 Isaac Trindade-Santos
Isaac Trindade-Santos Kátia de Meirelles Felizola Freire
Kátia de Meirelles Felizola Freire- Laboratório de Ecologia Pesqueira, Departamento de Engenharia de Pesca e Aquicultura, Universidade Federal de Sergipe, São Cristóvão, Brazil
Fish reproductive biology plays an important role for fishery management, especially in developing countries. The aim of this study was to compile all available information and analyze reproduction patterns of marine fishes in three Large Marine Ecosystems (LMEs): North, East, and South Brazil Shelves. We tested the hypothesis that the onset and duration of spawning season differ among these three LMEs; compared the ratio between length at first maturity and asymptotic length with the global trend observed; analyzed sex ratios; and tested whether females allocate more energy into reproduction than males. The following data were compiled from published sources and “gray” literature: sex ratio, spawning season, gonadosomatic indices (GSI), and length at first maturity (Lm). The reproductive load was estimated as Lm/L∞. The median extension of the spawning season in the North, East, and South Brazil Shelves were 6.5, 6.0, and 5.0 months, respectively, with higher frequency during austral summer in South Brazil. Marine fishes from these three LMEs can be grouped in summer and non-summer spawners. About 96% of the cases the reproductive load was between 0.3 and 0.8, which is slightly shifted toward smaller values, compared with the global range of 0.4–0.9. Gonadosomatic indices for females were higher. Contrary to some expectations, there is seasonality in the reproduction of tropical fishes. However, seasonality is stronger in southern populations. Size at first maturity is not efficiently used as a tool for fisheries management in the ecosystems analyzed.
Introduction
Fish reproductive biology is crucial for fishery management (Jakobsen et al., 2009), especially in developing countries such as Brazil, where managers rely on size at first maturity and the onset and duration of spawning season for managing fisheries (Dias Neto, 2010a). Despite the importance of fish biology, the lack or scarcity of fishery data in some countries leads to overexploitation of the stocks and management failure in some cases (Kinas, 1996; Alves and Minte-Vera, 2012). Furthermore, subsidies drive the limits of exploitation beyond sustainability (Abdallah and Sumaila, 2007; Castello, 2007; Dias Neto, 2010b). Spawning season has a temporal scale, with most fishes exhibiting one or two spawning seasons a year (Bye, 1984; Cushing, 1990). The success of reproductive activities depends on the match between spawning season and the best conditions for larval survival (Cushing, 1973; Lowerre-Barbieri et al., 2011). Hence, it is fundamental to have basic biological data in order to conduct a reliable management system.
Large Marine Ecosystems (LMEs) are defined as ecosystems with similar bathymetry, hydrography, productivity and dependent populations at trophic level (Ekau and Knoppers, 2003). The vast extension of the Brazilian coast comprises 17 coastal states and encompasses three LMEs: North Brazil Shelf, East Brazil Shelf, and South Brazil Shelf. Thus, maintaining a continuous system of data collection of fishery statistics and basic biological information has been a continuous challenge. Besides, Brazil has the highest richness of freshwater fishes in the world (about 3139 species) and one of the highest richness of marine fish species (1207) (Froese and Pauly, 2015). FishBase, an electronic encyclopedia with biological information about fishes, indicates, e.g., that local information related to reproduction (size at first maturity) is available for only 5.4% of Brazilian fish species (Froese and Pauly, 2015). In principle, this information could be derived from meta-analysis (Borenstein et al., 2009). However, before this is undertaken, many of the empirical values that have been obtained so far should be made available.
Currently, some results of important studies and/or surveys are not published, remaining in the format of theses, technical reports, and/or proceedings of local congresses or conferences, often in language not widely read by the international scientific community. A study by Stergiou and Tsikliras (2006) showed that hypotheses can be more straightforwardly tested after gray literature has been accessed. Godinho and Lamas (2010) recently reviewed all information available on the reproduction of Brazilian freshwater fishes. However, for Brazilian marine fishes the only review found was by Ferraz-Dias (1989).
Thus, this study was proposed with the main objective of compiling and analyzing all available information (published or not) on reproduction for marine fishes of the three Brazilian LMEs. We analyzed information available on length at first maturity and spawning season for Brazilian marine fishes and how it has been used for fisheries management. We tested the hypothesis that the onset and duration of spawning season differ among the three LMEs that comprise Brazil; compared the ratio between length at first maturity and asymptotic length with the global trend observed; analyzed sex ratios; and tested whether females allocate more energy into reproduction than males.
Materials and Methods
The basis for our analysis of reproductive patterns of marine fishes from Brazil was FishBase (see www.fishbase.org). This database was complemented through a compilation of data available until 2013 from published sources (local, national, and international journals, and books) and from “gray” literature (theses, reports, conference proceedings) (see Supplementary Material). Within each LME, the following data were compiled: the onset and duration of spawning period (months), methods used to determine spawning period (histological or macroscopic observation, or fishers' ecological knowledge), length at first maturity (Lm, cm) (Vazzoler, 1996), sex-ratio (female:male) by month and/or year, and gonadosomatic indices using equation (a) GSI = (gonad weight/somatic weight) × 100 (Isaac-Nahum and Vazzoler, 1987; Wootton, 1998) or some variation, equation (b) GSI = (gonad weight × 103/fork length3) (Schaefer and Orange, 1956), or equation (c) GSI = (gonad weight/total lengthb) × 106, where “b” is the power of the length-weight relationship (Vazzoler, 1996), with all units in centimeters and grams. We analyzed the proportional difference between GSI of females and males using the highest or mean values reported in each study [(female GSI − male GSI) × 100/(female GSI)]. In order to estimate the reproductive load [Lm/L∞; (Froese and Binohlan, 2000)], growth parameters [L∞(cm), K (year−1) and to(years)] were also gathered. For comparison purposes, we used data available in FishBase for neighboring countries, i.e., Guyana, Suriname, and French Guiana.
Data on reproduction were complemented by information on sample size, size range of the specimens sampled (cm), length type [total (TL), standard (SL), fork (FL), disc width (DW) (all in cm), minimum and maximum maturity size and age, spawning frequency (total or partial), sampling frequency (weekly, monthly, seasonally), fishing gears used (gillnet, trawl net, longline, casting net, trap), and sampling period. Reproduction data were allocated to each of the three LMEs based on information (geographic coordinates of sampling location) presented in each publication considering the limits provided by Ekau and Knoppers (2003):
• North Brazil Shelf (LME #17)—starts at Caribbean Sea (Orinoco delta) and extends eastwards to the Parnaíba River estuary in Brazil. Includes part of Venezuela, Guiana, Suriname, and French Guiana, along with two coastal states from northern Brazil (Amapá and Pará) and one from northeastern Brazil (Maranhão).
