 Shuiwang Duan
Shuiwang Duan Yuxiang He
Yuxiang He Sujay S. Kaushal
Sujay S. Kaushal Thomas S. Bianchi
Thomas S. Bianchi Nicholas D. Ward
Nicholas D. Ward Laodong Guo
Laodong Guo- 1Department of Geology and Earth System Science Interdisciplinary Center, University of Maryland, College Park, MD, USA
- 2National Water Center, National Weather Service, National Oceanic and Atmospheric Administration, Silver Spring, MD, USA
- 3Department of Geological Sciences, University of Florida, Gainesville, FL, USA
- 4Whitney Laboratory for Marine Bioscience, University of Florida, St. Augustine, FL, USA
- 5School of Freshwater Sciences, University of Wisconsin-Milwaukee, Milwaukee, WI, USA
Prior to discharging to the ocean, large rivers constantly receive inputs of dissolved organic carbon (DOC) from tributaries or fringing floodplains and lose DOC via continuous in situ processing along distances that span thousands of kilometers. Current concepts predicting longitudinal changes in DOC mainly focus on in situ processing or exchange with fringing floodplain wetlands, while effects of heterogeneous watershed characteristics are generally ignored. We analyzed results from a 17-year time-series of DOC measurements made at seven sites and three expeditions along the entire Mississippi River main channel with DOC measurements made every 17 km. The results show a clear downstream decrease in DOC concentrations that was consistent throughout the entire study period. Downstream DOC decreases were primarily (~63–71%) a result of constant dilutions by low-DOC tributary water controlled by watershed wetland distribution, while in situ processing played a secondary role. We estimate that from 1780 to 1980 wetland loss due to land-use alterations caused a ca. 58% decrease in in DOC concentrations in the tributaries of the Mississippi River. DOC reductions caused by watershed wetland loss likely impacted the capacity for the river to effectively remove nitrogen via denitrification, which can further exacerbate coastal hypoxia. These findings highlight the importance of watershed wetlands in regulating DOC longitudinally along the headland to ocean continuum of major rivers.
Introduction
Draining an area of ~28% of the Earth's surface, the 25 largest rivers in terms of discharge are responsible for over 40% of the global flux of DOC from rivers to the oceans (McKee, 2003). Within major river systems, DOC also plays an important role in driving microbially-mediated denitrification (Philip and Townsend, 2010) and greenhouse gas emissions (Raymond et al., 2013), and affecting the transport and toxicity of metals and trace organic contaminants (Aiken et al., 2011). A limited understanding of the fundamental processes controlling spatiotemporal variability of river DOC at the basin-scale restricts our ability to accurately predict the influence of future scenarios (e.g., land use-land cover and climate change) on fluvial DOC levels. Current concepts predicting mechanisms for DOC variations from the headwaters to the mouth of major rivers primarily focus on in situ processing (Vannote et al., 1980) and hydrologic exchange with fringing floodplain wetlands (Sedell et al., 1989). In situ processing causes DOC loss as a result of microbial degradation coupled with photochemical oxidation (Amon and Benner, 1997; Ward et al., 2013) and sorption onto minerals (Aufdenkampe et al., 2001), whereas autochthonous inputs to riverine DOC are generally more labile and thus largely recycled (Stanley et al., 2012). Fringing floodplains can considerably augment river DOC levels (Tockner et al., 1999; Hedges et al., 2000) owing to high levels of both algal and aquatic plant production. Relatively speaking, the effects of heterogeneous watershed characteristics have been generally ignored in river continuum approaches characterizing riverine DOC dynamics (Vannote et al., 1980; Sedell et al., 1989). This is particularly surprising when considering it is well known that DOC concentrations vary with climate zone (Schlesinger and Melack, 1981) and land use -land cover (LULC) (Butman et al., 2015). Wetland coverage is often the best predictor of DOC concentrations in rivers and streams (Gergel et al., 1999; D'Amore et al., 2016). However, LULC (e.g., wetland distribution) is seldom used to interpret DOC changes in large river systems. Models and theoretical concepts (e.g., SPARROW Shih et al., 2010 and the urban watershed continuum Kaushal and Belt, 2012) have been developed to link river water quality to watershed characteristics, but relative contributions of in situ vs. watershed controls on riverine DOC are rarely quantified.
Controls on river DOC may change in watersheds that are highly impacted by complex human alterations, such as North America's largest river, the Mississippi River (MR; Figure 1). For example, Butman et al. (2015) determined that 8% of the DOC present in rivers today is radiocarbon-depleted (i.e., millennial aged) organic carbon that has been mobilized as a result of human disturbances. The MR has been greatly altered during the last few centuries by: (1) large-scale conversion of watershed wetlands to farmlands (Dahl and Johnson, 1991), (2) construction of high and low dams in primary tributaries, and (3) an extensive network of flood-control levees (Wiener et al., 1996). Dam constructions generally increase the in situ processing time of riverine DOC, while artificial levees and large-scale conversion of watershed land use may affect DOC inputs from wetlands in floodplain and watershed. Prior studies in the Mississippi River have reported a downstream trend of decreasing DOC concentrations during three seasons from 1991 to 1992 (Leenheer et al., 1995). Based on the sparse temporal variability in the above study, it is unclear whether this trend is a consistent phenomenon in the MR (Duan et al., 2007, 2014). Such observations of a downstream decrease is different from other large river systems, where DOC concentrations increase or there are no apparent changes along the downstream gradient (Lara et al., 1998; Hedges et al., 2000; Wu et al., 2007). Reasons for these differences among the rivers (e.g., differential watershed characteristics vs. fundamental differences in in situ processing mechanisms) remain unclear.
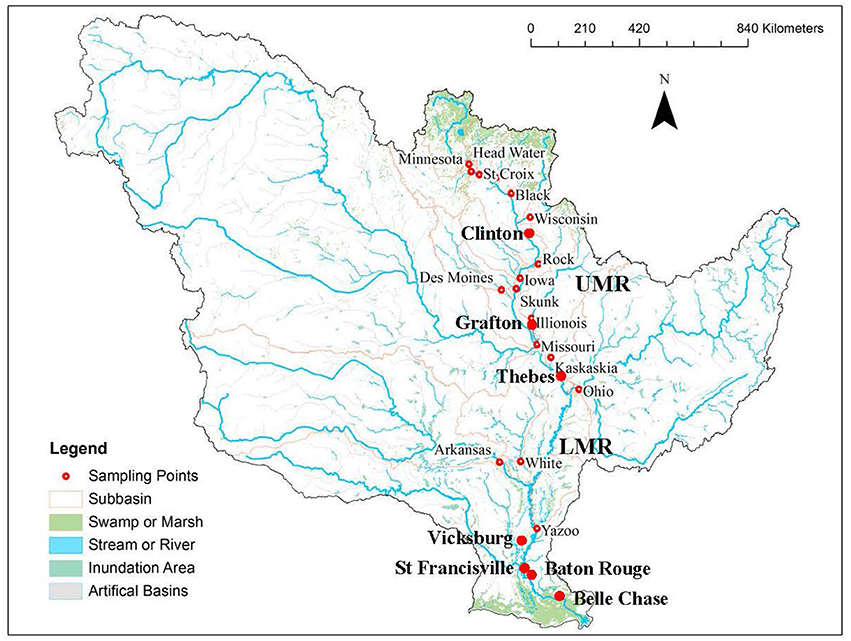
Figure 1. Map for U.S. Geological Survey monitoring stations on the Mississippi River main stem (large dots) and the primary tributaries (small dots), and distribution of wetland (in green) in the Mississippi River watershed. Thebes was the dividing point of the upper and lower Mississippi River (UMR and LMR).
