 Alexander G. Watts1*
Alexander G. Watts1* Peter E. Schlichting2
Peter E. Schlichting2 Shawn M. Billerman3,4
Shawn M. Billerman3,4 Brett R. Jesmer3,4
Brett R. Jesmer3,4 Steven Micheletti5Marie-Josée Fortin1W. Chris Funk6
Steven Micheletti5Marie-Josée Fortin1W. Chris Funk6 Paul Hapeman7
Paul Hapeman7 Erin Muths8
Erin Muths8 Melanie A. Murphy4,9*
Melanie A. Murphy4,9*- 1Department of Ecology & Evolutionary Biology, University of Toronto, Toronto, ON, Canada
- 2Department of Natural Resources Management, Texas Tech University, Lubbock, TX, USA
- 3Department of Zoology and Physiology, University of Wyoming, Laramie, WY, USA
- 4Program in Ecology, University of Wyoming, Laramie, WY, USA
- 5School of Biological Sciences, Washington State University, Pullman, WA, USA
- 6Graduate Degree Program in Ecology, Department of Biology, Colorado State University, Fort Collins, CO, USA
- 7Department of Biology, Central Connecticut State University, New Britain, CT, USA
- 8Fort Collins Science Center, U.S. Geological Survey, Fort Collins, CO, USA
- 9Department of Ecosystem Science and Management, University of Wyoming, Laramie, WY, USA
Heterogeneous landscapes and fluctuating environmental conditions can affect species dispersal, population genetics, and genetic structure, yet understanding how biotic and abiotic factors affect population dynamics in a fluctuating environment is critical for species management. We evaluated how spatio-temporal habitat connectivity influences dispersal and genetic structure in a population of boreal chorus frogs (Pseudacris maculata) using a landscape genetics approach. We developed gravity models to assess the contribution of various factors to the observed genetic distance as a measure of functional connectivity. We selected (a) wetland (within-site) and (b) landscape matrix (between-site) characteristics; and (c) wetland connectivity metrics using a unique methodology. Specifically, we developed three networks that quantify wetland connectivity based on: (i) P. maculata dispersal ability, (ii) temporal variation in wetland quality, and (iii) contribution of wetland stepping-stones to frog dispersal. We examined 18 wetlands in Colorado, and quantified 12 microsatellite loci from 322 individual frogs. We found that genetic connectivity was related to topographic complexity, within- and between-wetland differences in moisture, and wetland functional connectivity as contributed by stepping-stone wetlands. Our results highlight the role that dynamic environmental factors have on dispersal-limited species and illustrate how complex asynchronous interactions contribute to the structure of spatially-explicit metapopulations.
Introduction
A fundamental goal of ecology is to understand how environmental variation influences species persistence, abundance, and gene flow (Cushman, 2006; Gomez-Rodriguez et al., 2009; Goldberg and Waits, 2010). Landscape heterogeneity is defined by fluctuations in environmental conditions that range from relatively invariable (e.g., topography, soil texture) to highly variable (e.g., rainfall, vegetation abundance) over multiple spatial and temporal scales. For many species, these abiotic conditions are necessary for species survival, recruitment, (Fahrig, 2003; Ewers and Didham, 2007), and dispersal (Girdner and Larson, 1995; Driscoll, 1997; Schwartz and Jenkins, 2000; Banks et al., 2004; Mokany, 2007). Yet it remains unclear to what extent variation in suitable conditions over space and time affects gene flow, population genetic structure, and genetic diversity of natural populations.
Functional connectivity, the degree to which the environment impedes or facilitates the movement of individuals among resource patches (Taylor et al., 1993; Bélise, 2005), is linked to genetic connectivity between populations existing in spatially-explicit habitat patches (Brown and Kodric-Brown, 1977; Tallmon et al., 2004). Geographic distance is expected to play a significant role in the explanation of genetic distance between a pair of occupied sites (McRae, 2006), assuming dispersal is limited over large distances. Yet the functional connectivity of a species may be dependent on environmental characteristics within- and between-habitat patches, whereby landscape condition may create resistance to gene flow in addition to animal movement.
Within-habitat characteristics (e.g., vegetation, resource abundance, presence of conspecifics) affect dispersal by influencing the production and survival of migrants (Banks et al., 2004), while between-habitat patch factors (e.g., inter-patch matrix: complex topography, vegetative cover, risk of predation) affect the probability of colonization and establishment (Stow and Sunnucks, 2004) in destination habitat patches. Temporal fluctuations in these environmental characteristics may mediate the complex ecological interactions that influence demographic and genetic processes within and between natural populations (Gomez-Rodriguez et al., 2009; Velo-Antón et al., 2013), especially for dispersal-limited species. It is therefore expected that fluctuating, heterogeneous landscapes will affect species functional connectivity corresponding to either beneficial or detrimental effects on demographic and dispersal thresholds essential for species persistence and genetic diversity (Schwartz and Jenkins, 2000; Scherer et al., 2012). Quantified values of functional connectivity can help characterize complex spatio-temporal interactions between landscape composition and configuration, population genetic structure, and genetic connectivity of a population.
Amphibians are exemplary model species to assess genetic connectivity in spatially and temporally variable landscapes because they are dispersal-limited, patch-dependent species (Gamble et al., 2007) sensitive to changes in vegetation and fluctuating hydrologic conditions. Juveniles leave ephemeral wetlands after metamorphosis, usually as wetlands are drying (Semlitsch, 2008) and are then subject to the spatio-temporal dynamics of the within-patch matrix. Successful recolonization of wetland habitat patches is more likely between neighboring patches than distant, isolated patches (Driscoll, 1997; Smith and Green, 2005; Rozenfeld et al., 2008) especially if the between-patch matrix is resistant to movement. Within- and between-patch environmental fluctuations may significantly influence amphibian occupancy of surrounding wetland patches (Scherer et al., 2012) altering amphibian population dynamics and genetic structure. However, these interactions between variable abiotic conditions and amphibian population genetic structure are understood poorly despite significant implications for population persistence, species diversity, and metapopulation dynamics as the environment changes and the climate warms. Thus, our goal was to test the effects of fluctuating environmental factors on functional connectivity of an amphibian population using a landscape genetics approach.
We evaluated the effect of spatio-temporal variation in wetland availability on the genetic connectivity of a population of boreal chorus frogs (Pseudacris maculata) in the high mountain wetlands of the Northern Rocky Mountains in Larimer County Colorado. Across the species range, boreal chorus frogs breed in primarily ephemeral wetlands with emergent vegetation in spring and summer. Individuals then disperse to wet meadows to forage during the summer and early fall (Weyrauch and Grubb, 2004). Spencer (1964) described the species' relatively low dispersal ability (~600 m average dispersal maxima) and variation in site occupancy within years across our study area. Given pond-breeding behavior and low dispersal capacity, boreal chorus frogs are well-suited for measuring environmental limits to genetic connectivity. Moreover, snowpack has the potential to have a direct effect on seasonal wetland availability for breeding montane amphibian populations (Corn and Muths, 2002; Pilliod et al., 2002). Dependent on winter snowpack levels, spring and summer snowmelt is expected to affect the hydrologic and vegetative conditions necessary for amphibian productivity and dispersal success among wetlands. Is it also expected that precipitation-driven snowmelt variation may alter the spatial configuration of the wetland habitat (Corn, 2005). We therefore consider spatio-temporal snowpack variation as an important potential driver of the hydrological conditions that could influence amphibian genetic connectivity in montane regions.
We predicted genetic connectivity of P. maculata among sampled breeding wetlands by within- and between-wetland predictors, as well as wetland connectivity predictors, using gravity models. Gravity models are network models parameterized to include landscape-based attraction and resistance factors to predict genetic distance. Our gravity models also incorporated functional connectivity predictors of species movement in spatially-explicit habitat networks to further predict how landscape spatial heterogeneity affects genetic connectivity. Additionally, we investigated the functional connectivity predictors independently from the within- and between-wetland predictors to evaluate the variation in spatio-temporal wetland structure on the potential connectivity of the wetland network. We hypothesized that spatial heterogeneity in habitat structure and inter-annual variation in snowpack directly control wetland availability in a given year and predict that isolation by distance does not explain amphibian genetic connectivity as well as spatio-temporal precipitation and functional connectivity predictors.
