 Mar Bosch-Belmar1,2*
Mar Bosch-Belmar1,2* Giacomo Milisenda3
Giacomo Milisenda3 Albert Girons4Valentina Taurisano1
Albert Girons4Valentina Taurisano1 Stefano Accoroni5
Stefano Accoroni5 Cecilia Totti5
Cecilia Totti5 Stefano Piraino1,2*†
Stefano Piraino1,2*† Verónica Fuentes6†
Verónica Fuentes6†- 1Dipartimento di Scienze e Tecnologie Biologiche ed Ambientali, Università del Salento, Lecce, Italy
- 2Consorzio Nazionale Interuniversitario per le Scienze del Mare, Rome, Italy
- 3Dipartimento Terra e Ambiente, CNR—IAMC, Mazara del Vallo, Italy
- 4ICTIOVET S.L., Barcelona, Spain
- 5Dipartimento di Scienze della Vita e dell'Ambiente, Università Politecnica delle Marche, Ancona, Italy
- 6Institut de Ciències del Mar, ICM-CSIC, Barcelona, Spain
In recent years, caged finfish mariculture across European seas suffered production losses by severe fish mortality, following episodic outbreaks of invertebrate cnidarian stingers. Due to their stinging cells and injectable venoms, medusozoan jellyfish, or drifting propagules of polyp colonies at high density may impair caged fish health through toxic effects on vulnerable tissues of gills and skin, and related secondary bacterial infections. Gill disorders in European sea bass (Dicentrarchus labrax) fish farms along the Spanish Mediterranean coast are commonly reported, but regular monitoring of the frequency of cnidarian outbreaks and their potential impacts on caged fish is still poorly enforced. In this study, two sea bass mariculture farms in Southern Spain (Málaga; Almería) were monitored biweekly for zooplankton, phytoplankton and fish gills condition, over 13 or 30 months for the Málaga and Almería facilities, respectively, within the period 2012–2014. Significant, direct correlations were found among low water temperature, recorded fish mortalities, and high abundances of planktonic cnidarians, particularly of the hydrozoan siphonophores Muggiaea atlantica and M. kochii, and the larval stage of Ectopleura larynx, a common member of cage biofouling communities. A significant relationship between cnidarian densities and the quantitative scoring of gill pathology was also observed. In addition, high densities of long-bristled planktonic diatoms (Chaetoceros spp.) coincided with a major fish mortality event (April 2012, Almería farm). Standardised monitoring of plankton dynamics and composition may help in promoting response capacities of Mediterranean mariculture managers to fish health challenges (such as stinging plankton blooms) by (a) improving diagnostic tools and preventative countermeasures and (b) supporting the development of science-based spatial planning and sustainable growth of coastal mariculture.
Introduction
With 29% of wild fish stocks exploited at biologically unsustainable levels, and global capture fisheries almost at same yield in the last 30 years, marine aquaculture has undergone a rapid expansion worldwide, being now the major source of increasing fish food supply, with large potential for economic growth (FAO, 2016). Stinging jellyfish proliferations can affect marine fish farming (Baxter et al., 2011a; Rodger et al., 2011a; Purcell et al., 2013), with negative repercussions on highly productive coastal and offshore systems.
In recent years, facilities from Asia, Australia, North and South America reported sudden fish mortalities as consequences of episodic blooms of different jellyfish species (Palma et al., 2007; Doyle et al., 2008; Willcox et al., 2008; Purcell et al., 2013). Physical contact of sensitive fish tissues (skin and gills) with conspicuous semeostomeae scyphozoan jellyfish Pelagia noctiluca and Aurelia aurita has been identified as the primary agent of repeated mass fish deaths (in 2007, 2010, 2013, and 2014) at salmon facilities in northern Europe (Doyle et al., 2008; FIS, 2014; Marcos-López et al., 2016). Stinging jellies are washed up on fish cages by currents, and can be either broken in pieces or their early stages may be small enough to enter the net cages and get in touch with caged fishes. In parallel, a few studies reported the impact of less conspicuous planktonic stingers on mariculture, with best evidence on the hydrozoan siphonophore Muggiaea atlantica and the microscopic hydromedusae Solmaris corona and Phialella quadrata as determinants of fish mortalities in Irish and Scottish fish farms (Baxter et al., 2011a; Fitridge and Keough, 2013; Purcell et al., 2013).
Additional issues arise for aquaculture from biofouling. Together with the restriction of water flow through net occlusions, the development of large hydrozoan polyp colonies on cage nets eventually lead to seasonal release of free-living propagules inside the cages, such small jellyfish (e.g., Obelia spp.), or the larvae of progenetic hydroids (e.g., the tubulariid Ectopleura larynx; Baxter et al., 2012). Tiny and almost transparent medusae, siphonophores, and hydrozoan larvae may easily remain unnoticed to aquaculture operators, but they can develop high density blooms, be inhaled by fish, and inflict severe damage due to nematocyst discharge and consequent tissues envenomation (Fosså et al., 2003). A range of morphological injuries caused by contacts with cnidarian stinging cells have been described on several cultured fish species (Baxter et al., 2011b; Bosch-Belmar et al., 2016b; Marcos-López et al., 2016). Available evidence shows that a few hours of exposure to stinging cells may lead to severe damage at cellular, tissue and systemic levels, including changes in fish metabolic performances (Bosch-Belmar et al., 2016a).
Gill disorders are considered to be a major problem for fish aquaculture and are usually referred to any of different causes, including jellyfish, phytoplankton, parasites, bacteria, and viruses (Rodger, 2007). To date, several marine fish farms in northern European seas have started plankton monitoring as standard preventative tool against potential impacts. On the contrary, little or no plankton monitoring activities have been launched in Mediterranean mariculture facilities.
Since 2009, two off-shore fish farms located in southern Spain (Almería and Málaga) suffered repeated mortality events of caged European sea bass (Dicentrarchus labrax), for which no causative agent was clearly identified. Specifically, bacterial and known viral diseases, parasites or hypoxic events in the fish cages were excluded by veterinary controls. Injured sea bass showed clinical signs similar to those described by Rodger et al. (2011a) in farmed salmons exposed to jellyfish blooms in the Irish Sea: lethargic behavior, atypical swimming close to the surface, visible respiratory distress, interrupted feeding, and dead or moribund individuals. For this reason, in 2012 a combined plankton vs. fish gill survey was launched at both aquaculture facilities to [I] disclose the spatial and seasonal variability of the plankton community accessing the fish cages; and [II] test the hypothesis of a direct relationship between the occurrence of potentially harmful plankton (i.e., stinging species) and fish health.
