 Concepción Marcos1
Concepción Marcos1 David Díaz2Katharina Fietz3Aitor Forcada4
David Díaz2Katharina Fietz3Aitor Forcada4 Amanda Ford5
Amanda Ford5 José Antonio García-Charton1Raquel Goñi2Philippe Lenfant6Sandra Mallol2David Mouillot7
José Antonio García-Charton1Raquel Goñi2Philippe Lenfant6Sandra Mallol2David Mouillot7 María Pérez-Marcos8Oscar Puebla9Stephanie Manel10
María Pérez-Marcos8Oscar Puebla9Stephanie Manel10 Angel Pérez-Ruzafa1*
Angel Pérez-Ruzafa1*- 1Department of Ecology and Hydrology, Regional Campus of International Excellence “Mare Nostrum”, University of Murcia, Murcia, Spain
- 2Centro Oceanográfico de Baleares, Instituto Español de Oceanografía, Palma de Mallorca, Spain
- 3Geomar Helmholtz Centre for Ocean Research Kiel, Kiel, Germany
- 4Department of Marine Sciences and Applied Biology, University of Alicante, Alicante, Spain
- 5School of Agriculture, Geography, Environment, Ocean and Natural Sciences (SAGEONS), University of the South Pacific, Suva, Fiji
- 6Université de Perpignan Via Domitia, Centre de Formation et de Recherche sur les environnements Méditerranéens, UMR 5110, 58 Avenue Paul Alduy, Perpignan, France
- 7MARBEC, Université de Montpellier, CNRS, IFREMER, IRD, Montpellier, France
- 8Biological Pest Control and Ecosystem Services Laboratory, Instituto Murciano de Investigación y Desarrollo Agrario y Alimentario (IMIDA), La Alberca, Spain
- 9Ecology Department, Leibniz Centre for Tropical Marine Research (ZMT), Bremen, Germany
- 10CEFE, University of Montpellier, CNRS, EPHE-PSL University, IRD, Montpellier, France
Marine protected areas (MPAs) are globally important environmental management tools that provide protection from the effects of human exploitation and activities, supporting the conservation of marine biological diversity, habitats, ecosystems and the processes they host, as well as resources in a broad sense. Consequently, they are also expected to manage and enhance marine ecosystem services and material, non-material, consumptive and non-consumptive goods, and benefits for humans. There is however certain confusion on what constitutes an ecosystem service, and it is not always easy to distinguish between them and societal benefits. The main nuance is that an ecosystem service is the aptitude an ecosystem has or develops naturally or as consequence of a management action, and that manifests through its own properties (productivity, diversity, stability, quality of its key parameters, etc.), while a societal benefit is the economic or other profitability (emotional, educational, scientific, etc.) that humans obtain from said service or quality. In this work, 268 publications, together with our own experiences in the different investigations carried out in the MPAs that are part of the BiodivERsA3-2015-21 RESERVEBENEFIT European project, have been selected, reviewed and discussed to analyze the knowledge status of the expected ecosystem services of MPAs and the societal benefits derived from them, sometimes providing information on their evidence, when they exist. We define and classify the effects of protection, ecosystem services and societal benefits and elaborate a conceptual model of the cause-effect relationships between them.
Introduction
Conservation of the marine environment–its physical and ecological functioning and its biodiversity–is of utmost importance for maintaining global natural processes of the planet, regulating its responses to major future challenges such as the minimization of and adaptation to climate changes, and from the point of view of guaranteeing societal goods and benefits (Roberts et al., 2017; Pantzar et al., 2018).
Different designations like “marine reserve,” “no-take zone” or “fish box” have mostly been considered important tools in the management and protection of fishery resources (Pérez-Ruzafa et al., 2017). However, there is a broader concept, that of Marine Protected Area (MPA), which includes a greater number of objectives and regulations despite in many cases offering a lower degree of protection (Roberts and Hawkins, 2000; Gell and Roberts, 2003a; Costello and Ballantine, 2015). Following Reuchlin-Hugenholtz and McKenzie (2015), an MPA is a “marine space designated and effectively managed to protect marine ecosystems, processes, habitats, and species, which can contribute to the restoration and replenishment of resources for social, economic, and cultural enrichment.” This term thus could encompass many other concepts that are used in a similar sense, although sometimes involving a greater or lesser degree of restrictions, such as the aforementioned marine reserve, fully protected marine area, no-take zone, fish box or fishery closure area, marine sanctuary, ocean sanctuary, marine park or locally managed marine area, and could also include Special Areas of Conservation or Sites of Special Scientific Interest. In this review we refer to MPAs in this broad meaning, although on some occasions we will refer to marine reserves when the original bibliographic data does so.
The main MPA global policy targets aim to protect by 2020 at least 10% of coastal and marine areas (Convention on Biological Diversity of Aichi, Japan, 2010, www.cbd.int accessed 12/02/20), and the urgency of increasing the ocean area covered by ecologically representative and well-connected MPA systems to at least 30% by 2030 is underlined in the IUCN World Parks Congress of Sydney, Australia, 2014 (Charles et al., 2016; Krueck et al., 2017). In Europe, the European Marine Strategy Framework Directive aims to establish a network of MPAs as one of the main protection measures to maintain and improve the sustainable use of European marine waters, the biodiversity and biological connectivity, the quality and occurrence of habitats and the distribution and abundance of species (EC, 2008). These objectives could be achieved if the design of MPA networks took into account the recommendations based on a reserve size that optimizes the minimum cost of surveillance and maintenance, the maximum protection efficiency (estimated at around 600 ha for the no take area), and the effective biomass export distances (estimated at about 5–15 km) (Pérez-Ruzafa et al., 2008a, 2017).
Nevertheless, the amount of established protected area in the oceans clearly falls short of global policy targets. This percentage currently amounts to 5.7% of implemented zones (www.mpatlas.org accessed 12/02/20), and only 2.6% is in strongly implemented or fully protected areas (Sala et al., 2018; www.mpatlas.org accessed 12/02/20). In the case of European seas, by the end of 2016, 10.8% of their surface had been designated as MPAs, although with important differences between geographical areas (27.1% of the Greater North Sea vs. 2.9% of the Aegean-Levantine Sea) (www.eea.europa.eu/themes/water/europes-seas-and-coasts/assessments/marine-protected-areas, accessed 12/02/20).
Many existing MPAs are, however, not implemented in fact and only exist as “paper parks” where legislation is not enforced, the necessary surveillance is not present, management resources are lacking, and management plans are inactive or deficient, or do not comply with the regulations in place (Gilman, 1997; Roberts and Hawkins, 2000; Gell and Roberts, 2003a; Reker, 2015; Garcia-Rubies et al., 2017; Pieraccini et al., 2017).
Ecosystem-based management is one of the basic supports in the theoretical application of MPAs as management tools for the marine environment (Browman et al., 2004; Halpern et al., 2010; Long et al., 2015). It implies a broad and integrative approach scale, where all ecosystem components and processes are considered, including those in which people intervene. The success of an MPA must, therefore, be able to be verified at the ecosystem level. However, it is not easy either to find indicators, cause-effect relationships or conceptual schemes, nor to assess the effectiveness of MPAs at this scale where multiple factors and variables fluctuate in space and time.
The key ecosystem-based management principles that are most frequently acknowledged in the literature, according to Long et al. (2015) review, include “adaptive management,” “stakeholder involvement,” and “recognize coupled social-ecological systems.” This shows that beyond scientific knowledge of ecosystem processes, the success of MPAs also implies the recognition by local communities (Di Franco et al., 2016), in particular those whose activities are linked to the protected area, the implication of the involved administrations that must make an effort to provide resources for an effective protection, and a global awareness that understands that protection is a transversal tool that must be coordinated and affects and is affected by the many uses and activities that take place in a certain area.
One of the most effective ways to accept, value, and promote the protection of a space is to understand which elements and processes are involved and what the consequences of that protection are. These are couched in terms of benefits, both economic and not, that it produces directly or indirectly to human societies.
Knowledge of goods and benefits, supported by the physical and ecological processes and the existing biodiversity in an area, will allow us to justify and better fulfill the objectives of protection and guarantee it within a framework of medium and long-term sustainability. From the maintenance of species of fishing interest to the maintenance of the connectivity between populations and of genetic heterogeneity and diversity, there are many ecosystem services and human benefits provided by healthy marine habitats (Haines et al., 2018). However, not all of these goods, services and societal benefits are well-known, and the information is dispersed in a multitude of works focused on particular case studies that refer mainly to the effects of protection on some specific aspect of the biology of a single species or assemblage. Furthermore, the bibliography that reviews them together, and put them in the context of the protection of a coastal area, is still scarce (Potts et al., 2014; Van den Belt and Cole, 2014; Saunders et al., 2015; Olander et al., 2018; Schratzberger et al., 2019; Armoškaite et al., 2020).
The first problem to face is the existing confusion and inconsistency on what means and constitutes an ecosystem service (Olander et al., 2018). Frequently “services,” “goods,” or “benefits” are used as being synonyms, and in the literature there is such a proliferation of terms, like abiotic or biotic provisioning, service capacity or demand, human related activities, ecological process or products from ecological systems, that it is not always easy to distinguish between ecosystem services and societal benefits (De Groot et al., 2002, 2010; Fisher et al., 2007; Fisher and Turner, 2008; Haines-Young and Potschin, 2013, 2016; Villamagna et al., 2013; La Notte et al., 2017; Newton et al., 2018; Pérez-Ruzafa et al., 2019). In this way there is even confusion and inconsistency over what is meant by “ecosystem service” indicator (Johnston et al., 2012; Bauer and Johnston, 2013; Boyd and Krupnick, 2013; Boyd et al., 2016). Many of the definitions are tautological and include the defined term in the definition, while others include the terms that are intended to differentiate (such as benefit) in the definition of the other (service) (Table 1). Other works define new concepts like Ecosystem Service capacity as an ecosystem's potential to deliver services based on biophysical properties, social conditions, and ecological functions, or differentiate between final or end services from intermediate services, without defining what they specifically consider an ecosystem service (Villamagna et al., 2013). Although some works have contributed to clarify the framework (e.g., Mace et al., 2012), they also use the term benefits as definition of services. Therefore, as some of the more recent main reviews and assessments on the topic recognize, the problem of categorizing ecosystems services is still not completely resolved (Haines-Young and Potschin, 2016).
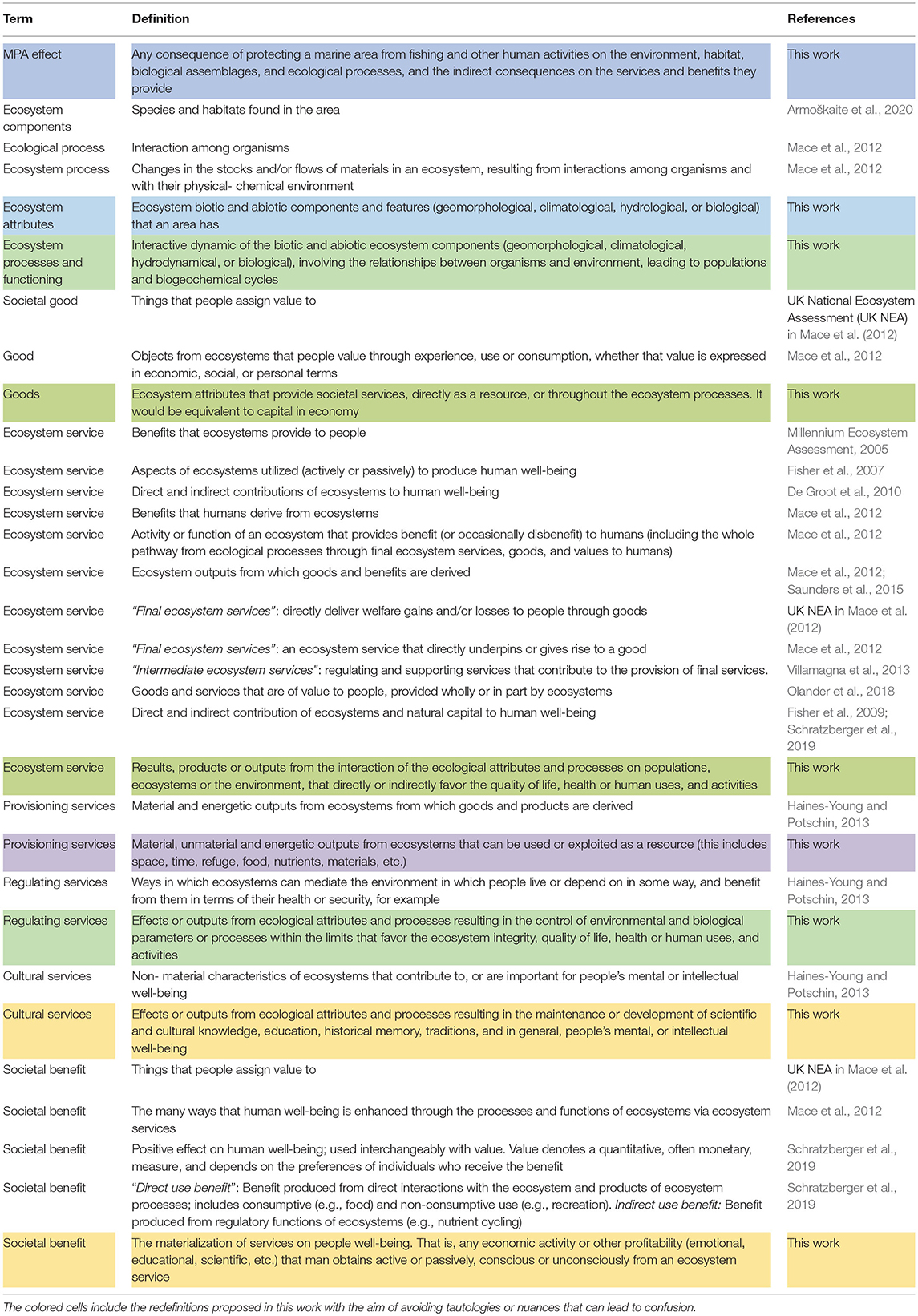
Table 1. Terms and definitions related to the effects of protection, ecosystem services, and societal benefits provided by marine protected areas (MPAs) used in the literature.
This work aims to review the knowledge and evidences of the status on the goods, ecosystem services and benefits of MPAs for the society, framing them in context of the processes and biodiversity that sustain them and highlighting some of the main threats that they face. This work has been carried out based on an extensive bibliographic review and on the data and conclusions of numerous previous studies by the authors themselves.
Materials and Methods
Terms and Definitions
Before analyzing the state of knowledge and the evidence of ecosystem services and benefits produced by MPAs, we have defined the main terms used (protection effect, ecosystem attribute, ecosystem process, ecosystem good, ecosystem service, provisioning, regulating and cultural services, and societal benefit) in the context of main previous definitions and trying to solve some ambiguities and tautologies found in them (Table 1).
Perhaps the main differences between our definitions and previous ones lie in the nuance of the potential vs. the realization of the benefit. An ecosystem service is the aptitude or qualification that an ecosystem has, or develops, as a result of the interactions and processes that take place between their biological and environmental components and features that directly or indirectly can favor the quality of life, health or human uses and activities. A societal benefit is the effect that it produces throughout any economic activity or other profitability (well-being, emotional, educational, scientific, saving costs, preventing diseases or increasing health, etc.) that man obtains, through an active use or passive enjoyment, conscious or unconsciously from an ecosystem service or quality.
Accordingly, we have differentiated the ecological attributes and processes that a coastal area has or develops and are enhanced by the protection constituting its capital or goods, from the services they provide, and these from the societal benefits that they may manifest (Figure 1).
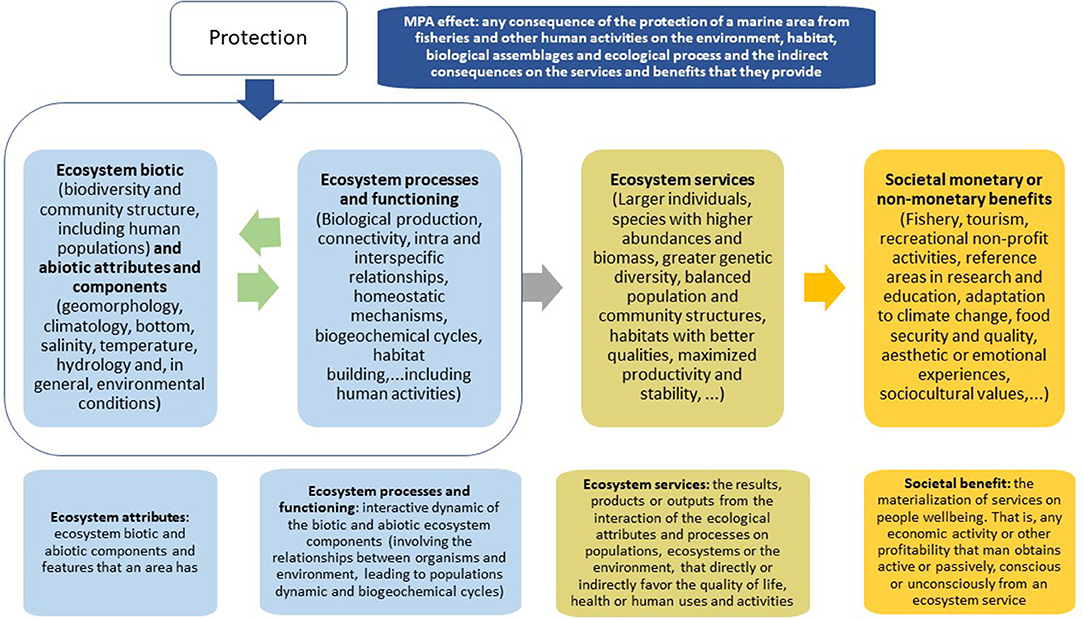
Figure 1. Basic scheme that shows the sequence of relationships between the components of an ecosystem that condition its functioning, the services they provide and the benefits that humans can obtain in the case of an MPA. The definitions of MPA effect, ecosystem attribute, process and functioning, ecosystem service, and societal benefit are included. A clear difference is established between the services that an ecosystem offers and the benefits, economic or non-economic, material or not, that man obtains.
Literature Review
A comprehensive review of the scientific literature on the effects of protection in MPAs and the ecosystem services and societal benefits that they provide have been conducted on peer-reviewed articles and, less frequently, books and technical reports, academic theses or gray literature produced in the last 20 years.
Following the scheme of Moher et al. (2009), records to be analyzed were identified through a search in all databases of the Web of Science (WoS), entering the keywords “marine protected area + effect” (n = 3,910), “marine protected area + ecosystem service” (n = 851) and “marine protected area + benefit” (n = 1,722) in the TOPIC field, for a period between January 2000 to November 2020. The searches produced a total of n = 5,511 articles after removing duplicates (Figure 2).
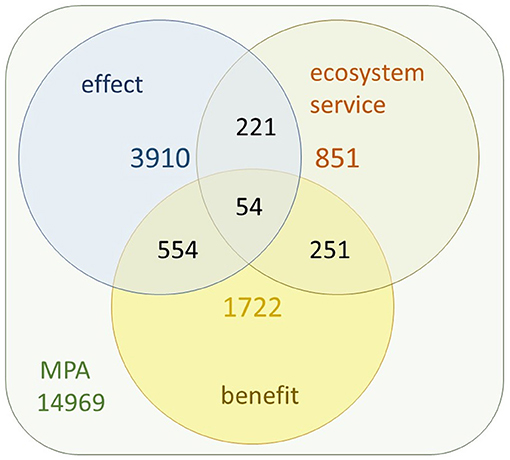
Figure 2. Number of articles identified searching the concepts “marine protected area + effect,” “marine protected area + ecosystem service,” and “marine protected area + benefit” and their respective intersections in the TOPIC field of the Web of Science database.
A list of 248 different terms or variations of them (like habitat, habitat complexity, key habitat, individual size, size at maturity, abundance, species richness, genetic, genetic diversity, genetic structure, life history, food web, top down, resilience, etc.) was selected according to our experience and with a preliminary analysis of the main review papers on the effects of protection in marine areas (88 terms or variations of them) or expected ecosystem services (108 terms) and benefits (65 terms). These terms were used for refined additional searches in the respective references list (from the previous step) obtained for effects, services and benefits. Only 17 terms or variations were common to the different searches. To ensure a direct relationship between some terms in the works, and not only that the two terms appear independently cited, we have used expressions such as protected near/4 ecosyst* in the search. In other words, the separation in the text between protected and ecosystem was not greater than four words. We refer to these searches in the results and discussion as closely related terms. The complete list of terms and their variations, the corresponding search criteria and the total number of articles found in each case can be found in Supplementary Material Table S1.
As some works not published in the journals included in WoS (including the gray bibliography) may contain interesting evidences for a qualitative review of the main protection effects, services and benefits, additional records were identified through a Google search (“marine protected area effect,” “marine protected area benefit,” and “marine protected area ecosystem service”), considering the first five pages of the results obtained in each search (n = 146). These works have not been included in the quantitative statistical analysis of the WoS searches.
The information has been supplemented with data from the different authors' research teams, based on the available information in the eight RESERVEBENEFIT (BiodivERsA3-2015-21 Cofund European project) MPA case studies in the western Mediterranean (Cerbère-Banyuls-sur-Mer, Cap de Creus, Llevant de Mallorca-Cala Rajada, Islas Columbretes, Nord de Menorca, Cabo de Palos-Islas Hormigas, Tabarca, and Cabo de Gata) (Figure 3, Table 2), including scientific articles, technical reports, PhDs and others (n = 62). All these case studies are effective MPAs with more than 13 years of functioning, with management plans, enforcement and scientific surveillance. RESERVEBENEFIT case studies' data and expert knowledge of those responsible for their surveillance were also used for a comparative analysis with the results of the bibliographic search.
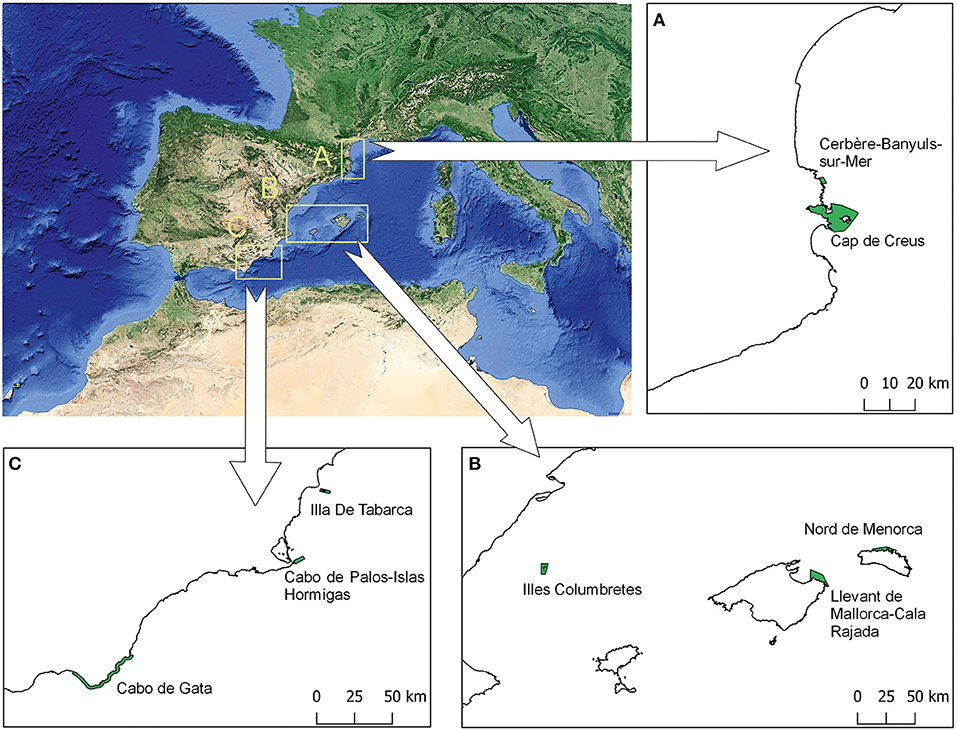
Figure 3. Location of the eight western Mediterranean marine protected areas, all of them RESERVEBENEFIT project case studies, that have contributed to the results of this study.