Methods
Field Collection
Permits for field data and tissue collection were provided by the United States Forest Service and the Colorado Division of Wildlife. All animal procedures in this study followed recommendations of the Colorado State University Institutional Animal Care and Use Committee; this study was approved and permitted by the Colorado Division of Wildlife (# 09HP957) and Colorado State University Institutional Animal Care and Use Committee (# 09107A02).
We used a stratified random sample design (by elevation) to select sites, augmented by sites used in the Spencer study (Spencer, 1964). Of the 35 surveyed sites in the study area, 22 were occupied in 2009–2010. Eighteen sites yielded a sufficient number of samples to be used in the analysis (n = 322 individuals, Table 1). Of the 18 sites with sufficient sample sizes for genetic analysis, 14 were selected via the random stratification (9/14 of these were occupied in Spencer, 1964) while 4 were “augmentation” sites (occupied during the Spencer study but not part of the random stratification; Spencer, 1964; Corn and Muths, 2002). Wetlands varied in area (200–20,000 m2) and were visited 1–3 times with the goal of obtaining 30 samples (buccal swabs from adults, Goldberg et al., 2003) or tail clips from larvae (Murphy et al., 2010b; Figure 1). Wetland depths were categorized as < 1 m, 1–2 m, and >2 m. Vegetation cover was estimated by perceived percentage vegetative cover during sampling. pH and conductivity were sampled at each wetland.
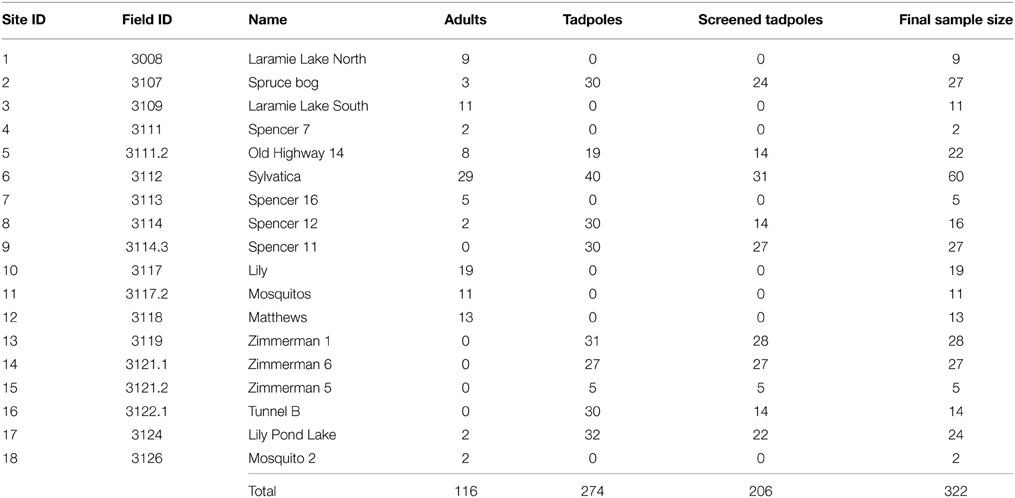
Table 1. Boreal chorus frog (Pseudacris maculata) sample sizes per studied wetland.
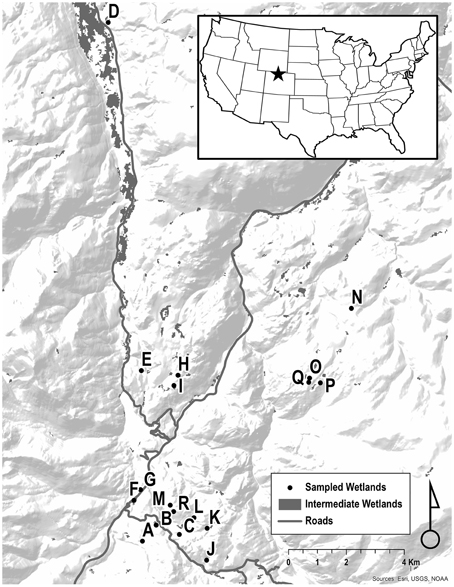
Figure 1. Study area: Headwaters of the Cache La Poudre River and Laramie River, Colorado, USA. Sampled breeding wetlands are shown as letters.
Genetic Data
DNA was extracted from tissue samples using a Qiagen DNeasy96 tissue kit with minor modifications to manufacturer's protocol (Murphy et al., 2010b). We generated multi-locus genotypes (n = 322, loci = 12, Lemmon et al., 2011, Appendix S1) using the Qiagen multiplex kit, an Applied Biosystems 3730 automated sequencer, and scored fragments with Gene Marker 1.91 (SoftGenetics). We implemented a number of measures for quality control: at least 2 negative controls per DNA extraction, 2 negative controls (no DNA) per each PCR amplification, amplified a known genotype in each PCR reaction, re-amplified all rare alleles (< 5% frequency), and re-amplified in at least 10% of samples to assess accuracy of genotyping. When using larvae, varying sample size of full siblings may bias estimates of allele frequencies (Goldberg and Waits, 2009). Therefore, we estimated clusters of full siblings for each sample location (Wang, 2004) and subsampled each sibling cluster (n) where n is the number of individuals in the smallest sibling cluster for that location (Goldberg and Waits, 2009; Murphy et al., 2010b). We tested for Hardy–Weinberg proportions and gametic phase disequilibrium and estimated genetic distance using Dps (Bowcock et al., 1994) with Microsatellite Analyser (MSA) (Dieringer and Schlötterer, 2003) measured for all pairwise comparisons.
Gravity Models
Gravity models (Fotheringham and O'Kelly, 1989) are network-based models that incorporate landscape data that potentially influence genetic connectivity, and factors potentially influencing amphibian population dynamics. In this framework, functional connectivity (Tij, 1-genetic distance, dependent variable) is modeled as a response of three type of independent variables: spatial distribution (distance between sites, w), at-site (network nodes, v) characteristics representing production of flow and between-site (network edges, c) characteristics describing resistance to flow (Equation 1; Murphy et al., 2010a, 2015). Parameter estimates for independent variables are μ, α, and β respectively, where α and β may represent a vector of variables (Anderson, 1979; Fotheringham and O'Kelly, 1989).
Geographic distance was included in all gravity models as gravity models (“spatial interaction models”) assume spatial autocorrelation (Anderson, 1979). To fit the gravity models as singly constrained, we linearized the equation by taking the natural log and fit using mixed effects models (Murphy et al., 2010a) where each site has an independent estimated intercept (k) but global estimate of all independent variables (Murphy et al., 2010a). Singly constrained gravity models balance information content and effective sample size (Fotheringham and O'Kelly, 1989); they also account for non-independence of pair-wise genetic distances (Murphy et al., 2015).
To include spatially-explicit measures of functional connectivity, we included three sets of predictors: (a) within-habitat (v) and (b) between-wetland environmental predictors (c), and by (c) spatio-temporal wetland connectivity predictors (c). Our spatio-temporal wetland connectivity predictors are quantified using three wetland networks: (1) a spatial network, to evaluate connectivity of 18 occupied wetlands; (2) a temporal network, to evaluate effects of annual fluctuations in hydroperiod; and (3) a stepping-stone network, to evaluate the effect of all 128 potential wetlands in the study area on genetic connectivity. All candidate models within 2 ΔAIC (Akaike's information criterion, AIC; Akaike, 1974; Burnham and Anderson, 2002) were considered the top models of connectivity. A null model of isolation by distance (distance alone) was used as a baseline for comparison. Model validation techniques for gravity models are limited. However, our goal was not to infer but to evaluate the relative contribution of within- and between-wetland versus wetlands connectivity predictors over space and time in predicting genetic distance. Accordingly, we included ΔAIC values per single predictor in the top six models as a proxy of predictor contribution to top-ranking gravity models.
