 Claire Daisy Bonneville
Claire Daisy Bonneville Solène Derville
Solène Derville Jolanda A. Luksenburg
Jolanda A. Luksenburg Marc Oremus5
Marc Oremus5 Claire Garrigue
Claire Garrigue- 1UMR 250/9220 ENTROPIE, IRD, Université de La Réunion, Université de la Nouvelle-Calédonie, CNRS, Ifremer, Laboratoire d’Excellence-CORAIL, BPA5, Nouméa, New Caledonia
- 2Opération Cétacés, Nouméa, New Caledonia
- 3Institute of Environmental Sciences, Leiden University, Leiden, Netherlands
- 4Department of Environmental Science and Policy, George Mason University, Fairfax, VA, United States
- 5WWF-France Antenne Nouvelle-Calédonie, Nouméa, New Caledonia
Understanding population structure and habitat use of poorly known cetacean species is a first step toward scientifically informed management decisions. In the southern range of New Caledonia (South Pacific), a long-term dataset of Indo-Pacific bottlenose dolphins (Tursiops aduncus) encounters primarily during winter seasons 1997 to 2019 (473 group observations) was used to assess social structure, habitat use and potential threats. A total of 338 individuals were photographically identified, forming three distinct communities in the south-west lagoon, the south lagoon and the Isle of Pines. Mark-recapture histories revealed that the three communities were weakly connected and might be considered as independent management units. Suitable habitats were estimated with presence-only distribution models relative to topographic and seabed substrate predictors. Habitat suitability increased with proximity to coasts or reefs, at shallow depth, and over muddy bottom. These habitats had various levels of protection and were used by humans, mostly in the south-west lagoon. External injuries were interpreted to determine natural interactions and potential anthropogenic threats. The prevalence in injuries did not vary among the three areas. A substantial proportion of injuries related to propeller hits was reported, representing a total of 16.7% (34 of 204) of all injuries observed on dolphins. The three communities of Indo-Pacific bottlenose dolphins revealed in the southern part of New Caledonia are particularly vulnerable due to their insularity, their coastal habitat use and the low levels of connectivity found among them. In a context of increasing maritime traffic, fishing and recreational activities, this study provides a useful baseline to the urgent assessment of the conservation status of dolphins in New Caledonia.
Introduction
Connectivity is known to positively influence populations by favoring gene flow (Wright, 1943). In the absence of connectivity, insularity may result in higher susceptibility of small populations to inbreeding (Charlesworth and Charlesworth, 1987) and ultimately increase the risk of local species extirpation, especially when these populations face threats from human activities. Non-migratory insular species such as lagoon-living cetaceans are particularly at risk as their gene flow may only be ensured by punctual and rare immigration events (Oremus et al., 2015).
The Indo-Pacific bottlenose dolphin (Tursiops aduncus, Ehrenberg 1833) is one of the most frequently observed cetacean species in the Indian and Pacific Ocean coastal waters. Three phylogenetic units specific to African waters, to the Pacific Ocean and to the Arabian Sea have been highlighted within the species (Hale et al., 2000; Oremus et al., 2015; Amaral et al., 2017; Gray et al., 2018). They are characterized by local ecological adaptations, as well as reproductive isolation and minimal or absent gene flow. Mean ranges of Indo-Pacific bottlenose dolphins vary among regions or islands (Chilvers, 2001; Kiszka et al., 2012) and can rise up to 150 km2 (Sprogis et al., 2016). Despite their capacity to travel long distances, Indo-Pacific bottlenose dolphins may be genetically structured at a fine scale leading to the separation of distinct populations (Ansmann et al., 2012; Manlik et al., 2019). In particular cases, regional adaptations to environmental parameters can in turn lead to morphological differences between populations (van Aswegen et al., 2019).
Generally, this species occupies both continental shelf waters and oceanic island waters (Wang and Yang, 2009; Braulik et al., 2019), using a diversity of habitats such as rocky and coral reef areas, sandy bottoms and seagrass beds (Sprogis et al., 2018a). In some regions, the Indo-Pacific bottlenose dolphin shows a marked preference for waters with shallow depth, flat seabed, and in close proximity to the coast, suggesting that these environmental conditions may be important predictors of their spatial distribution (Ross et al., 1987; Gross et al., 2009; Kiszka et al., 2012). These parameters directly impact the abundance of preys (Torres et al., 2008; Torres and Read, 2009; Sprogis et al., 2016), and that of predators (Heithaus and Dill, 2006; Sprogis et al., 2018b), thus influencing habitat use patterns of dolphins. In turn, spatial distribution results in varying levels of exposure to anthropogenic threats.
Bycatch in fishing gear probably represents the most important threat to small cetaceans (Kiszka et al., 2008; Mansur et al., 2012) and has considerably increased over recent decades (DeMaster et al., 2001). In addition, collisions with boats may also constitute an important threat to dolphins (Van Waerebeek et al., 2008; Avila et al., 2018). Due to their coastal habitats, Indo-Pacific bottlenose dolphins are particularly at risk, many of them showing scars from nets or propellers (Kiszka et al., 2008; Mansur et al., 2012). In 2019, the conservation status set by the IUCN Red List was upgraded from “Data Deficient” to “Near Threatened” (Braulik et al., 2019), considering the small and coastal range of the species and the vulnerability to human threats such as entanglement in fishing gear, or habitat loss and degradation. However, data are still insufficient in many regions (Cottarel et al., 2016) to estimate the diversity of threats and their current impact on dolphins at a local scale, especially in tropical lagoon remote islands.
New Caledonia (South Pacific) is the eastern limit of the Indo-Pacific bottlenose dolphin’s range (Braulik et al., 2019). The species is encountered around the main island of New Caledonia “Grande Terre” and into the western coasts of the Isle of Pines. The species belongs to the Pacific Ocean phylogenetic unit and only two mitochondrial DNA haplotypes (364 bp control region) have been detected among 88 individuals sampled in different parts of the archipelago (Oremus et al., 2015). This low level of genetic diversity suggests that the species colonized the region a few thousand years ago and then became isolated by distance (Oremus et al., 2015). Considering its geographic isolation, the eastern limit to the species’ range and low genetic diversity, there is an urgent need to evaluate the current status of the Indo-Pacific bottlenose dolphin in New Caledonia. Moreover, separate communities have been detected through photo-identification and genetic sampling all around New Caledonia (Oremus et al., 2009), thus highlighting the need to separately estimate vulnerability of subpopulations at very small scale.
In the southern range of New Caledonia, maritime traffic has increased significantly since 1995, thus influencing the potential risk of negative interaction with marine mammals in this region (Bourgogne et al., 2018). In this study, we first used photo-identification to test for demographic structuring within the study area, using social community analysis. Second, habitat use was assessed in order to identify priority conservation areas where dolphin presence may conflict with coastal anthropogenic activities. Finally, injuries found on the dorsal fin or on the body of identified individuals were described and interpreted as indicators of anthropogenic threats (Luksenburg, 2014). This study constitutes a first step toward conservation of a poorly-known species to ensure its protection in a remote island of the South Pacific.
Materials and Methods
Study Area
New Caledonia is an archipelago located in the southwest Pacific Ocean east of Australia (21°30′S 165°30′E, Figure 1). The main island, the Grande Terre, is about 400 km long and 50–80 km wide and is surrounded by a large lagoon delimited by 1,600 km of barrier reef considered as one of the world’s largest lagoons. The study area is located at the southeastern tip of the Grande Terre (Figure 1). It includes the coastal waters surrounding Nouméa, the capital of the island, which is the most populated and industrialized city of New Caledonia. The southern part of the lagoon includes the less populated Ouen Island, the Prony Bay and a wide Integral Marine Protected Area (IMPA), the Merlet reserve (IUCN category 1a, 17,000 ha). The Isle of Pines is separated from the Grande Terre by the Sarcelle pass. Since 2008, a large part of the provincial Park “Great South Lagoon” is a UNESCO World Heritage Zone (UNESCO, 2009).
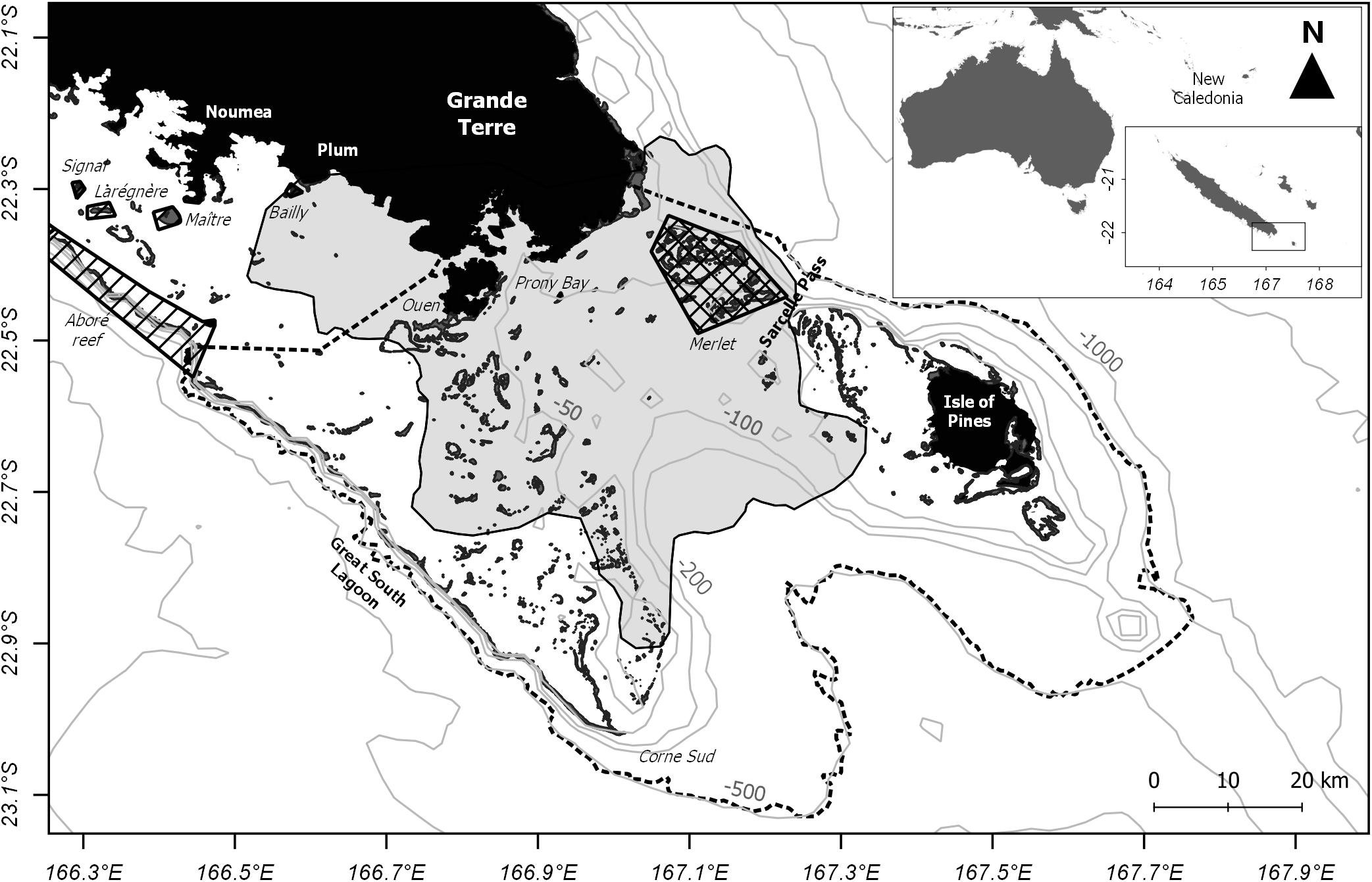
Figure 1. Map of the study area in the southern range of New Caledonia (South Pacific). Land and shallow reefs are shown in black. Winter study area with recorded effort during humpback whale season is shown in light gray. Delimitation of the provincial Park “Great South Lagoon” is shown with a dotted line. Polygons represent Integral Marine Protected Areas (gridded) or natural reserves with restrictions (hashed). Light gray lines represent 50–1000 m isobaths.