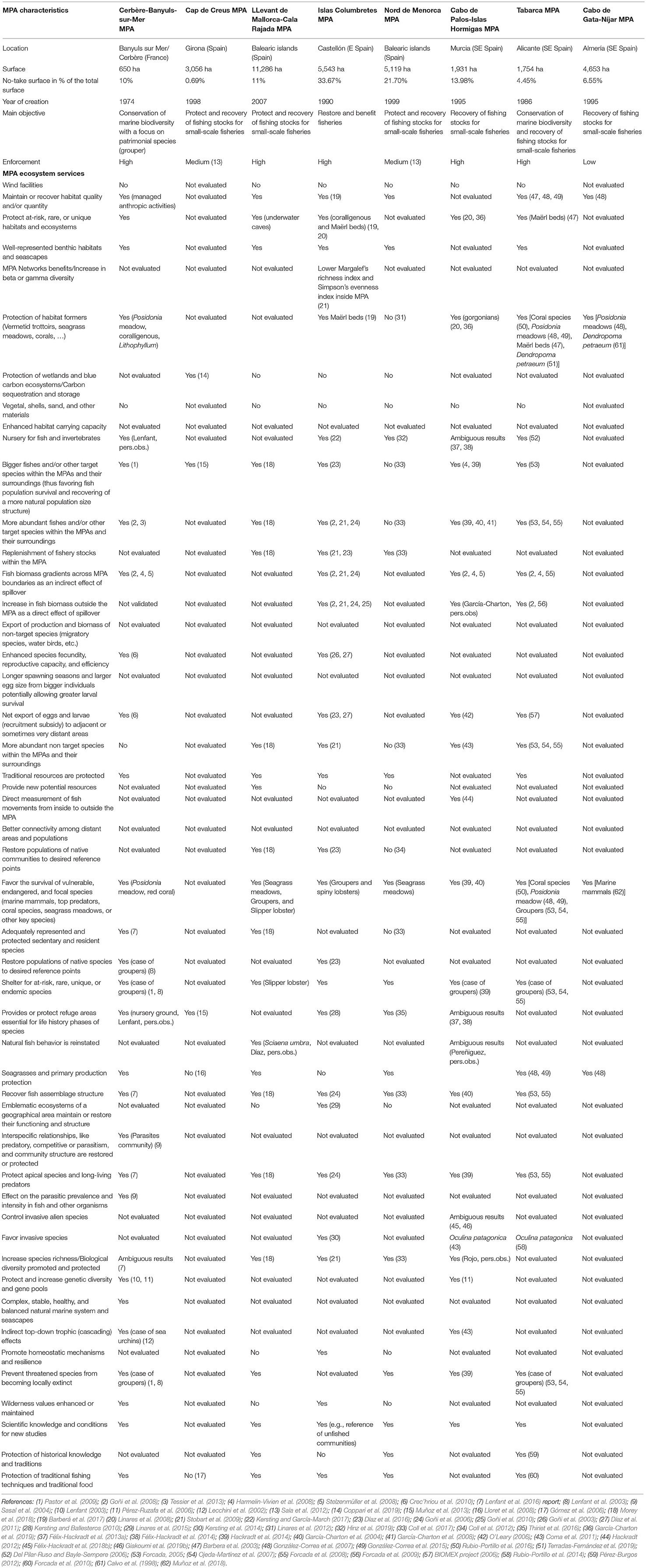
Table 2. Attributes, processes, ecosystem services, and societal benefits associated with Marine Protected Areas in eight MPAs in the western Mediterranean.
After duplicates were eliminated, a total of n = 5,646 works were screened through their titles and/or abstracts, and we then excluded those that were too general or not adjusted to the review topic records, or where the data did not seem sufficiently supported or seemed to be outside the scope of the review. The final number of studies selected for the qualitative analyses, discussion and to build the synthesis table (Table 3) and the conceptual cause-effect diagram (Figure 8) of this work was n = 268.
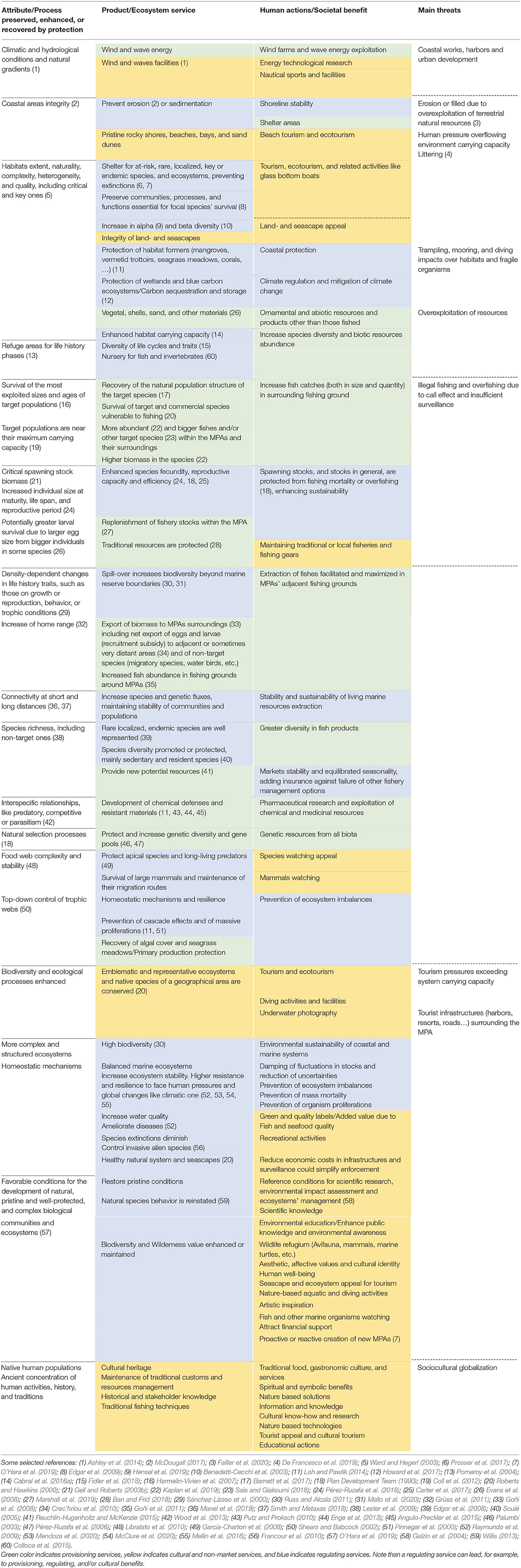
Table 3. Processes, ecosystem services, and societal benefits associated with marine protected areas.
For the organization and classification of the ecosystem services provided by MPAs in the synthesis table (Table 3), we have based on the general scheme of Common International Classification of Ecosystem Services proposed by the EEA (Haines-Young and Potschin, 2013), progressively adapting it according to the bibliography consulted and our own information.
To compare the effects and services observed in the RESERVEBENEFIT MPAs with the results of the bibliographic review, we built a matrix including the effects and services for each MPA, the average score for all of them and the results obtained in the WoS review. Data were expressed as a percentage of total cases (total number of MPAs in the case study, Table 2, or total citations found in the respective searches MPA+services or MPA+effects recorded in Supplementary Table 1). The data were transformed with the square root and analyzed with Principal Components Analysis (PCA) using Canoco v.5.
Results and Discussion
Among the 14,969 WoS articles including marine protected areas, 3,910 refer to effects, 851 to ecosystem services and 1,722 to benefits. Only 54 of them include all three terms. The interactions between the other combinations of terms can be seen in Figure 2.
The protection of marine areas generates a series of effects (Figure 4) and provides a wide spectrum of valuable ecosystem services, many of them reflected in human welfare and benefits (Table 3, Figure 5). However, the identification and classification of these services as well as their verification and assessment, and the cause-effect relationships between marine protection, ecological processes and benefits are a complex issue, and literature with this focus remains scarce (Leenhardt et al., 2015; Pérez-Ruzafa et al., 2019). It is more difficult if we take into account that a process that generates a service can, at the same time, produce effects on the ecosystem components, changing their properties, and/or facilitating an additional service.
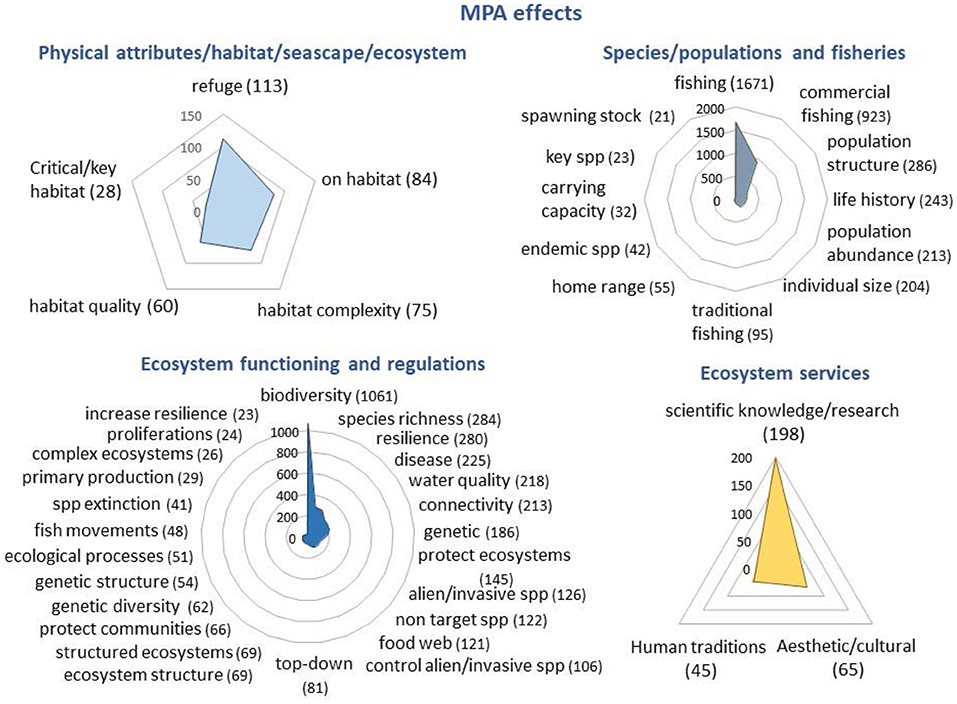
Figure 4. Graphic representation of the occurrence (number of articles) of main protection effects in MPAs according to the articles recorded in the Web of Science. Only search terms exceeding 1% of the total number of articles including the term “effect” are represented.
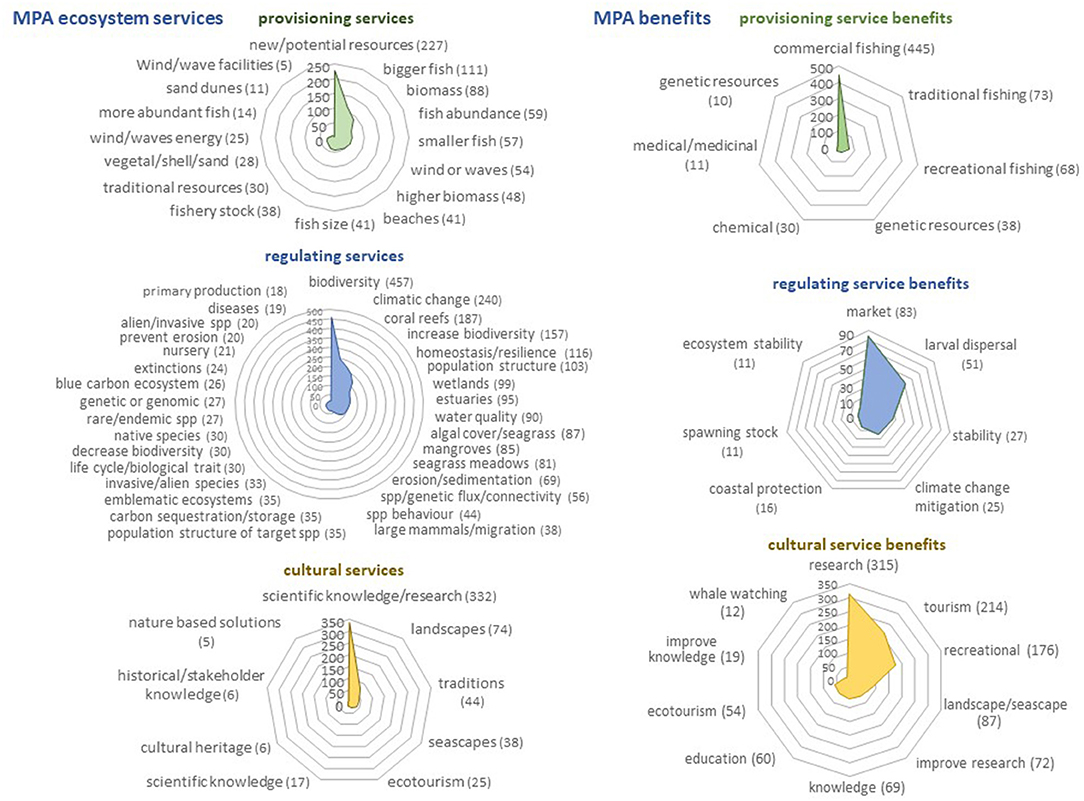
Figure 5. Graphic representation of the occurrence (number of articles) of main Ecosystem services and societal benefits produced by MPAs according to the articles recorded in the Web of Science. Only search terms exceeding 1% of the total number of articles including the terms “service” and “benefit”, respectively, are represented.
Furthermore, given the diversity of management measures applied in the numerous existing protection designations, the different efficiency of each one of them, the particularity of the environmental and connectivity conditions at each site and the different types and intensity of human pressure in the areas adjacent to reserves (Mazaris et al., 2019; Nickols et al., 2019), among other sources of variability (Pendleton et al., 2018), it is not possible to generalize for all protection designations or situations. Therefore, it cannot be guaranteed that the mere application of the term “protected” will imply that a certain good or service will be improved or produced, and many empirical studies on protection, ecosystem services and societal benefits relationships are needed until a clear and consistent doctrine on these issues is achieved. The importance of spatial and temporal scales is crucial. The size and location of the MPAs will condition the type and magnitude of the effects obtained (Claudet et al., 2008), and these effects also manifest themselves at different spatial scales, from the most local (few km) to large (thousands of km) scales. Young MPAs in many cases will not have had the possibility to yet fully develop all their response mechanisms (Vandeperre et al., 2011), and it should be also considered that ineffective protection could maintain this initial status regardless of the actual time elapsed.
Ecosystem services are hence inconstant in space and time, as are the societal benefits obtained from them. Assessing these dynamics and this heterogeneity constitutes one of the main challenges of future studies.
The main ecosystem services of MPAs and their benefits for man resulting from this review have been compiled in Table 3 and the most commonly documented ones are briefly discussed below.
Main Effects, Ecosystem Services, and Benefits Produced by MPAS
The frequency with which the different terms appear in the works collected in WoS are shown in Supplementary Table 1. Figures 4, 5 represent the number of papers in which the most frequently used terms appear. Fishing (42.7%), biodiversity (27.1%), population structure (7.3%), species richness (7.3%), resilience (7.2%), life history (6.2%), disease (5.8%), water quality (5.6%), connectivity (5.5%), individual size (5.2%), genetic (4.8%), protected ecosystems (3.7%), alien or invasive species (3.2%), food web (3.1%) or the habitat as a refuge (2.9%) are the most frequent between MPA effects. Biodiversity (53.6%), scientific knowledge or research (38.9%), climate change (28.1%), new potential resources (26.6%), coral (24.9%), increase diversity (18.4%), homeostasis and resilience (13.6%), population structure (12%), bigger fish (13%), or biomass (10,3%) are the most recorded in papers including services. The most frequently cited ecosystems linked to services are coral reefs (21.9%), wetlands (11.6%), estuaries (11.1%), mangroves (10%) and seagrass meadows (9.5%). Finally, commercial fishing (25.8%), research (18.3%), and tourism (12.4%) are the most frequent terms among benefits. In general, even for some effects, services or benefits usually considered important and assumed to be clearly associated with MPAs, the number of articles that mention them is very low and very few exceed 5% of the published works. As an example, we can mention effects like export eggs/larvae (0.2%), cascade-effect (0.1%), prevent extinction (0.08%) or invasive species (0.03%), services like spill-over (1.4%), species fecundity (0.47%), at risk or rare species (0,23%), biomass export (0.12%), benefits like traditional fishing gears (0.25%), spiritual, artistic or aesthetic (<0.3%), cultural or historical knowledge (0%). This suggests that there is still much experimental work to be carried out in marine reserves to be able to generalize and quantify the provision of these services.
MPAs Maintain or Recover Habitat Quality and/or Quantity
This is one of the effects most attributed to the protection of marine areas. However, its frequency in the scientific literature depends on the habitat attributes considered. Effects “on the habitat” are present in 2.15% (84 articles) of the total of 3,910 articles that include “marine protected area + effect” key concepts. Complexity is highlighted in 1.9% (75) and the term habitat quality in 1.5% (60), while naturalness does not appear at all.
MPAs contribute to improving or, at least, avoiding the degradation of two fundamental aspects of the habitat, spatial heterogeneity and structural complexity (Stephenson et al., 2019). The first is the mosaic of types of habitats that can occur on the seabed (sandy or muddy bottoms, rocky bottoms, macrophyte meadows) and constitutes the horizontal dimension of space (García-Charton and Pérez-Ruzafa, 1998). Structural complexity refers to the vertical dimension and is related to roughness, the presence of large blocks, medium-sized blocks, and stones, changes in slope or depth differences in a given space (García-Charton and Pérez-Ruzafa, 2001). Both aspects contribute to increasing the carrying capacity of the ecosystem including multiple characteristics of physical structure, such as the variation of elements or the size of cavities (Tokeshi and Arakaki, 2012) providing refuge, food, and space. And, at the same time, both are complementary; while heterogeneity favors species richness and diversity (especially if there is some equality in the extension of the different types), vertical complexity is mainly associated with the abundance of individuals (García-Charton and Pérez-Ruzafa, 1998; Willis et al., 2005; Forcada et al., 2009; Tokeshi and Arakaki, 2012; St.Pierre and Kovalenko, 2014). The effects of habitat structure manifest at different spatial scales and have effects upon different faunal guilds (nektonic fish, benthic fish, macroinvertebrates, meiofauna.) according to the scale at which they are considered. Thus, for example, the dominance of medium-sized blocks (between 1 and 2 m in diameter) favors the abundance and richness of benthic or demersal fish and starfish or urchins, while the abundance of stones (<1 m in diameter) favors ophiuroids, small sea urchins, or gobiids (García-Charton and Pérez-Ruzafa, 1998; Entrambasaguas et al., 2008). As a result, habitat structure is a determinant factor for processes like settlement, recruitment, growth, depredation, or successful reproduction (Botsford, 2001; García-Charton and Pérez-Ruzafa, 2001) and can determine the different effectivity of protection on different species (Forcada et al., 2009).
MPAs may avoid the degradation and homogenization of the habitat produced by human uses like trawling, dredging or pumping sediments, and, on the other hand, they favor the recovery of species that are habitat builders. The effects of protection on the complexity and heterogeneity of the habitat are variable and will take place at different scales depending on the nature of the substrate (rocky vs. soft bottoms, geologic vs. biogenic structures). Once the aggressions have ceased, while on rocky bottoms the protection can hardly recover the structure of blocks and rocks, however, both in hard and soft bottoms, hydrodynamism can act in restoring the sedimentary dynamic, and biological structures can recover their natural growth. There are a large number of works that analyze the effects of trawling on communities and the nature of the sediments (Kaiser et al., 2002; Thrush and Dayton, 2002; Ramalho et al., 2020; Silveira et al., 2020). Stephenson et al. (2019) established a conceptual relationship between disturbance, loss of habitat heterogeneity and consequences on ecological functioning and ecosystem services through the loss of substrate burrowers and the role that they play on the biogeochemical process and nutrient and oxygen fluxes. On the other hand, protection can also enhance temporal stability of habitat characteristics (Fraschetti et al., 2013). On the coast, protecting beach habitats from human disturbance restore seabirds breeding (Lafferty et al., 2006). At regional biogeographic scale, the protection of habitats (mainly the underrepresented) in a network of representative MPAs is considered essential to sustain global biodiversity (Abdulla et al., 2008).
In this context, the protection of biogenic habitats such as mangroves, coral reefs, coralligenous, maërl, and gorgonian biocenosis (D'Onghia et al., 2010; Costanzo et al., 2021), vermetid platforms or “trottoirs” (Chemello et al., 2000), or seagrass meadows (Waycott et al., 2009), take on all their importance.
However, despite the importance of habitat recognized in the reviewed works, the analysis of its explicit link with ecosystem services or benefits is scarce. Most papers that discuss the effects of protection mention the words service or benefit in a general way, or they refer benefits to particular species or attributes, but not to human societies in the sense of our review. This may indicate that ecological relationships and processes in marine reserves are not always studied in the explicit context of the ecosystem services and benefits they produce and their quantification, although they would undoubtedly be implicit. References to habitats as services are scarce, although there are differences in terms like “protect beaches” (0.35%) or “protect dunes” (0.23%) and “protect rocky shores” (0%). “Rocky shore” appears more frequently without the term “protect” (0.58%), but it is often referred to generically. Although only 0.11% of the studies appear under the term “habitat formers,” some of the ecosystems most mentioned, within the articles that mention MPA services, are represented by habitat-forming species such as the above mentioned “coral reefs” (21.9%), “mangroves” (9.96%), or “seagrass meadows” (9.5%). Vermetid trottoirs, conformed by reef-building vermetid gastropods, considered an endangered key intertidal habitat of the warmest areas of the Mediterranean Sea and modulators of coastal geomorphological processes (Chemello et al., 2000; Naylor and Viles, 2002; Templado et al., 2016), have not turned up in searches.
Some aspects of the habitat, like the complexity of geological hard bottoms, are not possible to recover simply with protection and would probably require active restoration measures, but as mentioned, MPAs can contribute to improve the quality of the habitats they protect and restore other degraded attributes. Habitat extent and quality, which determine population carrying capacity (García-Charton and Pérez-Ruzafa, 1998, 1999), are good criteria for protection strategies and can help achieve both conservation and fisheries objectives (Cabral et al., 2016a).
Given the generalized degradation and increasing pressures on coastal ecosystems, and the inherent asymmetry of ecological succession, in which recovery processes are always slower than degradation changes (Margalef, 1997), MPAs should be selected looking for those habitats with a higher quality. In this way, these will serve as a reservoir and can help to recover those that are now more degraded. In a second step it will be possible to start also protecting the degraded ones so that they can be recovered as well.
Although the response of the different species can manifest at different spatio-temporal scales, habitat changes have immediate consequences on the functioning of the communities and the ecosystem as a whole. Many expected processes and services depend directly on the habitat meeting minimum requirements, but, at the same time, the habitat will not be able to provide them if there are other external limiting factors, e.g., inadequate connectivity (Andrello et al., 2017; Magris et al., 2018) or human and environmental pressures in the surrounding areas, that negatively affect the state of populations or communities. This leads Cabral et al. (2016a) to state that “protecting sites on the basis of sources, sinks, or other centrality measures of connectivity becomes optimal only in limited situations, that is, when larval production is not dependent on the adult population” and that “carrying capacity is crucial in the evolution of larval metapopulations” (Cabral et al., 2016a). MPAs could protect and restore key habitats that are required for vulnerable, endangered, and focal species' survival, including commercial species (Roberts and Hawkins, 2000; Pomeroy et al., 2004). This includes refuge areas or those that could be essential for life history phases of species (Pomeroy et al., 2004). Some examples focus on priority habitats such as Posidonia oceanica in the Mediterranean (González-Correa et al., 2007, 2015; Fraschetti et al., 2013; Padiglia et al., 2018). However, protection is not always effective and may be insufficient for habitat restoration. Montefalcone et al. (2009) detected poor health and degradation of P. oceanica meadows on the Liguria coast (NW Mediterranean) related to human impacts, such as activities producing turbidity, that are not mitigated by protection measures.
Furthermore, the priority could be given to the protection of emblematic, vulnerable or underrepresented habitats, especially those that have a restricted distribution (Roberts and Hawkins, 2000; Ward and Hegerl, 2003). Abdulla et al. (2008) paid attention to the habitat heterogeneity of the Mediterranean basin and underlined that many areas and habitats are still underrepresented in MPAs, in particular, in the southern and eastern parts that host ecological habitats of high conservation value, including those being home to highly mobile marine vertebrates, sessile invertebrates or seagrass meadows of great interest.
MPAs also protect habitats from damage caused by fishing gear, or other detrimental effects due to exploitative or non-exploitative human uses (like the impact of divers or mooring), and stimulate their recovery (Ward et al., 2001; Reuchlin-Hugenholtz and McKenzie, 2015), However, it is important to highlight that recreational fishing or other traditional and local artisanal fisheries are frequently allowed within some zones of MPAs, like in the buffer zones or in the case of fish boxes, in which cases the vigilance and fullfilment of established rules are highly important, and bearing in mind that protection based exclusively on regulatory measures is often inefficient and that such measures can only really be effective if they are associated to a no-take zone (Pérez-Ruzafa et al., 2017).