In this study, DOC data from three intensive samplings along the Mississippi River were reanalyzed along with a 17 year time-series of DOC measurements starting in 1997 in an effort to unravel the processes regulating longitudinal changes in DOC concentrations in the MR. Further, we compare our results to longitudinal DOC changes in the world's major rivers to determine the importance of unique watershed characteristics. A combined mass balance and regression approach was used to estimate the relative contributions from in situ processing vs. tributary inputs, which are most likely controlled by watershed land use and climate. We hypothesize that watershed wetland distribution controls DOC levels of tributaries and DOC longitudinal changes along the Mississippi River. Decreases in DOC as a result of wetland loss may have large implications on the role that wetlands play in controlling DOC abundance, denitrification, and nitrogen fluxes from major rivers to coastal waters, such as the Gulf of Mexico (Schramm et al., 2009).
Materials and Methods
Water Quality Data
Past measurements of DOC and temperature made during three intensive samplings along the MR main channel from 1991 to 1992, which were used here for comparison and reanalysis, were collected from U.S. Geological Survey Open-File Report 94-4191 (Leenheer et al., 1995) and Open-File Report 94-523 (Moody, 1995), respectively. These expeditions were conducted in summer 1991 (June 23 to July 1), fall 1991 (September 25 to October 4), and spring 1992 (March 25 to April 4). Samples were taken near the center of the river approximately every 17 km from downriver to upriver between New Orleans and Minneapolis. DOC concentrations were also measured in the 17 primary tributaries up to six times (twice for the 3 seasons). Water samples were collected from just below water surface and DOC was analyzed using a high-temperature catalytic oxidation method (Leenheer et al., 1995).
Long-term water quality data were collected from the U.S. Geological Survey (USGS) National Stream Quality Accounting Network (NASQAN) webpage (http://water.usgs.gov/nasqan/). The data were obtained for all available sites on the Mississippi River main stem (Clinton, Grafton, Thebes, Vicksburg, St. Francisville, Baton Rouge, and Belle Chasse; Figure 1) and its primary tributaries (Iowa River at Wapello, Des Moines River at Keosauqua, Illinois River at Valley City, the Missouri River at Hermann, the Ohio River at Dam 53 near Grand Chain, the Arkansas River at David D Terry Lock and Dam below Little Rock, and the Yazoo River below Steele Bayou near Long Lake). Data at most sites covered the whole period of water years of 1997 to 2013 except the Des Moines River (2009–2013), and the LMR at Vicksburg (2008–2013), Baton Rouge (2008–2013), and Belle Chasse (2007–2013). Water sampling frequency varied but generally ranged from 6 up to 24 times per year; the most frequent collections occurred during spring high flow period. Methods for sample collections and measurements of water temperatures and DOC are available at Wilde (2006) and Brenton and Arnett (1993)—the same as those used in Leenheer et al. (1995).
Estimating Contributions of Tributaries Inputs vs. In situ Processing
Long-term USGS DOC concentration data (with water discharges) were used to separate contributions of tributary inputs from that of in situ processing to the longitudinal DOC decreases in the Upper Mississippi River (UMR, from Clinton to Thebes) and the Lower Mississippi River (LMR, from Thebes to St. Francisville) based on DOC mass balances. Mean daily water discharge data were assembled from the records of the U.S. Army Corps of Engineers (USACE; http://www.usace.army.mil/) or the USGS (http://waterdata.usgs.gov/usa/nwis/). In each river section (UMR or LMR), we used conservative mixing of tributaries with main stem water to estimate DOC concentrations at the downriver station; fractions explained and unexplained by conservative mixing were tributary inputs and in-situ transformation, respectively. That is, contributions from tributary inputs were the difference between in situ DOC measurements at the upstream station and DOC estimates for the downriver station. Meanwhile, contributions of in situ transformation were the difference between the DOC estimates and measurements at the downstream station. Positive values of the contributions from tributary inputs and in situ processing suggest dilutions by tributaries and in situ loss, while negative values imply increases in concentrations by tributary inputs and in situ production/floodplain inputs.
In order to estimate DOC concentrations of the UMR and LMR at the downstream stations, daily DOC fluxes of each input station (upstream main stem or tributaries) and output station were initially estimated from discrete DOC concentration measurements and continuous average daily flow using the USGS LOADEST method (http://water.usgs.gov/software/loadest/). LOADEST estimates constituent loads in streams and rivers by developing a regression model, given a time series of streamflow, constituent concentrations, and additional data inputs. The model is well documented in the published literature and is accepted as a valid means of calculating annual solute loads from a limited number of water quality measurements. We used measurements of DOC concentrations and streamflow during the same dates for model calibrations. The models, which contained up to seven parameters, captured the dependence of concentrations on discharge and season and any long-term trends. Daily discharge data for the study period were prepared for LOADEST models so that daily fluxes of DOC were estimated for the whole study period, once the model had been calibrated. From the returning files, results from the adjusted maximum likelihood estimates (AMLE) were used to modify loading equations to correct for transformation bias. Additionally, a period-weighted approach28 was used to verify above LOADEST method. In this approach, a measured DOC concentration was assumed to represent a period around which the sample was collected; this allowed for connecting measured concentrations through time by a piecewise linear function.
The estimated DOC daily loads were used for DOC mass balances for the UMR and LMR before a mean travel time from each input station to the output station was known. The travel time was obtained by finding the best correlations between daily discharge data at an input and an output station (Duan et al., 2010). More specifically, daily water discharges of the two stations were used for correlation analysis, with data of the input station moved forward a few days until the highest significance level of the correlation was achieved. The number of days that data needed to be adjusted was taken as the mean residence time between the two stations. This approach for travel time was verified by comparing water discharge inputs and output of each river section shown as in Duan et al. (2010). Once residence time from each input site was known, the daily water discharges and DOC loads of the input stations were re-assembled by moving them forward by the number of days that represented their mean travel times to the output station.
Mass balances of water discharges were initially conducted, and the unbalanced discharges were assumed to come from ungauged small tributaries. These discharges were then multiplied by mean daily DOC concentrations of ungauged tributaries (assumed to be averages of the selected tributaries) to estimate daily DOC inputs from these tributaries. Finally, total inputs of daily water discharges and DOC loads (gauged and ungauged) were then summarized, and daily DOC concentrations at the downriver stations were obtained by dividing total daily DOC inputs by total daily water inputs.
Watershed Land Use Land Cover and Climate Data
To identify controls of watershed characteristics on DOC concentrations of the primary tributaries, correlation analyses were conducted between mean DOC concentrations of the tributaries and their watershed LULC and climatic parameters (air temperature, precipitation and runoff ratio). Relationships were tested using Pearson's correlations (p < 0.05, Pearson's correlation). DOC data collected during the three expeditions of 1991–1992 were used, because more tributaries (17 in total) were sampled during these expeditions than the USGS sites with long-term DOC data (only seven) (Figure 1). For most tributaries, the data were representative because DOC was measured up to six times a year across seasons. National Land Cover Dataset 1992 (NLCD 1992; http://www.epa.gov/mrlc/nlcd.html) was used for land use data collection in order to match the time when DOC concentrations were measured. NLCD 1992 data were a 21-class land cover classification scheme applied consistently over the United States.