Data Collection
Dolphin data were acquired at sea from 1997 to 2019 (Supplementary Table 1). The great majority of dolphin data was collected in the south lagoon during boat-based surveys aimed at monitoring humpback whales in austral winter, from July to September (Garrigue et al., 2001). These surveys were conducted from a 6-m rigid-hulled inflatable boat in Beaufort sea states ≤3 and survey effort was recorded (as shown in Figure 1, adapted from Derville et al., 2019). Other sightings (16%) were collected outside this season.
For each sighting of a Indo-Pacific bottlenose dolphin group, basic information was collected including the GPS position at detection of the group and the number of individuals. When possible, a focal follow was initiated during which photographs of the right and/or left sides of the dorsal fins were taken. We defined a group as an association of individuals moving in the same direction, engaged in similar behavioral activities, or interacting with each other within a radius of 100 m (McHugh et al., 2011). Each group was either simply “recorded,” when only basic information was reported or “studied,” when individuals were photographed for the purpose of identification.
Photo-Identification
Individuals were identified using photo-identification (Würsig and Jefferson, 1990). For each individual dolphin photographed in a group, the best photograph of the right and/or left sides of the dorsal fin was selected and used for photo-identification. Photographs were selected based on two criteria: the distinctiveness of the dorsal fin (for recognition of unique individuals) and the quality of the photograph (for judging the presence and nature of any injuries). The level of recognition (I) was based on the analysis of the marks on the dorsal fin and ranked from 1 to 4. A high value corresponded to the presence of significant marks or scars allowing an easy distinction of individuals. The quality of the photograph (Q) depended on four criteria, each ranked from 1 to 5: the sharpness, the exposure, the orientation and the percentage of the dorsal fin visible in the photograph. The overall quality Q of the photograph corresponded to their average. A high value represented a dorsal fin picture with excellent quality. Only photographs with I ≥ 2 and Q ≥ 2 were used in this study. The selected photographs were then compared using the open-source R package “finFindR” (Thompson et al., 2019). This software is capable of isolating fins in photographs, tracing the trailing edge of the fin, and producing a sorted list of likely identities from a reference catalog of known individuals. Here, the reference catalog was built by comparing trailing edges of dorsal fins from all dolphins photographed in New Caledonia. When a dorsal fin was not found with finFindR, another comparison was conducted manually to avoid potential recognition errors of the software. Once the lack of match with the catalog was cross-checked, the dorsal fin corresponding to a new unique individual was added to the reference catalog.
Connectivity
Social interactions between individuals observed at least twice(Bejder et al., 1998) were investigated using SOCPROG 2.9 (Whitehead, 2009). Individuals observed in the same group were considered associated (Whitehead, 2009). The half-weighted association index (HWI, Cairns and Schwager, 1987) was used, as it accounts for incomplete photo-identification of individuals within groups (Parra et al., 2011; Smith et al., 2016; Baker et al., 2018; Genov et al., 2019). A social network was built from an association matrix using NetDraw 2.158 (Borgatti, 2002) to assess social communities. In order to estimate if individuals associate at random or if they show preferred or non-preferred associations, the permutation test developed by Bejder et al. (1998) and modified by Whitehead (2009) was applied. This test is based on the standard deviation of the mean HWI and permutation is set on individuals among groups inside each sampling period. If individuals do not associate at random, the standard deviation of the observed mean HWI is significantly higher than that of the random mean HWI (p < 0.05; Gowans et al., 2007; Whitehead, 2009). Association matrices were randomly permuted 20,000 times.
Habitat Modeling
In order to characterize habitats of the Indo-Pacific bottlenose dolphins, a series of environmental variables were acquired, based on preferences identified in other regions of the world: distance to the coast, distance to the reefs, water column depth, seabed slope and sedimentology (Torres et al., 2008; Gross et al., 2009; Sprogis et al., 2018a; Vargas-Fonseca et al., 2018). The distance to the coast (DIS_COAST in meters) corresponded to the distance to the nearest land (excluding the islets), considering only the “Grande Terre,” the Ouen Island and the Isle of Pines (Figure 1). Alternative descriptors of reef habitats were derived from geomorphological maps provided by Andréfouët et al. (2008): the distance to all reefs except for the fringing reefs (DIS_REEF in meters), the distance to the barrier reef only (DIS_BAR in meters) and the distance to the intermediate reef patches only (DIS_PATCH in meters). The fringing reefs that surround the coastline were excluded from this analysis because their effect cannot be differentiated from that of the coast. Bathymetry (DEPTH in meters) was provided by the DTSI (2016) at a resolution of 100 × 100 m. The non-hydrographed areas were subsequently filled through extrapolation of satellite and aerial composite imagery (J. Lefèvre, IRD, pers. com.). The slope (SLOPE in°) was derived from bathymetric charts. Finally, the sedimentology was acquired from maps produced by P. Douillet (IRD, pers. com.) according to the method developed by Chevillotte et al. (2005). The percentage of sedimentary muddy clay (SED in%) in the bottom sediments was spatially interpolated from surveys obtained after (Debenay, 1987) and (Chardy et al., 1988).
To model dolphin habitat use, we accounted for the spatial heterogeneity of research effort in the southern range of New Caledonia by using the so-called “target” method (Phillips et al., 2009) that approximates the research effort at sea. According to this method, the points of presence of “the species of interest” (i.e., Indo-Pacific bottlenose dolphins) were compared to the control points of another species observed with the same spatially biased research effort, called the “target species” (i.e., humpback whales). As the great majority of Indo-Pacific bottlenose dolphin sightings were made during winter surveys aimed at studying humpback whales, the “target” method was particularly suitable to our dataset. However, in this approach calibration of the models was restricted to Indo-Pacific bottlenose dolphin observations made during such surveys.
Generalized additive models (GAM; Hastie and Tibshirani, 1990) were used to model and predict the distribution of suitable Indo-Pacific bottlenose habitat in the study area. This approach models the non-linear relationships between a binomial variable (presence and control points) and a series of explanatory variables. The probability of presence of a group of dolphins was modeled according to environmental variables, as well as a smooth of location (latitude/longitude projected in a UTM58S coordinate system). This spatial variable was included in the model using a Gaussian process (Wood, 2017), which allows to model variations in probability of presence on a very fine scale in a two-dimensional field. The Gaussian process relied on a Matern correlation function with kappa equal to 1.5. The range parameter of the Gaussian process was set to 10 km. Three types of models were tested: a spatial model including only a smooth of location (M1), environmental models including only environmental variables (M2.1, M2.2, M2.3, and M2.4) and a combined model including both a smooth of location and environmental variables (M3.4, see Supplementary Table 2). Including a smooth of location is a common approach to approximate other unavailable variables such as social drivers of distribution. Environmental models and spatial models may both provide valuable insights. Comparing the predictions from both types of models may help reveal suitable areas not currently used by a species or that have been subject to less survey effort. Hence, the combined model Menv–spa was derived from the best environmental model Menv and they were both used to produce predicted maps of habitat suitability.
Cross-correlation among explanatory variables was checked beforehand, to ensure that Spearman correlation coefficients between pairs of variables would be below 0.7. The effect of DIS_COAST and either DIS_PATCH or DIS_REEF were modeled separately or as an interaction term, assuming that dolphins may show variable and complex habitat use patterns with respect to these geomorphic characteristics of the lagoon environments. Indeed, dolphins were hypothesized to regroup in shallow waters either in proximity to the coast or in proximity to intermediate reef patches. Variables were modeled with penalized thin plate regression splines optimized with a Restricted Maximum Likelihood and limited basis size to prevent overfitting (Wood, 2017). The basis size parameter k was limited to 3 for environmental variables and to 50 for the two-dimension smooth of location. Variable selection across environmental models was conducted with a shrinkage approach which adds an extra penalty to each smoother and penalizes them non-significant variables to zero (Marra and Wood, 2011). In addition, the Akaike Information Criterion (AIC, Burnham and Anderson, 2004) and the percentage of deviance explained by the models was used to select between alternative environmental models (with or without interactions, with DIS_PATCH or with DIS_REEF) and compare them to a null model (M0, see Supplementary Table 2; Friedman, 2001). Finally, Moran’s I was computed in the residuals of the selected models Menv and Menv–spa to check for the absence of spatial autocorrelation (Dormann et al., 2007), using a Monte-Carlo simulation with 1000 permutations.
All statistical analyses were performed using the open-source program R (RCoreTeam, 2020), specifically with the mgcv R package (version 1.8–31). Map illustrations were produced with QGIS 3.4.0 (QGIS Development Team, 2018).
External Injury Interpretation
Photographs of all recorded individuals were examined for external injuries on the dorsal fin and body. All marks (wounds, scars, and nicks) on the leading edge of the dorsal fin were considered as an injury (Luksenburg, 2014). Marks on the trailing edge of the dorsal fin were considered as an injury if the mark was deeper than 5% of the dorsal fin width (Luksenburg, 2014). Injuries were classified in one of the ten following categories: (a) linear severed dorsal fin, (b) non-linear severed dorsal fin, (c) straight, deep cut, (d) opposing cuts, (e) parallel cuts, (f) collapse, (g) obtuse, short, cut-like indentation, (h) indentation, (i) round cut, (j) crescent shaped cut (shark inflicted bite wound in Luksenburg, 2014), and (k) cookie-cutter shark bite wound. A new category was added to this classification: (l) wide v-shaped cut. This wound is characterized by a wide v-shaped cut which is more wide than deep (see Supplementary Figure 1 for description and examples). Individuals may bear multiple injuries on different parts of their body, which may have been attributed to different categories. The origin of each injury was assessed on a case-by-case basis and was classified as natural, human-related or unidentified origin. Categories (b), (i), (k) may be attributed to natural origin, (a), (d), (e) to human-related origin and (c), (f), (g), (h), (j), (l) to natural or human-related origin. Natural injuries were assigned to intra/inter-specific interactions with unidentified species, or to shark attacks when the crescent shape and/or teeth marks were evident. Human-related injuries were assigned to propeller hits or fishing gear when possible. The Pearson’s χ2 homogeneity test was used to estimate whether the percentages differed significantly according to social communities identified in the study area.