Protecting habitats is also related to the term benefit (2.85%), generally referring to the enhancement of species like fish (Rosenberg et al., 2000; D'Onghia et al., 2010), seabirds (Lafferty et al., 2006; Burdon et al., 2017) or to the improvement of functional connectivity (Turgeon et al., 2010), and indirectly to fisheries, recreation or conservation, respectively. In the case of birds, a simple barrier that reduces the disturbance and impact of trampling on nesting areas can mean a significant increase in the population density and the existence of active breeding (Lafferty et al., 2006).
MPAs Contribute to Maintain Adequately Represented and Protected Resident and/or Focal Species and Communities
3.71% (145 articles) of the studies that contemplate these effects include closely related the terms protected and ecosystems. In addition, endemic (1.07%), at risk (0.49%), key (0.59%), focal (0.23%), or rare (0.18%) species are represented in the results.
Regarding ecosystem services, there are more mentions for endemic species (0.82%), key species (0.35%), or rare or at-risk species (0.23% each term), and focal species are only listed as such in 0.11%. The number of publications related to unique or rare ecosystems (0.82%) or ecosystems and communities at risk (0.23 and 0.11%, respectively) are in the same order.
There is evidence that MPAs favor the survival and restoration or maintenance of populations of native species and communities at desired reference points, especially those very vulnerable to the effects of fishing and, many times, at sites and/or life history stages where they become vulnerable (Roberts and Hawkins, 2000; Pomeroy et al., 2004).
It has generally been assumed that the effectiveness of MPAs in protecting sedentary and resident species is greater than in protecting those that are highly mobile and migratory (Soulé, 2005). However, as the meta-analysis carried out by Claudet et al. (2010a) on 40 data sets from 12 European marine reserves concluded, contrary to previous theoretical approaches, also mobile species with wide home ranges can benefit from protection just as much as the sedentary ones. This has been also confirmed in fish species that show high individual variability in residence and in the spatial range of their movements (Maggs et al., 2013; Andrzejaczek et al., 2020). Some authors have suggested that this may be due to a modification of the behavior of fish within protected areas, with a higher movement rate in the fishing grounds than in the MPA (Jiao et al., 2018).
The effects of the protection are especially important in the case of apical species and large long-lived predators vulnerable to fishing whose protection allows fish assemblage structure to be recovered (García-Charton et al., 2008), at the same time that natural fish behavior is reinstated (Willis, 2013). In the French Mediterranean, grouper (Epinephelus marginatus) has benefited from the protection of MPAs, which became true refuges when the species was overexploited. Since the 2000s, in all French MPAs, an almost constant increase in their numbers has been observed (e.g. Cerbère-Banyuls-sur-Mer MPA, Lenfant et al., 2003). The protection of groupers has also been verified in the Llevant de Majorca-Cala Rajada, Columbretes islands or Cabo de Palos MPAs (Hackradt et al., 2014; Table 2). In the case of silky sharks (Carcharhinus falciformis) in the Indian Ocean, despite their high mobility, the large proportion of juveniles, coupled with the high site fidelity observed in some individuals, in agreement with historical fishery data, suggests that MPAs could provide considerable conservation benefits for the species, particularly during early life history stages (Curnick et al., 2020).
The establishment of MPAs could be based on the identification of areas of interest for species of importance for conservation or focal biodiversity targets (Hooker et al., 2011). In this sense, beyond the harvested species, marine mammals (Gormley et al., 2012), top predators (Hooker et al., 2011), coral reefs (Topor et al., 2018), or other key species could benefit from the restoration of ecological balances in protected areas.
Marine biodiversity risks are spatially heterogeneous resulting from a combination of geographic, taxonomical and human impact factors, where MPAs play a definitive role in protecting at-risk and pristine species and ecosystems and in preventing extinctions (O'Hara et al., 2019) although, as mentioned above, protection may not be sufficient if other measures are not also adopted.
Species protected by MPAs may also be rare, localized or endemic species (Pomeroy et al., 2004). A good example can be the Galapagos Marine Reserve (Ecuador) where 41 threatened marine species, 25 of them being endemic fish, mollusk, crustacean, echinoderm, coral or macroalgal species, find shelter and protection, showing significant increases in abundance and distribution inside the MPA (Edgar et al., 2008). This service of protection must be underlined, as rare species often are the first to disappear under human-induced collapses of populations and species, and they play a crucial role in the maintenance of ecological processes increasing the potential breadth of functions provided by ecosystems (Mouillot et al., 2013).
Facilitate Adaptation to Climate Change of Marine Ecosystems
Climate change is a threat to oceans and marine biodiversity, increasing the challenge for marine conservation (Rilov et al., 2019). Global change or climate change together are among the terms that are most frequently mentioned in works related to MPA ecosystem services (28.14%). Different authors agree that MPAs are not a specific tool against warming, but they can provide resilience and contribute to healthier ecosystems in the context of the global role of the ocean (Roberts et al., 2017; Sala and Giakoumi, 2018). Managed marine ecosystems could reduce losses in carbon sequestration and storage, could buffer against uncertainty in management and/or inadequate management, and could also buffer environmental fluctuations, directional changes or extreme events (Simard et al., 2016; Roberts et al., 2017). They should work from a precautionary principle that could be extended to coastal and global management, help enforce marine conservation, reduce biodiversity loss, help safeguard critical ecological processes that support planetary life and improve recovery prospects in a scenario of increasing greenhouse gas emissions (Roberts et al., 2017; Rilov et al., 2019).
Marine ecosystems, especially coastal wetlands and species such as seagrasses and mangroves, are important carbon sinks (Russi et al., 2016). However, due to human uses and activities, these ecosystems are being degraded, undermining their role as sinks and running the risk of becoming sources of carbon dioxide emissions (Russi et al., 2016). Some studies provide data on loss rates, for example in coastal wetlands (Barbier et al., 2011) or kelp forests (Ling et al., 2009), and show evidence of their connection to the lack of protection. One major aim of many MPAs is the reduction of coastal ecosystem loss, the protection of emblematic ecosystems and wetlands and, in this way, the protection of the so-called “blue carbon ecosystems,” which have become a great priority in marine management (Howard et al., 2017; Roberts et al., 2017). As evidence of how MPAs can contribute to recover key blue carbon ecosystems, Mangos and Claudot (2013) estimated that Posidonia meadows would increase annually between 1 and 2% in an increasing-protection scenario due to the improvement of marine environmental quality.
MPAs Favor Larger Individuals
Fishery pressure is normally highest on the largest individuals of a population. Consequently, the increase in larger fish is one of the most frequently observed MPA effects, and this mainly translates into bigger individuals for traditional or local fisheries allowed in buffer areas (Pérez-Ruzafa et al., 2017) or for fisheries in their surroundings when biomass export occurs. Further than this immediate effect, the main effect of reducing fishing mortality is increasing life span. When life expectancy increases, the growth rates of individuals tend to decrease, but as the age of first maturity is delayed, the period dedicated to growth also lengthens and the individuals reach larger sizes before reproduction (Pérez-Ruzafa et al., 2018) canceling the effect of negative fishing selection (Plan Development Team, 1990). These effects related to the size of organisms, mainly fish, are mentioned in numerous studies and under different aspects, both in studies on protection effects (individual size 5.22%, size at maturity 0.33%), as in those referring to ecosystem services (bigger fish 13%, fish size 4.81%).
However, as fish mobility and home range may increase with body size and age (Grüss et al., 2011), Alemany et al. (2013) predicted a larger positive MPA-effect on juvenile size rather than on adults, particularly if nursery grounds are located inside their boundaries. In fact, they observed an increase in the mean total length of juveniles of 11% inside the Patagonian protected area studied. This agrees with the results of Edgar and Barrett (1997) for Tasmanian coastal MPAs, where authors observed a significant general increase in mean size of animals of 10%, and Willis et al. (2003) for snapper, Pagrus auratus, in northern New Zealand marine reserves where the individuals larger than the minimum legal size were estimated to be 14 times more abundant in protected than in fished areas. Furthermore, in his revision of 73 empirical MPA studies, Halpern (2003) found that 83% of MPAs had larger individuals inside than outside the MPA for carnivorous fish, and 89% had larger plankton and benthic invertebrate eating fish.
MPA-driven increases in individual size have been detected in numerous other studies (Sánchez-Lizaso et al., 2000; Stobart et al., 2009; Chirico et al., 2017; Rolim et al., 2019; see Table 2) not only for fish but also for other groups of organisms such as shellfish (Edgar and Barrett, 1999), sea urchins (Castilla and Pino, 1996), lobsters (MacDiarmid and Breen, 1993; Kelly et al., 2000; Díaz et al., 2011, 2016) as well as other invertebrates and algae (Lester et al., 2009; Fenberg et al., 2012).
However, this effect is not always evident as external fishing pressure may decrease average sizes mainly in the case of adults if, for example, the MPA is relatively small for the home range of the target species (Alemany et al., 2013), or due to the density-dependent growth rates of some species.
Increase in Species Abundance and Biomass Favor the Exportation of Fishable Biomass Out of the MPAs
When managed appropriately, MPAs protect ecosystem processes and components, providing areas where species are able to reproduce, spawn and grow to their adult size in a relatively undisturbed environment. The increase in abundance and/or biomass of fish or other exploited species inside MPAs are among their most proven effects and have been widely used as indicators of their effectiveness (Ward et al., 2001; Goñi et al., 2006, 2008, 2010; García-Charton et al., 2008; Planes et al., 2008; Stobart et al., 2009; Sciberras et al., 2013; Sala and Giakoumi, 2018; Rolim et al., 2019). This is reflected in the number of works that mention the effects of protection on abundance (population abundance 5.45%) or those about services associated with it (fish abundance 6.92%), with the most frequent studies highlighting its increase (more abundant fish 1.64%) without missing those that highlight its decrease in some species (less abundant fish 0.59%). The increase in size, mentioned above, and in abundance translate into an increase in biomass also collected with high frequency among ecosystem services (biomass 10.32%; higher biomass 5.63%), and as an increase in fish stocks (4.45%).
These effects have been proven for fish throughout the world (Gell and Roberts, 2003a,b; Willis et al., 2003; Williamson et al., 2004; Guidetti and Sala, 2007; Harmelin-Vivien et al., 2008; Goñi et al., 2011; Taylor et al., 2012; Giakoumi et al., 2017). Moreover, other harvested populations may also show these benefits, as proven, for example, in Palinurus elephas (Goñi et al., 2001), Homarus gammarus (Hoskin et al., 2011), Pectinidae (Howarth et al., 2011), Patella ferruginea (Meier, 2003), Mytilus galloprovincialis (Jacquet, 1999), and Paracentrotus lividus (Lecchini et al., 2002; Gianguzza et al., 2006; Pais et al., 2007). On numerous occasions the appearance of this positive effect is related with the surveillance and management enforcement of MPAs (Guidetti et al., 2008; Ceccherelli et al., 2011; Edgar et al., 2014; Di Franco et al., 2016; Haines et al., 2018).
But although, broadly, as mentioned above, total abundance and biomass are greater inside MPAs, and more clearly within no-take areas (Pérez-Ruzafa et al., 2017; Kaplan et al., 2019), many authors draw attention to the fact that this effect could be different for target or non-target species (Rolim et al., 2019), and also that the measurement of these parameters may be greatly affected by species mobility and by their fishing extraction in the surrounding areas of MPAs (Goñi et al., 2008), mainly if the integral reserve is not large enough (Pérez-Ruzafa et al., 2008a). Moreover, some studies on echinoderm species, and mainly on non-target ones, point out that a trophic cascade effect can cause a decrease in their abundance and biomass due to the increase in predation pressure following the recovery of predatory fish populations after protection (Sciberras et al., 2013).
Beyond these changes in the abundance of species, Blowes et al. (2020), studying 43 protected and 41 fished sites in the Mediterranean, have found that increased evenness (relative abundance of species) played the predominant role in changes in community structure in response to protection.
One of the most anticipated benefits of MPAs is that the effects of protection on populations and the size structure within the reserve translate into a net export of biomass to adjacent fishing grounds, through the spillover of larvae and adults (Plan Development Team, 1990; Rakitin and Kramer, 1996; Russ and Alcala, 1996; Kramer and Chapman, 1999; Lubchenco et al., 2003). However, although the biomass gradient between the interior and exterior of the reserves is usually evident, numerous studies have found difficulties to demonstrate such export (Harmelin-Vivien et al., 2008; Lester et al., 2009; Di Lorenzo et al., 2016). Much evidence comes from the behavior of fishermen and the benefits of fishing in the vicinity of reserves (McClanahan and Mangi, 2000; Murawski et al., 2005; Goñi et al., 2008, 2011; Stelzenmüller et al., 2008; Vandeperre et al., 2011; Cabral et al., 2016b) and others from the modeling of the displacement of the home-range of species across a density gradient (Gerber et al., 2003; Kellner et al., 2007; Pérez-Ruzafa et al., 2008a; Grüss, 2014). However, both, modeling and field studies agree that the spatial scale at which spillover from a marine reserve is effective is relatively short in the case of adult individuals, ranging, depending on species, from a few kilometers to <100 km (Harmelin-Vivien et al., 2008; Pérez-Ruzafa et al., 2008a).
Larval dispersal has been reported from 10 to 100 km for invertebrates and 50 to 200 km for fish (Palumbi, 2004). Le Port et al. (2017) reported a homogeneous increase in Chrysophrys auratus recruitment up to 40 km from the MPA. Other studies report even shorter average dispersal distance (<5–15 km), depending on the hydrographic and geomorphologic characteristics of the sites, highlighting self-recruitment as a common phenomenon (Jessopp and McAllen, 2007; D'Aloia et al., 2015; Green et al., 2015; Hopf et al., 2016).
MPAs Rehabilitate Exploited Species Natural Populations' Structure and Maintain or Restore Their Reproductive Potential
The imbalance in the population structure of sizes and sex is one of the negative effects of overfishing and, from the first calls to refocus fisheries management, its recovery has been one of the main objectives of the creation of marine reserves (Plan Development Team, 1990). This is reflected in the high number of studies on the effect of protection that include population structure (7.31%) and its consequences on the reproductive potential of the population (spawning stock 0.54%). The same occurs with those studies on ecosystem services where the concepts population structure (1.76%), population structure of target species (4.10%), and species fecundity or reproductive capacity (0.47%) stand out.
Barnett et al. (2017) quantified the extent of age truncation in 63 fished populations across five ocean regions, finding that the proportion of individuals in the oldest age classes decreased significantly in 79–97% of populations compared to historical or unfished values, respectively. MPAs avoid selective fishing pressure that normally acts on larger and older individuals. In this way, protection provides areas where fish are able to recover their population structure, and where it is possible to keep all the size/age classes of the populations represented in abundances that reflect natural conditions, which in turn protects the classes with a greater reproductive potential (Plan Development Team, 1990). These authors use the well-known example that one 61 cm red snapper (12.5 kg) potentially produces the same number of eggs (9,300,000) as 212 females at 42 cm (1.1 kg each) (Plan Development Team, 1990).
Improving and maintaining species' natural size and age structures within MPAs leads to a greater reproductive efficiency, which is normally associated with a larger size and/or age of the first reproduction (Berkeley et al., 2004), and can also reduce the probability of Allee effects (Drake and Kramer, 2011), especially in species with low mobility. Aalto et al. (2019) found that MPA networks reduced the risk of collapse following catastrophic events involving 75-90% mortality, while populations often continued to decline in the non-protected areas. MPAs or MPA networks are likely to be the most effective management tool for protecting old-growth age and thus stock productivity and stability (Hixon et al., 2014). Moreover, in the case of a successive hermaphrodite species such as the white seabream, Diplodus sargus, Lenfant (2003) showed that a marine reserve allows the age of sexual inversion to be preserved. The white seabream is a protandrous species, being male during its first reproduction, and becoming female only around 5/6 years old. Indeed, this author showed, on the basis of the age estimated from otoliths, that the females were younger outside the Cerbère/Banyuls natural marine reserve. This tends to show that protection allows for a later sex change and thus older and larger females in the marine reserve, ensuring greater reproductive potential within the protected area.
Evans et al. (2008) detected that the overall mean batch fecundity per unit area of Lutjanus carponotatus in different reserves in the Australian Great Barrier Reef was ~2.5 times greater than that in fished areas. This greater batch fecundity, together with longer spawning seasons and potentially greater larval survival due to larger egg size from bigger individuals, should benefit not only the no-take area itself but also surrounding fished areas (Evans et al., 2008). Of course, time from protection and species size and characteristics influence the importance of this effect.
Thus, protection of critical spawning stock biomass of species threatened by fishery-related depletion are expected to benefit fisheries via the net export of eggs and larvae (recruitment subsidy) from reserves to adjacent or sometimes very distant areas, depending on currents and seasonal climatic factors, helping to repopulate fishing territories (Gell and Roberts, 2003a,b; Ward and Hegerl, 2003).
Goñi et al. (2003) showed that the index of spawning potential of the lobster Palinurus elephas in the Western Mediterranean fishing areas was 5–18% that of the Columbretes marine reserve, closed to fishing for 13 years. Later on, Díaz et al. (2011) showed that this marine reserve, occupying 18% of the regional lobster fishing habitat, was responsible for over 80% of its egg production.
Carter et al. (2017), calculating the annual egg production per unit area for the commercially important Plectropomus leopardus, on fished and no-take reserves throughout the Great Barrier Reef in Australia, found that egg production within no-take areas was up to 152% greater compared to areas open to fishing, although they found areas with 56% less acting as recruitment sinks. Many factors as geographic region, hydrodynamic conditions, protection status, fish size or population density affect egg production which is spatially variable (García-Charton et al., 2008; Carter et al., 2017; Félix-Hackradt et al., 2018a). Moreover, increased egg production from no-take areas may be irrelevant if recruitment is already at saturation levels, and post-recruitment processes such as food availability and predation determine or limit adult populations (Evans et al., 2008).
Nevertheless, although site attached species can be expected to increase in abundance inside MPAs, there is little known about recruitment effects, understood as the process by which an individual is added to the population. The review by Planes et al. (2000) found an exceptionally low number of studies specifically addressing recruitment processes in MPAs. The review of recruitment studies conducted in MPAs in the north-western Mediterranean showed no difference in survival of newly-settled littoral fish between MPAs and areas outside of them, while for older recruits mortality was higher inside the MPA, probably due to the greater abundance and size of predators. In a multispecies study carried out in the Southwestern Mediterranean, Félix-Hackradt et al. (2018a) found that post-larvae abundance of commercial species were negatively related to protected areas, while the distribution of juveniles did not show any protection effect, due to a high spatial variability. Multiple factors can preclude to find effect of protection on recruitment. Félix-Hackradt et al. (2013a) found that densities of settlers could not be predicted from post-larval abundances due to high early postsettlement mortality rates in most species. Such rates of mortality showed also a high interspecific variability, mediated by the habitat complexity and direct and indirect MPA effects (Félix-Hackradt et al., 2013b). However, in a relatively small MPA (5.2 km2) in New Zealand, Le Port et al. (2017) showed evidence that snapper (Chrysophrys auratus) recruitment increased by up to 10.6% in an area of up to 400 km2 around the MPA.
MPAs Maintain Existing Resources and Provide New Potential Ones
MPAs maintain and protect biodiversity and prevent its loss (Pomeroy et al., 2004), also increasing species richness, protecting directly from extraction or by creating favorable conditions for natural biological communities different from those present in fishing territories (Roberts and Hawkins, 2000). Lester et al. (2009), in a comprehensive study of 149 scientific publications of 124 different marine reserves located throughout the world, found a 21% average increases for species richness, coinciding with the results of 25% obtained by Halpern (2003). In addition to this, Lester et al. (2009) suggested that MPA studies tend to quantify richness using species counts over a relatively small sample area (e.g., transect) probably underestimating this parameter. Overexploitation of fishing resources frequently leads to a shift in target species. Many species that constituted important fisheries decades ago have been replaced by others, generally pelagic (Pérez-Ruzafa, 2003; Fisher and Frank, 2004). The recovery of species richness and biodiversity in general opens the door, not only to the recovery of previously exploited species, but to new resources that diversify both fishing pressure and social demand.
However, in a recent study in Brazil coastal marine reserves, Rolim et al. (2019) pointed out that species richness of target and non-target fish was correlated with habitat complexity but not with protection status, relating this to the effect of a greater diversity of niches. We should take into account that MPAs are normally located in rocky areas or areas of greater structural complexity, within which the carrying capacity of habitats and the availability of effective niches are greater, and their effect may be adding or blurring that of protection. This reinforces the idea that fishing reserves must be designed or located according to the characteristics of target species, and that the carrying capacity of habitats is a fundamental factor explaining the occupation of the territory, species richness, the densities reached by species and the possible observation of movements against expected density/biomass gradients or spill-in (Langebrake et al., 2012).
MPAs Protect Genetic Diversity and Gene Pools
Since the Palumbi (2003) article highlighting the importance of considering genetic diversity in the protection of marine areas, the first work that quantified the genetic effects of fishing protection in three marine reserves in the western Mediterranean was that of Pérez-Ruzafa et al. (2006). Since then, references to the genetic structure of populations have increased and works have begun to relate it to connectivity, mainly as a good tool to measure the former parameter, in reserve networks. The terms “genetic or genomic,” “genetic diversity,” and “genetic structure” appear in 4.76, 1.59, and 1.38%, respectively, of the works on the effects of protection. On the other hand, species flux or genetic flux or connectivity are present in 6.57% of those works linked to ecosystem services. In the articles related to benefits, the terms resources and genetic are named as coexisting terms in 2.21% of them, although a direct relationship in the near four words appeared only in 0.58%.
When fisheries or artificial selection pressures are removed from the affected portion of a population, genetic diversity should be favored (Plan Development Team, 1990). This genetic diversity could be enhanced and maintained in the population by normal dispersal mechanisms and, then, in assessing this effect it is crucial to take into account the connectivity between MPAs and how it can be optimized depending on the size, location or distances between them (Pérez-Ruzafa et al., 2006; Gaines et al., 2010; Huserbråten et al., 2013; Pujolar et al., 2013; Sahyoun et al., 2016).
Connectivity can be defined as the exchange of individuals among marine populations (Cowen et al., 2006) and can take place through their dispersal as larvae, juveniles, or adults. When individuals reproduce successfully, demographic connectivity thereafter translates into genetic connectivity between MPAs or between them and their surrounding areas (Manel et al., 2019). These last authors, after a meta-analysis of 130 articles containing information on 243 species, found that marine dispersal has been extensively documented at short distances (mostly <40 km), that few recent empirical studies have demonstrated dispersal of fish at larger spatial scales (up to 400 km), but point out that these estimates were limited by the maximum sampling distance of the studies that seem not to be far-reaching enough. Connectivity patterns over greater distances remain challenging to validate empirically but have potentially important consequences in terms of reserve design and benefits (Palumbi, 2003; Andrello et al., 2017; Balbar and Metaxas, 2019; Manel et al., 2019).
MPAs Can Restore, Protect or Change Interspecific Relationships, and Community Structure
It is expected that protected areas can re-establish lost interspecific interactions, such as predatory ones, causing community changes (Guidetti, 2006). In two marine reserves in northeastern New Zealand, Babcock et al. (1999) reported a higher trophic complexity supported by an increased primary and secondary productivity compared to non-protected areas. However, studies on the effects of protection on interspecific interactions are still scarce (0.23%) and they are not reflected in the search for ecosystem services. In old and well-enforced MPAs, it is expected that the biomass of high-level predators reaches their carrying capacity, restoring top-down regulation processes that also could control the spreading of some alien and invasive fish species within MPA boundaries (Pomeroy et al., 2004; Francour et al., 2010). However, the majority of existing MPAs are either young and/or not well-enforced (Gill et al., 2017) and complementary management actions, such as species-targeted removals, should be adopted in these cases to effectively control or mitigate invasive fish population impacts on MPAs (Giakoumi et al., 2019a). These authors warn that, currently, invasive fish populations do better in Mediterranean MPAs than in unprotected sites.
Other works revised different species relations including competitive interactions (Baskett et al., 2007) or parasitism (Wood et al., 2013), although studies with these approaches are still scarce.