Materials and Methods
Study Sites
Aquaculture facilities were located 206 km apart in the Málaga Bay and the Almería Gulf in the Alboran Sea (Figure 1), a transitional area connecting the Atlantic Ocean to the Western Mediterranean. Characterized by high hydrodynamics, upwelling of cold, nutrient-rich subsurface waters (Sarhan et al., 2000), the Alboran Sea basin is a highly productive region in the upper 200 m layer for zooplankton (Thibault et al., 1994) and near-surface phytoplankton (Sanchez-Vidal et al., 2004; Navarro et al., 2011).
Figure 1. Fish farms locations in the Spanish coast of the Alboran Sea: (1) Málaga Bay and (2) Almería Gulf.
Both fish farms are off-shore facilities for European sea bass (D. labrax) and Gilthead sea bream (Sparus aurata) production in floating cages, from 15 g fry to commercial size (300–500 g). Floating cages of 25 m in diameter and 10 m in depth were placed at sites with water depth ranging 30–50 m.
Sampling and Sample Processing
Monitoring was performed from January 2012 to June 2014 for the Almería facility and from June 2013 to June 2014 for the Málaga fish farm. Temperature and fish stocks mortality data were provided by the veterinary staff of the facilities. Temperature was recorded daily at 5 m depth by a sensor (Oxyguard) located in the cages and fish mortality was reported on a weekly basis. Only mortality events without a clearly identified causative agent and with fish pathologies as described by Rodger et al. (2011a) were considered for further analyses.
Zooplankton samples were collected biweekly, using a net of 200-μm mesh with a filtering cod-end and a digital flow meter (Hydrobios, model 438110) to determine the volume of filtered water. To characterize the potentially harmful plankton community in the facility area, three replicate vertical net hauls were carried out at each sampling sites, located in the close proximity of cage nets (two cages in Almería, one cage in Málaga), and near the northern and southern perimetric buoys of the two facilities (at 350–500 m distance from fish cages), from −10 m depth to surface. In a preceding 8-month pilot study with the same sampling methodology at the Almería fish farm, no significant differences in monthly plankton densities inside and outside the fish cages (F1 = 1.90, p = 0.07) were recorded. Thus, the plankton survey in this study was carried out immediately outside the cages, to prevent any stress on the farmed fish by the sampling activities. A number of 12 or 9 samples for each sampling time were collected in the Almería and Málaga fish farms, respectively.
All zooplankton samples were preserved in a 4% neutral buffered formalin solution. In the laboratory, gelatinous zooplankton was quantified (individuals m−3) and sorted in 5 different taxa (Cnidaria, Polychaeta, Larvacea, Thaliacea, Chaetognatha). Cnidarian zooplankton was identified to genus or species level in most cases. Crustacean taxa were not taken in consideration in this study because there is no available evidence of crustaceans, apart from easily detectable ectoparasites, affecting fish health through gill, and skin lesions.
Phytoplankton samples were collected at the same time and sites as zooplankton, by use of a Lund tube (a weighted polyethylene tube 2 cm in diameter) of 5 m length (Lund and Talling, 1957). From the total sample, a subsample of 500 ml was taken after homogenization, and was preserved by adding 1 ml of neutral Lugol's Iodine solution. In laboratory, a variable subsample volume (25–63 ml depending on the abundance observed) was settled in an Utermöhl chamber (Edler and Elbrächter, 2010). Species identification and counting were performed using an inverted microscope (Zeiss Axiovert 135) equipped with phase contrast at 400x magnification, in 10–30 random fields, in order to obtain a significant cell number. For each sample, abundances were expressed as number of cells per liter (cells l−1). Results were screened for species potentially harmful for farmed fish.
Five to ten D. labrax specimens were randomly sampled for gill analysis from monitoring cages routinely once a month and every time that mortality in cages was recorded. During customary welfare analyses, qualified veterinary staff of the farms randomly caught fish specimens by hand net (avoiding moribund specimens to ensure best representation of the caged population), and immediately killed them by a combination of rapid clove oil anesthesia followed by chilling in ice/water slurry, in accordance with the best practices for welfare and stunning/killing procedures on cultured sea bass/sea bream in the Mediterranean (EFSA, 2009; Panagiotis et al., 2014). By working on dead animals provided by the fish farms, this study did not require approval by any ethics committee according to University of Salento guidelines and Italian national regulations.
Gill samples were taken from the second gill arch and were fixed in 10% neutral-buffered formalin. Samples were embedded in paraffin and 4-μm sections were stained with hematoxylin and eosin standard protocol. Slides were examined microscopically at 50x, 100x, and 400x magnifications. The gill score protocol created by Mitchell et al. (2012) was used to quantify gill damage. The index criteria for gill histopathology were lamellar hyperplasia, lamellar fusion, cellular anomalies (necrosis or sloughing), and lamellar oedema. A score from 0 to 3 was assigned for each parameter depending on injury extent (1: <10% affected surface, 2: 10–50%, and 3: >50% of the gill epithelium injured). Ancillary criteria, such as hypertrophy, hemorrhage and the presence of specific pathogens, were assigned a score of 0 or 1. Pathogens affecting sea bass included the parasites Cryptocaryon sp., the monogenea Diplectanum sp., filamentous bacteria Tenacibaculum sp. and Epiteliocystis intracellular bacterial colonies. The injury scores were summed and total scores between 1 and 3 were considered to be typical of gills regularly observed in off-shore marine-farmed fish (A. Girons, pers. obs.).
Statistical Analyses
Multiple analyses of variance—(Anderson, 2001) PRIMER software—were performed in order to test for differences in the gelatinous zooplankton assemblage among covariates (date, farm, sampling site). The experimental design was composed by three factors: (1) “Date,” fixed with 12 levels; (2) “location,” fixed and orthogonal with 2 levels; and (3) “sampling site,” fixed and orthogonal with 2 levels. Moreover, the species that contributed most to the similarity in each location were characterized using the SIMPER routine (Clarke, 1993).
An ordination of the zooplankton community based on the density of each group was obtained with a principal component analysis (PCA) in order to display graphically samples aggregation by taxa and to perform a preliminary assessment between biotic and abiotic variables. The PCA analysis was performed with the R-language function Princomp, which is available in the Vegan library (Oksanen et al., 2005) of the R software platform. Generalized linear models (GLM) with a binomial distribution and a logit link were used to describe the relationship between different densities of the gelatinous zooplankton (independent variable) and the presence/absence of mortality events (response variable).
During the monitoring period four atypical mortality events—i.e., not referable to known bacteria, viruses, parasites or hypoxia events—were recorded in the Almería facility (April 2012, May 2012, January 2013, and March 2014) and only one in the Málaga fish farm (November–December 2013). For this reason, analyses focused on the relation between stocks mortality and plankton densities over time were performed only for Almería.
GLM model with a Poisson error family and a logit link was applied to test for a relationship between the total fish gill score and cnidarian density in both facilities, after checking for the normality and homoscedastic condition. The same model and distribution was employed to describe the variation in density of harmful cnidarian species among different temperature values in the Almería facility. The relationship between cnidarian density and temperature in the Málaga fish farm was investigated through a generalized additive models (GAMs) with Poisson distribution, because of the non-linear relationship between variables (Wood, 2006). These analyses were performed using the free statistical software R, version 3.2.3 (http://cran.r-project.org).
