is a systematic difference in positive and negative correlations. In children and the machine opponent, all clusters were negatively correlated to the largest cluster, and positively with each other; see Figures 7A, B. However, the pathway (maximum sequence) was only positively correlated with the largest cluster in human children, r = 0.32, showing that creating a longer place sequence contributed to building the largest cluster, but not in the child machine, r = 0.04. The shorter pathways were negatively correlated with the much smaller clusters (<10 places).
In adults, most of the associations of the clusters with the maximum sequence and the next move showed similar correlational patterns as in children, see Figures 8A, B. Again, the smaller clusters were negatively correlated with the largest cluster but positively correlated with each other. And again, longer sequences were positively correlated with the largest cluster in human adults but not in the machine opponent, indicating a human strategy. Likewise, pathways and next move were significantly correlated in humans, r = −0.68, but not as much in the machine opponent, r = −0.20.
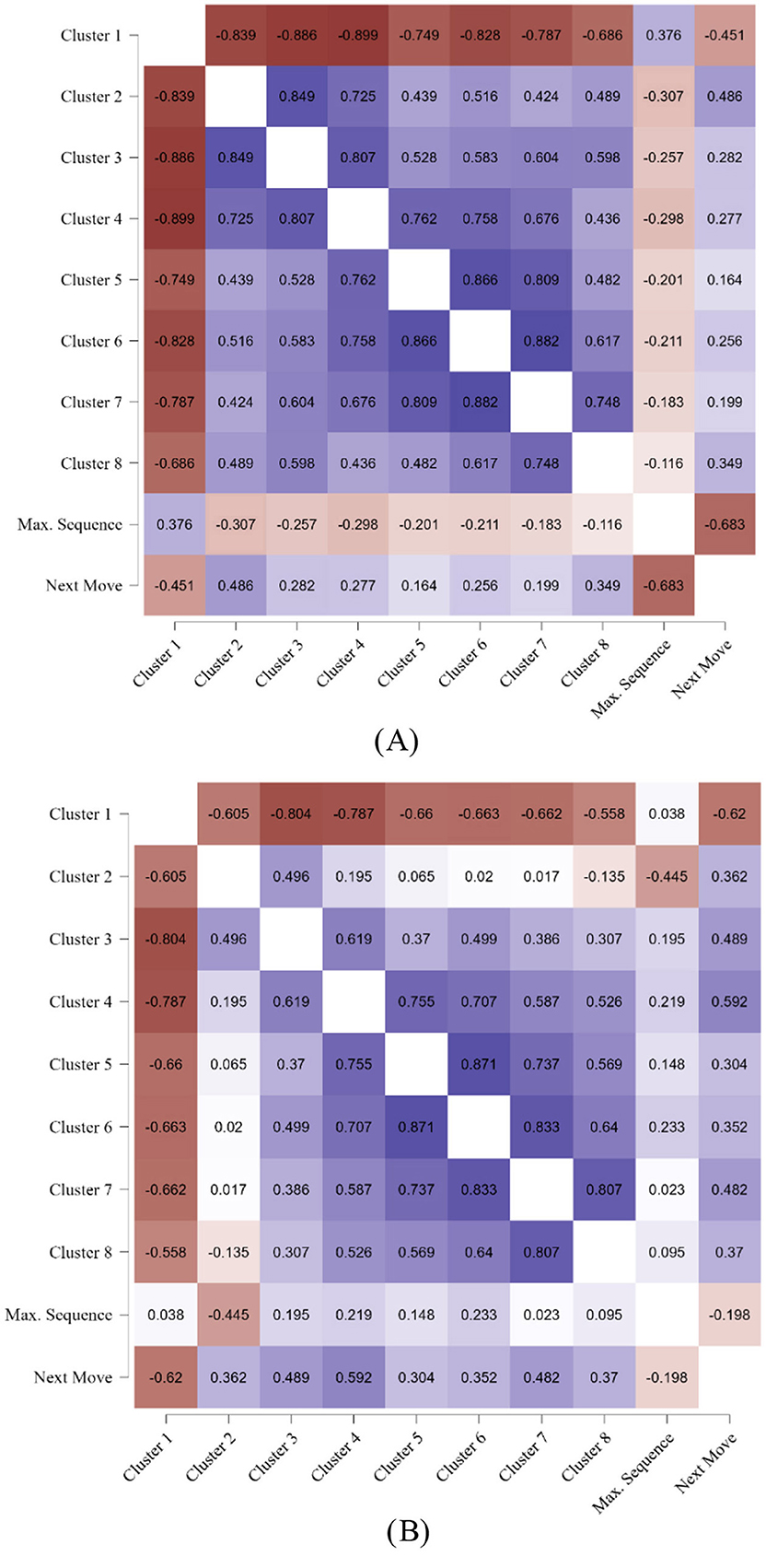
Figure 8. Adults' correlations between clusters, pathway length, and distance of the next move. (A) Adults. (B) Machine Opponent.
Regarding the machine, near-zero correlations occurred between cluster 2, that is, the first cluster in which the machine had created a larger cluster than the humans, and the smaller clusters 5, 6, and 7. However, this should not be overinterpreted because when comparing it to the cluster correlations of the children's machine opponent in Figure 7B, the correlations of cluster 2 are also smaller as visible by the lighter color.
A real difference to the correlations of the children, however, was that in both, the human adults and their machine opponent, the next move was pronouncedly negatively associated with the largest cluster, not just in the human players. We remember that for the whole sample, the machine made moves that were, on average, approximately five places away from the previous move, while humans colored in three places away from the previous one. There was a trend for an age group, p < 0.083, which was not further explored, but it could be that the age effect did not reach significance because there were four age groups. When comparing the collapsed age groups of children with adults in a univariate 2 (human/machine) × 2 (age group) mixed ANOVA with distance of the next move as dependent variable and children/adults as between-subjects factor, indeed a significant interaction of age by machine showed, F(1, 158) = 4.10, p < 0.045, = 0.03, in addition to the highly significant main effect of the machine exploring further in the array, F(1, 158) = 217.93, p < 0.001, = 0.59.
