 Ana C. Prizon1
Ana C. Prizon1 Daniel P. Bruschi2*
Daniel P. Bruschi2* Luciana A. Borin-Carvalho1Andréa Cius1Ligia M. Barbosa1
Luciana A. Borin-Carvalho1Andréa Cius1Ligia M. Barbosa1 Henrique B. Ruiz3Claudio H. Zawadzki3Alberto S. Fenocchio4Ana L. de Brito Portela-Castro1
Henrique B. Ruiz3Claudio H. Zawadzki3Alberto S. Fenocchio4Ana L. de Brito Portela-Castro1- 1Laboratório de Citogenética de Vertebrados, Departamento de Biotecnologia, Genética e Biologia Celular, Universidade Estadual de Maringá, Maringá, Brazil
- 2Laboratório de Citogenética Animal e Mutagênese Ambioental, Departamento de Genética, Universidade Federal do Paraná, Curitiba, Brazil
- 3Departamento de Biologia/Nupélia, Universidade Estadual de Maringá, Maringá, Brazil
- 4Facultad de Ciencias Exactas, Químicas y Naturales, Universidad Nacional de Misiones, Posadas, Argentina
Only one species of armored catfish, Ancistrus cirrhosus Valenciennes 1836, has been historically described in the basin of the Paraná River, from Misiones (Argentina). However, the ample variation found in the morphology and coloration of the populations sampled in the tributaries of the Brazilian state of Paraná makes it difficult to establish the real taxonomic status and evolutionary history of the Ancistrus specimens, suggesting that A. cirrhosus is not the only species found in this basin. By combining data on mitochondrial DNA (COI gene) and chromosomal markers from different Ancistrus populations, totaling 144 specimens, in the tributaries of the Paraná, and specimens from Misiones (type-locality of A. cirrhosus), we detected five distinct evolutionary lineages. All the specimens were 2n = 50, but had four distinct karyotype formulae. The results of the Generalized Mixed Yule Coalescent (GYMC) and the genetic distances (uncorrected P-values) between lineages ranged from 3 to 5%. Clusters of 18S rDNA were observed in a single chromosome pair in seven populations of Ancistrus, but at different positions, in some cases, in synteny with the 5S rDNA sites. Multiple 5S sites were observed in all populations. Overall, the cytogenetic data reinforce the genetic evidence of the diversification of lineages, and indicate the existence of candidate species in the study region. The evidence indicates that at least four candidate species of the Ancistrus may coexist in the Paraná basin besides A. cirrhosus. Overall, our results provide a comprehensive scenario for the genetic variation among Ancistrus populations and reinforce the conclusion that the true diversity of the freshwater fish of the Neotropical regions has been underestimated.
Introduction
The inclusion of genetic data in studies of taxonomy and evolution has had a profound influence on our understanding of the unique diversity of species in the Neotropical region (Pereira et al., 2011, 2013). The combination of approaches has altered our perceptions of the region's biological diversity and contributed to an increase in the rate of discovery of new species, especially in cryptic lineages. In this context, the delimitation of species using DNA sequences can be highly efficient, enabling the more systematic documentation of species diversity (e.g., Yang and Rannala, 2010; Ence and Carstens, 2011; Fujisawa and Barraclough, 2013). For example, the Generalized Mixed Yule Coalescent (GMYC) method has been designed to delimit potential lineages using information on a single locus, that is, this method considers that the mutations arising in one species cannot spread readily into another species (Ahearn and Templeton, 1989; Barraclough et al., 2003; De Queiroz, 2007). The premise of the GMYC method is that independent evolution leads to the emergence of distinct genetic clusters, separated by longer internal branches, optimizing the set of nodes that defines the transition between inter- and intra-specific processes (Barraclough et al., 2003).
Fish are an excellent candidate group for the application of integrative taxonomical methods, which combine different lines of evidence (DNA sequences, chromosomal data, and morphological features) to define taxonomic status. Most surveys of Neotropical freshwater fish have focused on major river basins, even though a large proportion of the total diversity comprises small species, found in minor rivers and streams. These species are often highly endemic and occupy a wide variety of microhabitats, providing enormous potential for diversification (Viana et al., 2013). Castro (1999) referred to the identification of the cryptic diversity of small freshwater fish as a major challenge for Neotropical Ichthyology. The reliable definition of species and their ranges is also essential to conservation strategies (Angulo and Icochea, 2010). In this context, the rivers and streams of the Paraná River basin provide an interesting study area for the evaluation of the degree to which the taxonomic diversity of these environments has been underestimated.
The Ancistrini is composed of 29 genera, with a total of 217 recognized species (Fisch-Muller, 2003). With 69 species, Ancistrus Kner, 1854 is one of the most diverse Ancistrini groups, the second richest in species of the Loricariidae (Ferraris, 2007; Bifi et al., 2009; Froese and Pauly, 2017). Cytogenetic data on Ancistrus are still scarce, and restricted to species found in the basins of the Paraguay River in Mato Grosso, and the Amazon, in Manaus (de Oliveira et al., 2009; Mariotto et al., 2011, 2013; Favarato et al., 2016; Prizon et al., 2016). While the cytogenetics of Ancistrus species from other river basins are still unknown, considerable variability has been found in this genus, with diploid (2n) numbers of 34, 38, 40, 42, 44, 48, 50, and 54 chromosomes. Surveys of the upper Paraná River have revealed the presence of a single species of Ancistrus in this basin, identified as Ancistrus cirrhosus (Langeani et al., 2007). However, the considerable variation in the morphology and coloration observed in the specimens collected in the tributaries of the Paraná River hampers the reliable identification of the Ancistrus species found in this region. Thus, given the karyotypic diversity of Ancistrus and the assumption that cryptic diversity exists in the rivers and streams of the Upper Paraná River basin, we combine chromosomal data and DNA sequences to evaluate the taxonomic status of these populations.
Materials and Methods
Biological Samples
A total of 144 Ancistrus specimens were collected in ten rivers of the Paraná River basin (Table 1, Figure 1). Specimen collection was authorized by the Brazilian Environment Ministry through its Biodiversity Information and Authorization System (SISBIO), under license number 36575-1. The protocols used in this study were submitted to the Ethics Committee on the use of animals in research (CEUA) of the Universidade Estadual de Maringá (UEM) and approved under case number 013/2009. Voucher specimens were deposited in the ichthyological collection of the Limnology, Ichthyology and Aquaculture Research Center (Nupélia) at Universidade Estadual de Maringá, Paraná, Brazil. The catalog numbers are provided in Table 1.
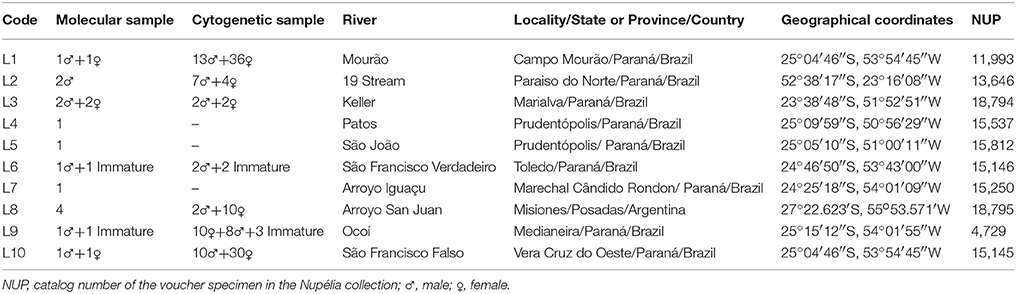
Table 1. Details of the Ancistrus populations and specimens sampled in the study area of the upper Paraná basin.
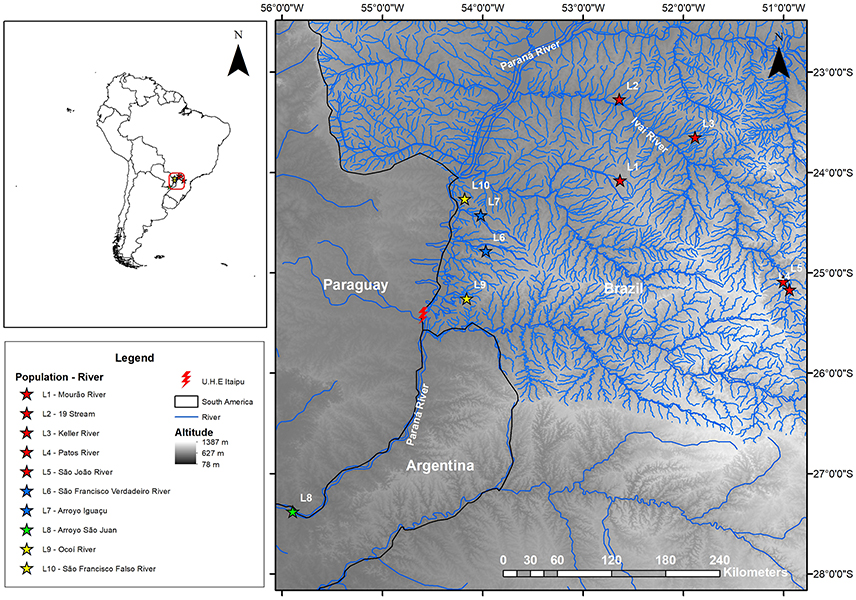
Figure 1. Topographic map showing the sites for the collection of specimens from the Ancistrus populations sampled in the Paraná River basin, in the present study. Each color represents a different clade recovered in our phylogenetic inferences (see Figure 2): red star, populations in clade I; blue star, populations in clade II; green star, populations in clade IV; and yellow star, populations in clade V. L1, Mourão River; L2, 19 Stream; L3, Keller River; L4, Patos River; L5, São João River; L6, São Francisco Verdadeiro River; L7, Arroyo Iguaçu; L8, Ancistrus cirrhosus; L9, Ocoí River; L10, São Francisco Falso River.
