 Alexander Owusu Boakye1,2
Alexander Owusu Boakye1,2 Christian Obirikorang1,2
Christian Obirikorang1,2 Anthony Afum-Adjei Awuah1,2
Anthony Afum-Adjei Awuah1,2 Evans Asamoah Adu1,2Doris Winter3Eric Ebenezer Boham1,2Hakim Alani1
Evans Asamoah Adu1,2Doris Winter3Eric Ebenezer Boham1,2Hakim Alani1 Sylvester Kofi Newton1Nana Safi Toure Almoustapha1
Sylvester Kofi Newton1Nana Safi Toure Almoustapha1 James Deke1,4Welbeck Odame Dzadey1
James Deke1,4Welbeck Odame Dzadey1 Louis Adu-Amoah1,2Sally-Ann Kroduah1,5
Louis Adu-Amoah1,2Sally-Ann Kroduah1,5 Mary Ama Grant1Gracelyn Asare1Amos Amoako-Adusei1Wibke Loag3
Mary Ama Grant1Gracelyn Asare1Amos Amoako-Adusei1Wibke Loag3 Jenny Kettenbeil3Yaw Adu Sarkodie4Ebenezer Oduro-Mensah6Alfred Edwin Yawson7Stephen Apanga8Rose Odotei Adjei1,5Austin Gideon Adobasom-Anane1,5Eva Lorenz3,9Aurélia Souares10,11Oumou Maiga-Ascofaré1,3,9
Jenny Kettenbeil3Yaw Adu Sarkodie4Ebenezer Oduro-Mensah6Alfred Edwin Yawson7Stephen Apanga8Rose Odotei Adjei1,5Austin Gideon Adobasom-Anane1,5Eva Lorenz3,9Aurélia Souares10,11Oumou Maiga-Ascofaré1,3,9 Jürgen May3,9,12
Jürgen May3,9,12 Nicole S. Struck3,9*†John Humphery Amuasi1,3,5,12*†
Nicole S. Struck3,9*†John Humphery Amuasi1,3,5,12*†- 1Kumasi Centre for Collaborative Research in Tropical Medicine (KCCR), Kwame Nkrumah University of Science and Technology, Kumasi, Ghana
- 2Department of Molecular Medicine, School of Medicine and Dentistry, College of Health Sciences, Kwame Nkrumah University of Science and Technology (KNUST), Kumasi, Ghana
- 3Infectious Disease Epidemiology, Bernhard Nocht Institute for Tropical Medicine, Hamburg, Germany
- 4Department of Clinical Microbiology, School of Medicine and Dentistry, College of Health Sciences, Kwame Nkrumah University of Science and Technology (KNUST), Kumasi, Ghana
- 5Department of Global and International Health, School of Public Health, College of Health Sciences, Kwame Nkrumah University of Science and Technology (KNUST), Kumasi, Ghana
- 6Ga East Hospital, Accra, Ghana
- 7Department of Community Health, Medical School, University of Ghana, Accra, Ghana
- 8Department of Community Health and Preventive Medicine, School of Medicine, University for Development Studies, Tamale, Ghana
- 9German Center for Infection Research (DZIF), Partner Site Hamburg-Borstel-Lübeck-Riems, Hamburg, Germany
- 10German Center for Infection Research (DZIF), Partner Site Heidelberg, Heidelberg, Germany
- 11Heidelberg Institute of Global Health (HIGH), Heidelberg University Hospital, Heidelberg, Germany
- 12Department of Tropical Medicine I, University Medical Centre Hamburg Eppendorf (UKE), Hamburg, Germany
Background: Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), enters human cells using the angiotensin-converting enzyme 2 (ACE-2) receptor. ACE2 single nucleotide polymorphisms (SNPs) can influence susceptibility by affecting viral binding or gene expression. This study investigated the association between ACE2 SNPs, rs2285666 and rs2106809, and the SARS-CoV-2 infection susceptibility in a Ghanaian population.
Methods: Genomic DNA was extracted, using a magnetic bead-based method, from blood samples of a random-subset of 1,334 participants drawn from a two-stage cluster, population-based household cross-sectional SARS-CoV-2 IgG seroprevalence survey. Data collected included, socio-demographic characteristics, medical history, vaccination, and smoking status. Genotyping of the ACE2 SNPs was performed using Allele-Specific Oligonucleotide Polymerase Chain Reaction (ASO-PCR) combined with melting curve analysis. Logistic regression models were utilized to assess the association between the ACE2 SNPs and the susceptibility to SARS-CoV-2 infection
Results: The median age of participants was 33 [Interquartile range (IQR) = 24–46] years. Females accounted for the majority of the sampled population, 64.3%. SARS-CoV-2-IgG seropositivity was (58.4%, 95%CI: 52.6%–64.2%) among the male population and (54.1%, 95%CI: 49.54%–58.61%) in the female population. There were no significant differences in overall allele or genotype frequencies of ACE2 SNPs between SARS-CoV-2 IgG seropositive and seronegative individuals for both females and males. Among females, those with the T allele of ACE2 rs2285666 had a 38% decreased susceptibility to SARS-CoV-2 infection under the dominant [adjusted odds ratio (aOR) = 0.62; 95%CI = 0.45–0.85, P = 0.003] and heterozygous advantage models (aOR = 0.62; 95%CI = 0.45–0.86, P = 0.004), after adjusting for confounders, but not thee recessive model (aOR = 0.41; 95%CI = 0.03–5.22, P = 0.490). No significant association was observed among males. Overall, the ACE2 rs2106809 was not associated with the susceptibility to SARS-CoV-2 infection in both males and females.
Conclusion: This study found no association between ACE2 rs2106809 genetic variant and susceptibility to SARS-CoV-2 infection, whilst the rs2285666 T-allele was associated with a decreased frequency for SARS-CoV-2 infection among Ghanaian females. These findings enhance our understanding of genetic factors influencing SARS-CoV-2 susceptibility, which could help identify at-risk populations and inform more targeted public health interventions in future outbreaks.
Introduction
The severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) was identified as the causative agent of the recent global pandemic, coronavirus disease 2019 (COVID-19) (Zhu et al., 2020). Declared as a pandemic in early 2020 (WHO, 2020b), this outbreak significantly disrupted global health systems, societal norms, and economic structures, resulting in widespread morbidity and mortality. As of June 2024, over 700 million confirmed cases and 7.1 million deaths have been recorded globally. Through a combination of scientific innovation, global collaboration, and coordinated public health efforts, the world has made remarkable progress in recovering from the COVID-19 pandemic. The rapid development, approval, and rollout of vaccines have played a critical role in the process. As such, focus has shifted from emergency response to managing COVID-19 as an endemic situation, with the WHO officially announcing it no longer a Public Health Emergency of International Concern (PHEIC) (Statement on the fifteenth meeting of the IHR, 2005). Despite the progress made, it is important to remain vigilant, as the virus still looms as a formidable challenge to global health systems due to its ability to evolve and persist, and also the risk of other emerging infections remains.
The SARS-CoV-2 virus penetrates human cells via attachment of the trimeric SARS-CoV-2 spike (S) protein receptor binding domain (RBD) to the angiotensin-converting enzyme 2 (ACE2) (Hoffmann et al., 2020; Letko et al., 2020; Wan et al., 2020; Zhou et al., 2020). Host genetics has long been known to play a key function in establishing the outcome of host-pathogen interactions (susceptibility or protection) and subsequently influencing infection outcomes (mild, moderate, or severe) (De Silva and Stumpf, 2004; Samson et al., 1996; Martinson et al., 1997). Such genetic factors also contribute to population-specific traits, potentially explaining disparities in disease burden and severity across different populations, as highlighted in the context of COVID-19.
Researchers have suggested that genetic changes including single nucleotide polymorphisms (SNPs) within ACE2 could either alter the binding affinity to the virus or gene expression levels, thereby influencing an individual’s susceptibility or protection from infection (Martínez-Gómez et al., 2022; Möhlendick et al., 2021; Mahmood et al., 2022; Liu et al., 2016). While there is ample research exploring the impact of host genetics, particularly, ACE2 SNPs on SARS-CoV-2 infection in Europe, Asia, and other global regions, there is considerable knowledge gap regarding these genetic associations within the African population. This is due to the absence of a well-established and robust laboratory technology pipeline such as high throughput next-generation sequencing and genotyping approaches, which are essential to conducting similar studies (Petersen et al., 2022). Additionally, the distinct genetic diversity of Africa, shaped by early human origins (Stringer, 2016), endemic diseases (Adimulam et al., 2023), and a large socio-cultural structure (Schlebusch et al., 2012), distinguishes it from other populations (Campbell and Tishkoff, 2008; Yu et al., 2002). Consequently, the current global evidence derived from existing studies lacks comprehensive representation of genetic diversity, limiting our ability to draw consistent conclusions across populations. In sub-Saharan Africa, several SNPs have been associated with infectious diseases. For example, constant exposure to the parasite in malaria-endemic regions have driven the natural selection for protective genetic variations (Dhangadamajhi et al., 2010). The unique genetic background raises critical concerns about the generalizability of genetic findings across diverse global populations, including African populations.