Post-hoc t-tests (two-tailed) of the interaction showed that the two machine opponents carried out exactly the same average next move, Ms = 5.0, t(77) = 0.17, p = 0.865, while children explored less (M = 2.79) than adults (M = 3.30), t(77) = −2.16, p = 0.034.
Discussion
Children are not always systematic in their spatial categorization of regions, especially not those with special educational needs such as autism and attention-deficit hyperactivity disorder (Lange-Küttner and Kochhar, 2020). They initially construct just object–place units (Lange-Küttner, 2006; Metzger, 1956; Stiles and Tada, 1996), a strategy which is based on an aggregate space concept (Lange-Küttner, 2009), and only later attribute regions to objects that match in some way (Lange-Küttner, 2006). The current study investigates the binding of places into a common region. This is different from binding matching objects into common regions as it involves creating areas without considering objects. As such, it is perhaps similar to the board game Settlers of Catan, where areas are created by each player (Guhe and Lascarides, 2014) and it also does not have a necessity to consider for existing boundaries (Veracini, 2013). Likewise, in the game Monopoly, streets (and thus pathways) can be traded without consideration of objects (Maulyda et al., 2020). Whether objects in a region match or not is often a matter of contention, if not war (Flanagan and Jakobsson, 2023). Nevertheless, understanding and computing area is a core subject in the teaching of geometry for a long time (Jones and Tzekaki, 2016; Luchins and Luchins, 1947), and it is assumed that a natural geometry already belongs to infants' core knowledge (Spelke, 2011). These basic geometric intuitions are already present in animals and can give rise to more abstract representations of area (Spelke and Lee, 2012). While much of this research is based on perception (Izard et al., 2022; Odic et al., 2013), the current study uses a constructive and interactive task. An especially important feature of this new task is that while the computer system, as the interaction partner, had no choice but to randomly select an empty place, children could have selected to color in places either randomly or with a strategy.
The first hypothesis, that young children would also be able to create large clusters as they are sensitive to practical, aggregate space could be confirmed. The study clearly showed that they were able and motivated to build large areas in a grid coloring-in game against a randomly behaving computer system. Like adults, children between 4 and 10 years created one large area and several smaller ones. The younger age groups could also have shown an inclination toward coloring in isolated places because theoretically, it would have been possible to either color-in randomly like the computer, or to construct several clusters of the same size. However, a large primary area was larger when colored in by human participants, children and adults, while the smaller clusters were larger when colored in by the machine. Nevertheless, both humans and the machine produced one large area and multiple smaller clusters. The six smaller clusters were a logical consequence of one large area that imposed strong spatial restraints, with an effect size of 0.83. The one large area built by the machine was an antagonist result of the human strategy: As the human player creates one large area, it leaves the rest of the array to the machine, which would consequently have the other area to itself.
This result is very much in accordance with the reasoning of preoperational children who were thinking in action-based spatial pathways rather than about areas in geometric terms and argued that cows have more to eat when grazing on a large empty meadow than on a meadow that is occupied by several houses dispersed in this area (Piaget et al., 1948). The dispersed houses would need several walk-arounds while a row of terraced houses would just be one entity. Likewise, if the humans would not have embarked on building a large area, the machine could not have had the large empty swathe of the remaining space at its disposal to build an area matching in size, just by chance, without a concept, just with randomly coloring in places. An area can emerge from randomly filled-in places or as a result of intentionally planned clusters.
The advantage of the humans for the largest cluster was the first indicator that they were following a strategy, and thus, we tested several other parameters to prove that humans were actually following a strategy in this coloring-in game. The second hypothesis predicted that sequences of places forming pathways would emerge in all human participants, while the machine could not form pathways because of the programmed random coloring-in. Also, the second hypothesis was confirmed as both children and adults built on average sequences that were twice as long as those of the machine player who created only very short occasional ones connecting just about two places.
Also, the third hypothesis was confirmed, namely, that the average distance between places made by humans and the machine may be similar, but the average distance to the next move would be larger in the machine opponents. Indeed, the difference in average distance of the machine and of human adults was significant but with a very small effect size of 0.06–0.08. In contrast, the average distance of the next consecutive move following coloring-in a place showed a much larger effect size of 0.79. Humans colored-in about three places farther away while the next move of the machine was approximately five places away. This result showed that spatial exploration in humans was much more limited, if not inhibited.
That both children and adults showed less spatial exploration and created longer pathways clearly indicates that spatial heuristics were used. How these spatial heuristics were functioning was analyzed by looking at the contingencies between area, spatial pathways (place sequence length), and spatial exploration. The fourth hypothesis that the areas that the machine generates would be the result of places that incidentally aggregated into areas, while the areas that the children and adults generate would be intentional, could be confirmed as the places that clustered into areas significantly correlated with spatial exploration and pathways.
With respect to children, the pattern of correlations between cluster size, spatial exploration, and next move showed very clear differences from the machine. In children, their pathway length and next move were correlated with all the clusters whatever their size, while the machine showed no, or systematically lower correlations between clusters, pathway length, and spatial exploration. This indicates that the children monitored the clusters that they had created to plan their next move, while the machine clusters necessarily emerged randomly.
Surprisingly, this strong difference between human and machine players was not apparent in adults and their machine opponents. There were only significant correlations between clusters and the average distance in human adults but not in the machine. However, both the adults and the machine showed significant correlations between clusters and spatial exploration as per distance of the next consecutive move. Further investigation showed that here the only age difference between adults and children could be found. There was a small but significant difference between children, with 2.8 places away for the next move, and adults, showing a greater distance of 3.3 places away for the next move. Thus, adults showed somewhat more spatial exploration than children, which may be an indicator of more machine-like exploration in adults. This on average half-place difference may have been contributing to similar-sized correlations between area and next move in adults and their machine opponents. Further research needs to test the hypothesis of an inhibited spatial exploration of humans compared to the unrestrained spatial exploration of a machine.