Results
Harmful Zooplankters
The overall abundance of gelatinous zooplankton was tested for differences over the sampling period, between fish farms, and between sampling sites (near-cages or perimetric sites) at each farm. A total number of 608 zooplankton samples were analyzed, with identification of 26 taxa of cnidarians (25 hydrozoans and 1 scyphozoan) (Supplementary Table I). Differences in total gelatinous zooplankton densities between near-cages and perimetric sampling sites were not significant (p > 0.05); however, the gelatinous zooplankton community changed significantly between facilities and over time (Table 1, Supplementary Figures 1, 2).
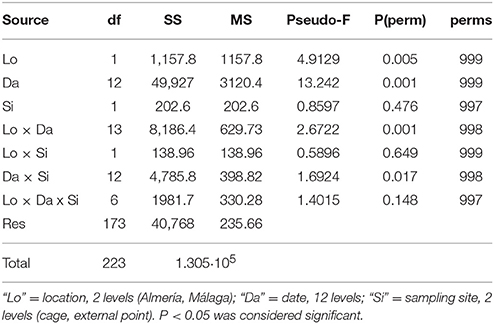
Table 1. Multiple analyses of variance (PERMANOVA) of the gelatinous zooplankton community comparing the Almería and Málaga aquaculture facilities (location), sampling sites (cage, external point), and sampling dates (date).
Four potentially harmful cnidarian species, previously known to be involved in mass mortalities of farmed salmons in northern Europe, were present in the Almería and Málaga fish farms: M. atlantica, P. quadrata, S. corona, and P. noctiluca. The hydrozoan siphonophore M. atlantica was present in both facilities, with peaks in spring 2012 (Almería), autumn 2013 (Málaga) and March 2014 (Almería and Málaga fish farms), with high abundances of both the polygastric colonial stages and the free living sexual stage (eudoxid) (up to 60 and 98 stages m−3, respectively). The hydromedusae P. quadrata and S. corona occurred at low densities with autumn abundance peaks (between 3 and 8 ind m−3) in 2013 in both facilities. The ephyra stages of the scyphozoan P. noctiluca were observed throughout the monitoring period at low-medium densities (from 1.50 to 15 ind m−3) in both facilities.
SIMPER analysis (Table 2) showed the cnidarian assemblage differed between facilities, with an average dissimilarity of 88%. The most representative species at the Almería fish farm were Podocorynoides minuta, Aglaura hemistoma, and Obelia dichotoma, while the most abundant species were O. dichotoma, M. atlantica, and M. kochii. In the Málaga fish farm, the most representative species were M. atlantica, P. minuta, and Eucheilota paradoxica. These species were also the most abundant, together with Stauridiosarsia gemmifera. Additional species typical of offshore waters were also recorded at low densities in both facilities (i.e., the trachymedusa Solmundella bitentaculata and the siphonophores Chelophyes appendiculata and Abylopsis tetragona).
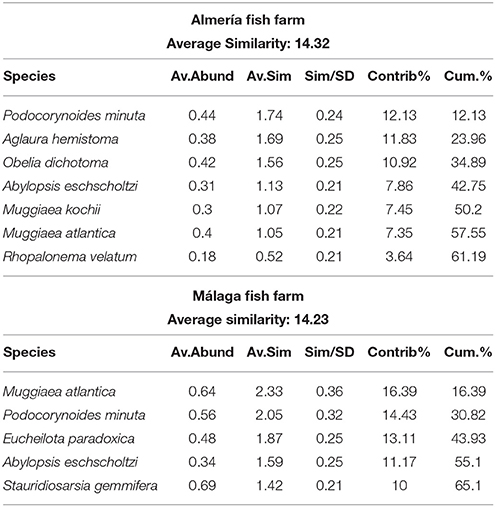
Table 2. SIMPER analysis on cnidarian species at both facilities.
Relationships with Fish Mortalities
To display potential taxon-specific relationships among fish mortalities recorded in the Almería fish farm, different gelatinous zooplankton groups and temperature, a PCA model was carried out (Figure 2), which displayed similar spatial trends in the PCA environment between fish mortalities and cnidarians.
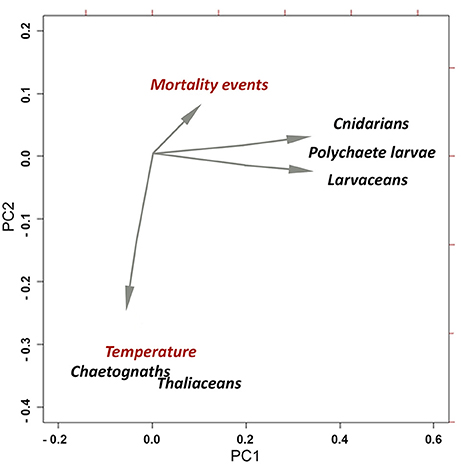
Figure 2. Principal component analysis (PCA) of gelatinous zooplankton (cnidarians, polychaete larvae, larvaceans, chaetognaths, and thaliaceans), fish mortality and temperature in the Almería aquaculture facility.
GLM analyses showed that the only significant and positive relationship with fish mortality in the Almería facility were due to coincidence with cnidarian abundances (Z1 = 4.04, p = 5·10−5; Figure 3) when the average temperature was 15.75 ± 0.52°C (mean ± SE). Among the identified cnidarians, each of three species were significantly related with fish mortalities: the calycophoran siphonophores M. kochii (Z1 = 3.35, p = 0.02) and M. atlantica (Z1 = 2.55, p = 0.01), and the free-living actinula larvae of the hydroid E. larynx (Z1 = 2.46, p = 0.01; Figure 4). Furthermore, fish mortalities occurred during coincident, multispecific blooms of cnidarians (Supplementary Figure 1). Temperature was significantly and negatively related with mortality (Z1 = −5.69, p = 1.24·10−8) and equally significantly and negatively related with the above cnidarian species (Z1 = −20.33, p = 2·10−16; Figure 5).
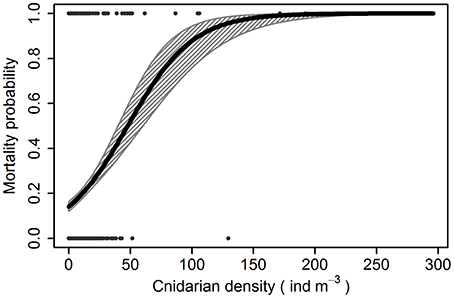
Figure 3. Probability of a fish mortality event at different densities of cnidarians in the Almería facility. The dots at the top and bottom axes represent the mortality presence/absence over the monitoring.
