 Violeta López-Márquez1
Violeta López-Márquez1 José Templado1
José Templado1 David Buckley1,2,3
David Buckley1,2,3 Ilaria Marino4
Ilaria Marino4 Elisa Boscari4Dragos Micu5
Elisa Boscari4Dragos Micu5 Lorenzo Zane4,6
Lorenzo Zane4,6 Annie Machordom1*
Annie Machordom1*- 1Museo Nacional de Ciencias Naturales – Consejo Superior de Investigaciones Científicas, Madrid, Spain
- 2Centre D’Estudis Avançats de Blanes – Consejo Superior de Investigaciones Científicas, Girona, Spain
- 3Departamento de Biología (Unidad de Genética), Universidad Autónoma de Madrid, Madrid, Spain
- 4Department of Biology, University of Padova, Padova, Italy
- 5National Institute for Marine Research and Development “Grigore Antipa”, Constanta, Romania
- 6Consorzio Nazionale Interuniversitario per le Scienze del Mare, Rome, Italy
Genetic connectivity studies are essential to understand species diversity and genetic structure and to assess the role of potential factors affecting connectivity, thus enabling sound management and conservation strategies. Here, we analyzed the patterns of genetic variability in the marine snail Gibbula divaricata from five coastal locations in the central-south Adriatic Sea (central Mediterranean) and one in the adjacent northern Ionian Sea, using 21 described polymorphic microsatellite loci. Observed and expected heterozygosity varied from 0.582 to 0.635 and 0.684 to 0.780, respectively. AMOVA analyses showed that 97% of genetic variation was observed within populations. Nevertheless, significant, although small, genetic differentiation was found among nearly all of the pairwise FST comparisons. Over a general pattern of panmixia, three groups of populations were identified: eastern Adriatic populations, western Adriatic populations, and a third group represented by the single northern Ionian Sea population. Nonetheless, migration and gene flow were significant between these groups. Gibbula divaricata is thought to have a limited dispersal capacity related to its lecithotrophic trochophore larval stage. Our results indicated high levels of self-recruitment and gene flow that is mainly driven through coastline dispersion, with populations separated by the lack of suitable habitats or deep waters. This stepping-stone mode of dispersion together with the high levels of self-recruitment could lead to higher levels of population structuring and differentiation along the Adriatic Sea. Large effective population sizes and episodic events of long-distance dispersal might be responsible for the weak differentiation observed in the analyzed populations. In summary, the circulation system operating in this region creates natural barriers for dispersion that, together with life-history traits and habitat requirements, certainly affect connectivity in G. divaricata. However, this scenario of potential differentiation seems to be overridden by sporadic events of long-distance dispersal across barriers and large effective population sizes.
Introduction
Connectivity among populations determines the dynamics of metapopulation systems, how genetic diversity arises and is maintained within species, and the adaptability and resilience of populations to environmental changes, among other factors (Botsford et al., 2001). Defining connectivity patterns for marine organisms, however, is a challenging task since factors that affect connectivity (e.g., life history traits, habitat, hydrological regime, occurrence of geological/topographical boundaries, layout of coastline, etc.) act at very different geographic and temporal scales (Villamor et al., 2014). For instance, processes related to dispersion, which ultimately determine patterns of connectivity, are highly linked to the biology and ecology of species, but they are also contingent on the evolutionary history of the group and the geological history of the inhabited area. More taxon-specific analyses are, therefore, needed to better understand how dispersion and connectivity of marine species are shaped through time and geographic space, and the evolutionary and ecological consequences they have on species.
In this study, we focus on Gibbula divaricata, a common species of shallow-water trochid gastropods (commonly known as top shells). This species was recently reassigned to the genus Steromphala Gray, 1847 (Affenzeller et al., 2017) due to previous descriptions of Gibbula as a paraphyletic group (Williams et al., 2010; Uribe et al., 2017). However, this reassignment lacks strong phylogenetic support indicating that more complete genetic and morphological analyses of the genus are still needed. Therefore, we have chosen to maintain the original genus name. Gibbula divaricata is distributed throughout the Mediterranean Sea (Templado, 2011) on shallow, sheltered rocky bottoms, including coastal lagoons, harbors and artificial hard structures. This species has also been cited along the coasts of the Black Sea (Anistratenko, 2005). It typically has a patchy pattern of distribution, with dense populations found in more favorable habitats but not in sandy beaches and rocky coastlines exposed to waves. Trochid gastropods produce lecithotrophic trochophore larvae that remain in the plankton for a short period (Hickman, 1992), usually less than 10 days (Underwood, 1972). Gibbula divaricata releases eggs into the water where fertilization takes place. Encapsulated larval development is fast: larvae hatch from the egg capsules 12 h post-fertilization and then remain in the plankton for a few days (Chukhchin, 1960). Although the exact duration of the planktotrophic larval stage of G. divaricata has not yet been determined, data from a closely related species, Gibbula varia (see Barco et al., 2013), indicate that development takes approximately 4 days from fertilization to complete metamorphosis and settlement (Samadi and Steiner, 2010). Given its distribution and presumptive limited dispersal capacity, G. divaricata represents an ideal organism to study genetic patterns at various geographic scales, and the potential historical and contemporary factors that have led to current biogeographic and genetic patterns.
This study focuses on central and southern populations of G. divaricata in the Adriatic basin. This basin represents a distinct marine sub-region with a priority status for marine spatial planning. This region is characterized by the existence of three cyclonic gyres (the north, the central and the southern Adriatic sub-gyres) and high differences on the bathymetry, temperature, chlorophyll-like pigment concentration, among others factors that differentiate up to six different regions (Barale et al., 2005). These factors have probably influenced the existence of biogeographical regions characterized by different categories of marine biota (Bianchi and Morri, 2000). However, some studies showed that these potential biogeographic boundaries cannot be generalized across species (Villamor et al., 2014).
The establishment of a large transboundary marine protected area has been proposed for the region (Bastari et al., 2016). The central-south sampling area was established by a pilot study within the European project COCONET (EU Seventh Framework Programme), which aims to acquire broad knowledge of marine protected areas using various approaches. Consequently, our sampling of G. divaricata was mainly confined to this predefined area. Genetic studies of other species in this area that have larval stages of varying durations have shown differing patterns of genetic differentiation, such as the fishes Mullus barbatus (Garoia et al., 2004), Diplodus sargus (Di Franco et al., 2012; Pujolar et al., 2013), Tripterygion delaisi (Koblmüller et al., 2015), Scorpaena porcus (Boissin et al., 2016) and Symphodus tinca (Carreras et al., 2017), the green crabs Carcinus maenas and C. aestuarii (Marino et al., 2011; Schiavina et al., 2014), the marble crab Pachygrapsus marmoratus (Fratini et al., 2016), the sea urchin Paracentrotus lividus (Paterno et al., 2017), and the seagrass Posidonia oceanica (Jahnke et al., 2017). These studies provide a good framework to compare and test the homogeneity or heterogeneity of dispersion and connectivity patterns within the same region.
The biology and dispersal capacity of G. divaricata, along with the distinctive physical features of the Adriatic Sea, provide a complex scenario in which to study population connectivity. Furthermore, small-scale processes are known to be important in generating patterns in benthic assemblages, supporting the idea that small-scale spatial variance contribute to regional- and broad-scale patterns of variation (Fraschetti et al., 2005).
In this study, we used microsatellite markers previously developed for G. divaricata (López-Márquez et al., 2016) to analyze central and southern Adriatic populations of the species in order to identify patterns in population structure, estimate connectivity at regional and local scales, and detect putative barriers to dispersion. Our analyses of genetic connectivity provide insight on the patterns of genetic structuring of G. divaricata in the central-south Adriatic Sea and point to contemporary factors (e.g., life history, habitat, currents, topography) potentially involved in such structuring. Genetic connectivity studies also provide valuable information for conservation and management plans in the area. For instance, coastal erosion and subsidence, among other factors such as the positive eustatism of the sea, will continue to affect the coastline for years to come (Brambati, 1992). These changes could result in the loss of suitable habitats for the species, and in the establishment of new barriers to dispersal and biogeographic boundaries along the Adriatic coastline, all of which can be highlighted by studying connectivity.
Materials and Methods
Study Area
The Adriatic Sea (Figure 1) is the most continental basin of the Mediterranean Sea (excluding the Black Sea) and is connected to the Ionian Sea through the Otranto Channel, which is 74 km wide. This latitudinally elongated sea is about 800 km in length (in its major axis, oriented from SE to NW) and has a mean width of 180 km. The Adriatic Sea is divided into three sectors: (1) the northern sector is shallow (average depth = 40 m) and has a gentle slope, (2) the middle sector (average depth = 140 m) has two depressions that reach depths of up to 260 m, and (3) the southern sector is where a wide depression is found (1,200 m deep) (Artegiani et al., 1996).
