 Florian R. Kirchner1
Florian R. Kirchner1 Katharina Littringer2
Katharina Littringer2 Simon Altmeier1
Simon Altmeier1 Van Du T. Tran3Franziska Schönherr1
Van Du T. Tran3Franziska Schönherr1 Christina Lemberg1Marco Pagni3
Christina Lemberg1Marco Pagni3 Dominique Sanglard4
Dominique Sanglard4 Nicole Joller2
Nicole Joller2 Salomé LeibundGut-Landmann1*
Salomé LeibundGut-Landmann1*- 1Section of Immunology, Vetsuisse Faculty, University of Zurich, Zurich, Switzerland
- 2Institute of Experimental Immunology, University of Zurich, Zurich, Switzerland
- 3Vital-IT Group, Swiss Institute of Bioinformatics, Lausanne, Switzerland
- 4Institute of Microbiology, University of Lausanne and University Hospital Center, Lausanne, Switzerland
Controlled immune activation in response to commensal microbes is critical for the maintenance of stable colonization and prevention of microbial overgrowth on epithelial surfaces. Our understanding of the host mechanisms that regulate bacterial commensalism has increased substantially, however, much less data exist regarding host responses to members of the fungal microbiota on colonized surfaces. Using a murine model of oropharyngeal candidiasis, we have recently shown that differences in immune activation in response to diverse natural isolates of Candida albicans are associated with different outcomes of the host-fungal interaction. Here we applied a genome-wide transcriptomic approach to show that rapid induction of a strong inflammatory response characterized by neutrophil-associated genes upon C. albicans colonization inversely correlated with the ability of the fungus to persist in the oral mucosa. Surprisingly, persistent fungal isolates showed no signs of a compensatory regulatory immune response. By combining RNA-seq data, genetic mouse models, and co-infection experiments, we show that attenuation of the inflammatory response at the onset of infection with a persistent isolate is not a consequence of enhanced immunosuppression. Importantly, depletion of regulatory T cells or deletion of the immunoregulatory cytokine IL-10 did not alter host-protective type 17 immunity nor did it impair fungal survival in the oral mucosa, indicating that persistence of C. albicans in the oral mucosa is not a consequence of suppressed antifungal immunity.
Introduction
Opportunistic infections with fungi are an increasing cause of morbidity and mortality worldwide. C. albicans is one of the most important disease-causing fungi in humans. It is found as a commensal in the human gastrointestinal and genital tracts with a large proportion of healthy individuals being carriers, but it may become pathogenic under certain conditions. Disease symptoms range from mild to severe superficial infections of the oral and vaginal mucosae, the skin and the nails, affecting millions of people worldwide (1–5). More rarely, C. albicans causes systemic diseases associated with high mortality rates (6). The development of C. albicans mucosal infections is mainly attributed either to defects in host cellular immunity, including those resulting from primary or secondary immunodeficiency, or to changes in the normal microbiota that may be caused by antibiotic treatment (3, 5). Defects in epithelial barrier integrity are also associated with infections, highlighting the importance of intact epithelial function for preventing fungal entry into the tissue (7). In addition to host factors, C. albicans infections may be favored by increased expression of virulence attributes of the fungus. Genetic variations within the species of C. albicans that result in phenotypic diversity within the fungus have been found to modulate its pathogenicity at epithelial surfaces and systemically (8–12). The decision between C. albicans commensalism and disease is thus the result of a fine balance between fungal virulence and host defense mechanisms.
The experimental model of oropharyngeal candidiasis (OPC) in mice has been widely used to study the interaction of C. albicans with the host at mucosal surfaces in vivo (13). It allowed unraveling antifungal immune mechanisms against C. albicans, such as the interleukin 17 (IL-17)-pathway (14), and to explore the pathogenicity of C. albicans mutants at the mucosal surface (15, 16).
Using the common laboratory strain SC5314 for infection in this model only partially reflects the situation of OPC in humans (where C. albicans is a commensal). In wild type mice, this highly virulent C. albicans isolate is rapidly cleared from the oral mucosa (17). However, we have recently shown that depending on the fungal isolate, the murine oral mucosa can be colonized with C. albicans for a prolonged period of time without immunosuppression of the host (12), thus mimicking the situation in humans.
Our initial investigations of the differences between acute and persistent OPC in mice have revealed major differences in the host response that was induced by different isolates of C. albicans (12). Most strikingly, the differential degree of inflammation that was triggered by virulent strains (such as strain SC5314) compared to persistent strains (such as strain 101) correlated with the differential outcome of infection (12). However, the basis of the differential host response triggered by different strains remains unclear.
Here, we applied an unbiased genome-wide approach to obtain a comprehensive view on the host response to two functionally divergent fungal strains and to assess differences in the host response that might modulate the balance between fungal persistence and rapid clearance. Moreover, we addressed whether persistent C. albicans had a propensity to trigger an immunosuppressive response in the host that might be responsible for the curbed immune activation and in turn allow prolonged colonization in the host.
Materials and Methods
Mice
Foxp3-GFP.KI x Il10-Thy1.1 reporter mice (18, 19), DEREG mice (20) and Il10−/− mice (21) were obtained from Vijay Kuchroo (Brigham and Women's Hospital, Harvard Medical School, Boston, MA, USA), Manfred Kopf (ETH Zurich, Switzerland), and the Swiss Immunological Mouse Repository, respectively and bred at the Laboratory Animal Service Center (University of Zürich, Switzerland). Wildtype (WT) C57BL/6J mice were purchased from Janvier Elevage. All mice were kept in specific pathogen-free conditions and used in sex- and age-matched groups at 6–15 weeks of age. DEREG mice were treated with 1 μg diphtheria toxin i.p. on day 11 and 13 post-infection. Where indicated, 0.125 mg anti-CD25 antibody (clone PC-61.5.3, BioXCell) or the corresponding isotype control were administered i.p. per mouse 7 days prior to infection.
Fungal Strains and OPC Infection Model
C. albicans strains SC5314 (22) and 101 (12) were grown in YPD medium at 30°C and 180 rpm for 15–18 h. Mice were infected sublingually with 2.5 × 106 C. albicans yeast cells as described (23), without immunosuppression. In co-infection experiments, mice were infected with 1.25 × 106 yeast cells of each strain, i.e., with a total of 2.5 × 106 yeast cells. For determination of the fungal burden, the tongue of euthanized animals was removed, homogenized in sterile 0.05% NP40 in H2O for 3 min at 25 Hz using a Tissue Lyzer (Qiagen) and serial dilutions were plated on YPD agar containing 100 μg/ml Ampicillin. Infection of mice with strain 101 resulted in detectable fungal loads in the tongue and in the feces for >60 days, in some mice >1 year.
Preparation of Tongue Epithelial Sheets and RNA Isolation
The tongue was cut in half to obtain the dorsal part, which was then freed from muscle tissue with a scalpel and floated on PBS containing 2.86 mg/ml dispase II (Roche) for 60 min with the epithelial side facing upwards to separate the epithelial tissue from the lamina propria. Epithelial sheets were incubated in RNAlater (Sigma-Aldrich) for 1 min immediately after isolation and then grinded in liquid N2. Three epithelial sheets were pooled for the generation of each RNA-seq replicate and two replicates were generated.
RNA isolation from epithelial sheets was done by combining two phase separations and DirectZol™ RNA MiniPrep kit (Zymo Research) as described (24). DNase treatment was performed off-column using the DNA-free™ Kit (Life Technologies). RNA integrity was determined using a 2100 Bioanalyzer system (Agilent Technologies) according to the manufacturer's instructions. Samples were only included in the study if the RNA integrity value (RIN) was above 7.5 and no obvious degradation was observed.
Preparation of cDNA Libraries and Sequencing
cDNA libraries were generated using the SureSelect multiplexed sequencing kit with strand-specific RNA library preparation for Illumina (Agilent Technologies) according to the manufacturer's instructions. In brief, mRNA was purified using poly(A) beads, fragmented and double-stranded cDNA with ligated adapters was generated. The library was amplified using primers that match the adapters. During this step, RNA-seq indexes were inserted into the libraries for multiplexing. The collected double-strand cDNA was then amplified and indexed in a separate PCR. RNA quality, fragment size, and cDNA concentration were determined using a fragment analyzer automated CE system (Advanced Analytical) and a Qubit fluorometer (Invitrogen).
cDNA libraries were subjected to cluster generation using the Illumina TruSeq PE cluster kit v3 reagents and sequenced on the Illumina HiSeq 2500 system with TruSeq SBS kit v3 reagents at the Lausanne Genomic Technologies Facility (LGTF).