To recap the machine strategy, the average distance between places was not very different in the humans and the machine, but the machine showed a highly significant advantage in reaching out across the array. The machine with its random coloring-in code expanded its coloring of places significantly further than humans in consecutive moves, both in the 10 × 10 grid and in the pilot study in an especially pronounced way in the larger 20 × 20 grid: The machine colored in places at double the distance compared to humans. Regarding adults, for psychologists believing in evolution (Silverman et al., 2007), rewards in spatial exploration (Meder et al., 2021) may be seen as more important rather than intellectual progress of spatial concepts from topological aggregate to Euclidean space. However, the maps for geographical areas needed for such exploration did improve over the centuries (Black, 2000). In developmental psychology, initially the development of the ability to use maps as indicators in reality was investigated (Downs and Liben, 1990; Liben and Downs, 2001), but more recent research investigated how the drawing of maps develops in children (Swienton et al., 2023). These authors showed that even 6–14-year-olds rarely used scales, legends, or arrows for the north direction, showing a lack of Euclidean information, but instead compensated with a mix of written labels or descriptive words on their maps. Thus, it is even more impressive that area building using aggregates of places did not differ in children and adults in this task.
It may be not only that computers beat humans in all kinds of board games (Purves, 2019) but also that machines work more similarly to adults' minds than to children's minds. Turing (1950) suggested building machines that would function like children's minds as they are constantly learning. He assumed that a computer successfully mimicking a child would use reasoning and decision-making at various developmental stages, instead of being conditioned with punishments and rewards. However, the background to Turing's suggestion may have been the much more limited computing power that machines had at his time.
There were limitations in the pilot research as we used a large grid for adults and a smaller grid for children. It is a common problem in developmental psychology how to equate task difficulty between age groups (Lange-Küttner, 2012): Increasing task difficulty for older age groups makes performance not directly comparable anymore, but using the same task may create ceiling effects in the older age groups. In the pilot study to the current research, it was originally assumed that there was no qualitative difference between grids of different sizes, and we followed board game practice to let children interact with a smaller board. The pilot showed that it was not possible to just divide the adults' scores by 4 (100 vs. 400 places) because (1) the adjusted score, when the 20 × 20 grid was used, underestimated adults' clusters by about half a place; (2) place sequences were about the double length in the 20 × 20 grid compared to the 10 × 10 grid and thus the grid size adjustment division factor of 4 would have actually shrunk the adult scores below the children's scores; (3) there was also a very large effect of the grid size on the average distance, which was double in the larger grid; and (4) a most striking result was that adults would make a next move at a comparable distance in either grid, while the machine opponents reached out farther and twice as far. Thus, these grid size effects were interesting, but the comparison also showed that no one-factor equation solution was possible; hence, the children and adults were compared here on the same-sized grid.
Neuropsychological insights and future directions
The discussion of this report should also include future research perspectives. Thus, this section considers some research studies that could give cues about the likely neuropsychological basis of the interactive game that revealed the construction of pathways and areas as participants navigated the two-dimensional grid. There are some developmental studies about the neuropsychological basis of spatial pathways in children and adults (e.g., Benear et al., 2022; Bohbot et al., 2012; Bostelmann et al., 2020; Botdorf et al., 2022; DeMaster et al., 2013) that need to be considered in some detail.
Many spatial tasks use objects or other cues as landmarks and then investigate whether these are used, ignored, or do not make a difference (e.g., Iaria et al., 2003; Lange-Küttner et al., 2023). Different from adults, young children often use landmarks for orientation (e.g., Bohbot et al., 2012), and it is assumed that spatial layout learning without spatial cues needs development until at least age 10 (Lange-Küttner, 2024; Leplow et al., 2003). A life-span study with twelve age groups from ages 6 to 89 showed that adolescents between 13 and 15 years were the fastest (but not the most accurate) in distance judgments of three objects toward oneself, the most accurate were young adults in their twenties, while the best allocentric estimations between objects in the field were made by adults in their 30s (Ruggiero et al., 2016). Thus, this study showed that the development of spatial judgments can be very protracted. Indeed, empty fields without boundaries like oceans are the hardest for spatial memory, but spatial boundaries that constitute even just a simple frame delimit the memory search space and improve spatial memory (Lange-Küttner, 2013).
The most common neuropsychological brain substrates found in navigation are the subcortical hippocampus and caudate nucleus, as well as the medial prefrontal cortex (e.g., Benear et al., 2022; Dahmani and Bohbot, 2015). Developmental differences were also found in regions in the posterior parietal cortex, the anterior prefrontal cortex, and the insula (DeMaster et al., 2013). A recent meta-analysis of 25 studies and 1,357 participants showed that in children and adolescents, correlations between hippocampal volume and memory tasks ranged from −0.36 to 0.48, but the hippocampal volume was not predictable by recognition vs. recall, delay, grouping (spatial categorization), age, sample size, or gender (Botdorf et al., 2022).
When researching navigation, there is also the problem of small-scale space vs. large environmental space. In real space, spatial navigation requires motion of the body in various directions, while small-scale spatial navigation does not require changes in perspective (Wang et al., 2014). However, from a neuropsychological perspective, the distinction between large- and small-scale space is not absolute as there are also overlapping brain substrates (Li et al., 2019).2 A recent meta-analysis of Li et al. (2019, Figure 3) showed subcortical hippocampal activity in large-scale environments but extensive neocortical activation in studies investigating small-scale spatial ability. Li et al. came to the conclusion that small-scale spatial abilities encompass motor imagery, working memory, the control of fingers and hands, arithmetic, and calculation, as well as matching. Large-scale spatial ability would involve virtual environments, scenes, and places, as well as autobiographical, episodic, and semantic memory.