Mean air temperature and precipitation during a close period (1990–1995) were obtained from the NCEP North American Regional Reanalysis (NARR) dataset and the forcing data for the North Land Assimilation System (NLDAS). The near-surface mean air temperature was adjusted vertically to account for terrain height difference between NARR and NLDAS. Precipitation was a product of temporal disaggregation applied directly on the NLDAS grid based on gauge-only CPC daily precipitation analysis, and orographically-adjusted according to the widely used PRISM climatology.
Sub-watersheds of the 17 primary tributaries were divided from the Mississippi River basin. Boundary polygons of each watershed were generated based on USGS 8-digit hydrologic unit code (HUC). The LULC, annual mean air temperature and precipitation of each sub-watershed were extracted from data of the whole Mississippi watershed according to its boundary polygon using the ArcGIS spatial analyst tools. Annual runoff of each sub-watershed was calculated by dividing annual water discharge with watershed drainage area, and the annual runoff was then divided by annual precipitation to estimate runoff ratio (runoff / precipitation × 100%); data of water discharge and drainage area were from http://waterdata.usgs.gov/usa/nwis/.
Results
Longitudinal Changes in DOC Concentrations along the MR
Results showed that the downstream decrease in DOC concentrations along the main stem was consistent throughout the study period. High-resolution spatial data of 1991–1992 (Figures 2A–C) demonstrated that DOC concentrations decreased gradually, except for two rapid drops shortly after the confluence of major tributaries (Missouri and Ohio Rivers). DOC concentrations of major tributaries also displayed a longitudinal decreasing trend (Figures 2A–C; r2 ranging from 0.52 to 0.74, p < 0.05). To be consistent with data of 1991–1992, long-term USGS DOC data (1997–2013) were presented seasonally. These new records showed that DOC concentrations in the main stem consistently decreased downstream of MR headwaters across seasons throughout the study period (Figures 2D–G; r2 ranging from 0.87 to 0.96, depending on seasons, p < 0.05). The coefficients of variation were minor (C.V. = 0.4–10%) showing strong confidence.
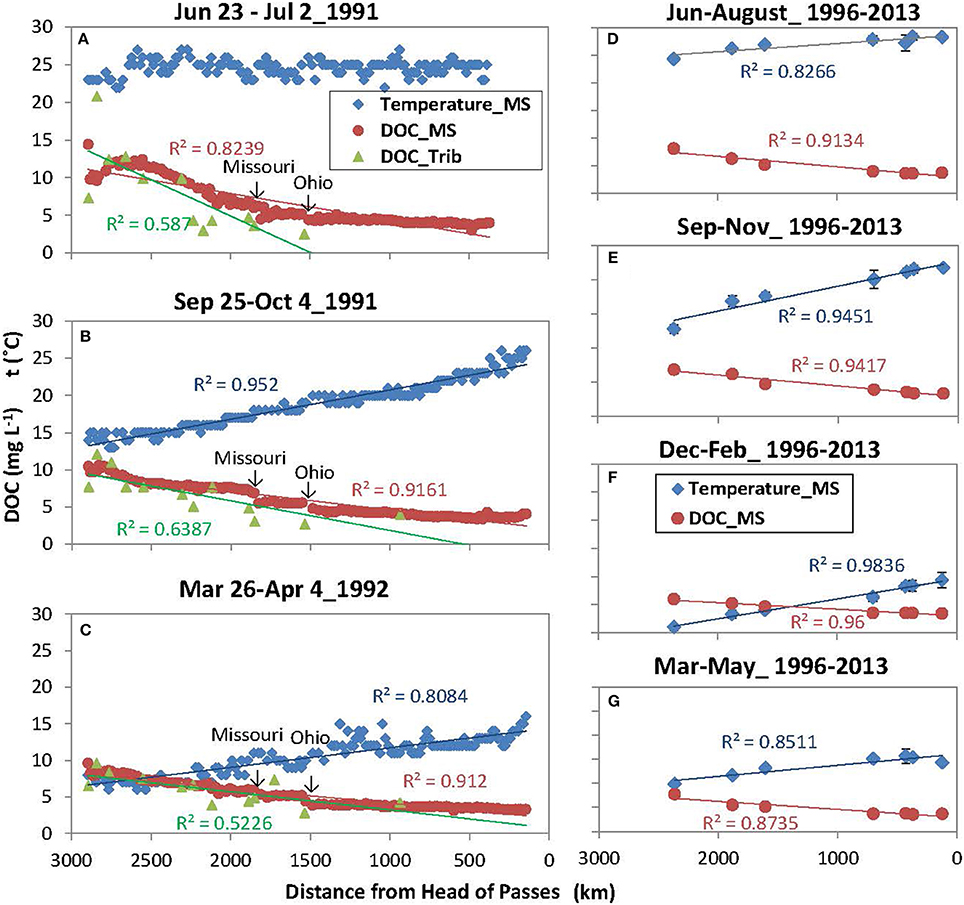
Figure 2. Downstream changes in dissolved organic carbon (DOC) concentrations and water temperatures in the Mississippi River (MS) and its tributaries (Trib) during three expeditions from 1991–1992 (A–C, left) and 1997–2013 (D–G, right). Regression lines are shown for statistically significant trends (p < 0.05). 1991–1992 measurements were obtained from the U.S. Geological Survey Open-File Report 94–4191 and Open-File Report 94–523.
Interestingly, the decreases in DOC concentrations were generally accompanied by downstream increases in water temperatures across seasons (one exception in Figure 2A) (Figures 2B–G; r2 ranging from 0.81 to 0.98, p < 0.05). As a result, DOC concentrations and water temperatures were negatively correlated (r2 ranging from 0.80 to 0.94, p < 0.05; one except of June-July 1991), but the slope changed with seasons suggesting complex temperature effects (Figure 3).
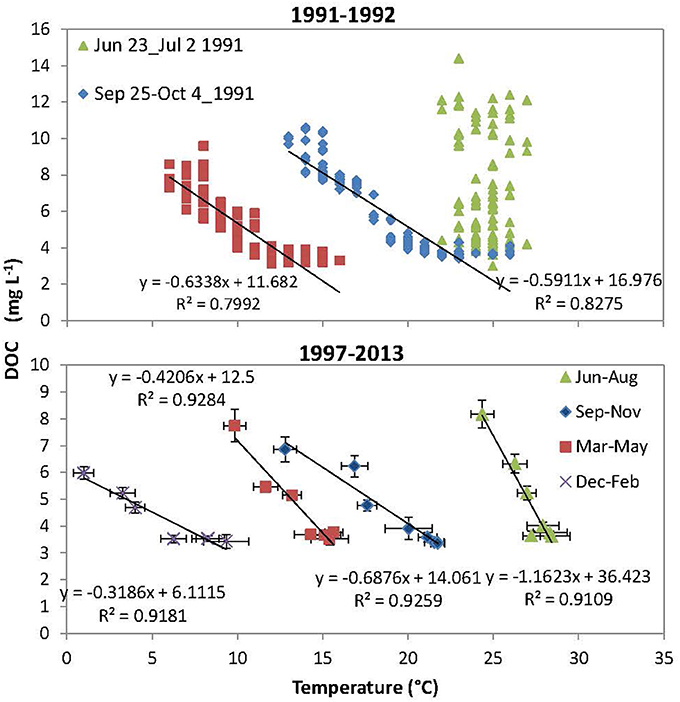
Figure 3. Correlations between DOC concentration and water temperature at the Mississippi River main stem during the three longitudinal sampling expeditions of 1991–1992 (upper) and during four seasons of 1997–2013 (bottom). Bars showed standard errors for every season.