Within- and Between-site Predictors
Wetland characteristics are expected to control the number of potential migrants. Therefore, we included within-wetland (node) variables potentially important for recruitment: peripheral habitat (ratio of meadow to forest), run-off (impervious surfaces), water accumulation potential (compound topographic index), precipitation timing (precipitation ratio), site accessibility (relative slope position), and conductivity (see Table 2 for description and justification of variables). All within-wetland characteristics were measured within a 100 m buffer surrounding wetland edge. Between-wetland (edge between nodes) variables are those that are hypothesized to promote or resist dispersal: habitat (ratio of meadow to forest), roads (impervious surfaces), water accumulation potential (compound topographic index), precipitation timing (precipitation ratio), and topographic complexity (surface relief ratio; see Table 2). We tested for correlations among these variables and did not include any variables with an R2 > 0.7. We found no significant collinearity between remaining predictors using a VIF threshold of 5. Between-wetland variables were evaluated for the 18 primary wetlands using a saturated network (i.e., each wetland is connected to all other wetlands) where the sensitivity of land cover types between wetlands was analyzed at multiple spatial scales, measured as bandwidths along each network edge [30 (minimum resolution), 60, 120, and 240 m buffers, Murphy et al., 2010a,b]. Selection of the best bandwidth to use was evaluated using AIC.
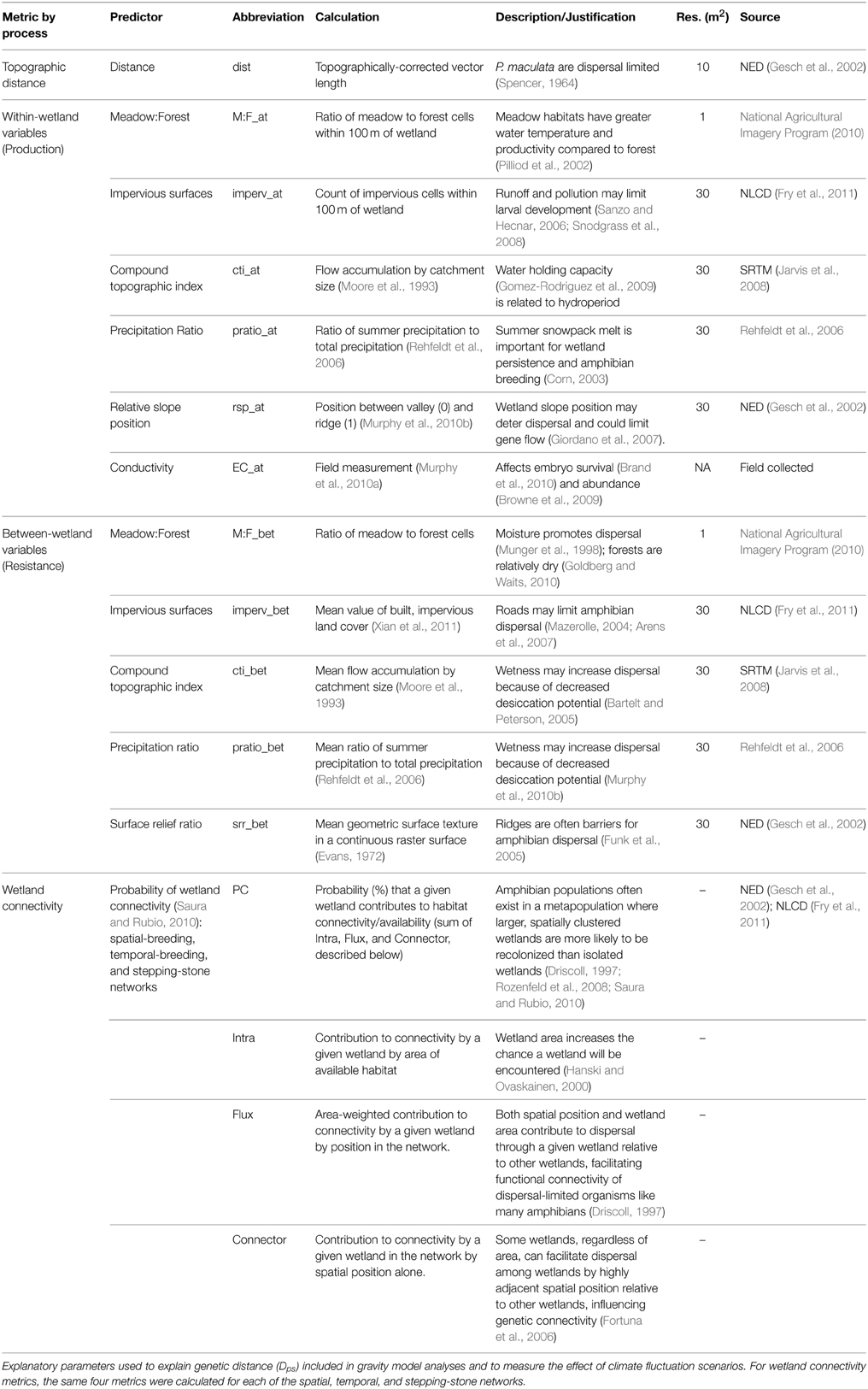
Table 2. Within-wetland, between-wetland, and wetland connectivity predictors.
Wetland Connectivity Predictors
We incorporated wetland connectivity predictors in the gravity models that represented wetland composition and configuration data (i.e., wetland area, spatial position in landscape). These functional connectivity metrics quantify the functional capacity of the landscape to inhibit or facilitate movement and, consequentially, gene flow (Taylor et al., 1993). Functional connectivity predictors were calculated per wetland based on three types of network: (a) a spatial breeding network (“spatial-breeding”) measuring static functional connectivity of 18 breeding wetlands; (b) a temporal network (“temporal-breeding”), measuring fluctuating connectivity per breeding wetland over time, dependent on snowpack variation; and (c) a spatial wetland network (“stepping-stone”) measuring per wetland connectivity considering the sum of 110 additional suitable stepping-stone habitat between 18 breeding wetlands (LinkageMapper v 0.9, McRae and Kavanagh, 2011) in ArcGIS 10.0 (ESRI, 2011).
Spatial-breeding network
We calculated functional connectivity of 18 breeding wetlands for one static sampling period using the spatial-breeding network to incorporate the effect of spatially-explicit habitat structure on genetic connectivity in the gravity models. We developed the network using frog-occupied wetlands as graph nodes and Euclidean distance between wetlands as graph edges. For each wetland, we evaluated the probability of connectivity (Saura and Rubio, 2010, Appendix S2), quantified as four per-wetland connectivity metrics: PC (overall), composed of the sum of the three sub-metrics: Intra (probability of wetland connectivity calculated by habitat area alone), Flux (probability wetland connectivity calculated by spatial position, weighted by area), and Connector (probability of wetland connectivity calculated by spatial position alone, Table 2). These metrics were constrained by a dispersal kernel, calculated using the maximum observed dispersal distance of P. maculata (~600 m, Spencer, 1964), but assuming that some individuals have the capacity to surpass this maximum distance (5% of individuals).
Temporal-breeding network
To evaluate the effect of interannual variation in wetland hydroperiod on P. maculata genetic connectivity, we calculated functional connectivity metrics for 18 breeding wetlands over time using the temporal-breeding network. For the temporal-breeding network, using the same frog-occupied wetlands as the spatial-breeding network, we modified the “availability” property of each wetland according to the amount of snowpack (SNOTEL 1979-2010 Station CO05J37S, National Water and Climate Center). We classified observed wetland permanence (Amburgey et al., 2014), a qualitative proxy of ability for wetlands to maintain a suitable hydroperiod for amphibian productivity, under varying snowpack depths. We used the average snotel snowpack depth over recorded years (average depth = 63.5 cm) as a median threshold for wetland permanence (Low permanence < 63.5 cm snow; High permanence > 63.5 cm snow; Neutral = no differences related to snowpack). We designed this novel, qualitative method to classify what conditions were most productive for chorus frogs based on snowpack for a given site: (1) wetlands that are productive only when snowpack is low and breeding areas are available due to decreased water depth (Low); (2) wetlands that are productive only when snowpack is high and breeding areas are available due to increased water depth (High); and (2) wetlands where production is not influenced by the amount of water from snowpack (Neutral). We then attributed the sum of years of available hydroperiod per snowpack category occurring from 1979–2010: “low” wetlands productive 15 of 33 years, “high” wetlands productive 18 of 33 years, and “neutral” wetlands productive in all 33 years. Temporal functional connectivity predictors were quantified using probability of connectivity metrics, as described for the spatial network.