Isolation, Amplification, and Sequencing of the DNA
Genomic DNA was extracted from the liver, muscle tissue, or a cell suspension of a subset of the sample (Table 1) using the TNES method, as applied by Bruschi et al. (2012). A fragment of the mitochondrial cytochrome C oxidase subunit I (COI) gene was amplified by polymerase chain reaction (PCR) using the primers: FishF1 (5′-TCAACCAACCACAAAGACATTGGCAC-3′), FishR1 (5′-TAGACTTCTGGGTGGCCAAAGAATCA-3′) (Ward et al., 2005). The solution for the amplification reaction included 20 ng/μl of the DNA template, 7 pmol of the forward and reverse primers, 10 mM of dNTPs, 1 U Taq DNA Polymerase, 1.5 mM MgCl2, and 1x PCR buffer (200 mM Tris, pH 8.4, 500 mM KCL). The amplification protocol was 5 min−94°C/(94°C/30 s−60°C/1 min−72°C/30 s) 35 cycles/10 min−72°C. The amplified products were purified using Exonuclease I (10 units) and SAP (1 unit), incubated for 45 min 37°C, followed by denaturation at 85°C for 10 min (Applied Biosystems, Santa Clara, CA, USA), as recommended by the manufacturer. The samples were then used directly as templates for sequencing in an automatic ABI/Prism DNA sequencer (Applied Biosystems, Foster City, CA, USA) with the BigDye Terminator kit (Applied Biosystems, Foster City, CA, USA), as recommended by the manufacturer. The DNA samples were sequenced bidirectionally and were edited in Bioedit version 7.2.5 (http://www.mbio.ncsu.edu/bioedit/page2.html) (Hall, 1999).
Phylogenetic Inferences and the Delimitation of Species
The phylogenetic relationships among the populations were inferred from the matrix of the 459-bp sequence of the COI gene. The dataset was complemented with 35 sequences of Ancistrus and one sequence of the sister group Lasiancistrus available in GenBank (Supplementary File 1). The outgroup was Pseudolithoxus sp., which was chosen based on the arrangement reported by Lujan et al. (2015). The sequence was aligned using Clustal W in BioEdit, version 7.2.5.0 (Thompson et al., 1994). The initial alignments were checked visually and adjusted wherever necessary. The dataset was used for phylogenetic reconstruction by Bayesian inference (BI) and the Maximum Parsimony (MP) approach.
Bayesian Inference (BI) method was applied to the dataset, which were divided into three partitions according to codon position for mit-COI. The best model of nucleotide evolution for each nucleotide partition was determined using Akaike Information Criterion (AIC) with the software jModelTest v2.1.6 (Guindon and Gascuel, 2003; Darriba et al., 2012). The BI was performed with the software Mr. Bayes 3.2.6 (Ronquist and Huelsenbeck, 2003), as available in the CIPRES Science Gateway 3.1 (Miller et al., 2010). BI was implemented using two independent runs, each starting from random trees, with four simultaneous independent chains, and performed 10,000,000 generations, keeping one tree every 1,000th generation. Of all trees sampled, 20% were discarded as burn-in and checked by the convergence criterion (frequencies of average standard deviation of split < 0.01) with Tracer v.1.6 (Rambaut et al., 2014), while the remaining were used to reconstruct a 50% majority-rule consensus tree and to estimate Bayesian posterior probabilities (BPP) of the branches. A node was considered to be strongly supported if it had a BPP ≥ 0.95, while moderate support was considered when BPP ≥ 0.9.
The MP analysis was implemented in TNT v1.1 (Goloboff et al., 2003) using a heuristic search method with tree bisection-reconnection (TBR) swapping and 100 random additional replicates. The bootstrap values of the branches inferred in this analysis were calculated with 1,000 non-parametric pseudoreplicates.
The genetic distances among and within species were calculated using the Kimura-2-Parameter (K2P) and p-distance model (Kimura, 1980) implemented in MEGA v 6.0. A neighbor-joining (NJ) tree of K2P distances was created to provide a graphic representation of the patterning of divergence between species with the software MEGA v 7.0 (Kumar et al., 2016). We also applied the General Mixed Yule-coalescent (GMYC) method to delineate species using single-locus sequence data. The GMYC requires a fully resolved and ultrametric tree as input for the analysis and combines a coalescence model of intraspecific branching with a Yule model for interspecific branching to estimate species boundaries and provide statistical confidence intervals to evaluate the sequences of the clusters recovered. Ultrametric trees were constructed by a BI tree in BEAST2 2.4.0 (Drummond et al., 2006; Drummond and Rambaut, 2007). We conducted three independent runs using different priors, that is, the Yule, relaxed clock, and constant coalescent models. An ultrametric gene tree was obtained for each prior. An XML file was produced using the BEAUti2 v2.4.5 interface with the following settings: GTR+G+I substitution model, previously inferred by MrMODELTEST (Nylander et al., 2004), empirical base frequencies, four gamma categories, all codon positions partitioned with unlinked base frequencies and substitution rates. The MCMC chain was 10 million generations long, and was logged every 1,000 generations. The Estimated Sample Sizes (ESS) and trace files of the runs were evaluated in Tracer v1.6. The resulting logs were analyzed in TREEANNOTATOR 2.4.4, with 25% burn-in, maximum clade credibility trees with a 0.5 posterior probability limit, and node heights of the target tree. The splits package (https://r-forge.r-project.org/R/?group_id=333) in R was used for the GMYC calculations, using the single-threshold strategy and default scaling parameters.
Cytogenetic Analysis
All specimens were anesthetized and euthanized by an overdose of clove oil (Griffiths, 2000). Mitotic chromosomes were obtained from kidney cells according to Bertollo et al. (1978). The AgNORs were revealed by the silver nitrate impregnation technique (Howell and Black, 1980). The regions of heterochromatin were determined by the C-banding technique (Sumner, 1972) and stained with propidium iodide according to the method of Lui et al. (2012). Physical mapping of the 5S rDNA and 18S rDNA sequences was carried out by fluorescence in situ hybridization (FISH) according to Pinkel et al. (1986), with probes obtained from Leporinus elongatus Valenciennes, 1850 (Martins and Galetti, 1999) and Prochilodus argenteus Spix et Agassiz, 1829 (Hatanaka and Galetti, 2004). We also isolated and cloned the rDNA 5S gene of the Ancistrus sample from Keller River using a more specific DNA probe for this gene. The genomic DNA extracted for the molecular analyses was used as the template for the amplification reaction, using the primers 5S-A (5′-TACGCCCGATCTCGTCCGATC-3′) and 5S-B (5′-CAGGCTGGTATGGCCGTAAGC-3′) (Pendas et al., 1994). The products of this amplification were isolated in the 1.5% agarose gel, then purified with an EasyPure® Quick Gel Extraction kit. These sequences were inserted into a linearized cloning vector (pJET1.2/blunt) by the CloneJET PCR Cloning kit (Thermo Scientific) and cloned in Escherichia coli. Ten clones with the insert containing the 5S rDNA sequence were selected for sequencing. The 5S rDNA nucleotide sequences were edited using BioEdit software and compared with sequences from GenBank database (www.ncbi.nlm.nih.gov).
Hybridization was conducted under high stringency conditions (77%), and the probes were labeled by nick translation with digoxigenin-11-dUTP (5S rDNA) and biotin-16-dUTP (18S rDNA). The 5S rDNA probes obtained from the recombinant plasmids were labeled by PCR using the fluorochrome digoxigenin-11-dUTP. The solution for labeling reaction included 20 ng/μl of the DNA template, 7 pmol of the forward and reverse primers, 4 mM of dNTPs, 1 mM of dig-11-dUTP, 1 U Taq DNA Polymerase, 1.5 mM MgCl2, and 1x PCR buffer (200 mM Tris, pH 8.4, 500 mM KCL). The hybridization signals were detected using anti-digoxigenin-rhodamine for the 5S rDNA probe and avidin-FITC (fluorescein isothiocyanate) for the 18S rDNA probe. The chromosomes were counterstained with DAPI. Double staining was carried out with chromomycin A3 (CMA3) and DAPI, according to Schweizer (1976). The metaphases were photographed using an epifluorescence microscope and adjusted for best contrast and brightness using the Adobe Photoshop CS6 software.