RNA-Seq Data Analysis
RNA-seq purity-filtered reads were adapter- and quality-trimmed with cutadapt (v1.2.1) (25) and filtered for low complexity with seq_crumbs (v0.1.8) (https://bioinf.comav.upv.es/seq_crumbs). After alignment against the mouse genome GRCm38.p4 using STAR (v2.4.2a), htseq-count (v0.5.4p3) (26) was used to summarize the number of read counts per gene locus. Genes with counts fewer than one per million in all samples were removed from the statistical analysis, yielding 13,855 remaining genes. Data normalization and differential expression analysis were performed in R (v3.2.2) as follows. Normalization of read count data was performed with the edgeR package using the TMM (trimmed mean of M-values) method (27). Normalized read count data was transformed to log2-counts with the voom transformation (28). A linear model was applied to the transformed data using the limma package (29) on all conditions, all in duplicates. The contrasts representing the difference between the infected and naïve conditions were studied, including conditions at 9 h, 1 day, 3 days, 7 days post-infection with strains SC5314 and 101, respectively. P-values produced from the differential analysis were adjusted using Benjamini & Hochberg correction (30). Genes were considered to be differentially regulated if their expression was altered by a factor of at least 2-fold with adjusted p-values < 0.05 (false discovery rate, FDR). Gene Ontology (GO) enrichment analysis was done with the topGO package (version 2.32.0) using the fisher statistic and weight01 algorithm. This analysis was restricted to all GO terms that are offspring of GO:0002376 (immune system processes) and to genes that are differentially regulated in at least one contrast. An additional enrichment analysis was performed with the MetaCoreTM software (Thomson Reuters, https://clarivate.com/products/metacore) on all GO terms belonging to biological process. Heat maps and hierarchical clustering were generated with the Morpheus software (Broad Institute, https://software.broadinstitute.org/morpheus) using the distance of one minus Pearson correlation and the average linkage mode. The data analyzed here are accessible under the NCBI BioProject accession number PRJNA491801.
RNA Isolation From Total Tongue and Quantitative RT-PCR
Isolation of total RNA from bulk tongues was carried out according to standard protocols using TRI Reagent® (Sigma Aldrich). cDNA was generated by RevertAid reverse transcriptase (Thermo Fisher). Quantitative PCR was performed using SYBR Green (Roche) and a QuantStudio 7 Flex (Life Technologies) instrument. The primers used for qPCR are listed in Table S1. All qRT-PCR assays were performed in duplicate and the relative expression (rel. expr.) of each gene was determined after normalization to β-actin transcript levels.
Preparation of Lymph Node Cells for T Cell and Treg Analysis by Flow Cytometry
Cervical lymph nodes were removed and single cell suspensions were prepared by digestion with DNase I (2.4 mg/ml, Roche) and Collagenase I (2.4 mg/ml, Invitrogen) in PBS for 15 min at 37°C. For inducing cytokine production by primed T cells, 106 cervical lymph node cells were re-stimulated for 6 h with 1 × 105 MutuDC1940 cells (31) pulsed with 2.5 × 105/ml heat-killed C. albicans or left unpulsed. Brefeldin A (10 μg/ml, AppliChem) was added for the last 5 h to inhibit the secretory pathway. Intracellular IL-17 and IFN-γ production was then analyzed by flow cytometry.
Preparation of Tongue Cells for Flow Cytometry
For quantification of neutrophils, tongues were removed and digested with DNase I (2.4 mg/ml, Roche), and Collagenase IV (4.8 mg/ml) in PBS for 45 min at 37°C. For analysis of Tregs, mice were anesthetized with a sublethal dose of Ketamin (100 mg/kg), Xylazin (20 mg/kg), and Acepromazin (2.9 mg/kg) and perfused by injection of PBS into the right heart ventricle prior to removing the tongue. Tongues were cut in half and the underlying muscle tissue was carefully removed using a scalpel. The remaining tongue tissue was cut into small pieces and digested with DNase I (2.4 mg/ml, Roche), Collagenase IV (2.4 mg/ml) and in some case Trypsin (1 mg/ml) in PBS for 45 min at 37°C. Single cell suspensions were obtained by passing the digested tissue through a 70 μm strainer using icecold PBS supplemented with 1% FCS and 2 mM EDTA and then stained for flow cytometry.
Flow Cytometry
Single cell suspensions of tongue and lymph nodes were stained in PBS supplemented with 1% FSC, 5 mM EDTA, and 0.02% NaN3. LIVE/DEAD Near IR stain (Life Technologies) was used for exclusion of dead cells. The antibodies for surface and intracellular cytokine staining are listed in Table S2. For intracellular cytokine staining, cells were fixed and permeabilized using BD Cytofix/Cytoperm reagent (BD Biosciences) and subsequently incubated in Perm/Wash buffer (BD Biosciences). All extracellular and intracellular staining steps were carried out on ice.
For intranuclear Foxp3 staining, cells were fixed/permeabilized for 40 min at RT using the Foxp3 Staining Buffer Set (eBioscience) and subsequently stained for 40 min at RT in BD Perm/Wash buffer (BD Biosciences), Cells were acquired on a FACS Gallios (Beckman Coulter), a SP6800 Spectral Analyzer (Sony) or a BD LSRFortessa (BD Biosciences). The data were analyzed with FlowJo software (FlowJo LLC). The gating of the flow cytometric data was performed according to the guidelines for the use of flow cytometry and cell sorting in immunological studies (32), including pre-gating on viable and single cells for analysis. Absolute cell numbers of lymphoid cell populations were calculated based on a defined number of counting beads (BD Biosciences, Calibrite Beads), which were added to the samples before flow cytometric acquisition.
Histology
For histology, tissue was fixed in 4% PBS-buffered paraformaldehyde overnight and embedded in paraffin. Sagittal sections (9 μm) were stained with Periodic-acidic Schiff (PAS) reagent and counterstained with Haematoxilin and mounted with Pertex (Biosystem, Switzerland) according to standard protocols. Images were acquired with a digital slide scanner (NanoZoomer 2.0-HT, Hamamatsu) and analyzed with NDP.view2.
Results
The Host Response to C. albicans Strain 101 Is Delayed Compared to Strain SC5314
To obtain genome-wide information about the host response to C. albicans in the oral mucosa, we performed a transcriptomic analysis. We used the two functionally distinct C. albicans strains SC5314 and 101 to assess strain-specific differences between acute and persistent infection in mice at 9 h, 1 day, 3 days and 7 days post-infection. To enrich for host tissue in direct contact with the fungus, we isolated epithelial sheets from the tongue by dispase II-mediated digestion of the basal membrane. Epithelial sheets were strongly enriched for epithelial cells compared to the proportion found in the bulk tongue, but they also contained tissue-resident and infiltrated immune cells as evidenced by histology and flow cytometry (Figure S1).
Epithelial sheets were subjected to RNA isolation, cDNA library generation and sequencing. Analysis of the number of differentially expressed genes over time in comparison to uninfected conditions revealed a rapid onset of the transcriptional response to the virulent strain SC5314 between 9 and 24 h post-infection, whereas a slower kinetic was observed after infection with the persistent strain 101, which started rising after day 1 post-infection and increased continuously until day 7 post-infection (Figures 1A–C). Interestingly, in case of strain SC5314, the number of differentially expressed genes increased until day 3 post-infection (Figures 1A–C), even though the fungal burden is known to decline rapidly at this time point for this specific isolate (33). The genes differentially expressed on day 3 post-infection largely differed from those regulated earlier (Figure S2). Finally, by day 7, only very few genes were changed in comparison to uninfected controls (Figures 1A–C, Figure S2) indicating that homeostasis was rapidly restored after clearance of strain SC5314.
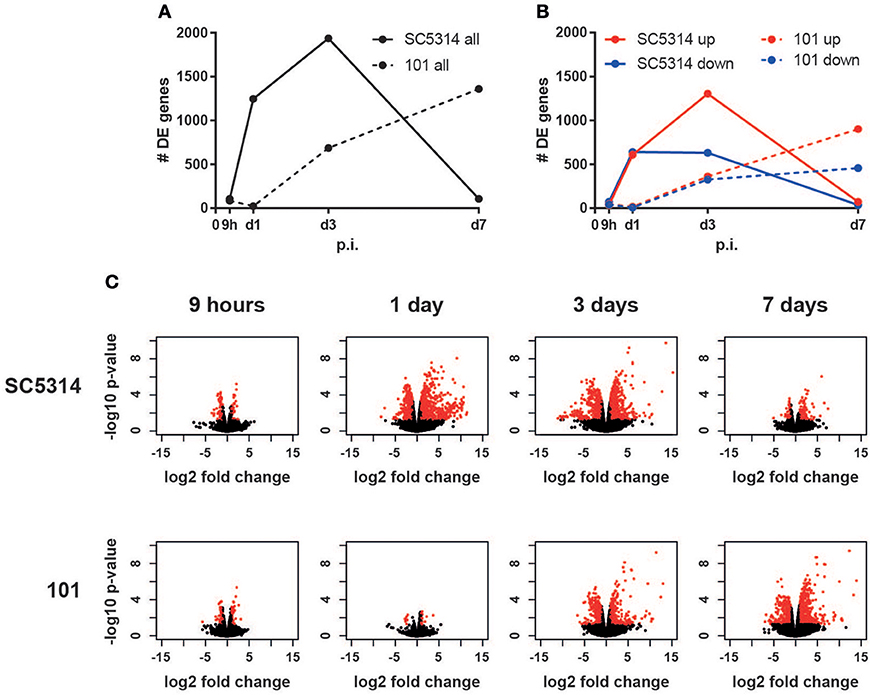
Figure 1. The host response to C. albicans strain 101 is delayed compared to strain SC5314. Epithelial sheet from C. albicans strain SC5314- and strain 101-infected WT mice were subjected to RNA-seq analysis. (A) Graph showing the total number of differentially expressed (DE) genes at the indicated time points compared to naïve controls. (B) Separate display of the up- and down-regulated genes at the indicated time points. (C) Volcano plots displaying the fold changes and the FDR of all genes detected in each condition separately. Genes with FDR < 0.05 and fold change < −2 or > 2 are marked in red.
