 Richard J. Sanders1
Richard J. Sanders1 Stephanie A. Henson1*
Stephanie A. Henson1* Adrian P. Martin1Tom R. Anderson1Raffaele Bernardello1Peter Enderlein2Sophie Fielding2
Adrian P. Martin1Tom R. Anderson1Raffaele Bernardello1Peter Enderlein2Sophie Fielding2 Sarah L. C. Giering1
Sarah L. C. Giering1 Manuela Hartmann1Morten Iversen3
Manuela Hartmann1Morten Iversen3 Samar Khatiwala4
Samar Khatiwala4 Phyllis Lam5Richard Lampitt1Daniel J. Mayor1
Phyllis Lam5Richard Lampitt1Daniel J. Mayor1 Mark C. Moore5Eugene Murphy2Stuart C. Painter1
Mark C. Moore5Eugene Murphy2Stuart C. Painter1 Alex J. Poulton1Kevin Saw1Gabriele Stowasser2Geraint A. Tarling2Sinhue Torres-Valdes1
Alex J. Poulton1Kevin Saw1Gabriele Stowasser2Geraint A. Tarling2Sinhue Torres-Valdes1 Mark Trimmer6George A. Wolff7
Mark Trimmer6George A. Wolff7 Andrew Yool1Mike Zubkov1
Andrew Yool1Mike Zubkov1- 1National Oceanography Centre, Southampton, UK
- 2British Antarctic Survey, Cambridge, UK
- 3MARUM - Center for Marine Environmental Sciences, University of Bremen, Bremen, Germany
- 4Department of Earth Sciences, University of Oxford, Oxford, UK
- 5Ocean and Earth Science, National Oceanography Centre Southampton, University of Southampton, Southampton, UK
- 6School of Biological and Chemical Sciences, Queen Mary University of London, London, UK
- 7School of Environmental Sciences, University of Liverpool, Liverpool, UK
The ocean's biological carbon pump plays a central role in regulating atmospheric CO2 levels. In particular, the depth at which sinking organic carbon is broken down and respired in the mesopelagic zone is critical, with deeper remineralization resulting in greater carbon storage. Until recently, however, a balanced budget of the supply and consumption of organic carbon in the mesopelagic had not been constructed in any region of the ocean, and the processes controlling organic carbon turnover are still poorly understood. Large-scale data syntheses suggest that a wide range of factors can influence remineralization depth including upper-ocean ecological interactions, and interior dissolved oxygen concentration and temperature. However, these analyses do not provide a mechanistic understanding of remineralization, which increases the challenge of appropriately modeling the mesopelagic carbon dynamics. In light of this, the UK Natural Environment Research Council has funded a programme with this mechanistic understanding as its aim, drawing targeted fieldwork right through to implementation of a new parameterization for mesopelagic remineralization within an IPCC class global biogeochemical model. The Controls over Ocean Mesopelagic Interior Carbon Storage (COMICS) programme will deliver new insights into the processes of carbon cycling in the mesopelagic zone and how these influence ocean carbon storage. Here we outline the programme's rationale, its goals, planned fieldwork, and modeling activities, with the aim of stimulating international collaboration.
Motivation
The oceanic production, export to depth and remineralization of organic carbon constitutes the biological carbon pump, the suite of processes that currently store enough carbon in the ocean's interior to keep atmospheric CO2 200 ppm lower than it would otherwise be (Parekh et al., 2006). The size of this stored carbon pool is dictated by the sinking flux of organic carbon into the ocean interior and its residence time in the deep ocean, which is a function of the depth at which sinking carbon is remineralized (Kwon et al., 2009), with deeper remineralization leading to increased storage.
Multiple mechanisms contribute to the flux of organic matter to the ocean interior. The majority of studies focus on sinking particles, such as fecal pellets, marine snow aggregates, and individual organisms. Fecal pellets play a significant role at some times of the year, with their flux to the seafloor on occasions representing up to 70% of the total flux (Manno et al., 2015). Aggregates can also contribute large fluxes, especially in the aftermath of diatom blooms (e.g., Martin et al., 2011). The assumption is often made, based on Bishop et al. (1977), that the bulk of the flux is carried by large, rapidly sinking aggregates. However, recent observations (Alonso-Gonzalez et al., 2010; Riley et al., 2012; Villa-Alfageme et al., 2014) suggest that, on occasion, a significant fraction of flux sinks much more slowly. Our understanding of this slow sinking fraction is in its infancy, although a model study suggests that its importance may be widespread, with knowledge of its variability key to understanding what fraction of primary production is exported, i.e., the export efficiency (Henson et al., 2015).