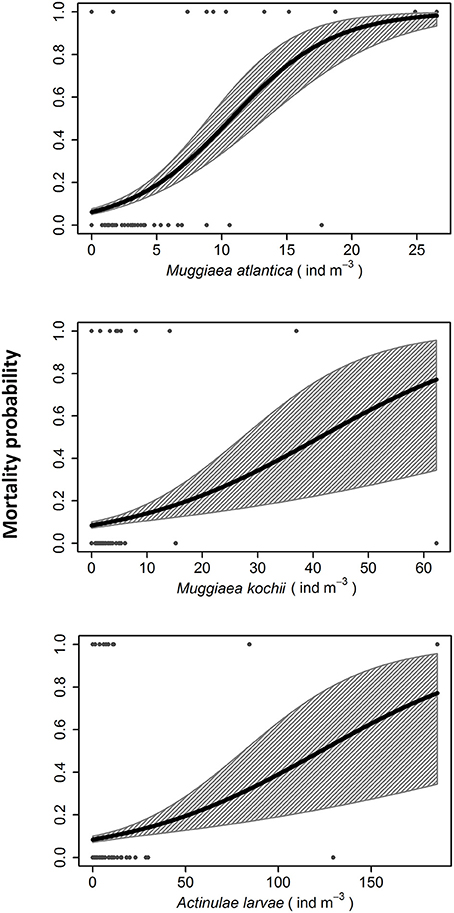
Figure 4. Probability of fish mortality event at different densities of identified harmful cnidarian species (Muggiaea atlantica, Muggiaea kochii, and Ectopleura larynx actinula larvae). The dots at the top and bottom axes represent the mortality presence/absence over the monitoring.
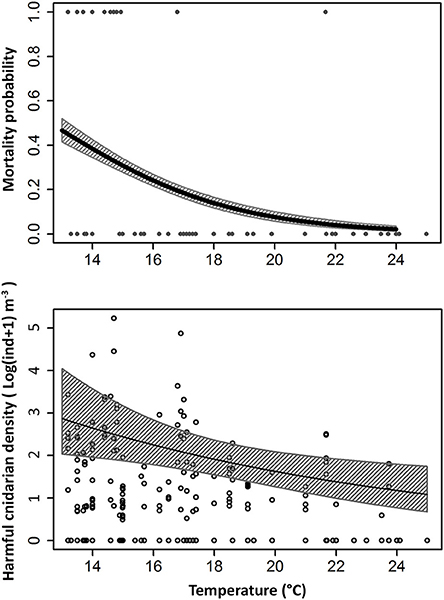
Figure 5. Probability of fish mortality event at different water temperatures (above) and relationship between harmful cnidarian densities and water temperature (below).
In the Málaga facility, significant inverse relation between cnidarian density and temperature was detected (χ2 = 780.50, p = 2·10−16). The highest cnidarian densities were recorded at temperatures ranging from 15 to 19°C, but drastically decreased above 20°C. Temperature was 16.30 ± 0.45°C when fish mortality was recorded (Nov-Dec 2013), paralleled by high abundance of M. atlantica (polygastric and eudoxid stages) and O. dichotoma medusae.
Harmful Phytoplankters
More than 70 taxa of phytoplankton were identified in collected samples from both facilities. Several microalgae genera known to be potentially harmful for fish health were recorded at high densities in both fish farms. Among diatoms, a number of Chaetoceros species (i.e., C. lorenzianus, C. laciniosus, and C. didymus) occurred several times throughout the monitoring period in both facilities, with a density peak of 8.43·105 cells l−1 in April 2012 in the Almería fish farm, when fish mortality was recorded. The silicoflagellate Dictyocha speculum was also found at high densities in the Málaga installation (2.60·103 cells l−1) and Pseudo-nitzschia spp. occurred during 2012, with an abundance peak in August (3.1·105 cells l−1), but the presence of both species did not coincide with fish mortality events.
Gill Damage
Histological analyses from both facilities revealed moderate to severe damage in the fish gills, including generalized inflammation of gill epithelium, lamellar hyperplasia and fusion, oedema and in many cases necrotic patches with advanced bacterial infection (Figure 6). Gill scoring analysis showed different peaks of severe gill damage in the sampled fish. Regression between histological scores and total cnidarian densities was significant and positive (Z1 = 4.34, p = 1.44·10−5) for the Almería fish farm (Figure 7). High farmed fish mortalities were recorded four time over the monitoring (April–May 2012, January 2013, and March 2014), but elevated scores were also observed in November 2012 and April 2014 (Figure 8).
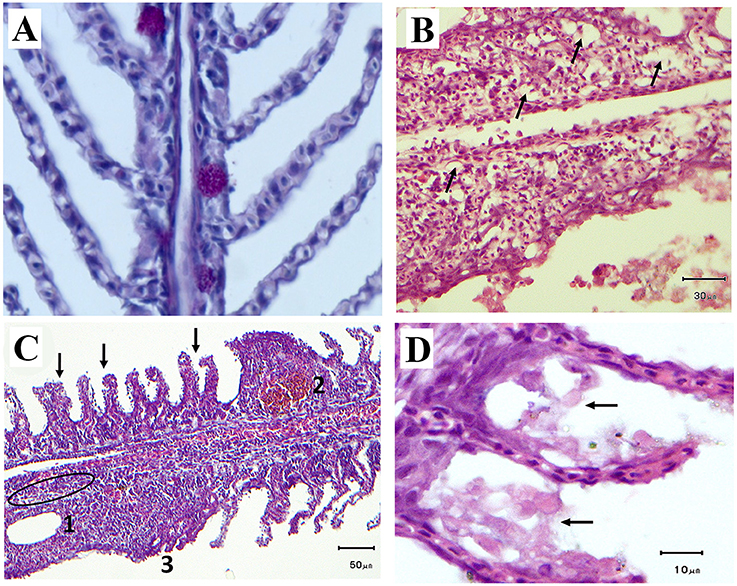
Figure 6. (A) Healthy gills with undamaged primary lamellae and presence of mucous cells (400x); (B–D). Pathological features in gills from sampled Dicentrarchus labrax at aquaculture facilities in Spain; (B) Lamellar oedema (black arrows); (C) Hyperplasia of primary lamellae (black arrows), lymphocytes accumulation (black circle), and 1, lacunae formation; 2, circulatory disturbances (telangectasia); 3, lamellar fusion; (D) cellular degeneration.