Quantitative and Qualitative Differences in the Host Response to Virulent and Persistent OPC
Given the delayed response observed to the persistent strain 101 compared to the virulent strain SC5314, we were interested whether the two responses were qualitatively comparable, regardless of kinetics of the observed changes. While a large number of genes were selectively regulated by SC5314, there was also a considerable overlap between the two responses, whereby most co-regulated genes were delayed by one to two time points in case of strain 101 (Figure S2). Likewise, at the level of the regulated biological processes, as defined at the level of gene ontology, there was a good concordance of the response to the two fungal strains, again with slower kinetics and lower FDR in case of strain 101 (Figure S3). The processes that were most strongly induced early after infection comprised those linked to antimicrobial response and immunity, while on day 3 post-infection with strain SC5314, metabolic processes dominated. The processes that were most strongly repressed by both strains comprised those linked to development and negative regulation of signaling (Figure S3).
Among the immune system processes that were most significantly regulated by strain SC5314, some were found to have a comparably high FDR in the response to strain 101, including GO terms linked to chemotaxis and cell migration. This strain-specific difference was also reflected at the level of the genes that were annotated to these processes: genes encoding for the neutrophil chemokines CXCL1, CXCL2, and CXCL5 as well as IL-1β were strongly induced in response to SC5314 on day 1 post-infection, but very poorly in response to 101 at any time point analyzed (Figure 2). Even after 60 days of infection, when mice were still highly colonized with strain 101, expression of these factors remained as low as in uninfected controls (Figures S4A–B). This was consistent with the curbed recruitment of neutrophils to the tongue infected with strain 101 in comparison to strain SC5314 (12). In some mice, strain 101 persisted for over 1 year without causing inflammation or triggering neutrophil recruitment (Figures S4C–E).
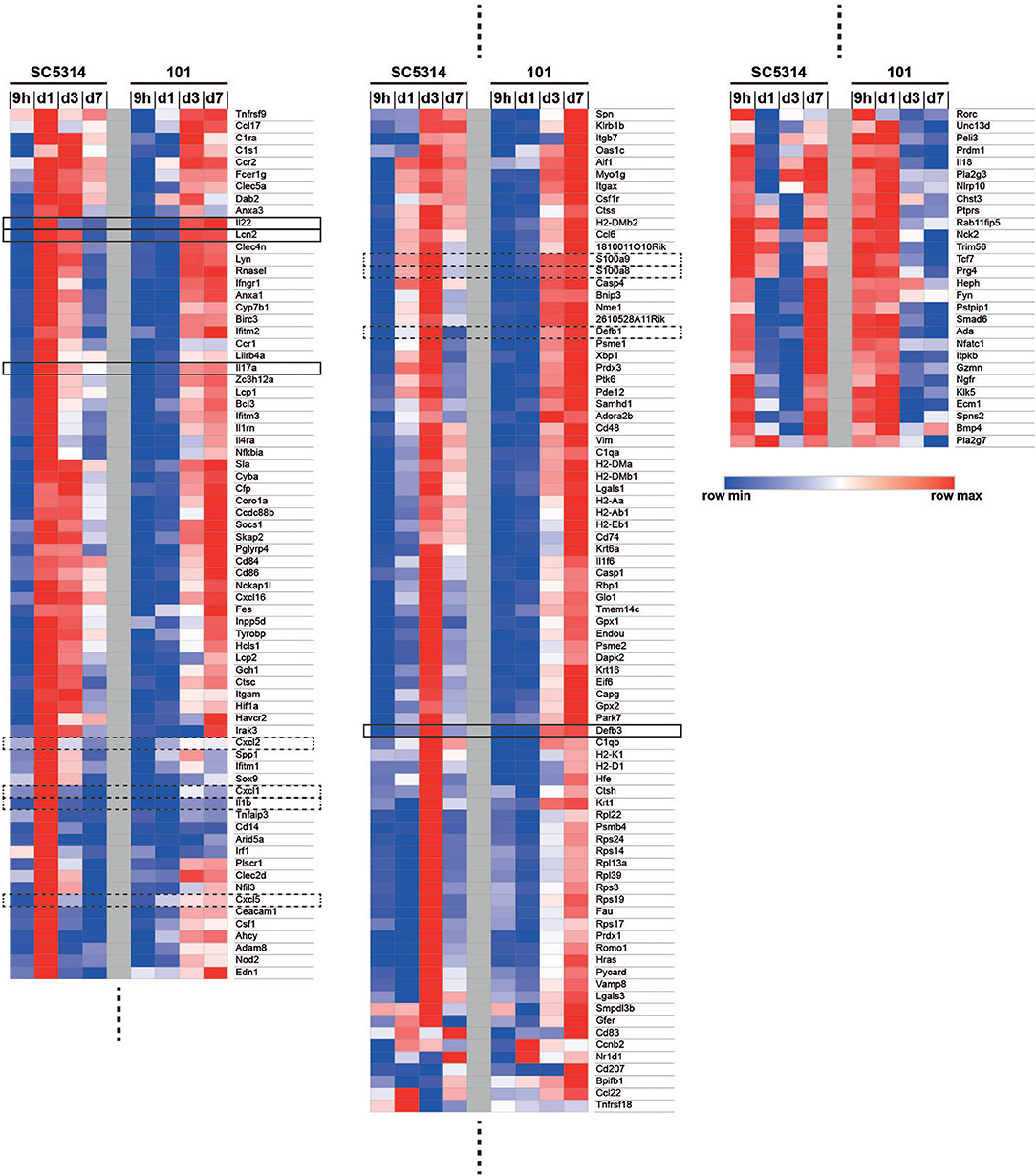
Figure 2. Co-regulated and differentially regulated genes in the mucosal response to strains 101 and SC5314. The GO term enrichment analysis on genes that are differentially regulated in at least one contrast for the 62 GO terms that are offspring of GO: 0002376 (immune system processes) with p < 0.05 yielded 182 genes. Genes were ordered by hierarchical clustering of their log2 fold changes, which are displayed in a heat map with a row-scaling color scheme.
Among the genes annotated to immune system processes that were significantly regulated (FDR < 0.05), we observed many that were regulated to a comparable degree by both fungal strains, including genes associated with the IL-17 pathway (Figure 2). Conserved induction of the IL-17 pathway by both fungal strains was also confirmed at the level of IL-17 target genes, including those coding for S100a8, S100a9, Lipocalin 2, β-defensin-3, and β-defensin-1. They reached highest expression levels on day 1–3 post-infection in case of strain SC5314, and on day 3–7 in case of strain 101, respectively (Figure 2). Defb3 and Defb1 genes were an exception to this pattern as their expression did not rise before day 3 in either condition (Figure 2). Overall, this confirmed what we previously observed in response to these two fungal strains (Figure S4F) (12).
C. albicans Strain 101 Does Not Actively Suppress the Host Response at the Onset of Infection
In search of an explanation for the delayed and more limited host response to strain 101 in comparison to strain SC5314, we examined whether expression of immune regulatory and immunosuppressive genes that might curb the inflammatory response predominated during infection with strain 101. Surprisingly, such genes were not found to be significantly induced in the RNA-seq data set by any of the two strains. Moreover, in a GO process enrichment analysis, processes associated with immune regulation such as “negative regulation of immune response” or “negative regulation of immune system processes” were not altered significantly. This was also consistent with the absence of an increase in IL-10 or TGF-β transcript levels by RT-qPCR in epithelial sheets (Figure 3A). This analysis may not have captured the full spectrum of the antifungal response, as the epithelial sheets used for RNA isolation and sequencing/RT-qPCR do not represent the full complexity of the mucosal immune system and some cells may have been lost during preparation of the epithelial sheets (and others might act at a distance). We therefore quantified IL-10 and TGF-β expression levels in the bulk tongue of 101-infected compared to naïve mice (Figure 3B).