Thus, which cognitive and hence brain processes may be relevant for the grid game? The grid game involves constructive spatial strategies without a memory component, which should be an advantage because children need the perceptual input that predicted hippocampal activation and, in turn, memory retrieval (DeMaster et al., 2013). The grid game also has the advantage that the places have visible boundaries and conform with young children's concept of aggregate space (Lange-Küttner, 2009) and thus it is suitable for children and adults alike (Lange-Küttner, 2009; Lange-Küttner and Beringer, 2023). The grid game requires attention to the to-be-colored-in places in the empty spaces in the entire array, the self-constructed pathways, and the colored-in places that dynamically appear due to the actions of the system player and may represent obstacles that constrain room for maneuver. Thus, it can be assumed that besides the hippocampus, neocortical networks would play an important role in the grid game: The development of attention to dynamic displays from 7 to 11 years of age and to adulthood showed a gradual increase of activation in a frontoparietal network, and more specifically, an age-related increase of the mid-cingulate gyrus activation, which is involved in cognitive control accompanied by a decrease in the involvement of the visual cortex (Wolf et al., 2018), which shows increasingly less reliance on perception.
Conclusion
In conclusion, this is the first study that developed and tested a digital interactive grid game for children and adults. In short, the current study compares children and adults with a randomly behaving machine when playing an interactive game of coloring in places in a grid. Interestingly, both children and adults used long place sequences and short consecutive moves to create large clusters, while the machine, with its fixed random place selection, had only short place sequences and longer consecutive moves at its disposal yet also created one large cluster and several small ones. This speaks to the logic of spatial constraints determined by filled vs. empty space (Lange-Küttner, 2024). Future variations of the task could show how children and adults would react if the computer had a sequencing strategy at its disposal, or investigate the length of a game if the players could overwrite already-colored-in places of the system player. One could hypothesize that such modifications would sharpen competition and that the length of the game would develop along the same lines as exploration for reward. This study contributes to theories about the development of spatial cognition in children showing an early matured ability to construct pathways and areas that is careful and circumspect. In addition, it showed that young children's practical spatial reasoning is a reasonable approach in its own right in addition and not inferior to geometric spatial concepts.
Author's note
I thank Sophie Clare Michailovic for organizing the data collection at Parkside Primary School in North London, United Kingdom, the head teacher for giving permission, and the parents and children for taking part. This study was carried out while the first author worked at London Metropolitan University, United Kingdom. The second author wrote the code for the Cognition Lab experiment and its evaluation. Henriette Mehl and Carmen Pütz-Bonnéhs at the University of Greifswald, Germany, helped with the literature search. The Leibniz Institute for Educational Trajectories, Bamberg, Germany, further supported the research. Parts of the study were presented in 2022 on the 22nd conference of the European Society for Cognitive Psychology in Lille, France, and in 2023 on the 52nd Annual Conference of the Jean Piaget Conference in Madrid, Spain.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: OSF Depository link (https://osf.io/tjgkn/).
Ethics statement
The studies involving humans were approved by London Metropolitan University, Psychology Department Ethical Board. The studies were conducted in accordance with the local legislation and institutional requirements. The approved consent letter was uploaded on the online experiment (further details see Procedure). Informed consent for children's participation in the study was provided by their legal guardians/next of kin. Adult participants provided informed consent by themselves.
Author contributions
CL-K: Conceptualization, Investigation, Writing – original draft, Resources, Validation, Project administration, Data curation, Writing – review & editing, Supervision, Visualization, Formal analysis, Methodology. JB: Visualization, Data curation, Writing – review & editing, Conceptualization, Investigation, Software, Resources.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Conflict of interest
JB was employed by BeriSoft Inc.
The remaining author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Correction note
This article has been corrected with minor changes. These changes do not impact the scientific content of the article.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fdpys.2025.1596481/full#supplementary-material
Footnotes
1. ^The term move is used here, even if no piece is moved like in chess, to denote that a player is taking a turn.
2. ^An interesting aspect of the common brain activation of both large- and small-scale spatial abilities (Li et al., 2019) was that these were located at the cerebral mantle of the brain, which appear to be the most compromised in general brain atrophy (Pini et al., 2016, Figure 4), especially in younger adults with Alzheimer's disease. Because some cases of Alzheimer's disease exist without hippocampal atrophy, Pini et al. (2016) have hypothesized that dementia can start selectively in various regions of the brain and then generalizes. Bohbot et al. (2007) could indeed show that covariation in brain atrophy could be found for the hippocampus with its surrounding brain areas, for instance, the amygdala and the perirhinal, the entorhinal, and even with the orbitofrontal cortices.
References
Agini, S. (2025). How a Random Chess Variant Won Over the World's Best Players. The FT Magazine. Available online at: https://www.ft.com/content/24561f88-bc6f-46c5-b44d-e1eba1e02363 (Accessed June 26, 2025).
Bar-Hillel, M., and Wagenaar, W. A. (1991). The perception of randomness. Adv. Appl. Math. 12, 428–454. doi: 10.1016/0196-8858(91)90029-I
Benear, S. L., Horwath, E. A., Cowan, E., Camacho, M. C., Ngo, C. T., Newcombe, N. S., et al. (2022). Children show adult-like hippocampal pattern similarity for familiar but not novel events. Brain Res. 1791:147991. doi: 10.1016/j.brainres.2022.147991
Black, J. (2000). Maps and History: Constructing Images of the Past. New Haven; London: Yale University Press.