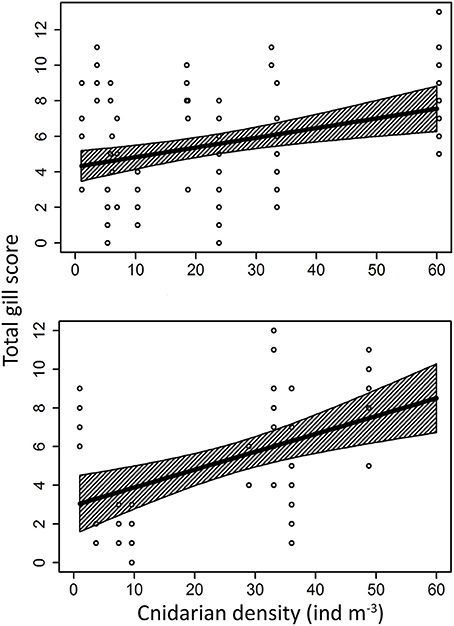
Figure 7. Generalized linear model between total gill score and cnidarian density for the Almería facility (above) and the Málaga fish farm (below).
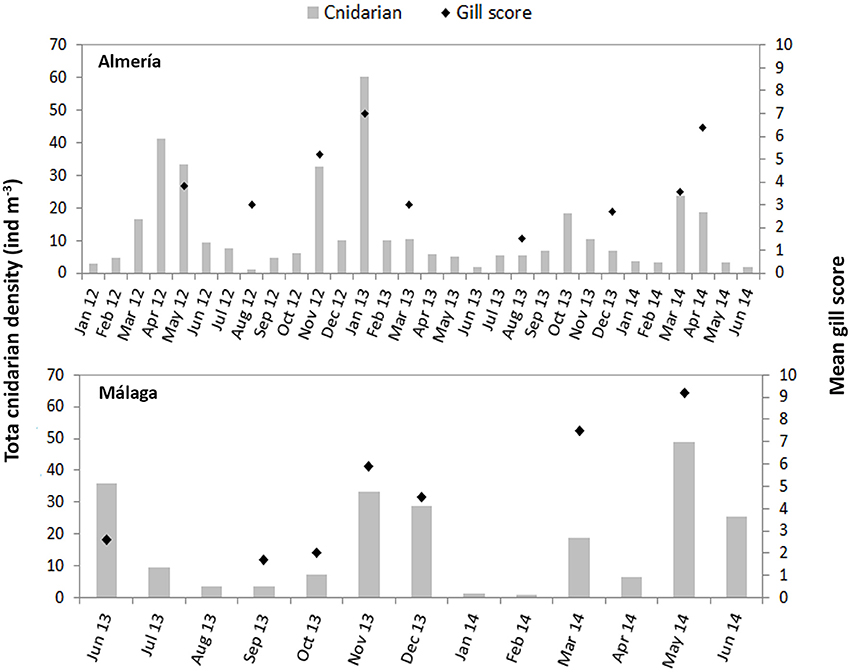
Figure 8. Cnidarian densities and gill scores over time for the Almería and Málaga fish farms.
Significant and positive regression between gill scores and cnidarian densities was also observed for the Málaga fish farm (Z1 = 5.03, p = 4.83·10−7; Figure 7). The only fish mortality event recorded in this facility was at the end of November and continued in December 2013, but cnidarian density peaks associated with high gill damage scores were also observed in March and May 2014 (Figure 8).
Discussion
To our knowledge, this is the first survey on potentially harmful plankton species carried out to investigate their role as hidden causative agents of farmed fish mortalities in Mediterranean aquaculture facilities. Farmed sea bass mortalities in the two facilities were more clearly related to spring and autumn water temperature and the peaks of abundance of three hydrozoan species, the siphonophores M. atlantica and M. kochii and the actinula larvae of the fouling hydroid colonies of E. larynx.
Muggiaea atlantica was previously identified as a potentially harmful species for marine aquaculture in northern Europe, together with S. corona and P. quadrata (Baxter et al., 2011a). Those species were also identified in samples from the two Mediterranean facilities but statistical analyses showed non-significant correlation of both hydromedusae densities and fish mortalities (Z1 = −0.01, p = 0.99 and Z1 = −0.002, p = 0.10 for S. corona and P. quadrata, respectively). High abundances of Muggiaea spp. eudoxid stages were recorded several times in both fish farms. Each calycophoran siphonophore polygastric stage asexually produces several eudoxids, a sexually-reproductive stage that feeds with stinging tentacles, as is the polygastric stage. High abundances of eudoxids could be detrimental for farmed fish health because hundreds of cnidocytes are present in each tentacle and may severely damage sensitive fish tissues. Mortality recorded in November-December 2013 in the Málaga facility coincided with a Muggiaea spp. reproductive event, reaching high abundances of polygastric colonies and tens of eudoxids in the water column.
The appearance of the microscopic actinula larvae of E. larynx were also correlated with fish kill events in the Almería facility. The actinulae were present when temperatures were low, from January to May and from November to December in both facilities. Over the course of the monitoring, several reproductive periods were observed, with the released actinula larvae reaching very high densities in the water during January 2013 (>200 ind m−3) when thousands of fish died (Supplementary Figure 1B). This hydroid usually forms part of cage fouling community in the North Sea where is a severe problem for fish farms (Guenther et al., 2010; Carl et al., 2011; Baxter et al., 2012) and also in the Mediterranean Sea aquaculture facilities (facilities staff, pers. obs.). Guenther et al. (2010) demonstrated that after washing to clean the nets, this species regrows and occludes the net apertures rapidly. Underwater cleaning of the net cages resulted in higher numbers of E. larynx actinulae and polyps in the water column that could significantly affect caged fish health by injuring gill tissue after contact (Carl et al., 2011; Baxter et al., 2012).
The scyphozoan mauve stinger P. noctiluca is the most abundant and one of the most venemous jellyfish both in the Mediterranean Sea and in eastern Atlantic waters (Russell, 1970), with the potential to reproduce all year long in some areas, such as the Strait of Messina, Italy (Milisenda et al., in press). Due to its envenomation potential and large population outbreaks, it is renowned as one of the most dangerous species for the European mariculture fish farms (Marcos-López et al., 2016). In zooplankton samples, P. noctiluca ephyra stages, deriving from recent metamorphosis of planula larvae, were observed at densities up to 15 ind m−3 several times in both facilities (mainly in February, June, and October). Adult P. noctiluca medusa were not collected by the vertical hauls during the monitoring period, however their occurrence at low density (≤1 ind m−3) were visually recorded by the facility operators (unpublished data). In preceding years, the monitored facilities endured large aggregations of P. noctiluca adult medusae, leading in 2011 to severe fish mortalities (10 tons) with associated economic consequences (Bosch-Belmar et al., 2017).
Obelia dichotoma is another common hydrozoan species in Mediterranean coastal areas (Bouillon et al., 2004; González-Duarte et al., 2016). This species was observed in the Almería and Málaga facilities throughout the year, both as hydroids and medusae forming part of biofouling and zooplankton communities. The small hydromedusae were found at high densities in the Almería facility (maximum of 197.35 ind m−3) when fish stock mortality was recorded in April 2012. However, despite its high concentrations, a significant relationship of this species with the recorded fish kill events was not found.