Results
Individual Identification and Social Connectivity
In total, 397 groups of Indo-Pacific bottlenose dolphins were recorded during 812 survey days (associated with humpback whale survey effort) mostly focused in the south lagoon in austral winter (Figure 1). Among these 397 groups, 160 were studied. Other sightings completed the dataset with 76 groups recorded and 45 studied. In total, 473 groups were recorded, among which 205 were studied.
The number of groups recorded, groups studied and photographs allowing individual identification are the highest in the south lagoon (Table 1). From the 1,567 photographs that passed the quality control (I ≥ 2 and Q ≥ 2), 338 unique individuals were identified. A total of 64 unique dolphins were photo-identified in the south-west lagoon, 240 in the south lagoon and 39 around the Isle of Pines. Recapture rate varied between the three areas (χ2 = 49.8, p < 0.001) and was the highest in the south lagoon (73%), followed by the Isle of Pines (41%) and the south-west lagoon (24%).
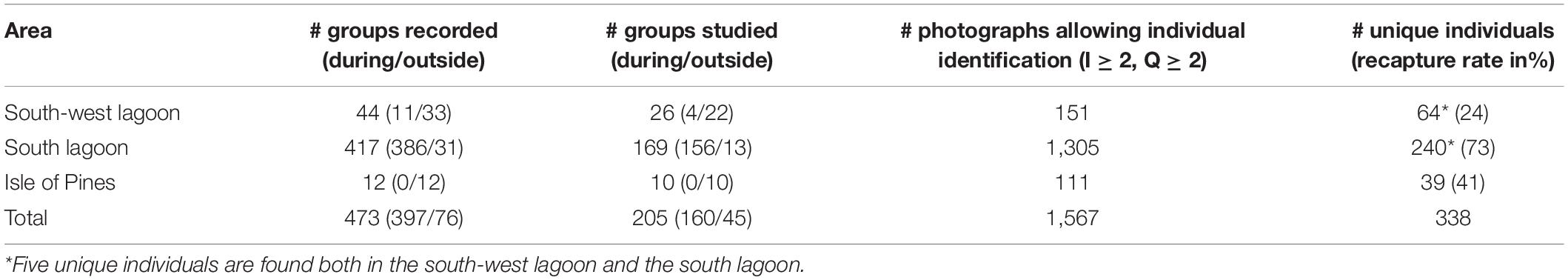
Table 1. Number of groups recorded (during/outside the humpback whale winter season), groups studied (during/outside of the humpback whale winter season), photographs allowing individual identification and unique individuals photo-identified in the south-west lagoon, the south lagoon and the Isle of Pines.
Given that few individuals were resighted in the south-west lagoon and in the Isle of Pines (Supplementary Table 3), social network analysis was performed over 185 individuals sighted at least twice. The standard deviation of the observed mean HWI was significantly higher than that of the randomly generated data (observed mean HWI = 0.0393 ± 0.1097; random mean HWI = 0.0392 ± 0.1079; p < 0.001). This statistic indicated that individuals tend to have preferred associations.
Results highlighted the presence of three communities of dolphins in the study area (Figure 2). The Isle of Pines community (IoPc: red) did not show any connection with the other two communities. Within the IoPc, a total of 16 individuals were identified and only observed around the Isle of Pines. Within the south community (Sc: blue), 157 individuals were identified and observed in an area delimited by Ouen Island to the west and Merlet IMPA to the east. Finally, 11 individuals were identified in the south-west community (SWc: yellow) and observed around Nouméa and Plum. One individual was socially linked to individuals from both Sc and SWc (green, Figure 2). Out of the four observations of this dolphin (Supplementary Figure 2), two were in interaction with individuals from SWc, one with individuals from Sc and one with an individual from an unknown community (i.e., never recaptured). A small number of individuals (n = 4) belonging to the Sc was occasionally observed west of Ouen Island or around Plum, but never in interaction with individuals belonging to SWc (Supplementary Figure 2).
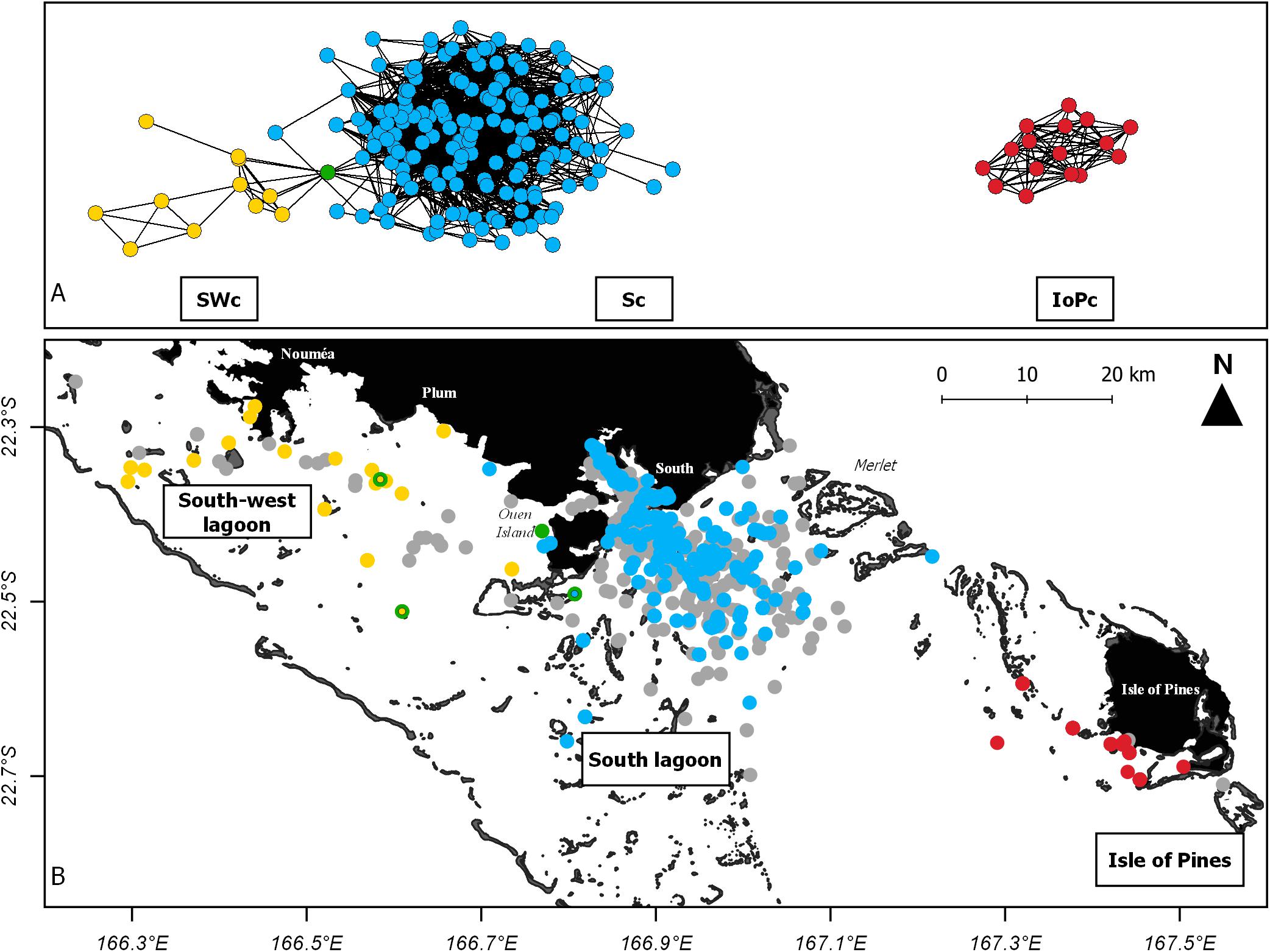
Figure 2. Population structure of Indo-Pacific bottlenose dolphins photo-identified in the southern range of New Caledonia between 1997 and 2019 (winter seasons mostly). (A) Social network of interactions among individuals between three communities: the south-west community (SWc in yellow), the south community (Sc in blue) and the Isle of Pines community (IoPc in red). The green point represents an individual interacting with other dolphins from two different communities (SWc and Sc). (B) Group positions where individuals were observed. Three out of four positions of the green point represents interactions with SWc (yellow center) and Sc (blue center). Gray points represent groups recorded that have not been studied or those containing individuals observed only once.
Habitat Suitability and Use
Binomial statistical models were used to determine the environmental variables likely to predict the distribution of the groups of Indo-Pacific bottlenose dolphins in austral winter. Only the observations of dolphins made during humpback whale-dedicated surveys were taken into account for this analysis (n = 397), with 2.8% observed in the south-west lagoon, 97.2% in the south lagoon and none in the Isle of Pines (Supplementary Figure 3). In comparison, 1,560 humpback whale group positions were used as background points in the binomial models. Except for DIS_REEF and DIS_PATCH that were highly correlated (Spearman coefficient = 0.98), all other variables were related with Spearman coefficients below 0.7, with the strongest correlation being found between DEPTH and DIS_COAST (Spearman coefficient = −0.67). Residuals from the models Menv and Menv–spa showed no significant pattern of spatial autocorrelation (Moran’s I, Menv: statistic = −0.0005, p = 0.352; Menv–spa: statistic = −0.0007, p-value = 0.818).
In the best environmental model Menv (Table 2), distance to the coast and intermediate reef patches (edf = 8.6, χ2 = 32.0, p < 0.001), distance to barrier reef (edf = 1.3, χ2 = 4.7, p = 0.025), bathymetry (edf = 1.1, χ2 = 80.3, p < 0.001) and percentage of sedimentary muddy clay (edf = 0.9, χ2 = 7.6, p = 0.001) significantly influenced the presence of dolphins in austral winter (12.7% explained deviance, Supplementary Table 2, Supplementary Figure 4 and Table 2). Based on these environmental variables, the Menv model predicted suitable habitats in the main south lagoon region, but also suggested potentially suitable habitats in the south-west lagoon and the Isle of Pines, even though these areas had been subject to less survey effort (Figure 3A). In the south-west lagoon, the waters most suitable for the presence of Indo-Pacific bottlenose dolphins were predicted in the coastal shallow waters around Nouméa and Plum, the reefs south of Plum and south-west of Ouen Island. In the Isle of Pines, the waters most suitable were located all around the island, close to the coast.