Bohbot, V. D., Lerch, J., Thorndycraft, B., Iaria, G., and Zijdenbos, A. P. (2007). Gray matter differences correlate with spontaneous strategies in a human virtual navigation task. J. Neurosci. 27:10078. doi: 10.1523/JNEUROSCI.1763-07.2007
Bohbot, V. D., McKenzie, S., Konishi, K., Fouquet, C., Kurdi, V., Schachar, R., et al. (2012). Virtual navigation strategies from childhood to senescence: evidence for changes across the life-span. Front. Aging Neurosci. 4:28. doi: 10.3389/fnagi.2012.00028
Bostelmann, M., Lavenex, P., and Banta Lavenex, P. (2020). Children five-to-nine years old can use path integration to build a cognitive map without vision. Cogn. Psychol. 121:101307. doi: 10.1016/j.cogpsych.2020.101307
Botdorf, M., Canada, K. L., and Riggins, T. (2022). A meta-analysis of the relation between hippocampal volume and memory ability in typically developing children and adolescents. Hippocampus 32, 386–400. doi: 10.1002/hipo.23414
Bremner, J. G. (1984). Errors towards the perpendicular in children's copies of angular figures: a test of the bisection interpretation. Perception 13, 117–128. doi: 10.1068/p130117
Brugger, P., Monsch, A. U., Salmon, D. P., and Butters, N. (1996). Random number generation in dementia of the Alzheimer type: a test of frontal executive functions. Neuropsychologia 34, 97–103. doi: 10.1016/0028-3932(95)00066-6
Carsley, D., Heath, N. L., and Fajnerova, S. (2015). Effectiveness of a classroom mindfulness coloring activity for test anxiety in children. J. Appl. Schl. Psychol. 31, 239–255. doi: 10.1080/15377903.2015.1056925
Dahmani, L., and Bohbot, V. D. (2015). Dissociable contributions of the prefrontal cortex to hippocampus- and caudate nucleus-dependent virtual navigation strategies. Neurobiol. Learn. Memory 117, 42–50. doi: 10.1016/j.nlm.2014.07.002
DeMaster, D., Pathman, T., and Ghetti, S. (2013). Development of memory for spatial context: hippocampal and cortical contributions. Neuropsychologia 51, 2415–2426. doi: 10.1016/j.neuropsychologia.2013.05.026
Downs, R. M., and Liben, L. S. (1990). Getting a bearing on maps: the role of projective spatial concepts in map understanding by children. Childrens Environ. Quart. 7, 15–25.
Falk, R. (1981). “The perception of randomness,” in Fifth International Conference for the Psychology of Mathematical Education, Vol. 1 (Grenoble, France), 222–229.
Fitzpatrick, M. J., and McPherson, B. J. (2010). Coloring within the lines: gender stereotypes in contemporary coloring books. Sex Roles 62, 127–137. doi: 10.1007/s11199-009-9703-8
Flanagan, M., and Jakobsson, M. (2023). Playing Oppression: The Legacy of Conquest and Empire in Colonialist Board Games. Cambridge, MA: MIT Press. doi: 10.7551/mitpress/11779.001.0001
Flavell, J. H., Green, F. L., Herrera, C., and Flavell, E. R. (1991). Young children's knowledge about visual perception: lines of sight must be straight. Br. J. Dev. Psychol. 9, 73–87. doi: 10.1111/j.2044-835X.1991.tb00863.x
Freeman, N. H., and Adi-Japha, E. (2008). “Pictorial intention, action and interpretation,” in Drawing and the Non-Verbal Mind: A Life-Span Perspective, eds. C. Lange-Küttner and A. Vinter (Cambridge: Cambridge University Press), 104–120. doi: 10.1017/CBO9780511489730.006
Gobet, F., De Voogt, A., and Retschitzki, J. (2004). Moves in Mind. The Psychology of Board Games. Hove: Psychology Press. doi: 10.4324/9780203503638
Guhe, M., and Lascarides, A. (2014). “Game strategies for The Settlers of Catan,” in IEEE Conference on Computational Intelligence and Games (Dortmund: IEEE). doi: 10.1109/CIG.2014.6932884
Hentschel, M., Averbeck, B. B., and Lange-Küttner, C. (2022). The role of IQ and social skills in coping with uncertainty in 7- to 11-year-old children. Zeitschrift Entwicklungspsychol. Pädagogische Psychol. 54, 105–123. doi: 10.1026/0049-8637/a000256
Hentschel, M., Averbeck, B. B., and Lange-Küttner, C. (2023). The difference an explanation makes: verbal debrief effects when children cope with uncertainty. Cogn. Dev. 66:101317. doi: 10.1016/j.cogdev.2023.101317
Herman, J. F. (1980). Children's cognitive maps of large-scale spaces: effects of exploration, direction, and repeated experience. J. Exp. Child Psychol. 29, 126–143. doi: 10.1016/0022-0965(80)90096-X
Hund, A. M., and Plumert, J. M. (2005). The stability and flexibility of spatial categories. Cogn. Psychol. 50, 1–44. doi: 10.1016/j.cogpsych.2004.05.002
Iaria, G., Petrides, M., Dagher, A., Pike, B., and Bohbot, V. D. (2003). Cognitive strategies dependent on the hippocampus and caudate nucleus in human navigation: variability and change with practice. J. Neurosci. 23:5945. doi: 10.1523/JNEUROSCI.23-13-05945.2003
Izard, V., Pica, P., and Spelke, E. S. (2022). Visual foundations of Euclidean geometry. Cogn. Psychol. 136:101494. doi: 10.1016/j.cogpsych.2022.101494
Jones, K., and Tzekaki, M. (2016). “Research on the teaching and learning of geometry,” in The second Handbook of Research on the Psychology of Mathematics Education, eds. Á. Gutiérrez, G. C. Leder, and P. Boero (Leiden: Brill), 109–149. doi: 10.1007/978-94-6300-561-6_4
Kline, J. A., VanRyzin, K., Davis, J. C., Parra, J. A., Todd, M. L., Shaw, L. L., et al. (2020). Randomized trial of therapy dogs versus deliberative coloring (art therapy) to reduce stress in emergency medicine providers. Acad. Emerg. Med. 27, 266–275. doi: 10.1111/acem.13939
Kubovy, M., and Gilden, D. (1991). “Apparent randomness is not always the complement of apparent order,” in The Perception of Structure: Essays in Honor of Wendell R. Garner, eds. G. R. Lockhead and J. R. Pomerantz (Washington, DC: American Psychological Association), 115–127. doi: 10.1037/10101-006