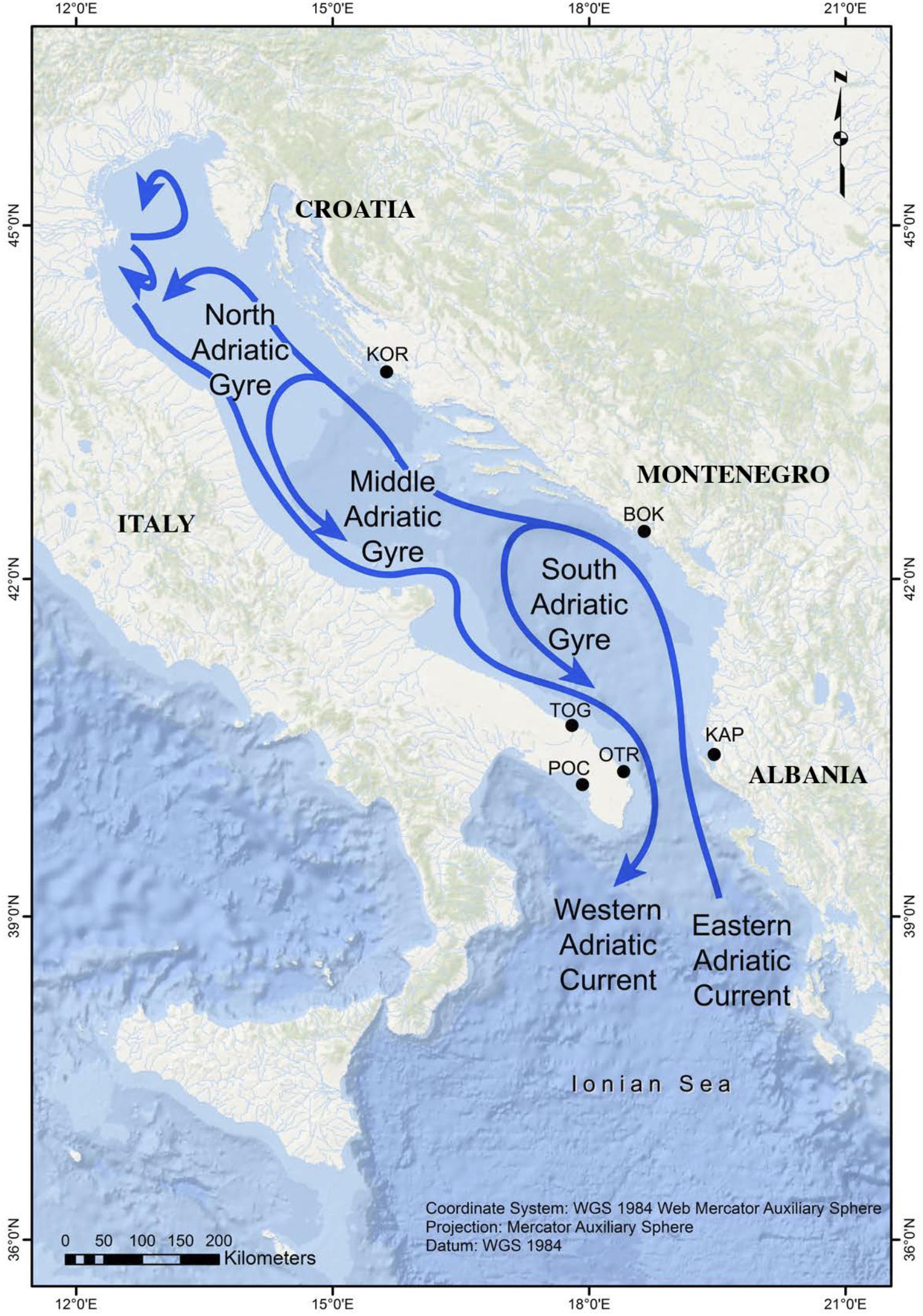
Figure 1. Map showing the sampling locations in the Adriatic Sea and its main surface currents (adapted from Melià et al., 2016).
The predefined study area is comprised of eight localities, most of which are located in the southern sector. However, no G. divaricata specimens were found on Tremiti (42° 8′ 4.23″N; 15°31′38.59″E) or Othonoi (39° 50′ 29.60″N; 19° 24′ 8.92″E) islands, which, at least in the latter case, is likely due to the presence of Gibbula rarilineata, and its occupation of suitable habitats. The six remaining localities (Table 1 and Figure 1) are separated by distances ranging from 73 to 590 km (crossing the sea). Three locations along the eastern Adriatic coast (Karaburun Peninsula in Albania = KAP, Boka Kotorska in Montenegro = BOK, and Murter in Kornati National Park in Croatia = KOR) are up to 360 km apart. The other three localities are located along the Apulian coast of Italy, with two, separated by about 90 km, in the Adriatic side (Torre Guaceto = TOG and Otranto = OTR) and the third in the Gulf of Taranto (Porto Cesareo = POC) in the northern Ionian Sea. The localities OTR and POC are separated from each other by about 120 km of coastline, circumventing the end of the Apulian Peninsula. The localities KAP and OTR occupy opposite sides of the south Adriatic Sea (73 km apart crossing the sea) across the Otranto Sill.

Table 1. Location of Gibbula divaricata samples.
The coast of Apulia extends for several hundred kilometers and is characterized by long and regular stretches of calcarenitic rock, divided by sandy beaches, with a gentle slope. It is also exposed to W–NW winds. The eastern Adriatic coast is irregular, with a series of large and small islands and a bathymetric profile comprised of rocky, steeply sloping areas, which can result in abrupt changes in habitat types.
The general surface circulation in the Adriatic Sea is cyclonic with a flow to the northwest along the eastern side and coming back to the southeast along the western side (Russo and Artegiani, 1996). Ionian waters enter on the eastern flank of the Otranto Channel and flow north-westward off Albania and Croatia by the Eastern Adriatic Current (EAC) (Poulain and Hariri, 2013). The EAC current re-circulates along the Italian coast, forming the permanent South Adriatic Gyre, which is the principal site where the Adriatic Deep Water forms. The Western Adriatic Current (WAC) is a strong coastal current that flows toward the southeast, with its surface waters eventually exiting through the western side of the Otranto Channel. Intra basin-scale circulation is dominated by seasonal cyclonic gyres of variable intensity, with waters from the EAC splitting and crossing the basin (Figure 1). The sub-gyre of the southern Adriatic tends to persist year-round (Russo and Artegiani, 1996). In conclusion, the mean cyclonic circulation favors alongshore connections and westward cross-Adriatic transport via the northern arms of the central and southern Adriatic sub-gyres (Carlson et al., 2016). This oceanographic pattern, which divides the area into three sub-regions that are connected to a larger circulation system and a series of temporal connections, greatly influences seascape connectivity within the basin (Melià et al., 2016). On the basis of large-scale connectivity patterns observed using different metrics, Dubois et al. (2016) noted that the Adriatic Sea has a persistent larval sink along the Puglia coast and the Gulf of Taranto, meaning that this area depends on other (source) populations to maintain its own populations.
Sample Collection
Between 16 and 35 specimens of G. divaricata were collected from each locality (Table 1). Live specimens (kept alive on ice) were photographed prior to fixation with ethanol. As the operculum of trochid gastropods can sometimes prevent thorough fixation of the inner tissues, shells were first cracked with a small chisel or a vice clamp. The mollusc tissues were then removed and preserved in vials with absolute ethanol, and the shells were conserved for further morphological identification. The samples were stored at 4°C prior to DNA extraction. Intact shells, if available, were deposited into the Malacological Collection at the Museo Nacional de Ciencias Naturales in Madrid (MNCN 15.05/80173).
DNA Extraction, Microsatellite Amplification, and Genotyping
Genomic DNA was extracted from a small piece of foot tissue. DNA was purified using the QIAGEN BioSprint 15 DNA Blood Kit (Qiagen), according to the manufacturer’s protocol, including a RNase treatment. DNA was quantified with the Quant-iT dsDNA HS Assay using a Qubit fluorometer, according to the manufacturer’s instructions. Aliquots of 2 ng/μl were prepared for subsequent genotyping analyses. DNA quality was also checked on 1% agarose gels. All individuals were identified to the species level by molecular and (when possible) morphological determination. Molecular identification was made by DNA barcoding following Barco et al. (2013). A 658 bp fragment at the 5′ end of cytochrome c oxidase subunit I (COI) was amplified using the primers LCO1491 (Folmer et al., 1994) and COI-H (Machordom et al., 2003). Sequences were compared with those available in GenBank using the BLAST algorithm (Altschul et al., 1997).
Microsatellites specific for G. divaricata (López-Márquez et al., 2016) were initially tested using nested-PCR conditions following the protocol developed for the mollusc Panopea abbreviata and the nemertean Malacobdella arrokeana (Ahanchédé et al., 2013; Alfaya et al., 2014, respectively). This consisted in a two steps PCR: in the first round, a forward primer with a tail at its 5′ end (PaulAn, see Acevedo et al., 2009) and a reverse primer were used. The product of this first reaction was used as template for the second, where the primers were PaulAn fluorescently 5′ end labeled with 6-FAM, NED, PET or VIC, and the reverse.
To amplify all populations, a three-primer PCR procedure (Vartia et al., 2014) was used. That included the forward primer with the tail at its 5′ end, the reverse primer, and the fluorescently 5′ end labeled tail. Alleles sizes were determined relative to the ABI GS500 LIZ standard. Based on electropherogram patterns and polymorphisms, 23 of the 26 loci described by López-Márquez et al. (2016) were selected for further analysis.
Data Analysis and Genetic Diversity
We used MICRO-CHECKER v2.2.3 (Van Oosterhout et al., 2004) to test for the presence of null alleles (with a 95% confidence interval) and scoring errors. When the estimated frequency of the putative null allele was higher than 5% and/or the heterozygosity deficit was not corrected after considering the adjusted genotypes, the involved loci were eliminated.
Allelic richness was estimated for a standardized sample of 16 individuals per population using the R package STANDARICH (Alberto, 2006). Allelic diversity (Na), observed (Ho) and expected (He) heterozygosities, Hardy–Weinberg equilibrium (HWE) within populations for each locus and linkage disequilibrium were calculated using GENEPOP v4.0 (Raymond and Rousset, 1995) and GenAlEx 6.0 (Peakal and Smouse, 2006). In the statistical analyses, p-values were adjusted with Benjamini and Hochberg’s false discovery rate (Benjamini and Hochberg, 1995; Benjamini and Yekutieli, 2001).
Inbreeding coefficient values (FIS) were estimated for each population with GenAlEx. We also tested the influence of null alleles and genotyping failures on FIS with INEST 2.2 (Chybicki and Burczyk, 2009). Inbreeding coefficients (for each population and the mean for the six populations pooled together) and the limit of the highest density posterior interval were calculated. To detect the existence of inbreeding effects in our data, a Bayesian approach using two different models (individual inbreeding models) was performed (with 50,000 burn-in cycles; 500,000 cycles overall; and 250 retained at each update). The first model, nbf, considered null alleles, inbreeding coefficients, and genotyping failures; the second model, nb, did not consider the inbreeding coefficients. To estimate the best-fit model, the deviance information criterion (DIC) was used.
Genetic relatedness was estimated by calculating pairwise relatedness in GenAlEx, considering all samples as a single population. Lynch and Ritland (1999) estimator and mean values were also assessed.