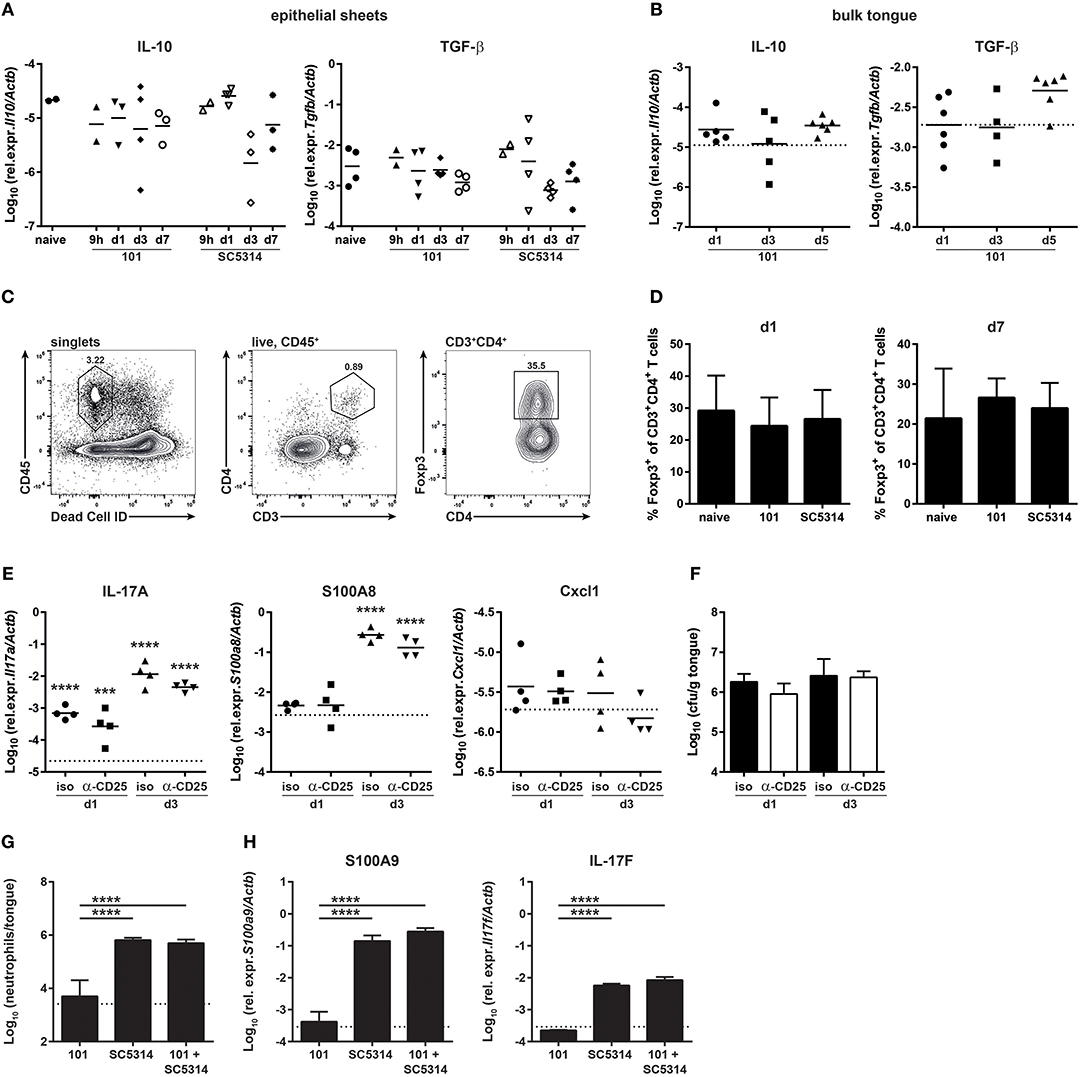
Figure 3. The persistent strain 101 does not suppress the antifungal host response. (A,B) Relative expression of Il10 (left) and Tgfb1 transcripts (right) in epithelial sheets (A) or in bulk tongue tissue (B) of mice that were infected with strain 101 or SC5314 for the indicated period of time. Each symbol represents a pool of epithelial sheets from three animals each (A) or a single mouse (B). The geomean of each group is indicated. Data are pooled from two independent experiments each. (C,D) % Foxp3+ Tregs within the viable CD45+CD3+CD4+ population in the tongue of mice that were infected with strain 101 or SC5314 for the indicated period of time. Representative FACS plots and the gating strategy are shown in C, summary plots showing the mean + SD of data pooled from two independent experiments with 3–4 animals per group are shown in D. (E,F) Mice were treated with anti-CD25 or isotype control antibody prior to infection with strain 101. Relative expression of Il17a (left), S100a8 (middle), and Cxcl1 (right) (E) and tongue fungal loads (F) in the bulk tongue tissue at the indicated time point after infection are shown. In (E), each symbol represents a single animal, The geomean of each group is indicated. The dotted line represents transcript levels in naïve animals (mean of 8 animals). In (F), each bar is the geomean + SD of 4 animals per group. (G,H) WT mice were infected sublingually with a 1:1 mixture of strain 101 and strain SC5314 or with each strain alone. Tongues were harvested on day 1 post-infection and analyzed for the infiltration of Ly6G+Ly6CloCD11b+ neutrophils by flow cytometry (G) and for the expression of S100a9 and Il17f (H) transcript by RT-qPCR. Data are the mean + SD of 3–4 mice per group. Graphs display data representative of one out of two independent experiments. The dotted line represents the detection limit. Statistics were calculated using one-way ANOVA. In (E), statistics compare infected to naïve groups. ***p < 0.001, ****p < 0.0001.
We further looked at regulatory T cells (Tregs) and whether they contribute to suppression of the early host response to strain 101. Immune suppression might be undetectable at the RNA level as transcriptional changes may occur in rare cell populations that are lost in resolution in the whole tissue. Tregs are more abundant in the oral mucosa compared to other organs (34). Detection of lymphocytes in the tongue is challenging due to the small number of cells and the high degree of autofluorescence of this tissue (37, 38). Using a refined protocol of tissue preparation and flow cytometry we were able to detect a small number of Foxp3+ T cells in the tongue which were present at comparable frequencies in naïve and infected animals (Figures 3C–D). Moreover, depletion of Tregs prior to infection (Figure S5) did not alter the kinetics of the epithelial response to strain 101 or allow expression of inflammatory genes such as those regulating the neutrophil response (Figure 3E), nor did it affect the fungal load (Figure 3F). This indicates that Tregs are not responsible for the limited and delayed host response at the onset of persistent OPC.
Finally, we tested the capacity of strain 101 to suppress the host response in the oral mucosa with an unbiased approach using a co-infection experiment for which we infected mice with a 1:1 mix of strain 101 and strain SC5314. We questioned whether the presence of strain 101 would lead to a reduced overall host response in the tongue due to a dominant regulatory response. This was however not the case: the induction of a rapid inflammatory response was as efficient after co-infection as after infection with strain SC5314 alone. The recruitment of neutrophils to the site of infection as well as the induction of Il17f and S100a9, two of the most strongly induced genes by SC5314 on day 1 post-infection, was comparable in both conditions (Figures 3G–H). These data show that strain 101 is not actively repressing the antifungal host-response and that fungal persistence is not a consequence of suppressed immune activation.
The Treg Compartment Is Normal During Persistent C. albicans Infection
Albeit with delay and more restricted in comparison to strain SC5314, C. albicans strain 101 does induce a pronounced immune response in the host, characterized by a prominent IL-17 signature. While induction of the type 17 response does not lead to clearance of strain 101 from the oral mucosa, IL-17 signaling is essential for controlling persistence and preventing fungal overgrowth (12). Importantly, unlike infection with strain SC5314, the type 17 response to strain 101 infection is maintained over a prolonged period of time (Figures 4A,B). Given the potential of type 17 immunity to cause immunopathology and provoke tissue damage (35), tight regulation by mechanisms such as those mediated by Tregs might be necessary.