Lange-Küttner, C. (2006). Drawing boundaries: from individual to common region. The development of spatial region attribution in children. Br. J. Dev. Psychol. 24, 419–427. doi: 10.1348/026151005X50753
Lange-Küttner, C. (2009). Habitual size and projective size: the logic of spatial systems in children's drawings. Dev. Psychol. 45, 913–927. doi: 10.1037/a0016133
Lange-Küttner, C. (2010a). Gender-specific developmental pathways for boys and girls: the Wertheimer Common-Region-Test can predict spatial memory. Eur. J. Dev. Sci. 4, 46–66. doi: 10.3233/DEV-2010-4104
Lange-Küttner, C. (2010b). Ready-made and self-made facilitation effects of arrays: Priming and conceptualization in children's visual memory. Swiss J. Psychol. 69, 189–200. doi: 10.1024/1421-0185/a000023
Lange-Küttner, C. (2012). The importance of reaction times for Developmental Science: what a difference milliseconds make. Int. J. Dev. Sci. 6, 51–55. doi: 10.3233/DEV-2012-11089
Lange-Küttner, C. (2013). Array effects, spatial concepts, or information processing speed: what is the crucial variable for place learning? Swiss J. Psychol. 72, 197–217. doi: 10.1024/1421-0185/a000113
Lange-Küttner, C. (2024). Visual and Motor Cognition in Infants and Children: What Develops and What Stays the Same. London: Routledge. doi: 10.4324/9780429270604
Lange-Küttner, C., Averbeck, B. B., Hentschel, M., and Baumbach, J. (2021). Intelligence matters for stochastic feedback processing during sequence learning in adolescents and young adults. Intelligence 86:101542. doi: 10.1016/j.intell.2021.101542
Lange-Küttner, C., Averbeck, B. B., Hirsch, S. V., Wießner, I., and Lamba, N. (2012). Sequence learning under uncertainty in children: self-reflection vs. self-assertion. Front. Psychol. 3:127. doi: 10.3389/fpsyg.2012.00127
Lange-Küttner, C., and Beringer, J. (2023). How to create an area when playing against a machine. Int. J. Hum. Comput. Interact. 39, 3361–3376. doi: 10.1080/10447318.2022.2096737
Lange-Küttner, C., Collins, C. L., Ahmed, R. K., and Fisher, L. E. (2023). Rich and sparse figurative information in children's memory for colorful places. Dev. Psychol. 59, 256–271. doi: 10.1037/dev0001483
Lange-Küttner, C., and Kochhar, R. (2020). Fine motor skills and unsystematic spatial binding in the Common Region Test: under-inclusivity in autism spectrum disorder and over-inclusivity in attention-deficit hyperactivity disorder. J. Motor Learn. Dev. 8, 544–568. doi: 10.1123/jmld.2019-0033
Lange-Küttner, C., and Küttner, E. (2015). How to learn places without spatial concepts: does the what-and-where reaction time system in children regulate learning during stimulus repetition? Brain Cogn. 97, 59–73. doi: 10.1016/j.bandc.2015.04.008
Leplow, B., Lehnung, M., Pohl, J., Herzog, A., Ferstl, R., and Mehdorn, M. (2003). Navigational place learning in children and young adults as assessed with a standardized locomotor search task. Br. J. Psychol. 94, 299–317. doi: 10.1348/000712603767876244
Li, Y., Kong, F., ji, M., Luo, Y., Lan, J., and You, X. (2019). Shared and distinct neural bases of large- and small-scale spatial ability: a coordinate-based activation likelihood estimation meta-analysis. Front. Neurosc. 12:1021. doi: 10.3389/fnins.2018.01021
Liben, L. S. (1982). Children's large-scale spatial cognition: Is the measure the message? New Dir. Child Adolesc. Dev. 1982, 51–64. doi: 10.1002/cd.23219821507
Liben, L. S., and Downs, R. M. (2001). “Geography for young children: maps as tools for learning environments,” in Psychological Perspectives on Early Childhood Education: Reframing Dilemmas in Research and Practice, ed. S. L. Golbeck (London: Erlbaum), 220–252.
Long, B., Simson, J., Buxó-Lugo, A., Watson, D. G., and Mehr, S. A. (2023). How games can make behavioural science better. Nature 613, 433–436. doi: 10.1038/d41586-023-00065-6
Luchins, A. S., and Luchins, H. (1947). A structural approach to the teaching of the concept of area in intuitive geometry. J. Educ. Res. 40, 528–533. doi: 10.1080/00220671.1947.10881546
Maulyda, M. A., Hidayati, V. R., and Erfan, M. (2020). Monopoly game media as an effort to improve calculation problem solving ability in elementary school students. J. Innov. Math. Learn. 3, 199–207. doi: 10.22460/jiml.v3i4.p199-207
Meder, B., Wu, C. M., Schulz, E., and Ruggeri, A. (2021). Development of directed and random exploration in children. Dev. Sci. 24:e13095. doi: 10.1111/desc.13095
Metzger, W. (1956). Die Entwicklung der Gestaltauffassung in der Zeit der Schulreife I, II [The development of Gestalt concepts in school beginners]. Westermanns Pädagogische Beiträge, 8, 531, 603–615.
Odic, D., Libertus, M. E., Feigenson, L., and Halberda, J. (2013). Developmental change in the acuity of approximate number and area representations. Dev. Psychol. 49, 1103–1112. doi: 10.1037/a0029472
Palmer, S. E. (1992). Common region: a new principle of perceptual grouping. Cogn. Psychol. 24, 436–447. doi: 10.1016/0010-0285(92)90014-S
Perner, J., Kohlmann, R., and Wimmer, H. (1984). Young children's recognition and use of the vertical and horizontal in drawings. Child Dev. 55, 1637–1645. doi: 10.2307/1130033
Piaget, J. (1977). “The role of action in the development of thinking,” in Knowledge and Development. Vol. 1, Advances in Research and Theory, eds. F. W. Overton and J. MacCarthy Gallagher (New York, NY: Plenum Press), 17–42. doi: 10.1007/978-1-4684-2547-5_2
Piaget, J., and Inhelder, B. (1948). La représentation de l'espace chez l'enfant. Paris: Presses Universitaires de France.