Results
Phylogenetic Inferences and Delimitation of Species
The phylogenetic reconstructions based on the MP and BI approaches produced the basic topology of the dataset (Figures 2, 3). Topology inferred from the Bayesian analysis performed with the software Mr. Bayes 3.2.6. and from Neighbor-Joining were presented in the supplementary material (Figures S1, S2, respectively). These analyses recovered five clades from the Ancistrus populations of the Paraná basin. The first clade (clade I) comprises five populations: Ancistrus sp. “Mourão River” (L1) + Ancistrus sp. “19 Stream” (L2) + Ancistrus sp. “Keller River” (L3) + Ancistrus sp. “Patos River” (L4) + Ancistrus sp. “João River” (L5). The genetic distance analysis returned low uncorrected P-distances among these populations, ranging from 0.0 to 1.8% (Table 2). The second clade (clade II) included the Ancistrus populations from São Francisco Verdadeiro River (L6) and Ancistrus sp. from Arroyo Iguaçu (L7), separated by a genetic distance of 2.0% (Table 2). Two sequences deposited in Genbank as A. “cirrhosus” included in our dataset were recovered in the clade III. Clade IV included only the population from Arroyo San Juan (L8), Argentina, and was considered to represent the nominal A. cirrhosus due to its proximity to the type-locality of this species. Clade V consisted of the Ancistrus sp. “Ocoí River” (L9) + Ancistrus sp. “São Francisco Falso River” (L10) populations. The uncorrected P-distance between these populations (1.2%) was also relatively low (Table 2). In all cases, by contrast, the genetic distances (uncorrected P-values) between clades were at least 3%, being 3% between clades I–II, I–III, II–III, II–IV, III–IV; 4% between clades I–IV, II–V, III–V, IV–V and finally a genetic distance of 5% between clades I–V.
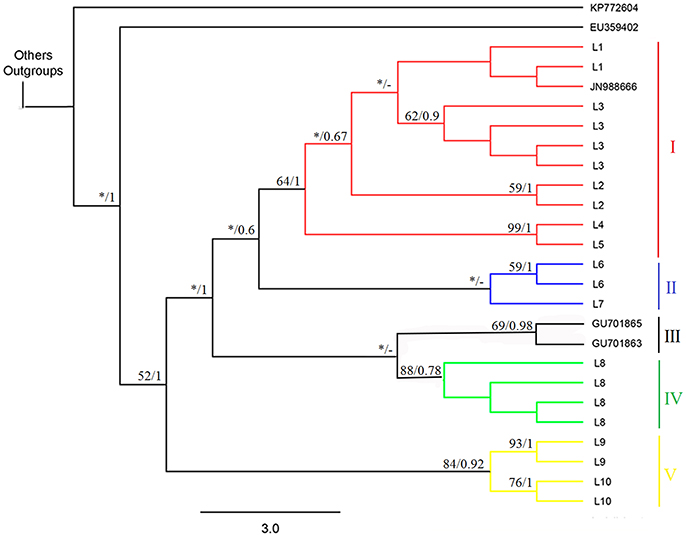
Figure 2. Strict consensus cladogram produced by the Maximum Parsimony analysis showing the intrageneric relationships of the Ancistrus species from the Paraná basin, based on a 459-bp sequence of the COI gene. The numbers above the branches show the bootstrap support and Bayesian posterior probabilities, respectively. Each color represents one of the five evolutionary lineages recovered in the analysis. The terminal species with alphanumerical identifiers were obtained from GenBank (Supplementary File 1). The asterisks indicate nodes with bootstrap values of < 50. The trace indicates that the node has not been recovered by Bayesian inference. Codes: L1, Mourão River; L2, 19 Stream; L3, Keller River; L4, Patos River; L5, São João River; L6, São Francisco Verdadeiro River; L7, Arroyo Iguaçu; L8, Ancistrus cirrhosus; L9, Ocoí River; L10, São Francisco Falso River.
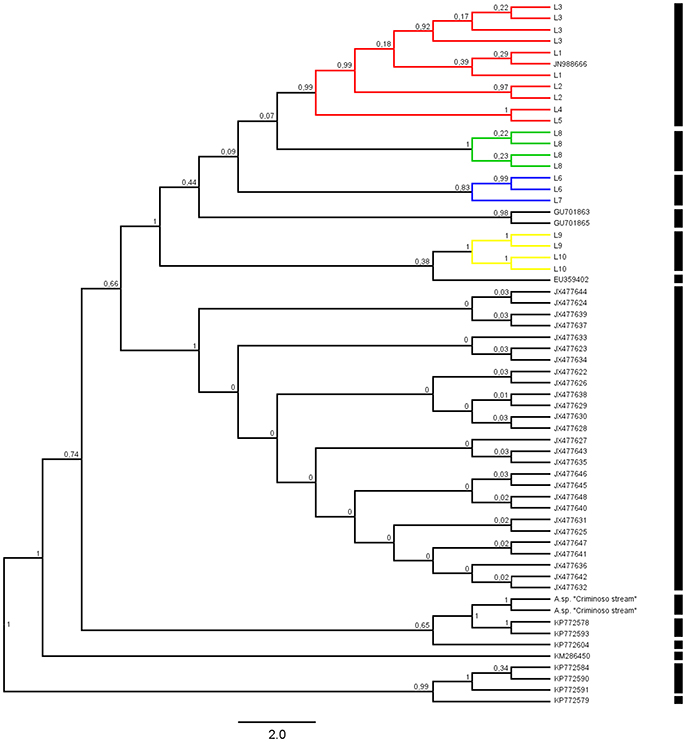
Figure 3. Bayesian consensus tree for the Ancistrus species of the Paraná River basin produced in BEAST2 2.4.0 and obtained from 10 million generations. Posterior probabilities are shown above the branches. Each color represents one of the five evolutionary lineages recovered. The results of the GMYC method using the Yule model of species delimitation are shown as black bars to the right of the phylogeny. Species access numbers are specified in the supplementary material (Supplementary File 1). Codes: L1, Mourão River; L2, 19 Stream; L3, Keller River; L4, Patos River; L5, São João River; L6, São Francisco Verdadeiro River; L7, Arroyo Iguaçu; L8, Ancistrus cirrhosus; L9, Ocoí River; L10, São Francisco Falso River.
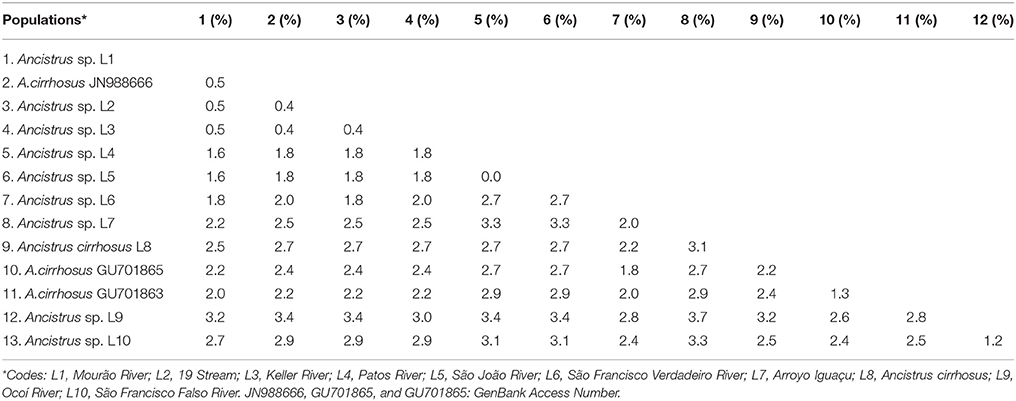
Table 2. Uncorrected pairwise distances between the mitochondrial COI sequences of the Ancistrus populations from the Paraná basin.
The ultrametric trees obtained using the Yule and Constant Coalescent priors were congruent in the GMYC analysis, recognizing 13 separate entities and identifying nine clusters in our dataset (including outgroup sequences). Considering only the Ancistrus populations, the focus of the present study, the sequences were grouped into five well-supported clusters (Figure 3), which corresponded exactly to the major clades recovered in our phylogenetic inferences (BI and MP). The Yule and Constant Coalescent priors of the branching rates indicated that the likelihood of the null model (i.e., that all the sequences belong to the same species) was 412.5381 and 421.5504, respectively, with a likelihood of 421.9409 and 430.6968, respectively, for the GMYC model (i.e., the existence of distinct species). The difference is highly significant, indicating the presence of more than one species in our sample. The analysis based on the Relaxed clock priors presented low scores in Tracer and did not produce a reliable interpretation of the phylogenetic relationships among the populations, and was not considered in the species delimitation tests.