A range of other processes also transfer organic carbon into the ocean interior including advection and mixing of dissolved organic carbon (DOC), diel vertical migration (DVM) of zooplankton, and production by chemosynthesis. Whilst DOC transport is a surface intensified process, chemosynthesis and DVM have the potential to directly supply organic carbon to deeper waters. DOC transport can form a significant (~50%) fraction of total export flux in the oligotrophic subtropical gyres (Carlson et al., 2010; Pan et al., 2014). During DVM, zooplankton swim to the surface at night to feed and then return to depth during the day where they respire and egest some of the material they have ingested at the surface and release some of it as DOC and particulate organic carbon (POC). This is the so called “active flux” which can be a significant term in mesopelagic carbon budgets (equivalent to 2–40% of the gravitational POC flux at 150 m; Steinberg et al., 2008; Giering et al., 2014). Chemosynthesis—new production by chemolithotrophic bacteria and archaea, which may be enhanced in anoxic regions (Juniper and Brinkhurst, 1986)—uses reduced inorganic substrates (e.g., sulfides and ammonia) as an energy source and CO2 as a carbon source. Chemosynthesis may significantly contribute to the mesopelagic carbon budget (Alonso-Saez et al., 2010; Herndl and Reinthaler, 2013), with rates of chemoautotrophic CO2 fixation comparable to heterotrophic production (Reinthaler et al., 2010).
Because of the complexity and multitude of processes contributing to export flux and remineralization, closing the mesopelagic carbon budget has proved challenging until recently, with estimates of organic carbon sources and sinks differing by up to an order of magnitude (Steinberg et al., 2008; Burd et al., 2010). However, in 2014, the first balanced oceanic mesopelagic carbon budget was published for the Northeast Atlantic Ocean (Giering et al., 2014, hereafter G14). A key result of G14 was that, whereas both zooplankton and microbes consume sinking particles in approximately equal measure, the zooplankton serve primarily to fragment this material. It is possible that this fragmentation increases the material's nutritive value by promoting the growth of microbes (Mayor et al., 2014). G14 demonstrated that a consequence of the fragmentation is that respiration by prokaryotes is the dominant sink for organic carbon.
Whilst G14 demonstrated that we now have the tools to understand and measure mesopelagic processes, it only considered a single site in the North Atlantic and only balanced the budget over the entire mesopelagic zone, probably due to inadequate representation of DVM or to uncertainties associated with the calculation of bacterial respiration. Given this state of the research field, we identify the following priorities for biological carbon pump research:
(1) To achieve a mechanistic understanding of the key processes in, and overall function of, the mesopelagic system;
(2) To close the budget (carbon supply vs. carbon demand) at other, contrasting locations;
(3) To balance the budget in discrete layers within the mesopelagic to identify the vertical phasing and interplay of processes;
(4) To represent mesopelagic remineralization dynamically in IPCC class models in order to improve simulations of ocean carbon storage.
Controls on Mesopelagic Remineralization
Most efforts to understand the large scale controls on interior remineralization have focussed on the sinking flux, quantifying its loss via the transfer efficiency (TE; flux at depth relative to flux at the base of the mixed layer), calculated from surface observations and deep trap flux data. Francois et al. (2002) and Henson et al. (2012) both suggest that TE is low at high latitudes and high at low latitudes. They explain this pattern in terms of surface biological properties, suggesting that highly productive communities, typically found in cool regions, have low surface recycling rates, but also low TE due to the sinking of highly labile aggregates which are remineralized efficiently in the mesopelagic. In contrast, communities residing in warm oligotrophic waters, have high values of TE due to high levels of surface recycling and the production of refractory aggregates. This picture is further complicated by the ballast hypothesis (Armstrong et al., 2002), which suggests that minerals drive deep flux by increasing aggregate density and by potentially protecting POC from degradation, since both high and low latitude regions produce biominerals, predominantly opal, and calcite, respectively (Francois et al., 2002). Recent studies, however, have only found evidence of ballast acting to increase aggregate density, rather than performing any protective function (Iversen and Robert, 2015).
In contrast, other work has focussed on water column temperature and dissolved oxygen (DO) concentrations as key controls over interior processes since both play first-order roles in respiration. A synthesis of all available estimates of the flux attenuation length scale (the depth at which ~1/3 of the export flux remains) obtained from drifting neutrally buoyant sediment traps demonstrated that cold environments have longer length scales (Marsay et al., 2015; Figure 1A), consistent with the known role of temperature in regulating respiration and viscosity (Yvon-Durocher et al., 2012; Taucher et al., 2014). This is, however, counter to the patterns found by Francois et al. (2002) and Henson et al. (2012).