ACE2-rs2285666 (C>T) and ACE2-rs2106809 (A>G) have been recognized as important SNPs that regulates ACE2 expression levels, which may, in turn affect the number of available receptor sites for SARS-CoV-2 virus attachment to cause an infection, thereby potentially conferring resistance to infection (Dhangadamajhi et al., 2010; De et al., 2021).
Ghana reflects the broader trends of Sub-Saharan Africa, with significant genetic diversity, unique epidemiological profile, and a history of malaria endemicity (WHO, 2023b). Investigating ACE2 SNPs in the Ghanaian population may reveal insights into COVID-19 susceptibility patterns and inform therapeutic strategies for future (coronavirus) outbreaks.
Investigating the association between ACE2 SNPs and the susceptibility to SARS-CoV-2 infection using SARS-CoV-2 IgG seropositivity, determined through robust methods as a proxy allows for a more inclusive and practical solution, particularly in resource-limited settings like Ghana. Other diagnostic methods, such as PCR testing, though the reference standard, are expensive and logistically difficult to scale, leading to testing strategies that primarily target specific groups rather than the broader population in such settings (COVID-19, 2024). While PCR testing detects acute infections, seropositivity provides evidence of past infections and thus captures the cumulative exposure to SARS-CoV-2 over time. This broader temporal coverage makes seropositivity particularly suitable for studying genetic susceptibility to infection in population-based studies.
By leveraging SARS-CoV-2 IgG serostatus from a highly representative population sample, this approach provides an alternative framework for conducting genetic association studies. Unlike traditional case-control studies, which require carefully matched controls and active cases, this alternative framework addresses the challenges of control selection–particularly in situations with high rates of asymptomatic cases, as seen with COVID-19 (Zhao et al., 2020; Oran and Topol, 2020). Based on this framework, this study hypothesizes that ACE2 SNPs are associated with SARS-CoV-2 infection susceptibility in the Ghanaian population. Specifically, it aims to (a) assess the sex-specific genetic association of ACE2-rs2285666 (C>T) and ACE2-rs2106809 (A>G) with SARS-CoV-2 IgG seropositivity, and (b) evaluate their genotypic and allelic frequencies.
Materials and methods
Study design and population
The study was a sub-study of a larger SARS-CoV-2 IgG seroprevalence study in Ghana, conducted in accordance with the previously published SeroCoV protocol (Lorenz et al., 2021). This protocol aligned with the World Health Organization’s standardized guidelines for seroepidemiological studies (WHO, 2020a).
Briefly, a non-random subset of participants was drawn from a two-stage cluster population-based household cross-sectional study where participants were recruited from three major cities–namely, Kumasi (Struck et al., 2022), Accra, and Tamale–covering the period from February 2021, and February 2022. The survey comprised three phases: the first phase between February 2021, and March 2021, in Kumasi, the second phase between June 2021, and October 2021, in Accra, and the third phase between November 2021, and February 2022, in Tamale.
The study included individuals aged 10 years and above who resided in households within selected sampling frame, provided informed consent for genetic analysis, and had their SARS-CoV-2 IgG seropositivity results, and complete demographic and SNP genotyping data available from the main study. To ensure that seropositivity reflected natural SARS-CoV-2 infection rather than vaccine-induced immunity, individuals who had received at least one of any of the rolled-out COVID-19 vaccination at the time of the main study were excluded.
Detection of SARS-CoV-2-specific IgG seropositivity
SARS-CoV-2-specific IgG antibodies were measured in participant plasma samples using a sensitive and specific qualitative enzymed-linked immunosorbent assay (ELISA), following manufacturer’s guidelines.
Fcγ-receptor-based anti-SARS-CoV-2-specific IgG ELISA kit was developed by the Diagnostics Development Laboratory (DDL) at the Bernhard Nocht Institute for Tropical Medicine (BNITM), Hamburg, Germany (patent EP2492689) (Schmitz et al., 2011), and used to determine SARS-CoV-2 serostatus in the presented study. This ELISA operates by capturing antigen-antibody complexes via a solid-phase-bound Fcγ-receptor (FcγR), with a truncated SARS-CoV-2 nucleocapsid protein (NCP) as the target antigen and has proven to be highly specific. The assay was tested and validated using a serum panel derived from 35 COVID-19 patients with 213 longitudinal samples, as well as a control panel of negative samples containing 790 preCOVID-19 samples sourced from a variety of geographical regions, including Europe (Germany), Asia (Laos), and Africa (Ghana, Madagascar, and Nigeria), designed to address the issue of limited specificity of commercialized ELISA assays among samples originating from malaria-endemic countries (Deschermeier et al., 2022; Emmerich et al., 2021). Results were interpreted based on criteria set by the manufacturer, according to the batch specific certificate of analysis for the positive and negative controls used in the assay.
Genomic DNA extraction
Following blood collection during the initial study, samples were centrifuged, and the cell pellet resuspended in 8M Urea for stable storage at room temperature. Genomic DNA was extracted using the Mag Maxi DNA extraction kit (Biosearch technologies, United Kingdom), according to the manufacturer’s instructions. Briefly, equal volumes of sample and lysis buffer were mixed and incubated overnight at 55°C with 20 µL protease. Magnetic particles were added, and bound DNA was washed, eluted, and measured using NanoDrop®.
Primer design and information
The Modified Tetra-Primer Assay (MTPA) PCR primer design tool was used to design primers (Tanha et al., 2015) using GenBank sequence of ACE2 rs2285666 and ACE2 rs2106809 (NC_000023.10:g.15610348C>T, and NC_000023.11:g.15599938A>G), respectively.
For each SNP, the MTPA PCR primer design tool revealed two sets of primers (two outer and two inner primers) (Tanha et al., 2015; Baris et al., 2013; Etlik et al., 2008). Two distinct primer sets are employed to amplify two smaller and allele-specific fragments. The outer primers amplify the common, much larger DNA fragment containing the SNP, whereas the inner primers (allele-specific primers) amplify the two allele-specific fragments - wild-type and mutant-type allele-specific fragments - concurrently. The mutant allele-specific fragment is amplified by the forward inner primer and the reverse outer primer, while the wildtype allele-specific fragment is amplified by the reverse inner primer and the forward inner primer.
The NCBI online Primer Blast program at (https://www.ncbi.nlm.nih.gov/tools/primer-blast/) and the online Oligo-Analyzer 3.1 tool by Integrated DNA Technologies (IDT) (https://eu.idtdna.com/calc/analyzer) was employed to assess the specificity of the selected primer. The primers used are shown in Supplementary Table S1.
Genotyping of ACE2 rs2285666 and rs2106809
Genotyping was performed using real-time PCR on the LineGene 9600 thermocycler (Hangzhou Bioer Technology Co. Ltd, China). For each ACE2 SNP, two separate allele-specific reactions were prepared to target the mutant and wildtype alleles with concentrations, as specified in Supplementary Table S1.
PCR amplification conditions were set up as follows: initial denaturation at 95°C for 5 min. For rs2285666, amplification continued with 40 cycles of 95°C for 10 s (denaturation), 50.9°C for 20 s (annealing), and 72°C for 30 s (extension). For rs2106809, 30 cycles were performed with an annealing temperature of 55°C. After amplification, allele-specific fragment product melting curves were analysed by first heating the samples to 95°C for 15 s, cooling to 65°C for 1 min, and then gradually increasing the temperature to 97°C at a rate of 0.2°C/min for 15 s, while continuously measuring the change in fluorescence. The thermocycler calculated the negative derivative of the fluorescence change and generated a melting curve for each sample. Melting peaks were visualized as the negative derivative of fluorescence versus temperature (-dF/dT) of the melting curve for amplification products.
In each PCR run, specific known genotyped control samples were used: NA18499 (CT) and NA19118 (CC) for rs2285666, and HG02769 (AG) and NA19118 (AA) for rs2106809. Control samples were selected from the National Human Genome Research Institute (NHGRI) repository, using genotype data available through the Ensembl database (https://www.ensembl.org/), and were obtained from the Coriell Institute for Medical Research (http://www.coriell.org/). These controls were essential for validating primer specificity and optimizing PCR conditions, enabling precise differentiation between heterozygous and homozygous genotypes. For details on control samples used, see Supplementary Table S2. As an additional verification step, PCR-specific products were run on 1.5% agarose gel electrophoresis to confirm the presence and band sizes of products obtained (Supplementary Figure S1).
Statistical analysis
Data analyses were carried out with the Statistical Package for the Social Sciences (SPSS) version 26.0 (IBM Corp. Released 2019. IBM SPSS Statistics for windows, Version 26.0. Armonk, N.Y: IBM Corp) and R studio software (R: The R Project for Statistical Computing).
The Chi-square test was used to assess Hardy-Weinberg Equilibrium (HWE) analysis for the two SNPs and compare the genotype and allele frequencies of rs2285666 (C>T) and rs2106809 (A>G) between SARSCoV-2 IgG-seropositive and seronegative participants. SARS-CoV-2 IgG seropositivity was used as a proxy to assess the susceptibility to SARS-CoV-2 infection. To determine the association between ACE2 SNPs and infection susceptibility, logistic regression was used to estimate odds ratios (ORs) and 95% confidence intervals (CIs). This analysis considered each SNP as a predictive variable while adjusting for potential confounders, including participant age, underlying health conditions, and smoking status. Additionally, the study site was included as an offset variable to account for potential variations in seropositivity due to differences in sampling times across the three study sites.