Since abundance, size or age of target populations increase in MPAs, density-dependent changes in life history traits are also expected within the protected areas or their surroundings (Sánchez-Lizaso et al., 2000; García-Charton et al., 2008). Although some life traits are better documented, such as those on growth or reproduction, other aspects, including behavior or feeding, remain little known (Claudet et al., 2010a; Fidler et al., 2018). Some works highlight the indirect effects that occur as a result of protection. In this sense, trophic cascades have been studied for predatory species (fishes, crabs, sea stars) mainly preying on invertebrates, and their indirect effect on algae (Pinnegar et al., 2000; Micheli et al., 2005; Guidetti, 2006; O'Sullivan and Emmerson, 2011).
Emblematic Ecosystems of a Geographical Area Are Adequately Represented and Protected, Maintaining or Restoring Their Functioning and Structure
MPAs protect marine ecosystems by eliminating or minimizing unnatural threats and human pressures inside and/or outside them, facilitating their recovery (Roberts and Hawkins, 2000; Pomeroy et al., 2004). In this sense, O'Hara et al. (2019) distinguish between proactive protection, where ecologically important areas merit protection from future degradation, from reactive protection, where areas of elevated risk would benefit from protection to mitigate existing threats. The objective to strongly protect 30% of the ocean by 2030 should be channeled through a common strategy that takes advantage of knowledge about MPA functioning to improve marine conservation efforts and policies (Hameed et al., 2017). This aspect is a well-considered item in the literature. Thus, the term protected ecosystems appears in 145 articles (3.71%) of the publications that consider the effects of MPAs. Other terms such as structured ecosystems (1.76%), protected communities (1.69%), or complex ecosystems (0.66%) also appear. However, terms such as pristine ecosystems (0.05%), pristine communities (0.03%) or well-protected communities and well-protected ecosystems, both with no records, are much scarcer. The protection of a marine area maintains or restores natural equilibrium and ecosystem balance, and thus the ecosystem structure and complexity and the processes and functions essential for focal species and habitats (Pomeroy et al., 2004; Sala and Giakoumi, 2018). However, the inherent complexity of ecosystems and their wide spatiotemporal scales of changes mean that specific responses of systems to protection or other changes in management cannot easily be extrapolated from one region to another, and have to be assessed empirically and locally (Edgar et al., 2007).
Increase Ecosystem Stability and Promote Resilience
MPAs can restore ecosystems to more complex and resilient states (Sala and Giakoumi, 2018) allowing for better defense against anthropic and natural disturbances and pressures, including climate change (Simard et al., 2016). Numerous articles mention the resilience among the effects of protection (280 articles, 7.16%) and some of them clearly specify that it increases resilience (0.59%) and none that protection decreases it. The homeostatic mechanisms or the resilience (13.60% both together) are, in fact, among the most cited ecosystem services, only after those related to biodiversity (53.58%) or climate change (18.41%). The benefits also include the stability linked to ecosystems (0.64%).
Although some works show empirical evidence of enhanced spatio-temporal stability of communities in MPAs (Fraschetti et al., 2013), ecosystem complexity does not allow us to make generalizable or well-quantified statements in this regard and there is very limited evidence on it as an emerging property of the systems (Bates et al., 2019). In fact, resilience relies on many of the mechanisms described above, such as increases in body size, condition and physiological performance, increases in predators, herbivores or parasites, in genetic and species diversity, habitat complexity, spatial connectivity or ecosystem functions, among the most important, and any improvement in these characteristics is supposed to improve the resilience of the system (Carr et al., 2018; Bates et al., 2019; Kroeker et al., 2019) although with different degrees of evidence for each aspect (Bates et al., 2019).
Currently, there are many authors who draw attention to the fact that resilience management needs to be considered in coupled social-ecological systems and that interdisciplinary approaches are critical for understanding potential effectiveness of MPAs and new marine strategies for climate and other global changes adaptation (Jones, 2014; Kroeker et al., 2019).
Control Biological Invasions
This aspect is still controversial. A relatively high number of articles mention the effects of protection related to alien or invasive species (126 articles, 3.22%) and a majority link them directly with its control (2.71%). Also, among the benefits of protection, 3.9% of the works mention the terms introduced or invasive species. However, paradoxically, only in 0.03% of the studies is this aspect linked to the ecosystem services offered by MPAs.
Giakoumi and Pey (2017) conducted a literature survey to synthesize the available information on the performance of alien/invasive species in MPAs at a global scale. Information on the effects of protection on these species is available for only 11% of the marine biogeographic regions, and most of the information they reviewed referred to mollusks and algae. The same authors concluded that MPAs can have a negative effect on alien/invasive species, finding a significant negative effect on half the species, whereas 33% of the species were positively affected. This fact and the precedent revision by Burfeind et al. (2013) that obtained different conclusions, suggest that this issue still needs more investigation.
It is worth highlighting the difference that may exist between young and old protected areas, which may explain that invasive fish populations could be favored in some Mediterranean MPAs (Otero et al., 2013; Giakoumi et al., 2019a). Francour et al. (2010) hypothesized that within old MPAs a new non indigenous fish will be top-down controlled by top predators and trophic cascades. This is once again a fact that must be considered to more effectively address protected areas management.
MPAs Act as Reference Areas in the Management of Marine Ecosystems
Scientific knowledge and research are among the most considered cultural ecosystem services in the articles reviewed (5.06%), and especially among those that reflect the benefits of the protection (38.92%).
MPAs mean the existence of “reference” or “control” areas, not damaged and intact, which can serve as a point of comparison in the proper management of marine ecosystems (Roberts and Hawkins, 2000; Willis and Millar, 2005; Wilson et al., 2010; Kay and Wilson, 2012). They allow the development of research in natural and/or non-impacted ecosystems that can be used in experimental sampling designs and as reference conditions for environmental impact and ecological status assessments (García-Charton et al., 2008; Lester et al., 2009; Fenberg et al., 2012; Díaz et al., 2016). Moreover, they facilitate stock assessment models and studies, the determination of natural mortality rates for different life-history stages (USA National Research Council/Committee on the Evaluation, Design, and Monitoring of Marine, Reserves, and Protected Areas in the U. S. Ocean Studies Board Commission on Geosciences, Environment and Resources, 2001), or the study of natural species' relationships.
MPAs Enhance or Maintain Wilderness Values
The concept of wilderness could be used in a variety of ways. In this context, it can be considered a biological descriptor, referring to places that are ecologically pristine (Casson et al., 2016). Possibly, oceans and coastal water places that possess wilderness qualities are not as identified and valued as terrestrial areas (Casson et al., 2016) and this must be considered a challenge to address. In our review, terms such as natural communities (0.33%), wilderness (0.28%), or natural ecosystems (0.26%) are recorded by a certain number of articles referring the effects of protection, but they are relatively scarce. The same happens with the articles referring to the benefits where wilderness and pristine conditions appear only in 0.94 and 0.47%, respectively.
Edgar et al. (2014), indicated that the observed 80% reduction in biomass of large fishes outside effective MPAs, coincides with the threshold value used by the International Union for Conservation of Nature (IUCN Standards, and Petitions Working Group, 2018) to categorize species as Critically Endangered for Red List assessments, suggesting that this is an evidence that MPAs are a very effective tool for preserving populations currently at risk.
However, restoring wilderness in heavily anthropized areas that have been extensively overexploited can be a difficult task. MPAs that are located in what can be considered wild regions, although they normally include extensive areas, are generally found in remote places and their species and communities can be unique and with specific compositions different from those of coastal MPAs, which are usually small in size. Thus, although there is currently a growing trend to declare MPAs with these characteristics, and it could be considered that this would enhance the recovery of anthropized and distant areas in a global network of MPAs (Graham and McClanahan, 2013), their effectiveness may be doubtful (Edgar et al., 2014) and, in any case, also subject to the efficiency of long-distance connectivity (Manel et al., 2019).
Some authors also warn of the risk that the new MPAs end up focusing on places that are remote, little used for the exploitation of resources, which makes it easier to avoid anthropic pressures and reduce stakeholders' conflicts and surveillance costs, but that end up being “residual” for commercial uses, so really, they do not protect truly threatened species and communities (Devillers et al., 2015). This possibility is clear when the reasons for protection focus solely on the conservation of biodiversity or the uniqueness of ecosystems. However, unlike in terrestrial ecosystems, most marine reserves, especially in the Atlantic-Mediterranean region, have been precisely created to safeguard, not so much biodiversity, but fishing activity and its yields. Another of the great advantages of marine reserves in warm and temperate areas, such as the Mediterranean, is that, at the same time, they are very attractive and highly profitable for recreational tourism activities and diving (Pérez-Ruzafa et al., 2017). Also interesting is that, with proper regulation of uses and spatial zonation, these activities can be profitable and there can be compatibility between them and biodiversity objectives if the spatial design always includes effective no-take areas. These circumstances cause the demand for marine reserves in exploited and tourist areas to rise exponentially (MedPAN and SPA/RAC, 2021).
Therefore, it is important to plan the creation of MPAs with a global vision of the multiple conservation objectives. Management measures must be taken into account in order to recover reasonable levels of wilderness attributes in the most anthropized coastal areas and also protect them in the areas that still maintain the same due to their isolation from human pressures.
MPAs Produce Economic Benefits for Local Populations
While in the terrestrial environment one of the main objectives of protection has been the conservation of biodiversity, the marine environment protection has been primarily linked to the management and maintenance of fishing activity (Roberts et al., 2001; Haines et al., 2018; Carr et al., 2019). Therefore, one of the most evaluated effects and the most expected or assumed services are on fishing performance. Commercial fishing (23.61%) and traditional fishing (2.43%) are widely included in the articles on the effects of protection. In the same way, fish stocks frequently appear in the works on ecosystem services (4.46%). Commercial fishing is also the most recorded benefit in the bibliography (25.84%), but it is interesting to note that among the benefits produced by MPAs, recreational fishing is nearly as mentioned (3.95%) as traditional fishing (4.24%).
In addition to fisheries, the other main market benefits of MPAs that have been most documented are those related to tourism (Alban et al., 2008, 2011; Rees et al., 2015; Viana et al., 2017; Haines et al., 2018; Davis et al., 2019), although numerous models are theoretical. In fact, marine reserves, and the effects they produce on the ecosystem, have shown a great capacity to reconcile fishing with other activities like tourism (Pérez-Ruzafa et al., 2008b). In this way, among the benefits of protection, tourism (12.43%) and related activities (ecotourism, 3.14%; recreational activities, 10.2%; artistic activities, 0.35%) are almost at the same level as fishing yields. Among nature observation activities, only whale watching has a significant number of items (0.70%), followed by fish watching (0.23%). However, the information on these aspects is not very wide, and in many cases depends on the geographical area and its socioeconomic characteristics, and on the size, age, human uses and exploitation models, data availability, etc., of the MPAs, giving rise to very heterogeneous valuations and results.
Fishing benefits both from the spread of individuals of commercial species from mature MPAs with high population density, and from the protection of their reproductive function, as demonstrated in the case of fish (Goñi et al., 2008, 2010; Forcada et al., 2009; Stobart et al., 2009; Vandeperre et al., 2011) and other species such as lobster (Díaz et al., 2011). From this fishing point of view, an MPA may potentially raise prices due to the variation of fish quantities landed, due to a quality impact based in sizes and species composition of the landings or due to better marketing opportunities and the added MPA brand (Alban et al., 2008). It should be noted that small-scale fisheries are a very important economic activity for local communities in a large part of MPAs, and that the co-management and involvement of these communities are a very important factor for their success (Niccolini et al., 2019).
Roncin et al. (2008), studying 12 MPAs in southern Europe, estimated the average yearly landed value, considering inside and outside the protected area, between 18,000 and 40,000 € per boat for commercial fishing, although Columbretes MPA appeared as an outlier with 234,000 € per boat. In the same study, they concluded that local incomes generated by MPAs amounted to a mean annual value of 710,000 € regarding the commercial fishing industry (generating 54 jobs per MPA on average), 88,000 € (two jobs) for recreational fishing and 551,000 € (13 jobs) for scuba-diving activities, although the standard deviations for the studied MPAs were high (Roncin et al., 2008). Mangos and Claudot (2013), analyzing Kuriat Islands (Tunisia), Cap de Creus (Spain), and Kas Kekova (Turkey) MPAs provided results for commercial fishing between 763,000 and 30,915,000 € per year, between 2,334,000 and 7,584,000 € for recreational fishing, and between 440,000 and 27,387,000 € for scuba-diving, confirming the variability mentioned above.
Some MPAs also offer underwater educational trails within their perimeter. This is the case of the Marine Natural Reserve of Cerbère-Banyuls-sur-Mer which has created an underwater trail to educate the public, in general, but especially tourists. This activity has generated an increase in tourist activity at the beaches of the reserve that completes the economic impact of fishing and diving. This increase in visits to swimming areas could have a negative effect on biodiversity. However, a study on this underwater trail showed that the impact of visitors was negligible on the fixed flora (from the beach where the underwater trail starts) and on the fish fauna (presence in the same densities as on the others reserve sites) (Claudet et al., 2010b).
It is important to indicate that, although MPAs redistribute benefits to some extent, they have overall positive effects in their immediate surroundings and on the local and traditional economic development (Mangos and Claudot, 2013; Ban and Frid, 2018).
Finally, other aspects such as medical (0.64%), genetic (0.58%) or pharmaceutical resources (0.17%), are starting to appear among the benefits of protection.
General Non-monetary, Sociocultural, and Educational Values
General non-monetary, sociocultural, and educational services and benefits are no less recognized than monetary ones in the case of MPAs (Fletcher et al., 2014a,b; Rosales, 2018). Scientific knowledge and research are recorded in a prominent way among the protection effects (5.06%), services (38.92%), and recognized as one of the main benefits (18.29%) (see e.g. Haines et al., 2018).
The articles on the effects of protection also underline the aesthetic and cultural aspects (1.66%), and those regarding services underline landscapes (8.68%), seascapes (4.45%), and the preservation of traditions (5.16%). Among the benefits, we find also landscapes and seascapes (5.05%), knowledge (4%), and education (3.48%).
When an MPA is working properly, it is expected that the availability of locally caught sea products for public consumption improves, and so nutritional needs of coastal residents in MPA-influenced areas are better served. At the same time, food security usually gets better while the economic status and relative wealth of coastal residents and/or resource users can also improve. Also, in a general sense, the household occupational and income structure tend to stabilize or diversify due to the appearance of new activities or due to a lower unpredictability in obtaining managed marine resources, which, in short, enhances the quality of life of populations related to the protected area (Plan Development Team, 1990; Reuchlin-Hugenholtz and McKenzie, 2015).
The measurement of non-market, non-use, or indirect benefits generated by MPAs is more challenging because they are related to goods or activities that are not associated with a market price, and there is less tangibility of their value for people (Davis et al., 2019). These include, for example, those benefits related to maintaining future fishing opportunities, educational opportunities or aesthetic experiences (Angulo-Valdés and Hatcher, 2010), the value of knowing that the site is protected for future generations, and for the species that live there in their own right (Kumar, 2010), some sociocultural aspects, cultural features, or historical sites linked to coastal areas. In the same way, value is given to the understanding of local knowledge and traditions, or non-commercial recreational activities, such as diving, snorkeling, photography, and the enjoyment of aesthetic values or emotional experiences (Ledoux and Turner, 2002). MPAs also offer opportunities for marine research, providing unique undisturbed sites for education and research, especially for comparison to areas altered by human activities (Plan Development Team, 1990), expand the scientific understanding of marine processes and ecosystems, and facilitate and improve the transmission of this knowledge to the public (Kenchington et al., 2003). In this sense, an important objective is the understanding and compatibility between management and local cultures and lifestyles, and the participation of local communities in management. In fact, research is considered a key element for the management and for harmonizing conservation with other human activities in the constituent act of the creation of marine reserves in countries such as New Zealand (Ministry for the Environment, 2016) and for decision-making on fisheries management in the European Union through its policy support program projects such as EMPAFISH (European Marine Protected Areas as tools for FISHeries management and conservation; https://cordis.europa.eu/project/id/6539/reporting/es). There are some studies that have analyzed the non-commercial value of MPAs through surveys in which users showed their willingness to pay (WTP) for aspects related to enjoyment or the environmental quality of the site. Ledoux and Turner (2002) found a users' WTP between 3.70 and 9.30 US$ per year for preventing further deterioration in Buccoo Reef Marine Park in Tobago. In a similar order of magnitude, Gelcich et al. (2013) estimated the users' WTP for an entry fee associated to management and protection of Lafken Mapu Lahual MPA in Chile between 3.77 and 4.38 US$. Börger et al. (2014) found that citizens were willing to pay between £7.76 per year on average for a 25% increase in species diversity on the Dogger Bank offshore marine area in the UK, Yu et al. (2018) estimated an average annual WTP of $27.4 for Putuo Islands MPA and of $34.3 for Nanji Islands MPA, both in China, and Niccolini et al. (2019) found that nature-based tourists would be willing to pay a daily fee between 4.11 and 7.78 € to fund marine conservation projects. In fact, the five Egyptian MPAs of the Red Sea sustain themselves through tourism daily fees, and while the money reinvested in each MPA is <10% of what it generates, the rest of the income is used to maintain other protected areas (marine and terrestrial) of the whole Egyptian network (Samy et al., 2011).
In general, there is consensus that visitors are willing to pay a premium price to support marine environment conservation, and the amount of this WTP could be influenced by social-demographic characteristics including but not limited to age, sex, education level, residence, work, environmental awareness, or personal income (Velasco et al., 2018; Yu et al., 2018; Lee et al., 2019).
Some Conclusions from the Bibliographic Review, Data and Expert Experience in the Western Mediterranean MPAs
MPAs are globally important management tools that provide protection from the effects of human exploitation and activities, supporting the conservation of marine resources, habitats and the processes they host, biological diversity, and exploited species such as commercial fish stocks (Ward and Hegerl, 2003). As a consequence, they also are expected to manage and enhance ecosystem services and material, non-material, consumptive or non-consumptive, market or non-market benefits for humans (Sanchirico et al., 2002; Leenhardt et al., 2015). However, reference to ecosystem services derived from MPAs, in the vast majority of the articles, is limited to assuming such benefits in the introduction.
The fact that a term is included in the title, abstract or keywords of an article on the effects, ecosystem services or benefits of protection, does not imply its evaluation or quantification, even for the most widely assumed and studied MPA effects. The assumption that they produce ecosystem services and benefits to society is mentioned generically in the introduction and sometimes in the discussion, but is very rarely quantified or specifically analyzed. However, studies that analyse the effects of protection with appropriate experimental designs and provide evidence and data are essential to provide an empirical basis for understanding ecosystem services (Table 3) and informing cause-effect relationships (Figure 8).
In this context, it is interesting to compare the results of the bibliographic search with the works and opinions of researchers who have been responsible for long-term monitoring of some of the most significant marine reserves in the western Mediterranean, such as those in the RESERVEBENEFIT project (Table 2, Figure 6). The results of this analysis have shown a certain coherence, and the variability or differences may be perfectly related to the heterogeneity of the marine reserves themselves, the main uses that take place in their surroundings, the time that protection has been active and the effectiveness of the management actions and surveillance.
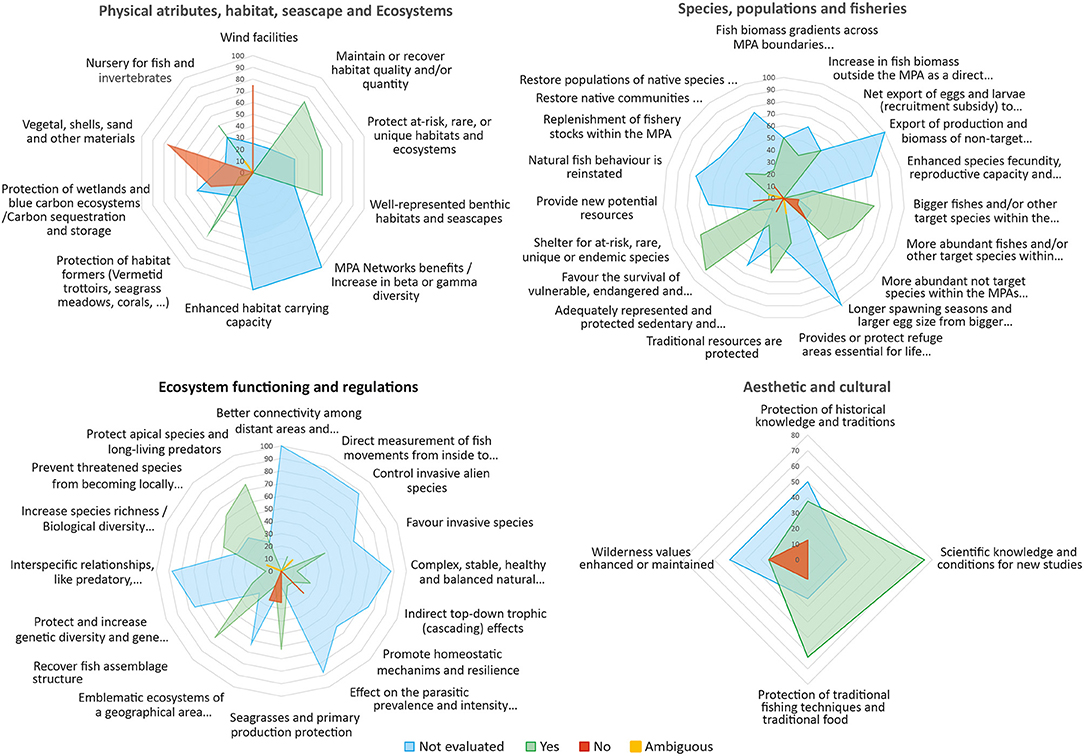
Figure 6. Frequency of occurrence (%) of main Ecosystem services and societal benefits provided by eight of the main Western Mediterranean MPAs included in RESERVEBENEFIT project (see Table 2).
The first two axis of the performed PCA explained a cumulative 58% of the variation; the first axis explained 39.43%. The graph in Figure 7 represents the 37 of the 49 terms included in the analyses that passed the rule of having a contribution equal to or higher than 10%.
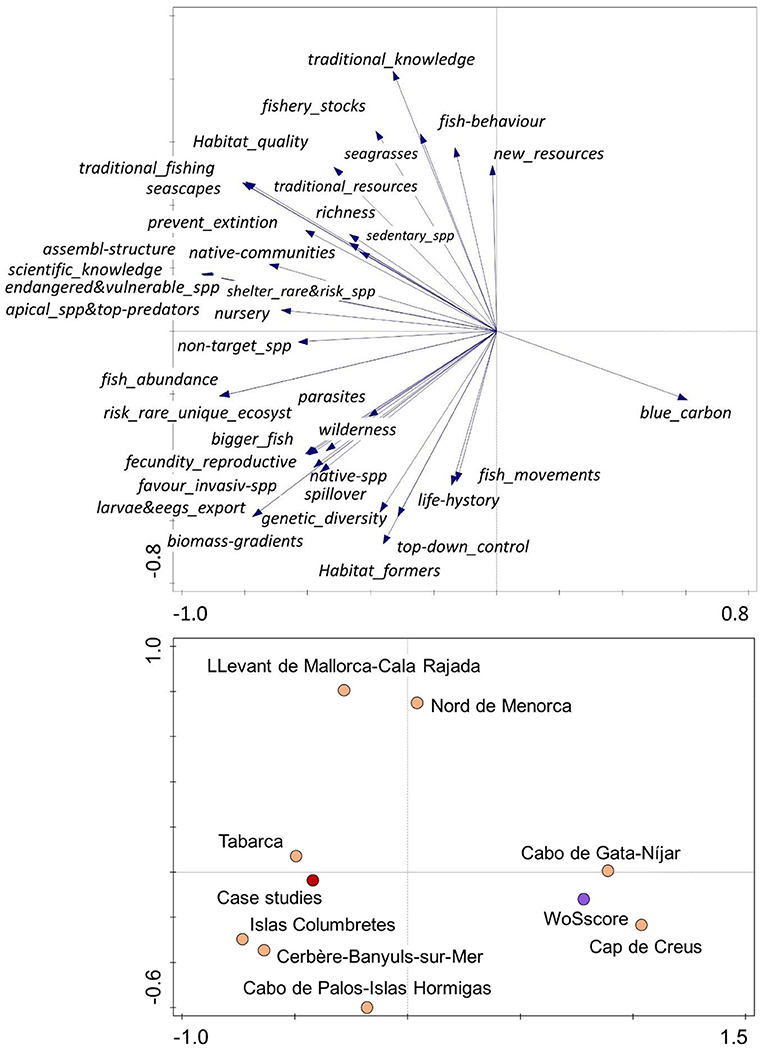
Figure 7. Biplot representations of the first two axes of the PCA performed on the scores matrix of protection effects and ecosystem services observed in RESERVEBENEFIT MPA case studies and according to the percentage of occurrence of the same terms in the articles recorded in the Web of Science (WoS). Above: ordination of the 37 of the 49 terms included in the analyses that passed the rule of having a contribution equal or higher than 10%. Below: ordination of the Western Mediterranean RESERVEBENEFIT MPAs (orange dots) and the WoS scores. Case studies red dot correspond to the average score of all MPAs.