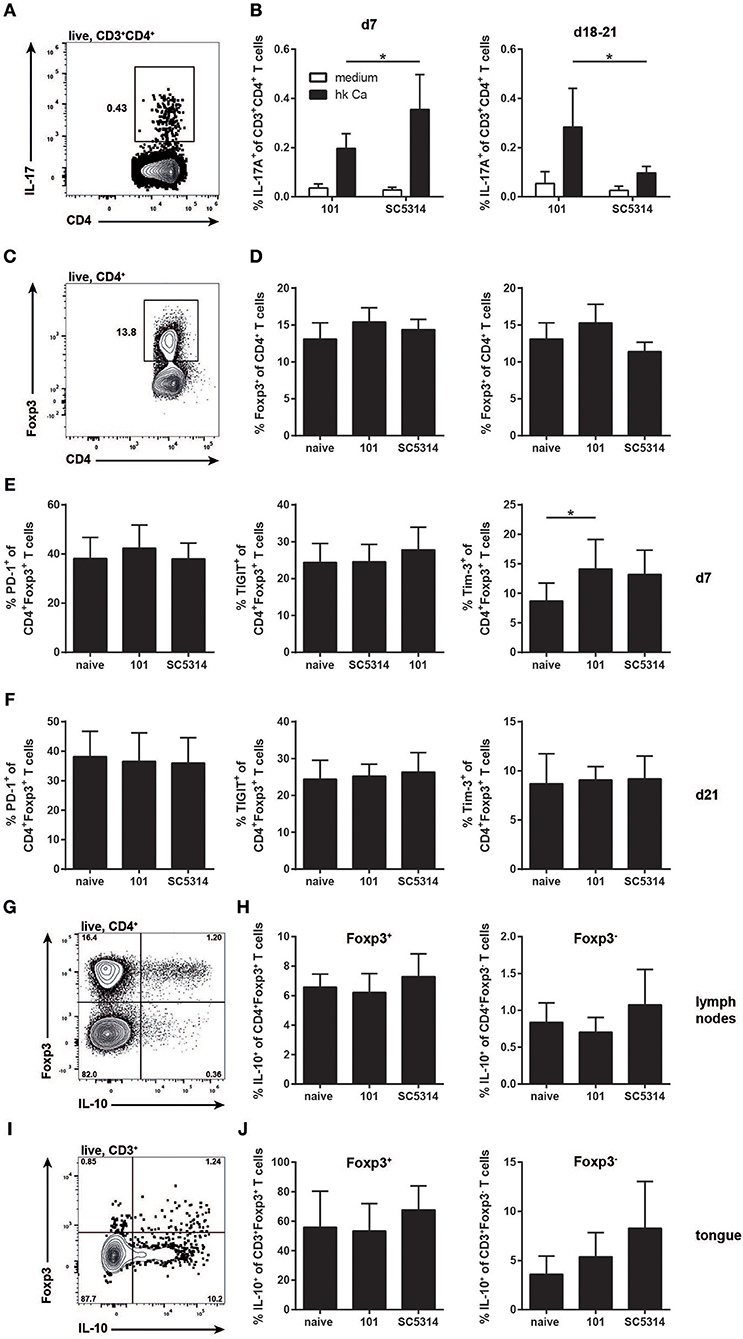
Figure 4. The Treg response during persistent colonization of the oral mucosa with C. albicans. (A–F) WT mice were sublingually infected with strain 101 or SC5314 and cervical lymph node cells were analyzed on day 7 or day 18–21 as indicated. (A,B) Lymph node cells were re-stimulated with MutuDC1940 cells that were pulsed with heat-killed C. albicans or left unpulsed for 5 h in the presence of Brefeldin A. IL-17A production by CD3+CD4+ cells was analyzed by intracellular cytokine staining and flow cytometry. (C,D) The frequency of Foxp3-expressing cells within the CD4+ lymphocyte compartment was assessed by flow cytometry. (E,F) PD-1, TIGIT, and Tim-3 expression by CD4+Foxp3+ Treg cells was analyzed by flow cytometry. (G–J) Il10-Thy1.1 reporter mice were sublingually infected with strain 101 or SC5314 or left naive. IL-10 expression by Foxp3+ Tregs and Foxp3− effector T cells was assessed in the cervical lymph nodes (G,H) and in the tongue (I,J) on day 21 post-infection by flow cytometry. Cells were pregated on CD90+CD4+ (G,H) or on CD45.2+CD3+ (I,J), respectively. Representative FACS plots are shown in (A, C, G, I); summary plots with data pooled from at least 2 experiments with 6–9 animals per infected group and 3–4 animals per naive group are shown in (B, D–F, H, J), with the exception of the right plot in (D), where data are from a single experiment with 3 animals per group. In B, Statistics were calculated using t-test. In (D–F, H, J), statistics were calculated using one-way ANOVA. *p < 0.05.
We thus turned to the analysis of Tregs at later time points of infection to assess their role during persistent colonization of the oral mucosa with C. albicans. Treg numbers did not change significantly over the course of 3 weeks of infection and there was also no difference in Treg frequencies between strain 101 and strain SC5314-infected mice (Figures 4C,D). Treg frequencies remained unchanged even after 100 days of C. albicans colonization with strain 101 (data not shown). We also examined possible functional differences in Tregs in the different settings. Tregs are characterized by the expression of co-inhibitory receptors. We have shown previously that Tim-3 expression is induced on Tregs in a type 17-polarized environment (36) and this was also observed here (Figure 4E). Differences in Tim-3 expression by Tregs were detected on day 7 post-infection, but were lost at later time points (Figures 4E,F). The expression of other co-inhibitory receptors remained unchanged during the course of OPC with either strain of C. albicans (Figures 4E,F).
Next we assessed IL-10 expression by Tregs, a key effector molecule of these cells with a central role in preventing inflammation-mediated tissue damage. IL-10 is very difficult to track by intracellular staining and flow cytometry. Therefore, we made use of Il10-Thy1.1 reporter mice (18). These mice also contain a GFP-reporter for Foxp3 (19), making the need for intranuclear staining of Foxp3 dispensable. Infecting these mice with C. albicans revealed a strong expression of IL-10 by Foxp3+ cells and to a lesser degree by Foxp3− cells (Figures 4G,H). However, again no significant differences in IL-10 production were observed between Tregs from naïve and infected animals, if anything, there was a trend toward higher IL-10 in SC5314 infected mice (Figures 4G,H).
Next, we analyzed IL-10 production by Tregs in the tongue. A prominent fraction of those also expressed IL-10, as assessed by the Thy1.1 reporter (Figures 4I,J). In addition, we detected some IL-10 expression by Foxp3-negative T cells. Albeit with some variation, these populations were detected in all samples analyzed, including the naïve tongue and that of animals infected with either strain of C. albicans (Figures 4I,J).
Tregs and IL-10 Are Not Required for C. albicans Persistence in the Murine Oral Mucosa
Although we did not observe significant changes in the Treg compartment during C. albicans colonization of the oral mucosa, the Tregs present might still modulate the antifungal host response to C. albicans and thereby contribute to fungal persistence in the oral mucosa. We therefore examined DEREG mice (20) in which Tregs can be ablated by means of diphtheria toxin injection. We chose to initiate Treg depletion at a time point when stable fungal colonization and burden-regulating IL-17 immunity was already established (Figure 5A). To our surprise, we did not observe any significant changes in the degree and the quality of the effector T cell response to C. albicans (Figure 5B) or in the growth of the fungus on the mucosa (Figure 5C) upon Treg depletion.
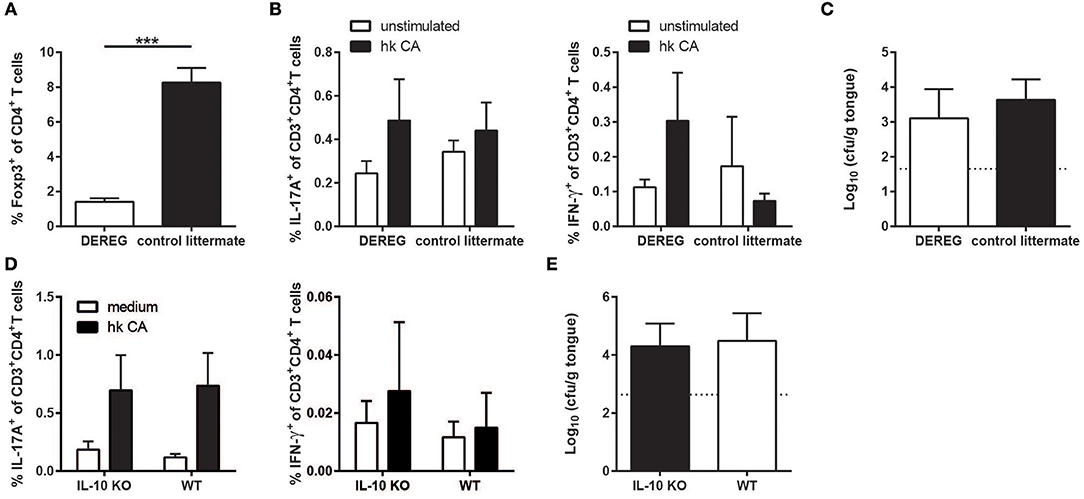
Figure 5. The absence of Tregs or IL-10 does not compromise persistence of strain 101 in the oral epithelium. (A–C) DEREG mice and control littermates were sublingually infected with C. albicans strain 101 and treated with diphtheria toxin on day 11 and 13 after the antifungal response was fully established. One day after the last treatment, the mice were sacrificed for analysis. (A) Treg depletion efficiency was analyzed in the cervical lymph nodes by flow cytometry. Data are the % of Foxp3+ cells within the population of CD4+ viable cells. (B) Lymph node cells were re-stimulated with MutuDC1940 cells that were pulsed with heat-killed C. albicans or left unpulsed for 5 h in the presence of Brefeldin A. IL-17A (left) and IFN-γ (right) production by CD3+CD4+ cells was analyzed by intracellular cytokine staining and flow cytometry. (C) The fungal burden was determined by plating tongue homogenates on YPD agar. Each bar represents the mean + SD of 3 to 4 mice per group. Data are from one out of two independent experiments. (D,E) IL-10-deficient mice and WT controls were sublingually infected with C. albicans strain 101 and analyzed on day 9 post-infection. (D) Lymph node cells were re-stimulated and analyzed for IL-17 (left) and IFN-γ (right) production as in B. (E) Tongue fungal burdens were analyzed as in C. Each bar represents the mean + SD of 8–9 mice per group pooled from two independent experiments. Statistics were calculated using unpaired t-Test. ***p < 0.001.
Because IL-10 production was not limited to the Treg compartment, we also assessed the impact of IL-10 itself on the antifungal response and on C. albicans colonization levels separately by means of IL-10-deficient mice (21). Similarly to what we observed under Treg-depleting conditions, IL-10 deficiency did not affect the extent or the quality of the C. albicans-specific Th17 response in animals infected with strain 101 (Figure 5D). Consistent with this, fungal loads in the tongue were also unchanged in IL-10 knockout mice compared to WT controls (Figure 5E). Together, these findings indicate that immune regulation by Tregs and IL-10 is dispensable for persistent C. albicans colonization of the oral mucosa.