Given the ACE2 gene is located on the human X chromosome (gene locus Xp22.2), its inheritance patterns differ between sexes—males (XY) having a single copy (hemizygous) and females (XX) having two copies (homozygous or heterozygous). Therefore, separate analyses were performed to account for potential sex-specific differences. Various inheritance models, including dominant, heterozygous advantage, recessive, and allele models, were considered in the association analysis for each SNP. For all tests, the statistical significance level was set at α = 0.05. All P-values were adjusted for multiple testing using the Benjamini–Hochberg (BH) procedure, where appropriate.
Results
Socio-demographic and clinical characteristics of study participants
1,993 participants from 910 households in Kumasi, Accra, and Tamale were eligible for analysis. However, 354 subjects had received at least one dose of the COVID-19 vaccination and 308 did not have complete SNP genotyping data. As a result, data from 1,334 participants and 691 households were included in this analysis, with the majority (64.3%) being female subjects. To determine the relationship between ACE2 SNPs and the susceptibility to SARS-CoV-2 infection, two groups were compared: 55.6% (742) SARS-CoV-2 IgG seropositive, and 44.4% (592) SARS-CoV-2 IgG seronegative.
The median age was 33 (IQR = 24–46) years among SARS-CoV-2-IgG seronegative and 31 (IQR = 23–45) for SARS-CoV-2-IgG seropositive individuals. SARS-CoV-2-IgG seropositive and SARS-CoV-2-IgG seronegative participants varied considerably across study sites (except for Accra), demographic groups, and socio-behavioral factors (Table 1). Overall, females predominated (64.3%). Males demonstrated a seropositivity rate of 58.4%, (95%CI: 52.6%–64.2%) while females reported a seropositivity rate of (54.1%, 95%CI: 49.54%–58.61%), when seropositivity was analysed within sex. Education levels differed across participants, with primary education being the most frequently reported, and a higher proportion of seropositive individuals having no formal education compared to seronegatives. Nearly 41.5% were engaged in high-risk occupations, while the vast majority had no underlying health conditions (87.2%) and did not smoke (97.3%). Clinically, 42.7% reported no symptoms in the 12 months before the study, while 26.8% were paucisymptomatic and 30.4% symptomatic. Reported exposure to confirmed COVID-19 cases was low (5.5%), though room sharing (80.5%) and outdoor work (43.3%) were common. Most participants had 10 to fewer than 50 daily contacts, and 41.0% reported a history of travel. The full version of the table, including 95% confidence intervals is available in the Supplementary Table S3.
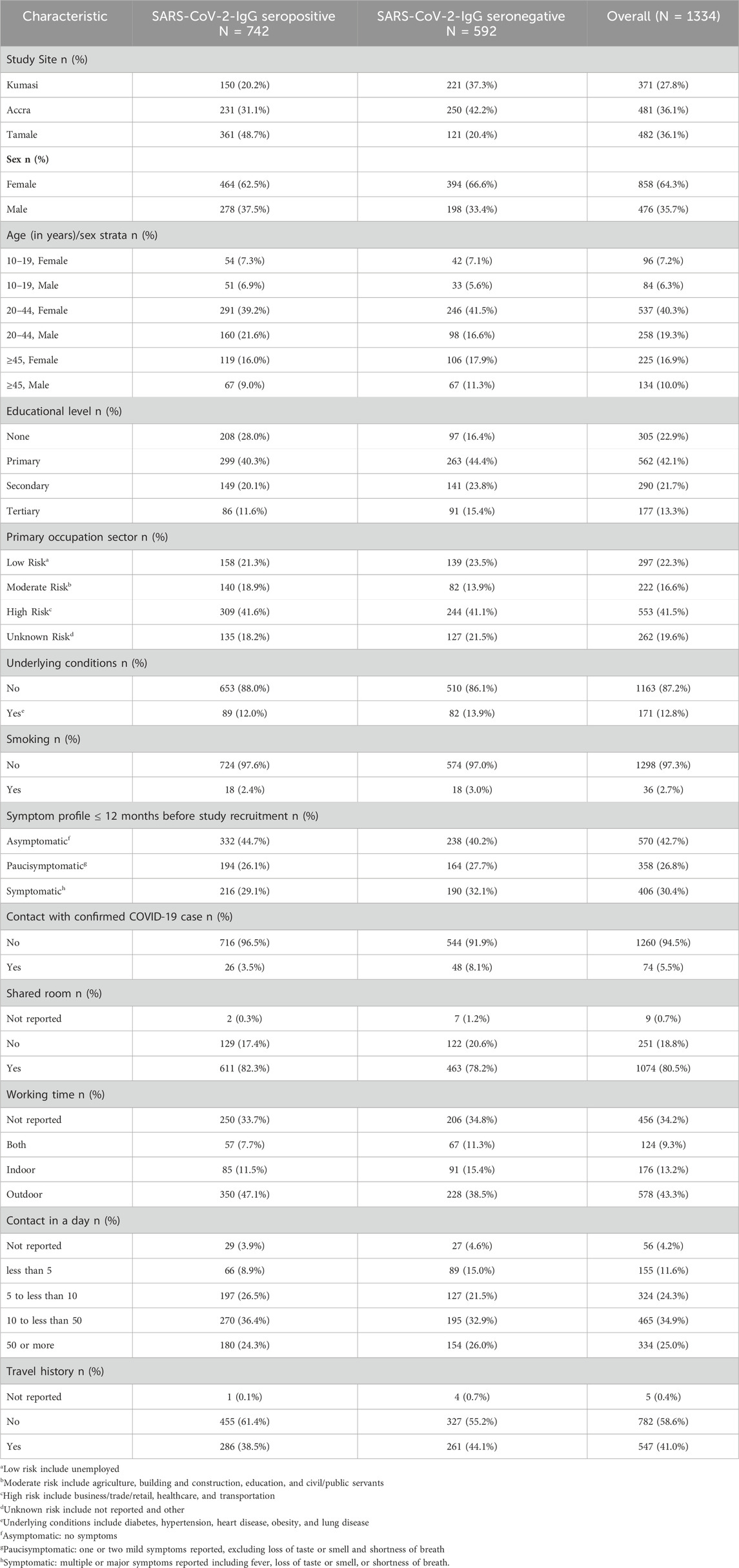
Table 1. Socio-demographic and clinical characteristics of study participants.
Genotype and allele frequencies of ACE2 SNPs
Table 2 shows the genotype and allele frequency distribution of ACE2 SNPs among the study population. Males (XY) were represented by allele frequencies due to their hemizygosity, while females (XX) were represented by genotype frequency (homozygous or heterozygous). The distribution for both SNPs were consistent with Hardy-Weinberg equilibrium, with (P = 0.697) for rs2285666 and (P = 0.366) for rs2106809.
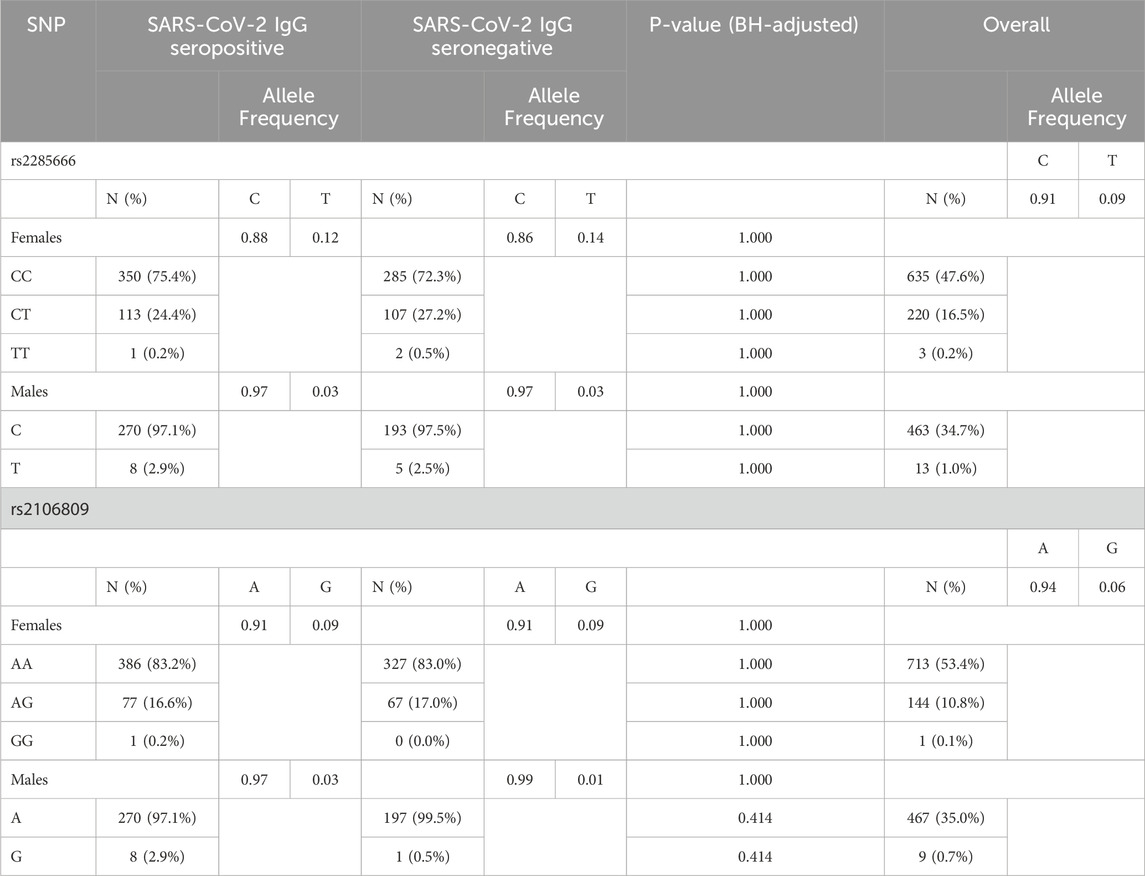
Table 2. Genotype and allele frequency distributions among study participants.