Piaget, J., Inhelder, B., and Szeminska, A. (1948). La geometrie spontanee de l'enfant. Paris: Presses Universitaires de France.
Pini, L., Pievani, M., Bocchetta, M., Altomare, D., Bosco, P., Cavedo, E., et al. (2016). Brain atrophy in Alzheimer's Disease and aging. Ageing Res. Rev. 30, 25–48. doi: 10.1016/j.arr.2016.01.002
Purves, D. (2019). What does AI's success playing complex board games tell brain scientists? Proc. Natl. Acad. Sci. U. S. A. 116, 14785–14787. doi: 10.1073/pnas.1909565116
Ruggiero, G., D'Errico, O., and Iachini, T. (2016). Development of egocentric and allocentric spatial representations from childhood to elderly age. Psychol. Res. 80, 259–272. doi: 10.1007/s00426-015-0658-9
Silver, D., Huang, A., Maddison, C. J., Guez, A., Sifre, L., van den Driessche, G., et al. (2016). Mastering the game of Go with deep neural networks and tree search. Nature 529, 484–489. doi: 10.1038/nature16961
Silverman, I., Choi, J., and Peters, M. (2007). The Hunter-Gatherer theory of sex differences in spatial abilities: data from 40 countries. Arch. Sex. Behav. 36, 261–268. doi: 10.1007/s10508-006-9168-6
Spelke, E. S. (2011). “Natural number and natural geometry,” in Space, Time and Number in the Brain, eds. S. Dehaene and E. M. Brannon (San Diego, CA: Academic Press), 287–317. doi: 10.1016/B978-0-12-385948-8.00018-9
Spelke, E. S., and Lee, S. A. (2012). Core systems of geometry in animal minds. Philos. Trans. R. Soc. B Biol. Sci. 367, 2784–2793. doi: 10.1098/rstb.2012.0210
Stiles, J., and Tada, W. L. (1996). Developmental change in children's analysis of spatial patterns. Dev. Psychol. 32, 951–970. doi: 10.1037/0012-1649.32.5.951
Swienton, H. A., Giordano, A., and Hagelman III, R. R. (2023). What do children draw when asked to draw a map? Results of a mental map experiment. Cartographica 58, 116–143. doi: 10.3138/cart-2022-0016
Tokuhisa, S., and Kamiyama, Y. (2010). “The world is canvas: a coloring application for children based on physical interaction,” in IDC 10 Proceedings of the 9th International Conference on Interaction Design and Children (ACM Digital Library), 315–318. doi: 10.1145/1810543.1810601
Towse, J. N., and McLachlan, A. (1999). An exploration of random generation among children. Br. J. Dev. Psychol. 17, 363–380. doi: 10.1348/026151099165348