Harmful algal blooms produced by dinoflagellates, diatoms, silicoflagellates, raphidophytes, and prymnesiophytes (Granéli and Turner, 2006), are often associated to fish mortality episodes worldwide (Treasurer et al., 2003; Burridge et al., 2010). In our study, some phytoplankton taxa previously related with farmed fish kill events were recorded at high densities in both fish farms (Cembella et al., 2002; Treasurer et al., 2003). Chaetoceros is a colonial diatom genus characterized by long setae, which either can clogging gills causing asphyxia or can penetrate the gill tissues causing histological damages (Smayda, 2006). Fish kills attributed to Chaetoceros occurred in Canada, USA and Scotland where abundances of 105 cells l−1 were recorded (Rensel, 1992; Treasurer et al., 2003). In analyzed samples, we recorded abundances of the same order of magnitude only during one farmed fish mortality event recorded in April 2012 in the Almería fish farm, therefore Chaetoceros should be taken into account as plankton species that may had contributed to fish mortality events.
On this respect, it is worth noting fish mortalities in the two Spanish facilities were detected in months characterized by either large peaks of abundance of renowned harmful species (Muggiaea spp., E. larynx) or by the coincident abundance of different plankters not always recognized as harmful taxa (including phytoplankters, P. noctiluca ephyrae, small hydromedusae) (Supplementary Figures 1, 2). This evidence may lead to the hypothesis that concurrent planktonic cnidarians (and even organisms with long stiff bristles, such as diatom phytoplankton cells) may also play a potential impact as guild (Piraino et al., 2002; Boero et al., 2008) because of their cumulative stinging potential, rather than as single species. Overall, low-medium densities of high number of mild stingers might produce similar effects as high concentrations of a single powerful stinger, both in open water and in fish farms. What makes the difference between low, medium, or highly harmful plankters? For cnidarians, it has been suggested that jellyfish stings of some species maybe more harmful than others not only because of chemical differences among venom types but also due to differential cnidocyst structure, with particular reference to the length of different stinging filaments, which may penetrate prey tissues at different depths (Purcell, 1997; Kitatani et al., 2015). If this would be confirmed as a general rule, it might be possible to predict the potential toxicity and harm for fish farms of cnidarian plankters (jellyfish, larvae, hydroid propagules) by investigating their cnidome composition, abundance, and cnidocyst morphology.
Total cnidarian densities were related with temperatures, with reduced densities during summer period, especially for species with a pelago-benthic life cycle. Previous observations revealed a typical summer impoverishment of the Mediterranean Sea filter feeders, including the disappearance of small hydrozoan colonies, entering in a resting phase as dormant creeping stolons (Boero and Fresi, 1986; Bavestrello et al., 2006; González-Duarte et al., 2013). In the Almería farm, the fish mortalities were significantly related to cold months (Figure 5). In the Málaga facility, even though it was not possible to perform regression analyses, the unique recorded mortality event occurred at December 2013, coincident with high cnidarian densities (up to 30 ind m−3).
Gill scoring presented a significant positive relationship with cnidarian abundances, and demonstrated the existence of severe gill disorders related with high cnidarian densities even when fish mortalities were not recorded. For example, in April 2014 in Almería and May 2014 in the Málaga fish farm, gill scores were 6 ± 1.10 and 9.20 ± 0.60, respectively, and jellyfish occurred in medium-high densities (19.71 ± 3.07 ind m−3 in the Almería pen and 48.80 ± 10.79 ind m−3 in the Málaga facility) but no fish mortalities were reported in these periods. Experimental studies with the scyphozoan A. aurita showed gill epithelium recovery required 2 weeks after even brief (10 h) contact between jellyfish and salmonids (Baxter et al., 2011b). Equally, laboratory experiments carried out by Bosch-Belmar et al. (2016b), demonstrated partial recovery of sea bream gill tissue after 3 weeks from fish exposure to medium densities of P. noctiluca. These gill disorders used to pass unnoticed, but the consequences of this stressful situation (repeated contact with jellyfish) could be severe, and even if fish gill epithelium could recovered, fish metabolic performances may be affected and in the long term also fish growth could be compromised (Rodger et al., 2011a,b; Bosch-Belmar et al., 2016a).
This study allowed to identify some blooming and potentially injuring hydrozoan jellyfish species occurring in the monitored mariculture facilities, and to determine the range of temperatures boosting jellyfish proliferations. The optimal thermal values are typical of spring-autumn months, when small sea bass juveniles (15 g in weight) are usually transferred in cages at sea. As a consequence, most mortalities were recorded in different stocks with fish weights between 20 and 70 g. The interaction of biotic and abiotic factors together with the changing fish metabolism adapting to new conditions could create severe stressful situation for fingerling fish. Reduction of fish stock numbers or variation in juveniles' cages introduction times could represent some of the preventive actions to minimize the impact of jellyfish blooms on farmed fish health. Due to the growth of the aquaculture sector and the increased frequency of jellyfish blooms in some coastal waters, the negative interactions of stinging jellyfish on caged finfish is expected to become a substantial issue producing highly relevant economic losses (Purcell et al., 2013). Regular plankton monitoring will be crucial to gather site-specific information on potential harmful species, including their seasonal occurrence and densities, and predict potential impacts of harmful plankters in fish gill disorders and mortalities.
Author Contributions
MB, VF, and SP conceived the study. Plankton and fish samplings were performed by MB and fish farms staff. Histological analysis of gill samples was carried out by AG. Zooplankton quantification and identification was performed by MB and VT, while phytoplankton screening and identification by CT and SA. MB and GM performed all statistical analysis and figures. MB, VF, and SP wrote the manuscript and all co-authors commented on and provided edits to the original manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We wish to thank Mariló López, M. Carmen Marín and all aquaculture facilities staff for their availability and excellent assistance during samplings. Furthermore, we are especially grateful to Dr. Jennifer E. Purcell and Dr. Jan Marcin Weslawski for critical revisions on the manuscript; Dr. Antonio Canepa and Dr. Emily Baxter for their help in monitoring design and cnidarians identification. This work has received funding from the European Union's projects MED-JELLYRISK (grant n. I-A/1.3/098 - ENPI CBCMED programme), VECTORS (Vectors of Change in Oceans and Seas Marine Life, Impact on Economic Sectors, grant n. 266445, FP7th programme) and CERES (Climate Change and European Aquatic Resources, grant n. 678193, Horizon 2020 programme).