The analysis did not reveal significant differences in overall allele or genotype frequencies between SARS-CoV-2 IgG seropositive and seronegative individuals, for both females and males. For ACE2 rs2285666, genotype distribution was 75.4% CC, 24.4% CT, and 0.2% TT in SARS-CoV-2-IgG seropositive females and 97.1% C and 2.9% T in SARS-CoV-2-IgG seropositive males, with a minor allele frequency (MAF) of 0.12 and 0.03 for the T-allele in females and males, respectively. The genotypes for ACE2 rs2106809 in SARS-CoV-2 IgG seropositive individuals were distributed as follows: 83.2% AA, 16.6% AG, 0.2% GG in females and 97.1% A and 2.9% in males. The minor allele frequency (MAF) for the G allele was found to be 0.09 and 0.03 in SARS-CoV-2-IgG seropositive females and males, respectively.
Association between ACE2 SNPs and the susceptibility to SARS-CoV-2 infection
The association between ACE2 SNPs and the susceptibility to SARS-CoV-2 infection is shown in Table 3. To account for sex-specific differences, association analysis was conducted separately for males and females. Briefly, there was no evidence for an association between ACE2 - rs2285666 and rs2106809 SNPs and the susceptibility to SARS-CoV-2 infection under the crude model. After adjusting for age, underlying conditions, and smoking, with the study site set as an offset variable to account for study site-specific variations the analysis revealed sex-specific patterns for ACE2 rs2285666 and the susceptibility to SARS-CoV-2 infection. Among females, those with at least one copy of the T allele showed 38% reduced odds of susceptibility to SARS-CoV-2 infection under both the dominant (aOR = 0.62; 95%CI = 0.45–0.85, BH-adjusted P = 0.017) and heterozygous advantage models (aOR = 0.62; 95%CI = 0.45–0.86, BH-adjusted P = 0.017). The effect estimate under the recessive model was similar in direction but had substantially wider confidence intervals (aOR = 0.41; 95%CI = 0.03–5.22, BH-adjusted P = 0.637), reflecting the small number of individuals with two copies of the T allele. Among males, no clear direction of associations was found under the dominant (aOR = 1.00; 95% CI = 0.30–3.32; BH-adjusted P = 0.998), and recessive models (aOR = 1.00; 95% CI = 0.30–3.32; BH-adjusted P = 0.998). Notably, in the allelic model, where the C allele was treated as the effect allele, an increased odds of susceptibility to SARS-CoV-2 infection was observed (aOR = 1.62, BH-adjusted P = 0.017), which aligns with the protective role associated with the T allele observed under the dominant and heterozygous models. These findings may be explained by a potential sex-specific genetic effect, with the T allele of ACE2 rs2285666 being more protective against SARS-CoV-2 infection in females than males (Table 3).
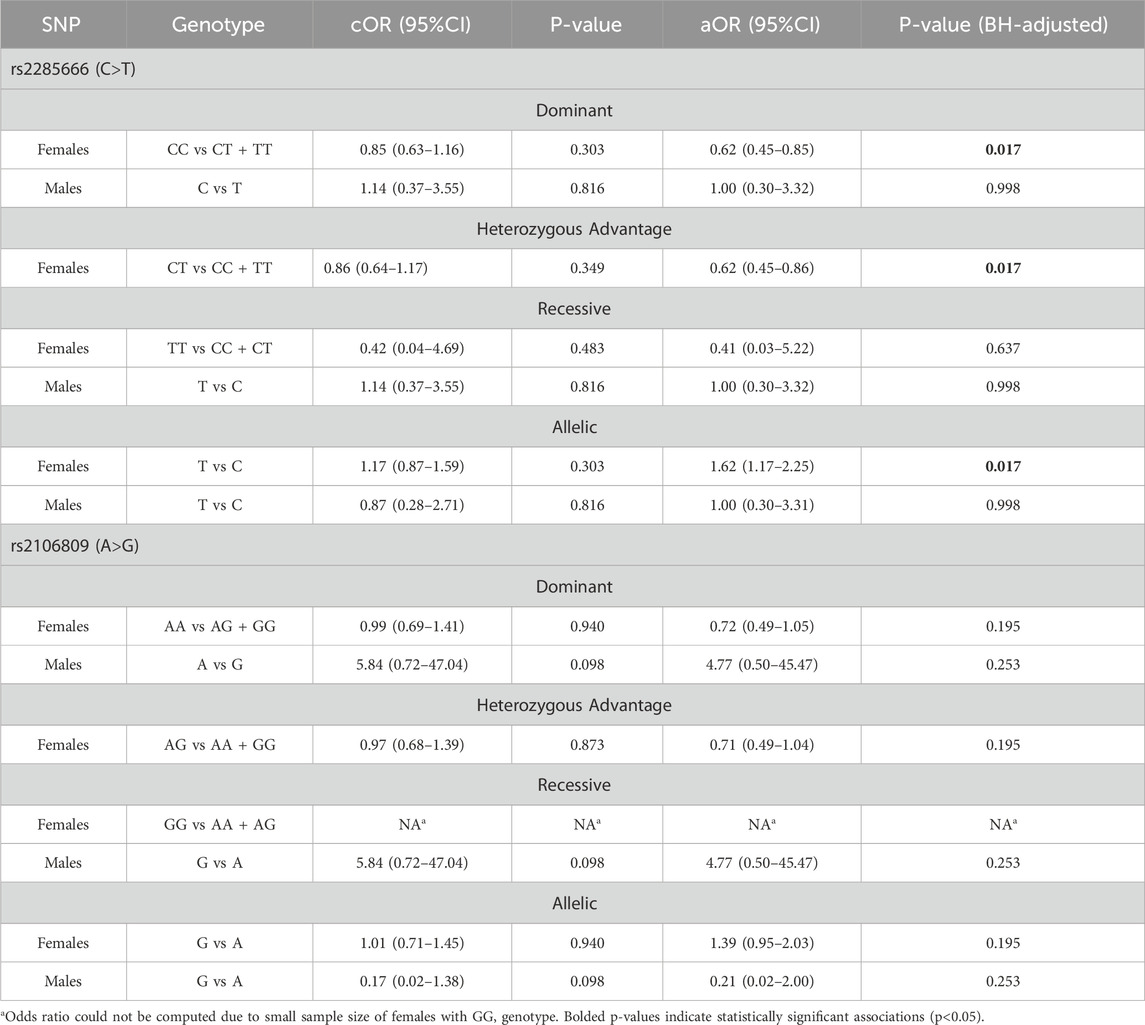
Table 3. Association between rs2285666 and rs2106809 and susceptibility to SARS-COV-2 infection.
Discussion
The COVID-19 pandemic presented with varying infection rates between and within populations (Bialek et al., 2020; WHO, 2023a). Factors contributing to this disparity, includes behavioral (Betsch et al., 2020; Chinazzi et al., 2020; Courtemanche et al., 2020), social (Dorn et al., 2020; Lewer et al., 2020; Yancy, 2020), physiological (Kellam and Barclay, 2020; Guan et al., 2020), and biological variables (Kellam and Barclay, 2020; Eshetie et al., 2023). As of June 26, 2022, there were 542,018,955 confirmed cases globally, with WHO regional disparities in distribution (WHO, 2023): 1.7% in the African region, 29.9% in the Americas, 4.0% in the Eastern Mediterranean, 41.8% in Europe, 10.8% in South-East Asia, and 11.7% in the Western Pacific. In Ghana, over 100,000 confirmed cases and 1,450 deaths were recorded during the same period, reflecting the pandemic’s varied impact across regions and populations (GHS, 2023).