Contributions of Tributary Inputs and In situ Processing to DOC Reductions
The contributions of tributary inputs and in situ processing to downstream DOC decreases were positive across all months (Figure 4), indicating that the downstream reductions of DOC concentrations were a combined effect of tributary dilution (by low-DOC water) and in situ loss. The contributions from tributary dilutions were positively correlated with water discharge in the LMR (Figure 4B; r2 = 0.81, p < 0.01), as well as in the UMR but the correlation was less significant (Figure 4A; two outliers of March and June; r2 = 0.63, p < 0.05). The contributions of in situ processing also exhibited pronounced seasonal variations. In the LMR section, this seasonal pattern strictly followed that of water temperatures (Figure 4D; r2 = 0.88, p < 0.05), but it was positively correlated with both water temperature and water discharge in the UMR (Figure 4C; r2 = 0.36 and 0.55, p < 0.05).
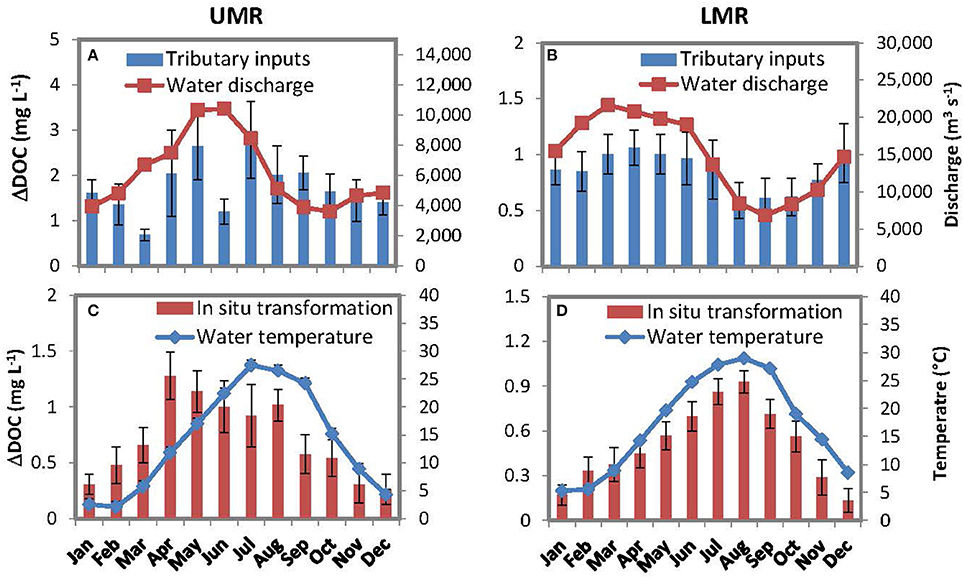
Figure 4. Monthly-averaged river discharge and contributions of tributary inputs to the downstream DOC decreases in the UMR and LMR from January 1997–September 2013 (A,B), and monthly-averaged water temperature and contributions of in situ transformation to the downstream DOC decreases (C,D). All values are presented in averages with standard errors.
Summing for the entire study period, we found that the effects of tributary dilution accounted for 71 and 63% of the DOC deceases in the UMR and LMR, respectively. The effects of tributary dilution were 2.4 times and 1.7 times that of in situ processing losses.
Control of Watershed Wetland Distribution on DOC Decreases
Patterns of DOC spatial variability across the Mississippi tributaries tightly followed that of watershed wetland coverage, with both parameters showing a downstream decrease from north-to- south (Figure 5A). In particular, DOC concentrations of the 17 primary tributaries were positively correlated with watershed wetland coverage (Figure 5D; r2 = 0.88, p < 0.05), whereas no significant correlation was found with any other type of land-use regime. Although tributary DOC concentrations also varied inversely with temperature, precipitation and runoff ratio (Figures 5B,C), the correlation coefficients were much lower (r2 = 0.57, 0.23, and 0.03, respectively). Moreover, we found watershed wetland coverage (wetland %) can be regressed negatively with mean air temperature (Temp) and runoff ratio (runoff%), but positively with annual precipitation (Pptn) with an r2 of 0.80 (Equation 1 and Figure 5E):
Where the standard errors of the estimates of the intercept and the three coefficients of the equation were 2.67, 0.32, 0.007, and 0.111, respectively.
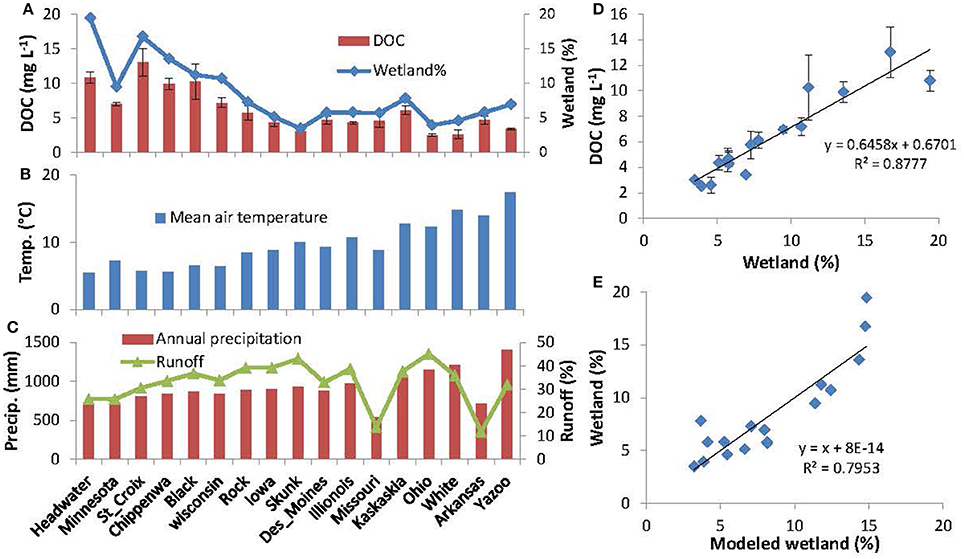
Figure 5. Downstream changes in DOC concentration in tributaries, watershed wetland coverage (wetland %), mean air temperature, annual precipitation, and runoff ratio (A–C); correlations between tributary DOC concentrations and watershed wetland area coverage (D); and comparison between watershed wetland % and modeled wetland % with Equation 1 (E). DOC concentrations of the tributaries were obtained from U.S. Geological Survey Open-File Report 94-4191. DOC concentrations were measured up to six times at different times for a year and presented as averages with standard errors.
Discussion
Consistent Longitudinal Pattern of DOC Concentrations along the Mississippi River
Our data suggest that the longitudinal decreases in DOC concentrations along the Mississippi River showed a consistent pattern. The downstream decreases were not only observed in the three high-resolution spatial data of 1991–1992 (Figures 2A–C), but also the long-term USGS DOC data (1997–2013) with intensive measurements (biweekly to monthly) (Figures 2D–G). Reviews on all available longitudinal DOC data of major rivers of the world (Table 1) found that the consistent downstream decreases in DOC concentrations along the MR were globally unique. Most rivers showed no obvious gradients or features (e.g., Lena; Lara et al., 1998, Columbia; Prahl et al., 1998, Paraná; Depetris and Kempe, 1993, and Yellow; Zhang et al., 2013), while other rivers displayed an increase or decrease only in a river section (e.g., Yangtze; Wu et al., 2007; Zhang et al., 2014 and Colorado; Miller, 2012). Consistent downstream increases occurred in the Amazon River (Hedges et al., 2000) and the St. Lawrence below the Great Lakes (Massicotte and Frenette, 2011). The unique longitudinal DOC pattern along the MR suggests that either fundamental in situ process are differentially expressed in the MR, or that watershed characteristics are the primary control on longitudinal DOC changes.