• East Brazil Shelf (LME #16)—starts at the Parnaíba River estuary in northeastern Brazil and extends east and southwards to Cape São Tomé in the state of Rio de Janeiro. Thus, nine Brazilian states were included in this LME (Piauí, Ceará, Rio Grande do Norte, Paraíba, Pernambuco, Alagoas, Sergipe, and Bahia in northeastern Brazil, and Espírito Santo in southeastern Brazil).
• South Brazil Shelf (LME #15)—includes states in southeastern Brazil (Rio de Janeiro and São Paulo) and in the southern region (Paraná, Santa Catarina, and Rio Grande do Sul).
Statistical difference among medians was tested by the Mood median test (Zar, 2010). Mann-Whitney was used to test the difference among two or more samples when assumptions for similar parametric tests were not satisfied (Zar, 2010). Bray-Curtis similarity index was used in a cluster analysis to group spawning months (Clarke and Warwick, 1994). A significance level of 0.05 was used in all tests. All analyses were performed using the R software (R Core Team, 2014).
Results
General Data, Length at First Maturity and Reproductive-Load
The total number of reproduction-related cases compiled in this study was 906, including spawning, length at first maturity (Lm), minimum (Min. Lm) and maximum (Max. Lm) length at first maturity, sex ratio, and gonadosomatic index (GSI). Overall we used reproduction data from 268 sources (178 compiled in this study and 90 available in FishBase) (Table 1). The best studied LME in Brazil, regarding the reproduction of marine fishes, is South Brazil Shelf (LME#15), followed by East Brazil Shelf (LME#16), and North Brazil Shelf (LME#17) (Figure 1). It is worth to point out that the high number of spawning data for East Brazil Shelf originated from a single paper by Alves and Lima (1978), who presented spawning data for 83 species in the state of Ceará based on fishers' ecological knowledge. The number of studies on reproduction of Brazilian marine fishes rose from the 1950s to mid-2000s (Figure 2), except for a sharp decrease in the mid-1990s. Approximately 20% of all sources used originated from “gray” literature (theses, reports, and conference proceedings), while the remaining sources were scientific articles or books.
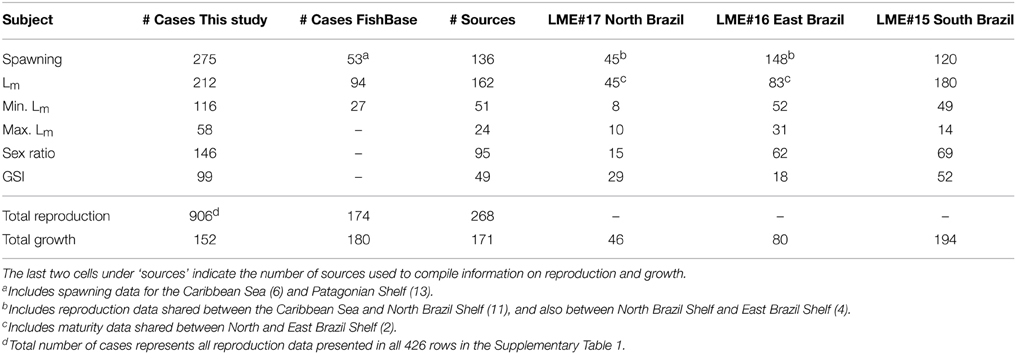
Table 1. Total number of cases on reproduction and growth compiled in this study and from FishBase: spawning months, length at first maturity (Lm), minimum and maximum length at first maturity as defined in Froese and Pauly (2000), sex ratio (F:M), gonadosomatic index (GSI), and growth parameters (L∞, K, and to).
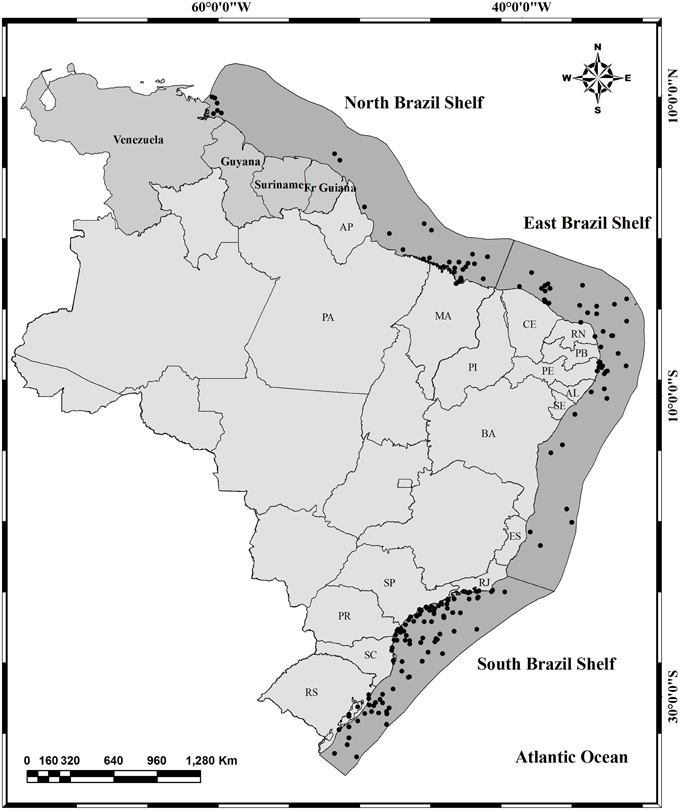
Figure 1. Map of the three Brazilian LMEs of Brazil: North Brazil Shelf, East Brazil Shelf, and South Brazil Shelf (dark gray). Black circles represent localities for which data on spawning, length at first maturity and/or sex ratio were compiled in this study or available in FishBase. In light gray are presented all Brazilian states and the coastal states are: Amapá (AP), Pará (PA), Maranhão (MA), Piauí (PI), Ceará (CE), Rio Grande do Norte (RN), Paraíba (PB), Pernambuco (PE), Alagoas (AL), Sergipe (SE), Bahia (BA), Espírito Santo (ES), Rio de Janeiro (RJ), São Paulo (SP), Paraná (PR), Santa Catarina (SC), and Rio Grande do Sul (RS).