Stepping-stone network
To evaluate the effect of neighboring wetlands on P. maculata genetic connectivity we included all potential breeding sites (110 additional wetlands, National Land Cover Dataset, 30 m resolution, Fry et al., 2011) to represent nodes in the spatial-breeding network. Only wetlands greater in area than the smallest neutral breeding wetland (>400 m2). The resulting stepping-stone network is the sum of the 18 sampled breeding wetlands and 110 wetlands located within the study region and were considered potentially suitable for intermediate habitat, for 128 nodes. We considered all selected wetlands to be neutral to snowpack variation for the stepping-stone network. We calculated functional connectivity predictors as described in the spatial-breeding network only for our 18 sampled, occupied wetlands to quantify the effect of neighboring wetland habitat on functional connectivity on our focal sites.
Results
Genetic Data
All 11 microsatellite loci were polymorphic, with between 7 and 26 alleles per locus (× = 14.16) and heterozygosity from 0.212 to 0.788 (× = 0.483) by locus (Table S1). Likely due to substructure, loci were not in global Hardy–Weinberg equilibrium (HWE) or linkage equilibrium. However, when considered on a pond-by-pond basis, no single locus was consistently out of HWE or LD, indicating that deviations are unlikely to be due to null alleles. Global GST over all loci was 0.215 and was highly significant. Pair-wise genetic differentiation metrics consistently revealed generally significant levels of genetic differentiation with 65% of pairwise GST comparisons significant after Bonferroni correction (p-value 0.05), 84% of non-significant values were sites with less than 700 meters separation. Pairwise GST values ranged from 0 to 0.370. Dps also indicated genetic structure in our study area, ranging from 0.312 to 0.943 (× = 0.553).
Gravity Models
The 30 m bandwidth of between-wetland factors had the lowest competing AIC scores and thus was used for all gravity analyses. The model set with the most support (8 models ΔAIC values < 2) included distance between sites (distance) with within-site moisture (pratio, cti) and between-site resistance (topography as measure by srr, pratio; Table 3, Appendix S3). Models including surface relief ratio within-wetland or precipitation ratio within-wetland as independent predictors resulted in ΔAIC values < 2 but were not the top-ranking models. Functional connectivity predictors from the stepping-stone network (i.e., stepping-stone wetland connectivity) were also important predictors in explaining the genetic connectivity among P. maculata-occupied wetlands (PC_steppingstone_at, ΔAIC < 2; Table 3) though did not explain genetic connectivity as well independently (Table 3). Some measures of wetness within-wetland (cti or pratio) were included in six of the eight top models. Surface relief ratio was relevant in all of the competing models while moisture (pratio) was the only other metric describing differences between-wetlands that was present in the top models (three of the eight models). Distance alone (null hypothesis) was not in the top set of models (Table 3).
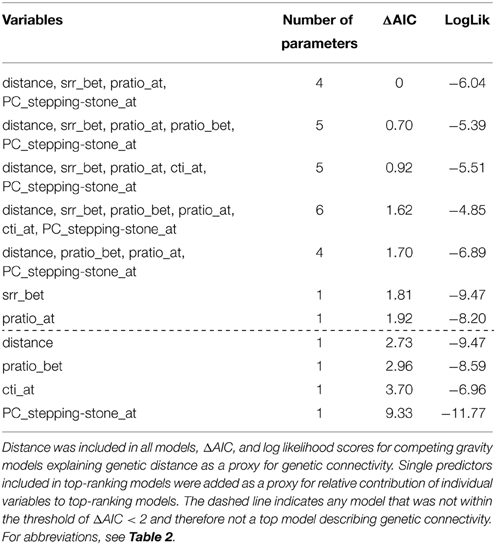
Table 3. Gravity models that best explain genetic distance.
Wetland Connectivity
Considering only wetland connectivity metrics in predicting genetic distance, we found that the presence of stepping-stone wetlands (stepping-stone network) was the highest contributor to the top models explaining genetic connectivity (Table 3). Percent-values of wetland connectivity measured using the spatial-breeding, temporal-breeding, and stepping-stone networks were variable perwetland (0.29–45.25%; 0.00–68.47%; 0.00–0.41%, respectively) (Table 4). For the spatial-breeding network, wetland area and wetland spatial position in the network (connectivity characteristics represented by the Flux connectivity metric) contributed most to wetland connectivity (Table 4, Figure 2A) whereby site “I” represented the most functionally connected wetland spatially (PC = 45.24%; Figure 2A) due to wetland availability alone (represented by the Intra metric). Conversely, for the temporal-breeding network, only wetland availability (measured by the Intra connectivity metric) contributed to overall wetland connectivity (Figure 2B) whereby site “C” represented the most functionally-connected wetland over time (PC = 68.47%). In the stepping-stone network, functional connectivity quantities (PC-values) for all 18 sampled sites were low (relatively low Flux, Intra and connection values) but variable. The Connector metric did not contribute to overall wetland connectivity (PC < 0.01%) for any wetlands in any of the three types of networks.
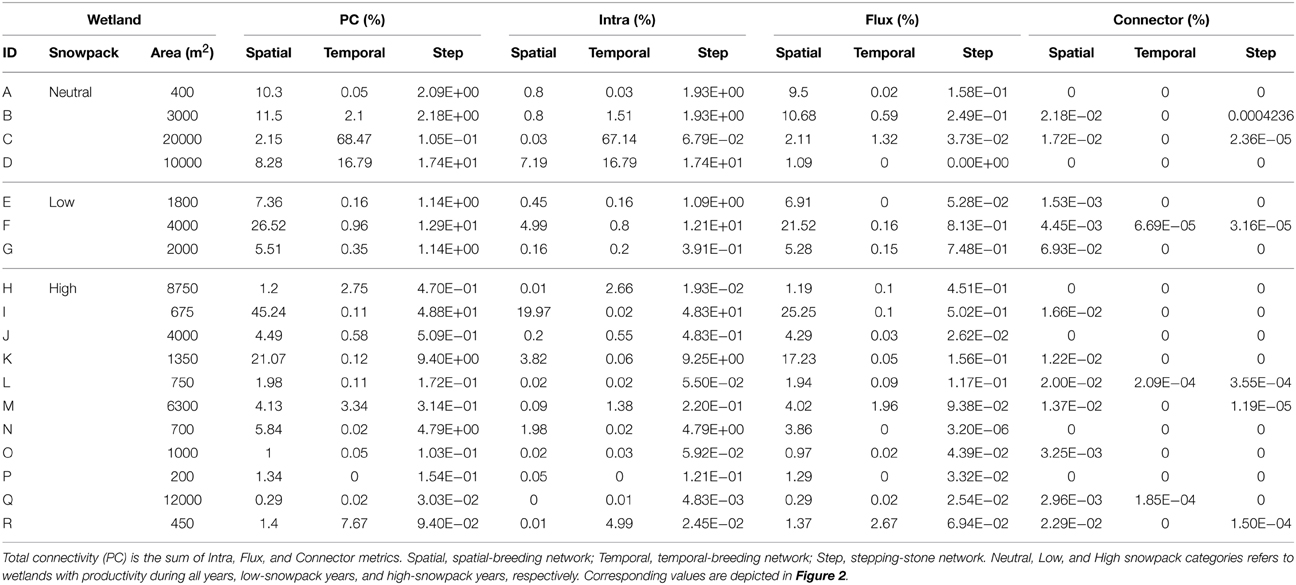
Table 4. Functional connectivity, quantified as probability of connectivity, per wetland.