Cytogenetic Analysis
Cytogenetic data were obtained for the populations from the Mourão, Keller, São Francisco Verdadeiro, São Francisco Falso, and Ocoi Rivers, and 19 Stream (Brazil), and Arroyo San Juan (Argentina). All the specimens analyzed presented a diploid number of 2n = 50 chromosomes, although four distinct karyotype formulae were detected (Table 3, Figure 4), together with variation in the location of the 18S and 5S rDNA sites (Figure 5). Interestingly, these formulae corresponded strongly with the major clades recovered in our genetic analysis, with exception of the clade III that have no karyotype available. A chromosome heteromorphism found in all the males of Stream 19 (L2) and Keller River (L3) populations were consistent with an XX/XY system. The X chromosome is a large metacentric (second pair) while the Y chromosome is a small subtelocentric (Figure 4).
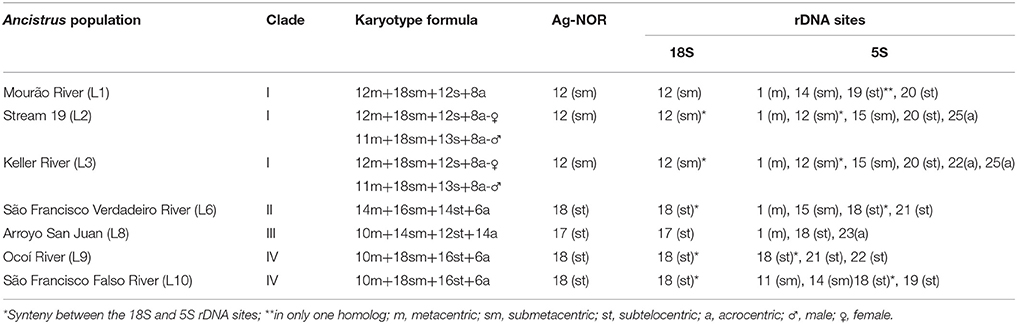
Table 3. Cytogenetic parameters of the Ancistrus populations sampled in the different rivers of the Paraná basin.
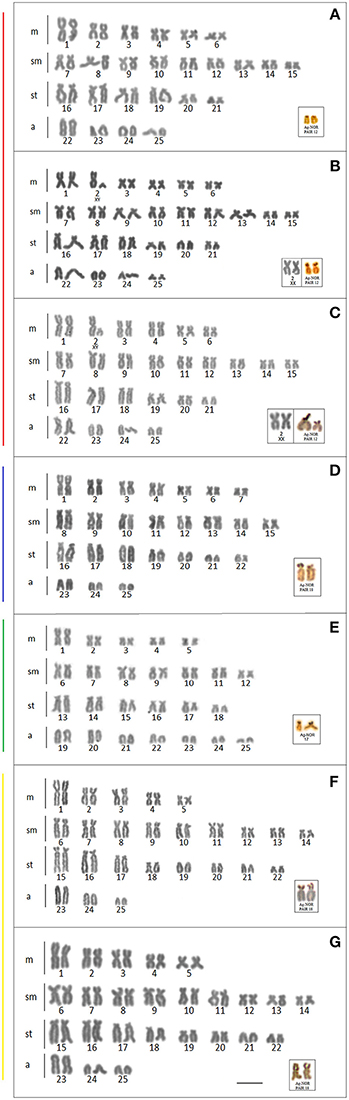
Figure 4. Karyotype of the Ancistrus populations sampled in different rivers of the Paraná basin, stained with Giemsa. The configuration of the silver nitrate-stained nucleolar organizing regions (Ag-NORs) are shown in the box. Each color of the side bars represents one of the evolutionary lineages recovered in the analysis, according to Figure 2. (A) L1: Mourão River; (B) L2: 19 Stream; (C) L3: Keller River; (D) L6: São Francisco Verdadeiro River; (E) L8: Ancistrus cirrhosus; (F) L9: Ocoí River; (G) L10: São Francisco Falso River. Bar = 10 μm.

Figure 5. Chromosomes of the Ancistrus populations of species after dual color-FISH showing the 5S rDNA (red) and 18S rDNA (green) sites. Each color of the side bars represents one of the evolutionary lineages recovered in the analysis, according to Figure 2. (A) L1: Mourão River; (B) L2: 19 Stream; (C) L3: Keller River; (D) L6: São Francisco Verdadeiro River; (E) L8: Ancistrus cirrhosus; (F) L9: Ocoí River; (G) L10: São Francisco Falso River. Bar = 10 μm.
The analysis of the heterochromatin revealed C-positive blocks at the pericentromeric and subterminal positions in a number of different chromosomal pairs of the populations allocated to clade I, with a conspicuous block coinciding with the NOR site of pair 12 in all karyotypes (Figures 6A–C). The X chromosome (pair 2, metacentric) from 19 Stream had a C-positive block in the pericentromeric region, while we detected heterochromatin blocks in the pericentromeric, interstitial, and subterminal regions of the X chromosome from Keller River. The Y chromosome of the Keller River population had a weak heterochromatin block in the subterminal region of the long arm, which was not found in the Y chromosome from 19 Stream (Figures 6B,C). In clades II, IV, and V, considerable variation was found in the amount and distribution of constitutive heterochromatin, which was concentrated primarily in the pericentromeric and subterminal regions of the chromosomes (Figures 6D–G). In the population from the São Francisco Falso River (clade V), in addition, we detected an interstitial heterochromatin block in pair 17, as well as a much larger amount of heterochromatin distributed throughout the chromosomes in comparison with the other populations (Figure 6G). Conspicuous blocks of heterochromatin were also found in the nucleolar pairs of all the populations analyzed in clades II and IV (pair 18) (Figures 6D,E).
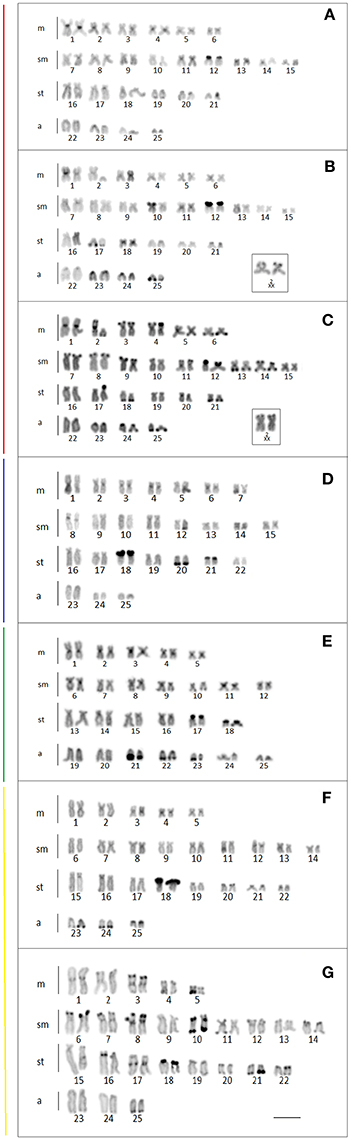
Figure 6. Karyotypes of the Ancistrus populations showing the distribution of the heterochromatin after C-banding. Each color of the side bars represents one of the evolutionary lineages recovered in the analysis, according to Figure 2. (A) L1, Mourão River; (B) L2, 19 Stream; (C) L3, Keller River; (D) L6, São Francisco Verdadeiro River; (E) L8, Ancistrus cirrhosus; (F) L9, Ocoí River; (G) L10, São Francisco Falso River. Bar = 10 μm.
The base-specific fluorochrome staining enhanced the distinctive composition of some heterochromatin blocks. In particular, staining with Chromomycin A3 revealed a richness of G and C in the subterminal and pericentromeric heterochromatin blocks of all the populations analyzed, and provides additional features for the comparative analysis. As expected, conspicuous C-positive areas were detected in the NOR-bearing chromosomes (pair 12, 17, and 18) (Figure S3).
Discussion
Molecular Phylogenetic Inferences Reveal a Number of Distinct Lineages in the Paraná Basin
The phylogenetic reconstructions and cytogenetic analyses presented in this study both detected the presence of five major clades among the Ancistrus populations surveyed, pointing to the existence of at least five lineages within the Paraná basin in Brazil previously undetected. Historically, all Ancistrus from the Paraná River basin were assigned to a single species, A. cirrhosus, which was described by Valenciennes (1836) from specimens collected in the Province of Misiones and Buenos Aires (Argentina), although the diagnostic traits are weakly defined (Langeani et al., 2007). Despite the morphological variations and wide geographic distribution, the Ancistrus populations of the Paraná basin have been identified invariably as A. cirrhosus. This fact has intrigued the ichthyologists with regard to the taxonomic status of many specimens, using only morphological and meristic characters. Therefore, the data obtained in the present study recovered at least five independent lineages (clades I–V) in Paraná basin. Given the proximity of the Arroyo San Juan to the type-locality of A. cirrhosus, this population (clade IV) was identified as nominal species A. cirrhosus. In this context, the other four clades (I, II, III, and V) can be categorized as “candidate species” in the terminology of Vieites et al. (2009). This interpretation is also supported by the results of the GMYC method, and the genetic distance detected among populations, considering a 2% threshold for interspecific differentiation.