Discussion
Immunoregulatory mechanisms such as those mediated by Tregs and IL-10 are essential for maintaining tolerance and preventing excessive immune responses to commensal microbes. In this study we analyzed the role of immune regulation during C. albicans colonization. Our data indicate that fungal persistence in the oral mucosa is not associated with immunosuppression that would blunt a strong host response and prevent rapid fungal elimination at the onset of infection, as it is observed during OPC with the highly virulent strain SC5314. Rather, the failure of strain 101 to induce an early inflammatory response appears to result from intrinsic properties of the strain and how it interacts with the host and in particular with the epithelium.
The epithelial response to C. albicans in turn instructs the extent of neutrophil trafficking to the site of infection (39). The fungal determinants that are responsible for the strain-specific differences at the interface with the epithelium and that underlie the differential behavior in contact with the host may comprise differences in filamentation (40), tissue invasion (12), and the production of virulence factors such as the recently identified peptide toxin candidalysin (15). Candidalysin has been linked to mucosal immune activation and neutrophil recruitment (15, 41) with an important contribution of IL-1 family cytokine-dependent chemokine induction (39, 42, 43). Our previous work has shown reduced expression of the ECE1 gene (which encodes for candidalysin) by strain 101 in contact with epithelial cells (12), and this difference to strain SC5314 may at least in part be responsible for the observed interspecies differences. The identification of relevant fungal factors underlying the differential host response is subject of ongoing research.
Induction of an inflammatory response by the highly virulent C. albicans strain SC5314 in epithelial cells and tissues has been observed in previous transcriptomic studies (44–47). However, our study is the first, to our knowledge, that assesses the kinetics of the transcriptional response to two functionally distinct C. albicans isolates in the murine oral mucosa in vivo. While host genes associated with the neutrophil response are differentially regulated during infection with strains SC5314 and 101, many other immune genes including those linked to the IL-17 pathway, are expressed to comparable levels in response to the two C. albicans strains under investigation, albeit with delay in case of strain 101. By day 3, IL-17 and related cytokines are expressed at comparable levels in both SC5314- and 101-infected tongues. Similarly, Th17 differentiation was indistinguishable after day 7 post-infection. The differential activation of the epithelial-inflammatory response and the IL-17 pathway by strain 101 underlines the notion that the two host pathways constitute two distinct modules of the antifungal response in the oral mucosa that are regulated largely independently. It also supports our earlier observation that the recruitment of neutrophils to the infected epithelium is not compromised in absence of IL-17 signaling (33), and strong IL-17 induction in response to strain 101 on the other hand is not sufficient for bringing neutrophils to the site of infection (12).
The uncoupling of the IL-17 pathway from the epithelial/neutrophil response is also consistent with our recent finding that induction of IL-17 is instructed by dendritic cells and in particular by the Langerin+ subset of dendritic cells that have the unique property in the C. albicans-infected oral mucosa to co-produce the three major IL-17-instructing cytokines IL-23, IL-1, and IL-6 (37). The IL-17 response may be further modulated via epithelial sensing of C. albicans and IL-1 family cytokines that are released from the epithelium in response to candidalysin in case of infection with strain SC5314 (42, 43).
β-defensin-3 is a prominent target gene of IL-17 and it was proposed to act as the major antifungal effector molecule during OPC (48). Strong induction in response to both strains SC5314 and 101 was confirmed in our RNA-seq analysis. However, surprisingly, β-defensin-3 expression was delayed when compared to the kinetics of IL-17 induction in case of SC5314. It reached its maximum levels only by day 3 post-infection when the fungus was already nearly cleared from the oral mucosa. Together with the observation that the lack of β-defensin-3 resulted in a slightly less severe loss of fungal control than IL-17 signaling deficiency (48), this suggests that additional IL-17-dependent antimicrobial effector molecules are likely involved.
Based on the observation that the IL-17 pathway is crucial for regulating fungal growth independently of the fungal isolate (12), it is surprising that strain 101 is able to resist the continuously activated IL-17-mediated immunity and to persist in the oral mucosa. Its predominant localization to the stratum corneum may limit its exposure to host immune effectors. Further, it is surprising that the prolonged IL-17 response does not trigger signs of tissue damage despite the strong pro-inflammatory potential of the cytokine. The IL-17 pathway can drive severe immunopathology in barrier tissues under certain conditions, as seen for instance in the skin of psoriatic patients and in murine models of psoriasis (49, 50). Th17 cells have been divided in different subtypes that differ in their pathogenic potential (51). Here, we show that a small proportion of all T helper cells in the lymph nodes and in the tongue of mice infected with strain SC5314 or 101 do express IL-10. However, a comprehensive analysis of the cytokine profile of C. albicans-specific Th17 cells during persistent and acute OPC remains to be determined in the future.
The generation of Th17 cells was proposed to be supported by Tregs. This might be through the provision of TGF-β (19, 52, 53), but also through the consumption of IL-2, which negatively regulates Th17 differentiation (54). As such, Pandiyan et al. showed in an adoptive transfer system that co-transfer of Tregs with effector T cells enhances Th17 differentiation in response to C. albicans (55). In our study, depletion of Tregs in wild type mice did not affect the induction of C. albicans-specific Th17 cells, nor did it affect already established type 17 immunity when Tregs were depleted at a later time point of persistent colonization. Similar results were also obtained in mice lacking IL-10, one of the key regulatory cytokines and a potent suppressor of Th17 cells (56). Together, our data thus indicate that immune regulation does not make an essential contribution to the balance of a protective non-pathological Th17 response and fungal persistence in our model.
In conclusion, our study indicates that the attenuated host response to C. albicans strain 101 is well-tared to allow fungal persistence in the oral mucosa by preventing elimination of the fungus but also uncontrolled fungal overgrowth as well as avoiding the development of immunopathology.
Ethics Statement
All mouse experiments described in this study were conducted in strict accordance with the guidelines of the Swiss Animal Protection Law and were performed under protocols approved by the Veterinary office of the Canton Zurich, Switzerland (license number 201/2012 and 183/2015). All efforts were made to minimize suffering and ensure the highest ethical and human standards.
Author Contributions
FRK, KL, SA, NJ, and SL-L: conceptualization; SA and DS: methodology; FRK, KL, SA, FS, and CL: investigation; FRK, KL, SA, VDTT, FS, and CL: formal analysis; NJ, DS, and SL-L: validation and project administration; FRK, KL, VDTT, and SL-L: visualization; MP, NJ, DS, and SL-L: funding acquisition; MP, NJ, and SL-L; supervision; FRK and SL-L: writing-original draft; KL, SA, VDTT, NJ, and DS: writing-review and editing.
Conflict of Interest Statement
SA currently works for Novartis but on a different topic and the data are restricted to his previous role in the LeibundGut-lab.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank Manfred Kopf, Vijay Kuchroo and Casey Weaver for mice; the staff of the Laboratory Animal Service Center of University of Zurich for animal husbandry; the Lausanne Genomic Technologies Facility for sequencing; and members of the LeibundGut-lab and the Joller-lab for helpful advice and discussions. This work was supported by the Swiss National Science Foundation (grant CRSII3_141848 to MP, DS, and SL-L, grant PP00P3_150663 to NJ), the Promedica Foundation (to SL-L), the Fonds zur Förderung des akademischen Nachwuchses of the Zurich University Association (to SL-L.) and the European Research Council (grant 677200 to NJ).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2019.00330/full#supplementary-material
Figure S1. Characterization of tongue epithelial sheets. Epithelial sheets were isolated from infected tongues as described in the Methods and analyzed by microscopy after periodic acid-Schiff staining (A) or digested with trypsin to obtain single cell suspensions and analyzed by flow cytometry (B) for their composition. In (A), the black arrow indicates epithelial cells; the white arrow indicates cellular infiltrates; the white arrow head indicates C. albicans hyphae. The numbers indicate the % of cells in each gate. The representative data shown are from strain SC5314-infected animals.
Figure S2. Overlap of differentially expressed genes at different time points and by the different strains of C. albicans. Venn diagrams showing the number of differentially expressed genes in tongue epithelial sheets at each time point after infection with C. albicans strain SC5314 (A) or strain 101 (B). (C) Venn diagram showing the differentially expressed genes on day 1 and 3 after infection with strain SC5314 and on day 3 and 7 after infection with strain 101, respectively, i.e., the conditions with the highest number of gene expression activity. The numbers in the overlapping areas refer to the number of genes that were co-regulated in the respective conditions.
Figure S3. The host response to C. albicans in the tongue is dominated by changes in the immune response and metabolic processes. The heat maps show the upregulated (A) and downregulated (B) GO processes with the smallest FDR. The heat map was arranged by hierarchical clustering using the distance of one minus Pearson correlation and the average linkage mode.