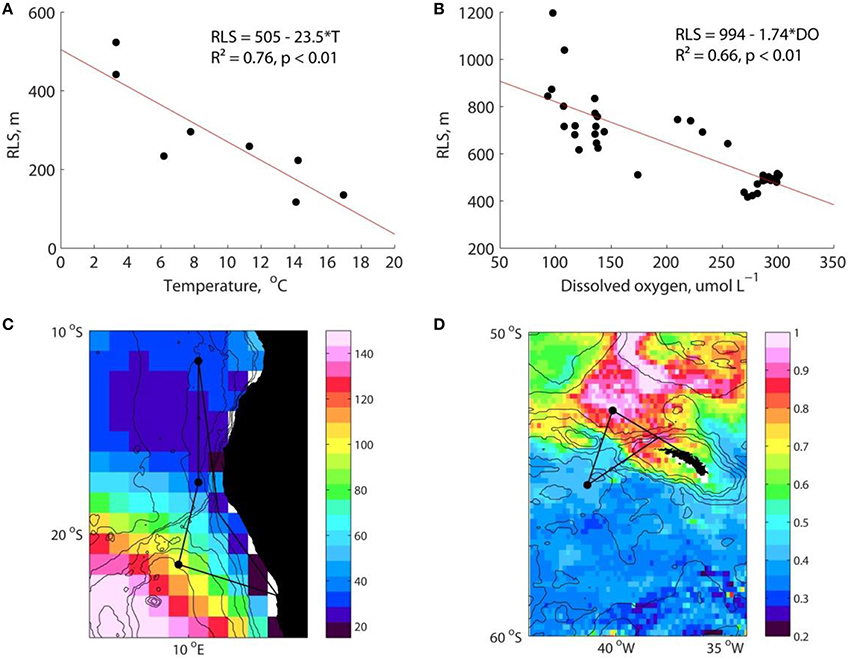
Figure 1. (A) Remineralization length scale (RLS) estimated from neutrally buoyant sediment traps deployed between 60 and 600 m depth plotted against 0−500 m mean temperature. Redrawn from a data compilation presented in Marsay et al. (2015). (B) RLS estimated from thorium-derived export flux at 100 m depth and moored sediment traps at 2000 m depth (from data presented in Henson et al., 2012) plotted against 0−500 m mean dissolved oxygen concentration. For both (A,B) environmental data are from the World Ocean Atlas 2013 (Locarnini et al., 2013; Garcia et al., 2014). Results from Ordinary Least Squares regression are shown. (C) Climatological minimum dissolved oxygen concentration (μmol L−1) in July in Benguela upwelling region (Garcia et al., 2014). (D) Climatological satellite-derived primary production (gC m−2) in December in the South Georgia region (estimated from MODIS data using the algorithm of Behrenfeld and Falkowski, 1997). In panels (C,D) black dots and lines show potential cruise track and major process stations.
Devol and Hartnett (2001) originally proposed that DO may be an important control on remineralization. A synthesis of attenuation length scale estimated from export flux and deep sediment traps (Henson et al., 2012) shows clearly that mesopelagic remineralization is much reduced at low DO concentrations (Figure 1B). Low DO concentration can cause oxic respiration in sinking aggregates to become diffusion rather than substrate limited, with a switch to nitrate respiration also possible (DO < 25 μM; Ploug, 2001; Ploug and Bergkvist, 2015). Additionally, modifications to DVM may occur as DO constrains DVM patterns (Bianchi et al., 2013). Zooplankton may avoid low DO waters, reducing the depth to which the active flux penetrates, and/or the depth of particle fragmentation.
A final possibility is that the feeding ecology and metabolic pathways of organisms in the interior play important roles. G14 infer that zooplankton permanently resident in the mesopelagic (i.e., ones which do not undergo DVM) fragment, rather than ingest, the majority of the particulate organic matter they encounter, converting much of it to smaller particles which are then respired by prokaryotes with a minor fraction remaining as, or being repackaged into, large particles such as fecal pellets. This fragmentation/repackaging process likely reduces mean sinking velocity of POC and may be associated with enhanced colonization and degradation rates (Buesseler and Boyd, 2009). Mayor et al. (2014) further speculate that zooplankton deliberately fragment detritus to stimulate the production of nutritious and harvestable microbial biomass from poor quality organic material, termed the “microbial gardening hypothesis.”
The lack of consensus about the relative importance of the processes involved in driving mesopelagic respiration and particle flux attenuation has the practical consequence of hindering the construction of an accurate representation of remineralization for use in global biogeochemical models. As a consequence, there is no consensus regarding how mesopelagic remineralization should be parameterized. No model can contain all possible processes and the argument for including particular processes is often made in isolation (tempered by computational efficiency) rather than from a perspective of how the processes interact. None have a parameterization grounded in an ecological and biogeochemical analysis of how the mesopelagic zone functions.
In light of the requirement for realistic model representations of mesopelagic processes to better quantify and constrain ocean carbon storage for the development of the UK Earth System Model (ESM), and other ESMs contributing to IPCC assessments, the UK NERC have funded a large integrated fieldwork and modeling programme aimed at understanding and parameterizing mesopelagic remineralization processes: Controls over Ocean Mesopelagic Interior Carbon Storage (COMICS). The importance of the biological carbon pump in global climate regulation and our current inability to model it reliably has implications for predictions of the carbon cycle; of the contemporary period, the past, and predictions for future climate change. As such we believe that a high-level overview of the work planned in COMICS will be of interest to a broad cross-section of the marine science community, both nationally and internationally.