Finally, we estimated effective population sizes (Ne) for the six populations, using linkage disequilibrium, heterozygosity excess, and molecular co-ancestry methods, as implemented in NeEstimatorv2.1 (Do et al., 2014). We estimated the Ne values together with their confidence intervals (CIs) either by a parametric chi-square approximation or by jackknifing over individuals.
Population Differentiation
Data from the 23 microsatellite loci genotyped were used to estimate genetic differentiation within and among populations. We calculated Wright’s fixation indices (FST) using Weir and Cockerham’s estimators with GENETIX v.4.03 (Belkhir et al., 2004). Standardized FST values were also obtained by recoding the data matrix, assuming different alleles in each population for each locus, while maintaining their observed allelic frequencies. The standardized FST was calculated by dividing the original FST value by the corresponding recoded one. Hedrick (2005) and Meirmans and Hedrick (2011) used this approach for GST estimators and indicated that it could be used for FST indices to correct the bias of FST dependency on within-population diversity. Principal coordinates analyses (PCoA) were performed in GenAlEx with the obtained FST values. Furthermore, we used the function betas β in R with the Hierfstat package to calculate population-specific FST values (Weir and Hill, 2002).
We searched for FST outliers with the software LOSITAN (Antao et al., 2008), testing each locus for deviations from neutral expectations of the relationship between heterozygosity and FST. Positive selection was inferred from the analysis if the given p-value was greater than 0.95. A Bayesian approach using BAYESCAN 2.01 (Foll, 2012) was performed with the following parameters: burn-in = 50,000, thinning interval = 30, number of outputted iterations = 5,000, number of pilot runs = 50, and length of pilot runs = 5,000. An R function called “plot_bayescan” was also run to plot and identify outliers. To compare the probability of a p-value, a Bayes factor was used, which provides a scale of evidence in favor of a selection model versus a neutral model following Jeffreys’s (1961) scale of evidence. We used STRUCTURE 2.2.3 (Pritchard et al., 2000) to infer the number of genetically differentiated populations (K) with the highest posterior probability. An admixture model was used under both the “popinfo” and the “popinfo” plus “location prior” functions with correlated allele frequencies. Twenty replicates per K were performed for K = 1–10 to calculate the mean log probability of the data [lnP(K)]. We exceed the number of analyzed populations to explore the potential existence of individuals belonging to locations outside the study area. MCMC iterations were set to 10,000 burn-in iterations and 100,000 sampled iterations. STRUCTURE Harvester (Earl and vonHoldt, 2012) was used to calculate ΔK values using the method proposed by Evanno et al. (2005). ARLEQUIN v3.5 (Excoffier et al., 2005) was then used to hierarchically quantify the molecular variance (AMOVA, n = 10,000 permutations) in the groups inferred by STRUCTURE. Clumpak (Kopelman et al., 2015) was used to compare the different results across the 20 replicates per K. A pattern of isolation by distance, through the correlation between pairwise multilocus differentiation [FST/(1 -FST)] and geographical distances (Ln distance), was assessed using the Mantel permutation test (10,000 permutations; Mantel, 1967) implemented in GenAlEx. Two methods were used to calculate geographical distances in kilometers: (1) Euclidean (straight line) distance (across the sea) between sampled locations and (2) the coastline distance between sampled locations.
Potential gene flow barriers were identified using BARRIER v.2.2 (Manni et al., 2004). Putative barriers were computed on a Delaunay triangulation, built with GPS coordinates, using Monmonier (1973) maximum difference algorithm with the pairwise FST matrix. The robustness of the identified barriers was tested with 100 resampled bootstrap matrices created with a R function (provided by Eric Petit, UMR ECOBIO CNRS).
We also investigated patterns of migration and individual ancestries among populations using Bayesian and coalescence tools. The three approaches used estimate migration at different time scales (among other parameters): Migrate-n estimates long-term or historical migration, BayesAss estimates recent migration, and GENCLASS2 identifies first generation migrants. For the first approach, implemented in Migrate-n 3.6 (Beerli and Palczewski, 2010), we ran some initial analyses with the six populations and an unrestricted pattern of migration between them to check for convergence and the adequacy of the priors for the mutation-scaled migration rates (M) and mutation-scaled effective population sizes (theta). We then ran a full analysis under a Brownian motion microsatellite model, generating the initial theta and M values using the default FST calculations, and uniform priors of 0–50 and 0–2000 for the two parameters, respectively. We ran one long MCMC chain with 105 steps recorded and an increment of 100, excluding 104 steps as burn-in. We also checked for the adequacy of this full population and migration model by comparing its probability (marginal likelihood) with that of three other demographic scenarios: panmixia [a single population (KAP+BOK+KOR+TOG+OTR+POC)], two populations [Adriatic (KAP+BOK+KOR) vs. Apulian (TOG+OTR) + Ionian (POC) sides], and three populations [Adriatic (KAP+BOK+KOR) vs. Apulian (TOG+OTR) vs. Ionian (POC) sides]. To estimate the marginal likelihood for each of these four scenarios, we ran new analyses with the same priors as above and four MCMC chains with a static heating scheme (temperatures: 1, 1.5, 3, 106). We also ran two analyses with asymmetrical models of gene flow, the first one favoring counter-clockwise dispersal, following the sea surface currents in the Adriatic, and the second one that goes against sea surface currents. These analyses were run in the Cipres Science Gateway v3.3 (Miller et al., 2010). We then compared the marginal likelihood (Bezier approximation score) of the six models and ranked them according to their probability calculated with log Bayes factors in mtraceR (Pacioni et al., 2015).
We ran a BayesAss analysis to estimate recent migration rates (m) between populations with the following parameters: number of iterations, 10,000,000; sampling frequency, 100; length of burn-in 1,000,000; delta allele, 0.1; delta migration, 0.2; and delta F, 0.1. Probabilities of exclusion and inclusion were also calculated. If the probability of exclusion was greater than 95%, an individual was excluded from its sampling site; it was re-assigned to one of the other sampled populations if the probability of inclusion was greater than 10% (Underwood et al., 2007). An individual was assumed to have originated from an unknown population if it was excluded from one population but not re-assigned to one of the others.
Finally, a Bayesian assignment method (Rannala and Mountain, 1997), implemented in GENECLASS2 (Piry et al., 2004), was performed to identify putative first-generation migrants among populations. Then, to compute individual probabilities of assignment to each population, a Monte Carlo resampling method (simulation algorithm Paetkau et al., 2004) was performed using 10,000 simulated individuals with a type I error of 0.05.
Results
Genetic Variability
The 23 loci analyzed were polymorphic for all populations. Standardized allelic richness over loci for each population ranged from 7.59 for OTR to 9.60 for POC (Table 2). The mean value over loci and populations was 8.64. Observed and expected heterozygosities ranged from 0.582 for TOG to 0.635 for BOK and from 0.684 for KAP to 0.780 for POC, respectively (with mean values of 0.6 and 0.722, respectively) (Table 2). All FIS values were positive and high (0.144–0.253), indicating a heterozygosity deficit. When correcting the effect of null alleles on this parameter, the values decreased (avg. Fi = 0.016–0.078), and all highest posterior density intervals (HPDI = 0–0.143) included the zero value. Almost all populations showed nb as the best-fit model, thus reducing the role of inbreeding in the differences between observed and expected heterozygosities. For BOK and KOR, nfb was the best-fit model, although differences in DIC values between the two models were less than 1 (Table 2).
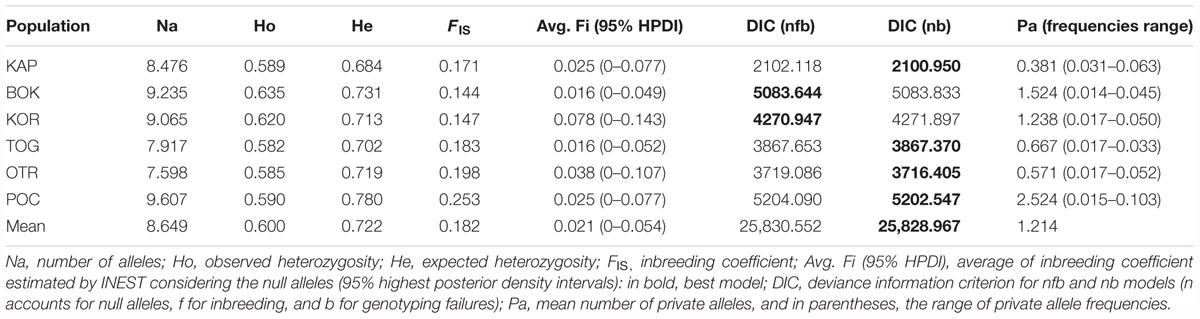
Table 2. Estimators of genetic diversity in 174 samples of Gibbula divaricata at 21 microsatellite loci.
Linkage disequilibrium among loci was not observed in any of the populations, implying that the 23 loci can be considered statistically independent. Even after Benjamini and Hochberg corrections, all localities had significant deviations from HWE for different loci, usually due to heterozygote deficits, which may be accounted for by the presence of null alleles. Two loci (Gd-L22 and Gd-L42) presented high frequencies for all studied locations and were, thus, removed from subsequent analyses. Problems associated with amplification, scoring or mutations in the regions where primers were designed may account for the presence of null alleles at these two loci. Other reasons may account for the deviation from HWE for the other loci. For example, 31 alleles were found for the locus Gd-5 in KOR, where 30 specimens were analyzed. These 31 alleles can be combined to give 496 different genotypes [n(n + 1)/2]. However, in order to interpret the resulting HWE χ2 tests accurately, the sample size and the expected number for all genotypes should be more than 50 and 5, respectively (Hedrick, 2010). The correction made by the exact test could not resolve this problem. No statistically significant differences were observed in pairwise FST and pairwise FST corrected for null alleles. Given this, null alleles were not considered hereafter, thus avoiding the artificial creation of shared (null) alleles among populations.
Despite being considered neutral markers, LOSITAN identified two outlier loci: Gd-L3 and Gd-L32 showed significantly higher FST values than neutral expectations, indicating positive selection. BAYESCAN showed four loci (Gd-L5, Gd-L16, Gd-L37, and Gd-L38) as putative outliers under negative selection. Given the lack of agreement between the two tests of neutrality, all loci were considered for downstream analyses.