Figure S4. Transcriptional response to C. albicans strain 101 during persistent oral infection. (A,B) Mice were infected with strain 101 for 60 days. Relative expression of Cxcl1 (left), Cxcl2 (middle), and Cxcl5 transcripts (right) in the bulk tongue tissue (A) and cfu in the feces (B) are shown. Each symbol represents one animal. The geomean of each group is indicated. (C–E) Mice were infected with strain 101 for 12–14 months. The fungal load in the feces (C) and in the tongue (D) are shown. (E) shows representative tongue histology stained with PAS from one of the still highly colonized mice. (F) Relative expression of S100a8 (left), Lcn2 (middle), and Defb1 transcripts (right) in the tongue of mice were infected with strains 101 or SC5314 for the indicated period of time. Each symbol represents one animal. The geomean of each group is indicated. Dotted line: geomean of a naive control group. Data are pooled from two independent experiments each, except for fecal cfu in (C), which are from one experiment only. Statistics were calculated using one-way ANOVA (comparison of infected to naïve groups). *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001.
Figure S5. Treg Depletion. % Foxp3+ cells within the splenic CD4+CD3+ population of mice treated with anti-CD25 antibody or isotype control as indicated.
Table S1. Oligonucleotides used in this study.
Table S2. Antibodies used in this study.
References
1. Brown GD, Denning DW, Gow NA, Levitz SM, Netea MG, White TC. Hidden killers: human fungal infections. Sci Transl Med. (2012) 4:165rv13. doi: 10.1126/scitranslmed.3004404
2. de Repentigny, Lewandowski D, Jolicoeur P. Immunopathogenesis of oropharyngeal candidiasis in human immunodeficiency virus infection. Clin Microbiol Rev. (2004) 17:729–59, table of contents. doi: 10.1128/CMR.17.4.729-759.2004
3. Goncalves B, Ferreira C, Alves CT, Henriques M, Azeredo J, Silva S. Vulvovaginal candidiasis: epidemiology, microbiology and risk factors. Crit Rev Microbiol. (2016) 42:905–27. doi: 10.3109/1040841X.2015.1091805
4. Kirkpatrick CH. Chronic mucocutaneous candidiasis. Pediatr Infect Dis J. (2001) 20:197–206. doi: 10.1097/00006454-200102000-00017
5. Puel A, Cypowyj S, Marodi L, Abel L, Picard C, Casanova JL. Inborn errors of human IL-17 immunity underlie chronic mucocutaneous candidiasis. Curr Opin Allergy Clin Immunol. (2012) 12:616–22. doi: 10.1097/ACI.0b013e328358cc0b
6. Pfaller MA, Diekema DJ. Epidemiology of invasive candidiasis: a persistent public health problem. Clin Microbiol Rev. (2007) 20:133–63. doi: 10.1128/CMR.00029-06
7. Struck MF, Gille J. Fungal infections in burns: a comprehensive review. Ann Burns Fire Disasters. (2013) 26:147–53.
8. Odds FC, Bougnoux ME, Shaw DJ, Bain JM, Davidson AD, Diogo D, et al. Molecular phylogenetics of Candida albicans. Eukaryot Cell. (2007) 6:1041–52. doi: 10.1128/EC.00041-07
9. MacCallum DM, Castillo L, Nather K, Munro CA, Brown AJ, Gow NA, et al. Property differences among the four major Candida albicans strain clades. Eukaryot Cell. (2009) 8:373–87. doi: 10.1128/EC.00387-08
10. Hirakawa MP, Martinez DA, Sakthikumar S, Anderson MZ, Berlin A, Gujja S, et al. Genetic and phenotypic intra-species variation in Candida albicans. Genome Res. (2015) 25:413–25. doi: 10.1101/gr.174623.114
11. Marakalala MJ, Vautier S, Potrykus J, Walker LA, Shepardson KM, Hopke A, et al. Differential adaptation of Candida albicans in vivo modulates immune recognition by dectin-1. PLoS Pathog. (2013) 9:e1003315. doi: 10.1371/journal.ppat.1003315
12. Schönherr FA, Sparber F, Kirchner FR, Guiducci E, Trautwein-Weidner K, Gladiator A, et al. The intraspecies diversity of C. albicans triggers qualitatively and temporally distinct host responses that determine the balance between commensalism and pathogenicity. Mucosal Immunol. (2017) 10:1335–50. doi: 10.1038/mi.2017.2
13. Hohl TM. Overview of vertebrate animal models of fungal infection. J Immunol Methods. (2014) 410:100–12. doi: 10.1016/j.jim.2014.03.022
14. Sparber F, LeibundGut-Landmann S. Interleukin 17-mediated host defense against Candida albicans. Pathogens. (2015) 4:606–19. doi: 10.3390/pathogens4030606
15. Moyes DL, Wilson D, Richardson JP, Mogavero S, Tang SX, Wernecke J, et al. Candidalysin is a fungal peptide toxin critical for mucosal infection. Nature. (2016) 532:64–8. doi: 10.1038/nature17625
16. Swidergall M, Solis NV, Lionakis MS, Filler SG. EphA2 is an epithelial cell pattern recognition receptor for fungal beta-glucans. Nat Microbiol. (2018) 3:53–61. doi: 10.1038/s41564-017-0059-5
17. Gladiator A, Wangler N, Trautwein-Weidner K, LeibundGut-Landmann S. Cutting edge: IL-17-secreting innate lymphoid cells are essential for host defense against fungal infection. J Immunol. (2013) 190:521–5. doi: 10.4049/jimmunol.1202924
18. Maynard CL, Harrington LE, Janowski KM, Oliver JR, Zindl CL, Rudensky AY, et al. Regulatory T cells expressing interleukin 10 develop from Foxp3+ and Foxp3- precursor cells in the absence of interleukin 10. Nat Immunol. (2007) 8:931–41. doi: 10.1038/ni1504
19. Bettelli E, Carrier Y, Gao W, Korn T, Strom TB, Oukka M, et al. Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells. Nature. (2006) 441:235–8. doi: 10.1038/nature04753
20. Lahl K, Loddenkemper C, Drouin C, Freyer J, Arnason J, Eberl G, et al. Selective depletion of Foxp3+ regulatory T cells induces a scurfy-like disease. J Exp Med. (2007) 204:57–63. doi: 10.1084/jem.20061852
21. Kuhn R, Lohler J, Rennick D, Rajewsky K, Muller W. Interleukin-10-deficient mice develop chronic enterocolitis. Cell. (1993) 75:263–74. doi: 10.1016/0092-8674(93)80068-P
22. Odds FC, Brown AJ, Gow NA. Candida albicans genome sequence: a platform for genomics in the absence of genetics. Genome Biol. (2004) 5:230. doi: 10.1186/gb-2004-5-7-230
23. Solis NV, Filler SG. Mouse model of oropharyngeal candidiasis. Nat Protoc. (2012) 7:637–42. doi: 10.1038/nprot.2012.011
24. Amorim-Vaz S, Tran Vdu, Pradervand S, Pagni M, Coste AT, Sanglard D. RNA enrichment method for quantitative transcriptional analysis of pathogens in vivo applied to the fungus Candida albicans. MBio (2015) 6:e00942-15. doi: 10.1128/mBio.00942-15
25. Martin M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet.jounal (2011) 17:10–2. doi: 10.14806/ej.17.1.200
26. Anders S, Pyl PT, Huber W. HTSeq–a Python framework to work with high-throughput sequencing data. Bioinformatics. (2015) 31:166–9. doi: 10.1093/bioinformatics/btu638
27. Robinson MD, Oshlack A. A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol. (2010) 11:R25. doi: 10.1186/gb-2010-11-3-r25
28. Law CW, Chen Y, Shi W, Smyth GK. voom: precision weights unlock linear model analysis tools for RNA-seq read counts. Genome Biol. (2014) 15:R29. doi: 10.1186/gb-2014-15-2-r29
29. Smyth GK. limma: Linear Models for Microarray Data. In: Gentleman R, Carey VJ, Huber W, Irizarry RA, Dudoit S, Editors. Bioinformatics and Computational Biology Solutions Using R and Bioconductor. Statistics for Biology and Health. New York, NY: Springer (2005). p. 397–420.