Fieldwork
The experimental approach of COMICS is to make observations of surface and mesopelagic community structure and functioning in contrasting ocean locations where strong local gradients in key parameters occur. A study site at South Georgia (SG) targets a region of large gradients in ecosystem structure, whilst a study site in the Benguela upwelling system has strong gradients in DO (Figures 1C,D).
South of SG, productivity is relatively low with the community being dominated by small phytoplankton taxa (Korb et al., 2012), and a high degree of surface recycling (Owens et al., 1991). In contrast, north of SG the Subantarctic Circumpolar Current Front interacts with the shelf and slope introducing iron to the water column (Holeton et al., 2005; Nielsdottir et al., 2014) and causing an intense phytoplankton bloom characterized by large diatoms (Korb et al., 2012) and decreased surface recycling. Carbon export is higher in the northern region (Rembauvillle et al., 2016), with silica:carbon ratios in exported particles also different at the two sites (Le Moigne et al., 2014), potentially due to the well documented control iron plays in regulating silica:nitrogen ratios in diatoms (Hutchins and Bruland, 1998; Takeda, 1998). This frames the hypothesis that greater surface recycling and higher silica:carbon ratios will result in a deeper penetration of sinking particles into the interior at the southern site. Both sites are well oxygenated, and we therefore expect a strong active flux associated with DVM (Bianchi et al., 2013).
The second field study will focus on the role of DO by targeting the Benguela region where coastal upwelling supports intense production of organic matter, the sinking and respiration of which leads to the formation of a pronounced low oxygen zone (Shannon and Nelson, 1996; Mohrholz et al., 2008). Occupation of stations with differing DO will allow the hypothesis to be tested that the reduced oxygen layer, which is evident just below the thermocline, causes a deeper penetration of organic carbon due either to changes in DVM, or to a reduction in aggregate respiration rates driven by low external DO and nitrate concentrations (Kalvelage et al., 2011; Füessel et al., 2012).
Natural gradients within the study regions will not allow the role of temperature to be directly tested, although comparisons between the systems will be feasible, with the caveat that DVM patterns are likely to differ in the two systems. Two research cruises are planned for 2017/18 when gradients in primary production (for South Georgia), and DO (for Benguela) are maximal. On each cruise a full suite of measurements will be collected to characterize the carbon supply to the mesopelagic and the processes that control its attenuation (Table 1).
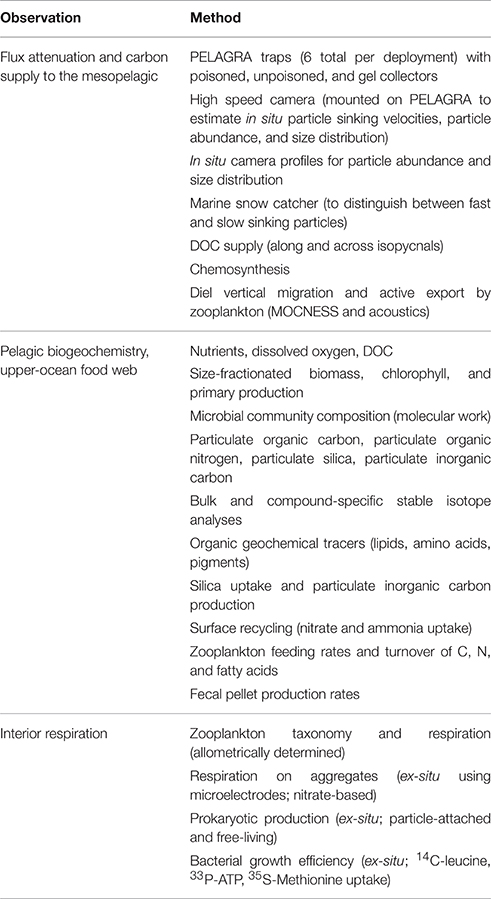
Table 1. Observations planned for the COMICS fieldwork phase.
Synthesis and Modeling
A hierarchical approach is required to achieve integration of the fieldwork into improved model forecasts. An initial data synthesis phase will bring together the estimates of organic matter supply and demand measured during the fieldwork and determine how these can be reconciled to produce a balanced carbon budget. This will highlight the processes that have the greatest influence on mesopelagic remineralization and which should therefore be the focus of initial 1-D modeling efforts. Creation of a detailed process model (an extension of G14 to include improved depiction of zooplankton and microbial dynamics) will permit development of a range of models with sufficient realism to represent mesopelagic functioning accurately, yet which are sufficiently simple that they can be run in a global climate model. The challenge is to produce an interior food web model with the minimum complexity required to reproduce the budgets of interior flux and respiration.