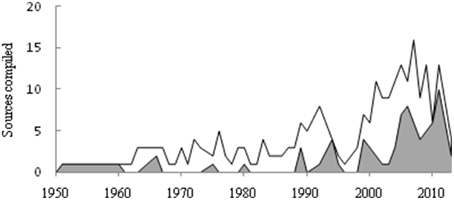
Figure 2. Number of studies on fish reproduction for Brazilian marine fishes. Language in which the studies were published: English (gray) or Portuguese (white).
In about 52% of the cases, a macroscopic method was used to define the maturity stages of the gonads. In 18% of them, a histological method was used; for the remaining data entries the method was not mentioned. For each method, different maturity scales were used for different species. Most of the histological analyses were carried out in the East Brazil Shelf (48% of all cases), followed by South Brazil Shelf (36%) and North Brazil Shelf (16%). Several values of Lm were available which allowed to estimate the reproductive load (Lm/L∞) for Brazilian marine fishes ranging from 0.20 (Trichiurus lepturus) to 0.82 (Rhizoprionodon lalandii). About 96% of the cases were between 0.3 and 0.8 (Figure 3).
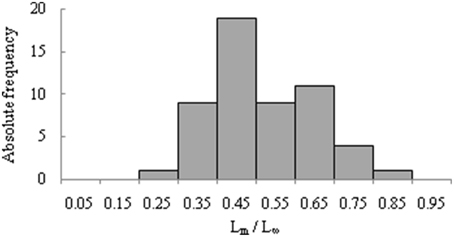
Figure 3. Frequency distribution of reproductive load (Lm/L∞) for Brazilian marine fishes.
Management policy in Brazil relies heavily on minimum landing size, associated to length at first maturity, and closed fishing seasons, even though other measures are also used. The Ministry of Fisheries and Aquaculture (MPA) and the Brazilian Institute for the Environment and Renewable Natural Resources (IBAMA) established closed seasons and/or minimum landing size for 54 species (MPA/IBAMA, 2014). Only closed fishing season was found for eight fish species: Centropomus undecimalis, Centropomus parallelus, Sardinella brasiliensis, Lutjanus purpureus, Brachyplatystoma vaillantii, Genidens genidens, and Genidens barbus. Only minimum landing size was used for 48 species. Both tools are used in different states and different time periods for three species: Pomatomus saltatrix, Centropomus undecimalis, and Centropomus parallelus.
Fish reproduction data originating from commercial landings represented about 31% of all studies for which this information was available, which decrease sampling costs. Single gears were used in 26% of the cases and multiple gears (2–4) in 13% of them. A high proportion of the studies did not present the sampling method. The most commonly used single gears were trawl net (31%), followed by gillnet, longline, beach seine, and others. Most studies measured total length (TL) of fishes, which is the most commonly used length type in fisheries science.
The studies extended for 1–19 years, with most of them carried out for 1–3 years (59%). Research proposals are funded in Brazil with a “standard” period of 3 years and this is evidenced here with the main duration of the studies. In a total of 70% of the studies that reported sampling frequency, it corresponded to monthly sampling (see Supplementary Table 1).
Analysis by Taxa
Information on reproduction of Brazilian fishes was available mainly for Perciformes, Myctophiformes, Carcharhiniformes, Clupeiformes, and Siluriformes. If we consider the proportion of species studied in relation to the total number species included in each order, we note that less than 60% of the species had at least one of their reproductive aspects analyzed. Reproduction data (i.e., sex ratio, Lm, spawning season) were available only for 21% of all 1207 marine species reported for Brazil in FishBase (Figure 4). Species with more data available for the entire Brazilian coast were: Macrodon ancylodon, Sardinella brasiliensis, Lutjanus purpureus, Micropogonias furnieri, and Rhizoprionodon lalandii. For North Brazil Shelf, Macrodon ancylodon and Lutjanus purpureus were the most studied species. Lutjanus purpureus and Alphestes afer were the most studied in the East Brazil Shelf, and Macrodon ancylodon and Sardinella brasiliensis in the South Brazil Shelf (Table 2). All these species are of commercial importance in their respective ecosystems.
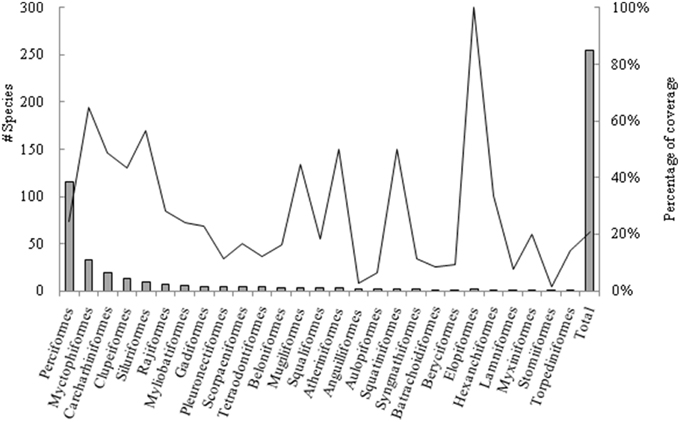
Figure 4. Number of marine fish species studied in terms of reproduction in Brazil per order (columns) and percentage of the number of species studied in relation the total number of species within each order (line).
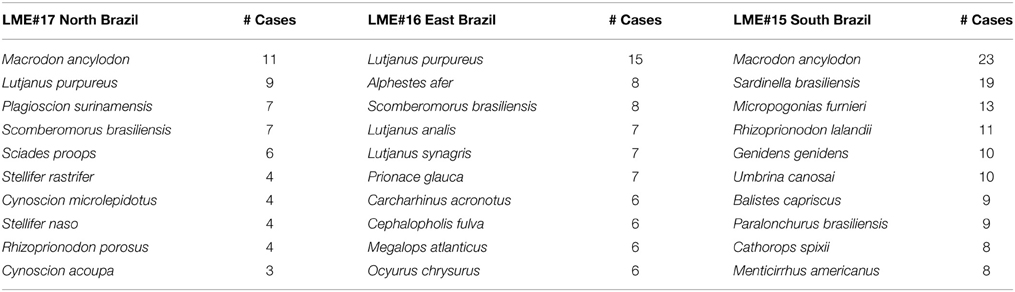
Table 2. Number of cases for the most studied species in each Brazilian Large Marine Ecosystem.