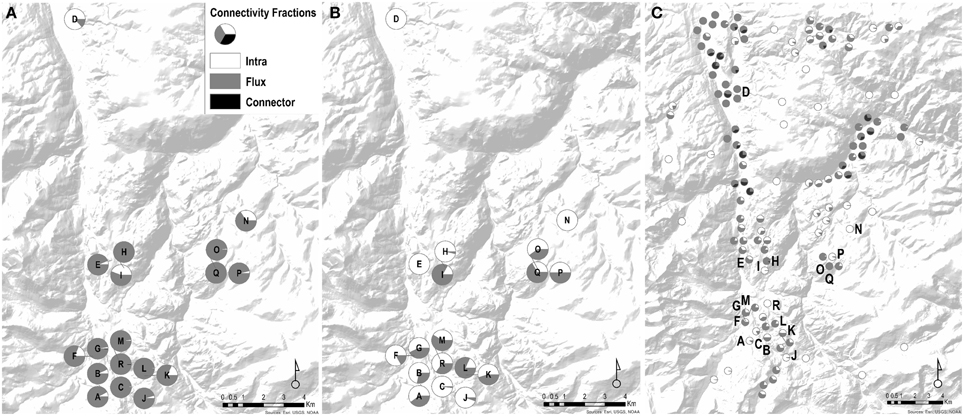
Figure 2. Networks designed for wetland connectivity of Pseudacris maculata. Pie charts represent the proportion of influence by Intra, Flux, and Connector to overall wetland connectivity. (A) Spatial-breeding network: all sampled sites were considered nodes. In this scenario, wetland connectivity is largely driven by Flux (availability + spatial position). (B) Temporal-breeding network: all sampled sites were considered nodes, but categorized as temporal_low, temporal_high, and temporal_neutral wetlands. Compared to the Spatial_breeding network, the temporal network is now driven by both Intra (availability) and Flux (availability + spatial position). (C) Stepping-stone network: 110 unoccupied sites within the region were added to the 18 sampled sites. In the stepping_stone network, the Connector fraction now becomes a driver of connectivity for sampled wetlands.
Discussion
Understanding how spatio-temporal variation in habitat composition and configuration influences species dispersal, colonization success, and gene flow is critical to predicting species demographic dynamics in changing landscapes. Variability in the state of environmental conditions on which many species depend may have significant consequences on individual development, population dynamics, and genetic diversity (Funk et al., 2005; Fortuna et al., 2006; Gamble et al., 2007). Considering future climatic and land cover changes, it is essential we understand how fluctuations in environmental factors affect species genetic connectivity toward prediction of demographic and genetic shifts. We compared these environmental factors to elements of functional connectivity that influenced dispersal and gene flow in a population of P. maculata over space and time, including climatic fluctuations to address the potential impact of climate warming on the genetic structure of populations. We found that complex interactions among covariates (i.e., within- and between-site moisture, between-wetland topographic complexity, underlying wetland connectivity and fluctuations in annual precipitation), have distinct and potentially critical roles in controlling genetic connectivity in boreal chorus frogs.
Within-wetland Factors
Within-wetland characteristics were important in determining genetic connectivity. Our results indicate stronger genetic connectivity among wetlands with higher levels of water recharge (measured by surface relief ratio and precipitation ratio), a greater capacity to hold water (measured as compound topographic index), and presence in all years (measured as neutral snowpack). Indeed, based on ΔAIC model results, surface relief and precipitation were moisture predictors that explained genetic connectivity independently better than other individual predictors (Table 3). These moisture predictors are related to hydroperiod duration indicating that resources are available for breeding, growth, and development (Gomez-Rodriguez et al., 2009). In an ephemeral habitat, the longer the hydroperiod, the higher the probability that offspring and thus potential migrants will be produced contributing to a greater probability of gene flow (Husband and Barrett, 2002). We therefore suggest that fluctuations in precipitation (snow) directly affect variability in wetland availability and indirectly affect the conditions necessary for frog productivity and dispersal (Driscoll, 1997; Schwartz and Jenkins, 2000). We also suggest that precipitation flux affects between-wetland conditions (i.e., topographic roughness) which could confound dispersal costs (Funk et al., 2005; Semlitsch, 2008) and affect gene flow. Thus, frog populations that are faced with annual variability in precipitation both within- and between wetlands may rely on asynchronous dispersal dynamics to maintain genetic diversity.
Between-wetland Factors
Two between-wetland characteristics influenced P. maculata genetic connectivity: topography and moisture. Habitat and landscape controls on functional connectivity are crucial factors that facilitate species persistence and genetic diversity (Funk et al., 2005). In a montane region with relatively high levels of topographic roughness, amphibians are particularly susceptible to genetic isolation (Gomez-Rodriguez et al., 2009; Murphy et al., 2010b). Dispersal between wetlands is facilitated by more moisture suggesting that desiccation risk is an important limiting factor to dispersal. Therefore, topographic roughness inhibits amphibian movement while high-moisture landscape matrix—controlled by snowmelt—facilitates movement for this dispersal-limited species.
The stepping-stone wetland connectivity metrics were considered most important in explaining genetic connectivity despite a much lower magnitude in values relative to the spatial-breeding and temporal-breeding networks. While Euclidean distance alone was not a significant predicator in any of the best models, the presence of wetland habitat between the 18 occupied wetlands was a significant factor. Therefore, wetland spatial heterogeneity likely interacts with fluctuating environmental characteristics to affect amphibian genetic connectivity. Stepping-stone habitat improves a given habitat's probability of connectivity regardless of that habitat's area (Saura and Rubio, 2010). Considering dispersal limitations of P. maculata, the presence of stepping stone wetlands possibly improves the probability of dispersal success and gene flow among occupied wetlands and is likely important when there is high topographic between-wetland resistance. This result demonstrates the importance of the underlying habitat spatial heterogeneity compared to fluctuations of critical environmental factors potentially influencing population dynamics of a dispersal-limited species.
Climatic Fluctuation and Genetic Connectivity
Our results indicate that winter snowfall is essential to the amount and quality of wetland that is available to P. maculata and that high snowpack results in consistent inter-annual habitat availability for frogs. Thus, high snowpack is associated positively with gene flow. Because chorus frogs produce large numbers of propagules and show little parental investment, more wetland should facilitate greater production and increased colonization success (Corn, 2005), especially if wetter between-wetland matrix is also available. Conversely, low snowpack might result in fewer available wetlands and fewer stepping-stone wetlands. Thus, low snowpack scenarios may reflect decreased habitat availability, reduced reproduction, decreased colonization and less gene flow. Global models of climate change predict changes in precipitation, both in frequency and amount, and are suggested to impact montane species dramatically (Corn, 2005; Castillo et al., 2014).
Metapopulation Dynamics
We argue that fluctuating environmental conditions in heterogeneous landscapes have a potential role in structuring spatially-explicit populations, and could be important drivers of metapopulation dynamics. Theoretically, a classic metapopulation structure is defined by interbreeding subpopulations linked by dispersal and extinction-colonization dynamics (Smith and Green, 2005). As metapopulations are dynamic, they are influenced strongly by complex and interacting landscape characteristics that affect reproduction and dispersal capacity of individuals (Hanski and Ovaskainen, 2000). Based on the results of our study, we suggest that dramatic changes in available habitat and resistance between habitat patches (i.e., wetlands) influence the functional connectivity of a metapopulation where dispersal is limited, likely controlling genetic connectivity among amphibian subpopulations. Twenty-two of the total 35 sites in the sampling region were occupied in 2009–2010 (we analyzed only 18 in this study due to sample size limitations). Thirteen of the 22 occupied sites in this study were also occupied in an earlier study (21 total occupied sites, Spencer, 1964). However, we found frogs at nine additional sites that were described as lacking frogs in the earlier study (Spencer, 1964) and did not find frogs in eight of the sites described as occupied by frogs in the earlier study. Notably, some of the original occupied sites sampled in the 1964 study were no longer holding water or were considered unsuitable in 2009–2010. These observations suggest that shifts in occupancy have taken place in this landscape over the past 40+ years, likely due to succession and fluctuating snowpack patterns affecting the hydroperiod and availability of suitable within- and between-wetland habitat, similar to changes observed in amphibian habitats elsewhere (McMenamin et al., 2008; Hossack et al., 2015). Annual variability in precipitation (timing and amount) influences a spatially-explicit metapopulation structure (Hanski, 2001) because temporal fluctuations in biotic and abiotic factors can modify the availability of population sources and sinks over space and time (Consentino et al., 2012). If annual variability in precipitation (i.e., snowpack) has significant control over habitat availability, then asynchronous dynamics among subpopulations may be required to consistently recolonize wetlands. We expect this effect because improved connectivity among wetlands increases the ability of individuals to disperse and promotes genetic diversity within the metapopulation.