Proper evaluation of the effect this foodweb model has on the global carbon cycle requires it to be run to equilibrium in a 3D framework to examine its predicted patterns of nutrient, carbon and oxygen distributions within the ocean interior. However, reaching equilibrium requires a spin-up of many thousands of years and is too computationally expensive to achieve using a full ESM. To sidestep this, the suitability of newly formulated alternative parameterizations for future climate projections will be evaluated by determining their ability to reproduce the global tracer field using the computationally efficient Transport Matrix Modeling approach (Khatiwala, 2007; Bernardello et al., in prep). This effectively represents ocean circulation and mixing via a computationally inexpensive matrix operation that allows significantly accelerated model spin-up. A mechanistic, particle-based modeling approach will also be pursued using the Transport Matrix Method, using the explicit simulation of a large ensemble of marine particles and coagulation and disaggregation processes (Jokulsdottir and Archer, 2016). Following this, the best parameterization(s) will be implemented in NEMO-MEDUSA (Yool et al., 2013), the coupled biophysical model selected for use within UKESM1, the UK contribution to the next IPCC assessment.
While it is clearly important to determine whether the fieldwork and initial modeling efforts result in different estimates of oceanic carbon storage, it is also important to understand how—and why—this may change, both in the past and into the future, and what such changes imply, both for our understanding of mesopelagic carbon cycling and, more broadly, the global carbon cycle and climate feedbacks (Kwon et al., 2009; Roth et al., 2014). To address this, forward simulations using the new parameterization will allow us to study how the functioning of the mesopelagic system may evolve as external forcing changes, and the consequences of this for carbon storage.
Perspective
International policy and strategy on climate change mitigation and adaptation rely on the most robust possible projections of future changes to CO2 uptake and storage. A key part of these projections lie in understanding the sustainability of, and reducing the impact on, existing environmental services, e.g., atmospheric CO2 modulation by the ocean. By improving predictions of future ocean carbon storage, COMICS will contribute essential knowledge for evidence-based policy-making in both of these areas.
COMICS has been carefully designed so that new insights into the mesopelagic carbon cycle produced by leading-edge fieldwork will be directly translated into testable novel model parameterizations, which will then be incorporated into the UK's community ESM. The insights we gain will potentially have far-reaching influence, firstly via our innovative fieldwork which will provide an unprecedented level of detail on mesopelagic carbon fluxes, and secondly via our development of new parameterizations for carbon remineralization. These can be incorporated into any suitable global biogeochemical climate model, and have the potential to influence future climate model development at the international level. The need to better understand and model mesopelagic processes is underscored by increasing international efforts, for example in the concurrent and complementary NASA programme EXPORTS (Siegel et al., 2016), which will focus on field studies and modeling to support and develop future satellite-derived estimates of upper ocean export fluxes and mesopelagic remineralization.
Mechanistic understanding of the processes that contribute to mesopelagic remineralization remains a major challenge for the oceanographic community, both for describing the present-day carbon cycle and for projecting how this will interact with global climate change into the future. The targeted fieldwork campaign and innovative modeling approach of COMICS will enable significant progress toward filling this knowledge gap.
Author Contributions
All authors are members of the COMICS project team and contributed to the programme's development.
Funding