For the most studied species, i.e., Macrodon ancylodon, we found 24 values of length at first maturity ranging from 18.6 cm in the North Brazil Shelf (state of Pará) to 29.0 cm in the South Brazil Shelf (states of Rio de Janeiro and São Paulo). The minimum landing size allowed for M. ancylodon is 25 cm for the entire Brazilian coast. However, some studies showed a higher length at first maturity compared with the minimum allowed landing size. The same scenario is observed for other species. We compared the compiled values of length at first maturity with all values of minimum landing size found for marine fishes in the Brazilian coast. In about 65% of the cases, minimum landing size was larger than the length at first maturity (Lm). In 27% of the cases, the values of minimum catch size established do not allow fishes to reproduce at least once in their lifetime: Genidens barbus, Macrodon ancylodon, Micropogonias furnieri, Mustelus fasciatus, Mycteroperca bonaci, Opisthonema oglinum, Prionotus punctatus, Sphyrna lewini, Trichiurus lepturus, Umbrina canosai, and Urophycis mystacea. Finally, in about 8% of the cases, these two parameters are exactly the same.
Twenty-seven orders were associated with at least one reproductive study. The most abundant orders without any reproductive information were: Ophidiiformes (47 species), Osmeriformes (18 species), Lophiiformes (14 species), Gobiesociformes (7 species), Lampriformes and Zeiformes (5 species each), and eight more orders with less than 5 species.
Spawning Season
The onset of the spawning season is usually defined by gonadosomatic indices. A total of 104 studies of GSI were compiled for Brazilian marine waters. About 83% of the studies used the standardized equation (a). Comparing all available GSI data from studies that reported GSI values for both sexes, we observed that for 35 out of 36 species females had higher GSI values than males. The maximum differences were observed for Cathorops spixii, followed by Cynoscion microlepidotus and Genidens genidens. The smallest differences between GSI for females and males were found for Diplodus argenteus argenteus, Maurolicus stehmanni, and Pomatomus saltatrix.
The median extension of the spawning season in the North, East and South Brazil Shelf were 6.5, 6.0, and 5.0 months, respectively (Figure 5). The Mood median test showed a statistical difference among these medians (χ2 = 7.213, p = 0.027). However, the Tukey multiple comparison tests were not able to detect where the differences were (Table 3). A total of 72% of the species were partial spawners (see Supplementary Table 1).
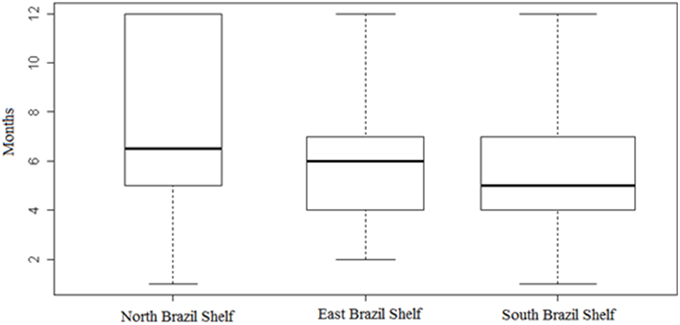
Figure 5. Number of months over which the spawning season extends for Brazilian marine fishes. Horizontal central lines show medians, boxes correspond to the interquartile range, and whiskers are maximum and minimum values.

Table 3. Multiple comparison Tukey test for differences among medians for duration of spawning season (SE = standard error = 3.791; q = standardized difference; q0.05,3 = 4.622).
In general spawning of marine fishes occurs all year around in Brazilian waters (Figure 6). However, a seasonal pattern was clear for the South Brazil Shelf, a subtropical region where the spawning peak occurred during the austral summer (Figure 6C). Cases compiled from Alves and Lima (1978), who used fishers' ecological knowledge to define the spawning season were excluded from our analysis to avoid bias (a spawning season of two months was considered for all species cited in that reference).
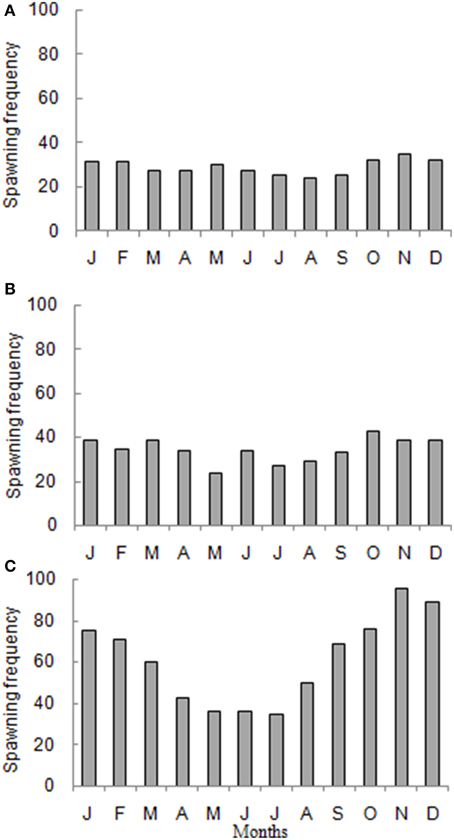
Figure 6. Percentage frequency of the number of species in spawning activity in each month for marine fishes in (A) North Brazil Shelf, (B) East Brazil Shelf, and (C) South Brazil Shelf.
A cluster analysis showed that Brazilian marine fishes can be grouped in two spawning groups: summer spawners and non-summer spawners (Figure 7). Summer spawners are species that usually spawn between November and March, i.e., Xiphias gladius, Macrodon ancylodon, Balistes capriscus, Etropus longimanus, and Cathorops spixii. Non-summer spawners include species that spawn between April and October such as Mugil curema, Cynoscion striatus, Atlantoraja platana, Holocentrus adscensionis, and Scomberomorus brasiliensis. A total of 25 species presented continuous spawning: 12 species in North Brazil Shelf, 4 in East Brazil Shelf, and 11 in South Brazil Shelf (total does not add up to 25 due to studies encompassing more than one LME). Most of these species belonged to the order Perciformes: Macrodon ancylodon, Lutjanus purpureus, Scomberomorus brasiliensis, Plagioscion surinamensis, Stellifer rastrifer, and Nicholsina ustausta. This order was followed by Clupeiformes, Rajiformes, Carcharhiniformes, Pleuronectiformes, and Stomiiformes.