ACE2 is the major receptor for SARS-CoV-2, which mediates cell entry (WHO, 2023a). Within the complex network of disease pathogenesis, SNPs in key genes have long been recognized as critical influencers affecting an individual’s susceptibility to infection (Gray et al., 2000). ACE2 SNPs have already been linked to various human illnesses, including hypertension (Bosso et al., 2020), heart failure (Chaoxin et al., 2013), malaria (Dhangadamajhi et al., 2010), and diabetes (Chaoxin et al., 2013). Recent findings have shown inconsistent findings on the association between ACE2 SNPs and the likelihood of SARS-CoV-2 infection, with an emphasis on Eurasian populations and limited investigations undertaken in African populations (Petersen et al., 2022). Differences in these findings may be attributed to genetic and environmental factors within the investigated populations (Price et al., 2010; Aschard et al., 2010), as well as variations in study design and methodology used (Visscher et al., 2017).
The two x-chromosomal ACE2 mutations rs2285666 (C>T) and rs2106809 (A>G) investigated in this study have been associated with elevated levels of angiotensin II (AngII), which is known to protect against severe malaria (Dhangadamajhi et al., 2010). Because AngII levels are regulated by ACE2 activity within the renin-angiotensin-aldosterone system (RAAS) (Ferrario, 2006), this indicates a decrease in ACE2 expression levels, also associated with these polymorphisms (Dhangadamajhi et al., 2010; Chen et al., 2018). However, analysis of publicly available GTEx data indicates that both rs2285666 and rs2106809 are associated with increased ACE2 expression in multiple tissues, particularly in the hypothalamus, nucleus accumbens, and pituitary, suggesting a more complex and tissue-specific regulatory effect than previously assumed (GTEx Portal, 2025). Membrane-bound (mACE2) and circulating or soluble ACE2 (cACE2 or sACE2), have been reported to retain catalytic activity and contain SARS-CoV-2 binding sites, which may contribute to onset of disease or disease progression (Medina-Enríquez et al., 2020; Yeung et al., 2021). Understanding how these polymorphisms modulate ACE2 expression across different tissues, particularly in populations with varied genetic backgrounds is needed.
The overall genotype and allele frequency distribution were comparable between SARS-CoV-2-IgG seropositive and seronegative groups for both females and males. However, ACE2-rs2285666 (T-allele) was found to be associated with protection from SARS-CoV-2 infection among the female population. This association was observed after adjusting for age, underlying medical conditions, participants’ smoking status, and study site sampling variations. In males, no discernible association was observed. This finding may be explained by a sex-specific functional role of the T-allele. Dhangadamajhi et al., (2010) observed that the T allele reduces ACE2 expression levels in women, specifically (Dhangadamajhi et al., 2010). Factors, such as sex hormones (Lott et al., 2023) and diverse immune responses (Ghosh and Klein, 2017), may contribute to such an association. Future studies investigating the interplay between sex hormones, humoral responses, and genetics would help understanding sex-specific differences in susceptibility to SARS-CoV-2 infection. Consistent with the findings of the rs2285666 SNP in this study, Möhlendick et al., (2021) reported an almost twofold increased SARS-CoV-2 infection susceptibility among individuals carrying the GG genotype (OR = 1.91, 95%CI: 1.13–3.24; P = 0.02) or G allele (OR = 1.88, 95%CI: 1.12–3.16; P = 0.02) of the rs2285666 SNP. However, differences exist as the previous study employed a highly selected cohort of PCR-confirmed SARS-COV-2 cases and controls, while the current study used population-based SARS-CoV-2-specific IgG serosurvey data. In addition, the previous study did not consider possible sex-specific ACE2 SNP-associated SARS-CoV-2 susceptibility differences as observed in this study. The frequency of the T allele varies across populations, with a higher prevalence observed in Asian populations compared to European and African populations (Srivastava et al., 2020). This suggests that genetic susceptibility to SARS-CoV-2 infection may differ among ethnic groups. Although studies have reported higher SARS-CoV-2 infection rates in males than females (Do Nascimento et al., 2020; The COVID-19 Sex-Disaggregated Data Tracker, 2025; Conti and Younes, 2020; Rostami et al., 2021; Vahidy et al., 2021), the findings of this study should be interpreted within the broader context of multiple factors influencing infection susceptibility.
Regarding the ACE2 rs2106809 (A>G), there was no significant association with SARS-CoV-2 infection susceptibility in both sexes. Although this finding does not necessarily negate the importance of the rs2106809 SNP in SARS-CoV-2 infection, they underscore how factors such as age, underlying condition, and smoking shape an individual’s susceptibility. This accentuates the necessity for comprehensive and multifactorial methods in genetic studies. The lack of a significant association could be attributed to the unique genetic makeup of the Ghanaian population, suggesting that different populations may yield diverse results, underscoring the need for replicative studies. While other studies have explored how this SNP is associated with the severity of COVID-19 (Cafiero et al., 2021; Karakaş Çelik et al., 2021; Mohammadi-Berenjestanaki et al., 2023; Sabater Molina et al., 2022), this study specifically explored its predisposition to SARS-CoV-2 infection, with insufficient evidence available for making direct comparisons.
It is worth noting that several other critical genes have been implicated in COVID-19 genetic studies, in addition to ACE2, across different populations. Numerous loci have shown associations with both susceptibility to infection and disease severity. Early genome-wide association study (GWAS) highlighted the significance of genes such as SLC6A20, LZTFL1, CCR9, FYCO1, CXCR6, and XCR1 as key determinants of SARS-CoV-2 susceptibility (lead variant rs2271616) (Niemi et al., 2021). Furthermore, IFNAR2 and IL10RB have been linked to increased SARS-CoV-2 infection susceptibility (Kasela et al., 2021). Additional studies have further reported associations with the genes ABO, DPP9, HLA, OAS1, SLC22A31, SFTPD, AND CXCR6 (Ellinghaus et al., 2020; Pairo-Castineira et al., 2020; Pairo-Castineira et al., 2023). Integrating polygenic risk scores and fine-mapping approaches in COVID-19 genetic studies could help clarify the role of these loci in diverse genetic backgrounds.
This study benefited from a population-representative sample derived from a household-based cross-sectional survey and utilized a highly specific ELISA assay for estimating seroprevalence. This study also finds its strength by the use of a robust genotyping assay bolstered by the inclusion of already known genotyped samples as controls, improving its sensitivity and specificity for accurate and reliable SNP detection. However, the study has several limitations. While it aimed for a population-representative sample, the specific eligibility criteria set out for this study reduced its initial representativeness. The exclusion of vaccinated individuals may have selectively removed certain groups from the analyses; thus, findings should be interpreted cautiously. The relatively small sample size for males may have limited the ability to detect significant associations in this study. Additionally, the cross-sectional design of the main study only allows for the assessment of associations at a single point in time, lacking the ability to account for the temporal dynamics of the COVID-19 epidemic. Another limitation is the inability to draw firm conclusions about the functional consequences of the investigated ACE2 SNPs on ACE2 expression, which precludes the direct assessment of the mechanistic link between SARS-CoV-2 infection susceptibility. Furthermore, antibody waning may have led to the misclassification of some previously infected individuals as seronegative.
The extended time frame of sample collection may have introduced heterogeneity due to the emergence of SARS-CoV-2 variants which may have affected antibody detection. However, this variability was more pronounced in the SARS-CoV-2 Spike (S) protein. In our study, serological detection was based on antibodies to the NCP, which is more conserved across variants, thereby minimizing potential bias related to antigenic variation. The study also did not quantify SARS-CoV-2 IgG antibody levels in participant plasma. Additionally, using seropositivity as a proxy for infection may exclude individuals who succumbed to COVID-19 prior to serological sampling, potentially introducing bias toward non-severe cases. However, the primary aim of this study was to assess genetic associations with susceptibility to SARS-CoV-2 infection, rather than disease severity. Future research that includes hospitalized patients with severe disease could provide further insights into the role of ACE2 polymorphisms in COVID-19 severity, thereby enhancing our understanding of the genetic basis of the disease in Africa.
Recent advancements integrating COVID-19 GWAS with single-cell RNA sequencing (scRNA-seq) data have provided deeper insights into the immune cell types and critical genes associated with SARS-CoV-2 infection. Tools such as scPagwas (Ma et al., 2023) and scDRS (Zhang et al., 2022) have proven powerful in identifying immune pathways involved in the disease pathogenesis. These methodologies offer significant potential for investigating COVID-19 in populations like the Ghanaian cohort studied in this research. Future studies applying this approach could help further elucidate the complex immune interactions and gene-environment factors that influence COVID-19 susceptibility and disease outcomes in African populations.
Conclusion
The study concludes that ACE2 rs228566 but not rs2106809 is associated with an individual’s susceptibility to SARS-CoV-2 infection among the Ghanaian female population. The observed sex-specific association for the ACE2 rs2285666 T allele highlights the importance of considering host genetics and immunological differences in understanding SARS-CoV-2 vulnerability. While further research is needed to clarify the underlying mechanisms and to validate these findings across diverse populations, the results underscore the potential for incorporating genetic markers into clinical risk stratification models and public health planning. In resource-limited settings like Ghana, integrating host genetic data into national pandemic preparedness plans could enhance resource management by prioritizing vaccination deployments to vulnerable populations. This in turn could facilitate tailored public health responses through enhanced preventive measures, specialized care pathways, or design of sentinel surveillance systems for susceptible individuals. These findings support the expansion of population-based genomic surveillance to enable precision public health responses in low resource settings and beyond.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding authors.