COMICS is funded by a Large Grant from the UK Natural Environment Research Council (NE/M020835/1) to RS.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Alonso-Gonzalez, I. J., Aristegui, J., Lee, C., Sanchez-Vidal, A., Calafat, A., Fabres, J., et al. (2010). Role of slowly settling particles in the ocean carbon cycle. Geophys. Res. Lett. 37:L13608. doi: 10.1029/2010GL043827
Alonso-Saez, L., Galand, P. E., Casamayor, E. O., Pedros-Alio, C., and Bertilsson, S. (2010). High bicarbonate assimilation in the dark by Arctic bacteria. ISME J. 4, 1581–1590. doi: 10.1038/ismej.2010.69
Armstrong, R., Lee, C., Hedges, J., Honjo, S., and Wakeham, S. (2002). A new, mechanistic model for organic carbon fluxes in the ocean based on the quantitative association of POC with ballast minerals. Deep Sea Res. 49, 219–236. doi: 10.1016/S0967-0645(01)00101-1
Behrenfeld, M. J., and Falkowski, P. G. (1997). Photosynthetic rates derived from satellite-based chlorophyll concentration. Limnol. Oceanogr. 42, 1–20. doi: 10.4319/lo.1997.42.1.0001
Bianchi, D., Galbraith, E. D., Carozza, D. A., Mislan, K. A. S., and Stock, C. A. (2013). Intensification of open-ocean oxygen depletion by vertically migrating animals. Nat. Geosci. 6, 545–548. doi: 10.1038/ngeo1837
Bishop, J. K. B., Edmond, J. M., Ketten, D. R., Bacon, M. P., and Silker, W. B. (1977). Chemistry, biology, and vertical flux of particulate matter from upper 400 m of Equatorial Atlantic Ocean. Deep Sea Res. 24, 511–520. doi: 10.1016/0146-6291(77)90526-4
Buesseler, K. O., and Boyd, P. W. (2009). Shedding light on processes that control particle export and flux attenuation in the twilight zone of the open ocean. Limnol. Oceanogr. 54, 1210–1232. doi: 10.4319/lo.2009.54.4.1210
Burd, A. B., Hansell, D. A., Steinberg, D. K., Anderson, T. R., Arístegui, J., Baltar, F., et al. (2010). Assessing the apparent imbalance between geochemical and biochemical indicators of meso- and bathypelagic biological activity: What the@$#! is wrong with present calculations of carbon budgets? Deep Sea Res. 57, 1557–1571. doi: 10.1016/j.dsr2.2010.02.022
Carlson, C., Hansell, D. A., Nelson, N., Siegel, D., Smethie, W., Khatiwala, S., et al. (2010). Dissolved organic carbon export and subsequent remineralisation in the mesopelagic and bathypelagic realms of the North Atlantic basin. Deep Sea Res. 57, 1433–1445. doi: 10.1016/j.dsr2.2010.02.013
Devol, A. H., and Hartnett, H. E. (2001). Role of the oxygen-deficient zone in transfer of organic carbon to the deep ocean. Limnol. Oceanogr. 46, 1684–1690. doi: 10.4319/lo.2001.46.7.1684
Francois, R., Honjo, S., Krishfield, R., and Manganini, S. (2002). Factors controlling the flux of organic carbon to the bathypelagic zone of the ocean. Glob. Biogeochem. Cycles 16:GB1087. doi: 10.1029/2001GB001722
Füessel, J., Lam, P., Lavik, G., Jensen, M. M., Holtappels, M., Gunter, M., et al. (2012). Nitrite oxidation in the Namibian oxygen minimum zone. ISME J. 6, 1200–1209. doi: 10.1038/ismej.2011.178
Garcia, H. E., Locarnini, R. A., Boyer, T. P., Antonov, J. I., Baranova, O. K., Zweng, M. M., et al. (2014). World Ocean Atlas 2013, Vol. 3, Dissolved Oxygen, Apparent Oxygen Utilization, and Oxygen Saturation, ed S. Levitus; Technical ed A. Mishonov, NOAA Atlas NESDIS 75, p. 27.
Giering, S. L. C., Sanders, R., Lampitt, R. S., Anderson, T. R., Tamburini, C., Boutrif, M., et al. (2014). Reconciliation of the carbon budget in the ocean's twilight zone. Nature 507, 480–483. doi: 10.1038/nature13123
Henson, S. A., Sanders, R., and Madsen, E. (2012). Global patterns in efficiency of particulate organic carbon export and transfer to the deep ocean. Glob. Biogeochem. Cycles 26:GB1028. doi: 10.1029/2011GB004099
Henson, S. A., Yool, A., and Sanders, R. (2015). Variability in efficiency of particulate organic carbon export: a model study. Glob. Biogeochem. Cycles 29, 33–45. doi: 10.1002/2014GB004965
Herndl, G. J., and Reinthaler, T. (2013). Microbial control of the dark end of the biological pump. Nat. Geosci. 6, 718–724. doi: 10.1038/ngeo1921
Holeton, C. L., Nedelec, F., Sanders, R., Brown, L., Moore, C. M., Stevens, D. P., et al. (2005). Physiological state of phytoplankton communities in the Southwest Atlantic sector of the Southern Ocean, as measured by fast repetition rate fluorometry. Polar Biol. 29, 44–52. doi: 10.1007/s00300-005-0028-y