Estimates of pairwise relatedness among the samples ranged from -0.079 to 0.471; the mean value was -0.003. Plotting the indices, a unimodal distribution around zero was obtained (data not shown), indicating no relatedness among most specimens.
Effective population sizes for the six populations were, in general, very large (Supplementary Table S1). Either these Ne values were ‘infinite’ or the CIs included ‘infinite,’ meaning that the populations are large enough that genetic drift does not have a significant effect on the variation of the population genetic characteristics.
Population Differentiation
The global value of FST revealed a low but significant level of genetic differentiation (FST global = 0.0246, p < 0.0001; standardized FST global = 0.0964). Pairwise FST values ranged from negative values (that should be considered as zero) for KOR vs. KAP and TOG vs. OTR to 0.0460 (0.1738 for the standardized FST) for OTR vs. KAP (Table 3). Western Adriatic populations from the Apulian Peninsula (POC, OTR, and TOG) were significantly differentiated from eastern Adriatic populations (KAP, KOR, and BOK). The Ionian locality POC showed significant values with respect to the other localities. Population specific FST β ranged from -0.0469 for POC to 0.0661 for KAP (Supplementary Table S2).
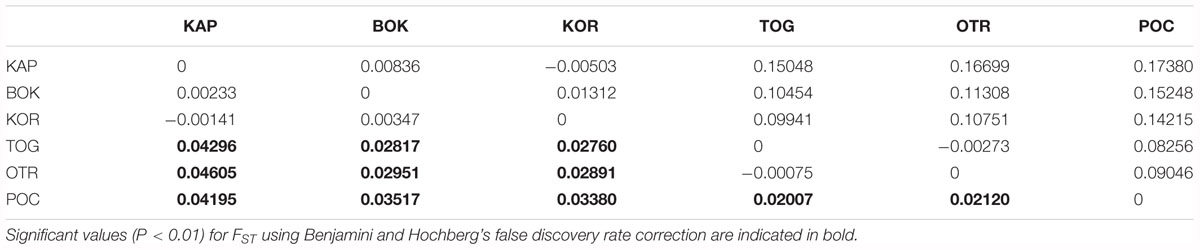
Table 3. Genetic distances shown as FST (below the diagonal) and standardized FST values (above the diagonal).
No significant association between genetic differentiation (FST) and geographic distance was revealed when using Euclidean distances between locations across the sea (Mantel test, P = 0.440, R = 0.00091). In contrast, a higher correlation was found when geographic distances were measured following the coastline between sampled locations (Mantel test, P = 0.050, R = 0.62582).
In the STRUCTURE analyses, two genetically differentiated clusters were detected under the functional parameters of both “popinfo” and “popinfo + location prior.” The highest value of likelihood was K = 2, supported by the highest ΔK values (ΔK = 17.77 and 22.28, respectively) (Supplementary Figure S1). The two clusters corresponded to populations belonging to the west (TOG, OTR, and POC) and east (KAP, BOK, and KOR) coasts of the Adriatic Sea, although most of the specimens from the western locations presented admixed ancestries (Figure 2A). Since the FST values indicated differentiation among the western populations, a second STRUCTURE analysis of this group was performed. POC was differentiated from the other western locations (TOG and OTR), a result that was supported by the highest ΔK value (ΔK = 7.80) (Figure 2B). In fact, POC has the greatest number of private alleles in almost all loci, some at frequencies around 10%.
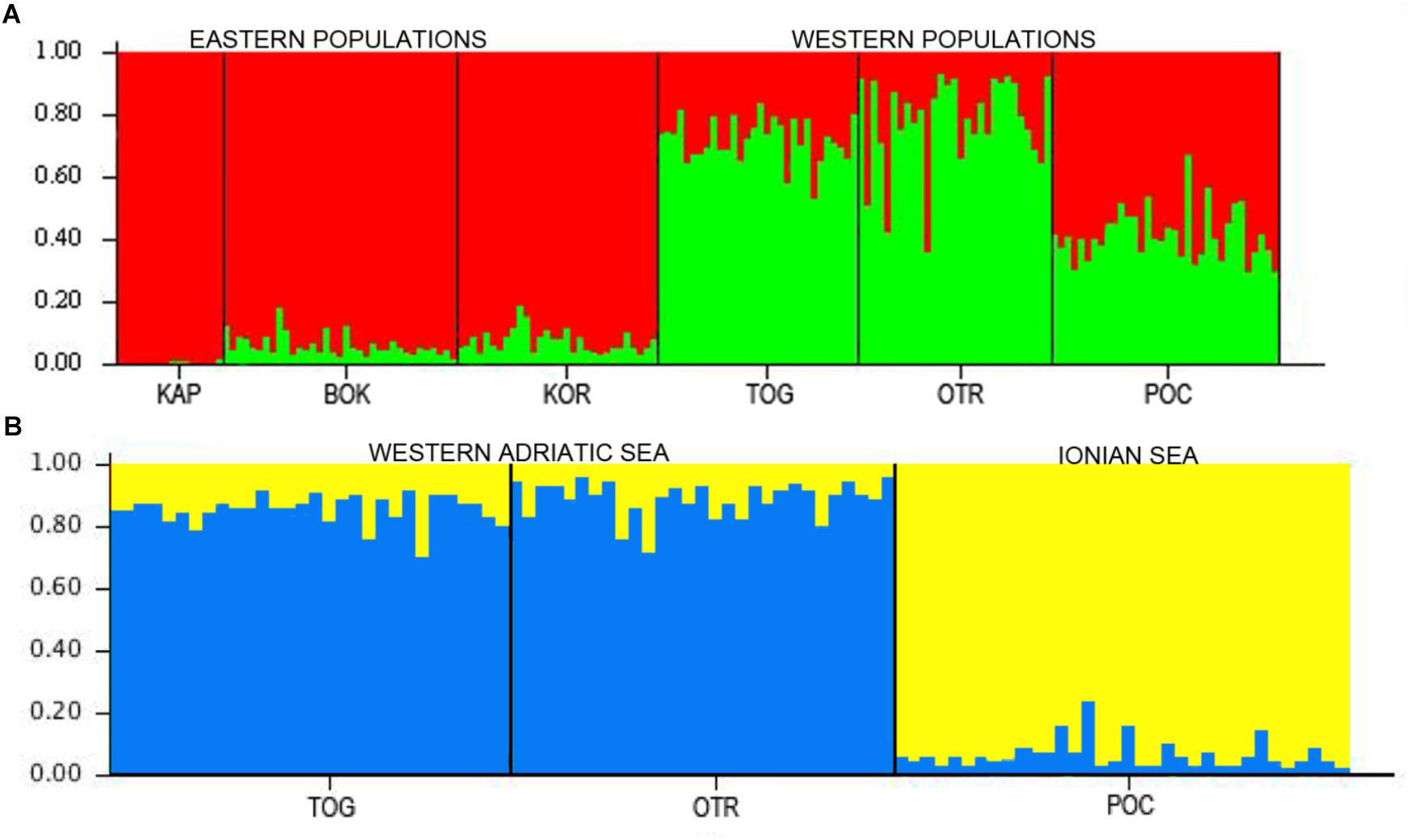
Figure 2. (A) STRUCTURE results for the six locations in the Adriatic and Ionian seas (K = 2). (B) STRUCTURE results for the three west locations in the Adriatic and Ionian seas (K = 2).
AMOVA analysis of the eastern and western groups revealed that 96.64% of the genetic variation originated within populations and only 2.31% among populations. The lowest percentage of variation (1.05%) was among populations within the two groups (Table 4).
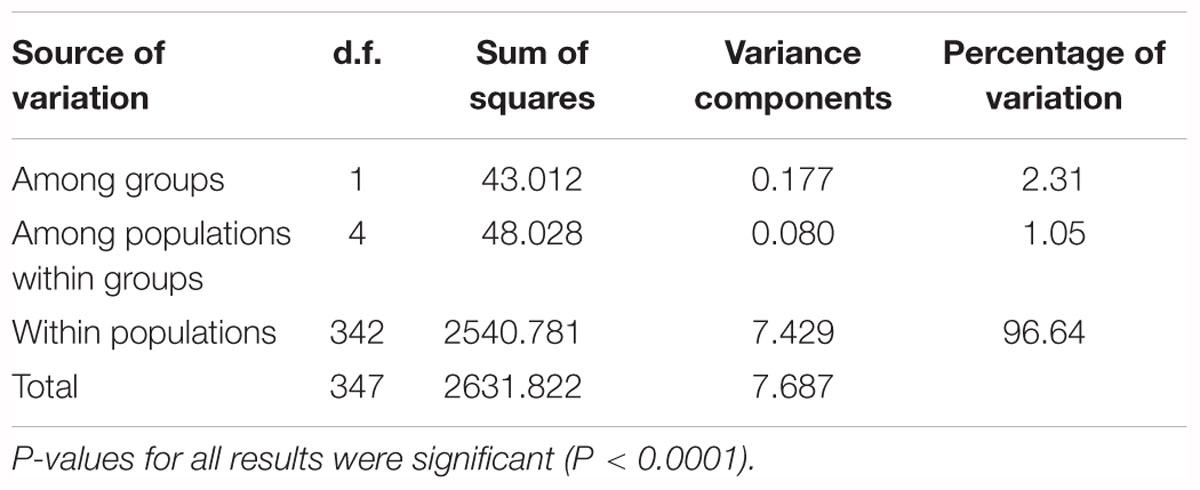
Table 4. AMOVA analyses for six populations of G. divaricata divided into two groups (east and west sides of Adriatic Sea).
The FST values among populations represented in the PCoA analysis showed that 96.81% of variation could be explained by the first two axes. The first axis of the PCoA clearly separated OTR, TOG and POC from the other Adriatic localities (Figure 3). Nevertheless, within this group, analysis of the second axis showed the well-supported division of POC from OTR and TOG.