The results show the variability of the effects depending on the MPAs. While there are some of the best-studied reserves that differ along the first axis from the results of the bibliographic analysis (WoSscore) and are grouped around the mean value of the case studies, others completely coincide with the WoS results, while others occupy an intermediate position and differ from both evaluations on the second axis. The negative part of axis 1 is characterized by the terms fish abundance, apical species and top-predators, conservation of at risk, rare and unique ecosystems, scientific knowledge, endangered and vulnerable species, or assemblage structure and traditional fishing, while the positive part is determined by the weight of the assessment of the role of protection in communities that contribute to blue carbon (carbon sequestration) having lower relative weight than the other terms. The positive end of axis two is determined by the contribution to traditional knowledge, fish stocks, fish behavior or the contribution of new resources, while the negative part is determined by the existence of studies on fish movements, the top-down control of the food web, the preservation of life cycle phases and habitat builders.
The main apparent result is that reality exceeds what is reflected in the bibliography. That is, all the services and protection effects evaluated, except the role in carbon sequestration, are more confirmed in the field data and direct experience of researchers working in the RESERVEBENEFIT case study MPAs than what is reflected in the bibliography. In some way, this is a consequence of how the scores have been built, since an effect found in an MPA has a value in the matrix of 100%, while in the literature this percentage is diluted among many publications that analyze other effects or services. The log transformation of the data reduces these differences, but they still remain in some way. However, the fact that a significant core of RESERVEBENEFIT MPAs is aggregated around the middle score indicates that the majority of services occur across a majority of MPAs. On the other hand, the fact that blue carbon is the only factor that does not follow this pattern may be determined by the circumstance that this service is more accentuated (and it is probably easier to assess, more expected to occur and has been more evaluated) in marshes, while the MPAs studied in RESERVEBENEFIT correspond to rocky coastlines and coves and studies on carbon sequestration by photophilic algae communities and seagrass meadows have been much scarcer and do not appear as proven evidence.
MPAs add precautionary management principles and insurance against failure of other management options (Plan Development Team, 1990; USA National Research Council/Committee on the Evaluation, Design, and Monitoring of Marine, Reserves, and Protected Areas in the U. S. Ocean Studies Board Commission on Geosciences, Environment and Resources, 2001). If they are appropriately managed and well-coordinated, considering the needs and wants of affected populations and stakeholders, they could simplify enforcement and attract financial support (Plan Development Team, 1990). They also enhance the public knowledge and environmental awareness regarding marine ecosystem values and functioning, and their acceptance of protection measures and the objectives of environmental and social sustainability.
Understanding the elements that make up ecosystems, including biodiversity, the processes that work in them, and how protection modifies them, is an essential pillar to understand the importance of MPAs and their role in the functioning and well-being of related human societies.
The protection of marine areas may generate a series of ecosystem services, from preventing erosion to providing shelter and pristine habitats for species, enhancing ecological processes, favoring carbon sequestration, preserving biodiversity, including genetic, or recovering and providing resources for humans (Table 3). These depend largely on the final objectives and the effectiveness of the protection and, more importantly, on the location, size, age, geomorphological and hydrodynamic conditions, and types of habitat and communities of the protected area. These ecosystem services can be translated into socio-economic benefits, including present and future benefits to sustainability.
However, classifying ecosystem attributes and processes based on the ecosystem services and benefits that they produce, or vice versa, is difficult due to the complex web of interactions that take place. Most attributes result in services that improve those attributes or regulate processes. At the same time, the resulting social benefits may be the consequence of the existence of more than one attribute or the interaction between different processes. For example, it has been quantitatively demonstrated that the presence of top predators and high reef complexity have an additive effect and can increase total fish abundance up to 300% (Hensel et al., 2019). For this reason, although the construction of tables such as the one produced for this work is useful because it facilitates the inventory of the different aspects, it is difficult to find a coherent order that reduces repetition. A more realistic view is facilitated by a cause-effect relationship diagram such as the one represented in Figure 8.
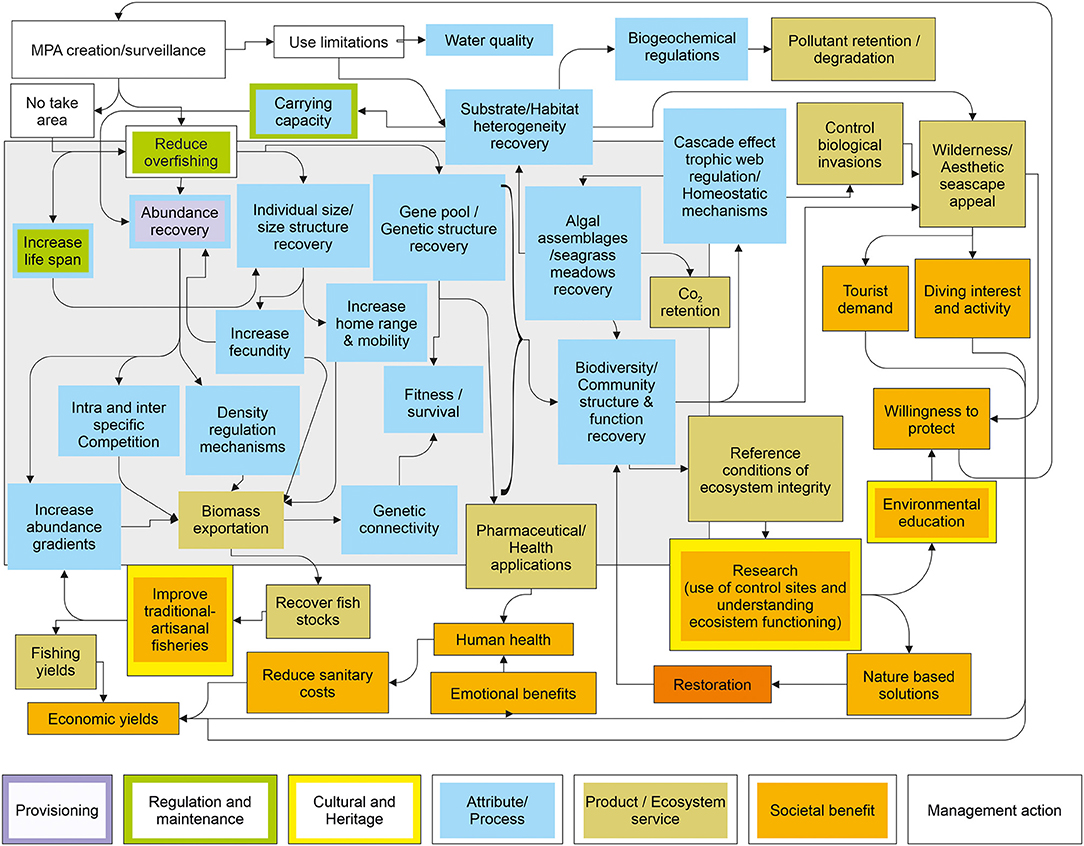
Figure 8. Cause effect relationships diagram considering human actions, MPA attributes and the effects of protection on biological and ecological processes and the ecosystems services and societal benefits provided.
The analysis of ecosystem services and benefits for society of some of the oldest and best-studied marine reserves in the Western Mediterranean, based on the direct knowledge of the researchers who have carried out scientific monitoring over the last decades, and on the data obtained during the same (Table 2, Figure 6), shows some heterogeneity in the provision or in the evidence of them. The expected services or benefits rarely are present in more than 80% of the eight case studies considered. We may expect that the basic ecological processes that regulate the functioning and provision of services provided by MPAs should be common to all geographic areas, and that differences found will be attributable to variation in the characteristics of each MPA. The comparative analysis carried out between the results of the bibliographic search and the case studies (all from the western Mediterranean) suggests such basic uniformity.
The clearest evidence that exists is for the recovery of individual populations of target or threatened species and their reproductive potential. However, many of the other services, like those related to the recovery of habitats, the carrying capacity of the system or the connectivity among distant areas, are not evaluated and verified in any of the case studies considered. Clearly, much more research work is needed to come up with a numerical modeling of the benefits of protection or the consequences of different management measures.
However, a conceptual model such as the one proposed in Figure 8 can help to understand the positive or negative synergies between various attributes or processes affected by protection measures. At the same time, it can serve to identify gaps in knowledge, in the evaluation of some of these interactions and in the ecosystem services that are expected to obtain or improve with such management measures.
Finally, it must be borne in mind that MPA networks could be managed cooperatively (e.g., coordinating the distribution of fishing effort in the fishing grounds between reserves or jointly planning monitoring programs and checking the effectiveness of management measures), and be designed to operate synergistically at various spatial scales (e.g., taking into account the optimum distance between MPAs depending on the spill-over and connectivity in the area). Moreover, networks could include a wide range of protection levels in their spatial ordination, with no-take areas designed according to the particular objectives of each MPA, where most of the uses are prohibited, and buffer zones with regulated uses, covering different conservation objectives and spatial ordination of human activities. These networks could be a tool that fulfill ecological objectives more effectively than individual sites could alone (WCPA/IUCN, 2007; Grorud-Colvert et al., 2014; Ospina-Alvarez et al., 2020), helping to more effectively achieve social and economic benefits (Klein et al., 2008).
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Author Contributions
AP-R and CM have designed the article and processed the data. All the authors have collaborated in the bibliographic review and provided the data from the case studies included in the RESERVEBENEFIT project and have contributed to the discussion and revision of the text.
Funding
This work has been funded through the BiodivERsA3-2015-21 Cofund RESERVEBENEFIT European project, with the national funder ANR (France), Formas (Sweden), DLR (Germany), and the Project PCIN-2016-139 of the Spanish Ministry of Economy and Competitiveness.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank all the RESERVEBENEFIT members for the fruitful discussions and exchanges that we have had throughout its development, and to the referees and the editor for their comments and suggestions that have helped to significantly improve this article.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2021.613819/full#supplementary-material
References
Aalto, E. A., Micheli, F., Boch, C. A., Espinoza Montes, J. A., Woodson, C. B., and De Leo, G. A. (2019). Catastrophic mortality, allee effects, and marine protected areas. Am. Natural. 193, 391–408. doi: 10.1086/701781
Abdulla, A., Gomei, M., Hyrenbach, D., Notarbartolo-di-Sciara, G., and Agardy, T. (2008). Challenges facing a network of representative marine protected areas in the Mediterranean: prioritizing the protection of underrepresented habitats. ICES J. Mar. Sic. 66, 22–28. doi: 10.1093/icesjms/fsn164
Alban, F., Appéré, G., and Boncoeur, J. (2008). Economic Analysis of Marine Protected Areas. A Literature Review. EMPAFISH Project, Booklet No. 3. Murcia: Editum.
Alban, F., Boncoeur, J., and Roncin, N. (2011). “Assessing the impact of marine protected areas on society's well-being: an economic perspective,” in Marine Protected Areas: A Multidisciplinary Approach, ed. J. Claudet (Cambridge: Cambridge University Press), 226–246. doi: 10.1017/CBO9781139049382.013
Alemany, D., Iribarne, O. O., and Acha, E. M. (2013). Effects of a large-scale and offshore marine protected área on the demersal fish assemblage in the Southwest Atlantic. ICES J. Mar. Sci. 70, 123–134. doi: 10.1093/icesjms/fss166
Andrello, M., Guilhaumon, F., Albouy, C., Parravicini, V., Scholtens, J., Verley, P., et al. (2017). Global mismatch between fishing dependency and larval supply from marine reserves. Nat. Commun. 8:16039. doi: 10.1038/ncomms16039
Andrzejaczek, S., Chapple, T. K., Curnick, D. J., Carlisle, A. B., et al. (2020). Individual variation in residency and regional movements of reef manta rays mobula alfredi in a large marine protected area. Mar. Ecol. Prog. Ser. 639, 137–153. doi: 10.3354/meps13270
Angulo-Preckler, C., Cid, C., Oliva, F., and Avila, C. (2015). Antifouling activity in some benthic Antarctic invertebrates by “in situ” experiments at deception island, Antarctica. Mar. Environ. Res. 105, 30–38. doi: 10.1016/j.marenvres.2015.02.001
Angulo-Valdés, J. A., and Hatcher, B. G. (2010). A new typology of benefits derived from marine protected areas. Mar. Policy 34, 635–644. doi: 10.1016/j.marpol.2009.12.002
Armoškaite, A., Purina, I., Aigars, J., Strāke, S., Pakalniete, K., Frederiksen, P., et al. (2020). Establishing the links between marine ecosystem components, functions and services: an ecosystem service assessment tool. Ocean Coast. Manage. 193:105229. doi: 10.1016/j.ocecoaman.2020.105229
Ashley, M. C., Mangi, S. C., and Rodwell, L. D. (2014). The potential of offshore windfarms to act as marine protected areas – a systematic review of current evidence. Mar. Policy 45, 301–309. doi: 10.1016/j.marpol.2013.09.002
Babcock, R. C., Kelly, S., Shears, N. T., Walker, J. W., and Willis, T. J. (1999). Changes in community structure in temperate marine reserves. Mar. Ecol. Prog. Ser. 189, 125–134. doi: 10.3354/meps189125
Balbar, A. C., and Metaxas, A. (2019). The current application of ecological connectivity in the design of marine protected areas. Glob. Ecol. Conserv. 17:e00569. doi: 10.1016/j.gecco.2019.e00569
Ban, N. C., and Frid, A. (2018). Indigenous peoples' rights and marine protected areas. Mar. Policy 87, 180–185. doi: 10.1016/j.marpol.2017.10.020
Barbera, C., Bordehore, C., Borg, J. A., Glémarec, M., Grall, J., Hall-Spencer, J. M., et al. (2003). Conservation and management of northeast Atlantic and Mediterranean maerl beds. Aquat. Conserv. Mar. Freshw. Ecosyst. 13, S65–S76. doi: 10.1002/aqc.569
Barberá, C., Mallol, S., Verges, A., Cabanellas-Reboredo, M., Díaz, D., and Goñi, R. (2017). Maerl beds inside and outside a 25-year-old no-take área. Mar. Ecol. Progr. Series 572, 77–90. doi: 10.3354/meps12110
Barbier, E. B., Hacker, S. D., Kennedy, C., Koch, E. W., Stier, A. C., and Silliman, B. R. (2011). The value of estuarine and coastal ecosystem services. Ecol. Monogr. 81, 169–193. doi: 10.1890/10-1510.1
Barnett, L. A. K., Branch, T. A., Ranasinghe, R. A., and Essington, T. E. (2017). Old-Growth fishes become scarce under fishing. Curr. Biol. 27, 2843–2848. doi: 10.1016/j.cub.2017.07.069
Baskett, M. L., Micheli, F., and Levin, S. A. (2007). Designing marine reserves for interacting species: Insights from theory. Biol. Conserv. 137, 163–179. doi: 10.1016/j.biocon.2007.02.013
Bates, A. E., Cooke, R. S. C., Duncan, M. I., Edgar, G. J., Bruno, J. F., Benedetti-Cecchi, L., et al. (2019). Climate resilience in marine protected areas and the ‘protection paradox’. Biol. Conserv. 236, 3015–3314. doi: 10.1016/j.biocon.2019.05.005
Bauer, D. M., and Johnston, R. J. (2013). The economics of rural and agricultural ecosystem services: purism versus practicality. Agric. Res. Econ. Rev. 42, iii–xv. doi: 10.1017/S1068280500007589
Benedetti-Cecchi, L., Bertocci, I., Micheli, F., Maggi, E., Fosella, T., and Vaselli, S. (2003). Implications of spatial heterogeneity for management of marine protected areas (MPAs): examples from assemblages of rocky coasts in the northwest Mediterranean. Marine Environmental Research 55, 429–458. doi: 10.1016/S0141-1136(02)00310-0
Berkeley, S. A., Hixon, M. A., Larson, R. J., and Love, M. S. (2004). Fisheries sustainability via protection of age structure and spatial distribution of fish populations. Fisheries 29, 23–32. doi: 10.1577/1548-8446(2004)29[23:FSVPOA]2.0.CO;2
BIOMEX project (2006). Assessment of BIOMass Export From Marine Protected Areas & Its Impacts on Fisheries in the Western Mediterranean Sea. (QLRT-2001-0891) Final report, Perpignan, 500.
Blowes, S. A., Chase, J. M., Di Franco, A., Frid, O., Gotelli, N. J., Guidetti, P., et al. (2020). Mediterranean marine protected areas have higher biodiversity via increased evenness, not abundance. J. Appl. Ecol. 57, 578–589. doi: 10.1111/1365-2664.13549
Börger, T., Hattam, C., Burdon, D., Atkins, J. P., and Austen, M. C. (2014). Valuing conservation benefits of an offshore marine protected area. Ecol. Econ. 108, 229–241. doi: 10.1016/j.ecolecon.2014.10.006
Botsford, L. W. (2001). Physical influences on recruitment to California Current invertebrate populations on multiple scales. ICES J. Mar. Sci. 58, 1081–1091. doi: 10.1006/jmsc.2001.1085
Boyd, J., and Krupnick, A. J. (2013). Using ecological production theory to define and select environmental commodities for nonmarket valuation. Agric. Res. Econ. Rev. 42, 1–32. doi: 10.1017/S1068280500007590
Boyd, J., Ringold, P. L., Krupnick, A., Johnston, R. J., Matt, W., and Hall, K. (2016). Ecosystem services indicators: improving the linkage between biophysical and economic analyses. Int. Rev. Environ. Resourc. Econ. 8, 225–279. doi: 10.1561/101.00000073
Browman, H. I., Cury, P. M., Hilborn, R., Jennings, S., Lotze, H. K., Mace, P. M., et al. (2004). Perspectives on ecosystem-based approaches to the management of marine resources. Mar. Ecol. Prog. Ser. 274, 269–303. doi: 10.3354/meps274269
Burdon, D., Potts, T., Barbone, C., and Mander, L. (2017). The matrix revisited: a bird's-eye view of marine ecosystem service provision. Mar. Policy 77, 78–89. doi: 10.1016/j.marpol.2016.12.015
Burfeind, D. D., Pitt, K. A., Connolly, R. M., and Byers, J. E. (2013). Performance of non-native species within marine reserves. Biol. Invas. 15, 17–28. doi: 10.1007/s10530-012-0265-2
Cabral, R. B., Gaines, S. D., Johnson, B. A., Bell, T. W., and White, C. (2016b). Drivers of redistribution of fishing and non-fishing effort after the implementation of a marine protected area network. Ecol. Appl. 27, 416–428. doi: 10.1002/eap.1446
Cabral, R. B., Gaines, S. D., Lim, M. T., Atrigenio, M. P., Mamauag, S. S., Gerold, C., et al. (2016a). Siting marine protected areas based on habitat quality and extent provides the greatest benefit to spatially structured metapopulations. Ecosphere 7:e015331v. doi: 10.1002/ecs2.1533
Calvo, M., Templado, J., and Penchaszadeh, P. E. (1998). Reproductive biology of the gregarious mediterranean vermetid gastropod Dendropoma petraeum. J. Mar. Biol. Assoc. 78, 525–549. doi: 10.1017/S0025315400041606
Carr, M. H., Bell, K. L. C., Leary, P., Sagar, H. L., and Tucker, S. (2018). How MPAs Enhance the Resilience of Coastal Marine Ecosystems and their Services. Supplemental Report by the Marine Protected Areas Federal Advisory Committee's Ecosystem Team. MD: Silver Spring, 24.
Carr, M. H., White, J. W., Saarman, E., Lubchenco, J., Milligan, K., and Caselle, J. E. (2019). Marine Protected Areas exemplify the evolution of science and policy. Oceanography 32, 94–103. doi: 10.5670/oceanog.2019.315
Carter, A. B., Davies, C. R., Emslie, M. J., Mapstone, B. D., Russ, G. R., Tobin, A. J., et al. (2017). Reproductive benefits of no-take marine reserves vary with region for an exploited coral reef fish. Sci. Rep. 7:e9693. doi: 10.1038/s41598-017-10180-w
Casson, S. A., Martin, V. G., Watson, A., Stringer, A., and Kormos, C. F. (Eds). (2016). Wilderness Protected Areas: Management Guidelines for IUCN Category 1b Protected Areas. Gland: IUCN.
Castilla, J. C., and Pino, C. (1996). The small-scale fishery of the red sea urchin, Loxechinus albus, in Chile and the Management and Exploitation Area of Caleta El Quisco. Out Shell 5, 5–8.
Ceccherelli, G., Pais, A., Pinna, S., Sechi, N., and Chessa, L. A. (2011). Human impact on Paracentrotus lividus: the result of harvest restrictions and accessibility of locations. Mar. Biol. 158, 845–852. doi: 10.1007/s00227-010-1611-5
Charles, A., Westlund, L., Bartley, D. M., Fletcher, W. J., García, S., Govan, H., and Sanders, J. (2016). Fishing livelihoods as key to marine protected areas: insights from the World Parks Congress. Aquat. Conserv. 26(Suppl. 2), 165–184. doi: 10.1002/aqc.2648
Chemello, R., Dieli, T., and Antonioli, F. (2000). Il Ruolo Dei “Reef” a Molluschi Vermetidi Nella Valutazione Della Biodiver-Sità. Mare e Cambiamenti Globali. Roma: Quaderni ICRAM, 105–118.
Chirico, A. A. D., McClanahan, T. R., and Eklöf, J. S. (2017). Community- and government-managed marine protected areas increase fish size, biomass and potential value. PLoS ONE 12:e0182342. doi: 10.1371/journal.pone.0182342
Claudet, J., Lenfant, P., and Schrimm, M. (2010b). Snorkelers impact on fish communities and algae in a temperate marine protected area. Biodivers. Conserv. 19, 1649–1658. doi: 10.1007/s10531-010-9794-0
Claudet, J., Osenberg, C. W., Benedetti-Cecchi, L., Domenici, P., García-Charton, J. A., Pérez-Ruzafa, A., et al. (2008). Marine reserves: size and age do matter. Ecol. Lett. 11, 481–489. doi: 10.1111/j.1461-0248.2008.01166.x
Claudet, J., Osenberg, C. W., Domenici, P., Badalamenti, F., Milazzo, M., Falcón, J. M., et al. (2010a). Marine reserves: Fish life history and ecological traits matter. Ecol. Applic. 20, 830–839. doi: 10.1890/08-2131.1
Coll, J., García-Rubies, A., Morey, G., and Grau, A. M. (2012). The carrying capacity and the effects of protection level in three marine protected areas in the Balearic islands (NW Mediterranean). Sci. Mar. 76, 809–826. doi: 10.3989/scimar.03531.02H
Coll, J., Morey, G., Verger, F., and Navarro, O. (2017). La Reserva Marina del Nord de Menorca. Seguiment de les poblacions de peixos de substrat rocós en el període 2000-2017. Palma de Mallorca: Direcció General de Pesca i Medi Marí-Govern de les Illes Balears – Tragsatec.