Hutchins, D. A., and Bruland, K. W. (1998). Iron-limited diatom growth and Si: N uptake ratios in a coastal upwelling regime. Nature 393, 561–564. doi: 10.1038/31203
Iversen, M., and Robert, M. (2015). Ballasting effects of smectite on aggregate formation and export from a natural plankton community. Mar. Chem. 175, 18–27. doi: 10.1016/j.marchem.2015.04.009
Jokulsdottir, T., and Archer, D. (2016). A stochastic, Lagrangian model of sinking biogenic aggregates in the ocean (SLAMS 1.0): model formulation, validation and sensitivity. Geosci. Model Dev. 9, 1455–1476. doi: 10.5194/gmd-9-1455-2016
Juniper, S. K., and Brinkhurst, R. O. (1986). Water-column dark CO2 fixation and bacterial-mat growth in intermittently anoxic Saanich Inlet. Br. Columb. Mar. Ecol. Prog. Ser. 33, 41–50. doi: 10.3354/meps033041
Kalvelage, T., Jensen, M. M., Contreras, S., Revsbech, N. P., Lam, P., Günter, M., et al. (2011). Oxygen sensitivity of anammox and coupled N-cycle processes in oxygen minimum zones. PLoS ONE 6:e29299. doi: 10.1371/journal.pone.0029299
Khatiwala, S. (2007). A computational framework for simulation of biogeochemical tracers in the ocean. Glob. Biogeochem. Cycles 21:GB3001. doi: 10.1029/2007GB002923
Korb, R. E., Whitehouse, M. J., Ward, P., Gordon, M., Venables, H. J., Poulton, A., et al. (2012). Regional and seasonal differences in microplankton biomass, productivity, and structure across the Scotia Sea: implications for the export of biogenic carbon. Deep Sea Res. 59, 67–77. doi: 10.1016/j.dsr2.2011.06.006
Kwon, E. Y., Primeau, F., and Sarmiento, J. L. (2009). The impact of remineralization depth on the air-sea carbon balance. Nat. Geosci. 2, 630–635. doi: 10.1038/ngeo612
Le Moigne, F. A. C., Pabortsava, K., Marcinko, C. L. J., Martin, P., and Sanders, R. J. (2014). Where is mineral ballast important for surface export of particulate organic carbon in the ocean?. Geophys. Res. Lett. 41, 8460–8468. doi: 10.1002/2014GL061678
Locarnini, R. A., Mishonov, A. V., Antonov, J. I., Boyer, T. P., Garcia, H. E., Baranova, O. K., et al. (2013). World Ocean Atlas 2013, Vol. 1, Temperature, ed S. Levitus; Technical ed A. Mishonov, NOAA Atlas NESDIS 73, p. 40.
Manno, C., Stowasser, G., Enderlein, P., Fielding, S., and Tarling, G. (2015). The contribution of zooplankton faecal pellets to deep carbon transport in the Scotia Sea (Southern Ocean). Biogeosciences 12, 1955–1965. doi: 10.5194/bg-12-1955-2015
Marsay, C. M., Sanders, R. J., Henson, S. A., Pabortsava, K., Achterberg, E. P., Lampitt, R., et al. (2015). Attenuation of sinking particulate organic carbon flux through the mesopelagic ocean. Proc. Natl. Acad. Sci. U.S.A. 112, 1089–1094. doi: 10.1073/pnas.1415311112
Martin, P., Lampitt, R. S., Perry, M. J., Sanders, R., Lee, C., D'Asaro, E., et al. (2011). Export and mesopelagic particle flux during a North Atlantic spring diatom bloom. Deep Sea Res. 58, 338–349. doi: 10.1016/j.dsr.2011.01.006
Mayor, D. J., Sanders, R., Giering, S. L. C., and Anderson, T. R. (2014). Microbial gardening in the ocean's twilight zone: detritivorous metazoans benefit from fragmenting, rather than ingesting, sinking detritus. Bioessays 36, 1132–1137. doi: 10.1002/bies.201400100
Mohrholz, V., Bartholomae, C. H., van der Plas, A. K., and Lass, H. U. (2008). The seasonal variability of the northern Benguela undercurrent and its relation to the oxygen budget on the shelf. Continen. Shelf Res. 28, 424–441. doi: 10.1016/j.csr.2007.10.001
Nielsdottir, M., Bibby, T., Moore, C. M., Hinz, D., Sanders, R., Whitehouse, M. J., et al. (2014). Seasonal and spatial dynamics of iron availability in the Scotia Sea. Mar. Chem. 130, 62–72. doi: 10.1016/j.marchem.2011.12.004
Owens, N. J. P., Priddle, J., and Whitehouse, M. J. (1991). Variations in phytoplanktonic nitrogen assimilation around South Georgia and in the Bransfield Strait (Southern Ocean). Mar. Chem. 35, 287–304. doi: 10.1016/S0304-4203(09)90023-8
Pan, X., Achterberg, E. P., Sanders, R., Poulton, A. J., Oliver, K. I. C., and Robinson, C. (2014). Dissolved organic carbon and apparent oxygen utilization in the Atlantic Ocean. Deep Sea Res. 85, 80–87. doi: 10.1016/j.dsr.2013.12.003
Parekh, P., Dutkiewicz, S., Follows, M. J., and Ito, T. (2006). Atmospheric carbon dioxide in a less dusty world. Geophys. Res. Lett. 33:L03610. doi: 10.1029/2005GL025098