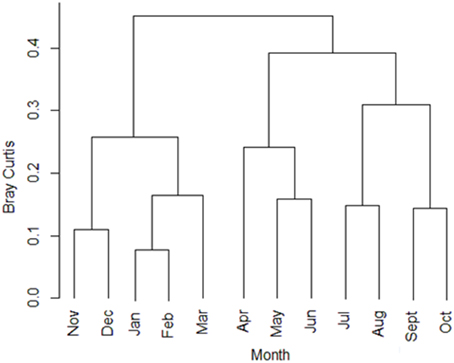
Figure 7. Cluster analysis of spawning months over which Brazilian marine fishes reproduce (based on Bray-Curtis similarity matrix).
Sex Ratio (F:M)
Sex ratio is important information to assess changes in the structure of populations and their reproductive potential. This ratio may vary during the life cycle due to successive events acting differently over individuals of each sex (Vazzoler, 1996). Even though the number of cases for this ratio was low, mainly for North Brazil Shelf, a higher number of females were observed in all three LMEs (65% of all cases) (Figure 8). The ultimate cases were Zapteryx brevirostris (F:M = 17.0), Urophycis brasiliensis (9.1), Cephalopholis fulva (5.8), Squalus megalops (5.6), and Squalus mitsukurii (4.6). On the other hand, we had a few cases where males were more abundant, with more extreme examples such as: Carcharhinus limbatus (F:M = 0.2), Prionace glauca (0.2), Lutjanus analis (0.3), Rhizoprionodon porosus (0.3), Galeocerdo cuvier (0.4), and Mustelus canis (0.4).
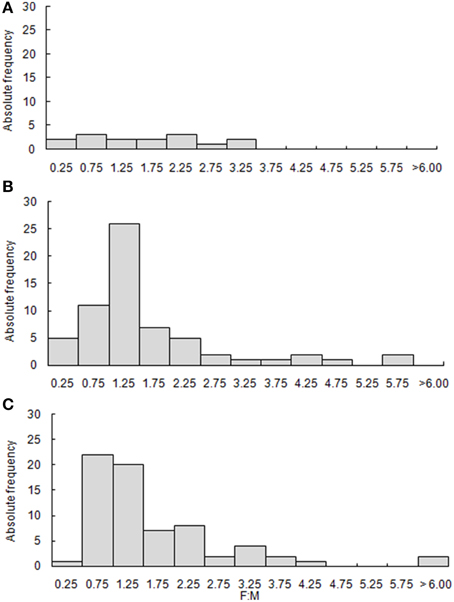
Figure 8. Frequency distribution of the number of females (F) in relation to the number of males (M) sampled for the three Brazilian LMEs. (A) North Brazil Shelf; (B) East Brazil Shelf; and (C) South Brazil Shelf.
Discussion
Knowledge on fish population dynamics is crucial to understand how fishes react under anthropogenic pressure. The management of exploited fish populations is possible only if basic biological data is available, particularly reproduction and growth data. This analysis showed that studies about reproduction of marine fishes are more concentrated in the South Brazil Shelf and that only a small percentage of all marine species have more than minimum information on their reproduction. Overcoming this situation requires more attention from universities, government, granting institutions, and industry. A proper management system based on sound biological information should be of interest for all sectors, as the wealth of the stocks benefits all of them.
The observed values of reproductive load for the three LMEs studied of 0.3–0.8 are slightly shifted toward smaller values when compared with the range of 0.4–0.9 compiled by Froese and Binohlan (2000). The reproductive load is usually smaller in large fishes and larger in smaller fishes (Froese and Binohlan, 2000; Froese and Pauly, 2000). We should consider in this analysis Lm and L∞ parameters estimated for the same population, but usually they are not available. Thus, information was used for population preferentially in similar areas or otherwise in any other areas. Furthermore, environmental and fishery-related differences may affect the reproductive load as estimated here. The relative constancy of the Lm/L∞ is due to the fact that fishes have to reach a critical size before they are able to perceive environmental stimuli that induce them to spawn (Pauly, 2010). For those cases where Lm is currently unavailable for Brazilian waters, we may use the empirical equation [log10 (Lm) = −0.0782 + 0.8979·(log10L∞)] developed by Froese and Binohlan (2000) until local data is generated. This could be used by recreational fishers, for example, as many species caught do not have a minimum landing size defined yet (Freire et al., 2012).
However, it is important to investigate those orders above mentioned that have no information available on the reproduction of any species, as many of these species may be landed in Brazilian waters under general common names (Freire and Pauly, 2005) or have been discarded with no record. These results show that scientific efforts should be applied to better cover the basic biological information at least for some species of these orders that are caught in Brazilian waters, as fishing pressure on target species also impacts by-catch species (Daan, 1987; Magnússon, 1995; Jakobsen et al., 2009). And this may be more informative than continuing spending effort in studying reproduction of very well known orders and families within.
Fish populations are assessed and managed using length at first maturity and fecundity data to estimate the spawning stock biomass (SSB). The histological analysis of the gonads provides a more accurate estimation of those parameters (Vitale et al., 2006). Thus, fishery biologists should use standardized histological methods associated with macroscopic analysis to define maturity stages (Kjesbu et al., 2003; Vitale et al., 2006). However, traditional macroscopic analysis has been the most used method in developing countries despite being less accurate than histological analysis (Murua et al., 2003) due to lower costs involved and easier and faster results. Other methodological issue to be considered here is the selectivity of the gears. Highly selective gears such as gillnet and longline may lead to the estimation of biased parameters such as length at first maturity. Thus, priority should be given to studies using less selective gears such as trawl nets.
Minimum legal size limits are usually site-specific. This is appropriate, as the Brazilian coast spans a wide latitudinal range, from about 4°N (Cape Orange) to 33°S (Chuí), which generates differences in the reproductive patterns of the marine fishes among its three LME. Besides, size at first maturity usually presents differences between sexes and this may not be properly captured in the legislation (Tsikliras and Stergiou, 2014). The establishment of a “maximum size” could be an efficient management tool to protect the reproductive potential of the stocks (Froese, 2004) and has been recently discussed among recreational fishers for managing Brazilian stocks.
The absence of a seasonal pattern in the North Brazil Shelf, a tropical region, may be associated with the low number of data points compiled for that ecosystem. The East Brazil Shelf is essentially a tropical area, but includes two portions influenced by different currents (Ekau and Knoppers, 1999, 2003). This ecosystem also did not show a seasonal pattern as suggested by Longhurst and Pauly (2007). In these two cases, the absence of a seasonal pattern may also be influenced by improper definition of the study area or by the use of information originating from landings. However, the last factor was minimized using information related to fishing grounds and not to landing ports. Besides, the use of LMEs decreases the influence of such differences between fishing grounds and landing ports due to the larger extension of their areas.