Ethics statement
The study obtained ethical approval from the Committee of Human Research Publication and Ethics (CHPRE) at the School of Medical Sciences (SMS) at the Kwame Nkrumah University of Science and Technology (KNUST) in Kumasi, Ghana (Reference No: CHRPE/AP/218/20; date of approval: 01 February 2022) and the Ethical Commission of the Ärztekammer Hamburg, Germany (2020–10035-BO). The studies were conducted in accordance with the local legislation and institutional requirements. All data and samples used in this study was obtained from individuals who demonstrated their voluntariness to participate in the study with a signed or thumb-printed informed consent or assent form. Written informed consent for participation in this study was provided by the participants’ legal guardians/next of kin.
Author contributions
AOB: Data curation, Formal Analysis, Investigation, Methodology, Validation, Writing – original draft, Writing – review and editing. CO: Conceptualization, Data curation, Methodology, Project administration, Resources, Supervision, Validation, Writing – review and editing. AA-A: Conceptualization, Funding acquisition, Methodology, Project administration, Resources, Supervision, Validation, Writing – review and editing. EA: Data curation, Methodology, Validation, Writing – review and editing, Investigation. DW: Validation, Writing – review and editing, Investigation. EB: Data curation, Writing – review and editing, Investigation. HA: Data curation, Writing – review and editing, Investigation. SN: Data curation, Writing – review and editing, Investigation. NA: Data curation, Writing – review and editing, Investigation. JD: Data curation, Writing – review and editing, Investigation. WD: Data curation, Writing – review and editing, Investigation. LA-A: Data curation, Writing – review and editing, Investigation. S-KA: Data curation, Writing – review and editing, Investigation. MG: Data curation, Writing – review and editing, Investigation. GA: Data curation, Writing – review and editing, Investigation. AA-A: Data curation, Writing - review and editing, Investigation. WL: Methodology, Resources, Writing – review and editing. JK: Methodology, Resources, Writing – review and editing. YS: Methodology, Resources, Writing – review and editing. EO-M: Methodology, Resources, Writing – review and editing. AY: Methodology, Resources, Writing – review and editing. SA: Methodology, Resources, Writing – review and editing. RO: Data curation, Writing – review and editing, Investigation. AA-A: Data curation, Writing – review and editing, Investigation. EL: Investigation, Methodology, Resources, Writing – review and editing. AS: Methodology, Resources, Writing – review and editing. OM-A: Resources, Writing – review and editing. JM: Conceptualization, Funding acquisition, Project administration, Resources, Writing–review and editing. NS: Conceptualization, Validation, Project administration, Resources, Supervision, Writing–review and editing. JA: Conceptualization, Funding acquisition, Project administration, Resources, Supervision, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This sub-study did not receive any external funding. However, the main study, from which SARS-CoV-2 IgG seroprevalence data was derived, benefited from an open access funding supported by the German Ministry of Health (BMG), grant number ZMVI1-252oCOR001 and Ghana COVID-19 National Trust Fund, grant number NTD/COVID-19/19/F/F/006.
Acknowledgments
The authors gratefully acknowledge the support provided by the main study, which served as the foundation for this research. We extend our special thanks to the participants of the main study who willingly contributed and to the dedicated field staff who facilitated data collection and blood sample collection. Their contributions were invaluable to the success of this research.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2025.1555515/full#supplementary-material
References
Adimulam, T., Arumugam, T., Gokul, A., and Ramsuran, V. (2023). Genetic variants within SARS-CoV-2 human receptor genes may contribute to variable disease outcomes in different ethnicities. Int. J. Mol. Sci. 24, 8711. doi:10.3390/ijms24108711
Aschard, H., Hancock, D. B., London, S. J., and Kraft, P. (2010). Genome-wide meta-analysis of joint tests for genetic and gene-environment interaction effects. Hum. Hered. 70 (4), 292–300. doi:10.1159/000323318
Baris, I., Etlik, O., Koksal, V., Ocak, Z., and Baris, S. T. (2013). SYBR green dye-based probe-free SNP genotyping: introduction of T-Plex real-time PCR assay. Anal. Biochem. 441 (2), 225–231. doi:10.1016/j.ab.2013.07.007
Betsch, C., Wieler, L. H., and Habersaat, K.COSMO group (2020). Monitoring behavioural insights related to COVID-19. Lancet 395 (10232), 1255–1256. doi:10.1016/S0140-6736(20)30729-7
Bialek, S., Bowen, V., Chow, N., Curns, A., Gierke, R., Hall, A., et al. (2020). Geographic differences in COVID-19 cases, deaths, and incidence — United States, february 12–april 7, 2020. MMWR Morb. Mortal. Wkly. Rep. 69 (15), 465–471. doi:10.15585/mmwr.mm6915e4
Bosso, M., Thanaraj, T. A., Abu-Farha, M., Alanbaei, M., Abubaker, J., and Al-Mulla, F. (2020). The two faces of ACE2: the role of ACE2 receptor and its polymorphisms in hypertension and COVID-19. Mol. Ther. Methods Clin. Dev. 18, 321–327. doi:10.1016/j.omtm.2020.06.017
Cafiero, C., Rosapepe, F., Palmirotta, R., Re, A., Ottaiano, M. P., Benincasa, G., et al. (2021). Angiotensin system polymorphisms’ in SARS-CoV-2 positive patients: assessment between symptomatic and asymptomatic patients: a pilot study. Pharmgenomics Pers. Med. 14, 621–629. doi:10.2147/PGPM.S303666
Campbell, M. C., and Tishkoff, S. A. (2008). African genetic diversity: implications for human demographic history, modern human origins, and complex disease mapping. Annu. Rev. Genomics Hum. Genet. 9, 403–433. doi:10.1146/annurev.genom.9.081307.164258
Chaoxin, J. S. D. H. Y. G. R. W. C. T. Y., Daili, S., Yanxin, H., Ruwei, G., Chenlong, W., and Yaobin, T. (2013). The influence of angiotensin-converting enzyme 2 gene polymorphisms on type 2 diabetes mellitus and coronary heart disease. Eur. Rev. Med. Pharmacol. Sci. 17 (19), 2654–2659.
Chen, Y. Y., Zhang, P., Zhou, X. M., Liu, D., Zhong, J. C., Zhang, C. J., et al. (2018). Relationship between genetic variants of ACE2 gene and circulating levels of ACE2 and its metabolites. J. Clin. Pharm. Ther. 43 (2), 189–195. doi:10.1111/jcpt.12625
Chinazzi, M., Davis, J. T., Ajelli, M., Gioannini, C., Litvinova, M., Merler, S., et al. (2020). The effect of travel restrictions on the spread of the 2019 novel coronavirus (COVID-19) outbreak. Science 368 (6489), 395–400. doi:10.1126/science.aba9757
Conti, P., and Younes, A. (2020). Coronavirus COV-19/SARS-CoV-2 affects women less than men: clinical response to viral infection. J. Biol. Regul. Homeost. Agents 34 (2), 339–343. doi:10.23812/Editorial-Conti-3
Courtemanche, C., Garuccio, J., Le, A., Pinkston, J., and Yelowitz, A. (2020). Strong social distancing measures in the United States reduced the COVID-19 growth rate. Health Aff. (Millwood) 39 (7), 1237–1246. doi:10.1377/hlthaff.2020.00608
COVID-19 (2024). COVID-19 Updates | Ghana. Available online at: https://www.ghs.gov.gh/covid19/.