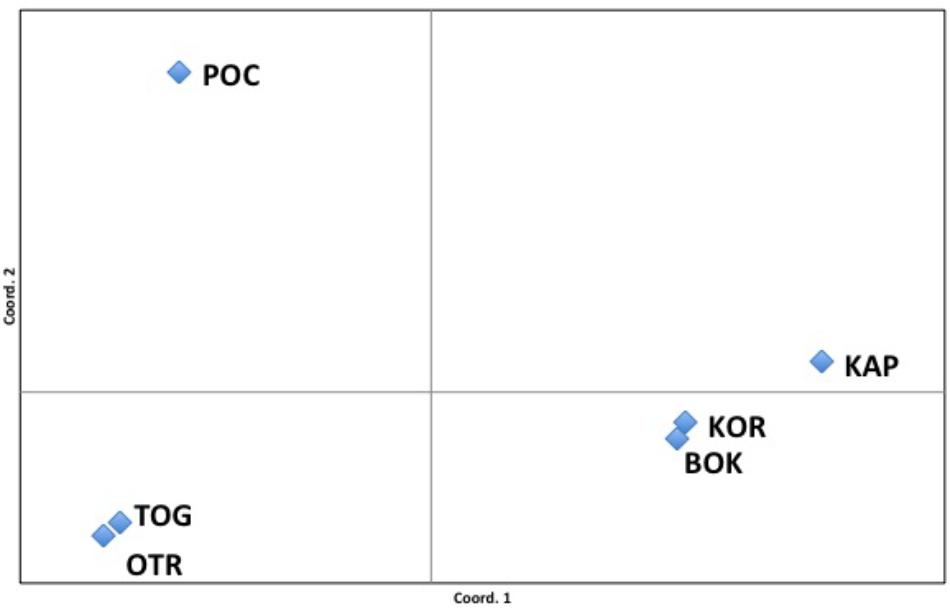
Figure 3. Principal coordinates analysis (PCoA). FST values among populations showed a variation of 96.81% explained by the two first axes.
Putative barriers of gene flow across the studied area were assessed with BARRIER. This analysis indicated a well-supported (bootstrap value = 96%) barrier between the east and the west coasts of the Adriatic Sea, dividing KAP, BOK, and KOR from TOG, OTR, and POC. The presence of a second barrier, separating POC from TOG and OTR, was also indicated but with lower support.
Migration rates among the six populations were estimated using a Bayesian coalescence approach (Migrate-n). The highest mutation-scaled migration rates (M) were generally found between populations on either the eastern or western sides of the Adriatic Sea [populations KAP, BOK, KOR or TOG, OTR, POC, respectively (Supplementary Table S3)], which seem to corroborate the results of the FST, STRUCTURE, and Mantel test analyses. Migration rates from POC to the other five populations are, however, consistently high. Dispersal from TOG and OTR to the other populations (except to each other) seems to have been historically constrained, as evidenced by the low M estimated for these two populations. However, when the marginal likelihood of the full migration model among the six populations was compared with those of the other demographic and gene flow scenarios (panmixia, two populations, three populations, asymmetric counter-clockwise gene flow, and asymmetric clockwise gene flow), the results favored panmixia as the most likely scenario (Supplementary Table S4), followed by the two asymmetric gene flow scenarios, the latter mimicking a stepping-stone model of dispersion along the coastline.
In the BayesAss analysis, the highest migration rates (m) were found between populations from either the eastern or western sides (i.e., from BOK to KAP and KOR and from TOG to OTR and POC, respectively) (Supplementary Table S5). Recent emigration from OTR and POC, however, was minimal. GENECLASS2 detected 30 individuals as potential first generation migrants, corresponding to 17.2% of the total number of individuals analyzed (Supplementary Table S6). Remarkably, in this analysis, the largest number of putative first generation migrants originated from unknown populations, followed by the western population of POC (with most emigrating to OTR).
Discussion
Connectivity among populations, and patterns of dispersal and gene flow, are primarily determined by the physical characteristics of the landscape occupied by a species and the biological life-history traits of that species. Connectivity and gene flow, in turn, shape the patterns of genetic structuring of a species, which are of profound ecological and evolutionary importance. In this study, we analyzed the genetic structure of the topshell Gibbula divaricata in the central and southern Adriatic Sea to infer its population structure and patterns of dispersal and gene flow. We found that, overall, populations seem well-connected and panmixia characterizes the species in the area. However, the presence of physical barriers to dispersal were also indicated in the area, and our results suggest that dispersion is mainly following the coastlines with help from the currents. One hypothesis is that this would eventually lead to high levels of genetic structuring and differentiation. However, this apparently is not the case in G. divaricata: we found evidence of weak though statistically significant differentiation among some of the populations, but overall panmixia is the rule. Large effective populations sizes and sporadic long-distance migration events among populations, as shown by the coalescence-based migration analyses, could explain this scenario. A large-scale study, nevertheless, would be necessary to estimate with more accuracy the patterns of gene flow, the demographic history of the species, and the precise physical environmental barriers influencing connectivity patterns in the species.
Overall, all of our analyses and tests consistently showed that connectivity was limited to some degree between the eastern and the western sites of the South Adriatic and, to a lesser extent, between Porto Cesareo in the Ionian Sea and the other two Italian localities along the Adriatic coast of the Apulian Peninsula. Taken together, our results indicate an Adriatic population (although panmictic) structured in two groups: one comprised of eastern populations and the other of western ones, although this last group showed a certain level of admixture (see Figure 2). Further analyses of the western populations revealed a third cluster, Porto Cesareo. A putative barrier between this location and the others was also found. This population showed the highest number of private alleles, some at frequencies near 10%, further supporting the presence of this third group. In any case, the barriers between eastern and western coasts of the Adriatic and between the Adriatic and Ionian coasts of Apulia are permeable as migrants were found among populations of each side (although most exchanges occurred between populations on the same coastline). Notably, the analyses of migration events indicate POC as a source population, both historically and currently. POC is the most differentiated population, and it is located at the edge of the Adriatic Sea. Therefore, the assigned migrants coming from POC (according to GENECLASS2) could be those considered as ‘unknown’ in the BayesAss analysis.
Genetic studies of the bentho-pelagic red mullet (Mullus barbatus) (Garoia et al., 2004) and the benthic black scorpionfish (Scorpaena porcus) (Boissin et al., 2016) have also revealed differences between the eastern and the western parts of the Adriatic basin. However, G. divaricata and S. porcus show differences in connectivity along the eastern Adriatic coast. Although we found no significant genetic differentiation of G. divaricata among our three sampled eastern populations, Boissin et al. (2016) showed strong differences among eastern populations of S. porcus, particularly between two samples from Croatia and Albania, highlighting how connectivity patterns cannot be generalized among species. A single biological trait (e.g., characteristics of the larval phase, self-recruitment or larval duration) can determine the capacity of larval exchange among populations and, hence, the extent and direction of dispersal (Melià et al., 2016). Likewise, synchronization between spawning and hydrodynamic conditions may also be a factor determining the extent of larval transport (Koeck et al., 2015).
We found no relationship between genetic and geographic distances when using Euclidean distances between locations; however, an association was found when geographic distances were measured following the coastline distances between sampled locations. Therefore, factors such as the current system, coastline layout, small-scale local hydrodynamic processes or favorable habitat availability are likely to have greater influence on the degree of connectivity between different local populations than distance itself.
Our data indicate that trochophore larvae of G. divaricata, although thought to have limited dispersal abilities, can eventually reach and settle in distant populations. The precise timing of larval development in G. divaricata is still unknown; however, other species of the genus take no longer than 10 days to reach complete metamorphosis (Underwood, 1972). Lagrangian models that simulate drifter trajectories in the Adriatic Sea support sporadic and unexpected migrations, which may account for some of our connectivity results as it only takes 5 days (consistent with the presumed duration of the larval stage of G. divaricata) for particles to drift between TOG and OTR or POC (Carlson et al., 2016). Migration between these sites and others (e.g., KOR and KAP) is highly unlikely following these models as it can take more than 40 days for particles to drift from TOG to KOR (Poulain, 2001; Bray et al., 2017), which is much longer than the pelagic larval period assumed for G. divaricata.
Migration from POC to the other populations and between populations on either side of the Adriatic Sea, however, were not consistent with predominant surface currents/biophysical models (Figure 1), although local gyres might explain these results. In a recent study of the connectivity patterns of a Mediterranean wrasse, Symphodus ocellatus, which also has a short larval phase of about 10 days (Macpherson and Raventos, 2006), and the biophysical models in the Adriatic, Melià et al. (2016) suggested the coexistence of a high local retention potential of propagules with very occasional intense movement across the sea basin. On the other hand, according to these same authors, communities were significantly more similar along a gradient of connectivity by ocean currents than with increasing distance in the region encompassing the southern Adriatic and Ionian seas. Likewise, in our analysis, more differences were found between physically closer populations (KAP and OTR) than between remote populations linked by these ocean currents (KAP and KOR).
Genetic analyses of a variety of species in the Adriatic Sea (Garoia et al., 2004; Di Franco et al., 2012; Pujolar et al., 2013; Schiavina et al., 2014; Boissin et al., 2016; Carreras et al., 2017) have indicated that realized dispersal (larvae that actually travel from one locality to another) is much less pronounced than potential dispersal (probability of larval transport from a source to destination location, as quantified by Lagrangian particle simulations) as evidenced by some degree of differentiation among sites for some species. Panmictic species may represent exceptions. For example, the marbled crab Pachygrapsus marmoratus, given its ubiquity and long pelagic larval duration, was shown to have high connectivity within the Adriatic and even more broadly throughout the Mediterranean (Fratini et al., 2016). The sea urchin Paracentrotus lividus showed a similar pattern, but in this case, connectivity was limited to the Adriatic and Ionian seas (Paterno et al., 2017). The shore crab Carcinus aestuarii (Schiavina et al., 2014), which is a highly dispersive species (in lagoons and estuaries), may also have limited connectivity in these seas as females release larvae in coastal waters after a brief migration toward the open sea (Mori et al., 1990). The specificity of this species to isolated habitats, along with their great dispersal capacity, implies that oceanographic conditions play an important role in larval dispersion. In such cases, the translocation of pre-competent larvae from nearshore to offshore and the shoreward return (by onshore advection) of competent larvae should be favored, replenishing the larval supply to this species’ specific shore habitats. In the case of G. divaricata, and in spite of its assumed low dispersal capacity, its specific habitat requirements (shallow sheltered rocky coast, including harbor areas), together with the coastal currents, likely favors a degree of dispersal along the coastline. The differentiation observed in the Ionian population may be due to, among other factors, the absence of sheltered habitats at intermediate points between OTR and POC. The southern area of the Apulian Peninsula consists of a high rocky coast exposed to intense wave action (Damiani et al., 1988) that may restrict connectivity in the species. Therefore, in species with restricted or specific environmental requirements, habitat patchiness may greatly influence population connectivity by affecting local larval production and recruitment success based on the proportion of favorable habitats (Pinsky et al., 2012; Anadón et al., 2013).