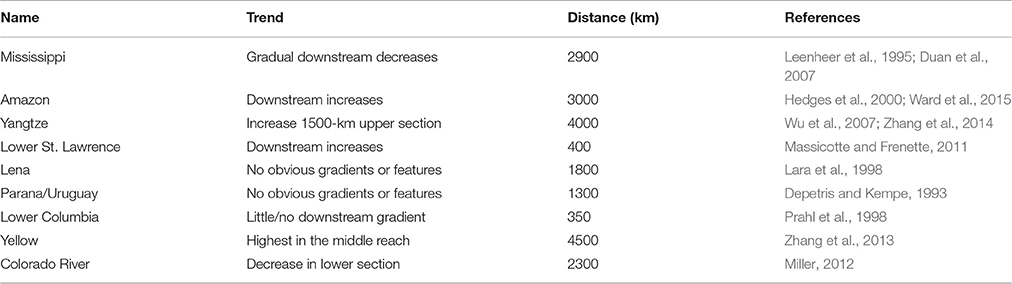
Table 1. Comparison for longitudinal changes in DOC concentrations along major rivers of the world.
The UMR is one of the three primary tributaries of the Mississippi River (the other are the Missouri, and the Ohio Rivers), and it represents neither the longest tributary nor the tributary carrying the most discharge. It seems that choosing either of the other tributaries as the origin of the LMR would change this story, since both are well known to have lower DOC concentrations than the upper MR. However, the UMR was chosen as the source, like a few other rivers (e.g., the Potomac River, USA), because it flows in the same direction as the main stem and thus crosses more climate zones and/or land-cover regimes. Therefore, the consistent downstream changes in DOC concentrations along the MR could be attributed to changes in climate zones and land-cover regimes reflecting DOC concentrations of tributaries.
Tributary Inputs as a Dominant Control on the DOC Longitudinal Changes
This study used a mass balance approach for isolating physical (e.g., dilution) and biogeochemical (e.g., bio/photodegradation) controls on DOC concentrations along the MR. To complete the mass balance it was necessary to assume that DOC concentrations of unsampled small tributaries (including inputs from floodplains) were equivalent to the average concentration observed in primary tributaries of the same region. This assumption was partially satisfied. First, despite large uncertainty, there was a regional pattern of DOC concentrations, i.e., higher in UMR watershed and lower in LMR watershed (Figure 5A). Moreover, although DOC inputs from floodplains can be substantial in natural lowland rivers (Mulholland and Kuenzler, 1979), the impact of floodplains should be much less because the Mississippi River (especially the LMR) was leveed and >90% of the floodplain was isolated from the river (Wiener et al., 1996). Our mass balance approach was also verified with seasonal data (Figure 4). For example, we found that contributions of in situ transformation were positively correlated with water temperature (Figures 4C,D). This relationship corresponds with the observed temperature dependence of both bacterial decomposition and photochemical oxidation (Davidson and Janssens, 2006; Porcal et al., 2015)—the two major pathways for in situ DOC loss in aquatic systems (Cole et al., 2007). The lower coefficient of correlation between DOC concentration and temperature in the UMR (4c) was probably due to additional DOC loss pathways, such as sorption onto particles (Aufdenkampe et al., 2001), DOC flocculation (Benedetti et al., 2003) and exchanges with floodplains (Tockner et al., 1999). Meanwhile, we also found that contributions from tributary dilutions on DOC decreases were all positively correlated with water discharge in both river sections (Figures 4A,B). This relationship matched well with the fact that DOC concentrations and water discharge are generally positively correlated in most tributaries of the Mississippi watershed (Irena et al., 2015).
Using the above mass balance approach, our results showed that tributary inputs are the primary control on riverine DOC longitudinal changes, while in situ processing play a secondary role. It was estimated that the effects of tributary dilution accounted for 71and 63% of the DOC deceases in the UMR and LMR, respectively, which is 2.4 times and 1.7 times that of in situ transformation losses, respectively. It is possible that tributaries dilute DOC concentrations in the MR because of the same longitudinal reduction phenomenon present within the tributaries themselves on a smaller scale, e.g., DOC concentrations decrease as the tributary flows toward the main channel (Figures 2A,C). On the other hand, although DOC concentrations and water temperature were negatively correlated downstream the river (Figure 3), this relationship did not support that the downriver decreases in DOC concentration was due to increasing in situ processing—that could be enhanced with higher temperatures at the downriver sites. This negative relationship did occur all the time (e.g., June-July 1991), and temperatures and DOC were not correlated if all-year data were included (Figure 3). In fact, when mean DOC in situ loss rates were estimated from data shown in Figure 4 (normalized to river length), we found that the in situ loss rates decreased from 0.93 μg L−1km−1 in the colder UMR to 0.43 μg L−1km−1 in the warmer LMR, in contrast to the temperature gradient. Thus, we hypothesize that temperature-dependent in situ processing were not the primary control on the downstream distribution of DOC in the Mississippi River.
Recent studies highlight the importance of in situ processing of DOC and CO2 emissions from many major river systems (Richey et al., 2002; Raymond et al., 2013). For example, it is reported that CO2 emissions from the Amazon River are an order of magnitude higher than fluvial DOC flux (Raymond et al., 2013). This finding, however, does not conflict with the results of this study. That is, although DOC in situ processing can be substantial, it may only modify the magnitude of DOC spatial variability but does not alter DOC longitudinal patterns along a river network, per se. Otherwise, one would expect that DOC concentrations would longitudinally decrease for most large rivers worldwide, which is not observed. Therefore, longitudinal DOC variability in major river systems are likely highly dependent on the structure of tributary networks and the LULC regime of the surrounding watershed.
Wetland Distribution Effects on River DOC Longitudinal Patterns
Data from this study showed that tributary DOC concentrations were tightly correlated with watershed wetland coverages, both of which decreased from headwater watersheds to the watersheds of the LMR (Figure 5A). Tributary DOC concentrations also varied inversely with temperature, precipitation and runoff ratio with less significance (Figures 5B,C), but it was more likely that these variables were controls for the formation of wetlands (Equation 1). That is, the occurrences of large areas of wetlands in the headwater sub-basins probably attributed to topographical depressions, cool climates, and abundant moisture in this region. Moreover, the negative relationship between wetland extent and air temperature also explains the complex relationship between DOC concentrations and water temperature, which varied seasonally (Figure 3). Therefore, wetland distribution (occurring mainly in headwaters) is likely the primary control on tributary DOC concentrations and downstream DOC decreases along the main stem of the MR. The tight coupling of tributary DOC concentrations with watershed wetland coverage observed here was also consistent with prior reviews showing occurrence of maximal DOC concentrations in wetlands (Schlesinger and Melack, 1981; Spitzy and Leenheer, 1991). This is also consistent with findings showing that wetland coverage was the best predictor of DOC concentration in rivers and streams (Mulholland and Kuenzler, 1979; Massicotte and Frenette, 2011).