The GMYC method has become one of the most popular tools for the delimitation of species based on single-locus data, and has been applied to the analysis of a number of poorly-known groups of organism (Barraclough et al., 2009; Monaghan et al., 2009; Marshall et al., 2011; Vuataz et al., 2011; Roxo et al., 2015). The GMYC method uses an ultrametric tree derived from the sequences to identify shifts in the branching rate from the Yule model (species) to the coalescent (population) process, with the algorithm computing the probability of splits between lineages in relation to speciation rates, thus identifying a cutoff value, at which species and populations split from one another (Powell, 2012). This approach was highly effective in the present study, allowing us to delimit five groups, which four corresponded exactly with the chromosomal data (unfortunately, no chromosomal data available to clade III), reinforcing the hypothesis that four “candidate species” exist in the Paraná River basin, besides A. cirrhosus as previously reported.
Armbruster (2004) found a sister group relationship between the Ancistrini and Pterygoplichthini tribes, which both belong to the subfamily Hypostominae. Ancistrus is the most species-rich genus of the tribe Ancistrini, although phylogenetic analyses of this genus are still scarce. Lujan et al. (2015) adopted a broad approach to the phylogenetic relationships of the Loricariidae, a Neotropical catfish family, but included few Ancistrus species, which reinforces the need for further research. Studies in systematics based on osteological (Schaefer, 1987) and molecular (Montoya-Burgos et al., 1998) data show that Ancistrus constitutes a monophyletic group of species. The lack of any robust phylogenetic tree for Ancistrus still limits our understanding of its intrageneric relationships, and more detailed analyses, with a more representative dataset, are needed to provide a more comprehensive understanding of the phylogenetic relationships of this genus.
The Chromosomal Data Reinforce the Hypothesis of Complete Lineage Divergence
Cytogenetic studies, together with molecular and biochemical analyses, may be useful for the identification of cryptic species (Nakayama et al., 2001; Milhomen et al., 2007). In the present study, in fact, the inclusion of a cytogenetic approach was crucial to the recognition of the “candidate species,” given that the distinct chromosomal formulae found in the four lineages (clades I, II, IV, and V) emphasizes their reciprocal monophyly and the lack of gene flow between them. The results obtained also complement the available cytogenetic data to the Ancistrus species, since these studies are currently restricted to the taxa found in the Paraguay (Mato Grosso) and Amazon (Manaus) basins (de Oliveira et al., 2007, 2008, 2009; Mariotto et al., 2011, 2013; Favarato et al., 2016; Prizon et al., 2016).
In the present study, a diploid number of 2n = 50 chromosomes was recorded in all the samples from the Paraná River basin. This number is within the range recorded for the genus, which vary from 2n = 34 in Ancistrus cuiabae to 2n = 54 in Ancistrus claro (Mariotto et al., 2009, 2013; Favarato et al., 2016; Prizon et al., 2016). Despite the homogeneity of the diploid number, the karyotype formula varied considerably among populations. The number of acrocentric chromosomes, for example, varied from three to four pairs in the populations of clades I, II, and V, to seven pairs in clade IV (Arroyo San Juan). This points evidences to the occurrence of chromosomal rearrangements, such as translocation and pericentric inversions, which did not affect the diploid number. Unfortunately, while we were able to identify these features, we were unable to trace the pathways of the transformations due to the lack of resolution in the internal topology of the Ancistrus lineages recognized here. In this case, further phylogenetic studies, based on a larger set of characters and a multi-locus dataset, may provide more definitive insights into the evolution of this group. Among all the ancistrinis species studied so far, the karyotypic evolution of Ancistrus is invariably associated with chromosomal rearrangements, which typically involve variation in the diploid number, given that this number ranges from 34 to 54 in this genus. As Artoni and Bertollo (2001) considered 2n = 54 to be the ancestral diploid number of the Loricariidae, karyotypic evolution in Ancistrus appears to have been associated with a reduction in the chromosome number. In fact, de Oliveira et al. (2009) suggested that the karyotypic evolution of this genus was predominantly involves by centric fusions.
In Ancistrus the occurrence of heteromorphic sex chromosomes has been well-documented in some species, including simple systems (Mariotto et al., 2004; Alves et al., 2006; Mariotto and Miyazawa, 2006) and multiples (de Oliveira et al., 2007, 2008) which also contributed to the karyotype evolution in the genus (Favarato et al., 2016). One peculiar feature observed in clade I was the heteromorphic sex chromosomes found in the populations of 19 Stream (L2) and the Keller River (L3), which are consistent with an XX/XY system. Surprisingly, however, this feature was not observed in the specimens from the Mourão River (L1), which were included in clade I and separated by low genetic distances from the populations of Stream 19 and the Keller River. The karyotypic formula of the Mourão River population differs from these two other populations only by the absence of the pair of sex chromosomes, and the GYMC analysis also identified these populations as a unique taxonomical unit. A similar result was obtained by Henning et al. (2011), combining cytogenetic and molecular data for different species of Eigenmannia, included two populations of E. virescens (Mogi-Guaçu and Tietê rivers), whose karyotypes with 2n = 38 differ by the presence of a pair sexual XX/XY (Tietê river population). The acrocentric X chromosome possesses a heterochromatinized distal region (Almeida-Toledo et al., 2001) and according to Henning et al. (2011), both populations (Mogi-Guaçu e Tietê rivers) were considered sister species. Furthermore, these authors concluded that seems likely that suppression of recombination in the homologous pair of acrocentric chromosomes and accumulation of heterochromatin on the X chromosome occurred after a recent geographical separation. In this context, we hypothesized that the heteromorphic sex chromosome found in the Ancistrus populations of 19 Stream and the Keller River represent a recent event which may have occurred after the geographical isolation of these populations from that of the Mourão River, together with the behavioral characteristics of these fish, which occupy specific microhabitats, form territories, and do not normally migrate (Power, 1984, 1990; Buck and Sazima, 1995). All these characteristics favor the fixation of chromosomal rearrangements and could be contribute to allopatric speciation on a micro scale (de Oliveira et al., 2009).
The hypothesis of the recent differentiation of the sex chromosome pair in the Ancistrus populations of clade I was also supported by the C-banding data. Heterochromatin is widely used in the identification of the sex chromosomes, and the addition or deletion of heterochromatin or the occurrence of a pericentric inversion involving one of the chromosomes have been postulated as important mechanisms in the origin of simple sexual chromosome systems in Neotropical fish (Almeida-Toledo et al., 2000). If we compare the Y chromosome of males from 19 Stream (L2) and Keller River (L3), it is possible to detect a discreet heterochromatin block in the subterminal region of the long arm of the Keller River males, which was not detected in the Y chromosomes from 19 Stream. The presence of the heteromorphic pair in the Ancistrus population of 19 Stream and Keller River suggests that chromosomal rearrangements (inversions), the loss of chromosomal material and, in the specific case of the Keller River, the presence of constitutive heterochromatin in the heteromorphic pair, may all be evidence of the recent origin of the Y chromosome, derived from a large metacentric, similar to the X chromosome. Pericentric inversions, followed by a loss of chromosomal material, have been suggest as a mechanism to explain the origin of the ZZ/ZW chromosome system in Ancistrus sp. Piagaçu (de Oliveira et al., 2007) and the XX/XY chromosome systems in Ancistrus sp. Purus and Ancistrus sp. Macoari (de Oliveira et al., 2009), given absence of the heterochromatin blocks in either the X or Y chromosomes.
The NOR mapping provided an excellent marker in the present study, being found in a single chromosome pair in each clade, a condition shared with most other Ancistrus species (Medeiros et al., 2016). The variation among clades in the NOR-bearing chromosome may be the result of chromosomal rearrangements occurring during chromosomal evolution. We also recorded synteny between the 18S and 5S rDNA sites in most populations of clades I, II, and V, except for the population from Mourão River (clade I). This synteny of the rDNA may represent the basal condition for the genus (Mariotto et al., 2011), as it is found in A. claro (2n = 54), although synteny of the ribosomal sites was not found in the population from Arroyo San Juan (clade IV). The position and distribution of the 5S rDNA sites varied considerably among the four clades, as they do in other Ancistrus species, occupying multiple sites in pericentromeric, interstitial, or terminal positions. This variation is considered to be an important reflection of the enormous karyotypic diversity found in the Ancistrus, which is seen as evidence of the apomorphic condition of this group (Medeiros et al., 2016). This variability, together with the disjunction of the ribosomal sites caused by rearrangements or mobile genetic elements appears to be a common condition among Neotropical fish species (de Oliveira et al., 2009). We observed heterochromatin in association with the ribosomal DNA sites (18 and 5S), recurrent characteristic in the karyotypes of Neotropical fishes (Vicari et al., 2003). The presence of heterochromatin, which contains large quantities of transposable elements (Dimitri et al., 2009) may facilitate transposition events, moving ribosomal genes around the genome (Moreira-Filho et al., 1984; Vicari et al., 2008; Gross et al., 2009, 2010). This may be one of the factors responsible for the presence of the multiple 5S ribosomal DNA sites found in the present study.