De, A., Tiwari, A., Dash, M., and Sinha, A. (2021). ACE2 mutation might explain lower COVID-19 burden in malaria endemic areas. Hum. Cell 34 (2), 702–705. doi:10.1007/s13577-021-00489-0
Deschermeier, C., Ehmen, C., von Possel, R., Murawski, C., Rushton, B., Amuasi, J., et al. (2022). Fcγ-receptor-based enzyme-linked immunosorbent assays for sensitive, specific, and persistent detection of anti-SARS-CoV-2 nucleocapsid protein IgG antibodies in human sera. J. Clin. Microbiol. 60 (6), e0007522. doi:10.1128/jcm.00075-22
De Silva, E., and Stumpf, M. P. H. (2004). HIV and the CCR5-Delta32 resistance allele. FEMS Microbiol. Lett. 241 (1), 1–12. doi:10.1016/j.femsle.2004.09.040
Dhangadamajhi, G., Mohapatra, B. N., Kar, S. K., and Ranjit, M. (2010). Gene polymorphisms in angiotensin I converting enzyme (ACE I/D) and angiotensin II converting enzyme (ACE2 C → T) protect against cerebral malaria in Indian adults. Infect. Genet. Evol. 10 (2), 337–341. doi:10.1016/j.meegid.2010.01.009
Do Nascimento, I. J. B., Cacic, N., Abdulazeem, H. M., von Groote, T. C., Jayarajah, U., Weerasekara, I., et al. (2020). Novel coronavirus infection (COVID-19) in humans: a scoping review and meta-analysis. J. Clin. Med. 9 (4), 941. doi:10.3390/jcm9040941
Dorn, A. van, Cooney, R. E., and Sabin, M. L. (2020). COVID-19 exacerbating inequalities in the US. Lancet 395 (10232), 1243–1244. doi:10.1016/S0140-6736(20)30893-X
Ellinghaus, D., Degenhardt, F., Bujanda, L., Buti, M., Albillos, A., Invernizzi, P., et al. (2020). Genomewide association study of severe covid-19 with respiratory failure. N. Engl. J. Med. 383 (16), 1522–1534. doi:10.1056/NEJMoa2020283
Emmerich, P., Murawski, C., Ehmen, C., von Possel, R., Pekarek, N., Oestereich, L., et al. (2021). Limited specificity of commercially available SARS-CoV-2 IgG ELISAs in serum samples of African origin. Trop. Med. & Int. Health 26 (6), 621–631. doi:10.1111/tmi.13569
Eshetie, S., Jullian, P., Benyamin, B., and Lee, S. H. (2023). Host genetic determinants of COVID-19 susceptibility and severity: a systematic review and meta-analysis. Rev. Med. Virol. 33 (5), e2466. doi:10.1002/rmv.2466
Etlik, O., Koksal, V., Tugba Arican-Baris, S., and Baris, I. (2008). An improved tetra-primer PCR approach for the detection of the FGFR3 G380R mutation responsible for achondroplasia. Mol. Cell Probes 22 (2), 71–75. doi:10.1016/j.mcp.2007.06.002
Ferrario, C. M. (2006). Role of angiotensin II in cardiovascular disease therapeutic implications of more than a century of research. J. Renin Angiotensin Aldosterone Syst. 7 (1), 3–14. doi:10.3317/jraas.2006.003
Ghosh, S., and Klein, R. S. (2017). Sex drives dimorphic immune responses to viral infections. J. Immunol. 198 (5), 1782–1790. doi:10.4049/jimmunol.1601166
GHS (2023). COVID-19 Updates | Ghana. Available online at: https://www.ghs.gov.gh/covid19/archive.php#.
Gray, I. C., Campbell, D. A., and Spurr, N. K. (2000). Single nucleotide polymorphisms as tools in human genetics. Hum. Mol. Genet. 9 (16), 2403–2408. doi:10.1093/hmg/9.16.2403
GTEx Portal (2025). GTEx Portal. Available online at: https://www.gtexportal.org/home/snp/rs2106809.
Guan, W. jie, Liang, W. hua, Zhao, Y., Liang, H. rui, Chen, Z. sheng, Li, Y. min, et al. (2020). Comorbidity and its impact on 1590 patients with COVID-19 in China: a nationwide analysis. Eur. Respir. J. 55 (5), 2000547. doi:10.1183/13993003.00547-2020
Hoffmann, M., Kleine-Weber, H., Schroeder, S., Krüger, N., Herrler, T., Erichsen, S., et al. (2020). SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 181 (2), 271–280. doi:10.1016/j.cell.2020.02.052
Karakaş Çelik, S., Çakmak Genç, G., Pişkin, N., Açikgöz, B., Altinsoy, B., Kurucu, İ. B., et al. (2021). Polymorphisms of ACE (I/D) and ACE2 receptor gene (Rs2106809, Rs2285666) are not related to the clinical course of COVID-19: a case study. J. Med. Virol. 93 (10), 5947–5952. doi:10.1002/jmv.27160
Kasela, S., Daniloski, Z., Bollepalli, S., Jordan, T. X., tenOever, B. R., Sanjana, N. E., et al. (2021). Integrative approach identifies SLC6A20 and CXCR6 as putative causal genes for the COVID-19 GWAS signal in the 3p21.31 locus. Genome Biol. 22 (1), 242. doi:10.1186/s13059-021-02454-4
Kellam, P., and Barclay, W. (2020). The dynamics of humoral immune responses following SARS-CoV-2 infection and the potential for reinfection. J. Gen. Virol. 101 (8), 791–797. doi:10.1099/jgv.0.001439
Letko, M., Marzi, A., and Munster, V. (2020). Functional assessment of cell entry and receptor usage for SARS-CoV-2 and other lineage B betacoronaviruses. Nat. Microbiol. 5 (4), 562–569. doi:10.1038/s41564-020-0688-y
Lewer, D., Braithwaite, I., Bullock, M., Eyre, M. T., White, P. J., Aldridge, R. W., et al. (2020). COVID-19 among people experiencing homelessness in England: a modelling study. Lancet Respir. Med. 8 (12), 1181–1191. doi:10.1016/S2213-2600(20)30396-9
Liu, D., Chen, Y., Zhang, P., Zhong, J., Jin, L., Zhang, C., et al. (2016). Association between circulating levels of ACE2-Ang-(1-7)-MAS axis and ACE2 gene polymorphisms in hypertensive patients. Med. (United States) 95 (24), e3876. doi:10.1097/MD.0000000000003876
Lorenz, E., Souares, A., Amuasi, J., Loag, W., Deschermeier, C., Fusco, D., et al. (2021). Seroprevalence of SARS-CoV-2 in urban settings in three sub-Saharan African countries (SeroCoV): a study protocol for a household-based cross-sectional prevalence study using two-stage cluster sampling. BMJ Open 11 (12), e056853. doi:10.1136/bmjopen-2021-056853
Lott, N., Gebhard, C. C. E., Bengs, S., Haider, A., Kuster, G. M., Regitz-Zagrosek, V., et al. (2023). Sex hormones in SARS-CoV-2 susceptibility: key players or confounders? Nat. Rev. Endocrinol. 19 (4), 217–231. doi:10.1038/s41574-022-00780-6
Ma, Y., Deng, C., Zhou, Y., Zhang, Y., Qiu, F., Jiang, D., et al. (2023). Polygenic regression uncovers trait-relevant cellular contexts through pathway activation transformation of single-cell RNA sequencing data. Cell Genomics 3 (9), 100383.doi:10.1016/j.xgen.2023.100383
Mahmood, Z. S., Fadhil, H. Y., Abdul Hussein, T. A., and Ad’hiah, A. H. (2022). Severity of coronavirus disease 19: profile of inflammatory markers and ACE (rs4646994) and ACE2 (rs2285666) gene polymorphisms in Iraqi patients. Meta Gene 31, 101014. doi:10.1016/j.mgene.2022.101014
Martínez-Gómez, L. E., Herrera-López, B., Martinez-Armenta, C., Ortega-Peña, S., Camacho-Rea, M. del C., Suarez-Ahedo, C., et al. (2022). ACE and ACE2 gene variants are associated with severe outcomes of COVID-19 in men. Front. Immunol. 13, 812940. doi:10.3389/fimmu.2022.812940
Martinson, J. J., Chapman, N. H., Rees, D. C., Liu, Y. T., and Clegg, J. B. (1997). Global distribution of the CCR5 gene 32-basepair deletion. Nat. Genet. 16 (1), 100–103. doi:10.1038/ng0597-100
Medina-Enríquez, M. M., Lopez-León, S., Carlos-Escalante, J. A., Aponte-Torres, Z., Cuapio, A., and Wegman-Ostrosky, T. (2020). ACE2: the molecular doorway to SARS-CoV-2. Cell & Biosci. 10 (1), 1–17. doi:10.1186/s13578-020-00519-8
Mohammadi-Berenjestanaki, H., Mohammadali, E., Khasayesi, M., Rafiei, A., Kashi, Z., Mirzaei-Ilali, N., et al. (2023). Association between angiotensin-converting enzyme-2 gene polymorphism (rs2106809) with severity and outcome of COVID-19 infection. Mol. Biol. Rep. 50 (8), 6669–6679. doi:10.1007/s11033-023-08493-3
Möhlendick, B., Schönfelder, K., Breuckmann, K., Elsner, C., Babel, N., Balfanz, P., et al. (2021). ACE2 polymorphism and susceptibility for SARS-CoV-2 infection and severity of COVID-19. Pharmacogenet Genomics. 31, 165–171. doi:10.1097/fpc.0000000000000436
Niemi, M. E. K., Karjalainen, J., Liao, R. G., Neale, B. M., Daly, M., Ganna, A., et al. (2021). Mapping the human genetic architecture of COVID-19. Nature 600 (7889), 472–477. doi:10.1038/s41586-021-03767-x
Oran, D. P., and Topol, E. J. (2020). Prevalence of asymptomatic SARS-CoV-2 infection: a narrative review. Ann. Intern Med. 173 (5), 362–367. doi:10.7326/M20-3012
Pairo-Castineira, E., Clohisey, S., Klaric, L., Bretherick, A. D., Rawlik, K., Pasko, D., et al. (2020). Genetic mechanisms of critical illness in COVID-19. Nature 591 (7848), 92–98. doi:10.1038/s41586-020-03065-y
Pairo-Castineira, E., Rawlik, K., Bretherick, A. D., Qi, T., Wu, Y., Nassiri, I., et al. (2023). GWAS and meta-analysis identifies 49 genetic variants underlying critical COVID-19. Nature 617 (7962), 764–768. doi:10.1038/s41586-023-06034-3
Petersen, D. C., Steyl, C., Scholtz, D., Baker, B., Abdullah, I., Uren, C., et al. (2022). African genetic representation in the context of SARS-CoV-2 infection and COVID-19 severity. Front. Genet. 13, 909117. doi:10.3389/fgene.2022.909117
Price, A. L., Zaitlen, N. A., Reich, D., and Patterson, N. (2010). New approaches to population stratification in genome-wide association studies. Nat. Rev. Genet. 11 (7), 459–463. doi:10.1038/nrg2813
Rostami, A., Sepidarkish, M., Fazlzadeh, A., Mokdad, A. H., Sattarnezhad, A., Esfandyari, S., et al. (2021). Update on SARS-CoV-2 seroprevalence: regional and worldwide. Clin. Microbiol. Infect. 27 (12), 1762–1771. doi:10.1016/j.cmi.2021.09.019
Sabater Molina, M., Nicolás Rocamora, E., Bendicho, A. I., Vázquez, E. G., Zorio, E., Rodriguez, F. D., et al. (2022). Polymorphisms in ACE, ACE2, AGTR1 genes and severity of COVID-19 disease. PLoS One 17 (2), e0263140. doi:10.1371/journal.pone.0263140
Samson, M., Libert, F., Doranz, B. J., Rucker, J., Liesnard, C., Farber, M., et al. (1996). Resistance to HIV-1 infection in Caucasian individuals bearing mutant alleles of the CCR-5 chemokine receptor gene. Nature 382 (6593), 722–725. doi:10.1038/382722a0
Schlebusch, C. M., Skoglund, P., Sjödin, P., Gattepaille, L. M., Hernandez, D., Jay, F., et al. (2012). Genomic variation in seven khoe-san groups reveals adaptation and complex african history. Science. 338 (6105), 374–379. doi:10.1126/science.1227721
Schmitz, H., Gabriel, M., and Emmerich, P. (2011). Specific detection of antibodies to different flaviviruses using a new immune complex ELISA. Med. Microbiol. Immunol. 200 (4), 233–239. doi:10.1007/s00430-011-0195-0
Srivastava, A., Bandopadhyay, A., Das, D., Pandey, R. K., Singh, V., Khanam, N., et al. (2020). Genetic association of ACE2 rs2285666 polymorphism with COVID-19 spatial distribution in India. Front. Genet. 11, 564741. doi:10.3389/fgene.2020.564741
Statement on the fifteenth meeting of the IHR (2005). Emergency committee on the COVID-19 pandemic. Available online at: https://www.who.int/news/item/05-05-2023-statement-on-the-fifteenth-meeting-of-the-international-health-regulations-%282005%29-emergency-committee-regarding-the-coronavirus-disease-%28covid-19%29-pandemic.