Sex ratio is important information to assess the structure of populations and their reproductive potential. Based on this study, there is a trend of predominance of females along the Brazilian coast, with predominantly higher GSI. Changes in sex ratio and GSI should be further investigated to verify how fisheries are affecting the fish populations, when a higher volume of data is available for each of the ecosystems analyzed. We also have to be aware that even a finer spatial scale may be necessary as some stocks have a distribution area smaller than the LMEs used here.
The Lesser guitarfish, Zapteryx brevirostris, with a sex ratio of 17.0:1 (F:M), represented an outlier. A higher proportion of females (4.2:1) for this species was also found by Abilhoa et al. (2007) in the same area. It is worth pointing out that five out of six cases with the largest proportion of males are sharks. For C. limbatus, Castillo-Géniz et al. (1998) also found a predominance of males (F:M = 0.38) in the Gulf of Mexico when dealing with landings originating from artisanal fisheries (gillnetters and longliners). For P. glauca, Castro and Mejuto (1995) found similar predominance of males, but Beerkircher et al. (2002) found an opposite ratio, with heavy predominance of females when dealing with data originating from longliners off the southeastern United States. Castro and Mejuto (1995) noticed a slight increase in the proportion of females from west to east off the western African coast, which would be comparable to our results. For R. porosus, Dallos et al. (2012) also found predominance of males when analyzing data from the Colombian Caribbean. Several factors are attributed to sex ratios that diverge from 1:1, as differential growth, differential mortality, migration, different distribution range, influence of gear selectivity, exploitation rate, discards, and temperature, even though differential growth rates have been identified by some authors as the main factor (Castro and Mejuto, 1995; Montealegre-Quijano and Vooren, 2009).
Lessa et al. (2004) called attention to the fact that differential growth rates have been used as an explanation even without empirical evidence in some regions. The effects of fishing pressure, along with biotic and abiotic factors, upon the structure of fish populations are widely studied (Jakobsen et al., 2009; Kleisner et al., 2010). Species that exhibit differential growth are more vulnerable to size selective gears, which may lead to unbalanced sex-ratios due to differences in fishing mortality rates (Fogarty and O'brien, 2009). With respect to elasmobranchs, the problem is more complex due to the large number of individuals discarded (Beerkircher et al., 2002). Conover and Voorhees (1990) pointed that differences of sex ratio in a given fish population tend to a balanced sex ratio through generations. These authors also indicated that high temperature observed during the late breeding season may cause most offspring produced to become males (observed for Menidia menidia under controlled experiments). Testing all these factors requires a high volume of data and it is also difficult to isolate the effect of confounding effects.
Even though there was one exception, we corroborated here the general pattern of higher gonadosomatic indices for females, as also found for Mediterranean fishes (Tsikliras et al., 2010). Hutchings (2002) pointed out that different anthropogenic pressure on different stocks of a given species leads to significant differences in GSI values. Macrodon ancylodon was the species with the highest number of GSI values compiled for Brazilian waters to test the effect suggested by Hutchings. From 1996 to 2006, GSI values for females (0.023) and males (0.004) increased to 0.74 and 0.13, respectively (using the same equation for GSI in both periods). We can infer that this change in GSI values for M. ancylodon in the North Brazil Shelf may be attributed to fishing pressure, as this species has a high commercial importance in this area (Camargo and Isaac, 2005; Santos, 2007). The larger differences found for marine catfishes, Cathorops spixii and Genidens genidens, are associated with the reproductive behavior of these two species, as the large oocyte size is related with the low fecundity and parental care (Vazzoler, 1996; Gomes et al., 1999; Favaro et al., 2005). On the other hand, Elagatis bipinnulata had a completely inverse pattern with a GSI of 33.8 for males and 31.8 for females (equation b).
Spawning season information and length at first maturity should be collected at different spatial and temporal scales to allow for the analysis of the influence of fisheries upon wild fish stocks in different ecosystems. As it stands up to now, the number of data available throughout the three LMEs is not high enough to allow for a proper comparison. However, some general patterns were clear, such as the predominance of females, which usually have higher investment in reproduction, spawning peak during the summer in the South Brazil Shelf, low use of information available on reproduction by managing authorities, missing information on reproduction for some orders, which deserve some attention that would lead to better understanding of reproduction patterns (cost-effectiveness), and mainly a mixture of methodologies used in the studies analyzed here that may influence the absence of clear seasonality in spawning for the East Brazil Shelf. It is important that all existing information compiled here is widely available and FishBase plays an important role in disseminating such information mainly for developing countries. Thus, all information gathered during this study will be incorporated in that database. We hope our results may lead into a more efficient use of the results of fish reproduction studies by managers responsible for the health of fish stocks in the three LMEs analyzed here.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to acknowledge Victoria Judith Isaac Nahum and Roberto Licandeo for providing references; Bruna Larissa Ferreira de Carvalho, Maria Camila dos Santos Bispo, Thaisa Accioly de Souza, and Danyhelton Douglas Dantas for helping with the data compilation; to Ronaldo Angelini for his collaboration with the FishBase project; to the FishBase team in Philipines, particularly to Emily Capuli, Crispina Binohlan, and Josephine Barile, for providing FishBase data; to the Ministry of Fisheries and Aquaculture (MPA) for partially funding this study; to the National Council for the Scientific and Technological Development (CNPq) for provinding a PIBIC scholarship for the first author; to Daniel Pauly, Ana Rosa da Rocha Araújo and Roberto Schwarz Jr. for reviewing an earlier version of this manuscript; and to Paulo T. C. Chaves and Joanne Morgan for their revision, which helped to improve the quality of this paper.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmars.2015.00038/abstract
References
Abdallah, P. R., and Sumaila, U. R. (2007). An historical account of Brazilian public policy on fisheries subsidies. Mar. Policy 31, 444–450. doi: 10.1016/j.marpol.2007.01.002
Abilhoa, V., Bornatowski, H., and Freitas, M. O. (2007). Some information on reproduction and embryonic development of the lesser guitarfish Zapteryx brevirostris in Southern Brazil. Acta Adriat. 48, 185–190.
Alves, D. C., and Minte-Vera, C. V. (2012). Scientometric analysis of freshwater fisheries in Brazil: repeating past errors? Rev. Fish Bio. Fisher. 23, 113–126. doi: 10.1007/s11160-012-9282-6
Alves, M. I. M., and Lima, H. H. (1978). Sobre a época de desova de alguns peixes marinhos no estado do Ceará, Brasil. Bolm. Ciênc. Mar. 30, 1–7.