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmars.2017.00240/full#supplementary-material
References
Anderson, M. J. (2001). A new method for non-parametric multivariate analysis of variance. Austral Ecol. 26, 32–46. doi: 10.1111/j.1442-9993.2001.01070.pp.x
Bavestrello, G., Puce, S., Cerrano, C., Zocchi, E., and Boero, N. (2006). The problem of seasonality of benthic hydroids in temperate waters. Chem. Ecol. 22, S197–S205. doi: 10.1080/02757540600670810
Baxter, E. J., Rodger, H. D., McAllen, R., and Doyle, T. K. (2011a). Gill disorders in marine-farmed salmon: investigating the role of hydrozoan jellyfish. Aquac. Environ. Interact. 1, 245–257. doi: 10.3354/aei00024
Baxter, E. J., Sturt, M. M., Ruane, N. M., Doyle, T. K., McAllen, R., and Rodger, H. D. (2012). Biofouling of the hydroid Ectopleura larynx on aquaculture nets in Ireland: Implications for finfish health. Fish Vet. J. 13, 17–29.
Baxter, E. J., Sturt, M. M., Ruane, N. M., Doyle, T. K., McAllen, R., Harman, L., et al. (2011b). Gill damage to Altantic salmon (Salmo salar) caused by the common jellyfish (Aurelia aurita) under experimental challenge. PLoS ONE 6:e18529. doi: 10.1371/journal.pone.0018529
Boero, F., and Fresi, E. (1986). Zonation and evolution of a rocky bottom hydroid community. Mar. Ecol. 7, 123–150. doi: 10.1111/j.1439-0485.1986.tb00152.x
Boero, F., Bouillon, J., Gravili, C., Miglietta, M. P., Parsons, T., and Piraino, S. (2008). Gelatinous plankton: irregularities rule the world (sometimes). Mar. Ecol. Prog. Ser. 356, 299–310. doi: 10.3354/meps07368
Bosch-Belmar, M., Azzurro, E., Pulis, K., Milisenda, G., Fuentes, V., Kéfi-Daly Yahia, O., et al. (2017). Jellyfish blooms perception in Mediterranean finfish aquaculture. Mar. Policy 76, 1–7. doi: 10.1016/j.marpol.2016.11.005
Bosch-Belmar, M., Giomi, F., Rinaldi, A., Mandich, A., Fuentes, V., Mirto, S., et al. (2016a). Concurrent environmental stressors and jellyfish stings impair caged European sea bass (Dicentrarchus labrax) physiological performances. Sci. Rep. 6:27929. doi: 10.1038/srep27929
Bosch-Belmar, M., M'Rabet, C., Dhaouadi, R., Chalghaf, M., Daly Yahia, M. N., Fuentes, V., et al. (2016b). Jellyfish stings trigger gill disorders and increase mortality in farmed Sparus aurata (Linnaeus, 1758) in the Mediterranean Sea. PLoS ONE 11:e0154239. doi: 10.1371/journal.pone.0154239
Bouillon, J., Medel, M. D., Pagès, F., Gili, J.-M., Boero, F., and Gravili, C. (2004). Fauna of the Mediterranean Hydrozoa. Sci. Mar. 68, 5–438. doi: 10.3989/scimar.2004.68s25
Burridge, L. E., Martin, J. L., Lyons, M. C., and LeGresley, M. M. (2010). Lethality of microalgae to farmed Atlantic salmon (Salmo salar). Aquaculture 308, 101–105. doi: 10.1016/j.aquaculture.2010.07.034
Carl, C., Guenther, J., and Sunde, L. M. (2011). Larval release and attachment modes of the hydroid Ectopleura larynx on aquaculture nets in Norway. Aquac. Res. 42, 1056–1060. doi: 10.1111/j.1365-2109.2010.02659.x
Cembella, A. D., Quilliam, M. A., Lewis, N. I., Bauder, A. G., Dell'Aversano, C., Thomas, K., et al. (2002). The toxigenic marine dinoflagellate Alexandrium tamarense as the probable cause of mortality of caged salmon in Nova Scotia. Harmful Algae 1, 313–325. doi: 10.1016/S1568-9883(02)00048-3
Clarke, K. R. (1993). Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 18, 117–143. doi: 10.1111/j.1442-9993.1993.tb00438.x
Doyle, T. K., De Haas, H., Cotton, D., Dorschel, B., Cummins, V., Houghton, J. D. R., et al. (2008). Widespread occurrence of the jellyfish Pelagia noctiluca in Irish coastal and shelf waters. J. Plankton Res. 30, 963–968. doi: 10.1093/plankt/fbn052
Edler, L., and Elbrächter, M. (2010). “The Utermöhl method for quantitative phytoplankton analysis,” in Microscopic and Molecular Methods for Quantitative Phytoplankton Analysis, eds B. Karlson, C. Cusack, and E. Bresnan (Paris: IOC Unesco, Manuals and Guides), 13–20.
EFSA (2009). Scientific opinion of the panel on animal health and welfare on a request from the European commission on welfare aspect of the main systems of stunning and killing of farmed sea bass and sea bream. EFSA J. 1010, 1–12. doi: 10.2903/j.efsa.2009.1010
FAO (2016). The State of World Fisheries and Aquaculture 2016. Rome Available online at: http://www.fao.org/3/a-i3720e.pdf
FIS (2014). Jellyfish Kills Thousands of Salmon in Scottish Farm. Fisheries Information Services Available online at: http://www.fis.com/fis/worldnews/worldnews.asp?monthyear/journalabbrev=12-2014&day=17&id=73474&l=e&country=&special=&ndb=1&df=1 [Accessed May 8, 2017].
Fitridge, I., and Keough, M. J. (2013). Ruinous resident: the hydroid Ectopleura crocea negatively affects suspended culture of the mussel Mytilus galloprovincialis. Biofouling 29, 119–131. doi: 10.1080/08927014.2012.752465
Fosså, J., Flood, P., Olsen, A., and Jensen, F. (2003). Småog usynlige, men plagsomme maneter av arten Muggiaea atlantica (Small and invisible, but troublesome jellyfish of the species Muggiaea Atlantica). Fisk. og Havet. 2, 99–103.