The effects of environmental controls on metapopulation dynamics are not limited to amphibians (Johst et al., 2002). For example, many turtle species are dispersal-limited and exhibit metapopulation structure (Souza et al., 2002), thus, environmental circumstances may be influencing their genetics. Similarly, fragmented forests affect patch colonization and metapopulation dynamics differently for three mammalian species depending on the species dispersal ability (Lawes et al., 2000), likely influencing genetic connectivity among subpopulations. Finally, African butterflies of the genus Bicyclus expressed coupled genetic and physiological plasticity in seasonally-fluctuating environments (Brakefield, 1997). In general, organismal dispersal capability seems to have an important role in population persistence and gene flow within fluctuating, dynamic landscapes and merits future research in landscape genetics.
Conclusions
Our study used landscape genetic and graph-theoretic connectivity methods to examine how interactions between habitat spatial heterogeneity and climatic variability can influence metapopulation dynamics. The inclusion of fluctuating habitat conditions on species dispersal, colonization, and genetic rescue effects is an essential contribution to advance our understanding of metapopulation ecology. Our work emphasizes the importance in expanding investigations of genetic signatures of populations in dynamic landscapes for multiple species, metapopulations, and metacommunities. Specifically, in a conservation context, these results hold considerable importance in predicting species future responses to human driven land-use and climate change. Considering the likelihood of future climate-driven shifts in precipitation, we expect that fluctuations in habitat availability will continue to affect the metapopulation capacity of P. maculata and other dispersal-limited pond-breeding species. We also expect that similar fluctuations may be observed in different landscapes. Empirical investigations of agricultural or urban environments where land use and climate changes may co-occur, such as other montane (Koscinski et al., 2009), agricultural (Youngquist and Boone, 2014), or urban (Hamer and Parris, 2011) landscapes, could be instructional in understanding how metapopulation dynamics are influenced in the face of modified habitat conditions. Reserve design strategies intended to maintain metapopulation persistence should consider underlying habitat spatial heterogeneity together with environmental conditions that influence the dispersal and genetic rescue of dispersal-limited species. Further, temporal data is an essential counterpart to addressing asynchronous metapopulation dynamics and may be the key driver to evaluating species persistence in future ephemeral landscapes. Thus, combined landscape genetic and graph-theoretic approaches to metapopulation ecology will help achieve a more holistic understanding of the complex landscape-climate interactions and species population persistence under dramatic environmental change.
Author Contributions
Conceived and designed the field work: MM, EM, WF. Performed the experiments: MM. Analyzed the genetic data: SB, BJ, SM, MM. Gravity modeling: MM, PS. Wetland connectivity: MF, AW. Wrote the paper: AW, PS, EM, MM. All authors provided substantial review and comments to written manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was part of the Landscape Genetics Distributed Graduate Seminar (2012 offering, organized by Helene Wagner, Lisette Waits and Melanie Murphy), funded by American Genetic Association, Canadian Institute for Ecology and Evolution, and Wyoming NASA Space Grant Consortium (NASA Grant #NNX10AO95H). Field and lab work were provided by the Colorado State University and United States Geological Survey. We acknowledge Jeffrey Evans for collaboration in developing gravity model R-code, Staci Amburgey and Robin Green for field work and Kim Hoke for study design. Unpublished microsatellite loci were provided by J. Van Buskirk. Permits were provided by the United States Forest Service and the Colorado Division of Wildlife. All animal procedures were permitted by the Colorado Division of Wildlife (# 09HP957) and Colorado State University Institutional Animal Care and Use Committee (# 09107A02). This is contribution number 515 of the U.S. Geological Survey Amphibian Research and Monitoring Initiative (ARMI). Use of trade, product, or firm names are descriptive and do not imply endorsement by the U.S. Government.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fgene.2015.00275
References
Akaike, H. (1974). A new look at the statistical model identification. IEEE Automat. Contr. 19, 716–723. doi: 10.1109/TAC.1974.1100705
Amburgey, S. M., Bailey, L. L., Murphy, M., Muths, E., and Funk, W. C. (2014). The effects of hydropattern and predator communities on amphibian occupancy. Can. J. Zool. 92, 927–937. doi: 10.1139/cjz-2014-0106
Arens, P., van der Sluis, T., van't Westende, W. P., Vosman, B., Vos, C. C., and Smulders, M. J. M. (2007). Genetic population differentiation and connectivity among fragmented Moor frog (Rana arvalis) populations in The Netherlands. Landscape Ecol. 22, 1489–1500. doi: 10.1007/s10980-007-9132-4
Anderson, J. E. (1979). A theoretical foundation for the gravity equation. Am. Econ. Rev. 69, 106–116.
Banks, S. C., Lawson, S. J., Finlayson, G. R., Lindenmayer, D. B., Ward, S. J., and Taylor, A. C. (2004). The effects of habitat fragmentation on demography and genetic variation in a marsupial carnivore. Biol. Conserv. 122, 581–597. doi: 10.1016/j.biocon.2004.09.013
Bartelt, P., and Peterson, C. R. (2005). Physical modeling operative temperature and evaporative water loss in amphibians. J. Therm. Biol. 30, 93–102. doi: 10.1016/j.jtherbio.2004.07.007
Bélise, M. (2005). Measuring landscape connectivity: the challenge of behavioral landscape ecology. Ecology 86, 1988–1995. doi: 10.1890/04-0923
Bowcock, A. M., Ruiz-Linares, A., Tomfohrde, J., Minch, E., Kidd, J. R., and Cavalli-Sforza, L. L. (1994). High resolution of human evolutionary trees with polymorphic microsatellites. Nature 368, 455–457. doi: 10.1038/368455a0
Brakefield, P. M. (1997). “Phenotypic plasticity and fluctuating asymmetry as responses to environmental stress in the butterfly Bicyclus anynana,” in Environmental Stress, Adaptation and Evolution, eds K. Bijlsma and V. Loeschcke (Berlin: Birkhäuser Basel), 65–78.
Brand, A. B., Snodgrass, J. W., Gallagher, M. T., Casey, R. E., and Van Meter, R. (2010). Lethal and sublethal effects of embryonic and larval exposure of Hyla versicolor to stormwater pond sediments. Arch. Environ. Contam. Toxicol. 58, 325–331. doi: 10.1007/s00244-009-9373-0
Brown, J. H., and Kodric-Brown, A. (1977). Turnover rates in insular biogeography: effect of immigration on extinction. Ecology 58, 445–449. doi: 10.2307/1935620
Browne, C. L., Paszkowski, C. A., Foote, A. L., Moenting, A., and Boss, S. M. (2009). The relationship of amphibian abundance to habitat features across spatial scales in the boreal plains. Ecoscience 16, 209–223. doi: 10.2980/16-2-3220
Burnham, K. P., and Anderson, D. R. (2002). Model Selection and Multimodel Inference: A Practical Information-theoretic Approach. Heidelberg; New York: Springer.