However, unlike G. divaricata, species such as C. aestuarii and S. porcus showed no significant genetic differences between Apulian localities in the Adriatic (Torre Guaceto and Otranto) and Ionian (Porto Cesareo) seas (Schiavina et al., 2014; Boissin et al., 2016, respectively). Likewise, the biophysical models that showed infrequent and weak connections linking the eastern and western sides of the Adriatic also showed moderately intense fluxes connecting the southernmost areas of Apulia with the eastern part of the Gulf of Taranto (Melià et al., 2016). These findings agree with other data obtained from surface drifter deployments and Lagrangian simulations performed in the same area to describe larval dispersal of the white seabream Diplodus sargus (Pujolar et al., 2013). Some authors have suggested a phylogeographical discontinuity of the Adriatic Sea given the genetic distinctiveness of the populations of several species inhabiting this basin (Patarnello et al., 2007; Maltagliati et al., 2010). Nevertheless, these genetic discontinuities (inferred from mitochondrial markers) seem to be more related to the effects of historical events, such as periods of very low sea level. Present-day coastal marine currents connecting the Adriatic and the Ionian seas favor larval exchange and contemporary gene flow, although these processes may be partially restricted in species with a supposed low dispersal capacity, such as G. divaricata.
Genetic homogeneity may be explained by a continuous and significant coastal marine current that connects different areas, thus promoting larval exchange. Surface drifter tracks and modeled particle trajectories indicate that the Adriatic circulation patterns sustain physical corridors as connection pathways within a large-scale functional cell of connectivity in the southern Adriatic (Boero et al., 2016). Likewise, several persistent larval sinks have been noted along the southern Italian shore (Bray et al., 2017). Though these studies are informative about Adriatic alongshore coastal currents and cyclonic gyres, they may not reflect actual larval pathways, which in hydrodynamic connectivity studies, are based on drifter trajectories. Rather, larval behaviors range from passively drifting to selectively using currents for dispersal. Marine larvae display a large repertoire of behaviors that can greatly affect dispersal patterns at different spatial scales, such as the ability to migrate vertically, swim horizontally, or undergo ontogenetic changes in behavior or those related to having distinctive sensory capabilities. These behaviors allow larvae to change their dispersal pattern by taking advantage of different oceanographic conditions (Pineda et al., 2007). Habitat selection (at small spatial scales) is determined by the differential responses of larvae to proximate environmental stimuli (Kingsford et al., 2002). Hence, the interaction between larval behavior and physical processes can influence dispersal patterns to either increase advection from the native population or enhance self-recruitment (Sponaugle et al., 2002).
In order to determine if connectivity is mediated through a continuous stretch of suitable habitats along the coastline, genetic analyses need to be combined with habitat mapping. Species with supposed low dispersal abilities, such as G. divaricata, may use patches of its specific habitat as stepping stones, thus allowing them to disperse over a wider range. As a result of stepping stone dispersion, ubiquitous shore species that have access to relatively continuous habitats would show high genetic connectivity, while species inhabiting patchy dispersed habitats, or isolated habitats such as estuaries and lagoons, would show low to no connectivity. This connectivity, in turn, can be modulated by the durations and behaviors of the early life stages of different species.
Overall, our findings indicate weak differentiation among the studied populations belonging to the western and the eastern coasts of the central-south Adriatic. Although significant, the extent of differentiation and the strength of barriers were low, and some level of migration was evident, which taken together suggest very recent isolation or a weak restriction to gene flow. Multiple physical and biological factors may be influencing this population structure. Therefore, much research is still needed to determine the extent to which these factors affect connectivity in this species, not only in the Adriatic region but throughout its distribution. In addition, knowledge of the life cycle phases of G. divaricata, and their ecological interactions, will shed light on the actual dispersal ability of the species and enlighten ongoing phylogeographic studies.
Connectivity is increasingly recognized as a key conservation objective because of its importance for species replenishment (Saenz-Agudelo et al., 2011). Changes in fertilization success, larval supply, and recruitment play a major role in connectivity and, therefore, in both the future population dynamics and the long-term viability of a species. Regrettably, habitat fragmentation and loss due to human activities are widely considered one of the strongest drivers of change at all levels of biodiversity, as well as in the structuring and functioning of marine coastal ecosystems (Templado, 2014). Populations become more isolated (due to loss of connectivity) and decrease in size and abundance due to habitat fragmentation and loss. Therefore, conservation measures should aim to guarantee the persistence and continuity of natural habitats. The establishment of a well-planned network of marine protected areas will also help to maintain connectivity among populations.
Author Contributions
VL-M conceived the research, conducted field work and laboratory and data analyses, and wrote the manuscript. JT and AM conceived the research, were responsible for resource acquisition, conducted field work and wrote and provided comments on the manuscript. DB collaborated in data analyses and reviewed the manuscript. LZ conducted field work, collaborated in data analyses, and provided comments on the manuscript. IM, EB, and DM conducted field work and reviewed the manuscript. This research is part of VL-M Ph.D. thesis, co-directed by JT and AM.
Funding
This research was funded by the European project CoCoNET “Towards COast to COast NETworks of marine protected areas (from the shore to the high and deep sea), coupled with sea-based wind energy potential” from the VII FP of the European Commission (grant agreement n° 287844) and the Spanish Ministry of Economy and Competitiveness (grant reference: CTM2014-57949-R).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are grateful to many people who generously helped to collect the samples: Toni Terlizzi and Giorgio Aglieri from the University of Salento, Chiara Papetti from the University of Padova, Vesna Mačić from the Institute of Marine Biology, University of Montenegro, and Stefania Bortoluzzi, Anita Zane, and Nicola Zane. Thanks to the Photography and Computational Biogeography laboratories of the MNCN and the Malacological collection curator, Rafael Araujo, and Javier de Andrés for their kind help. We are also thankful to Melinda Modrell for the conscientious revision of the language.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2019.00177/full#supplementary-material
References
Acevedo, I., Bloor, P., Cabezas, P., Toledo, C., Calvo, M., and Machordom, A. (2009). Development of tetranucleotide microsatellite markers for the cushion starfish, Asterina gibbosa, and cross-species amplification. Mol. Ecol. Resour. 9, 274–277. doi: 10.1111/j.1755-0998.2008.02267.x
Affenzeller, S., Haar, N., and Steiner, G. (2017). Revision of the genus complex Gibbula: an integrative approach to delineating the Eastern Mediterranean genera Gibbula Risso, 1826, Steromphala Gray, 1847, and Phorcus Risso, 1826 using DNA-barcoding and geometric morphometrics (Vetigastropoda, Trochoidea). Org. Divers. Evol. 17, 789–812. doi: 10.1007/s13127-017-0343-5
Ahanchédé, A., Alfaya, J. E. F., Andersen, L. W., Azam, D., Bautista, M. A. M., Besnard, A. L., et al. (2013). Permanent genetic resources added to molecular ecology resources database 1 August 2012–30 September 2012. Mol. Ecol. Resour. 13, 158–159. doi: 10.1111/1755-0998.12035
Alberto, F. (2006). StandArich v1.0: an R Package to Estimate Population Allelic Richness Using Standardized Sample Size [Internet]. Available at: http://www.ccmar.ualg.pt/maree/software.php?soft=sarich
Alfaya, J. E. F., López-Márquez, V., García-Jiménez, R., Fernández-Álvarez, F., Bigatti, G., and Machordom, A. (2014). Development and characterization of microsatellite loci in the entocommensal Malacobdella arrokeana (Nemertea: Bdellonemertea), from Patagonia (Argentina) and cross-amplification in 34 nemertean species. Conserv. Genet. Resour. 6, 147–150. doi: 10.1007/s12686-013-0029-z
Altschul, S. F., Madden, T. L., Schäffer, A. A., Zhang, J., Zhang, Z., Miller, W., et al. (1997). Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25, 3389–3402. doi: 10.1093/nar/25.17.3389
Anadón, J. D., Mancha-Cisneros, M. M., Best, B. D., and Gerber, L. R. (2013). Habitat-specific larval dispersal and marine connectivity: implications for spatial conservation planning. Ecosphere 4:82. doi: 10.1890/ES13-00119.1
Anistratenko, V. V. (2005). Lectotypes for Tricolia pullus, Gibbula divaricata and Theodoxus fluviatilis (Mollusca, Gastropoda) Revisited. Vestn. Zool. 39, 3–10.