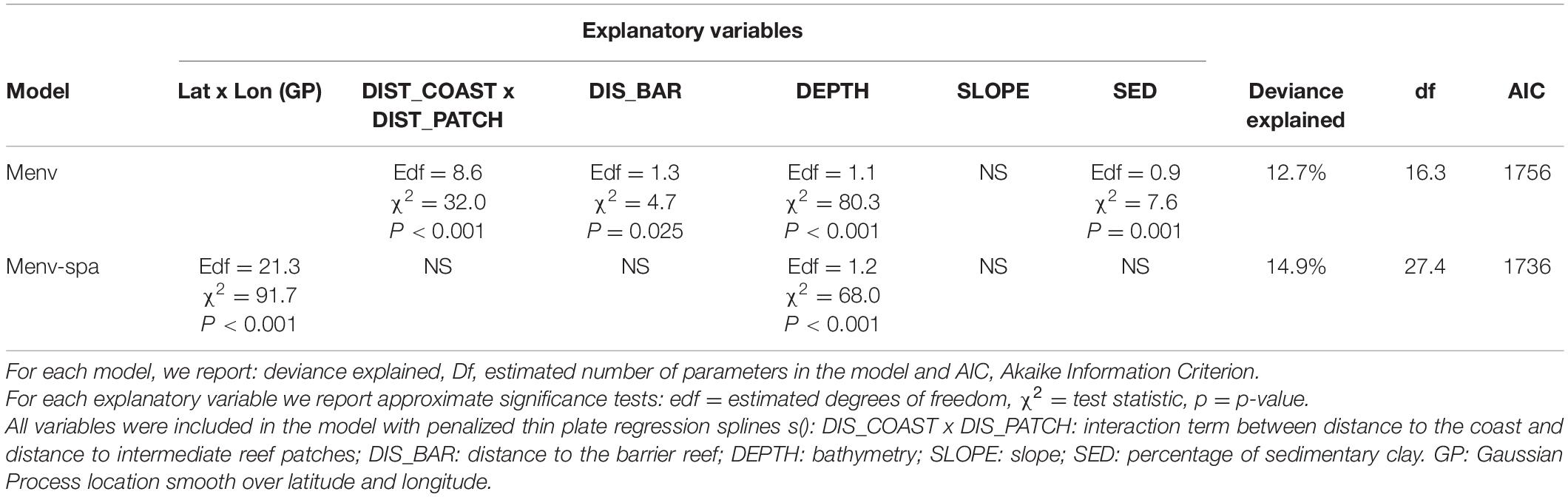
Table 2. Summary of the two generalized additive models selected to estimate dolphin habitat use: Menv (best environmental model) and Menv–spa (equivalent combined model with environmental and spatial predictors).
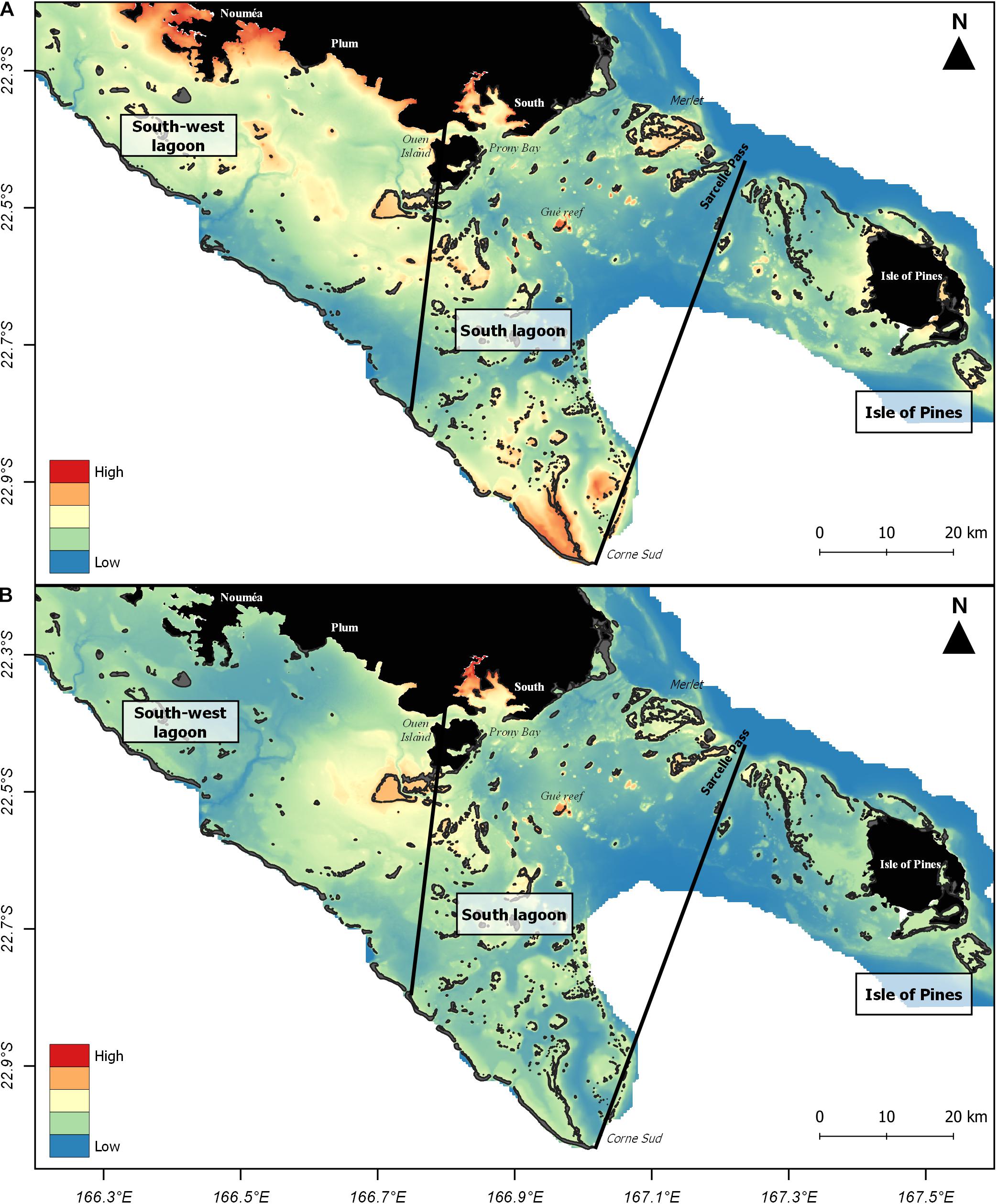
Figure 3. Predicted winter habitat suitability for Indo-Pacific bottlenose dolphins in the southern range of New Caledonia. (A) Environmental model Menv; (B) Combined environmental-spatial model Menv–spa. Approximate boundaries to the south-west lagoon, the south lagoon and the Isle of Pines are represented with black lines. Predicted habitat suitability is represented on a colored scale from high suitability (red) to low suitability (blue). Land and shallow reefs are shown in black.
In the equivalent combined model with environmental and spatial predictors Menv–spa, only the bathymetry (edf = 1.2, χ2 = 68.0, p < 0.001) and the smooth of location (edf = 21.3, χ2 = 91.7, p < 0.001) significantly influenced the presence of dolphins in austral winter (14.9% explained deviance, Supplementary Table 2, Supplementary Figure 5 and Table 2). Due to the effect of the smooth of location, this model primarily predicted suitable habitats in the south lagoon (Figure 3B) where most dolphin observations were concentrated. Based on this model, the waters most suitable to the presence of dolphins were the shallowest (0–50 m) located inside the Prony Bay and around reefs, particularly the Gué reef and the reef plateaus south of Ouen Island. The southeastern waters of Merlet IMPA also appeared to be suitable.
Levels and Origins of Injuries
Injuries (n = 204) were observed in the photographs of 131 out of 338 uniquely identified dolphins (38.8%, Table 3). At least 25% of the upper body was visible in 20.6% of cases and this proportion did not vary between the three geographical areas (χ2 = 4.5, p > 0.05). The proportion of animals with injuries did not vary geographically either (χ2 = 2.2, p > 0.05). Most of the injuries were observed on the dorsal fin only (82.4%). The most frequently observed injuries were round and oval cuts (i, 32.8%), followed by straight, deep cuts (c, 23.5%) and by wide v-shaped cuts (l, 13.7%).
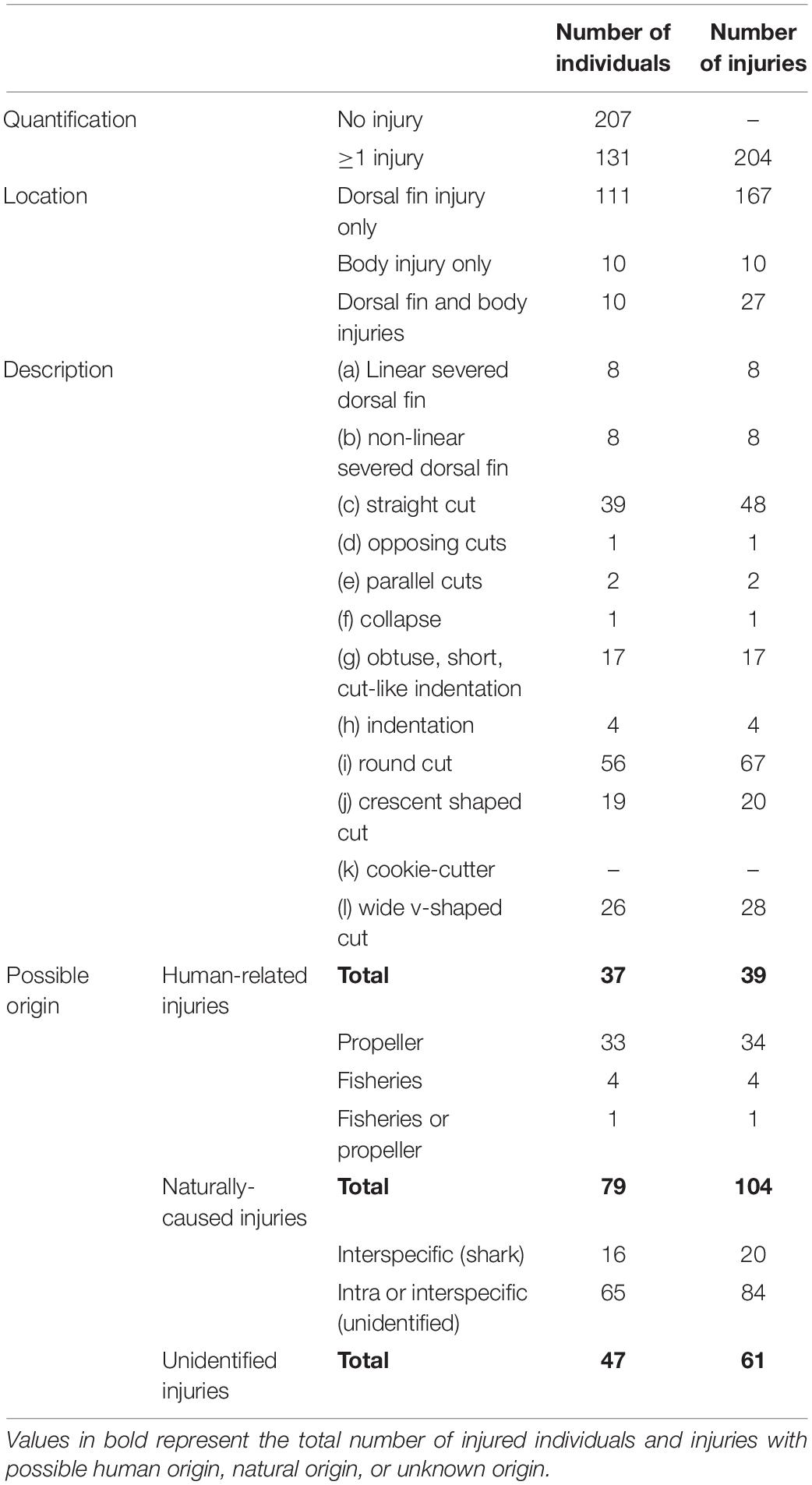
Table 3. Quantification of external injuries following (Luksenburg, 2014) in Indo-Pacific bottlenose dolphins photo-identified in the southern range of New Caledonia between 1997 and 2019.