With this relationship determined, wetland distribution can be used to interpret longitudinal patterns of DOC concentrations along major world rivers. For example, in more naturally flowing systems that have higher connectivity to floodplain wetlands, like the Amazon River, DOC increases downstream concurrent with high rates of in situ processing as a result of floodplain wetland inputs to the river (Hedges et al., 2000; Ward et al., 2015). Wetlands also play an important role in driving CO2 production in large rivers (Abril et al., 2014; Borges et al., 2015) due to aquatic plant respiration and enhanced in situ processing due to both breakdown of wetland-derived OC and processes, such as priming effects, which can stimulate the breakdown of terrestrially-derived OC in the presence of fresh autochthonous OC (Guenet et al., 2014; Bianchi et al., 2015; Ward et al., 2016). In contrast, in rivers where wetlands are primarily located in headwater sub-watersheds and floodplain wetland extent is limited (as a result of human development; Wiener et al., 1996), such as the MR (Figure 1), DOC concentrations decrease gradually from headwaters to the river mouth due to tributary dilution. Therefore, the effects of wetland distribution should be highlighted when we examine the controls on longitudinal changes in DOC concentrations along large rivers with respect to global and regional carbon budgets.
Historical Wetland Loss and Effect on Mississippi Watershed C and N
A significant fraction of wetland extent in the UMR has been lost during the last few centuries (1780–1980) due to large-scale conversion of wetlands to farmlands in the Ohio/MR Valleys and dam constructions in the UMR for navigation (Shih et al., 2010). In the LMR, a large fraction of floodplain wetlands have also been replaced with arable land, and human-made levees have resulted in a reduction of about 90% of the local inputs from the remaining floodplain wetlands in the region (Wiener et al., 1996). Based on EPA estimates of U.S. wetland extent in 1780 and 1980 for each state (Dahl, 1990), we calculated wetland losses for each tributary using ARC GIS. In total, wetland coverage of the MR watershed decreased from 8.8 to 3.1%, showing a wetland loss as high as 65% during this period. As previously discussed, such a drastic alteration to wetland extent likely played a key role in shaping the amount and composition of DOC that was exported from the watershed over the last several centuries, and helped form the downstream trends in DOC variability observed here in the present day. For example, based on the regression of tributary DOC concentrations with watershed wetland coverage (Figure 5D), we estimate that mean DOC concentrations of the MR tributaries decreased from 6.35 mg L−1 in 1780 to 2.66 mg L−1 in 1980 on average, having decreased by 58% or up to 3.69 mg L−1. Of course, there are many other factors in the basin that also affect carbon dynamics in the MR, including agricultural and urban development and channel modifications (e.g., dam constructions) throughout the UMR (Wiener et al., 1996). However, our data show agricultural or urban land cover was not correlated with river DOC concentrations. Dam construction may increase residence time for DOC in situ processing, but the effect of damming of riverine DOC is generally complicated (Wehr et al., 1997; Nadon et al., 2015) and thus less effective relative to watershed wetland loss.
Historical wetland loss and the associated DOC reductions may have further implications for the regional cycles of nutrients (nitrogen, phosphorus and silicon; Struyf and Conley, 2009). Reductions in wetlands would have local effects (e.g., nutrient sinks) as well as downstream effects from DOC export. For local effects, nitrogen, phosphorus and dissolved silicon can be retained via biological uptake or sedimentation, and high DOC exerts an important control on the occurrence of denitrification (the major process for nitrogen removal) by providing an energy source and anaerobic conditions (Philip and Townsend, 2010). Large amounts of DOC exported from wetland may cause anaerobic conditions in downstream rivers and nitrogen removal via denitrification. Thus, historical wetland loss in the UMR watershed and the LMR floodplain may have greatly reduced the capacity of this river system to retain nitrogen and other nutrients. Consequently, high nutrient yields from the croplands of MR watershed due to increased fertilizer application could be further enhanced by the wetland loss to some extent (Mitsch et al., 2005; Schramm et al., 2009). This finding has major implications for the implementation of restoration strategies to reduce nutrient fluxes from the MR to the Gulf of Mexico, which unfortunately continues to be ignored (Rabotyagov et al., 2014). In some cases these inundated wetland environments act as important sinks for river nitrogen, and their loss through land-use change can exacerbate hypoxia in coastal waters.
Future Perspectives
Consistent long-term monitoring of aquatic systems, along with mechanistic modeling is necessary for constraining how earth systems will react to changing conditions in the future (e.g., climate, land use land cover, and population density). While detailed mechanisms can be difficult to disentangle in large river systems, the basin-integrated signal present in the main channel of a large river allows the use of simple models, such as those presented here to elucidate large scale, broadly defined processes (e.g., in situ processing vs. tributary inputs) across large spatial scales (e.g., continental). Constraining the mechanisms that drive biogeochemical fluxes from the land to the sea on a conceptual and quantitative basis is essential to understanding how natural systems interact, the influence that human activities have on these processes, and how these processes have/will change in the future.
Author Contributions
SD, YH, and SK conceived of the modeling approach. SD compiled the DOC data from the literature and performed the model analyses. SD, TB, NW, and LG performed comparisons of the Mississippi River to other large rivers worldwide. All authors contributed to the writing and critical evaluation of the manuscript.
Funding
This research was supported by NASA grant NASA NNX11AM28G, NSF Awards EAR 1521224, DBI 0640300, CBET 1058502, and EAR-1426844. The Jon and Beverly Thompson Endowed Chair of Geological Science at the University of Florida.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Stuart Findlay provided helpful comments on an earlier version of the manuscript. We thank the USGS for the historic data evaluated here. Open access publication fees were supported in part by the Gordon and Betty Moore Foundation.
References
Abril, G., Martinez, J. M., Artigas, L. F., Moreira-Turcq, P., Benedetti, M. F., Vidal, L., et al. (2014). Amazon River carbon dioxide outgassing fuelled by wetlands. Nature 505, 395–398. doi: 10.1038/nature12797
Aiken, G. R., Gilmourb, C. C., Krabbenhoftc, D. P., and Oremd, W. (2011). Dissolved organic matter in the florida everglades: implications for ecosystem restoration. Crit. Rev. Env. Sci. Technol. 41, 217–248. doi: 10.1080/10643389.2010.530934
Amon, R., and Benner, R. (1997). Photochemical and microbial consumption of dissolved organic carbon and dissolved oxygen in the Amazon River system. Geochim. Cosmochim. Acta 60, 1783–1792. doi: 10.1016/0016-7037(96)00055-5
Aufdenkampe, A. K., Hedges, J. I., and Richey, J. E. (2001). Sorptive fractionation of dissolved organic nitrogen and amino acids onto fine sediments within the Amazon Basin. Limnol. Oceanogr. 46, 1921–1935. doi: 10.4319/lo.2001.46.8.1921
Benedetti, M. F., Mounier, S., Filizola, N., Benaim, J., and Seyler, P. (2003). Carbon and metal concentrations, size distributions and fluxes in major rivers of the Amazon Basin. Hydrol. Process. 17, 1363–1377. doi: 10.1002/hyp.1289
Bianchi, T. S., Thornton, D., Yvon-Lewis, S., King, G., Eglinton, T., Shields, M. R., et al. (2015). Positive priming of terrestrially-derived dissolved organic matter in a freshwater microcosm system. Geophys. Res. Lett. 42, 5460–5467. doi: 10.1002/2015gl064765
Borges, A. V., Abril, G., Darchambeau, F., Teodoru, C. R., Deborde, J., Vidal, L. O., et al. (2015). Divergent biophysical controls of aquatic CO2 and CH4 in the World's two largest rivers. Sci. Rep. 5:15614. doi: 10.1038/srep15614
Brenton, R. W., and Arnett, T. L. (1993). Methods of Analysis by the U.S. Geological Survey National Water Quality Laboratory - Determination of Dissolved Organic Carbon by UV-Promoted Persulfate Oxidation and Infrared Spectrometry, Open-File Report 92–480; U. S. Geological Survey.