Towse, J. N., and Neil, D. (1998). Analyzing human random generation behavior: a review of methods used and a computer program for describing performance. Behav. Res. Methods Instrum. Comput. 30, 583–591. doi: 10.3758/BF03209475
Treisman, M., and Faulkner, A. (1987). Generation of random sequences by human subjects: cognitive operations or psychological process? J. Exp. Psychol. Gen. 116, 337–355. doi: 10.1037//0096-3445.116.4.337
Veracini, L. (2013). Settlers of Catan. Settler Colon. Stud. 3, 131–133. doi: 10.1080/18380743.2013.761941
Vinter, A., Picard, D., and Fernandes, V. (2008). “Graphic syntax and representational development,” in Drawing and the Non-Verbal Mind: A Life-Span Perspective, eds. C. Lange-Küttner and A. Vinter (Cambridge: Cambridge University Press), 139–158. doi: 10.1017/CBO9780511489730.008
Wang, L., Cohen, A. S., and Carr, M. (2014). Spatial ability at two scales of representation: a meta-analysis. Learn. Individ. Diff. 36, 140–144. doi: 10.1016/j.lindif.2014.10.006
Wolf, K., Galeano Weber, E., van den Bosch, J. J. F., Volz, S., Nöth, U., Deichmann, R., et al. (2018). Neurocognitive development of the resolution of selective visuo-spatial attention: functional MRI evidence from object tracking. Front. Psychol. 9:1106. doi: 10.3389/fpsyg.2018.01106
Appendix
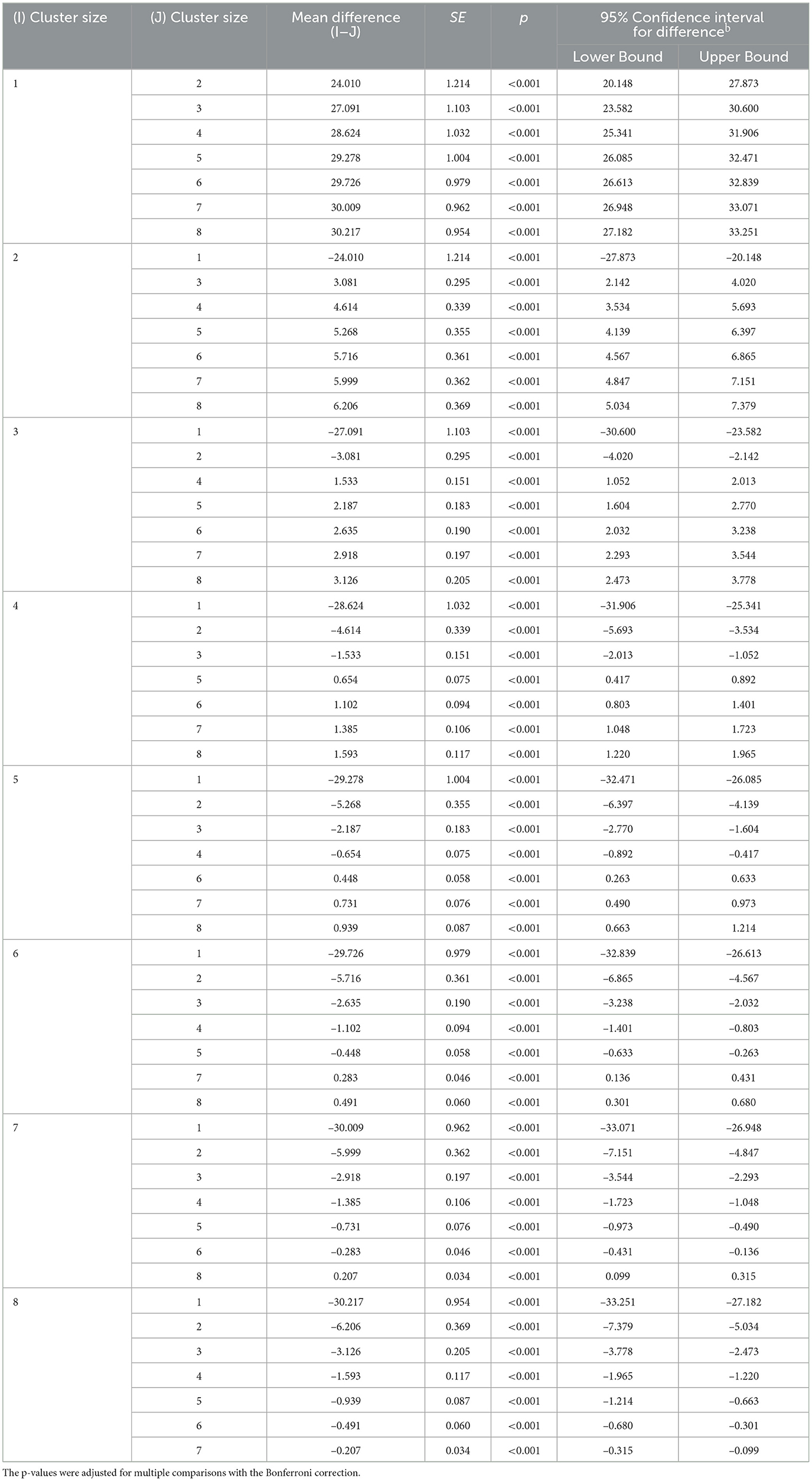
Table A1. Pairwise comparisons of the cluster.
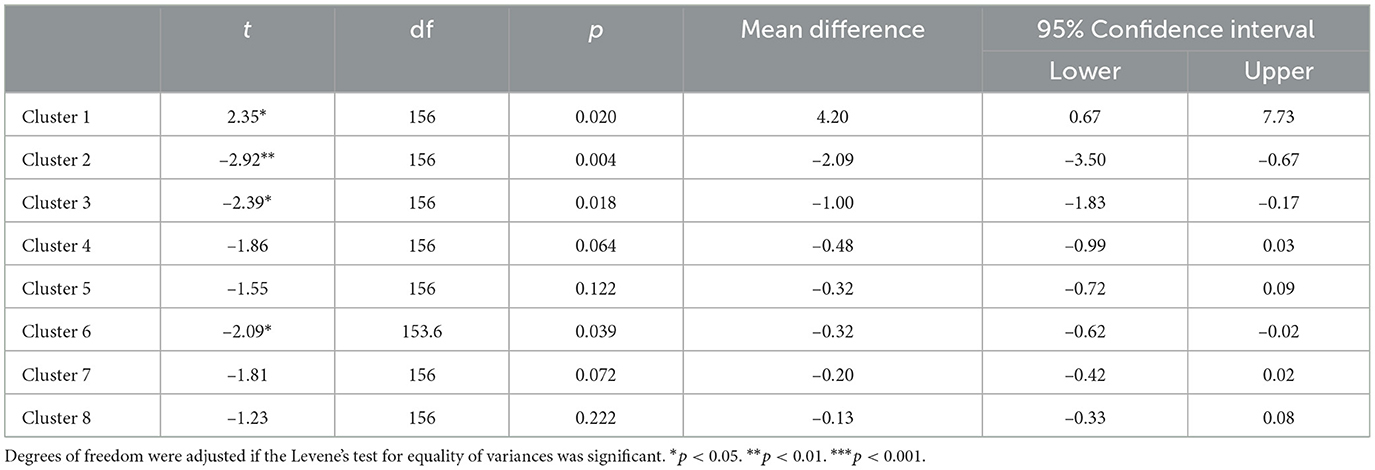
Table A2. Post-hoc t-tests (two-tailed) for independent samples (human/machine).
Keywords: spatial abilities, spatial concepts, Euclidean and aggregate space, places and pathways and areas, spatial exploration, human-machine interaction
Citation: Lange-Küttner C and Beringer J (2025) Spatial heuristics and random spatial exploration: children, adults, and the machine coloring-in places in the grid game. Front. Dev. Psychol. 3:1596481. doi: 10.3389/fdpys.2025.1596481
Received: 19 March 2025; Accepted: 30 June 2025;
Published: 07 August 2025; Corrected: 12 August 2025.
Edited by:
Marco Giancola, University of L'Aquila, ItalyReviewed by:
Victoria D. Chamizo, University of Barcelona, SpainAleksandar Trifunović, University of Belgrade, Serbia
Copyright © 2025 Lange-Küttner and Beringer. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Christiane Lange-Küttner, Yy5sYW5nZWt1ZXR0bmVyQHVuaS1icmVtZW4uZGU=
†ORCID: Christiane Lange-Küttner orcid.org/0000-0001-6021-8931
Jörg Beringer orcid.org/0000-0002-3270-6994