Among the dolphins with injuries, 28.2% (37 of 131) showed scars of probable human-related origin (Table 3 and Figure 4). Among them, a great majority of injuries was likely related to propeller hits (87.2% – 34 of 39 – representing 16.7% – 34 of 204 – of overall injuries, Figures 4.1A–C) and to fisheries interaction (10.3% – 4 of 39 – representing 2.0% – 4 of 204 – of overall injuries) (Figure 4.1D). In one case, the origin of the human-related injuries could not be specified (either fisheries or propeller).

Figure 4. Examples of injuries observed on Indo-Pacific bottlenose dolphins in the southern range of New Caledonia. Injuries were probably from human-related origin characterized by (1.A): straight and parallel deep cuts on the trailing edge generally caused by a propeller hit. (1.B): linearly severed top of dorsal fin generally caused by a propeller hit. (1.C): straight deep cut on the leading edge likely caused by a propeller hit. (1.D): opposite cuts on the tops of the dorsal fins generally caused by fishing gear interaction. Natural origin characterized by (2.E): v-shaped cut on the trailing edge mostly caused by inter or intra-specific interactions (2.F): crescent shaped bite marks on the flank of the body mostly caused by shark attacks.
The injuries observed on 60.3% (79 of 131) of injured individuals were assigned to a natural origin. In most cases, these injuries were attributed to inter or intra-specific interactions (80.8% – 84 of 104 – representing 41.2% – 84 of 204 – of overall injuries, Figure 4.2E) or to an evident shark attack (19.2% – 20 of 104 – representing 9.8% – 20 of 204 – of overall injuries, Figure 4.2F). Finally, the origin of some injuries remained unidentified (29.9%, 61 of 204). Prevalence in the origin of injuries did not vary between the three geographical areas (χ2 = 5.7, p > 0.05).
Discussion
Community Structure Increasing Vulnerability
Social network analysis contributes to understand connectivity between populations and dynamics in social species that may inform management (Whitehead, 2009; Smith et al., 2016). Our study has revealed that Indo-pacific bottlenose dolphins are socially structured in three communities in southern New Caledonia at least in winter when surveys were predominantly conducted. Communities were, respectively, sighted across a straight line distance of about 30 km. Based on photo-identification comparison and social network analysis, dolphins observed around the Isle of Pines appeared to form a separate community (IoPc), with no links to the neighboring communities of the south-west lagoon (SWc) and the south lagoon (Sc), while the latter two were weakly connected. On one hand, the isolation of IoPc could be influenced by the topography of the region: although the Sarcelle Pass that separates the Merlet IMPA from the north-west reefs of the Isle of Pines is quite shallow (about 60 m deep), it appears to be deep and unsuitable to Indo-Pacific bottlenose dolphin presence (Kiszka et al., 2012). Then, social components might also limit interactions between communities. For example, communities of Indo-Pacific bottlenose dolphins are known to be influenced by cultural transmission leading to genetic and habitat segregation (Kopps et al., 2014). On the other hand, the presence of coral reef complexes in the south-eastern limit of the Merlet IMPA and north-west of the Isle of Pines would constitute potential bridges for connectivity: these geographic structures are expected to be suitable to Indo-Pacific bottlenose dolphin and might be used for dispersion. Dolphin use of the Merlet IMPA cannot be confirmed due to restricted access but an increase in sampling effort in this area would help understand geographical limits of IoPc.
The weak social connectivity observed between the Sc and the SWc in winter may be explained by a few individuals either (1) conducting social interchange between communities or (2) simply occupying wider larger home ranges than others. The first hypothesis could lead to reproductive interchange between communities. However, this phenomenon cannot be assessed as to date only one dolphin was observed in interaction with more than one community. The second hypothesis is supported by four individuals from the Sc encountered on both sides of the Ouen Island without interacting with individuals from the SWc. Even if these spatial movements occurred both across and within years (e.g., 2009, 2018), this potential social connectivity is expected to be quite low as it represents only 2.1 and 7.8% of all individuals encountered in the south lagoon and the south-west lagoon, respectively. The social structure highlighted in our study during the winter months was established over all dolphins sighted at least twice, although most studies conducted on social structure of dolphins used individuals resighted at least five times to reduce data skew from individual rarely resighted (e.g., Chilvers and Corkeron, 2001; Moreno and Acevedo-Gutiérrez, 2016; Baker et al., 2018). Given the small sample sizes collected in the southwest lagoon and the Isle of Pines, we considered this choice more suitable to our dataset. The collection of additional data in these two regions and over other seasons (austral summer) will provide a more exhaustive view of Indo-Pacific bottlenose dolphins connectivity, as this species is known to display seasonally-varying social structure (Sprogis et al., 2018a).
Despite their inherent capacity to move over long distances, in an open and barrier-free marine environment, Indo-Pacific bottlenose dolphin communities often show fine-scale demographic and genetic structuring (Hoelzel, 1998; Möller et al., 2007; Wiszniewski et al., 2009, 2010; Oremus et al., 2015). Over the east coast of Australia, gene flow among communities of Indo-Pacific bottlenose dolphins was predominately governed by the capacity of communities to locally adapt to heterogeneous habitats (Wiszniewski et al., 2010). Previous studies performed in other regions of New Caledonia highlight an extremely low genetic diversity and the presence of multiple putative management units based on photo-identification and genetic data (Oremus et al., 2009). Further molecular analyses are therefore needed to confirm the population structure revealed by photo-identification in the southern range of New Caledonia.
Suitable Habitats Overlapping With Coastal Activities
Habitat models revealed that Indo-Pacific bottlenose dolphin presence in the southern range of New Caledonia in austral winter is primarily driven by seabed depth. Indeed, both the combined environmental-spatial and environmental models predicted that shallow waters (0–50 m) were most suitable, either when located in coastal areas (e.g., around Nouméa and Plum, or in the Prony Bay) or reef patches (e.g., Gué reef or the reef plateaus south of Ouen Island). Such habitat preferences are consistent with other studies conducted in tropical archipelagoes (Gross et al., 2009). They reveal that Indo-Pacific bottlenose dolphins take advantage of the diversity of habitats that is found in the south-west and south lagoons of New Caledonia. This diversity of biotopes is likely to be reflected in the distribution of prey that dolphins rely on, as observed in Indo-Pacific bottlenose dolphins (Heithaus and Dill, 2006) and common bottlenose dolphins (Torres et al., 2008; Torres and Read, 2009). Indo-Pacific bottlenose dolphins are known to feed on demersal and reef fishes, small and medium-sized neritic fishes and cephalopods (Amir et al., 2005). In New Caledonia, photographs attest that Indo-Pacific bottlenose dolphins may feed on neritic lagoon fish such as halfbeaks (Hemirhamphus far). Analysis of stomach content on one individual stranded in 1994 in the south-west lagoon has revealed that the species may locally feed on a variety of lagoon fishes (Rivaton, pers. com.). Analysis of 81 otoliths highlighted that the diet of this dolphin was constituted of fishes found on sandy and muddy bottoms (39.5% Scolopsis sp. Nemipteridae, 9.9% Saurida undosquamis Synodontidae, 2.5% Gerres longirostris Gerreidae, 2.5% Nemipterus sp. Nemiptiridae) and coastal brackish waters and coral reefs (39.5% Lethrinus rubrioperculatus Lethrinidae, 2.5% Lutjanus fulviflamma Lutjanidae, 2.5% Echenis naucrates Echeneidae). The heterogeneity of habitats used is therefore likely related to variations in prey availability and further investigation into their diet may help understand their fine scale distribution in the southern range of New Caledonia.
In the south-west lagoon, the environmental model predicted highly suitable habitats for Indo-Pacific bottlenose dolphins in the coastal areas of Nouméa and Plum. The importance of these areas for the species remains to be confirmed with more dedicated surveys, as data is currently sparse in the south-west lagoon. In the south lagoon, the relatively high probability of dolphin presence predicted in Prony Bay by both models can be explained by its shallow depths, combined with proximity to the coast and muddy bottoms. On the contrary, model predictions differed in the south lagoon area over the Corne Sud (Figure 3), which was predicted to be highly suitable only by the environmental model, despite few dolphin observations in this zone (Figure 2). The Corne Sud was not hypothesized to be intensively used by Indo-Pacific bottlenose dolphins and we suggest that this prediction was biased by the high proportion of sedimentary muddy clay in this area. Compared to the Prony Bay where the sediments are composed by terrigenous clay, the clays of the Corne Sud are carbonated and do not host the same biotope. Predictions made in this area should therefore be cautionary questioned until more surveys are conducted. Finally, in the Isle of Pines, predicted suitability was generally lower than in the other two regions. The highest predictions were found in the shallow coastal waters of the island and decreased rapidly as depth increased. Here again, model extrapolation should be carefully interpreted, as more data covering a wider period and other seasons would be needed to validate predictions, as the Indo-Pacific bottlenose dolphins are known to shift habitats seasonally (Fury and Harrison, 2011; Sprogis et al., 2018a).