Ploug, H. (2001). Small-scale oxygen fluxes and remineralization in sinking aggregates. Limnol. Oceanogr. 46, 1624–1631. doi: 10.4319/lo.2001.46.7.1624
Ploug, H., and Bergkvist, J. (2015). Oxygen diffusion limitation and ammonium production within sinking diatom aggregates under hypoxic and anoxic conditions. Mar. Chem. 176, 142–149. doi: 10.1016/j.marchem.2015.08.012
Reinthaler, T., van Aken, H. M., and Herndl, G. J. (2010). Major contribution of autotrophy to microbial carbon cycling in the deep North Atlantic's interior. Deep-Sea Research Part Ii-Topical Stud. Oceanogr. 57, 1572–1580. doi: 10.1016/j.dsr2.2010.02.023
Rembauvillle, M., Manno, C., Tarling, G., Blain, S., and Salter, I. (2016). Strong contribution of diatom resting spores to deep-sea carbon transfer in naturally iron-fertilised waters downstream of South Georgia. Deep Sea Res. I 115, 22–35. doi: 10.1016/j.dsr.2016.05.002
Riley, J. S., Sanders, R., Marsay, C., Le Moigne, F. A. C., Achterberg, E. P., Poulton, A., et al. (2012). The relative contribution of fast and slow sinking particles to ocean carbon export. Glob. Biogeochem. Cycles 26:GB1026. doi: 10.1029/2011GB004085
Roth, R., Ritz, S., and Joos, F. (2014). Burial-nutrient feedbacks amplify the sensitivity of atmospheric carbon dioxide to changes in organic matter remineralisation. Earth Syst. Dyn. 5, 321–343. doi: 10.5194/esd-5-321-2014
Shannon, L. V., and Nelson, G. (1996). “The Benguela: large scale features and processes and system variability,” in The South Atlantic Past and Present Circulation, eds G. W. H. Wefer, G. BergerSiedler, and D. J. Webb (Berlin; Heidelberg: Springer), 163–210.
Siegel, D., Buesseler, K. O., Behrenfeld, M. J., Benitez-Nelson, C. R., Boss, E., Brzezinski, M. A., et al. (2016). Prediction of the export and fate of global ocean net primary production: the EXPORTS science plan. Front. Mar. Sci. 3:22. doi: 10.3389/fmars.2016.00022
Steinberg, D. K., Van Mooy, B. A. S., Buesseler, K. O., Boyd, P. W., Kobari, T., Karl, D., et al. (2008). Bacterial vs. zooplankton control of sinking particle flux in the ocean's twilight zone. Limnol. Oceanogr. 53, 1327–1338. doi: 10.4319/lo.2008.53.4.1327
Takeda, S. (1998). Influence of iron availability on nutrient consumption ratio of diatoms in oceanic waters. Nature 393, 774–777. doi: 10.1038/31674
Taucher, J., Bach, L. T., Riebesell, U., and Oschlies, A. (2014). The viscosity effect on marine particle flux: a climate relevant feedback mechanism. Glob. Biogeochem. Cycles 28, 415–422. doi: 10.1002/2013GB004728
Villa-Alfageme, M., de Soto, F., Le Moigne, F. A. C., Giering, S. L. C., and Sanders, R. (2014). Observations and modeling of slow-sinking particles in the twilight zone. Glob. Biogeochem. Cycles 28, 1327–1342. doi: 10.1002/2014GB004981
Yool, A., Popova, E., and Anderson, T. R. (2013). MEDUSA-2.0: an intermediate complexity biogeochemical model of the marine carbon cycle for climate change and ocean acidification studies. Geosci. Model Dev. 6, 1767–1811. doi: 10.5194/gmd-6-1767-2013
Keywords: biological carbon pump, field campaign, science plan, ocean carbon cycle, biogeochemical model
Citation: Sanders RJ, Henson SA, Martin AP, Anderson TR, Bernardello R, Enderlein P, Fielding S, Giering SLC, Hartmann M, Iversen M, Khatiwala S, Lam P, Lampitt R, Mayor DJ, Moore MC, Murphy E, Painter SC, Poulton AJ, Saw K, Stowasser G, Tarling GA, Torres-Valdes S, Trimmer M, Wolff GA, Yool A and Zubkov M (2016) Controls over Ocean Mesopelagic Interior Carbon Storage (COMICS): Fieldwork, Synthesis, and Modeling Efforts. Front. Mar. Sci. 3:136. doi: 10.3389/fmars.2016.00136
Received: 06 June 2016; Accepted: 25 July 2016;
Published: 05 August 2016.
Edited by:
Javier Arístegui, University of Las Palmas de Gran Canaria, SpainReviewed by:
Il-Nam Kim, Incheon National University, South KoreaSergio Ruiz-Halpern, Southern Cross University, Australia
Copyright © 2016 Sanders, Henson, Martin, Anderson, Bernardello, Enderlein, Fielding, Giering, Hartmann, Iversen, Khatiwala, Lam, Lampitt, Mayor, Moore, Murphy, Painter, Poulton, Saw, Stowasser, Tarling, Torres-Valdes, Trimmer, Wolff, Yool and Zubkov. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Stephanie A. Henson, cy5oZW5zb25Abm9jLmFjLnVr