Conclusion
The cytogenetic data available for the genus Ancistrus indicate a highly heterogeneous pattern of chromosome evolution, marked by Robertsonian and non-Robertsonian rearrangements. While we do not have an exact understanding of the mechanisms that determine these rearrangements in natural populations, their fixation may either initiate or contribute to the divergence process, with specific implications for the utility of chromosomal characters for phylogenetic inference (Sites and Kent, 1994). As in other fish groups, the sex chromosomes, present in some Ancistrus species, may have contributed to high rates of evolution. The inferences obtained in the present study from a combined approach of molecular and cytogenetic analyses further corroborate the taxonomic complexity of this genus. This approach was especially important due to the lack of diagnostic features in the morphology of these fishes. The hidden diversity of the study populations was nevertheless decoded successfully by the combined approach, which allowed us to differentiate five distinct lineages of Ancistrus, reinforcing the hypothesis of the presence of at least four candidate species in the upper Paraná River basin, besides of the A. cirrhosus, previously described. Finally, our findings reinforce the observation that the true diversity of the freshwater fish of the Neotropical has been underestimated and improve our understanding of a regional diversity within Ancistrus genus.
Author Contributions
AP: provided chromosomal and molecular data, and drafted the manuscript; DB: designed and coordinated the study of molecular data and helped draft the manuscript; HR and CZ: collected specimens from Paraná state and helped to identify the specimens; AF: collected and processed material of specimens from Arroyo San Juan, Argentina; LB-C, AC, and LB: assisted in the execution and analysis of chromosomal banding; AdBP-C: designed and coordinated the study of cytogenetic data and helped draft the manuscript. All authors have read and approved the final manuscript.
Funding
This work was funded by CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, Brazil) and Fundação Araucária.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank the Brazilian agency CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, Brazil) for financial support, Maringá State University (UEM) and Limnology, Ichthyology and Aquaculture Research Center (Nupélia) for the logistic support, the collection, and identification of species. We also thank Prof. Dr. Mateus Arduvino Reck and Prof. Dr. Alessandra Valéria de Oliveira for their assistance and contribution in the programs of phylogeny.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2017.00185/full#supplementary-material
Figure S1. Topology inferred from the Bayesian analysis performed with the software Mr. Bayes 3.2.6. posterior probabilities are shown at each node. Scale bar represents the number of substitutions per site. The terminal species with alphanumerical identifiers were obtained from GenBank (Supplementary File 1). Codes: L1, Mourão River; L2, 19 Stream; L3, Keller River; L4, Patos River; L5, São João River; L6, São Francisco Verdadeiro River; L7, Arroyo Iguaçu; L8, Ancistrus cirrhosus; L9, Ocoí River; L10, São Francisco Falso River.
Figure S2. Chromosomes of the Ancistrus populations of species after CMA3 staining. Each color of the side bars represents one of the evolutionary lineages recovered in the analysis, according to Figure 2. (a) L1, Mourão River; (b) L2, 19 Stream; (c) L3, Keller River; (d) L6, São Francisco Verdadeiro River; (e) L8, Ancistrus cirrhosus; (f) L9, Ocoí River; (g) L10, São Francisco Falso River. Bar = 10 μm.
Figure S3. NJ dendrograma of the Ancistrus specimens. Node values = bootstrap test (1,000 pseudo replicas) are shown next to the branches. The terminal species with alphanumerical identifiers were obtained from GenBank (Supplementary File 1). Codes: L1, Mourão River; L2, 19 Stream; L3, Keller River; L4, Patos River; L5, São João River; L6, São Francisco Verdadeiro River; L7, Arroyo Iguaçu; L8, Ancistrus cirrhosus; L9, Ocoí River; L10, São Francisco Falso River.
References
Ahearn, J. N., and Templeton, A. R. (1989). Interspecific hybrids of Drosophila heteroneura and D. silvestris I. Courtship success. Evolution 43, 347–361. doi: 10.1111/j.1558-5646.1989.tb04232.x
Almeida-Toledo, L. F., Foresti, F., Daniel, M. F., and Toledo-Filho, S. A. (2000). Sex chromosome evolution in fish: the formation of the neo-Y chromosome in Eigenmannia (Gymnotiformes). Chromosoma 109, 197–200. doi: 10.1007/s004120050428
Almeida-Toledo, L. F., Foresti, F., Pequignot, E. V., and Daniel-Silva, M. F. Z. (2001). XX:XY sex chromosome system with X heterocromatinization: an early stage of sex chromosome differentiation in the Neotropic eletric eel Eigenmannia virescens. Cytogenet. Cell Genet. Res. 95, 73–78. doi: 10.1159/000057020
Alves, Á. L., Oliveira, C., Nirchio, M., Granado, A., and Foresti, F. (2006). Karyotypic relationships among the tribes of Hypostominae (Siluriformes: Loricariidae) with description of XO sex chromosome system in a Neotropical fish species. Genetica 128, 1–9. doi: 10.1007/s10709-005-0715-1
Angulo, A., and Icochea, J. (2010). Cryptic species complexes, widespread species and conservation: lessons from Amazonian frogs of the Leptodactylus marmoratus group (Anura: Leptodactylidae). Syst. Biodivers. 8, 357–370. doi: 10.1080/14772000.2010.507264
Armbruster, J. W. (2004). Phylogenetic relationships of the suckermouth armoured catfishes (Loricariidae) with emphasis on the Hypostominae and the Ancistrinae. Zool. J. Linn. Soc. 141, 1–80. doi: 10.1111/j.1096-3642.2004.00109.x
Artoni, R. F., and Bertollo, L. A. (2001). Trends in the karyotype evolution of Loricariidae fish (Siluriformes). Hereditas 134, 201–210. doi: 10.1111/j.1601-5223.2001.00201.x
Barraclough, T. G., Birky, C. W., and Burt, A. (2003). Diversification in sexual and asexual organisms. Evolution 57, 2166–2172. doi: 10.1111/j.0014-3820.2003.tb00394.x
Barraclough, T. G., Hughes, M., Ashford-Hodges, N., and Fujisawa, T. (2009). Inferring evolutionarily significant units of bacterial diversity from broad environmental surveys of single-locus data. Biol. Lett. 5, 425–428. doi: 10.1098/rsbl.2009.0091
Bertollo, L. A. C., Takahashi, C. S., and Moreira-Filho, O. (1978). Cytotaxonomic considerations on Hoplias lacerdae (Pisces, Erythrinidae). Rev. Bras. Genet. 1, 103–120.
Bifi, A., Pavanelli, G. C. S., and Zawadzki, C. H. (2009). Three new species of Ancistrus Kner, 1854 (Siluriformes: Loricariidae) from the Rio Iguaçu basin, Paraná State, Brazil. Zootaxa 2275, 41–59. doi: 10.5281/zenodo.191064
Bruschi, D. P., Busin, C. S., Siqueira, S., and Recco-Pimentel, S. M. (2012). Cytogenetic analysis of two species in the Phyllomedusa hypochondrialis group (Anura, Hylidae). Hereditas 149, 34–40. doi: 10.1111/j.1601-5223.2010.02236.x
Buck, S., and Sazima, I. (1995). An assemblage of mailed catfishes (Loricariidae) in southeastern Brazil: distribuition, activity, and feeding. Ichthyol. Explor. Freshw. 6, 325–332.
Castro, R. M. C. (1999). “Evolução da ictiofauna de riachos sul-americanos: padrões gerais e possíveis processos gerais e possíveis processos casuais,” in Ecologia de Peixes de Riachos, eds E. P. Caramashi and R. Mazzoni e Peres-Neto (Rio de Janeiro: PPGE Instituto de Biologia), 139–155.