Unexpected Threats in New Caledonia
This study constitutes the first attempt to describe and interpret injuries as indicators of natural and anthropogenic threats to Indo-Pacific bottlenose dolphins in New Caledonia. The proportion of individuals showing injuries was similar among the three areas and concerned more than a third of all individuals identified in the southern range of New Caledonia. Injuries of both natural and human-related origins were found in all three areas, with no significant geographical difference in proportion.
Most of the injuries observed on dolphins were attributed to a natural origin, due to social interactions between individuals. Indeed, marks resulting from such interactions are commonly found in gregarious species such as dolphins (McCann, 1974; MacLeod, 1998; Scott et al., 2005). Tooth rakes resulting from agonistic interactions are known to be due to sexual coercion and intra-sexual male competition (Scott et al., 2005; Marley et al., 2013).
Among the other injuries of natural origin, 19.2% (20 of 104) appeared to have been caused by evident inter-specific attacks, most likely due to sharks. None of these were related to cookie-cutter sharks (Isistius sp.), which is consistent to the known pelagic habitat of these shark species. Estimating which shark species may predate on Indo-Pacific bottlenose dolphins remains challenging (Heithaus, 2001; Smith et al., 2018; Sprogis et al., 2018b). In New Caledonia, large species of sharks such as tiger sharks (Galeocerdo cuvier) or bull sharks (Carcharhinus leucas) are present in the lagoon and are known to be involved in attacks on dolphins (Heithaus et al., 2017). Such predation marks have been observed in other Indo-Pacific bottlenose dolphin populations (Heithaus, 2001; Heithaus et al., 2017; Smith et al., 2018; Sprogis et al., 2018b). For instance, across 101 individuals studied in La Réunion Island, 20 showed shark-inflicted bite wounds (Heithaus et al., 2017), leading to an attack rate (19.8%) similar to our study. Other known predators such as killer whales (Orcinus orca) are less likely to affect dolphins in the New Caledonia lagoon as no injuries related to this species has been highlighted within our dataset and they have never been observed within the lagoon (Garrigue and Poupon, 2013; Derville et al., 2018).
Studies of human-related injuries provide good insights into anthropogenic impacts on cetacean population health. In this way, anthropogenic interactions with coastal dolphins increase the occurrence of injuries (Christiansen et al., 2016) and may affect survival, reproduction (Wells et al., 2008) and social interactions (Greenfield et al., 2020). Human-related injuries were observed in surprising proportions in all three study regions in the southern range of New Caledonia. Most of these were likely due to boat strikes. Indo-Pacific bottlenose dolphins are known to interact with vessels (Hawkins and Gartside, 2009) but boat strikes remain poorly documented. Strandings of Indo-Pacific dolphins have been associated with deadly propeller strikes in the past (Byard et al., 2012). Moreover, the prevalence of injuries related to boat strikes on the west coast of Florida was estimated at 6% for common bottlenose dolphins (Wells and Scott, 1997; Bechdel et al., 2009), against 9.8% (33 of 338) of all Indo-Pacific bottlenose dolphins identified in this study. In the southern range of New Caledonia, maritime traffic related to recreational use of the lagoon is growing, particularly within local MPAs during weekends and in austral summer (Gonson et al., 2016). It differs considerably between the three study regions, with a predominance in the south-west lagoon. Indeed, the latest human census conducted in 2019 showed that about 70% of the whole New Caledonian population was concentrated in the area of Nouméa1 in the south-west lagoon. Several marine protected areas exist within this region (natural reserves with restrictions, Figure 1), but recreational activities related to navigation remain allowed, such as motor-boats, sail-boats, jet-skis and taxi-boats (Gonson et al., 2017). In addition, the coastal waters around Nouméa are intensively used by recreational users, such as high-speed motorized vessels and engine-free nautical activities (e.g., kitesurf, windsurf), that likely increase collision risk. In the south lagoon, there are fewer recreational boats, mostly present during the whale-watching season in austral winter (Derville et al., 2019). Judging by the seemingly lower human use of the south lagoon, it is surprising that dolphins should bear as many injuries from propellers as in the south-west lagoon. Further investigation of dolphin injuries of human-related origin in no-go zones such as the Merlet IMPA would provide interesting insights.
Many studies suggest that bycatch is the dominant threat affecting Indo-Pacific bottlenose dolphins throughout their range (Kiszka et al., 2008; Amir, 2010; Mansur et al., 2012; Shirakihara and Shirakihara, 2012) and it has been estimated that in some populations, it is sufficiently high to cause declines (Braulik et al., 2019). Yet, in this study collisions with vessels appeared to be preponderant (87.2% of human-related origin injuries) compared to fishery-related injuries (10.3% of human-related origin injuries). Informal fisheries are known to occur in the south-west and south lagoons of New Caledonia (Jollit et al., 2010). Based on interviews with local fishermen from 2004 to 2005, fishing pressure appeared higher in coral habitats (mid-lagoon and nearby islet reefs), on the shore next to villages, by the reef passes and in Prony Bay (Jollit et al., 2010). This artisanal fishing does not seem to threaten dolphins as much as maritime traffic.
Most of the injuries were observed on the dorsal fin, probably because it is the part of the individual that most often emerges out of the water. Thus, the number of injuries on the body might be underestimated. We also acknowledge the difficulty of attributing a specific origin to an injury. Yet, using an objective classification system, tested in other study areas, should have limited potential interpretation biases (Luksenburg, 2014). In the end, this study constitutes an important contribution for the conservation of the species and will help the implementation of adequate management policy in the future.
Conclusion
The social structure and spatial distribution of Indo-Pacific bottlenose dolphins in the southern range of New Caledonia during the winter season has highlighted three distinct communities that should be considered as three independent management units, a suggestion supported by an extremely low genetic diversity which alters the resilience of this species in New Caledonia. Habitat use analysis further revealed that Indo-Pacific bottlenose dolphins mostly use coastal habitats, where anthropogenic activities are documented to be quite high. The assessment of anthropogenic threats has revealed that maritime traffic can constitute a threat to Indo-Pacific bottlenose dolphins in the southern range of New Caledonia by causing propeller hits. These new findings should be considered for the implementation of future management measures that will contribute to the conservation of Indo-Pacific bottlenose dolphins in New Caledonia.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
Ethical review and approval was not required for the animal study because this study is only based on data collected without animal manipulation.
Author Contributions
CB, SD, and CG: study design and writing the manuscript. CG, MO, and CB: data collection and data processing. CB, SD, and JL: data analysis. All authors critically reviewed the manuscript.
Funding
We thank Comité Consultatif Coutumier Environemental (CCCE) for funding this study within the TCHITU Project. We also thank Vale and the Province Sud for their financial contribution to the fieldwork carried on in the Great South Lagoon and Société Le Nickel for the data collected in the South West Lagoon.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank all the volunteers for their personal implication in the field through the Opération Cétacés association. We also especially thank Pascal Douillet and Christophe Chevillon for their help with sedimentary data, and the late Jacques Rivaton for the analysis of stranded dolphin stomach content. Finally, the two reviewers who contributed to improve the manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2021.606975/full#supplementary-material
Abbreviations
IMPA, Integral Marine Protected Area; IoPc, Isle of Pines community; Sc, South lagoon community; SWc, South-west lagoon community.
Footnotes
References
Amaral, A. R., Smith, B. D., Mansur, R. M., Brownell, R. L., and Howard, J. (2017). Oceanographic drivers of population differentiation in Indo-Pacific bottlenose (Tursiops aduncus) and humpback (Sousa spp.) dolphins of the northern Bay of Bengal. Conserv. Genet. 18, 371–381. doi: 10.1007/s10592-016-0913-7
Amir, O. A. (2010). Biology, Ecology and Anthropogenic Threats of Indo-Pacific Bottlenose Dolphins in East Africa. Ph.D. thesis, Stockholm University, Stockholm.
Amir, O. A., Berggren, P., Ndaro, S. G. M., and Jiddawi, N. S. (2005). Feeding ecology of the Indo-Pacific bottlenose dolphin (Tursiops aduncus) incidentally caught in the gillnet fisheries off Zanzibar, Tanzania. Estuar. Coast. Shelf Sci. 63, 429–437. doi: 10.1016/j.ecss.2004.12.006
Andréfouët, S., Chagnaud, N., Chauvin, C., and Kranenburg, C. (2008). Atlas of French Overseas Coral Reefs, 153. Available online at: http://umr-entropie.ird.nc/index.php/home/ressources/mcrmp (accessed February 2016).
Ansmann, I. C., Parra, G. J., Lanyon, J. M., and Seddon, J. M. (2012). Fine-scale genetic population structure in a mobile marine mammal: inshore bottlenose dolphins in Moreton Bay, Australia. Mol. Ecol. 21, 4472–4485. doi: 10.1111/j.1365-294X.2012.05722.x
Avila, I. C., Kaschner, K., and Dormann, C. (2018). Current global risks to marine mammals: taking stock of the threats. Biol. Conserv. 221, 44–58. doi: 10.1016/j.biocon.2018.02.021
Baker, I., O’Brien, J., McHugh, K. A., Ingram, S. N., and Berrow, S. (2018). Bottlenose dolphin (Tursiops truncatus) social structure in the Shannon Estuary, Ireland, is distinguished by age- and area-related associations. Mar. Mammal Sci. 34, 458–487. doi: 10.1111/mms.12462
Bechdel, S. E., Mazzoil, M. S., Murdoch, M. E., Howells, E. M., Reif, J. S., McCullosh, S. D., et al. (2009). Prevalence and impacts of motorized vessels on bottlenose dolphins (Tursiops truncatus) in the Indian River Lagoon, Florida. Aquat. Mamm. 35, 367–377. doi: 10.1578/AM.35.3.2009.367
Bejder, L., Fletcher, D., and Bräger, S. (1998). A method for testing association patterns of social animals. Anim. Behav. 56, 719–725. doi: 10.1006/anbe.1998.0802
Bourgogne, H., Derville, S., and Garrigue, C. (2018) Etude du Trafic Maritime Dans le Grand Lagon Sud afin D’apprécier les Risques de Collision et de Dérangement pour la Population de Baleines à Bosse de Nouvelle-Calédonie. Rapport non publié pour le CCCE. Nouméa: Opération Cétcés.