Castillo, J. A., Epps, C. W., Davis, A. R., and Cushman, S. A. (2014). Landscape effects on gene flow for a climate−sensitive montane species, the American pika. Mol. Ecol. 23, 843–856. doi: 10.1111/mec.12650
Consentino, B. J., Phillips, C. A., Schooley, R. L., Lowe, W. H., and Douglas, M. R. (2012). Linking extinction-colonization dynamics to genetic structure in a salamander metapopulation. Proc. Biol. Sci. 279, 1575–1682. doi: 10.1098/rspb.2011.1880
Corn, P. S., and Muths, E. (2002). Variable breeding phenology affects the exposure of amphibian embryos to ultraviolet radiation. Ecology 83, 2958–2963. doi: 10.1890/0012-9658(2002)083[2958:VBPATE]2.0.CO;2
Corn, P. S. (2003). Amphibian breeding and climate change: importance of snow in the mountains. Conserv. Biol. 17, 622–625. doi: 10.1046/j.1523-1739.2003.02111.x
Cushman, S. A. (2006). Effects of habitat loss and fragmentation on amphibians: a review and prospectus. Biol. Conserv. 128, 231–240. doi: 10.1016/j.biocon.2005.09.031
Dieringer, D., and Schlötterer, C. (2003). Microsatellite analyser (MSA): a platform independent analysis tool for large microsatellite data sets. Mol. Ecol. Notes 3, 167–169. doi: 10.1046/j.1471-8286.2003.00351.x
Driscoll, D. A. (1997). Mobility and metapopulation structure of Geocrinia alba and Geocrinia vitellina, two endangered frog species from southwestern Australia. Aust. J. Ecol. 22, 185–195. doi: 10.1111/j.1442-9993.1997.tb00658.x
Evans, I. S. (1972). “General geomorphometry, derivatives of altitude, and descriptive statistics,” in Spatial Analysis in Geomorphology, ed R. J. Chorley (New York, NY: Harper and Row), 17–90.
Ewers, R. M., and Didham, R. K. (2007). The effect of fragment shape and species' sensitivity to habitat edges on animal population size. Conserv. Biol. 21, 926–936. doi: 10.1111/j.1523-1739.2007.00720.x
Fahrig, L. (2003). Effects of habitat fragmentation on biodiversity. Annu. Rev. Ecol. Evol. Syst. 34, 487–515. doi: 10.1146/annurev.ecolsys.34.011802.132419
Fortuna, M. A., Gómez-Rodríguez, C., and Bascompte, J. (2006). Spatial network structure and amphibian persistence in stochastic environments. Proc. Biol. Sci. 273, 1429–1434. doi: 10.1098/rspb.2005.3448
Fotheringham, A. S., and O'Kelly, M. E. (1989). Spatial Interaction Models: Formulations and Applications. Dordrecht; Boston; London: Kluwer Academic Publishers.
Fry, J., Xian, G., Jin, S., Dewitz, J., Homer, C., Yang, L., et al. (2011). Completion of the 2006 National land cover database for the conterminous United States. Photogr. Eng. Remote Sens. 77, 858–864.
Funk, W. C., Blouin, M. S., Corn, P. S., Maxell, B. A., Pilliod, D. S., Amish, S., et al. (2005). Population structure of Columbia spotted frogs (Rana luteiventris) is strongly affected by the landscape. Mol. Ecol. 14, 483–496. doi: 10.1111/j.1365-294X.2005.02426.x
Gamble, L. R., McGarigal, K., and Compton, B. W. (2007). Fidelity and dispersal in pond-breeding amphibian, Ambystoma opacum: implications for spatio-temporal population dynamics and conservation. Biol. Conserv. 39, 247–257. doi: 10.1016/j.biocon.2007.07.001
Gesch, D., Oimoen, M., Greenlee, S., Nelson, C., Steuck, M., and Tyler, D. (2002). The National Elevation Dataset. Photogr. Eng. Remote Sens. 68, 5–11.
Girdner, S. F., and Larson, G. L. (1995). Effects of hydrology on zooplankton communities in high-mountain ponds, Mount Rainier National Park, USA. J. Plankton. Res. 17, 1731–1755. doi: 10.1093/plankt/17.9.1731
Giordano, A. R., Ridenhour, B. J., and Storfer, A. (2007). The influence of altitude and topography on genetic structure in the long-toed salamander (Ambystoma macrodactylum). Mol. Ecol. 16, 1625–1637. doi: 10.1111/j.1365-294X.2006.03223.x
Goldberg, C. S., Kaplan, M. E., and Schwalbe, C. R. (2003). From the frog's mouth: buccal swabs for collection of DNA from amphibians. Herp. Rev. 34, 220–221.
Goldberg, C. S., and Waits, L. P. (2009). Using habitat models to determine conservation priorities for pond-breeding amphibians in a privately-owned landscape of northern Idaho, USA. Biol. Conserv. 142, 1096–1104. doi: 10.1016/j.biocon.2009.01.025
Goldberg, C. S., and Waits, L. P. (2010). Comparative landscape genetics of two pond-breeding amphibian species in a highly modified agricultural landscape. Mol. Ecol. 19, 3650–3663. doi: 10.1111/j.1365-294X.2010.04673.x
Gomez-Rodriguez, C., Diaz-Paniqgua, C., Serrano, L., Florencio, M., and Porheault, A. (2009). Mediterranean temporary ponds as amphibian breeding habitats: the importance of preserving pond networks. Aquat. Ecol. 43, 1179–1191. doi: 10.1007/s10452-009-9235-x
Hamer, A. J., and Parris, K. M. (2011). Local and landscape determinants of amphibian communities in urban ponds. Ecol. Appl. 21, 378–390. doi: 10.1890/10-0390.1
Hanski, I., and Ovaskainen, O. (2000). The metapopulation capacity of a fragmented landscape. Nature 404, 755–758. doi: 10.1038/35008063
Hanski, I. (2001). Spatially realistic theory of metapopulation ecology. Naturwissenschaften 88, 372–381. doi: 10.1007/s001140100246
Hossack, B. R., Gould, W. R., Patla, D. A., Muths, E., Daley, R., Legg, K., et al. (2015). Amphibians on the U. S. Continental Divide: Status, trends, and the influence of beaver on occupancy dynamics in four national parks. Biol. Conserv. 187, 260–269.
Husband, B. C., and Barrett, S. C. H. (2002). Spatial and temporal variation in population size of Eichhornia paniculata in ephemeral habitats: implications for metapopulation dynamics. J. Ecol. 86, 1021–1031. doi: 10.1046/j.1365-2745.1998.00324.x
Jarvis, A., Reuter, H. I., Nelson, A., and Guevara, E. (2008). Hole-filled SRTM for the Globe Version 4, Available from the CGIAR-CSI SRTM 90m Database. Available online at: http://srtm.csi.cgiar.org
Johst, K., Brandl, R., and Eber, S. (2002). Metapopulation persistence in dynamic landscapes: the role of dispersal distance. OIKOS 98, 263–270. doi: 10.1034/j.1600-0706.2002.980208.x
Koscinski, D., Yates, A. G., Handford, P., and Lougheed, S. C. (2009). Effects of landscape and history on diversification of a montane, stream−breeding amphibian. J. Biogeogr. 36, 255–265. doi: 10.1111/j.1365-2699.2008.02009.x
Lawes, M. J., Mealin, P. E., and Piper, S. E. (2000). Patch occupancy and potential metapopulation dynamics of three forest mammals in fragmented afromontane forest in South Africa. Conserv. Biol. 14, 1088–1098. doi: 10.1046/j.1523-1739.2000.99120.x
Lemmon, M. E., Murphy, M., and Juenger, T. (2011). Identification and characterization of nuclear microsatellite loci for multiple species of chorus frogs (Pseudacris) for population genetic analyses. Conserv. Genet. Resour. 3, 233–237. doi: 10.1007/s12686-010-9330-2
Mazerolle, M. J. (2004). Amphibian road mortality in response to nightly variations in traffic intensity. Herpetologica 60, 45–53. doi: 10.1655/02-109
McMenamin, S. K., Hadly, E. A., and Wright, C. K. (2008). Climatic change and wetland desiccation cause amphibian decline in Yellowstone National Park. Proc. Natl. Acad. Sci. U.S.A. 105, 16988–16993. doi: 10.1073/pnas.0809090105
McRae, B. H., and Kavanagh, D. M. (2011). Linkage Mapper Connectivity Analysis Software. Seattle, WA: The Nature Conservancy. Available online at: http://www.circuitscape.org/linkagemapper
McRae, B. H. (2006). Isolation by resistance. Evolution 60, 1551–1561. doi: 10.1111/j.0014-3820.2006.tb00500.x
Mokany, K. (2007). How Communities Influence Ecosystem Processes: The Role of Species Diversity, Functional Diversity, and Functional Identity. Canberra, ACT: Australian National University.