Beerkircher, L. R., Cortés, E., and Shivji, M. (2002). Characteristics of shark bycatch observed on pelagic longlines off the southeastern United States, 1992-2000. Mar. Fish. Rev. 64, 40–49.
Borenstein, M., Hedges, L. V., Higgins, J. P. T., and Rothstein, H. R. (2009). Introduction to Meta-Analysis. Chichester, UK: John Wiley & Sons, Ltd. doi: 10.1002/9780470743386
Bye, V. J. (1984). “The role of environmental factors in the timing of reproductive cycles,” in Fish Reproduction: Strategies and Tactics, eds G. W. Potts and R. J. Wootton. (New York, NY: Academic Press), 187–205.
Camargo, M., and Isaac, V. J. (2005). Reproductive biology and spatio-temporal distribution of Stellifer rastrifer, Stellifer naso and Macrodon ancylodon (Sciaenidae) in the Caeté estuary, northern Brazil. Braz. J. Oceanogr. 53, 13–21. doi: 10.1590/S1679-87592005000100002
Castello, J. P. (2007). Gestão sustentável dos recursos pesqueiros, isto é realmente possível? PanamJAS 2, 47–52.
Castillo-Géniz, J. L., Márquez-Farias, J. F., Rodriguez de la Cruz, M. C., Cortés, E., and Cid del Prado, A. (1998). The Mexican artisanal shark fishery in the Gulf of Mexico: towards a regulated fishery. Mar. Freshwater Res. 49, 611–620. doi: 10.1071/MF97120
Castro, J. A., and Mejuto, J. (1995). Reproductive parameters of blue shark, Prionace glauca, and other sharks in the Gulf of Guinea. Mar. Freshwater Res. 46, 967–173. doi: 10.1071/MF9950967
Clarke, K. R., and Warwick, R. M. (1994). Change in Marine Communities: an Approach to Statistical Analysis and Interpretation. Plymouth: Plymouth Marine Laboratory.
Conover, D. O., and Voorhees, D. A. V. (1990). Evolution of a balanced sex ratio by frequency-dependent selection in a fish. Science 250, 1556–1559. doi: 10.1126/science.250.4987.1556
Cushing, D. H. (1973). Dependence of recruitment on parent stock. J. Fish. Res. Board Can. 30, 1965–1976. doi: 10.1139/f73-320
Cushing, D. H. (1990). “Plankton production and year-cass strength in fish populations: an update of the match/mismatch hypothesis,” in Advances in Marine Biology, Vol. 26, eds J. H. S. Blaxter and A. J. Southward (London: Academic Press Limited), 249–293.
Daan, N. (1987). Multispecies versus single-species assesment of North Sea fish stocks. Can. J. Fish. Aquat. Sci. 44, 360–370. doi: 10.1139/f87-337
Dallos, I. M., Álvarez, M. N., and Acerp, P. A. (2012). Aspectos biológicos de Rhizoprionodon lalandii y Rhizoprionodon porosus (Carcharhinidae - Carcharhiniformes) capturados mediante la pesca artesanal en Isla Fuerte, Caribe Colombiano. Bol. Invest. Mar. Cost. 41, 179–191.
Dias Neto, J. (2010a). Gestão do uso dos Recursos Pesqueiros Marinhos No Brasil. Brasília: Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis.
Dias Neto, J. (2010b). Pesca no Brasil e seus aspectos institucionais - um registro para o futuro. Rev. CEPSUL Biodivers. Cons. Mar. 1, 66–80.
Ekau, W., and Knoppers, B. (1999). An introduction to the pelagic system of the North-East and East Brazilian shelf. Arch. Fish. Mar. Res. 47, 113–132.
Ekau, W., and Knoppers, B. A. (2003). “A review and re-definition of the large marine ecosystems of Brazil,” in Large Marine Ecosystems of the World, eds G. Hempel and K. Sherman (Amsterdam: Elsevier Science), 234–248.
Favaro, L. F., Frehse, F. A., Oliveira, R. N., and Schwarz-Júnior, R. (2005). Reprodução do bagre amarelo, Cathorops spixii (Agassiz) (Siluriformes, Ariidae), da Baía de Pinheiros, região estuarina do litoral do Paraná, Brasil. Rev. Bras. Zool. 22, 1022–1029. doi: 10.1590/S0101-81752005000400030
Ferraz-Dias, J. (1989). Padrões Reprodutivos em Teleósteos da Costa Brasileira: uma Síntese. Master thesis, Universidade de São Paulo, São Paulo.
Fogarty, M. J., and O'brien, L. (2009). “Recruitment in marine fish populations,” in Fish Reproductive Biology: Implications for Assessment and Management, eds T. Jakobsen, M. J. Fogarty, B. A. Megrey, and E. Moksness (Oxford: Blackwell), 11–47. doi: 10.1002/9781444312133.ch1
Freire, K. M. F., Machado, M. L., and Crepaldi, D. (2012). Overview of inland recreational fisheries in Brazil. Fisheries 37, 484–493. doi: 10.1080/03632415.2012.731867
Freire, K. M. F., and Pauly, D. (2005). Richness of common names of Brazilian marine fishes and its effect on catch statistics. J. Ethnobiol. 25, 279–296. doi: 10.2993/0278-0771(2005)25[279:ROCNOB]2.0.CO;2
Froese, R. (2004). Keep it simple: three indicators to deal with overfishing. Fish Fish. 5, 86–91. doi: 10.1111/j.1467-2979.2004.00144.x
Froese, R., and Binohlan, C. (2000). Empirical relationships to estimate asymptotic length, length at first maturity and length at maximum yield per recruit in fishes, with a simple method to evaluate length frequency data. J. Fish Biol. 56, 758–773. doi: 10.1111/j.1095-8649.2000.tb00870.x
Froese, R., and Pauly, D. (2000). FishBase 2000: Concepts, Design and Data Sources. Los Baños: ICLARM.
Froese, R., and Pauly, D. (2015). FishBase. World Wide Web Electronic Publication. Available online at: http://www.fishbase.org (Accessed April 2015).