Colloca, F., Garofalo, G., Bitetto, I., Facchini, M. T., Grati, F., Martiradonna, A., et al. (2015). The seascape of demersal fish nursery areas in the North Mediterranean sea, a first step towards the implementation of spatial planning for trawl fisheries. PLoS ONE 10:e0119590. doi: 10.1371/journal.pone.0119590
Coma, R., Serrano, E., Linares, C., Ribes, M., Díaz, D., and Ballesteros, E. (2011). Sea urchins predation facilitates coral invasion in a marine reserve. PLoS ONE 6:e22017. doi: 10.1371/journal.pone.0022017
Coppari, M., Zanella, C., and Rossi, S. (2019). The importance of coastal gorgonians in the blue carbon budget. Sci. Rep. 9, 13550. doi: 10.1038/s41598-019-49797-4
Costanzo, L. G., Marletta, G., and Alongi, G. (2021). Ecological status of coralligenous macroalgal assemblages in the marine protected area (MPA) Isole Ciclopi (Ionian sea). Plants 10:329. doi: 10.3390/plants10020329
Costello, M. J., and Ballantine, B. (2015). Biodiversity conservation should focus on no take marine reserves. Trends Ecol. Evolut. 30, 507–509. doi: 10.1016/j.tree.2015.06.011
Cowen, R. K., Paris, C. B., and Srinivasan, A. (2006). Scaling of connectivity in marine populations. Science 311, 522–527. doi: 10.1126/science.1122039
Crec'hriou, R., Alemany, F., Roussel, E., Chassanite, A., Marinaro, J. Y., Mader, J., et al. (2010). Fisheries replenishment of early life taxa: potential export of fish eggs and larvae from a temperate marine protected area. Fish. Oceanogr. 19, 135–150. doi: 10.1111/j.1365-2419.2010.00533.x
Curnick, D. J., Andrzejaczek, S., Jacoby, D. M. P., Coffey, D. M., Carlisle, A. B., Chapple, T. K., et al. (2020). Behavior and ecology of silky sharks around the Chagos archipelago and evidence of Indian ocean wide movement. Front. Mar. Sci. 7:1045. doi: 10.3389/fmars.2020.596619
D'Aloia, C. C., Bogdanowicz, S. M., Francis, R. K., Majoris, J. E., Harrison, R. G., and Buston, P. M. (2015). Patterns, causes, and consequences of marine larval dispersal. Proc. Natl. Acad. Sci. U.S.A. 112, 13940–13945. doi: 10.1073/pnas.1513754112
Davis, K. J., Vianna, G. M. S., Meeuwig, J. J., Meekan, M. G., and Pannell, D. J. (2019). Estimating the economic benefits and costs of highly-protected marine protected areas. Ecosphere 10:e02879. doi: 10.1002/ecs2.2879
De Francesco, M. C., Carranza, M. L., Varricchione, M., Tozzi, F. P., and Stanisci, A. (2019). Natural protected areas as special sentinels of littering on coastal dune vegetation. Sustainability 11:5446. doi: 10.3390/su11195446
De Groot, R. S., Fisher, B., Christie, M., Aronson, J., Braat, L., Haines-Young, R., et al. (2010). “Integrating the Ecological and Economic Dimensions in Biodiversity and Ecosystem Service Valuation,” in The Economics of Ecosystems and Biodiversity (TEEB): Ecological and Economic Foundations, ed P. Kumar (London: Earthscan Routledge), 9–40.
De Groot, R. S., Wilson, M. A., and Boumans, R. M. J. (2002). A typology for the classification, description and valuation of ecosystem functions, goods and services. Ecol. Econ. 41, 393–408. doi: 10.1016/S0921-8009(02)00089-7
Del Pilar-Ruso, Y., and Bayle-Sempere, J. T. (2006). Diel and vertical movements of preflexion fish larvae assemblages associated with Posidonia oceanica beds. Sci. Mar. 70, 399–406. doi: 10.3989/scimar.2006.70n3399
Devillers, R., Pressey, R. L., Grech, A., Kittinger, J. N., Edgar, G. J., Ward, T., et al. (2015). Reinventing residual reserves in the sea: are we favouring ease of establishment over need for protection? Aquat. Conserv. Mar. Freshw. Ecosyst. 25, 480–504. doi: 10.1002/aqc.2445
Di Franco, A., Thiriet, P., Di Carlo, G., Dimitriadis, C., Francour, P., Gutiérrez, N. L., et al. (2016). Five key attributes can increase marine protected áreas performance for small-scale fisheries management. Sci. Rep. 6:e38135. doi: 10.1038/srep38135
Di Lorenzo, M., Claudet, J., and Guidetti, P. (2016). Spillover from marine protected areas to adjacent fisheries has an ecological and a fishery component. J. Nat. Conserv. 32, 62–66. doi: 10.1016/j.jnc.2016.04.004
Díaz, D., Mallol, S., Parma, A., and Goñi, R. (2016). A 25-year marine reserve as proxy for the unfished condition of an exploited species. Biol. Conserv. 203, 97–107. doi: 10.1016/j.biocon.2016.09.002
Díaz, D., Mallol, S., Parma, M., and Goñi, R. (2011). Decadal trend in lobster reproductive output from a temperate marine protected area. Mar. Ecol. Prog. Ser. 433, 149–157. doi: 10.3354/meps09182
D'Onghia, G., Maiorano, P., Sion, L., Giove, A., Capezzuto, F., Carlucci, R., et al. (2010). Effects of deep-water coral banks on the abundance and size structure of the megafauna in the Mediterranean sea. Deep Sea Res. Part II Top Stud. Oceanogr. 57, 397–411. doi: 10.1016/j.dsr2.2009.08.022
EC (2008). Directive 2008/56/Ec of the European Parliament and of the Council of 17 June 2008 establishing a framework for community action in the field of marine environmental policy (Marine Strategy Framework Directive). Off. J. Eur. Union L 164, 19–40.
Edgar, G. J., Banks, S., Bensted-Smith, R., Calvopiña, M., Chiriboga, A., Garske, L. E., et al. (2008). Conservation of threatened species in the Galápagos marine reserve through identification and protection of marine key biodiversity areas. Aquat. Conserv. Mar. Freshw. Ecosyst. 18, 955–968. doi: 10.1002/aqc.901
Edgar, G. J., and Barrett, N. S. (1997). Short term monitoring of biotic change in Tasmanian marine reserves. J. Exp. Mar. Biol. Ecol. 213, 261–279. doi: 10.1016/S0022-0981(96)02769-4
Edgar, G. J., and Barrett, N. S. (1999). Effects of the declaration of marine reserves on Tasmanian reef fishes, invertebrates and plants. J. Exp. Mar. Biol. Ecol. 242, 107–144. doi: 10.1016/S0022-0981(99)00098-2
Edgar, G. J., Barrett, N. S., and Stuart-Smith, R. D. (2009). Exploited reefs protected from fishing transform over decades into conservation features otherwise absent from seascapes. Ecol. Appl. 19, 1967–1974. doi: 10.1890/09-0610.1
Edgar, G. J., Russ, G. R., and Babcock, R. C. (2007). “Marine protected areas,” in Marine Ecology, eds S. D. Connell, and B. M. Gillanders (Oxford: Oxford University Press), 534–565.
Edgar, G. J., Stuart-Smith, R. D., Willis, T. J., Kininmonth, S., Baker, S. C., Banks, S., et al. (2014). Global conservation outcomes depend on marine protected areas with five key features. Nature 506, 216–220. doi: 10.1038/nature13022
Enge, S., Nylund, G. M., and Pavia, H. (2013). Native generalist herbivores promote invasion of a chemically defended seaweed via refuge-mediated apparent competition. Ecol. Lett. 16, 487–492. doi: 10.1111/ele.12072
Entrambasaguas, L., Pérez-Ruzafa, Á., García-Charton, J. A., Stobart, B., and Bacallado, J. J. (2008). Abundance, spatial distribution and habitat relationships of echinoderms in the Cabo Verde archipelago (eastern Atlantic). Mar. Freshw. Res. 59, 477–488. doi: 10.1071/MF07109
Evans, R. D., Russ, G. R., and Kritzer, J. P. (2008). Batch fecundity of Lutjanus carponotatus (Lutjanidae) and implications of no-take marine reserves on the great barrier reef, Australia. Coral Reefs 27, 179–189. doi: 10.1007/s00338-007-0309-8
Failler, P., Touron-Gardic, G., Drakeford, B., Sadio, O., and Traoré, M. S. (2020). Perception of threats and related management measures: the case of 32 marine protected areas in West Africa. Mar. Policy 117:103936. doi: 10.1016/j.marpol.2020.103936
Félix-Hackradt, F. C., Hackradt, C. W., Treviño-Otón, J., Pérez-Ruzafa, A., and García-Charton, J. A. (2013a). Temporal patterns of settlement, recruitment and post-settlement losses in a rocky reef fish assemblage in the South-Western Mediterranean sea. Mar. Biol. 160, 2337–2352. doi: 10.1007/s00227-013-2228-2
Félix-Hackradt, F. C., Hackradt, C. W., Treviño-Otón, J., Pérez-Ruzafa, A., and García-Charton, J. A. (2014). Habitat use and ontogenetic shifts of fish life stages at rocky reefs in South-western Mediterranean sea. J. Sea Res. 88, 67–77. doi: 10.1016/j.seares.2013.12.018
Félix-Hackradt, F. C., Hackradt, C. W., Treviño-Otón, J., Pérez-Ruzafa, A., and García-Charton, J. A. (2018a). Effect of marine protected areas on distinct fish life-history stages. Mar. Environ. Res. 140, 200–209. doi: 10.1016/j.marenvres.2018.06.012
Félix-Hackradt, F. C., Hackradt, C. W., Treviño-Otón, J., Segovia-Viadero, M., Pérez-Ruzafa, A., and García-Charton, J. A. (2013b). Environmental determinants on fish post-larval distribution in coastal areas of south-western Mediterranean Sea. Estuar. Coast. Shelf Sci. 129, 59–72. doi: 10.1016/j.ecss.2013.05.029
Félix-Hackradt, F. C., Sanchis-Martinez, A. M., Hackradt, C. W., Treviño-Otón, J., and García-Charton, J. A. (2018b). Distribution and ecological relations among the alien crab, Percnon gibbesi (H. Milne-Edwards 1853) and autochthonous species, in and out of an SW Mediterranean MPA. Hydrobiologia 806, 187–201. doi: 10.1007/s10750-017-3357-2
Fenberg, P. B., Caselle, J., Claudet, J., Clemence, M., Gaines, S., García-Charton, J. A., et al. (2012). The science of European marine reserves: status, efficacy and needs. Mar. Policy 36, 1012–1021. doi: 10.1016/j.marpol.2012.02.021
Fidler, R. Y., Carroll, J., Rynerson, K. W., Matthews, D. F., and Turingan, R. G. (2018). Coral reef fishes exhibit beneficial phenotypes inside marine protected areas. PLoS ONE 13:e0193426. doi: 10.1371/journal.pone.0193426
Fisher, B., Costanza, R., Turner, R. K., and Morling, P. (2007). Defining and Classifying Ecosystem Services for Decision Making. CSERGE Working Paper EDM 07–04. Norwich: University of East Anglia. Available online at: http://hdl.handle.net/10419/80264
Fisher, B., and Turner, R. K. (2008). Ecosystem services: classification for valuation. Biol. Conserv. 141, 1167–1169. doi: 10.1016/j.biocon.2008.02.019
Fisher, B., Turner, R. K., and Morling, P. (2009). Defining and classifying ecosystem services for decision making. Ecol. Econ. 68, 643–653. doi: 10.1016/j.ecolecon.2008.09.014
Fisher, J. A. D., and Frank, K. T. (2004). Abundance–distribution relationships and conservation of exploited marine fishes. Mar. Ecol. Prog. Ser. 279, 201–213. doi: 10.3354/meps279201
Fletcher, R., Baulcomb, C., Hall, C., and Hussain, S. (2014a). Revealing marine cultural ecosystem services in the Black sea. Mar. Policy 50, 151–161. doi: 10.1016/j.marpol.2014.05.001
Fletcher, S., Rees, S., Gall, S., Shellock, R., Dodds, W., and Rodwell, L. (2014b). Assessing the socio-economic benefits of marine protected areas. A Report for Natural Resources Wales by the Centre for Marine and Coastal Policy Research. Plymouth, MA: Plymouth University.
Forcada, A. (2005). Ictiofauna en Reservas Marinas: Influencia de la Estructura del Hábitat, Efecto de la Protección y Tamaño del Área Protegida. Alicante: Instituto Alicantino de Cultura Juan Gil-Albert, Publicaciones de la Diputación de Alicante, 194.
Forcada, A., Bayle-Sempere, J. T., Valle, C., and Sánchez-Jerez, P. (2008). Habitat continuity effects on gradients of fish biomass across marine protected area boundaries. Mar. Environ. Res. 66, 536–547. doi: 10.1016/j.marenvres.2008.08.003
Forcada, A., Valle, C., Bonhomme, P., Criquet, G., Cadiou, G., Lenfant, P., et al. (2009). Effects of habitat on spillover from marine protected areas to artisanal fisheries. Mar. Ecol. Prog. Ser. 379, 197–211. doi: 10.3354/meps07892
Forcada, A., Valle, C., Sanchez-Lizaso, J. L., Bayle-Sempere, J. T., and Corsi, F. (2010). Structure and spatio-temporal dynamics of artisanal fisheries around a Mediterranean marine protected area. ICES J. Mar. Sci. 67, 191–203. doi: 10.1093/icesjms/fsp234
Francour, P., Mangialajo, L., and Pastor, J. (2010). “Mediterranean marine protected areas and non-indigenous fish spreading,” in Fish Invasions of the Mediterranean Sea: Change and Renewal, ed D. Golani D, and B. Appelbaum-Golani (Sofia: Pensoft), 127–144.
Fraschetti, S., Guarnieri, G., Bevilacqua, S., Terlizzi, A., and Boero, F. (2013). Protection enhances community and habitat stability: evidence from a mediterranean marine protected area. PLoS ONE 8:e81838. doi: 10.1371/journal.pone.0081838
Gaines, S. D., White, C., Carr, M. H., and Palumbi, S. R. (2010). Designing marine reserve networks for both conservation and fisheries management. PNAS 107, 18286–18293. doi: 10.1073/pnas.0906473107
Galzin, R., Crec'hriou, R., Lenfant, P., and Planes, S. (2004). Marine protected areas: a laboratory for scientific research. Rev. Ecol. Terre. Vie 59, 37–48.
García-Charton, J. A., Barcala-Bellod, E., Cuadros-Casado, A., Orenes-Salazar, V., Pereñíguez-López, J. M., Rojo-Moreno, I., et al. (2019). Estudios de Seguimiento de las Reservas Marinas de Cabo de Palos – Islas Hormigas y Cabo Tiñoso 2018. Servicio de Pesca y Acuicultura, Comunidad Autónoma de la Región de Murcia.
García-Charton, J. A., and Pérez-Ruzafa, A. (1998). Correlation between habitat structure and a rocky reef fish assemblage in the Southwest Mediterranean. Mar. Ecol. 19, 111–128 doi: 10.1111/j.1439-0485.1998.tb00457.x
García-Charton, J. A., and Pérez-Ruzafa, A. (1999). Ecological heterogeneity and the evaluation of the effects of marine reserves. Fish. Res. 42, 1–20. doi: 10.1016/S0165-7836(99)00043-0
García-Charton, J. A., and Pérez-Ruzafa, A. (2001). Spatial pattern and the habitat structure of a Mediterranean rocky reef fish local assemblage. Mar. Biol. 138, 917–934. doi: 10.1007/s002270000524
García-Charton, J. A., Pérez-Ruzafa, A., Marcos, C., Claudet, J., Badalamenti, F., Benedetti-Cecchi, L., et al. (2008). Effectiveness of european atlanto-mediterranean MPAs: do they accomplish the expected effects on populations, communities and ecosystems? J. Nat. Conserv. 16, 193–221. doi: 10.1016/j.jnc.2008.09.007
García-Charton, J. A., Pérez-Ruzafa, A., Sánchez-Jerez, P., Bayle-Sempere, J., Reñones, O., and Moreno, D. (2004). Multi-scale spatial heterogeneity, habitat structure, and the effect of marine reserves on Western Mediterranean rocky reef fish assemblages. Mar. Biol. 144, 161–182. doi: 10.1007/s00227-003-1170-0
Garcia-Rubies, A., Cebrian, E., Schembri, P. J., Evans, J., and Macpherson, E. (2017). “Ecological effects and benefits of Mediterranean marine protected areas: management implications,” in Management of Marine Protected Areas: A Network Perspective, ed P. D. Goriup (Chichester: Wiley-Blackwell), 21–47. doi: 10.1002/9781119075806.ch2
Gelcich, S., Amar, F., Valdebenito, A., Castilla, J. C., Fernández, M., Godoy, C., et al. (2013). Financing marine protected areas through visitor fees: insights from tourist willingness to pay in Chile. Ambio 42, 975–984. doi: 10.1007/s13280-013-0453-z
Gell, F. R., and Roberts, C. M. (2003a). The Fishery Effects of Marine Reserves and Fishery Closures. Washington, DC: WWF-US.
Gell, F. R., and Roberts, C. M. (2003b). Benefits beyond boundaries: the fishery effects of marine reserves. Trends Ecol. Evolut. 18, 448–455. doi: 10.1016/S0169-5347(03)00189-7
Gerber, L. R., Botsford, L. W., Hastings, A., Possingham, H. P., Gaines, S. D., Palumbi, S. R., et al. (2003). Population models for marine reserve design: a retrospective and prospective synthesis. Ecol. Appl. 13, S47–S64. doi: 10.1890/1051-0761(2003)013[0047:PMFMRD]2.0.CO;2
Giakoumi, S., and Pey, A. (2017). Assessing the effects of marine protected areas on biological invasions: a global review. Front. Mar. Sci. 4:49. doi: 10.3389/fmars.2017.00049
Giakoumi, S., Pey, A., Difranco, A., Francour, P., Kizilkaya, Z., Arda, Y., et al. (2019a). Exploring the relationships between marine protected areas and invasive fish in the world's most invaded sea. Ecol. Appl. 29:e01809. doi: 10.1002/eap.1809
Giakoumi, S., Pey, A., Thiriet, P., Francour, P., and Guidetti, P. (2019b). Patterns of predation on native and invasive alien fish in mediterranean protected and unprotected areas. Mar. Environ. Res. 150:104792. doi: 10.1016/j.marenvres.2019.104792
Giakoumi, S., Scianna, C., Plass-Johnson, J., Micheli, F., Grorud-Colvert, K., Thiriet, P., et al. (2017). Ecological effects of full and partial protection in the crowded Mediterranean sea: a regional meta-analysis. Sci. Rep. 7:e8940. doi: 10.1038/s41598-017-08850-w
Gianguzza, P., Chiantore, M., Bonaviri, C., Cattaneo-Vietti, R., Vielmini, I., and Riggio, S. (2006). The effects of recreational Paracentrotus lividus fishing on distribution patterns of sea urchins at Ustica Island MPA (Western Mediterranean, Italy). Fish. Res. 81, 37–44. doi: 10.1016/j.fishres.2006.06.002
Gill, D. A., Mascia, M. B., Ahmadia, G. N., Glew, L., Lester, S. E., Barnes, M., et al. (2017). Capacity shortfalls hinder the performance of marine protected areas globally. Nature 543, 665–669. doi: 10.1038/nature21708
Gilman, E. L. (1997). Community based and multiple purpose protected areas: a model to select and manage protected areas with lessons from the Pacific islands. Coast. Manage. 25, 59–91. doi: 10.1080/08920759709362310
Gómez, S., Lloret, J., Demestre, M., and Riera, V. (2006). The decline of the artisanal fisheries in mediterranean coastal areas: the case of cap de creus (Cape Creus). Coast. Manage. 34, 217–232. doi: 10.1080/08920750500531389
Goñi, R., Adlerstein, S., Alvarez-Berastegui, D., Forcada, A., Reñones, O., Criquet, G., et al. (2008). Spillover from six western mediterranean marine protected areas: evidence from artisanal fisheries. Mar. Ecol. Prog. Ser. 366, 159–174. doi: 10.3354/meps07532
Goñi, R., Badalamenti, F., and Tupper, M. H. (2011). “Chapter 3: effects of marine protected areas on adjacent fisheries: evidence from empirical studies,” in Marine Protected Areas: A Multidisciplinary Approach, ed. J. Claudet (Cambridge: Cambridge University Press), 72–98. doi: 10.1017/CBO9781139049382.006
Goñi, R., Hilborn, R., Díaz, D., Mallol, S., and Adlerstein, S. (2010). Net contribution of spillover from a marine reserve to fishery catches. Mar. Ecol. Prog. Ser. 400, 233–243. doi: 10.3354/meps08419
Goñi, R., Quetglas, A., and Reñones, O. (2003). Size at maturity, fecundity and reproductive potential of a protected population of the spiny lobster Palinurus elephas (Fabricius, 1787) from the Western Mediterranean. Mar. Biol. 143, 583–592. doi: 10.1007/s00227-003-1097-5
Goñi, R., Quetglas, A., and Reñones, O. (2006). Spillover of lobster Palinurus elephas (Fabricius 1787) from a Western mediterranean marine reserve. Mar. Ecol. Prog. Ser. 308, 207–219. doi: 10.3354/meps308207
Goñi, R., Reñones, O., and Quetglas, A. (2001). Dynamics of a protected Western Mediterranean population of the European spiny lobster Palinurus elephas (Fabricius, 1787) assessed by trap surveys. Mar. Freshw. Res. 52, 1577–1587. doi: 10.1071/MF01208
González-Correa, J. M., Bayle Sempere, J. T., Sánchez-Jerez, P., and Valle, C. (2007). Posidonia oceanica meadows are not declining globally. Analysis of population dynamics in marine protected areas of the Mediterranean sea. Mar. Ecol. Prog. Ser. 336, 111–119. doi: 10.3354/meps336111
González-Correa, J. M., Sánchez-Lizaso, J. L., Fernández-Torquemada, Y., and Forcada, A. (2015). Long-term population dynamics in a healthy Posidonia oceanica meadow. Thalassas 31, 63–72.
Gormley, A. M., Slooten, E., Dawson, S., Barker, R. J., Rayment, W., du Fresne, S., et al. (2012). First evidence that marine protected areas can work for marine mammals. J. Appl. Ecol. 49, 474–480. doi: 10.1111/j.1365-2664.2012.02121.x
Graham, N. A. J., and McClanahan, T. R. (2013). The last call for marine wilderness? BioScience 63, 397–402. doi: 10.1525/bio.2013.63.5.13
Green, A. L., Maypa, A. P., Almany, G. R., Rhodes, K. L., Weeks, R., Abesamis, R. A., et al. (2015). Larval dispersal and movement patterns of coral reef fishes, and implications for marine reserve network design. Biol. Rev. 90, 1215–1247. doi: 10.1111/brv.12155
Grorud-Colvert, K., Claudet, J., Tissot, B. N., Caselle, J. E., Carr, M. H., Day, J. C., et al. (2014). Marine protected area networks: assessing whether the whole is greater than the sum of its parts. PLoS ONE 9:e102298. doi: 10.1371/journal.pone.0102298
Grüss, A. (2014). Modelling the impacts of marine protected areas for mobile exploited fish populations and their fisheries: what we recently learnt and where we should be going. Aquat. Living Res. 27, 107–133. doi: 10.1051/alr/2014013
Grüss, A., Kaplan, D. M., Guénette, S., Roberts, C. M., and Botsford, L. W. (2011). Consequences of adult and juvenile movement for marine protected areas. Biol. Conserv. 144, 692–702. doi: 10.1016/j.biocon.2010.12.015
Guidetti, P. (2006). Marine reserves reestablish lost predatory interactions and cause community changes in rocky reefs. Ecol. Appl. 16, 963–976. doi: 10.1890/1051-0761(2006)016[0963:MRRLPI]2.0.CO;2
Guidetti, P., Milazzo, M., Bussotti, S., Molinari, A., Murenu, M., Pais, A., et al. (2008). Italian marine reserve effectiveness: does enforcement matter? Biol. Conserv. 141, 699–709. doi: 10.1016/j.biocon.2007.12.013
Guidetti, P., and Sala, E. (2007). Community-wide effects of marine reserves in the Mediterranean sea. Mar. Ecol. Prog. Ser. 335, 43–56. doi: 10.3354/meps335043
Hackradt, C. W. (2012). Population Ecology and Mobility Patterns of Groupers (Serranidae: Epinephelinae) on Temperate Rocky Reefs on Southwestern Mediterranean Sea: Implications for their Conservation. [Ph.D., thesis], University of Murcia, Murcia.
Hackradt, C. W., García-Charton, J. A., Harmelin-Vivien, M., Pérez-Ruzafa, A., Le Diréach, L., Bayle-Sempere, J., et al. (2014). Response of rocky reef top predators (Serranidae: Epinephelinae) in and around marine protected areas in the Western mediterranean sea. PLoS ONE 9:e9820. doi: 10.1371/journal.pone.0098206
Haines, R., Hattam, C., Pantzar, M., and Russi, D. (2018). Study on the Economic Benefits of MPAs. Final report. Luxembourg: Publications Office of the European Union.
Haines-Young, R., and Potschin, M. (2013). Common International Classification of Ecosystem Services (CICES): Consultation on Version 4, August-December 2012. EEA Framework Contract No EEA/IEA/09/003
Haines-Young, R., and Potschin, M. (2016). “Categorisation systems: the classification challenge,” in Mapping Ecosystem Services, ed B. Burkhard, and J. Maes (Nottingham; Sofia: Pensoft Publishers), 42–45.