Moore, I., Gessler, P., Nielsen, G., and Petersen, G. (1993). “Terrain attributes and estimation methods and scale effects,” in Modeling Change in Environmental Systems, eds A. Jakeman, M. Beck, and M. McAleer (London: Wiley), 189–214.
Munger, J. C., Gerber, M., Madrid, K., Carroll, M. A., Petersenand, W., and Heberger, L. (1998). U.S. National wetland inventory classifications as predictors of the occurrence of Columbia spotted frogs (Rana luteiventris) and pacific treefrogs (Hyla regilla). Consevr. Biol. 12, 320–330. doi: 10.1046/j.1523-1739.1998.97012.x
Murphy, M. A., Dezzani, R., Pilliod, D. S., and Storfer, A. (2010a). Landscape genetics of high mountain frog populations. Mol. Ecol. 19, 3634–3649. doi: 10.1111/j.1365-294X.2010.04723.x
Murphy, M. A., Evans, J. S., and Storfer, A. (2010b). Quantifying Bufo boreas connectivity in Yellowstone National Park with landscape genetics. Ecology 91, 252–261. doi: 10.1890/08-0879.1
Murphy, M., Dyer, R. J., and Cushman, S. A. (2015). “Network models in landscape genetics,” in Landscape Genetics: Concepts, Methods, Applications, eds N. Balkenhol, L. P. Waits, S. A. Cushman, and A. Storfer (New Jersey: John Wiley & Sons, Ltd.), 165–182.
National Agricultural Imagery Program (NAIP). (2010). Geospatial Data Gateway. Available online at: http://datagateway.nrcs.usda.gov (Accessed February, 2012).
Pilliod, D. S., Peterson, C. R., and Ritson, P. I. (2002). Seasonal migration of Columbia spotted frogs (Rana luteiventris) among complementary resources in a high mountain basin. Can. J. Zool. 80, 1849–1862. doi: 10.1139/z02-175
Rehfeldt, G. E., Crookston, N. L., Warwell, M. V., and Evans, J. S. (2006). Empirical analyses of plant−climate relationships for the western United States. Int. J. Plant. Sci. 167, 1123–1150. doi: 10.1086/507711
Rozenfeld, A. F., Arnaud-Haond, S., Hernandez-Garcia, E., Eguiliuz, V. M., Serrao, E. A., and Duarte, C. M. (2008). Network analysis identifies weak and strong links in a metapopulation system. Proc. Natl. Acad. Sci. U.S.A. 105, 18824–18829. doi: 10.1073/pnas.0805571105
Sanzo, D., and Hecnar, S. J. (2006). Effects of road de-icing salt (NaCl) on larval wood frogs (Rana sylvatica). Environ. Pollut. 140, 247–256. doi: 10.1016/j.envpol.2005.07.013
Saura, S., and Rubio, L. (2010). A common currency for the different ways in which patches and links can contribute to habitat availability and connectivity in the landscape. Ecography 33, 523–537. doi: 10.1111/j.1600-0587.2009.05760.x
Scherer, R. D., Muths, E., and Noon, B. R. (2012). The importance of local and landscape-scale processes to the occupancy of wetlands by pond-breeding amphibians. Popul. Ecol. 54 487–498. doi: 10.1007/s10144-012-0324-7
Schwartz, S. S., and Jenkins, D. G. (2000). Temporary aquatic habitats: constraints and opportunities. Aquat. Ecol. 34, 3–8. doi: 10.1023/A:1009944918152
Semlitsch, R. D. (2008). Differentiating migration and dispersal processes for pond-breeding amphibians. J. Wild. Manag. 72, 260–267. doi: 10.2193/2007-082
Smith, M. A., and Green, D. M. (2005). Dispersal and the metapopulation paradigm in amphibian ecology and conservation: are all amphibian populations metapopulations? Ecography 28, 110–128. doi: 10.1111/j.0906-7590.2005.04042.x
Snodgrass, J. W., Casey, R. E., Joseph, D., and Simon, J. A. (2008). Microcosm investigations of stormwater pond sediment toxicity to embryonic and larval amphibians: variation in sensitivity among species. Environ. Pollut. 154, 291–297. doi: 10.1016/j.envpol.2007.10.003
Souza, F. L., Cunha, A. F., Oliveira, M. A., Pereira, G. A., and Reis, S. F. D. (2002). Estimating dispersal and gene flow in the neotropical freshwater turtle Hydromedusa maximiliani (Chelidae) by combining ecological and genetic methods. Genet. Mol. Biol. 25, 151–155. doi: 10.1590/S1415-47572002000200007
Spencer, A. W. (1964). The Relationship of Dispersal and Migration to Gene Flow in the Boreal Chorus Frog. Ph.D. dissertation, Colorado State University, Fort Collins, CO.
Stow, A. J., and Sunnucks, P. (2004). Inbreeding avoidance in Cunningham's skink (Egernia cunninghamii) in natural and fragmented habitat. Mol. Ecol. 13, 443–447. doi: 10.1046/j.1365-294X.2003.02060.x
Tallmon, D. A., Luikart, G., and Waples, R. W. (2004). The alluring simplicity and complex reality of genetic rescue. TREE 19, 489–496. doi: 10.1016/j.tree.2004.07.003
Taylor, P. D., Fahrig, L., Henein, K., and Merriam, G. (1993). Connectivity is a vital element of landscape structure. OIKOS 68, 571–573. doi: 10.2307/3544927
Velo-Antón, G., Parra, J. L., Parra-Olea, G., and Zamudio, K. R. (2013). Tracking climate change in a dispersal-limited species: reduced spatial and genetic connectivity in a montane salamander. Mol. Ecol. 22, 3261–3278. doi: 10.1111/mec.12310
Wang, J. (2004). Sibship reconstruction from genetic data with typing errors. Genetics 166, 1963–1979. doi: 10.1534/genetics.166.4.1963
Weyrauch, S. L., and Grubb, T. C. Jr. (2004). Patch and landscape characteristics associated with the distribution of woodland amphibians in an agricultural fragmented landscape: an information-theoretic approach. Biol. Conserv. 115, 443–450. doi: 10.1016/S0006-3207(03)00161-7
Xian, G., Homer, C., Dewitz, J., Fry, J., Hossain, N., and Wickham, J. (2011). The change of impervious surface area between 2001 and 2006 in the conterminous United States. Photogr. Eng. Remote Sens. 77, 758–762.
Keywords: boreal chorus frog (Pseudacris maculata), functional connectivity, dispersal, gravity model, landscape genetics, metapopulation dynamics, spatio-temporal dynamics
Citation: Watts AG, Schlichting PE, Billerman SM, Jesmer BR, Micheletti S, Fortin M-J, Funk WC, Hapeman P, Muths E and Murphy MA (2015) How spatio-temporal habitat connectivity affects amphibian genetic structure. Front. Genet. 6:275. doi: 10.3389/fgene.2015.00275
Received: 29 May 2015; Accepted: 14 August 2015;
Published: 08 September 2015.
Edited by:
Andrew Shirk, University of Washington, USAReviewed by:
Peter J. Prentis, Queensland University of Technology, AustraliaJeffrey Ryan Row, University of Waterloo, Canada
Copyright © 2015 Watts, Schlichting, Billerman, Jesmer, Micheletti, Fortin, Funk, Hapeman, Muths and Murphy. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alexander G. Watts, Department of Ecology & Evolution, University of Toronto, 25 Harbord St., Toronto, ON M5S 3G5, Canada,YWxleGFuZGVyLndhdHRzQG1haWwudXRvcm9udG8uY2E=;
Melanie A. Murphy, Department of Ecosystem Science and Management, Program in Ecology, University of Wyoming, 1000 E. University Ave., Laramie, WY 82071, USA,bWVsYW5pZS5tdXJwaHlAdXd5by5lZHU=