Braulik, G., Natoli, A., Kiszka, J., Parra, G., Plön, S., and Smith, B. (2019). Tursiops Aduncus, Indo-Pacific Bottlenose Dolphin. IUCN Red List Threat. Species e.T41714A5. doi: 10.2305/IUCN.UK.2019-3.RLTS.T41714A50381127.en
Burnham, K. P., and Anderson, D. R. (2004). Multimodel inference: understanding AIC and BIC in model selection. Sociol. Methods Res. 33, 261–304. doi: 10.1177/0049124104268644
Byard, R. W., Winskog, C., Machado, A., and Boardman, W. (2012). The assessment of lethal propeller strike injuries in sea mammals. J. Forensic Leg. Med. 19, 158–161. doi: 10.1016/j.jflm.2011.12.017
Cairns, S. J., and Schwager, S. J. (1987). A comparison of association indices. Anim. Behav. 35, 1454–1469. doi: 10.1016/S0003-3472(87)80018-0
Chardy, P., Chevillon, C., and Clavier, J. (1988). Major benthic communities of the south-west lagoon of New Caledonia. Coral Reefs 7, 69–75. doi: 10.1007/BF00301643
Charlesworth, D., and Charlesworth, B. (1987). Inbreeding depression and its evolutionary consequences. Annu. Rev. Ecol. Evol. Syst. 18, 237–268. doi: 10.1146/annurev.es.18.110187.001321
Chevillotte, V., Douillet, P., Cabioch, G., Lafoy, Y., Lagabrielle, Y., and Maurizot, P. (2005). Geomorphological evolution of the foreland of southwestern New Caledonia during the last glacial-interglacial episodes. C. R. Geosci. 337, 695–701. doi: 10.1016/j.crte.2005.02.008
Chilvers, B. L., and Corkeron, P. J. (2001). Trawling and bottlenose dolphins’ social structure. Proc. R. Soc. London B Biol. Sci. 268, 1901–1905. doi: 10.1098/rspb.2001.1732
Chilvers, B. L. (2001). Behavioural Ecology of Bottlenose Dolphins, Tursiops aduncus, in S.E. Queensland, Australia: Adaptations to Ecological and Anthropogenic Influences, 231. Ph.D. thesis, James Cook University, Townsville.
Christiansen, F., McHugh, K. A., Bejder, L., Siegal, E. M., Lusseau, D., McCabe, E. B., et al. (2016). Food provisioning increases the risk of injury in a long-lived marine top predator. R. Soc. Open Sci. 3:160560. doi: 10.1098/rsos.160560
Cottarel, G., Garrigue, C., Dulau, V. (2016). “Tursiops aduncus (Ehrenberg, 1832): grand dauphin de l’Indo-Pacifique,” in Atlas des Mammifères Sauvages de France: Mammifères Marins:IRD, (74), (Patrimoines Naturels; 74), Vol. 1, A. Savouré-Soubelet, S. Aulagnier, S. P. Haffner, F. Moutou, O. van Canneyt, J. B. Charrassin, et al. (Paris: MNHN), 296–299.
Debenay, J.-P. (1987). Sedimentology in the southwestern lagoon of New Caledonia, SW Pacific. J. Coast. Res. 3, 77–91.
DeMaster, D. P., Fowler, C. W., Perry, S. L., and Richlen, M. F. (2001). Predation and competition: the impact of fisheries on marine-mammal populations over the next one hundred years. J. Mammal. 82, 641–651. doi: 10.1644/1545-15422001082>0641:PACTIO<2.0.CO;2
Derville, S., Torres, L. G., Dodémont, R., Perard, V., and Garrigue, C. (2019). From land and sea, long-term data reveal persistent humpback whale (Megaptera novaeangliae) breeding habitat in New Caledonia. Aquat. Conserv. Mar. Freshw. Ecosyst. 29, 1697–1711. doi: 10.1002/aqc.3127
Derville, S., Torres, L. G., and Garrigue, C. (2018). Social segregation of humpback whales in contrasted coastal and oceanic breeding habitats. J. Mammal. 99, 41–54. doi: 10.1093/jmammal/gyx185
Dormann, C. F., McPherson, J. M., Araújo, M. B., Bivand, R., Bolliger, J., Carl, G., et al. (2007). Methods to account for spatial autocorrelation in the analysis of species distributional data: a review. Ecography 30, 609–628. doi: 10.1111/j.2007.0906-7590.05171.x
DTSI (2016). Atlas Bathymétrique de Nouvelle-Calédonie. Portail de L’information Géographique de Nouvelle-Calédonie. Available online at: http://www.geoportal.gouv.nc (accessed February 2016).
Friedman, J. (2001). Greedy function approximation: a gradient boosting machine. Ann. Stat. 29, 1189–1232. doi: 10.1214/009053606000000795
Fury, C. A., and Harrison, P. L. (2011). Seasonal variation and tidal influences on estuarine use by bottlenose dolphins (Tursiops aduncus). Estuar. Coast. Shelf Sci. 93, 389–395. doi: 10.1016/j.ecss.2011.05.003
Garrigue, C., Greaves, J., and Chambellant, M. (2001). Characteristics of the New Caledonian humpback whale population (Megaptera novaeangliae). Mem. Queensl. Museum 47, 539–546.
Garrigue, C., and Poupon, M. (2013). Guide D’identification des Mammifères Marins de Nouvelle-Calédonie. Nouvelle-Calédonie: Opération Cétacés, 128.
Genov, T., Centrih, T., Kotnjek, P., and Hace, A. (2019). Behavioural and temporal partitioning of dolphin social groups in the northern Adriatic Sea. Mar. Biol. 166, 1–14. doi: 10.1007/s00227-018-3450-8
Gonson, C., Pelletier, D., Alban, F., Giraud-carrier, C., and Ferraris, J. (2017). Influence of settings management and protection status on recreational uses and pressures in marine protected areas. J. Environ. Manage. 200, 170–185. doi: 10.1016/j.jenvman.2017.05.051
Gonson, C., Pelletier, D., Gamp, E., Preuss, B., Jollit, I., and Ferraris, J. (2016). Decadal increase in the number of recreational users is concentrated in no-take marine reserves. Mar. Pollut. Bull. 107, 144–154. doi: 10.1016/j.marpolbul.2016.04.007
Gowans, S., Würsig, B., and Karczmarski, L. (2007). The social structure and strategies of Delphinids: predictions based on an ecological framework. Adv. Mar. Biol. 53, 195–294. doi: 10.1016/S0065-2881(07)53003-8
Gray, H. W. I., Nishida, S., Welch, A. J., Moura, A. E., Tanabe, S., Kiani, M. S., et al. (2018). Cryptic lineage differentiation among Indo-Pacific bottlenose dolphins (Tursiops aduncus) in the northwest Indian Ocean. Mol. Phylogenet. Evol. 122, 1–14. doi: 10.1016/j.ympev.2017.12.027
Greenfield, M. R., McHugh, K. A., Wells, R. S., and Rubenstein, D. I. (2020). Anthropogenic injuries disrupt social associations of common bottlenose dolphins (Tursiops truncatus) in Sarasota Bay, Florida. Mar. Mammal Sci. 37, 29–44. doi: 10.1111/mms.12729
Gross, A., Kiszka, J., Van Canneyt, O., Richard, P., and Ridoux, V. (2009). A preliminary study of habitat and resource partitioning among co-occurring tropical dolphins around Mayotte, southwest Indian Ocean. Estuar. Coast. Shelf Sci. 84, 367–374. doi: 10.1016/j.ecss.2009.05.017
Hale, P. T., Barreto, A. S., and Ross, G. J. B. (2000). Comparative morphology and distribution of the aduncus and truncatus forms of bottlenose dolphin Tursiops in the Indian and Western Pacific Oceans. Aquat. Mamm. 26, 101–110. doi: 10.1016/b978-0-12-440280-5.50009-3
Hastie, T., and Tibshirani, R. (1990). “Generalized additive models,” in Monographs on Statistics and Applied Probability London: Chapman and Hall, 352.
Hawkins, E. R., and Gartside, D. F. (2009). Interactive behaviours of bottlenose dolphins (Tursiops aduncus) during encounters with vessels. Aquat. Mamm. 35, 259–268. doi: 10.1578/am.35.2.2009.259
Heithaus, M. R. (2001). Shark attacks on bottlenose dolphins (Tursiops aduncus) in Shark Bay, Western Australia: attack rate, bite scar frequencies, and attack seasonality. Mar. Mammal Sci. 17, 526–539. doi: 10.1111/j.1748-7692.2001.tb01002.x
Heithaus, M. R., and Dill, L. M. (2006). Does tiger shark predation risk influence foraging habitat use by bottlenose dolphins at multiple spatial scales? Oikos 114, 257–264. doi: 10.1111/j.2006.0030-1299.14443.x
Heithaus, M. R., Kiszka, J., Cadinouche, A., Dulau, V., Boucaud, V., and Pérez-Jorge, S. (2017). Spatial variation in shark-inflicted injuries to Indo-Pacific bottlenose dolphins (Tursiops aduncus) of the southwestern Indian Ocean. Mar. Mammal Sci. 33, 335–341. doi: 10.1111/mms.12346
Hoelzel, A. R. (1998). Genetic structure of cetacean populations in sympatry, parapatry, and mixed assemblages: implications for conservation policy. J. Hered. 89, 451–458. doi: 10.1093/jhered/89.5.451
Jollit, I., Léopold, M., Guillemot, N., David, G., Chabanet, P., Lebigre, J. M., et al. (2010). Geographical aspects of informal reef fishery systems in New Caledonia. Mar. Pollut. Bull. 61, 585–597. doi: 10.1016/j.marpolbul.2010.06.033
Kiszka, J., Pelourdeau, D., and Ridoux, V. (2008). Body scars and dorsal fin disfigurements as indicators of interaction between small cetaceans and fisheries around the Mozambique Channel island of Mayotte. West. Indian Ocean J. Mar. Sci. 7, 185–193.