Butman, D. E., Wilson, H. F., Barnes, R. T., Xenopoulos, M. A., and Raymond, P. A. (2015). Increased mobilization of aged carbon to rivers by human disturbance. Nat. Geosci. 8, 112–116. doi: 10.1038/ngeo2322
Cole, J. J., Prairie, Y. T., Caraco, N. F., McDowell, W. H., Tranvik, L. J., Striegl, R. G., et al. (2007). Plumbing the global carbon cycle: integrating inland waters into the terrestrial carbon budget. Ecosystems 10, 172–185. doi: 10.1007/s10021-006-9013-8
Dahl, T. E. (1990). Wetland Losses in the United States 1780's to 1980's; U.S. Washington, DC: Department of the Interior, Fish and Wildlife Service.
Dahl, T. E., and Johnson, C. E. (1991). Wetlands-Status and Trends in the Conterminous United States, Mid-1970's to Mid-1980's; U.S. Washington, DC: Fish and Wildlife Service.
D'Amore, D. V., Edwards, R. T., and Biles, F. E. (2016). Biophysical controls on dissolved organic carbon concentrations of Alaskan coastal temperate rainforest streams. Aquat. Sci. 78:381. doi: 10.1007/s00027-015-0441-4
Davidson, E. A., and Janssens, I. A. (2006). Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 440, 165–173. doi: 10.1038/nature04514
Depetris, P. J., and Kempe, S. (1993). Carbon dynamics and sources in the Paraná River. Limnol. Oceanogr. 38, 382–395. doi: 10.4319/lo.1993.38.2.0382
Duan, S. W., Allison, M. A., Bianchi, T. S., McKee, B. A., Shiller, A. M., Guo, L., et al. (2014). “Sediment, organic carbon, nutrients, and trace elements: sources, transport, and biogeochemical cycles in the lowermost Mississippi River,” in Biogeochemical Dynamics at Major River-Coastal Interfaces: Linkages with Global Climate Change, Vol. 16, eds T. S. Bianchi, M. A. Allison, and W. J. Cai (Cambridge: Cambridge University Press), 397–420.
Duan, S. W., Bianchi, T. S., Santschi, P. H., and Amon, R. M. W. (2010). Effects of tributary inputs and in-channel processes on nutrient export from the Mississippi and Atchafalaya Rivers. Mar. Freshwater Res. 61, 1029–1038. doi: 10.1071/MF09235
Duan, S. W., Bianchi, T. S., Shiller, A. M., and Dria, K. (2007). Variability in the bulk composition and abundance of dissolved organic matter in the lower Mississippi and Pearl rivers. J. Geophys. Res. Biogeosci. 112:G02024. doi: 10.1029/2006jg000206
Gergel, S. E., Turner, M. G., and Kratz, T. K. (1999). Dissolved organic carbon as an indicator of the scale of watershed influence on lakes and rivers. Ecol. Appl. 91, 1377–1390. doi: 10.1890/1051-0761(1999)009[1377:DOCAAI]2.0.CO;2
Guenet, B., Danger, M., Harrault, L., Allard, B., Jauset-Alcala, M., Bardoux, G., et al. (2014). Fast mineralization of land-born C in inland waters: first experimental evidences of aquatic priming effect. Hydrobiologia 721, 35–44. doi: 10.1007/s10750-013-1635-1
Hedges, J. I., Mayorga, E., Tsamakis, E., McClain, M. E., Aufdenkampe, A., Quay, P., et al. (2000). Organic matter in Bolivian tributaries of the Amazon River: a comparison to the lower mainstream. Limnol. Oceanogr. 45, 1449–1466. doi: 10.4319/lo.2000.45.7.1449
Irena, F., McKnight, D. M., Pellerin, B., Green, M. B., Bergamaschi, B., Aiken, G. R., et al. (2015). The river as a chemostat: fresh perspectives on dissolved organic matter flowing down the river continuum. Can. J. Fish. Aquat. Sci. 72, 1272–1285. doi: 10.1139/cjfas-2014-0400
Kaushal, S. S., and Belt, K. T. (2012). The urban watershed continuum: evolving spatial and temporal dimensions. Urban Ecosyst. 15, 409–435. doi: 10.1007/s11252-012-0226-7
Lara, R. J., Rachold, V., Kattner, G., Hubberten, H. W., Guggenberger, G., Skoog, A., et al. (1998). Dissolved organic matter and nutrients in the Lena River, Siberian Arctic: characteristics and distribution. Mar. Chem. 59, 301–309. doi: 10.1016/S0304-4203(97)00076-5
Leenheer, J. A., Barber, L. B. II, Rostad, C. E., and Noyes, T. I. (1995). Data on Natural Organic Substances in Dissolved, Colloidal, Suspended-Silt, and -Clay and Bed-Sediment Phases in the Mississippi River and Some of its Tributaries, 1991–1992. Report 94–4191, US Geological Survey Water Resources Investigations, Denver, CO.
Massicotte, P., and Frenette, J. J. (2011). Spatial connectivity in a large river system: Resolving the sources and fate of dissolved organic matter. Ecol. Appl. 21, 2600–2617. doi: 10.1890/10-1475.1
McKee, B. (2003). RiOMar: The Transport, Transformation and Fate of Carbon in River-Dominated Ocean Margins. Report of the RiOMarWorkshop, 1–3 November 2001. Tulane University, New Orleans.
Miller, M. P. (2012). The influence of reservoirs, climate, land use and hydrologic conditions on loads and chemical quality of dissolved organic carbon in the Colorado River. Water Resour. Res. 48:W00M02. doi: 10.1029/2012WR012312
Mitsch, W. J., Day, J. W., Zhang, L., and Lane, R. R. (2005). Nitrate-nitrogen retention in wetlands in the Mississippi River Basin. Ecol. Eng. 24, 267–278. doi: 10.1016/j.ecoleng.2005.02.005
Moody, J. A. (1995). Chemical Data for Water Samples Collected during Four Upriver Cruises on the Mississippi River between New Orleans, Louisiana, and Minneapolis, Minnesota, May 1990–April 1992: U.S. Report 94–523, Geological Survey Open-File.