Darriba, D., Taboada, G. L., Doallo, R., and Posada, D. (2012). jModelTest 2: more models, new heuristics and parallel computing. Nat. Methods 9:772. doi: 10.1038/nmeth.2109
de Oliveira, R. R., Feldberg, E., Anjos, M. B., and Zuanon, J. (2007). Karyotype characterization and ZZ/ZW sex chromosome heteromorphism in two species of the catfish genus Ancistrus Kner, 1854 (Siluriformes: Loricariidae) from the Amazon basin. Neotrop. Ichthyol. 5, 301–306. doi: 10.1590/S1679-62252007000300010
de Oliveira, R. R., Feldberg, E., Anjos, M. B., and Zuanon, J. (2008). Ocurrence of multiple sexual chromosomes (XX/XY1Y2 and Z1Z1Z2Z2/Z1Z2W1W2) in catfishes of the genus Ancistrus (Siluriformes, Loricariidae) from the Amazon Basin. Genetica 134, 243–249. doi: 10.1007/s10709-007-9231-9
de Oliveira, R. R., Feldberg, E., Anjos, M. B., and Zuanon, J. (2009). Mechanisms of chromosomal evolution and its possible relation to natural history characteristics in Ancistrus catfishes (Siluriformes: Loricariidae). J. Fish Biol. 75, 2209–2225. doi: 10.1111/j.1095-8649.2009.02450.x
De Queiroz, K. (2007). Species concepts and species delimitation. Syst. Biol. 56, 879–886. doi: 10.1080/10635150701701083
Dimitri, P., Caizzi, R., Giordano, E., Carmela Accardo, M., Lattanzi, G., and Biamonti, G. (2009). Constitutive heterochromatin: a surprising variety of expressed sequences. Chromosoma 118, 419–435. doi: 10.1007/s00412-009-0211-y
Drummond, A. J., Ho, S. Y., Phillips, M. J., and Rambaut, A. (2006). Relaxed phylogenetics and dating with confidence. PLoS Biol. 4:e88. doi: 10.1371/journal.pbio.0040088
Drummond, A. J., and Rambaut, A. (2007). BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 7:214. doi: 10.1186/1471-2148-7-214
Ence, D. D., and Carstens, B. C. (2011). SpedeSTEM: a rapid and accurate method for species delimitation. Mol. Ecol. Resour. 11, 473–480. doi: 10.1111/j.1755-0998.2010.02947.x
Favarato, R. M., Silva, M., Oliveira, R. R., Artoni, R. F., Feldberg, E., and Matoso, D. A. (2016). Cytogenetic diversity and the evolutionary dynamics of rDNA genes and telomeric sequences in the Ancistrus genus (Loricariidae: Ancistrini). Zebrafish 13, 103–111. doi: 10.1089/zeb.2015.1140
Ferraris, C. J. Jr. (2007). Checklist of catfishes, recent and fossil (Osteichthyes: Siluriformes), and catalogue of Siluriform primary types. New Zealand. Zootaxa 1418:628. doi: 10.11646/zootaxa.1418.1.1
Fisch-Muller, S. (2003). “Subfamily Ancistrinae (Armored catfishes),” in Check List of the Freshwater Fishes of South and Central America, eds R. E. Reis, S. O. Kullander Jr., and C. J. Ferraris (Porto Alegre: Museu de Ciências e Tecnologia), 373–400.
Froese, R., and Pauly, D. (2017). FishBase: World Wide Web Electronic Publication [Dataset]. Available online at: http://www.fishbase.org
Fujisawa, T., and Barraclough, T. G. (2013). Delimiting species using single-locus data and the generalized mixed yule coalescent (GMYC) approach: a revised method and evaluation on simulated datasets. Syst. Biol. 62, 707–724. doi: 10.1093/sysbio/syt033
Goloboff, P. A., Farris, J. S., and Nixon, K. C. (2003). T.N.T.: Tree Analysis Using New Technology. Program and Documentation. Available online at: http://www.lillo.org.ar/phylogeny/
Griffiths, S. P. (2000). The use clove oil as an anaesthetic and method for sampling intertidal rockpool fishes. J. Fish Biol. 57, 1453–1464. doi: 10.1111/j.1095-8649.2000.tb02224.x
Gross, M. C., Schneider, C. H., Valente, G. T., Martins, C., and Feldberg, E. (2010). Variability of 18S rDNA locus among Symphysodon fishes: chromosomal rearrangements. J. Fish Biol. 76, 1117–1127. doi: 10.1111/j.1095-8649.2010.02550.x
Gross, M. C., Schneider, C. H., Valente, G. T., Porto, J. I., Martins, C., and Feldberg, E. (2009). Comparative cytogenetic analysis of the genus Symphysodon (Discus Fishes, Cichlidae): chromosomal characteristics of retrotransposons and minor ribosomal DNA. Cytogenet. Genome Res. 127, 43–53. doi: 10.1159/000279443
Guindon, S., and Gascuel, O. (2003). A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 52, 696–704. doi: 10.1080/10635150390235520
Hall, T. (1999). Bioedit Program. Available online at: http://www.mbio.ncsu.edu/BioEdit/bioedit.html
Hatanaka, T., and Galetti, P. M. Jr. (2004). Mapping of the 18S and 5S ribosomal RNA genes in the fish Prochilodus argenteus Agassiz, 1829 (Characiformes, Prochilodontidae). Genetica 122, 239–244. doi: 10.1007/s10709-004-2039-y
Henning, F., Moyses, C. B., Calcagnotto, D., Meyer, A., and Almeida-Toledo, L. F. (2011). Independent fusions and recent origins of sex chromosomes in the evolution and diversification of glass knife fishes (Eigenmannia). Heredity 106, 391–400. doi: 10.1038/hdy.2010.82
Howell, W. M., and Black, D. A. (1980). Controlled silver staining of nucleolus organizer regions with a protective colloidal developer: a 1-step method. Experientia 36, 1014–1015. doi: 10.1007/BF01953855
Kimura, M. (1980). A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 16, 111–120. doi: 10.1007/BF01731581
Kumar, S., Stecher, G., and Tamura, K. (2016). MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 33, 1870–1874. doi: 10.1093/molbev/msw054
Langeani, F., Castro, R. M. C., Oyakawa, O. T., Shibatta, O. A., Pavanelli, C. S., and Casatti, L. (2007). Diversidade da ictiofauna do Alto Rio Paraná: composição atual e perspectivas futuras. Biota Neotrop. 7, 181–197. doi: 10.1590/S1676-06032007000300020
Lui, R. L., Blanco, D. R., Moreira-Filho, O., and Margarido, V. P. (2012). Propidium iodide for making heterochromatin more evident in the C-banding technique. Biotech. Histochem. 87, 433–438. doi: 10.3109/10520295.2012.696700
Lujan, N. K., Armbruster, J., Lovejoy, N. R., and López-Férnandez, H. (2015). Multilocus molecular phylogeny of the suckermouth armored catfishes (Siluriformes: Loricariidae) with a focus on subfamily Hypostominae. Mol. Phylogenet. Evol. 82, 269–288. doi: 10.1016/j.ympev.2014.08.020
Mariotto, S., Artoni, R. F., and Miyazawa, C. S. (2004). Occurence of sexual chromosome, of the type ZZ/ZW, in Ancistrus cf. dubius (Loricariidae, Ancistrinae) of the Paraguay River Basin, Mato Grosso, Brazil. Caryologia 57, 327–331. doi: 10.1080/00087114.2004.10589413
Mariotto, S., Centofante, L., Miyazawa, C. S., Bertollo, L. A. C., and Moreira-Filho, O. (2009). Chromosome polymorphism in Ancistrus cuiabae Knaack, 1999 (Siluriformes: Loricariidae: Ancistrini). Neotrop. Ichthyol. 7, 595–600. doi: 10.1590/S1679-62252009000400006
Mariotto, S., Centofante, L., and Moreira-Filho, O. (2013). Diversity and chromosomal evolution in the genus Ancistrus Kner, 1854 (Loricariidae: Ancistrini) from three hydrographic basins of Mato Grosso State, Brazil. Neotrop. Ichthyol. 11, 125–131. doi: 10.1590/S1679-62252013000100015
Mariotto, S., Centofante, L., Vicari, M. R., Artoni, R. F., and Moreira-Filho, O. (2011). Chromosomal diversification in ribosomal DNA sites in Ancistrus Kner, 1854 (Loricariidae, Ancistrini) from three hydrographic basins of Mato Grosso, Brazil. Comp. Cytogenet. 5, 289–300. doi: 10.3897/compcytogen.v5i4.1757
Mariotto, S., and Miyazawa, C. S. (2006). Ancistrus cf. dubius (Siluriformes, Ancistrinae), a complex of species. 1. Chromosomal characterization of four populations and occurrence of sex chromosomes of the type XX/XY, in the Pantanal Basin of Mato Grosso, Brazil. Caryologia 59, 299–304. doi: 10.1080/00087114.2006.10797929
Marshall, D. C., Hill, K. B., Cooley, J. R., and Simon, C. (2011). Hybridization, mitochondrial DNA phylogeography, and prediction of the early stages of reproductive isolation: lessons from New Zealand cicadas (Genus Kikihia). Syst. Biol. 60, 482–502. doi: 10.1093/sysbio/syr017
Martins, C., and Galetti, P. M. Jr. (1999). Chromosomal localization of 5S rDNA genes in Leporinus fish (Anostomidae, Characiformes). Chromosome Res. 7, 363–367. doi: 10.1023/A:1009216030316
Medeiros, L. A., Ginani, E. G., Sousa, L. M., Py-Daniel, L. H. R., and Feldberg, E. (2016). Cytogenetic analysis of Baryancistrus xanthellus (Siluriformes: Loricariidae: Ancistrini), an ornamental fish endemic to the Xingu River, Brazil. Neotrop. Ichthyol. 14:e150108. doi: 10.1590/1982-0224-20150108
Milhomen, S. S. R., Pieczarka, J. C., Crampton, W. G. R., Souza, A. C. P., Carvalho, J. R. Jr., et al. (2007). Differences in karyotype between two sympatric species of Gymnotus (Gymnotiformes: Gymnotidae) from the easern amazon of Brazil. Zootaxa 1397, 55–62. doi: 10.11646/zootaxa.1397.7
Miller, G. H., Pfeiffer, W., and Schwartz, T. (2010). “Creating the CIPRES Science Gateway for inference of large phylogenetic trees,” in Proceeding of the Gatway Computing Environments Workshop (New Orleands, LA), 1–8.