Antao, T., Lopes, A., Lopes, R. J., Beja-Pereira, A., and Luikart, G. (2008). LOSITAN: a workbench to detect molecular adaptation based on a Fst - outlier method. BMC Bioinformatics 9:323. doi: 10.1186/1471-2105-9-323
Artegiani, A., Bregant, D., Paschini, E., Pinardi, N., Raicich, F., and Russo, A. (1996). The Adriatic Sea general circulation. Part I: air-sea interactions and water mass structure. J. Phys. Oceanogr. 27, 1492–1514. doi: 10.1175/1520-0485(1997)027<1492:TASGCP>2.0.CO;2
Barale, V., Schiller, C., Tacchi, R., and Marechal, C. (2005). Trends and interactions of physical and bio-geo-chemical features in the Adriatic Sea as derived from satellite observations. Sci. Total Environ. 353, 68–81. doi: 10.1016/j.scitotenv.2005.09.060
Barco, A., Evans, J., Schembri, P. J., Taviani, M., and Oliverio, M. (2013). Testing the applicability of DNA barcoding for Mediterranean species of top-shells (Gastropoda, Trochidae, Gibbula s.l.). Mar. Biol. Res. 8, 785–793. doi: 10.1080/17451000.2013.765585
Bastari, A., Micheli, F., Ferretti, F., Pusceddu, A., and Cerrano, C. (2016). Large marine protected areas (LMPAs) in the Mediterranean Sea: the opportunity of the Adriatic Sea. Mar. Policiy. 68, 165–177. doi: 10.1016/j.marpol.2016.03.010
Beerli, P., and Palczewski, M. (2010). Unified framework to evaluate panmixia and migration direction among multiple sampling locations. Genetics 185, 313–326. doi: 10.1534/genetics.109.112532
Belkhir, K., Borsa, P., Chikhi, L., Raufaste, N., and Bonhomme, F. (2004). GENETIX 4.05, Logiciel Sous Windows TM Pour la Génétique des Populations. Laboratoire Génome Populations Interactions CNRS UMR5000. Available at: https://www.scienceopen.com/document?vid=9a3e2cf3-2971-405c-8297-258227c3cb30
Benjamini, Y., and Hochberg, Y. (1995). Controlling the false discovery rate: a practical and powerful approach to multiple testing. J. R. Stat. Soc. 57, 289–300. doi: 10.1111/j.2517-6161.1995.tb02031.x
Benjamini, Y., and Yekutieli, D. (2001). The control of the false discovery rate in multiple testing under dependency. Ann. Stat. 29, 1165–1188. doi: 10.1186/1471-2105-9-114
Bianchi, C. N., and Morri, C. (2000). Marine biodiversity of the mediterranean sea: situation, problems and prospects for future research. Mar. Pollut. Bull. 40, 367–376. doi: 10.1016/S0025-326X(00)00027-8
Boero, F., Foglini, F., Fraschetti, S., Goriup, P., Macpherson, E., Planes, S., et al. (2016). CoCoNet: towards coast to coast networks of marine protected areas (from the shore to the high and deep sea), coupled with sea-based wind energy potential. Scires It 6, 1–95.
Boissin, E., Micu, D., Janczyszyn-Le Goff, M., Neglia, V., Bat, L., Todorova, V., et al. (2016). Contemporary genetic structure and postglacial demographic history of the black scorpionfish, Scorpaena porcus, in the Mediterranean and the Black Seas. Mol. Ecol. 25, 2195–2209. doi: 10.1111/mec.13616
Botsford, L., Hastings, A., and Gaines, S. (2001). Dependence of sustainability on the configuration of marine reserves and larval dispersal distance. Ecol. Lett. 4, 144–150. doi: 10.1046/j.1461-0248.2001.00208.x
Brambati, A. (1992). “Origin and evolution of the Adriatic Sea,” in Marine Eutrophication and Population Dynamics, eds G. Colombo (Fredensborg: Olsen & Olsen), 327–346.
Bray, L., Kassis, D., and Hall-Spencer, J. M. (2017). Assessing larval connectivity for marine spatial planning in the Adriatic. Mar. Environ. Res. 125, 73–81. doi: 10.1016/j.marenvres.2017.01.006
Carlson, D. F., Griffa, A., Zambianchi, E., Suaria, G., Corgnati, L., Magaldi, M. G., et al. (2016). Observed and modeled surface Lagrangian transport between coastal regions in the Adriatic Sea with implications for marine protected areas. Cont. Shelf Res. 118, 23–48. doi: 10.1016/j.csr.2016.02.012
Carreras, C., Ordóñez, V., Zane, L., Kruschel, C., Nasto, I., Macpherson, E., et al. (2017). Population genomics of an endemic Mediterranean fish: differentiation by fine scale dispersal and adaptation. Sci. Rep. 7:43417. doi: 10.1038/srep43417
Chukhchin, V. D. (1960). “Pelagic larvae of Gastropod mollusks of the Black Sea,” in Akademiia Nauk SSSR, Trudy Sevastopol’ skoi Biologicheskoi Stantsii, No. 21, Vol. XIII, eds P. Chanley and R. Robertson, trans. E. C. Wells (Gloucester Point: Virginia Institute of Marine Science Gloucester Point), 92–113.
Chybicki, I. J., and Burczyk, J. (2009). Simultaneous estimation of null alleles and inbreeding coefficients. J. Hered. 100, 106–113. doi: 10.1093/jhered/esn088
Damiani, V., Bianchi, C. N., Ferretti, O., Bedulli, D., Morri, C., Viel, M., et al. (1988). Risultati di una ricerca ecologica sul sistema marino costiero pugliese. Thalassia Salentina 18, 153–169.
Di Franco, A., Coppini, G., Pujolar, J. M., De Leo, G. A., Gatto, M., Lyubartsev, V., et al. (2012). Assessing dispersal patterns of fish propagules from an effective mediterranean marine protected area. PLoS One 7:e52108. doi: 10.1371/journal.pone.0052108
Do, C., Waples, R. S., Peel, D., Macbeth, G. M., Tillett, B. J., and Ovenden, J. R. (2014). NeEstimator V2: re-implementation of software for the estimation of contemporary effective population size (Ne) from genetic data. Mol. Ecol. Res. 14, 209–214. doi: 10.1111/1755-0998.12157
Dubois, M., Rossi, V., Ser-Giacomi, E., Arnaud-Haond, S., López, C., and Hernández-García, E. (2016). Linking basin-scale connectivity, oceanography and population dynamics for the conservation and management of marine ecosystems. Glob. Ecol. Biogeogr. 25, 503–515. doi: 10.1111/geb.12431
Earl, D. A., and vonHoldt, B. M. (2012). STRUCTURE HARVESTER: a website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 4, 359–361. doi: 10.1007/s12686-011-9548-7
Evanno, G., Regnaut, S., and Goudet, J. (2005). Detecting the number of clusters of individuals using the software structure: a simulation study. Mol. Ecol. 14, 2611–2620. doi: 10.1111/j.1365-294X.2005.02553.x
Excoffier, L., Laval, G., and Schneider, S. (2005). ARLEQUIN (version 3.0): an integrated software package for population genetics data analysis. Evol. Bioinform. 1, 47–50. doi: 10.1177/117693430500100003
Folmer, O., Black, M., Hoeh, W., Lutz, R., and Vrijenhoek, R. (1994). DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 3, 294–299.
Fraschetti, S., Terlizzi, A., and Benedetti-Cecchi, L. (2005). Patterns of distribution of marine assemblages from rocky shores: evidence of relevant scales of variation. Mar. Ecol. Prog. Ser. 296, 13–29. doi: 10.3354/meps296013
Fratini, S., Ragionieri, L., Deli, T., Harrer, A., Marino, I. A. M., Cannicci, S., et al. (2016). Unravelling population genetic structure with mitochondrial DNA in a notional panmictic coastal crab species: sample size makes the difference. BMC Evol. Biol. 16:150. doi: 10.1186/s12862-016-0720-2
Garoia, F., Guarniero, I., Piccinetti, C., and Tinti, F. (2004). First microsatellite loci of red mullet (Mullus barbatus) and their application to genetic structure analysis of Adriatic shared stock. Mar. Biotechnol. 6, 446–452. doi: 10.1007/s10126-004-3045-x
Hedrick, P. W. (2005). A standardized genetic differentiation measure. Evolution 59, 1633–1638. doi: 10.1111/j.0014-3820.2005.tb01814.x
Hedrick, P. W. (2010). Genetics of Populations, 4th Edn. Burlington, MA: Jones and Bartlett Publishers.
Jahnke, M., Casagrandi, R., Melià, P., Schiavina, M., Shultz, S. T., Zane, L., et al. (2017). Potential and realized connectivity of the seagrass Posidonia oceanica and their implication for conservation. Divers. Distrib. 23, 1423–1434. doi: 10.1111/ddi.12633
Jeffreys’s, H. (1961). Theory of Probability (Oxford Classic Texts in the Physical Sciences), 3rd Edn. Oxford: Oxford University Press.
Kingsford, M. J., Leis, J. M., Shanks, A., Lindeman, K. C., Morgan, S. G., and Pineda, J. (2002). Sensory environments, larval abilities and local self-recruitment. Bull. Mar. Sci. 70, 309–340.
Koblmüller, S., Steinwender, B., Weiß, S., and Sefc, K. M. (2015). Gene flow, population growth and a novel substitution rate estimate in a subtidal rock specialist, the black-faced blenny Tripterygion delaisi (Perciformes, Blennioidei, Tripterygiidae) from the Adriatic Sea. J. Zool. Syst. Evol. Res. 53, 291–299. doi: 10.1111/jzs.12110
Koeck, B., Gerigny, O., Durieux, E. D. H., Coudray, S., Garsi, L.-H., Bisgambiglia, P.-A., et al. (2015). Connectivity patterns of coastal fishes following different dispersal scenarios across a transboundary marine protected area (Bonifacio strait, NW Mediterranean). Estuar. Coast. Shelf Sci. 154, 234–247. doi: 10.1016/j.ecss.2015.01.010
Kopelman, N. M., Mayzel, J., Jakobsson, M., Rosenberg, N. A., and Mayrose, I. (2015). Clumpak: a program for identifying clustering modes and packaging population structure inferences across K. Mol. Ecol. Resour. 15, 1179–1191. doi: 10.1111/1755-0998.12387
López-Márquez, V., García-Jiménez, R., Templado, J., and Machordom, A. (2016). Development and characterization of 26 novel microsatellite loci for the trochid gastropod Gibbula divaricata (Linnaeus, 1758), using Illumina MiSeq next generation sequencing technology. PeerJ 4:e1789. doi: 10.7717/peerj.1789
Lynch, M., and Ritland, K. (1999). Estimation of pairwise relatedness with molecular markers. Genetics 152, 1753–1766.
Machordom, A., Araujo, R., Erpenbeck, D., and Ramos, M. A. (2003). Phylogeograhy and conservation genetics of endangered European Margaritiferidae (Bivalvia: Unionoidea). Biol. J. Linn. Soc. 78, 235–252. doi: 10.1046/j.1095-8312.2003.00158.x
Macpherson, E., and Raventos, N. (2006). Relationship between pelagic larval duration and geographic distribution of Mediterranean littoral fishes. Mar. Ecol. Prog. Ser. 327, 257–265. doi: 10.3354/meps327257
Maltagliati, F., Di Giuseppe, G., Barbieri, M., Castelli, A., and Dini, F. (2010). Phygeography and genetic structure of the edible sea urchin Paracentrotus lividus (Echinodermata: Echinoidea) inferred from the mitochondrial cytochrome b gene. Biol. J. Linn. Soc. 100, 910–923. doi: 10.1111/j.1095-8312.2010.01482.x
Manni, F., Guerar, E., and Heyer, E. (2004). Geographic patterns of (genetic, morphologic, linguistic) variation: how barriers can be detected by using Monmonier’s algorithm. Hum. Biol. 76, 173–190. doi: 10.1353/hub.2004.0034
Mantel, N. (1967). The detection of disease clustering and a generalized regression approach. Cancer Res. 27, 209–220.