Halpern, B. S. (2003). The impact of marine reserves: Do reserves work and does reserve size matter? Ecol. Appl. 13, 117–137. doi: 10.1890/1051-0761(2003)013[0117:TIOMRD]2.0.CO;2
Halpern, B. S., Lester, S. E., and McLeod, K. L. (2010). Placing marine protected areas onto the ecosystem based management seascape. PNAS 107, 18312–18317. doi: 10.1073/pnas.0908503107
Hameed, S. O., Cornick, L. A., Devillers, R., and Morgan, L. E. (2017). Incentivizing more effective marine protected areas with the global ocean refuge system (GLORES). Front. Mar. Sci. 4, 208. doi: 10.3389/fmars.2017.00208
Harmelin-Vivien, M., García-Charton, J. A., Bayle-Sempere, J., Charbonnel, E., Le Diréach, L., Ody, D., et al. (2007). “Importance of marine reserves for the population dynamics of groupers (Epinephelinae) in the Western Mediterranean,” in Second Symposium on Mediterranean Groupers (Nice), 91–93.
Harmelin-Vivien, M., Le Diréach, L., Bayle-Sempere, J., Charbonnel, E., García-Charton, J. A., Ody, D., et al. (2008). Gradients of abundance and biomass across reserve boundaries in six Mediterranean marine protected areas: evidence of fish spillover? Biol. Conserv. 141, 1829–1839. doi: 10.1016/j.biocon.2008.04.029
Hensel, E., Allgeier, J. E., and Layman, C. A. (2019). Effects of predator presence and habitat complexity on reef fish communities in the Bahamas. Mar. Biol. 166:136. doi: 10.1007/s00227-019-3568-3
Hinz, H., Reñones, O., Gouraguine, A., Johnson, A. F., and Moranta, J. (2019). Fish nursery value of algae habitats in temperate coastal reefs. PeerJ 7:e6797. doi: 10.7717/peerj.6797
Hixon, M. A., Johnson, D. W., and Sogard, S. M. (2014). BOFFFFs: on the importance of conserving old-growth age structure in fishery populations. ICES J. Mar. Sci. 71, 2171–2185. doi: 10.1093/icesjms/fst200
Hooker, S. K., Cañadas, A., Hyrenbach, K. D., Corrigan, C., Polovina, J. J., and Reeves, R. R. (2011). Making protected area networks effective for marine top predators. Endanger. Species Res. 13, 203–218. doi: 10.3354/esr00322
Hopf, J. K., Jones, G. P., Williamson, D. H., and Connolly, S. R. (2016). Fishery consequences of marine reserves: short-term pain for longer-term gain. Ecol. Appl. 26, 818–829. doi: 10.1890/15-0348
Hoskin, M. G., Coleman, R. A., von Carlshausen, E., and Davis, C. M. (2011). Variable population responses by large decapod crustaceans to the establishment of a temperate marine no-take zone. Canad. J. Fish. Aquat. Sci. 68, 185–200. doi: 10.1139/F10-143
Howard, J., McLeod, E., Thomas, S., Eastwood, E., Fox, M., Wenzel, L., et al. (2017). The potential to integrate blue carbon into MPA design and management. Aquat. Conser. Mar. Freshw. Ecosyst. 27, 100–115. doi: 10.1002/aqc.2809
Howarth, L. M., Wood, H. L., Turner, A. P., and Beukers-Stewart, B. D. (2011). Complex habitat boosts scallop recruitment in a fully protected marine reserve. Mar. Biol. 158, 1767–1780. doi: 10.1007/s00227-011-1690-y
Huserbråten, M. B. O., Moland, E., Knutsen, H., Olsen, E. M., André, C., and Stenseth, N. C. (2013). Conservation, spillover and gene flow within a network of northern european marine protected areas. PLoS ONE 8:e73388. doi: 10.1371/journal.pone.0073388
IUCN Standards and Petitions Working Group. (2018). Guidelines for Using the IUCN Red List Categories and Criteria Version 7.0.
Jacquet, S. (1999). Etude de “l'effet réserve” de la Réserve Naturelle Marine de Cerbère-Banyuls à l'aide de l'espèce indicatrice: Mytilus galloprovincialis. Diplôme d'ingénieur ISIM (Montpellier. France) (45pp. +3 annexes).
Jessopp, M. J., and McAllen, R. J. (2007). Water retention and limited larval dispersal: implications for short and long-distance dispersers in marine reserves. Mar. Ecol. Prog. Ser. 333, 27–36. doi: 10.3354/meps333027
Jiao, J., Pilyugin, S. S., Riotte-Lambert, L., and Osenberg, C. W. (2018). Habitat-dependent movement rate can determine the efficacy of marine protected areas. Ecology 99, 2485–2495. doi: 10.1002/ecy.2477
Johnston, R. J., Schultz, E. T., Segerson, K., Besedin, E. Y., and Ramachandran, M. (2012). Enhancing the content validity of stated preference valuation: the structure and function of ecological indicators. Land Econ. 88, 102–120. doi: 10.3368/le.88.1.102
Jones, P. J. S. (2014). Governing Marine Protected Areas. Resilience Through Diversity. Oxon: Routledge. doi: 10.4324/9780203126295
Kaiser, M. J., Collie, J. S., Hall, S. J., Jennings, S., and Poiner, I. R. (2002). Modification of marine habitats by trawling activities: prognosis and solutions. Fish Fish. 3, 114–136. doi: 10.1046/j.1467-2979.2002.00079.x
Kaplan, K. A., Yamane, L., Botsford, L. W., Baskett, M., Hastings, A., Worden, S., et al. (2019). Setting expected timelines of fished population recovery for the adaptive management of a marine protected area network. Ecol. Appl. 29, e01949. doi: 10.1002/eap.1949
Kay, M. C., and Wilson, J. R. (2012). Spatially explicit mortality of California spiny lobster (Panulirus interruptus) across a marine reserve network. Environ. Conserv. 39, 215–224. doi: 10.1017/S0376892911000695
Kellner, J.B., Tetreault, I., Gaines, S.D., and Nisbet, R.M. (2007). Fishing the line near marine reserves in single and multispecies fisheries. Ecol. Appl. 17, 1039–1054. doi: 10.1890/05-1845
Kelly, S., Scott, D., MacDiarmid, A. B., and Babcock, R. C. (2000). Spiny lobster, Jasus edwardsii, recovery in New Zealand marine reserves. Biol. Conserv. 92, 359–369. doi: 10.1016/S0006-3207(99)00109-3
Kenchington, R., Ward, T., and Hegerl, E. (2003). The Benefits of Marine Protected Areas. Technical Report. Commonwealth of Australia.
Kersting, D., and Ballesteros, E. (2010). Gobius kolombatovici, Primera Cita en las Costas Ibéricas, Islas Columbretes (Mediterráneo Noroccidental). Alicante: XVI Simposio Ibérico de Estudios de Biología Marina, 181.
Kersting, D., and García-March, J. R. (2017). Long-term assessment of recruitment, early stages and population dynamics of the endangered Mediterranean fan mussel Pinna nobilis in the Columbretes islands (NW Mediterranean). Mar. Environ. Res. 130, 282–292. doi: 10.1016/j.marenvres.2017.08.007
Kersting, D. K., Ballesteros, E., De Caralt, S., and Linares, C. (2014). Invasive macrophytes in a marine reserve (Columbretes islands, NW Mediterranean): spread dynamics and interactions with the endemic scleractinian coral Cladocora caespitosa. Biol. Invas. 16, 1599–1610. doi: 10.1007/s10530-013-0594-9
Klein, C. J., Chan, A., Kircher, L., Cundiff, A. J., Gardner, N., Hrovat, Y., et al. (2008). Striking a balance between biodiversity conservation and socioeconomic viability in the design of marine protected areas. Conserv. Biol. 22, 691–700. doi: 10.1111/j.1523-1739.2008.00896.x
Kramer, D. L., and Chapman, M. R. (1999). Implications of fish home range size and relocation for marine reserve function. Environ. Biol. Fish. 55, 65–79. doi: 10.1023/A:1007481206399
Kroeker, K. J., Carr, M. H., Raimondi, P. T., Caselle, J. E., Washburn, L., Palumbi, S. R., et al. (2019). Assessing the potential role of marine protected areas and fisheries management approaches for resilience management in a changing ocean. Oceanography 32, 116–125. doi: 10.5670/oceanog.2019.318
Krueck, N. C., Ahmadia, G. N., Possingham, H. P., Riginos, C., Treml, E. A., and Mumby, P. J. (2017). Marine reserve targets to sustain and rebuild unregulated fisheries. PLoS Biol. 15:e2000537. doi: 10.1371/journal.pbio.2000537
Kumar, P., (ed.). (2010). The Economics of Ecosystems and Biodiversity (TEEB): Ecological and Economic Foundations. London: Earthscan; Routledge.
La Notte, A., D'Amato, D., Mäkinen, H., Paracchini, M. L., Liquete, C., Egoh, B., et al. (2017). Ecosystem services classification: a systems ecology perspective of the cascade framework. Ecol. Indicat. 74, 392–402. doi: 10.1016/j.ecolind.2016.11.030
Lafferty, K. D., Goodman, D., and Sandoval, C. P. (2006). Restoration of breeding by snowy plovers following protection from disturbance. Biodivers. Conserv. 15, 2217–2230. doi: 10.1007/s10531-004-7180-5
Langebrake, J., Riotte-Lambert, L., Osenberg, C. W., and Leenheer, P. D. (2012). Differential movement and movement bias models for marine protected areas. J. Mathe. Biol. 64, 667–696. doi: 10.1007/s00285-011-0407-7
Le Port, A., Montgomery, J. C., Smith, A. N. H., Croucher, A. E., McLeod, I. M., and Lavery, S. D. (2017). Temperate marine protected area provides recruitment subsidies to local fisheries. Proc. R. Soc. B Biol. Sci. 284:20171300. doi: 10.1098/rspb.2017.1300
Lecchini, D., Lenfant, P., and Planes, S. (2002). Estimation of reserve effect of sea urchin, Paracentrotus lividus in the marine protected area of cerbère-banyuls. Vie Milieu 52, 111–118.
Ledoux, L., and Turner, R. K. (2002). Valuing ocean and coastal resources: a review of practical examples and issues for further action. Ocean Coast. Manage. 45, 583–616. doi: 10.1016/S0964-5691(02)00088-1
Lee, C. H., Chen, Y. J., and Chen, C. W. (2019). Assessment of the economic value of ecological conservation of the kenting coral reef. Sustainability 11:5869. doi: 10.3390/su11205869
Leenhardt, P., Low, N., Pascal, N., Micheli, F., and Claudet, J. (2015). “Chapter 9 – the role of marine protected areas in providing ecosystem services,” in Aquatic Functional Biodiversity, eds A. Belgrano, G. Woodward, and U. Jacob (Academic Press), 211–239. doi: 10.1016/B978-0-12-417015-5.00009-8
Lenfant, P. (2003). Demographic and genetic structure of white seabream populations ( Diplodus sargus, Linnaeus, 1758) inside and outside a Mediterranean marine reserve. Compte Rendu Acad. Sci. Biol. 326, 751–760. doi: 10.1016/S1631-0691(03)00168-9
Lenfant, P., Lefranc, C., Pastor, J., Crec'hriou, R., Saragoni, G., and Payrot, J. (2016). Suivi des peuplements de poissons au sein de la réserve naturelle marine de Cerbère Banyuls et des zones périphériques, année 2016. CEFREM (UPVD-CNRS) - CG66, 57.
Lenfant, P., Louisy, P., and Licari, M. L. (2003). Recensement des mérous bruns (Epinephelus marginatus) de la réserve naturelle de Cerbère-Banyuls (France, Méditerranée Nord Occidentale). Cybium 27, 27–36.
Lester, S. E., Halpern, B. S., Grorud-Colvert, K., Lubchenco, J., Ruttenberg, B. I., Gaines, S. D., et al. (2009). Biological effects within no-take marine reserves: a global synthesis. Mar. Ecol. Prog. Ser. 384, 33–46. doi: 10.3354/meps08029
Libralato, S., Coll, M., Tempesta, M., Santojanni, A., Spoto, M., Palomera, I., et al. (2010). Food-web traits of protected and exploited areas of the Adriatic sea. Biol. Conserv. 143, 2182–2194. doi: 10.1016/j.biocon.2010.06.002
Linares, C., Cebria, E., and Coma, R. (2012). Effects of turf algae on recruitment and juvenile survival of gorgonian corals. Mar. Ecol. Prog. Ser. 452, 81–88. doi: 10.3354/meps09586
Linares, C., Coma, R., and Zabala, M. (2008). Restoration of threatened red gorgonian populations: an experimental and modelling approach. Biol. Conserv. 141, 427–437. doi: 10.1016/j.biocon.2007.10.012
Linares, C., Vidal, M., Canals, M., Kersting, D. K., Amblas, D., Aspillaga, E., et al. (2015). Persistent natural acidification drives major distribution shifts in marine benthic ecosystems. Proc. R Soc. B 282:20150587. doi: 10.1098/rspb.2015.0587
Ling, S. D., Johnson, C. R., Frusher, S. D., and Ridgway, K. R. (2009). Overfishing reduces resilience of kelp beds to climate-driven catastrophic phase shift. Proc. Natl. Acad. Sic. U.S.A. 106, 22341–22345. doi: 10.1073/pnas.0907529106
Lloret, J., Zaragoza, N., Caballero, D., and Riera, V. (2008). Impacts of recreational boating on the marine environment of cap de creus (Mediterranean sea). Ocean Coast. Manage. 51, 749–754. doi: 10.1016/j.ocecoaman.2008.07.001
Loh, T. L., and Pawlik, J. R. (2014). Chemical defenses and resource trade-offs structure sponge communities on caribbean coral reefs. Proc. Natl. Acad. Sic. U.S.A. 111, 4151–4156. doi: 10.1073/pnas.1321626111
Long, R. D., Charles, A., and Stephenson, R. L. (2015). Key principles of marine ecosystem-based management. Mar. Policy 57, 53–60. doi: 10.1016/j.marpol.2015.01.013
Lubchenco, J., Palumbi, S. R., Gaines, S. D., and Andelman, S. (2003). Plugging a hole in the ocean: the emerging science of marine reserves. Ecol. Appl. 13, S3–S7. doi: 10.1890/1051-0761(2003)013[0003:PAHITO]2.0.CO;2
MacDiarmid, A. B., and Breen, P. A. (1993). “Spiny lobster population changes in a marine reserve,” in Proceedings of the Second International Temperate Reef Symposium, eds. C. N. Battershill, D.R. Schiel, G. P. Jones, R. G. Creese, and A. B. MacDiarmid (Wellington: NIWA Marine), 47–56.
Mace, G. M., Norris, K., and Fitter, A. H. (2012). Biodiversity and ecosystem services: a multilayered relationship. Trends Ecol. Evolut. 27, 19–26. doi: 10.1016/j.tree.2011.08.006
Maggs, J. Q., Mann, B. Q., and Cowley, P. D. (2013). Reef fish display station-keeping and ranging behaviour in the pondoland marine protected area on the east coast of South Africa. Afr. J. Mar. Sci. 35, 183–193. doi: 10.2989/1814232X.2013.798152
Magris, R. A., Andrello, M., Pressey, R. L., Mouillot, D., Dalongeville, A., Martin, N., et al. (2018). Biologically representative and well-connected marine reserves enhance biodiversity persistence in conservation planning. Conserv. Lett. 11:e12439. doi: 10.1111/conl.12439
Manel, S., Loiseau, N., Andrello, M., Fietz, K., Goñi, R., Forcada, A., et al. (2019). Long-distance benefits of marine reserves: myth or reality? Trends Ecol. Evolut. 34, 342–354. doi: 10.1016/j.tree.2019.01.002
Mangos, A., and Claudot, M. A. (2013). Economic Study of the Impacts of Marine and Coastal Protected Areas in the Mediterranean. Valbonne: Plan Bleu Papers, 13. Plan Bleu
Margalef, R. (1997). “Our Biosphere,” in Excellence in Ecology Series, ed O. Kinne. Oldendorf: Ecology Institute.
Marshall, D. J., Gaines, S., Warner, R., Barneche, D. R., and Bode, M. (2019). Underestimating the benefits of marine protected areas for the replenishment of fished populations. Front. Ecol. Environ. 17, 407–413. doi: 10.1002/fee.2075
Mazaris, A. D., Kallimanis, A., Gissi, E., Pipitone, C., Danovaro, R., Claudet, J., et al. (2019). Threats to marine biodiversity in European protected areas. Sci. Total Environ. 677, 418–426. doi: 10.1016/j.scitotenv.2019.04.333
McClanahan, T. R., and Mangi, S. (2000). Spillover of exploitable fishes from a marine park and its effect on the adjacent fishery. Ecol. Appl. 10, 1792–1805. doi: 10.1890/1051-0761(2000)010[1792:SOEFFA]2.0.CO;2
McClure, E. C., Sievers, K. T., Abesamis, R. A., Hoey, A. S., Alcala, A. C., and Russ, G. R. (2020). Higher fish biomass inside than outside marine protected areas despite typhoon impacts in a complex reefscape. Biol. Conserv. 241, 108354. doi: 10.1016/j.biocon.2019.108354
McDougall, C. (2017). Erosion and the beaches of Negril. Ocean Coast. Manage. 148, 204–213. doi: 10.1016/j.ocecoaman.2017.08.008
MedPAN SPA/RAC. (2021). MAPAMED, the Database of MArine Protected Areas in the MEDiterranean - Release Notes for the 2019 edition. MedPAN and SPA/RAC. Available online at: http://www.mapamed.org/
Meier, C. (2003). Comparaison inter-annuelles et intersites des effectifs de Patella ferruginea dans la RNBB. Corte: IUP Génie de l'Environnement; Université de Corse.
Mellin, C., MacNeil, M. A., Cheal, A. J., Emslie, M. J., and Caley, M. J. (2016). Marine protected areas increase resilience among coral reef communities. Ecol. Lett. 19, 629–637. doi: 10.1111/ele.12598
Mello, H. L., Smith, A. M., Wood, A. C. L., and Tidey, E. J. (2020). Enhanced biodiversity and abundance of benthic invertebrate macrofauna in a New Zealand marine reserve. Aquat. Conserv. Mar. Freshw. Ecosyst. 30, 1854–1867. doi: 10.1002/aqc.3333
Mendoza, J. C., Clemente, S., and Hernández, J. C. (2020). Modeling the role of marine protected areas on the recovery of shallow rocky reef ecosystem after a catastrophic submarine volcanic eruption. Mar. Environ. Res. 155:104877. doi: 10.1016/j.marenvres.2020.104877
Micheli, F., Benedetti-Cecchi, L., Gambaccini, S., Bertocci, I., Borsini, C., Osio, G. C., et al. (2005). Cascading human impacts, marine protected areas, and the structure of mediterranean reef assemblages. Ecol. Monogr. 75, 81–102. doi: 10.1890/03-4058
Millennium Ecosystem Assessment (2005). Ecosystems and Human Well-being: Synthesis. World Resources Institute. Washington, DC: Island Press.
Ministry for the Environment (2016). A New Marine Protected Areas Act: Consultation Document. Wellington: Ministry for the Environment.
Moher, D., Liberati, A., Tetzlaff, J., Altman, D. G., and The PRISMA Group (2009). Preferred reporting items for systematic reviews and meta-analyses: the PRISMA statement. PLoS Med. 6:e1000097. doi: 10.1371/journal.pmed.1000097
Montefalcone, M., Albertelli, G., Morri, C., Parravicini, V., and Bianchi, C. N. (2009). Legal protection is not enough: Posidonia oceanica meadows in marine protected areas are not healthier than those in unprotected areas of the northwest Mediterranean sea. Mar. Pollut. Bull. 58, 515–519. doi: 10.1016/j.marpolbul.2008.12.001
Morey, G., Coll, J., Navarro, O., and Verger, F. (2018). La Reserva Marina del Llevant de Mallorca. Seguiment de les Poblacions de Peixos Vulnerables de Substrat Rocós en el Període 2008-2018. Madrid: Direcció General de Pesca i Medi Marí-Govern de les Illes Balears – Tragsatec.
Mouillot, D., Bellwood, D. R., Baraloto, C., Chave, J., Galzin, R., Harmelin-Vivien, M., et al. (2013). Rare species support vulnerable functions in high-diversity ecosystems. PLoS Biol. 11:e1001569. doi: 10.1371/journal.pbio.1001569
Muñoz, M., Lloret, J., and Vila, S. (2013). Effects of artisanal fisheries on the scorpaenids (Scorpaena spp.) reproduction in the marine protected area of Cap de Creus (NW Mediterranean). Fish. Res. 138, 146–151. doi: 10.1016/j.fishres.2012.07.023
Muñoz, M., Reul, A., García-Martínez, M. D. C., Plaza, F., Bautista, B., Moya, F., et al. (2018). Oceanographic and bathymetric features as the target for pelagic MPA Design: a case study on the cape of Gata. Water 10:1403. doi: 10.3390/w10101403
Murawski, S. A., Wigley, S. E., Fogarty, M. J., Rago, P. J., and Mountain, D. G. (2005). Effort distribution and catch patterns adjacent to temperate MPAs. ICES J. Mar. Sci. 62, 1150–1167. doi: 10.1016/j.icesjms.2005.04.005
Naylor, L. A., and Viles, H. A. (2002). A new technique for evaluating short-term rates of coastal bioerosion and bioprotection. Geomorphology 47, 31–44. doi: 10.1016/S0169-555X(02)00139-3
Newton, A., Brito, A. C., Icely, J. D., Delorez, V., Clara, I., Angus, S., et al. (2018). Assessing, quantifying and valuing the ecosystem services of coastal lagoons. J. Nat. Conserv. 44, 50–65. doi: 10.1016/j.jnc.2018.02.009
Niccolini, F., Marzo, D., Scipioni, S., Randone, M., Hogg, K., and Gomei, M. (2019). Ecosystem Services, Socio-Economic Values and Organizational Profiles of Mediterranean MPAs. Rome: WWF.
Nickols, K. J., White, J. W., Malone, D., Carr, M. H., Starr, R. M., Baskett, M. L., et al. (2019). Setting ecological expectations for adaptive management of marine protected areas. J. Appl. Ecol. 56, 2376–2385. doi: 10.1111/1365-2664.13463
O'Hara, C. C., Villaseñor-Derbez, J. C., Ralph, G. M., and Halpern, B. S. (2019). Mapping status and conservation of global at-risk marine biodiversity. Conserv. Lett. 12:e12651. doi: 10.1111/conl.12651
Ojeda-Martínez, C., Bayle-Sempere, J. T., Sánchez-Jerez, P., Forcada, A., and Valle, C. (2007). Detecting conservation benefits in spatially protected fish populations with meta-analysis of long-term monitoring data. Mar. Biol. 151, 1153–1161. doi: 10.1007/s00227-006-0557-0
Olander, L. P., Johnston, R. J., Tallis, H., Kagan, J., Maguire, L. A., Polasky, S., et al. (2018). Benefit relevant indicators: Ecosystem services measures that link ecological and social outcomes. Ecol. Indicat. 85, 1262–1272. doi: 10.1016/j.ecolind.2017.12.001
O'Leary, C. (2006). Spatial Distribution and Export of Ichthyoplankton from Mediterranean Marine Protected Areas. [Ph.D., tesis], University College Dublin, Dublin.
Ospina-Alvarez, A., de Juan, S., Alós, J., Basterretxea, G., Alonso-Fernández, A., Follana-Berná, G., et al. (2020). MPA network design based on graph theory and emergent properties of larval dispersal. Mar. Ecol. Prog. Ser. 650, 309–326. doi: 10.3354/meps13399
O'Sullivan, D., and Emmerson, M. (2011). Marine reserve designation, trophic cascades and altered community dynamics. Mar. Ecol. Prog. Ser. 440, 115–125. doi: 10.3354/meps09294
Otero, M., Cebrian, E., Francour, P., Galil, B., and Savini, D. (2013). Monitoring Marine Invasive Species in Mediterranean Marine Protected Areas (MPAs): A strategy and practical guide for managers. Málaga: IUCN.