Godinho, A. L., and Lamas, I. R. (2010). Reproductive ecology of Brazilian freshwater fishes. Environ. Biol. Fish. 87, 143–162. doi: 10.1007/s10641-009-9574-4
Gomes, I. D., Araújo, F. G., Azevêdo, M. C. C., and Passanha, A. L. M. (1999). Biologia reprodutiva dos bagres marinhos Genidens genidens (Valenciennes) e Cathorops spixii (Agassiz) (Siluriformes, Ariidae), na Baía de Sepetiba, Rio de Janeiro, Brasil. Rev. Bras. Zool. 16, 171–180. doi: 10.1590/S0101-81751999000600017
Hutchings, J. A. (2002). “Life histories of fish,” in Handbook of Fish Biology and Fisheries. Vol.1. Fish Biology, eds P. J. B. Hart and J. D. Reynolds (Malden, MA: Blackwell Pub.), 149–174.
Isaac-Nahum, V. J., and Vazzoler, A. E. A. M. (1987). Biologia reprodutiva de Micropogonias furnieri (Desmarest, 1823) (Teleostei, Sciaenidae). 2. Relação gonadossomática, comprimento e peso dos ovários como indicadores do período de desova. Bol. Inst. Oceano. 35, 123–134.
Jakobsen, T., Fogarty, M. J., Megrey, B. A., and Moksness, E. (2009). Fish Reproductive Biology: Implications for Assessment and Management. Oxford: Blackwell.
Kinas, P. G. (1996). Bayesian fishery stock assessment and decision making using adaptive importance sampling. Can. J. Fish. Aquat. Sci. 53, 414–423. doi: 10.1139/f99-076
Kjesbu, O. S., Hunter, J. R., and Witthames, P. R. (2003). Report of the Working Group on Modern Methods to Assess Maturiy and Fecundity in Warm- and Cold-water Fish and Squids. eds O. S. Kjesbu, J. R. Hunter and P. R. Witthames (Bergen: Fisken og Havet).
Kleisner, K. M., Walter, J. F. I., Diamond, S. L., and Die, D. J. (2010). Modeling the spatial autocorrelation of pelagic fish abundance. Mar. Ecol. Prog. Ser. 411, 203–213. doi: 10.3354/meps08667
Lessa, R., Santana, F. M., and Hazin, F. H. (2004). Age and growth of the blue shark Prionace glauca (Linnaeus, 1758) off northeastern Brazil. Fish. Res. 66, 19–30. doi: 10.1016/S0165-7836(03)00193-0
Lowerre-Barbieri, S. K., Ganias, K., Saborido-Rey, F., Murua, H., and Hunter, J. R. (2011). Reproductive timing in marine fishes: variability, temporal scales, and methods. Mar. Coast. Fish. 3, 71–97. doi: 10.1080/19425120.2011.556932
Magnússon, K. G. (1995). An overview of the multispecies VPA - theory and applications. Rev. Fish Biol. Fisher. 5, 195–212. doi: 10.1007/BF00179756
Montealegre-Quijano, S., and Vooren, C. M. (2009). Estimativas de mortalidade e tabelas de vida do tubarão-azul, Prionace glauca (Carcharhinidae), no sul do Brasil e águas internacionais adjacentes. Panam JA. S. 4, 476–489.
MPA/IBAMA (2014). Ministério da Pesca e Aquicultura - Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis Available online at: http://www.mpa.gov.br/ (Accessed February 2014).
Murua, H., Kraus, G., Saborido-Rey, F., Witthames, P. R., Thorsen, A., and Junquera, S. (2003). Procedures to estimate fecundity of marine fish species in relation to their reproductive strategy. J. Northwest Atl. Fish. Sci. 33, 33–54. doi: 10.2960/J.v33.a3
Pauly, D. (2010). “Gasping fish and panting squids: oxygen, temperature and the growth of water-breathing animals,” in Excellence in Ecology. Book 22, ed O. Kinne (Oldendorf/Luhe: International Ecology Institute), 216.
R Core Team (2014). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing. Available online at: http://www.R-project.org/ (Accessed February, 2014)
Santos, N. B. (2007). Biologia Reprodutiva de Peixes Cianídeos Capturados Nas Proximidades dos Terminais Portuários do Pará e Maranhão. Master Thesis, Universidade Federal do Pará, Belém.
Schaefer, M. B., and Orange, C. J. (1956). Studies of the sexual development and spawning of yellowfin tuna (Neothunnus macropterus) and skipjack (Katsuwonus pelamis) in the three areas of the Eastern Pacific Ocean, by examination of gonads. Inter. Amer. Trop. T. 1, 281–302.
Stergiou, K. I., and Tsikliras, A. C. (2006). Underrepresentation of regional ecological research output by bibliometric indices. Ethics Sci. Environ. Polit. 2006, 15–17.
Tsikliras, A. C., Antonopoulou, E., and Stergiou, K. I. (2010). Spawning period of Mediterranean marine fishes. Rev. Fish. Biol. Fisher. 20, 499–538. doi: 10.1007/s11160-010-9158-6
Tsikliras, A. C., and Stergiou, K. I. (2014). Size at maturity of Mediterranean marine fishes. Rev. Fish. Biol. Fisher. 24, 219–268. doi: 10.1007/s11160-013-9330-x
Vazzoler, A. E. A. M. (1996). Biologia da Reprodução de Peixes Teleósteos: Teoria e Prática. Maringá: Nupelia.
Vitale, F., Svedäng, H., and Cardinale, M. (2006). Histological analysis invalidates macroscopically determined maturity ogives of the Kattegat cod (Gadus morhua) and suggests new proxies for estimating maturity status of individual fish. ICES J. Mar. Sci. 63, 485–492. doi: 10.1016/j.icesjms.2005.09.001
Keywords: fish reproduction, population dynamics, sex ratio, size at first maturity, spawning season, gonadosomatic index, reproductive load
Citation: Trindade-Santos I and Freire KMF (2015) Analysis of reproductive patterns of fishes from three Large Marine Ecosystems. Front. Mar. Sci. 2:38. doi: 10.3389/fmars.2015.00038
Received: 13 March 2015; Accepted: 23 May 2015;
Published: 08 June 2015.
Edited by:
Athanassios C. Tsikliras, Aristotle University of Thessaloniki, GreeceReviewed by:
Paulo T. C. Chaves, Federal University of Paraná, BrazilJoanne Morgan, Fisheries and Oceans Canada, Canada
Copyright © 2015 Trindade-Santos and Freire. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kátia de Meirelles Felizola Freire, Laboratório de Ecologia Pesqueira, Departamento de Engenharia de Pesca e Aquicultura, Universidade Federal de Sergipe, Rua Mal. Rondon S/N, Jardim Rosa Elze, São Cristóvão, 49100-000 Sergipe, Brazil,a21mZnJlaXJlMjAxNUBnbWFpbC5jb20=