Marino, I. A. M., Pujolar, J. M., and Zane, L. (2011). Reconciling deep calibration and demographic history: bayesian inference of post glacial colonization patterns in Carcinus aestuarii (Nardo, 1847) and C. maenas (Linnaeus, 1758). PLoS One 6:e28567. doi: 10.1371/journal.pone.0028567
Meirmans, P. G., and Hedrick, P. W. (2011). Assessing population structure Fst and related measures. Mol. Ecol. Resour. 11, 5–18. doi: 10.1111/j.1755-0998.2010.02927.x
Melià, P., Schiavina, M., Rossetto, M., Gatto, M., Fraschetti, S., and Casagrandi, R. (2016). Looking for hotspots of marine metacommunity connectivity: a methodological Framework. Sci. Rep. 6:23705. doi: 10.1038/srep23705
Miller, M. A., Pfeiffer, W., and Schartz, T. (2010). Creating the CIPRES science gateway for inference of large phylogenetic trees. Gateway Comp. Environ. Workshop 2010, 1–8. doi: 10.1109/GCE.2010.5676129
Monmonier, M. S. (1973). Maximum-difference barriers: an alternative numerical regionalization method. Geogr. Anal. 5, 245–261. doi: 10.1111/j.1538-4632.1973.tb01011.x
Mori, M., Manconi, R., and Fanciulli, G. (1990). Notes on the reproductive biology of Carcinus aestuarii Nardo (Crustacea, Decapoda) from the lagoon of San Teodoro (Island of Sardinia, Italy). Riv. Idrobiol. 29, 763–774.
Pacioni, C., Hunt, H., Allentoft, M. E., Vaughan, T. G., Wayne, A. F., Baynes, A., et al. (2015). Genetic diversity loss in a biodiversity hotspot: ancient DNA quantifies genetic decline and former connectivity in a critically endangered marsupial. Mol. Ecol. 24, 5813–5828. doi: 10.1111/mec.13430
Paetkau, D., Slade, R., Burden, M., and Estoup, A. (2004). Direct, real-time estimation of migration rate using assignment methods: a simulation-based exploration of accuracy and power. Mol. Ecol. 13, 55–65. doi: 10.1046/j.1365-294X.2004.02008.x
Patarnello, T., Volckaert, F. A., and Castilho, R. (2007). Pillars of hercules: is the atlantic-mediterranean transition a phylogeographical break? Mol. Ecol. 16, 4426–4444. doi: 10.1111/j.1365-294X.2007.03477.x
Paterno, M., Schiavina, M., Aglieri, G., Ben Souissi, J., Boscari, E., Casagrandi, R., et al. (2017). Population genomics meet Lagrangian simulations: oceanographic patterns and long larval duration ensure connectivity among Paracentrotus lividus populations in the Adriatic and Ionian seas. Ecol. Evol. 7, 2463–2479. doi: 10.1002/ece3.2844
Peakal, R., and Smouse, P. E. (2006). GENEALEX 6: genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 6, 288–295. doi: 10.1111/j.1471-8286.2005.01155.x
Pineda, J., Hare, J. A., and Sponaugle, S. (2007). Larval transport and dispersal in the coastal ocean and consequences for population connectivity. Oceanography 20, 22–39. doi: 10.5670/oceanog.2007.27
Pinsky, M. L., Palumbi, S. R., Andréfouët, S., and Purkis, S. (2012). Open and closed seascapes: where does habitat patchiness create populations with high fractions of self-recruitment? Ecol. Appl. 22, 1257–1267. doi: 10.1890/11-1240.1
Piry, S., Alapetite, A., Cornuet, J. M., Paetkau, D., Baudouin, L., and Estoup, A. (2004). GENECLASS2: a software for genetic assignment and first-generation migrant detection. J. Hered. 95, 536–539. doi: 10.1093/jhered/esh074
Poulain, P. M. (2001). Adriatic Sea surface circulation as derived from drifter data between 1990 and 1999. J. Mar. Syst. 29, 3–32. doi: 10.1016/S0924-7963(01)00007-0
Poulain, P. M., and Hariri, S. (2013). Transit and residence times in the surface Adriatic Sea as derived from drifter data and Lagrangian numerical simulations. Ocean Sci. 10, 197–217.
Pritchard, J. K., Stephens, M., and Donnelly, P. (2000). Inference of population structure using multilocus genotype data. Genetics 155, 945–959.
Pujolar, J. M., Schiavina, M., Di Franco, A., Melià, P., Guidetti, P., Gatto, M., et al. (2013). Understanding the effectiveness of marine protected areas using genetic connectivity patterns and Lagrangian simulations. Divers. Distrib. 19, 1531–1542. doi: 10.1111/ddi.12114
Rannala, B., and Mountain, J. L. (1997). Detecting immigration by using multilocus genotypes. Proc. Natl. Acad. Sci. U.S.A. 94, 9197–9201. doi: 10.1073/pnas.94.17.9197
Raymond, M., and Rousset, F. (1995). GENEPOP (version 1.2): population genetics software for exact tests and ecumenicism. J. Hered. 86, 248–249. doi: 10.1093/oxfordjournals.jhered.a111573
Russo, A., and Artegiani, A. (1996). Adriatic Sea hydrography. Sci. Mar. 60(Suppl. 2), 33–43. doi: 10.1007/s10661-008-0350-2
Saenz-Agudelo, P., Jones, G. P., Thorrold, S. R., and Planes, S. (2011). Connectivity dominates larval replenishment in a coastal reef fish metapopulation. Proc. R. Soc. B. 278, 2954–2961. doi: 10.1098/rspb.2010.2780
Samadi, L., and Steiner, G. (2010). Expression of Hox genes during the larval development of the snail, Gibbula varia (L.)- further evidence of non-colinearity in molluscs. Dev. Genes Evol. 220, 161–172. doi: 10.1007/s00427-010-0338-0
Schiavina, M., Marino, I. A. M., Zane, L., and Melià, P. (2014). Matching oceanography and genetics at the basin scale. Seascape connectivity of the Mediterranean shore crab in the Adriatic Sea. Mol. Ecol. 23, 5496–5507. doi: 10.1111/mec.12956
Sponaugle, S., Cowen, R. K., Shanks, A., Morgan, S. G., Leis, J. M., Pineda, J., et al. (2002). Predicting self-recruitment in marine populations: biophysical correlates and mechanisms. Bull. Mar. Sci. 70, 341–375.
Templado, J. (2011). “Familia Trochidae,” in Moluscos marinos de Andalucía, eds S. Gofas, D. Moreno, and C. Salas (Málaga: Universidad de Málaga), 104–117.
Templado, J. (2014). “Future trends of mediterranean biodiversity,” in The Mediterranean Sea. Its History and Present Challenges, eds S. Goffredo and Z. Dubinsky (Berlin: Springer), 479–498.
Underwood, A. J. (1972). Spawning, larval development and settlement behaviour of Gibbula cineraria (Gatropoda: Prosobranchia) with a reppraisal of torsion in gastropods. Mar. Biol. 17, 341–349. doi: 10.1007/BF00366745
Underwood, J., Smith, L., Van Oppen, M., and Gilmour, J. (2007). Multiple scales of genetic connectivity in a brooding coral on isolated reefs following catastrophic bleaching. Mol. Ecol. 16, 771–784. doi: 10.1111/j.1365-294X.2006.03187.x
Uribe, J. E., Williams, S., Templado, J., Buge, B., and Zardoya, R. (2017). Phylogenetic relationships of Mediterranean and North-East Atlantic Cantharidinae and notes on Stomatellinae (Vetigastropoda: Trochidae). Mol. Phylogenet. Evol. 107, 64–79. doi: 10.1016/j.ympev.2016.10.009
Van Oosterhout, C., Hutchinson, W. F., Wills, D. P. M., and Shipley, P. (2004). MICROCHECKER: software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 4, 535–538. doi: 10.1111/j.1471-8286.2004.00684.x
Vartia, S., Collins, P. C., Cross, T. F., FitzGerald, R. D., Gauthier, D. T., McGinnity, P., et al. (2014). Multiplexing with three-primer PCR for rapid and economical microsatellite validation. Hereditas 151, 43–54. doi: 10.1111/hrd2.00044
Villamor, A., Costantini, F., and Abbiati, M. (2014). Genetic structuring across marine biogeographic boundaries in rocky shore invertebrates. PLoS One 9:e101135. doi: 10.1371/journal.pone.0101135
Weir, B. S., and Hill, W. G. (2002). Estimating F-statistics. Annu. Rev. Genet. 36, 721–750. doi: 10.1146/annurev.genet.36.050802.093940
Keywords: Adriatic Sea, genetic connectivity, Gibbula divaricata, microsatellites, dispersal capacity
Citation: López-Márquez V, Templado J, Buckley D, Marino I, Boscari E, Micu D, Zane L and Machordom A (2019) Connectivity Among Populations of the Top Shell Gibbula divaricata in the Adriatic Sea. Front. Genet. 10:177. doi: 10.3389/fgene.2019.00177
Received: 27 July 2018; Accepted: 18 February 2019;
Published: 08 March 2019.
Edited by:
Tian Tang, Sun Yat-sen University, ChinaReviewed by:
Lifeng Zhu, Nanjing Normal University, ChinaPeter J. Prentis, Queensland University of Technology, Australia
Rocío Pérez-Portela, Universidad Rey Juan Carlos, Spain
Copyright © 2019 López-Márquez, Templado, Buckley, Marino, Boscari, Micu, Zane and Machordom. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Annie Machordom, YW5uaWVAbW5jbi5jc2ljLmVz