Padiglia, A., Cadeddu, B., Ledda, F. D., Bertolino, M., Costa, G., Pronzato, R., et al. (2018). Biodiversity assessment in Western Mediterranean marine protected areas (MPAs): porifera of Posidonia oceanica meadows (Asinara island MPA) and marine caves (Capo Caccia–Isola Piana MPA) of Sardinia. Euro. Zool. J. 85, 409–422. doi: 10.1080/24750263.2018.1525440
Pais, A., Chessa, L. A., Serra, S., Ruiu, A., Meloni, G., and Donno, Y. (2007). The impact of commercial and recreational harvesting for Paracentrotus lividus on shallow rocky reef sea urchin communities in North-western Sardinia, Italy. Estuar. Coast. Shelf Sci. 73, 589–597. doi: 10.1016/j.ecss.2007.02.011
Palumbi, S. R. (2003). Population genetics, demographic connectivity, and the design of marine reserves. Ecol. Appl. 13, 146–158. doi: 10.1890/1051-0761(2003)013[0146:PGDCAT]2.0.CO;2
Palumbi, S. R. (2004). Marine reserves and ocean neighborhoods: the spatial scale of marine populations and their management. Annu. Rev. Environ. Res. 29, 31–68. doi: 10.1146/annurev.energy.29.062403.102254
Pantzar, M., Russi, D., Hooper, T., and Haines, R. (2018). Study on the Economic Benefits of Marine Protected Areas, Literature Review Analisis. ICF Consulting Services Limited, IEEP and PML. Executive Agency for Small and Medium-sized Enterprises (EASME) Contract No EASME/EMFF/2015/1.3.1.8/SI2.737373. European Union. Brussels: Publications Office of the European Union.
Pastor, J., Verdoit-Jarraya, M., Astruch, P., Dalias, N., Nelva Pasqual, J. S., Saragoni, G., et al. (2009). Acoustic telemetry survey of the dusky grouper (Epinephelus marginatus) in the marine reserve of Cerbère-Banyuls: informations on the territoriality of this emblematic species. Compte Rendu Biol. 332, 732–740. doi: 10.1016/j.crvi.2009.03.010
Pendleton, L. H., Ahmadia, G. N., Browman, H. I., Thurstan, R. H., Kaplan, D. M., and Bartolino, V. (2018). Debating the effectiveness of marine protected areas. ICES J. Mar. Sci. 75, 1156–1159. doi: 10.1093/icesjms/fsx154
Pérez-Burgos, J. M. (2012). Tabarca. Utopía y Realidad. Revista Canelobre, Instituto Alicantino de Cultura “Juan Gil-Albert.”
Pérez-Ruzafa, A. (2003). “Los recursos pesqueros en la Región de Murcia,” in Los Recursos Naturales de la Región de Murcia. Un Análisis Interdisciplinar, eds M. A. Esteve, M. Lloréns, M., and C. Martínez Gallur (Murcia: Servicio de Publicaciones de la Universidad de Murcia), 412–417.
Pérez-Ruzafa, A., García-Charton, J. A., and Marcos, C. (2017). North East Atlantic vs. mediterranean marine protected areas as fisheries management tool. Front. Mar. Sci. 4:245. doi: 10.3389/fmars.2017.00245
Pérez-Ruzafa, A., González-Wangüemert, M., Lenfant, P., Marcos, C., and García-Charton, J. A. (2006). Effects of fishing protection on the genetic structure of fish populations. Biol. Conserv. 129, 244–255. doi: 10.1016/j.biocon.2005.10.040
Pérez-Ruzafa, A., Marcos, C., García-Charton, J. A., and Salas, F. (2008b). European marine protected areas (MPAs) as tools for fisheries management and conservation. J. Nat. Conserv. 16, 187–192. doi: 10.1016/j.jnc.2008.09.008
Pérez-Ruzafa, A., Martín, E., Marcos, C., Zamarro, J. M., Stobart, B., Harmelin-Vivien, M., et al. (2008a). Modelling spatial and temporal scales for spill-over and biomass exportation from MPAs and their potential for fisheries enhancement. J. Nat. Conserv. 16, 234–255. doi: 10.1016/j.jnc.2008.09.003
Pérez-Ruzafa, A., Pérez-Marcos, M., and Marcos, C. (2018). From fish physiology to ecosystems management: Keys for moving through biological levels of organization in detecting environmental changes and anticipate their consequences. Ecol. Indicat. 90, 334–345 doi: 10.1016/j.ecolind.2018.03.019
Pérez-Ruzafa, A., Pérez-Ruzafa, I., Newton, A., and Marcos, C. (2019). “Coastal lagoons: environmental variability, ecosystem complexity and goods and services uniformity,” in Coasts and Estuaries, The Future, eds E. Wolanski, J. Day, M. Elliott, and R. Ramesh (Amsterdam: Elsevier), 253–276.
Pieraccini, M., Coppa, S., and De Lucia, G. A. (2017). Beyond marine paper parks? Regulation theory to assess and address environmental non-compliance. Aquat. Conserv. 27, 177–196. doi: 10.1002/aqc.2632
Pinnegar, J. K., Polunin, N. V. C., Francour, P., Badalamenti, F., Chemello, R., Harmelin-Vivien, M., et al. (2000). Trophic cascades in benthic marine ecosystems: lessons for fisheries and protected-area management. Environ. Conserv. 27, 179–200. doi: 10.1017/S0376892900000205
Plan Development Team (1990). The Potential of Marine Fishery Reserves for Reef Fish Management in the U.S. Southern Atlantic. Miami, FL: NOAA Technical Memorandum NMFS-SEFC-261.
Planes, S., Galzin, R., Garcia-Rubies, R. A., Goñi, R., Harmelin, J.-G., Le Diréach, L., et al. (2000). Effects of marine protected areas on recruitment processes with special reference to Mediterranean littoral ecosystems. Environ. Conserv. 27, 126–143. doi: 10.1017/S0376892900000175
Planes, S., García-Charton, J. A., Marcos, C., and Pérez-Ruzafa, A. (2008). Ecological Effects of Atlanto-Mediterranean Marine Protected Areas in the European Union. EMPAFISH Project, Booklet No. 1. Murcia: Editum.
Pomeroy, R. S., Parks, J. E., and Watson, L. M. (2004). How Is Your MPA Doing? A Guidebook of Natural and Social Indicators for Evaluating Marine Protected Area Management Effectiveness. Gland; Cambridge: IUCN. doi: 10.2305/IUCN.CH.2004.PAPS.1.en
Potts, T., Burdon, D., Jackson, E., Atkins, J., Saunders, J., Hastings, E., et al. (2014). Do marine protected areas deliver flows of ecosystem services to support human welfare? Mar. Policy 44, 139–148. doi: 10.1016/j.marpol.2013.08.011
Prosser, D. J., Nagel, J. L., Howlin, S., Marbán, P. R., Day, D. D., and Erwin, R. M. (2017). Effects of local shoreline and subestuary watershed condition on waterbird community integrity: influences of geospatial scale and season in the Chesapeake bay. Estuar. Coasts 41, 207–222. doi: 10.1007/s12237-017-0288-0
Pujolar, J. M., Schiavina, M., Di Franco, A., Meli,à, P., Guidetti, P., Gatto, M., et al. (2013). Understanding the effectiveness of marine protected areas using genetic connectivity patterns and lagrangian simulations. Diversi. Distribut. 19, 1531–1542. doi: 10.1111/ddi.12114
Putz, A., and Proksch, P. (2010). Chemical defence in marine ecosystems. Funct. Biotechnol. Plant Second. Metab. 162–213. doi: 10.1002/9781444318876.ch3
Rakitin, A., and Kramer, D. L. (1996). Effect of a marine reserve on the distribution of coral reef fishes in Barbados. Mar. Ecol. Prog. Ser. 131, 97–113. doi: 10.3354/meps131097
Ramalho, S. P., Lins, L., Soetaert, K., Lampadariou, N., Cunha, M. R., Vanreusel, A., et al. (2020). Ecosystem functioning under the influence of bottom-trawling disturbance: an experimental approach and field observations from a continental slope area in the west iberian margin. Front. Mar. Sci. 7:457. doi: 10.3389/fmars.2020.00457
Raymundo, L. J., Halford, A. R., Maypa, A. P., and Kerr, A. M. (2009). Functionally diverse reef-fish communities ameliorate coral disease. Proc. Natl. Acad. Sci. 106, 17067–17070. doi: 10.1073/pnas.0900365106
Rees, S. E., Mangi, S. C., Hattam, C., Gall, S. C., Rodwell, L. D., Peckett, F. J., et al. (2015). The socio-economic effects of a marine protected area on the ecosystem service of leisure and recreation. Mar. Policy 62, 144–152. doi: 10.1016/j.marpol.2015.09.011
Reker, J. (2015). Marine Protected Areas in Europe's Seas. An Overview and Perspectives for the Future. Luxembourg: EEA Report 3/2015, Publications Office of the European Union.
Reuchlin-Hugenholtz, E., and McKenzie, E. (2015). Marine Protected Areas: Smart Investments in Ocean Health. Gland: WWF.
Rilov, G., Mazaris, A. D., Stelzenmüller, V., Helmuth, B., Wahl, M., Guy-Haim, T., et al. (2019). Adaptive marine conservation planning in the face of climate change: what can we learn from physiological, ecological and genetic studies? Glob. Ecol. Conserv. 17:e00566. doi: 10.1016/j.gecco.2019.e00566
Roberts, C. M., Bohnsack, J. A., Gell, F., Hawkins, J. P., and Goodridge, R. (2001). Effects of marine reserves on adjacent fisheries. Science 294, 1920–1923. doi: 10.1126/science.294.5548.1920
Roberts, C. M., and Hawkins, J. P. (2000). Fully-Protected Marine Reserves: A Guide. Washington, DC: WWF Endangered Seas Campaign and University of York.
Roberts, C. M., O'Leary, B. C., McCauley, D. J., Cury, P. M., Duarte, C. M., Lubchenco, J., et al. (2017). Marine reserves can mitigate and promote adaptation to climate change. PNAS 114, 6167–6175. doi: 10.1073/pnas.1701262114
Rolim, F. A., Langlois, T., Rodrigues, P. F. C., Bond, T., Motta, F. S., Neves, L. M., et al. (2019). Network of small no-take marine reserves reveals greater abundance and body size of fisheries target species. PLoS ONE 14:e0204970. doi: 10.1371/journal.pone.0204970
Roncin, N., Alban, F., Charbonnel, E., Crec'hriou, R., de la Cruz Modino, R., Culioli, J. M., et al. (2008). Uses of ecosystem services provided by MPAs: how much do they impact the local economy? A southern Europe perspective. J. Nat. Conserv. 16, 256–270. doi: 10.1016/j.jnc.2008.09.006
Rosales, R. M. P. (2018). SEAT: measuring socio-economic benefits of marine protected areas. Mar. Policy 92, 120–130. doi: 10.1016/j.marpol.2018.02.026
Rosenberg, A., Bigford, T., Leathery, S., Hill, R., and Bickers, K. (2000). Ecosystem approaches to fishery management through essential fish habitat. Bull. Mar. Sci. 66, 535–542.
Rubio-Portillo, E., Izquierdo-Muñoz, A., Gago, J. F., Rosselló-Mora, R., Antón, J., and Ramos-Esplá, A. A. (2016). Effects of the 2015 heat wave on benthic invertebrates in the Tabarca marine protected area (southeast Spain). Mar. Environ. Res. 122, 135–142. doi: 10.1016/j.marenvres.2016.10.004
Rubio-Portillo, E., Vázquez-Luis, M., Izquierdo-Muñoz, A., and Ramos-Esplá, A. A. (2014). Distribution patterns of alien coral Oculina patagonica De Angelis D'Ossat, 1908 in western Mediterranean sea. J. Sea Res. 85, 372–378. doi: 10.1016/j.seares.2013.07.007
Russ, G. R., and Alcala, A. C. (1996). Do marine reserves export adult fish biomass? Evidence from Apo island, central Philippines. Mar. Ecol. Prog. Ser. 132, 1–9. doi: 10.3354/meps132001
Russ, G. R., and Alcala, A. C. (2011). Enhanced biodiversity beyond marine reserve boundaries: the cup spillith over. Ecol. Appl. 21, 241–250. doi: 10.1890/09-1197.1
Russi, D., Pantzar, M., Kettunen, M., Gitti, G., Mutafoglu, K., Kotulak, M., et al. (2016). Socio-Economic Benefits of the EU Marine Protected Areas. London; Brussels: Institute for European Environmental Policy (IEEP) & E.C. DG Environment.
Sahyoun, R., Guidetti, P., Di Franco, A., and Planes, S. (2016). Patterns of fish connectivity between a marine protected area and surrounding fished areas. PLoS ONE 11:e0167441. doi: 10.1371/journal.pone.0167441
Sala, E., Ballesteros, E., Dendrinos, P., Di Franco, A., Ferretti, F., Foley, D., et al. (2012). The Structure of mediterranean rocky reef ecosystems across environmental and human gradients, and conservation implications. PLoS ONE 7:e32742. doi: 10.1371/journal.pone.0032742
Sala, E., and Giakoumi, S. (2018). No-take marine reserves are the most effective protected areas in the ocean. ICES J. Mar. Sci. 75, 1166–1168. doi: 10.1093/icesjms/fsx059
Sala, E., Lubchenco, J., Grorud-Colvert, K., Novelli, C., Roberts, C., and Sumailae, U. R. (2018). Assessing real progress towards effective ocean protection. Mar. Policy 91, 11–13. doi: 10.1016/j.marpol.2018.02.004
Samy, M., Sánchez Lizaso, J. L., and Forcada, A. (2011). Status of marine protected areas in Egypt. Anim. Biodiversi. Conserv. 34, 165–177.
Sánchez-Lizaso, J. L., Goñi, R., Reñones, O., García-Charton, J. A., Galzon, R., Bayle, J. T., et al. (2000). Density dependence in marine protected populations: a review. Environ. Conserv. 27, 144–158. doi: 10.1017/S0376892900000187
Sanchirico, J. N., Cochran, K. A., and Emerson, P. M. (2002). Marine Protected Areas: Economic and Social Implications. Resources for the Future, Discussion Paper 02-26. Available online at: www.cbd.int/doc/case-studies/inc/cs-inc-rf-04-en.pdf
Sasal, P., Desdevises, Y., Durieux, E., Lenfant, P., and Romans, P. (2004). Parasites in marine protected areas: success and specificity of monogeneans. J. Fish Biol. 64, 370–379. doi: 10.1111/j.0022-1112.2004.00297.x
Saunders, J., Potts, T., Jackson, E., Burdon, D., Atkins, J. P., Hastings, E., et al. (2015). “Linking ecosystem services of marine protected areas to benefits in human wellbeing?,” in Coastal Zones Ecosystem Services, eds R. K. Turner and M. Schaafsma (Cham: Springer), 167–190. doi: 10.1007/978-3-319-17214-9_9
Schratzberger, M., Neville, S., Painting, S., Weston, K., and Paltriguera, L. (2019). Ecological and socio-economic effects of highly protected marine areas (HPMAs) in temperate waters. Front. Mar. Sci. 6, 749. doi: 10.3389/fmars.2019.00749
Sciberras, M., Jenkins, S. R., Kaiser, M. J., Hawkins, S. J., and Pullin, A. S. (2013). Evaluating the biological effectiveness of fully and partially protected marine areas. Environ. Evid. 2:4. doi: 10.1186/2047-2382-2-4
Shears, N. T., and Babcock, R. C. (2002). Marine reserves demonstrate top-down control of community structure on temperate reefs. Oecologia 132, 131–142. doi: 10.1007/s00442-002-0920-x
Silveira, S., Ortega, I., and Dumont, L. F. C. (2020). Artisanal trawling impact over prey availability and diet of estuarine megabenthic organisms in southern Brazil. Estuar. Coast. Shelf Sci. 237:106682. doi: 10.1016/j.ecss.2020.106682
Simard, F., Laffoley, D., and Baxter, J. M. (Eds.). (2016). Marine Protected Areas and Climate Change: Adaptation and Mitigation Synergies, Opportunities and Challenges. Gland: IUCN. doi: 10.2305/IUCN.CH.2016.14.en
Smith, J., and Metaxas, A. (2018). A decision tree that can address connectivity in the design of marine protected area networks (MPAn). Mar. Policy 88, 269–278. doi: 10.1016/j.marpol.2017.11.034
Soulé, M. E. (2005). “Foreword,” in Marine Conservation Biology: The Science of Maintaining the Sea's Biodiversity, eds E. A. Norse and L. B. Crowde (Washington, DC: Island Press), xi–xvi.
St.Pierre, J. I., and Kovalenko, K. E. (2014). Effect of habitat complexity attributes on species richness. Ecosphere 5:22. doi: 10.1890/ES13-00323.1
Stelzenmüller, V., Maynou, F., Bernard, G., Cadiou, G., Camilleri, M., Crec'hriou, R., et al. (2008). Spatial assessment of fishing effort around European marine reserves: implications for successful fisheries management. Mar. Pollut. Bull. 56, 2018–2026. doi: 10.1016/j.marpolbul.2008.08.006
Stephenson, F., Bulmer, R. H., Meredyth-Young, M., Meysick, L., Hewitt, J. E., and Lundquist, C. J. (2019). Effects of benthic protection extent on recovery dynamics of a conceptual seafloor community. Front. Mar. Sci. 6:607. doi: 10.3389/fmars.2019.00607
Stobart, B., Warwick, R., González, C., Mallol, S., Díaz, D., Reñones, O., et al. (2009). Long term and spillover effects of a marine protected area on an exploited fish community. Mar. Ecol. Prog. Ser. 384, 47–60. doi: 10.3354/meps08007
Taylor, B. M., McIlwain, J. L., and Kerr, A. M. (2012). Marine reserves and reproductive biomass: a case study of a heavily targeted reef fish. PLoS ONE 7:e3959. doi: 10.1371/journal.pone.0039599
Templado, J., Ritcher, A., and Calvo, M. (2016). Reef building Mediterranean vermetid gastropods: disentangling the Dendropoma petraeum species complex. Mediterran. Mar. Sci. 17, 13–31. doi: 10.12681/mms.1333
Terradas-Fernández, M., Zubcoff, J., and Ramos-Esplá, A. A. (2019). Early succession patterns in a Mediterranean vermetid reef. J. Sea Res. 152:101768. doi: 10.1016/j.seares.2019.101768
Tessier, A., Pastor, J., Francour, P., Saragoni, G., Crec'Hriou, R., and Lenfant, P. (2013). Video transects as a complement to underwater visual census to study reserve effect on fish assemblages. Aquat. Biol. 18, 229–241. doi: 10.3354/ab00506
Thiriet, P. D., Di Franco, A., Cheminée, A., Guidetti, P., Bianchimani, O., Basthard-Bogain, S., et al. (2016). Abundance and diversity of crypto- and necto-benthic coastal fish are higher in marine forests than in structurally less complex macroalgal assemblages. PLoS ONE 11:e0164121. doi: 10.1371/journal.pone.0164121
Thrush, S. F., and Dayton, P. K. (2002). Disturbance to marine benthic habitats by trawling and dredging: implications for marine biodiversity. Ann. Rev. Ecol. Systemat. 33, 449–473. doi: 10.1146/annurev.ecolsys.33.010802.150515
Tokeshi, M., and Arakaki, S. (2012). Habitat complexity in aquatic systems: fractals and beyond. Hydrobiologia 685, 27–47. doi: 10.1007/s10750-011-0832-z
Topor, Z. M., Rasher, D. B., Duffy, J. E., and Brandl, S. J. (2018). Marine protected areas enhance coral reef functioning by promoting fish biodiversity. Conserv. Lett. 12:e12638. doi: 10.1111/conl.12638
Turgeon, K., Robillard, A., Grégoire, J., Duclos, V., and Kramer, D. (2010). Functional connectivity from a reef fish perspective: behavioral tactics for moving in a fragmented landscape. Ecology 91, 3332–3342. doi: 10.1890/09-2015.1
USA National Research Council/Committee on the Evaluation Design, and Monitoring of Marine, Reserves, and Protected Areas in the U. S. Ocean Studies Board Commission on Geosciences, Environment and Resources. (2001). Marine protected areas: Tools for Sustaining Ocean Ecosystems. Washington, DC: National Academy Press.
Van den Belt, M., and Cole, A. (2014). Ecosystem Goods and Services in Marine Protected Areas (MPAs). Wellington: New Zealand Department of Conservation.
Vandeperre, F., Higgins, R. M., Sánchez-Meca, J., Maynou, F., Goñi, R., Martín-Sosa, P., et al. (2011). Effects of no-take area size and age of marine protected areas on fisheries yields: a meta-analytical approach. Fish Fish. 12, 412–426. doi: 10.1111/j.1467-2979.2010.00401.x
Velasco, A. M., Pérez-Ruzafa, A., Martínez-Paz, J. M., and Marcos, C. (2018). Ecosystem services and main environmental risks in a coastal lagoon (Mar Menor, Murcia, SE Spain): the perception of stakeholders. J. Nat. Conserv. 43, 180–189. doi: 10.1016/j.jnc.2017.11.002
Viana, D. F., Halpern, B. S., and Gaines, S. D. (2017). Accounting for tourism benefits in marine reserve design. PLoS ONE 12:e0190187. doi: 10.1371/journal.pone.0190187
Villamagna, A. M., Angermeier, P. L., and Bennett, E. M. (2013). Capacity, pressure, demand, and flow: a conceptual framework for analyzing ecosystem service provision and delivery. Ecol. Complex. 15, 114–121. doi: 10.1016/j.ecocom.2013.07.004
Ward, T., and Hegerl, E. (2003). Marine Protected Areas in Ecosystem-based Management of Fisheries. Canberra: Department of the Environment and Heritage, Natural Heritage Trust.
Ward, T. J., Heinemann, D., and Evans, N. (2001). The Role of Marine Reserves as Fisheries Management Tools. A Review of Concepts, Evidence and International Experience. Canberra, ACT: Bureau of Rural Sciences.
Waycott, M., Duarte, C. M., Carruthers, T. J. B., Orth, R. J., Dennison, W. C., Olyarnik, S., et al. (2009). Accelerating loss of seagrasses across the globe threatens coastal ecosystems. Proc. Natl. Acad. Sic. U.S.A. 106, 12377–12381. doi: 10.1073/pnas.0905620106
WCPA/IUCN (2007). Establishing Networks of Marine Protected Areas: A Guide for Developing National and Regional Capacity for Building MPA Networks. Non-technical summary report, WCPA/IUCN, Washington, DC.
Williamson, D. H., Russi, G. R., and Ayling, A. M. (2004). No-take marine reserves increase abundance and biomass of reef fish on inshore fringing reefs of the Great Barrier reef. Environ. Conserv. 31, 149–159. doi: 10.1017/S0376892904001262
Willis, S. C., Winemiller, K. O., and López-Fernández, H. (2005). Habitat structural complexity and morphological diversity of fish assemblages in a neotropical floodplain river. Oecologia 142, 284–295. doi: 10.1007/s00442-004-1723-z
Willis, T. J. (2013). Scientific and Biodiversity Values of Marine Reserves: A Review. DOC Research and Development Series 340. Wellington: Department of Conservation.
Willis, T. J., and Millar, R. B. (2005). Using marine reserves to estimate fishing mortality. Ecol. Lett. 8, 47–52. doi: 10.1111/j.1461-0248.2004.00692.x
Willis, T. J., Millar, R. B., and Babcock, R. C. (2003). Protection of exploited fish in temperate regions: high density and biomass of snapper Pagrus auratus (Sparidae) in northern New Zealand marine reserves. J. Appl. Ecol. 40, 214–227. doi: 10.1046/j.1365-2664.2003.00775.x
Wilson, J. R., Prince, J. D., and Lenihan, H. S.. (2010). A management strategy for sedentary nearshore species that uses marine protected areas as a reference. Mar. Coast. Fish. 2, 14–27. doi: 10.1577/C08-026.1
Wood, C. L., Micheli, F., Fernández, M., Gelcich, S., Castilla, J. C., and Carvajal, J. (2013). Marine protected areas facilitate parasite populations among four fished host species of central Chile. J. Anim. Ecol. 82, 1276–1287. doi: 10.1111/1365-2656.12104
Keywords: marine protected areas, MPA ecosystem services, MPA societal benefits, marine conservation, marine ecosystems management
Citation: Marcos C, Díaz D, Fietz K, Forcada A, Ford A, García-Charton JA, Goñi R, Lenfant P, Mallol S, Mouillot D, Pérez-Marcos M, Puebla O, Manel S and Pérez-Ruzafa A (2021) Reviewing the Ecosystem Services, Societal Goods, and Benefits of Marine Protected Areas. Front. Mar. Sci. 8:613819. doi: 10.3389/fmars.2021.613819
Received: 03 October 2020; Accepted: 22 March 2021;
Published: 01 June 2021.
Edited by:
Trevor Willis, Stazione Zoologica Anton Dohrn, ItalyReviewed by:
Neville Scott Barrett, University of Tasmania, AustraliaBarbara Horta E. Costa, University of Algarve, Portugal
Copyright © 2021 Marcos, Díaz, Fietz, Forcada, Ford, García-Charton, Goñi, Lenfant, Mallol, Mouillot, Pérez-Marcos, Puebla, Manel and Pérez-Ruzafa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Angel Pérez-Ruzafa, YW5nZWxwckB1bS5lcw==