Mulholland, P. J., and Kuenzler, E. J. (1979). Organic carbon export from upland and forested wetland watersheds. Limnol. Oceanogr. 24, 960–966. doi: 10.4319/lo.1979.24.5.0960
Nadon, M. J., Metcalfe, R. A., Williams, C. J., Somers, K. M., and Xenopoulos, M. A. (2015). Assessing the effects of dams and waterpower facilities on riverine dissolved organic matter composition. Hydrobiologia 744, 145–164. doi: 10.1007/s10750-014-2069-0
Philip, G. T., and Townsend, A. R. (2010). Stoichiometric control of organic carbon–nitrate relationships from soils to the sea. Nature 464, 1178–1181. doi: 10.1038/nature08985
Porcal, P., Dillon, P. J., and Molot, L. A. (2015). Temperature dependence of photodegradation of dissolved organic matter to dissolved inorganic carbon and particulate organic carbon. PLoS ONE 10:e0128884. doi: 10.1371/journal.pone.0128884
Prahl, F. G., Small, L. F., Sullivan, B. A., Cordell, J., Simenstad, C. A., Crump, B. C., et al. (1998). Biogeochemical gradients in the lower Columbia River. Hydrobiologia, 361, 37–52. doi: 10.1023/A:1003129124747
Rabotyagov, S. S., Campbell, T. D., White, M., Arnold, J. G., Atwood, J., Norfleet, M. L., et al. (2014). Cost-effective targeting of conservation investments to reduce the northern Gulf of Mexico hypoxic zone. Proc. Natl. Acad. Sci. U.S.A. 111, 18530–18535. doi: 10.1073/pnas.1405837111
Raymond, P. A., Hartmann, J., Lauerwald, R., Sobek, S., McDonald, C., Hoover, M., et al. (2013). Global carbon dioxide emissions from inland waters. Nature 503, 355–359. doi: 10.1038/nature12760
Richey, J. E., Melack, J. M., Aufdenkampe, A. K., Ballester, V. M., and Hess, L. L. (2002). Outgassing from Amazonian rivers and wetlands as a large tropical source of atmospheric CO2. Nature 416, 617–620. doi: 10.1038/416617a
Schlesinger, W. H., and Melack, J. M. (1981). Transport of organic carbon in the world's rivers. Tellus 33, 172–187. doi: 10.1111/j.2153-3490.1981.tb01742.x
Schramm, H. L., Cox, M. S., Tietjen, T. E., and Ezell, A. W. (2009). Nutrient dynamics in the lower Mississippi River floodplain: comparing present and historic hydrologic conditions. Wetlands 29, 476–487. doi: 10.1672/08-62.1
Sedell, J. R., Richey, J. E., and Swanson, F. J. (1989). “The river continuum concept: A basis for the expected ecosystem behavior of very large rivers?” in Proceedings of the International Large River Symposium, ed D. P. Dodge (Ottawa, ON: Canada Department of Fisheries and Oceans), 49–55.
Shih, J.-S., Alexander, R. B., Smith, R. A., Boyer, E. W., Schwarz, G. E., and Chung, S. (2010). An Initial SPARROW Model of Land Use and In-Stream Controls on Total Organic Carbon in Streams of the Conterminous United States, Open-File Report 2010–1276; U. S. Geological Survey.
Spitzy, A., and Leenheer, J. (1991). “Dissolved Organic Carbon in Rivers,” in Biogeochemistry of Major World Rivers, Scope 43, eds E. T. Degens, S. Kempe, and J. E. Richey (Chichester: J. Wiley & Sons), 213–232.
Stanley, E. H., Powers, S. M., Lottig, N. R., Buffam, I., and Crawford, J. T. (2012). Contemporary changes in dissolved organic carbon of human dominated rivers: is there a role for DOC management? Freshwater Biol. 57, 26–42. doi: 10.1111/j.1365-2427.2011.02613.x
Struyf, E., and Conley, D. J. (2009). Silica: an essential nutrient in wetland biogeochemistry. Front. Ecol. Environ. 7, 88–94. doi: 10.1890/070126
Tockner, K., Pennetzdorfer, D., Reiner, N., Schiemer, F., and Ward, J. V. (1999). Hydrological connectivity, and the exchange of organic matter and nutrients in a dynamic river-floodplain system (Danube, Austria). Freshwater Biol. 41, 521–535. doi: 10.1046/j.1365-2427.1999.00399.x
Vannote, R. L., Minshall, G. W., Cummins, K. W., Sedell, J. R., and Cushing, C. E. (1980). The River continuum concept. Can. J. Fish. Aquat. Sci. 37, 130–137. doi: 10.1139/f80-017
Ward, N. D., Bianchi, T. S., Sawakuchi, H. O., Gagne-Maynard, W., Cunha, A. C., Brito, D. C., et al. (2016). The reactivity of plant-derived organic matter and the potential importance of priming effects in the lower Amazon River. J. Geophys. Res. Biogeosci. 121, 1522–1539. doi: 10.1002/2016jg003342
Ward, N. D., Keil, R. G., Medeiros, P. M., Brito, D. C., Cunha, A. C., Dittmar, T., et al. (2013). Degradation of terrestrially derived macromolecules in the Amazon River. Nat. Geosci. 6, 530–533. doi: 10.1038/ngeo1817
Ward, N. D., Krusche, A. V., Sawakuchi, H. O., Brito, D. C., Cunha, A. C., Moura, J. M. S., et al. (2015). The compositional evolution of dissolved and particulate organic matter along the lower Amazon River-Óbidos to the ocean. Mar. Chem. 177, 244–256. doi: 10.1016/j.marchem.2015.06.013
Wehr, J. D., Lonergan, S. P., and Thorp, J. H. (1997). Concentrations and controls of dissolved organic matter in a constricted-channel region of the Ohio River. Biogeochemistry 38, 41–65. doi: 10.1023/A:1005708326368
Wiener, J. G., Fremling, C. R., Korschgen, C. E., Kenow, K. P., Kirsch, E. M., Rogers, S. J., et al. (1996). “Mississippi River,” in Status and Trends of the Nation's Biological Resources, eds M. J. Mac, P. A. Opler, C. E. P. Haecker, and P. D. Doran (Washington, DC: National Weather Research Center U.S. Geological Survey), 351–384.
Wilde, F. D (2006). Temperature (ver. 3.0): U.S. Geological Survey Techniques of Water-Resources Investigations, Book 9, Chap. A6., Sec. 6.1. Available online at: http://pubs.water.usgs.gov/twri9A6/
Wu, Y., Zhang, J., Liu, S. M., Zhang, Z. F., Yao, Q. Z., Hong, G. H., et al. (2007). Sources and distribution of carbon within the Yangtze River system. Estuar. Coast. Shelf Sci. 71, 13–25. doi: 10.1016/j.ecss.2006.08.016
Zhang, L., Xue, M., Wang, M., Cai, W.-J., Wang, L., and Yu, Z.-G. (2014). The spatiotemporal distribution of dissolved inorganic and organic carbon in the main stem of the Changjiang (Yangtze) River and the effect of the Three Gorges Reservoir. J. Geophys. Res. Biogeosci. 119, 741–757. doi: 10.1002/2012jg002230
Keywords: dissolved organic carbon, wetlands, Mississippi River Basin, river continuum, in-stream processes, land use-land cover
Citation: Duan S, He Y, Kaushal SS, Bianchi TS, Ward ND and Guo L (2017) Impact of Wetland Decline on Decreasing Dissolved Organic Carbon Concentrations along the Mississippi River Continuum. Front. Mar. Sci. 3:280. doi: 10.3389/fmars.2016.00280
Received: 26 September 2016; Accepted: 13 December 2016;
Published: 09 January 2017.
Edited by:
Javier Arístegui, University of Las Palmas de Gran Canaria, SpainReviewed by:
X. Antón Álvarez-Salgado, Spanish National Research Council, SpainJie Xu, South China Sea Institute of Oceanology (CAS), China
Copyright © 2017 Duan, He, Kaushal, Bianchi, Ward and Guo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shuiwang Duan, c2R1YW5AdW1kLmVkdQ==