Monaghan, M. T., Wild, R., Elliot, M., Fujisawa, T., Balke, M., Inward, D. J., et al. (2009). Accelerated species inventory on Madagascar using coalescent-based models of species delineation. Syst. Biol. 58, 298–311. doi: 10.1093/sysbio/syp027
Montoya-Burgos, J. I., Fisch-Muller, S., Weber, C., and Pawlowski, J. (1998). “Phylogenetic relationships of the Loricariidae (Siluriformes) based on mitochondrial rRNA gene sequences,” in Phylogeny and Classification of Neotropical Fishes, eds L. R. Malabarba, R. E. Reis, R. P. Vari, Z. M. S. Lucena, and C. A. S. Lucena (Porto Alegre: Edipucrs), 363–374.
Moreira-Filho, O. L., Bertollo, A. C., and Galetti, P. M. Jr. (1984). Structure and variability of nucleolar organizer regions in Parodontidae fish. Can. J. Genet. Cytol. 26, 564–568. doi: 10.1139/g84-089
Nakayama, C. M., Jégu, M., Porto, J. I. R., and Feldberg, E. (2001). Karyological evidence for a cryptic species of piranha within Serrasalmus rhombeus (Characidae, Serrasalminae) in the Amazon. Copeia 3, 866–869. doi: 10.1643/0045-8511(2001)001[0866:KEFACS]2.0.CO;2
Nylander, J. A., Ronquist, F., Huelsenbeck, J. P., and Nieves-Aldrey, J. L. (2004). Bayesian phylogenetic analysis of combined data. Syst. Biol. 53, 47–67. doi: 10.1080/10635150490264699
Pendas, A. M., Moran, P., Freije, J. P., and Garcia-Vazquez, E. (1994). Chromosomal mapping and nucleotide sequence of two tandem repeats of Atlantic salmon 5S rDNA. Cytogenet. Genome Res. 67, 31–36. doi: 10.1159/000133792
Pereira, L. H., Hanner, R., Foresti, F., and Oliveira, C. (2013). Can DNA barcoding accurately discriminate megadiverse Neotropical freshwater fish fauna? BMC Genet. 14:e20. doi: 10.1186/1471-2156-14-20
Pereira, L. H. G., Pazian, M. F., Hanner, R., Foresti, F., and Oliveira, C. (2011). DNA barcoding reveals hidden diversity in the Neotropical freshwater fish Piabina argentea (Characiformes: Characidae) from the Upper Paraná Basin of Brazil. Mitochondrial DNA 22, 87–96. doi: 10.3109/19401736.2011.588213
Pinkel, D., Straume, T., and Gray, J. W. (1986). Cytogenetic analysis using quantitative, high-sensitivity, fluorescence hybridization. Proc. Natl. Acad. Sci. U.S.A. 83, 2934–2938. doi: 10.1073/pnas.83.9.2934
Powell, J. R. (2012). Accounting for uncertainty in species delineation during the analysis of environmental DNA sequence data. Methods Ecol. Evol. 3, 1–11. doi: 10.1111/j.2041-210X.2011.00122.x
Power, M. E. (1984). The importance of sediment in the grazing ecology and size class interactions of an armored catfish, Ancistrus spinosus. Environ. Biol. Fishes 10, 173–181. doi: 10.1007/BF00001124
Power, M. E. (1990). Resource enhancement by indirect effects of grazers: armored catfish, algae, a sediment. Ecology 71, 897–904. doi: 10.2307/1937361
Prizon, A. C., Borin-Carvalho, L. A., Bruschi, D. P., Ribeiro, M. O., Barbosa, L. M., Ferreira, G. E., et al. (2016). Cytogenetic data on Ancistrus sp. (Siluriformes, Loricariidae) of the Paraguay River basin (MS) sheds light on intrageneric karyotype diversification. Comp. Cytogenet. 10, 625–636. doi: 10.3897/compcytogen.v10i4.8532
Rambaut, A., Suchard, M. A., Xie, D., and Drummond, A. J. (2014). Tracer v1.6. Available online at: http://tree.bio.ed.ac.uk/software/tracer/
Ronquist, F., and Huelsenbeck, J. P. (2003). MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 19, 1572–1574. doi: 10.1093/bioinformatics/btg180
Roxo, F. F., Ochoa, L. E., Costa-Silva, G. J., and Oliveira, C. (2015). Species delimitation in Neoplecostomus (Siluriformes: Loricariidae) using morphologic and genetic approaches. DNA Barcodes 3, 110–117. doi: 10.1515/dna-2015-0014
Schaefer, S. A. (1987). Osteology of Hypostomus plecostomus (Linnaeus), with a phylogenetic analysis of the loricariid subfamilies (Pisces: Siluroidei). Contribut. Sci. 394, 1–31.
Schweizer, D. (1976). Reverse fluorescent chromosome banding with chromomycin and DAPI. Chromossoma 58, 307–324. doi: 10.1007/BF00292840
Sites, J. W., and Kent, M. (1994). Chromosomal evolution, speciation, and systematics: some relevant issues. Herpetologica 50, 237–249.
Sumner, A. T. (1972). A simple technique for demonstrating centromeric heterochromatin. Exp. Cell Res. 75, 304–306. doi: 10.1016/0014-4827(72)90558-7
Thompson, J. D., Higgins, D. G., and Gibson, T. J. (1994). CLUSTAL W: improving the sensitivity of progressive multiple sequence alignments through sequence weighting, position specific gap penalties and weight matrix choice. Nucleic Acids Res. 22, 4673–4680. doi: 10.1093/nar/22.22.4673
Viana, D., Zawadzki, C. H., Oliveira, E. F., Vogel, H. F., and Graça, W. J. (2013). Structure of the ichthyofauna of the Bonito river, Ivaí river basin, upper Paraná river system, Brazil. Biota Neotrop. 13, 219–226. doi: 10.1590/S1676-06032013000200021
Vicari, M. R., Artoni, R. F., and Bertollo, L. A. C. (2003). Heterochromatin polymorphism associated with 18S rDNA. A diferrential pathway among the fish Hoplias malabaricus from Southern Brazil. Cytogenet. Genome Res. 101, 24–28. doi: 10.1159/000073413
Vicari, M. R., Noleto, R. B., Artoni, R. F., Moreira-Filho, O., and Bertollo, L. A. C. (2008). Comparative cytogenetics among species of the Astyanax scabripinnis complex. Evolutionary and biogeographical inferences. Genet. Mol. Biol. 31, 173–179. doi: 10.1590/S1415-47572008000200002
Vieites, D. R., Wollenberg, K. C., Andreone, F., Köhler, J., Glaw, F., and Vences, M. (2009). Vast underestimation of Madagascar's biodiversity evidenced by an integrative amphibian inventory. Proc. Natl. Acad. Sci. U.S.A. 106, 8267–8272. doi: 10.1073/pnas.0810821106
Vuataz, L., Sartori, M., Wagner, A., and Monaghan, M. T. (2011). Toward a DNA taxonomy of alpine Rhithrogena (Ephemeroptera: Heptageniidae) using a mixed Yule-coalescent analysis of mitochondrial and nuclear DNA. PLoS ONE 6:e19728. doi: 10.1371/journal.pone.0019728
Ward, R. D., Zemlak, T. S., Innes, B. H., Last, P. R., and Hebert, P. D. N. (2005). DNA barcoding Australia's fish species. Philos. Trans. R. Soc. Lond. B Biol. Sci. 360, 1847–1857. doi: 10.1098/rstb.2005.1716
Keywords: Ancistrus genus, cytotaxonomy, species delimitation, candidate species, chromosomal evolution
Citation: Prizon AC, Bruschi DP, Borin-Carvalho LA, Cius A, Barbosa LM, Ruiz HB, Zawadzki CH, Fenocchio AS and Portela-Castro ALdB (2017) Hidden Diversity in the Populations of the Armored Catfish Ancistrus Kner, 1854 (Loricariidae, Hypostominae) from the Paraná River Basin Revealed by Molecular and Cytogenetic Data. Front. Genet. 8:185. doi: 10.3389/fgene.2017.00185
Received: 12 August 2017; Accepted: 07 November 2017;
Published: 24 November 2017.
Edited by:
Roberto Ferreira Artoni, Ponta Grossa State University, BrazilReviewed by:
Sandra Mariotto, Instituto Federal de Educação, Ciência e Tecnologia de Mato Grosso, BrazilShane Lavery, University of Auckland, New Zealand
Copyright © 2017 Prizon, Bruschi, Borin-Carvalho, Cius, Barbosa, Ruiz, Zawadzki, Fenocchio and Portela-Castro. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Daniel P. Bruschi, ZGFuaWVscGFjaGVjb2JydXNjaGlAZ21haWwuY29t