González-Duarte, M. M., Megina, C., De Vito, D., Di Camillo, D. G., Puce, S., and Piraino, S. (2016). A unified assessment of marine Mediterranean assemblages: a lesson from benthic hydroids. Mar. Ecol. 37, 155–163. doi: 10.1111/maec.12271
González-Duarte, M. M., Megina, C., Piraino, S., and Cervera, J. L. (2013). Hydroid assemblages across the Atlantic-Mediterranean boundary: is the Strait of Gibraltar a marine ecotone? Mar. Ecol. 34, 33–40. doi: 10.1111/maec.12022
Guenther, J., Misimi, E., and Sunde, L. M. (2010). The development of biofouling, particularly the hydroid Ectopleura larynx, on commercial salmon cage nets in Mid-Norway. Aquaculture 300, 120–127. doi: 10.1016/j.aquaculture.2010.01.005
Kitatani, R., Yamada, M., Kamio, M., and Nagai, H. (2015). Length is associated with pain: Jellyfish with painful sting have longer nematocyst tubules than harmless jellyfish. PLoS ONE 10:e0135015. doi: 10.1371/journal.pone.0135015
Lund, J. W. G., and Talling, J. F. (1957). Botanical limnological methods with special reference to the algae. Bot. Rev. 23, 489–583. doi: 10.1007/BF02870144
Marcos-López, M., Mitchell, S. O., and Rodger, H. D. (2016). Pathology and mortality associated with the mauve stinger jellyfish Pelagia noctiluca in farmed Atlantic salmon Salmo salar L. J. Fish Dis. 39, 111–115. doi: 10.1111/jfd.12267
Milisenda, G., Martinez-Quintana, A., Fuentes, V. L., Bosch-Belmar, M., Aglieri, G., Boero, F., et al. (in press). Reproductive bloom patterns of Pelagia noctiluca in the Strait of Messina, Italy. Estuar. Coast. Shelf Sci. doi: 10.1016/j.ecss.2016.01.002
Mitchell, S. O., Baxter, E. J., Holland, C., and Rodger, H. D. (2012). Development of a novel histopathological gill scoring protocol for assessment of gill health during a longitudinal study in marine-farmed Atlantic salmon (Salmo salar). Aquac. Int. 20, 813–825. doi: 10.1007/s10499-012-9504-x
Navarro, G., Vzquez, Á., MacÍas, D., Bruno, M., and Ruiz, J. (2011). Understanding the patterns of biological response to physical forcing in the Alborn Sea (western Mediterranean). Geophys. Res. Lett. 38, 1–6. doi: 10.1029/2011GL049708
Oksanen, J., Kindt, R., Legendre, P., and O'Hara, R. (2005). Vegan: Community Ecology Package. Available online at: http://cran.r-project.org
Palma, S., Apablaza, P., and Silva, N. (2007). Hydromedusae (Cnidaria) of the Chilean southern channels (from the Corcovado Gulf to the Pulluche-Chacabuco Channels). Sci. Mar. 71, 65–74. doi: 10.3989/scimar.2007.71n165
Panagiotis, E. S., Aristeidis, T., Maria, A. C., Alkisti, B., Stelios, G. D., and Helen, M. (2014). Comparison of the effects of six stunning/killing procedures on flesh quality of sea bass (Dicentrarchus labrax, Linnaeus 1758) and evaluation of clove oil anaesthesia followed by chilling on ice/water slurry for potential implementation in aquaculture. Aquac. Res. 45, 1759–1770. doi: 10.1111/are.12120
Piraino, S., Fanelli, G., and Boero, F. (2002). Variability of species' roles in marine communities: change of paradigms for conservation priorities. Mar. Biol. 140, 1067–1074. doi: 10.1007/s00227-001-0769-2
Purcell, J. (1997). Pelagic cnidarians and ctenophores as predators: selective predation, feeding rates, and effects on prey populations. Ann. Inst. Oceanogr. 73, 125–137.
Purcell, J. E., Baxter, E. J., and Fuentes, V. L. (2013). “Jellyfish as products and problems of aquaculture,” in Advances in Aquaculture Hatchery Technology, eds G. Allan and G. Burnell (Oxford: Woodhead Publishing), 404–430.
Rensel, J. E. (1992). Harmful Effects of the Marine Diatom Chaetoceros Concavicornis on Atlantic salmon (Salmo salar). Ph.D. dissertation, University of Washington, Seattle.
Rodger, H. D. (2007). Gill disorders: an emerging problem for farmed Atlantic salmon (Salmo salar) in the marine environment? Fish Vet. J. 9, 38–48.
Rodger, H. D., Henry, L., and Mitchell, S. O. (2011a). Non-infectious gill disorders of marine salmonid fish. Rev. Fish Biol Fish. 21, 423–440. doi: 10.1007/s11160-010-9182-6
Rodger, H. D., Murphy, K., Mitchell, S. O., and Henry, L. (2011b). Gill disease in marine farmed Atlantic salmon at four farms in Ireland. Vet. Rec. 1, 1–4. doi: 10.1136/vr.d3020
Sanchez-Vidal, A., Calafat, A., Canals, M., and Fabres, J. (2004). Particle fluxes in the Almeria-Oran Front: control by coastal upwelling and sea surface circulation. J. Mar. Syst. 52, 89–106. doi: 10.1016/j.jmarsys.2004.01.010
Sarhan, T., García Lafuente, J., Vargas, M., Vargas, J. M., and Plaza, F. (2000). Upwelling mechanisms in the northwestern Alboran Sea. J. Mar. Syst. 23, 317–331. doi: 10.1016/S0924-7963(99)00068-8
Smayda, T. J. (2006). Scottish Executive Environment Group Harmful Algal Bloom Communities in Scottish Coastal Waters : Relationship to Fish Farming and Regional Comparisons - A Review. Report, University of Rhode Island, Kingston.
Thibault, D., Gaudy, R., and Le Fèvre, J. (1994). Zooplankton biomass, feeding and metabolism in a geostrophic frontal area (Almeria-Oran Front, western Mediterranean). Significance to pelagic food webs. J. Mar. Syst. 5, 297–311. doi: 10.1016/0924-7963(94)90052-3
Treasurer, J. W., Hannah, F., and Cox, D. (2003). Impact of a phytoplankton bloom on mortalities and feeding response of farmed Atlantic salmon, Salmo salar, in west Scotland. Aquaculture 218, 103–113. doi: 10.1016/S0044-8486(02)00516-1
Willcox, S., Moltschaniwskyj, N., and Crawford, C. (2008). Population dynamics of natural colonies of Aurelia sp. scyphistomae in Tasmania, Australia. Mar. Biol. 154, 661–670. doi: 10.1007/s00227-008-0959-2
Keywords: gelatinous zooplankton, marine aquaculture, European sea bass, gill disorders, southwestern Mediterranean
Citation: Bosch-Belmar M, Milisenda G, Girons A, Taurisano V, Accoroni S, Totti C, Piraino S and Fuentes V (2017) Consequences of Stinging Plankton Blooms on Finfish Mariculture in the Mediterranean Sea. Front. Mar. Sci. 4:240. doi: 10.3389/fmars.2017.00240
Received: 14 May 2017; Accepted: 18 July 2017;
Published: 02 August 2017.
Edited by:
Cosimo Solidoro, National Institute of Oceanography and Experimental Geophysics, ItalyReviewed by:
Jennifer Estelle Purcell, Western Washington University, United StatesJan Marcin Weslawski, Institute of Oceanology (PAN), Poland
Copyright © 2017 Bosch-Belmar, Milisenda, Girons, Taurisano, Accoroni, Totti, Piraino and Fuentes. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mar Bosch-Belmar, Ym9zY2gtYmVsbWFyQGNvbmlzbWEuaXQ=
Stefano Piraino, c3RlZmFuby5waXJhaW5vQHVuaXNhbGVudG8uaXQ=
†Senior Authors.