Stringer, C. (2016). The origin and evolution of Homo sapiens. Philosophical Trans. R. Soc. B Biol. Sci. 371 (1698), 20150237. doi:10.1098/rstb.2015.0237
Struck, N. S., Lorenz, E., Deschermeier, C., Eibach, D., Kettenbeil, J., Loag, W., et al. (2022). High seroprevalence of SARS-CoV-2 in Burkina-Faso, Ghana and Madagascar in 2021: a population-based study. BMC Public Health 22 (1), 1676–1679. doi:10.1186/s12889-022-13918-y
Tanha, H. M., Naeini, M. M., Rahgozar, S., Rasa, S. M. M., and Vallian, S. (2015). Modified tetra-primer ARMS PCR as a single-nucleotide polymorphism genotyping tool. Genet. Test. Mol. Biomarkers 19 (3), 156–161. doi:10.1089/gtmb.2014.0289
The COVID-19 Sex-Disaggregated Data Tracker (2025). Global health. Available online at: https://globalhealth5050.org/the-sex-gender-and-covid-19-project/the-data-tracker/?explore=variable&variable=Confirmed+cases.50/50
Vahidy, F. S., Pan, A. P., Ahnstedt, H., Munshi, Y., Choi, H. A., Tiruneh, Y., et al. (2021). Sex differences in susceptibility, severity, and outcomes of coronavirus disease 2019: cross-sectional analysis from a diverse US metropolitan area. PLoS One 16 (1), e0245556. doi:10.1371/journal.pone.0245556
Visscher, P. M., Wray, N. R., Zhang, Q., Sklar, P., McCarthy, M. I., Brown, M. A., et al. (2017). 10 Years of GWAS discovery: biology, function, and translation. Am. J. Hum. Genet. 101 (1), 5–22. doi:10.1016/j.ajhg.2017.06.005
Wan, Y., Shang, J., Graham, R., Baric, R. S., and Li, F. (2020). Receptor recognition by the novel coronavirus from wuhan: an analysis based on decade-long structural studies of SARS coronavirus. J. Virol. 94 (7), 001277-20. doi:10.1128/JVI.00127-20
WHO (2020a). Population-based age-stratified seroepidemiological investigation protocol for coronavirus 2019 (COVID-19) infection, 2019.
WHO (2020b). WHO Director-General’s opening remarks at the media briefing on COVID-19 - 11 March 2020. Available online at: https://www.who.int/director-general/speeches/detail/who-director-general-s-opening-remarks-at-the-media-briefing-on-covid-19---11-march-2020.
WHO (2023a). WHO coronavirus (COVID-19) dashboard | WHO coronavirus (COVID-19) dashboard with vaccination data. Available online at: https://covid19.who.int/.
Yancy, C. W. (2020). COVID-19 and african Americans. JAMA 323 (19), 1891–1892. doi:10.1001/jama.2020.6548
Yeung, M. L., Teng, J. L. L., Jia, L., Zhang, C., Huang, C., Cai, J. P., et al. (2021). Soluble ACE2-mediated cell entry of SARS-CoV-2 via interaction with proteins related to the renin-angiotensin system. Cell 184 (8), 2212–2228.e12. doi:10.1016/j.cell.2021.02.053
Yu, N., Chen, F. C., Ota, S., Jorde, L. B., Pamilo, P., Patthy, L., et al. (2002). Larger genetic differences within africans than between africans and eurasians. Genetics 161 (1), 269–274. doi:10.1093/genetics/161.1.269
Zhang, M. J., Hou, K., Dey, K. K., Sakaue, S., Jagadeesh, K. A., Weinand, K., et al. (2022). Polygenic enrichment distinguishes disease associations of individual cells in single-cell RNA-seq data. Nat. Genet. 54 (10), 1572–1580. doi:10.1038/s41588-022-01167-z
Zhao, H., Lu, X., Deng, Y., Tang, Y., and Lu, J. (2020). COVID-19: asymptomatic carrier transmission is an underestimated problem. Epidemiol. Infect. 148, e116. doi:10.1017/S0950268820001235
Zhou, T., Tsybovsky, Y., Gorman, J., Rapp, M., Cerutti, G., Chuang, G. Y., et al. (2020). Cryo-EM structures of SARS-CoV-2 spike without and with ACE2 reveal a pH-dependent switch to mediate endosomal positioning of receptor-binding domains. Cell Host Microbe 28 (6), 867–879. doi:10.1016/j.chom.2020.11.004
Keywords: angiotensin-converting enzyme 2 (ACE2), single nucleotide polymorphism (SNP), SARS-CoV-2 susceptibility, genetic epidemiological study, SARS-CoV-2 infection (COVID-19), candidate gene association study (CGAS)
Citation: Boakye AO, Obirikorang C, Afum-Adjei Awuah A, Adu EA, Winter D, Boham EE, Alani H, Newton SK, Almoustapha NST, Deke J, Dzadey WO, Adu-Amoah L, Kroduah S-A, Grant MA, Asare G, Amoako-Adusei A, Loag W, Kettenbeil J, Sarkodie YA, Oduro-Mensah E, Yawson AE, Apanga S, Odotei Adjei R, Adobasom-Anane AG, Lorenz E, Souares A, Maiga-Ascofaré O, May J, Struck NS and Amuasi JH (2025) Genetic association of ACE2 rs2285666 (C>T) and rs2106809 (A>G) and susceptibility to SARS-CoV-2 infection among the Ghanaian population. Front. Genet. 16:1555515. doi: 10.3389/fgene.2025.1555515
Received: 04 January 2025; Accepted: 07 May 2025;
Published: 26 May 2025.
Edited by:
Farren Briggs, University of Miami, United StatesReviewed by:
Yunlong Ma, University of Pennsylvania, United StatesYiran Shen, University of Michigan, United States
Copyright © 2025 Boakye, Obirikorang, Afum-Adjei Awuah, Adu, Winter, Boham, Alani, Newton, Almoustapha, Deke, Dzadey, Adu-Amoah, Kroduah, Grant, Asare, Amoako-Adusei, Loag, Kettenbeil, Sarkodie, Oduro-Mensah, Yawson, Apanga, Odotei Adjei, Adobasom-Anane, Lorenz, Souares, Maiga-Ascofaré, May, Struck and Amuasi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nicole S. Struck, bmljb2xlLmdpbGJlcmdlckBibml0bS5kZQ==; John Humphery Amuasi, YW11YXNpQGtjY3IuZGU=
†These authors have contributed equally to this work