Kiszka, J., Simon-Bouhet, B., Gastebois, C., Pusineri, C., and Ridoux, V. (2012). Habitat partitioning and fine scale population structure among insular bottlenose dolphins (Tursiops aduncus) in a tropical lagoon. J. Exp. Mar. Bio. Ecol. 41, 176–184. doi: 10.1016/j.jembe.2012.03.001
Kopps, A. M., Ackermann, C. Y., Sherwin, W. B., Allen, S. J., Bejder, L., Kru, M., et al. (2014). Cultural transmission of tool use combined with habitat specializations leads to fine-scale genetic structure in bottlenose dolphins. Proc. R. Soc. London B Biol. Sci. 281:20133245. doi: 10.1098/rspb.2013.3245
Luksenburg, J. A. (2014). Prevalence of external injuries in small cetaceans in aruban waters, Southern Caribbean. PLoS One 9:e88988. doi: 10.1371/journal.pone.0088988
MacLeod, C. D. (1998). Intraspecific scarring in odontocete cetaceans: an indicator of male “quality” in aggressive social interactions? J. Zool. 244, 71–77. doi: 10.1017/S0952836998001083
Manlik, O., Chabanne, D., Daniel, C., Bejder, L., Allen, S. J., and Sherwin, W. B. (2019). Demography and genetics suggest reversal of dolphin source-sink dynamics, with implications for conservation. Mar. Mammal Sci. 35, 732–759. doi: 10.1111/mms.12555
Mansur, R. M., Strindberg, S., and Smith, B. D. (2012). Mark-resight abundance and survival estimation of Indo-Pacific bottlenose dolphins, Tursiops aduncus, in the Swatch-of-No-Ground, Bangladesh. Mar. Mammal Sci. 28, 561–578. doi: 10.1111/j.1748-7692.2011.00520.x
Marley, S. A., Cheney, B., and Thompson, P. M. (2013). Using tooth rakes to monitor population and sex differences in aggressive behaviour in bottlenose dolphins (Tursiops truncatus). Aquat. Mamm. 39, 107–115. doi: 10.1578/AM.39.2.2013.107
Marra, G., and Wood, S. N. (2011). Practical variable selection for generalized additive models. Comput. Stat. Data Anal. 55, 2372–2387. doi: 10.1016/j.csda.2011.02.004
McHugh, K. A., Allen, J. B., Barleycorn, A. A., and Wells, R. S. (2011). Severe Karenia brevis red tides influence juvenile bottlenose dolphin (Tursiops truncatus) behavior in Sarasota Bay, Florida. Mar. Mammal Sci. 27, 622–643. doi: 10.1111/j.1748-7692.2010.00428.x
Möller, L. M., Wiszniewski, J., Allen, S. J., and Beheregaray, L. B. (2007). Habitat type promotes rapid and extremely localised genetic differenciation in dolphins. Mar. Freshw. Res. 58, 640–648. doi: 10.1071/MF06218
Moreno, K., and Acevedo-Gutiérrez, A. (2016). The social structure of Golfo Dulce bottlenose dolphins (Tursiops truncatus) and the influence of behavioural state. R. Soc. Open Sci. 3:160010. doi: 10.1098/rsos.160010
Oremus, M., Garrigue, C., Schaffar, A., and Baker, C. S. (2009). “Fine-scale demographic and genetic structure in insular Indo-Pacific bottlenose dolphins (Tursiops aduncus). Implications for conservation of local populations,” in Proceedings of the Society for Marine Mammalogy Biennal Conference, Quebec, QC.
Oremus, M., Garrigue, C., Tezanos-Pinto, G., and Scott Baker, C. (2015). Phylogenetic identification and population differentiation of bottlenose dolphins (Tursiops spp.) in Melanesia, as revealed by mitochondrial DNA. Mar. Mammal Sci. 31, 1035–1056. doi: 10.1111/mms.12210
Parra, G. J., Corkeron, P. J., and Arnold, P. (2011). Grouping and fission-fusion dynamics in Australian snubfin and Indo-Pacific humpback dolphins. Anim. Behav. 82, 1423–1433. doi: 10.1016/j.anbehav.2011.09.027
Phillips, S. J., Dudík, M., Elith, J., Graham, C. H., Lehmann, A., Leathwick, J., et al. (2009). Sample selection bias and presence-only distribution models: implications for background and pseudo-absence data. Ecol. Appl. 19, 181–197. doi: 10.1890/07-2153.1
QGIS Development Team (2018). QGIS Geographic Information System. Open source Geospatial Foundation Project. Version 2.18.13 Las Palmas. Available online at: http://qgis.osgeo.org (accessed January 2018
RCoreTeam (2020). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Ross, G. J. B., Cockcroft, V. G., and Butterworth, D. G. (1987). Offshore distribution of bottlenosed dolphins in Natal coastal waters and Algoa Bay, Eastern Cape. S. Afr. J. Zool. 22, 50–56. doi: 10.1080/02541858.1987.11448020
Scott, E. M., Mann, J., Watson-Capps, J. J., Sargeant, B. L., and Connor, R. C. (2005). Aggression in bottlenose dolphins: evidence for sexual coercion, male-male competition, and female tolerance through analysis of tooth-rake marks and behaviour. Behaviour 142, 21–44. doi: 10.1163/1568539053627712
Shirakihara, M., and Shirakihara, K. (2012). Bycatch of the Indo-Pacific bottlenose dolphin (Tursiops aduncus) in gillnet fisheries off Amakusa-Shimoshima Island, Japan. J. Cetacean Res. Manag. 12, 345–351.
Smith, F., Allen, S. J., Bejder, L., and Brown, A. M. (2018). Shark bite injuries on three inshore dolphin species in tropical northwestern Australia. Mar. Mammal Sci. 34, 87–99. doi: 10.1111/mms.12435
Smith, H., Frère, C., Kobryn, H., and Bejder, L. (2016). Dolphin sociality, distribution and calving as important behavioural patterns informing management. Anim. Conserv. 19, 462–471. doi: 10.1111/acv.12263
Sprogis, K. R., Christiansen, F., Raudino, H. C., Kobryn, H. T., Wells, R. S., and Bejder, L. (2018a). Sex-specific differences in the seasonal habitat use of a coastal dolphin population. Biodivers. Conserv. 27, 3637–3656. doi: 10.1007/s10531-018-1618-7
Sprogis, K. R., King, C., Bejder, L., and Loneragan, N. R. (2018b). Frequency and temporal trends of shark predation attempts on bottlenose dolphins (Tursiops aduncus) in temperate Australian waters. J. Exp. Mar. Bio. Ecol. 508, 35–43. doi: 10.1016/j.jembe.2018.08.008
Sprogis, K. R., Raudino, H. C., Rankin, R., McLeod, C. D., and Bejder, L. (2016). Home range size of adult Indo-Pacific bottlenose dolphins (Tursiops aduncus) in a coastal and estuarine system is habitat and sex-specific. Mar. Mammal Sci. 32, 287–308. doi: 10.1111/mms.12260
Thompson, J. W., Zero, V. H., Schwacke, L. H., Speakman, T. R., Quigley, B. M., Morey, J. S. M., et al. (2019). FinFindR: computer-assisted recognition and identification of bottlenose dolphin photos in R. bioRxiv [Preprint], 10. doi: 10.1101/825661
Torres, L. G., and Read, A. J. (2009). Where to catch a fish? The influence of foraging tactics on the ecology of bottlenose dolphins (Tursiops truncatus) in Florida Bay, Florida. Mar. Mammal Sci. 25, 797–815. doi: 10.1111/j.1748-7692.2009.00297.x
Torres, L. G., Read, A. J., and Halpin, P. (2008). Fine-scale habitat modeling of a top marine predator: do prey data improve predictive capacity? Ecol. Appl. 18, 1702–1717. doi: 10.1890/07-1455.1
UNESCO (2009). Lagoons of New Caledonia: Reef Diversity and Associated Ecosystems. U.W.H. Centre. Available online at: (whc.unesco.org/en/list/1115/documents/Accessed2018-06-20) (accessed June 20, 2018).
van Aswegen, M., Christiansen, F., Symons, J., Mann, J., Nicholson, K., Sprogis, K., et al. (2019). Morphological differences between coastal bottlenose dolphin (Tursiops aduncus) populations identified using non-invasive stereo-laser photogrammetry. Sci. Rep. 9:12235. doi: 10.1038/s41598-019-48419-3
Van Waerebeek, K., Baker, A. N., Félix, F., Gedamke, J., Iñiguez, M., Sanino, G. P., et al. (2008). Vessel collisions with small cetaceans worldwide and with large whales in the Southern Hemisphere, an initial assessment. Lat. Am. J. Aquat. Mamm. 6, 43–69. doi: 10.5597/lajam00109
Vargas-Fonseca, O. A., Kirkman, S. P., Conry, D., Rishworth, G. M., Cockcroft, V., and Pistorius, P. A. (2018). Distribution and habitat use of Indo-Pacific bottlenose dolphins Tursiops aduncus along the south coast of South Africa. Afr. J. Mar. Sci. 40, 439–450. doi: 10.2989/1814232X.2018.1547221
Wang, J. Y., and Yang, S. (2009). “Indo-Pacific bottlenose dolphin: Tursiops aduncus,” in Encyclopedia of Marine Mammals, 2nd Edn, eds W. F. Perrin, B. Würsig, and J. G. M. Thewissen (Amsterdam: Academic Press), 602–608.
Wells, R. S., Allen, J. B., Hofmann, S., Bassos-Hull, K., Fauquier, D. A., Barros, N. B., et al. (2008). Consequences of injuries on survival and reproduction of common bottlenose dolphins (Tursiops truncatus) along the west coast of Florida. Mar. Mammal Sci. 24, 774–794. doi: 10.1111/j.1748-7692.2008.00212.x
Wells, R. S., and Scott, M. D. (1997). Seasonal incidence of boat strikes on bottlenose dolphins near Sarasota, Florida. Mar. Mammal Sci. 13, 475–480. doi: 10.1111/j.1748-7692.1997.tb00654.x
Whitehead, H. (2009). SOCPROG programs: analyzing animal social structures. Behav. Ecol. Sociobiol. 63, 765–778. doi: 10.1007/s00265-008-0697-y
Wiszniewski, J., Allen, S. J., and Möller, L. M. (2009). Social cohesion in a hierarchically structured embayment population of Indo-Pacific bottlenose dolphins. Anim. Behav. 77, 1449–1457. doi: 10.1016/j.anbehav.2009.02.025
Wiszniewski, J., Beheregaray, L. B., Allen, S. J., and Möller, L. M. (2010). Environmental and social influences on the genetic structure of bottlenose dolphins (Tursiops aduncus) in Southeastern Australia. Conserv. Biol. 11, 1405–1419. doi: 10.1007/s10592-009-9968-z
Wood, S. (2017). Generalized Additive Models: An Introduction with R, 2nd Edn. New York, NY: CRC Press.
Keywords: odontocetes, New Caledonia, social communities, habitat, anthropogenic impacts, scars
Citation: Bonneville CD, Derville S, Luksenburg JA, Oremus M and Garrigue C (2021) Social Structure, Habitat Use and Injuries of Indo-Pacific Bottlenose Dolphins (Tursiops aduncus) Reveal Isolated, Coastal, and Threatened Communities in the South Pacific. Front. Mar. Sci. 8:606975. doi: 10.3389/fmars.2021.606975
Received: 16 September 2020; Accepted: 13 January 2021;
Published: 04 February 2021.
Edited by:
Jeremy Kiszka, Florida International University, United StatesReviewed by:
Kate R. Sprogis, Murdoch University, AustraliaJulian A. Tyne, South Atlantic Environmental Research Institute, Falkland Islands
Copyright © 2021 Bonneville, Derville, Luksenburg, Oremus and Garrigue. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Claire Daisy Bonneville, Y2xhaXJlLmJvbm5ldmlsbGVAaXJkLmZy