30. Benjamini Y, Hochberg Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Stat Soc Ser B. (1995) 57:289–300. doi: 10.1111/j.2517-6161.1995.tb02031.x
31. Fuertes Marraco, Grosjean F, Duval A, Rosa M, Lavanchy C, Ashok D, et al. Novel murine dendritic cell lines: a powerful auxiliary tool for dendritic cell research. Front Immunol. (2012) 3:331. doi: 10.3389/fimmu.2012.00331
32. Cossarizza A, Chang HD, Radbruch A, Akdis M, Andra I, Annunziato F, et al. Guidelines for the use of flow cytometry and cell sorting in immunological studies. Eur J Immunol. (2017) 47:1584–797. doi: 10.1002/eji.201646632
33. Trautwein-Weidner K, Gladiator A, Nur S, Diethelm P, LeibundGut-Landmann S. IL-17-mediated antifungal defense in the oral mucosa is independent of neutrophils. Mucosal Immunol. (2015) 8:221–31. doi: 10.1038/mi.2014.57
34. Park JY, Chung H, DiPalma DT, Tai X, Park JH. Immune quiescence in the oral mucosa is maintained by a uniquely large population of highly activated Foxp3(+) regulatory T cells. Mucosal Immunol. (2018) 11:1092–102. doi: 10.1038/s41385-018-0027-2
35. Eyerich K, Dimartino V, Cavani A. IL-17 and IL-22 in immunity: driving protection and pathology. Eur J Immunol. (2017) 47:607–14. doi: 10.1002/eji.201646723
36. Littringer K, Moresi C, Rakebrandt N, Zhou X, Schorer M, Dolowschiak T, et al. Common features of regulatory T cell specialization during Th1 responses. Front Immunol. (2018) 9:1344. doi: 10.3389/fimmu.2018.01344
37. Sparber F, Dolowschiak T, Mertens S, Lauener L, Clausen BE, Joller N, et al. Langerin+ DCs regulate innate IL-17 production in the oral mucosa during Candida albicans-mediated infection. PLoS Pathog. (2018) 14:e1007069. doi: 10.1371/journal.ppat.1007069
38. Sparber F, LeibundGut-Landmann S. Assessment of immune responses to fungal infections: identification and characterization of immune cells in the infected tissue. Methods Mol Biol. (2017) 1508:167–82. doi: 10.1007/978-1-4939-6515-1_8
39. Altmeier S, Toska A, Sparber F, Teijeira A, Halin C, LeibundGut-Landmann S. IL-1 coordinates the neutrophil response to C. albicans in the oral mucosa. PLoS Pathog. (2016) 12:e1005882. doi: 10.1371/journal.ppat.1005882
40. Moyes DL, Runglall M, Murciano C, Shen C, Nayar D, Thavaraj S, et al. A biphasic innate immune MAPK response discriminates between the yeast and hyphal forms of Candida albicans in epithelial cells. Cell Host Microbe. (2010) 8:225–35. doi: 10.1016/j.chom.2010.08.002
41. Richardson JPH, Willems ME, Moyes DL, Shoaie S, Barker KS, Tan SL, et al. Candidalysin drives epithelial signaling, neutrophil recruitment, and immunopathology at the vaginal mucosa. Infect Immun. (2017) 86:IAI.00645-17. doi: 10.1128/IAI.00645-17
42. Verma AH, Richardson JP, Zhou C, Coleman BM, Moyes DL, Ho J, et al. Oral epithelial cells orchestrate innate type 17 responses to Candida albicans through the virulence factor candidalysin. Sci Immunol. (2017) 2:eaam8834. doi: 10.1126/sciimmunol.aam8834
43. Verma AH, Zafar H, Ponde NO, Hepworth OW, Sihra D, Aggor FEY, et al. IL-36 and IL-1/IL-17 drive immunity to oral candidiasis via parallel mechanisms. J Immunol. (2018) 201:627–34. doi: 10.4049/jimmunol.1800515
44. Bruno VM, Shetty AC, Yano J, Fidel PL, Noverr MC, Peters BM. Transcriptomic analysis of vulvovaginal candidiasis identifies a role for the NLRP3 inflammasome. MBio. (2015) 6:e00182-15 doi: 10.1128/mBio.00182-15
45. Conti HR, Shen F, Nayyar N, Stocum E, Sun JN, Lindemann MJ, et al. Th17 cells and IL-17 receptor signaling are essential for mucosal host defense against oral candidiasis. J Exp Med. (2009) 206:299–311. doi: 10.1084/jem.20081463
46. Liu Y, Shetty AC, Schwartz JA, Bradford LL, Xu W, Phan QT, et al. New signaling pathways govern the host response to C. albicans infection in various niches. Genome Res. (2015) 25:679–89. doi: 10.1101/gr.187427.114
47. Moyes DL, Shen C, Murciano C, Runglall M, Richardson JP, Arno M, et al. Protection against epithelial damage during Candida albicans infection is mediated by PI3K/Akt and mammalian target of rapamycin signaling. J Infect Dis. (2014) 209:1816–26. doi: 10.1093/infdis/jit824
48. Conti HR, Bruno VM, Childs EE, Daugherty S, Hunter JP, Mengesha BG, et al. IL-17 receptor signaling in oral epithelial cells is critical for protection against oropharyngeal candidiasis. Cell Host Microbe. (2016) 20:606–17. doi: 10.1016/j.chom.2016.10.001
49. Hawkes JE, Chan TC, Krueger JG. Psoriasis pathogenesis and the development of novel targeted immune therapies. J Allergy Clin Immunol. (2017) 140:645–53. doi: 10.1016/j.jaci.2017.07.004
50. van der Fits L, Mourits S, Voerman JS, Kant M, Boon L, Laman JD, Cornelissen F, et al. Imiquimod-induced psoriasis-like skin inflammation in mice is mediated via the IL-23/IL-17 axis. J Immunol. (2009) 182:5836–45. doi: 10.4049/jimmunol.0802999
51. McGeachy MJ, Bak-Jensen KS, Chen Y, Tato CM, Blumenschein W, McClanahan T, et al. TGF-beta and IL-6 drive the production of IL-17 and IL-10 by T cells and restrain T(H)-17 cell-mediated pathology. Nat Immunol. (2007) 8:1390–7. doi: 10.1038/ni1539
52. Mangan PR, Harrington LE, O'Quinn DB, Helms WS, Bullard DC, Elson CO, et al. Transforming growth factor-beta induces development of the T(H)17 lineage. Nature. (2006) 441:231–4. doi: 10.1038/nature04754
53. Veldhoen M, Hocking RJ, Atkins CJ, Locksley RM, Stockinger B. TGFbeta in the context of an inflammatory cytokine milieu supports de novo differentiation of IL-17-producing T cells. Immunity. (2006) 24:179–89. doi: 10.1016/j.immuni.2006.01.001
54. Laurence A, Tato CM, Davidson TS, Kanno Y, Chen Z, Yao Z, et al. Interleukin-2 signaling via STAT5 constrains T helper 17 cell generation. Immunity. (2007) 26:371–81. doi: 10.1016/j.immuni.2007.02.009
55. Pandiyan P, Conti HR, Zheng L, Peterson AC, Mathern DR, Hernandez-Santos N, et al. CD4(+)CD25(+)Foxp3(+) regulatory T cells promote Th17 cells in vitro and enhance host resistance in mouse Candida albicans Th17 cell infection model. Immunity. (2011) 34:422–34. doi: 10.1016/j.immuni.2011.03.002
56. Huber S, Gagliani N, Esplugues E, O'Connor W, Jr, Huber FJ, Chaudhry A, et al. Th17 cells express interleukin-10 receptor and are controlled by Foxp3(-) and Foxp3+ regulatory CD4+ T cells in an interleukin-10-dependent manner. Immunity. (2011) 34:554–65. doi: 10.1016/j.immuni.2011.01.020
57. Huang XR, Chung AC, Wang XJ, Lai KN, Lan HY. Mice overexpressing latent TGF-beta1 are protected against renal fibrosis in obstructive kidney disease. Am J Physiol Renal Physiol. (2008) 295:F118–27. doi: 10.1152/ajprenal.00021.2008
58. Overbergh L, Giulietti A, Valckx D, Decallonne R, Bouillon R, Mathieu C. The use of real-time reverse transcriptase PCR for the quantification of cytokine gene expression. J Biomol Tech. (2003) 14:33–43.
59. Ivanov II, McKenzie BS, Zhou L, Tadokor CE, Lepelley A, Lafaille JJ, et al. The orphan nuclear receptor RORgammat directs the differentiation program of proinflammatory IL-17+ T helper cells. Cell. (2006) 126:1121–33. doi: 10.1016/j.cell.2006.07.035
60. Kundu P, Ling TW, Korecka A, Li Y, D'Arienzo R, Bunte RM, et al. Absence of intestinal PPARgamma aggravates acute infectious colitis in mice through a lipocalin-2-dependent pathway. PLoS Pathog. (2014) 10:e1003887. doi: 10.1371/journal.ppat.1003887
Keywords: Candida albicans, oropharyngeal candidiasis, immune regulation, persistence, IL-17, IL-10, regulatory T cells
Citation: Kirchner FR, Littringer K, Altmeier S, Tran VDT, Schönherr F, Lemberg C, Pagni M, Sanglard D, Joller N and LeibundGut-Landmann S (2019) Persistence of Candida albicans in the Oral Mucosa Induces a Curbed Inflammatory Host Response That Is Independent of Immunosuppression. Front. Immunol. 10:330. doi: 10.3389/fimmu.2019.00330
Received: 29 September 2018; Accepted: 08 February 2019;
Published: 27 February 2019.
Edited by:
Avi-Hai Hovav, Hebrew University of Jerusalem, IsraelReviewed by:
Julian Naglik, King's College London, United KingdomPushpa Pandiyan, Case Western Reserve University, United States
Copyright © 2019 Kirchner, Littringer, Altmeier, Tran, Schönherr, Lemberg, Pagni, Sanglard, Joller and LeibundGut-Landmann. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Salomé LeibundGut-Landmann, salome.leibundgut-landmann@uzh.ch