 Macarena Santos-Carvallo1,2,3*
Macarena Santos-Carvallo1,2,3* Fernanda Barilari2,4
Fernanda Barilari2,4 María José Pérez-Alvarez3,5,6
María José Pérez-Alvarez3,5,6 Laura Gutiérrez2,7,8
Laura Gutiérrez2,7,8 Guido Pavez2,3,9
Guido Pavez2,3,9 Héctor Araya10Cristobal Anguita11Claudia Cerda1,12
Héctor Araya10Cristobal Anguita11Claudia Cerda1,12 Maritza Sepúlveda2,3,13
Maritza Sepúlveda2,3,13- 1Programa de Magister en Áreas Silvestres y Conservación de la Naturaleza, Facultad de Ciencias Forestales y de la Conservación de la Naturaleza, Universidad de Chile, Santiago, Chile
- 2Centro de Investigación y Gestión de Recursos Naturales (CIGREN), Instituto de Biología, Facultad de Ciencias, Universidad de Valparaíso, Valparaíso, Chile
- 3Centro de Investigación Eutropia, Santiago, Chile
- 4Programa de Magister en Ciencias mención Biodiversidad y Conservación, Facultad de Ciencias, Universidad de Valparaíso, Valparaíso, Chile
- 5Escuela de Medicina Veterinaria, Facultad de Ciencias, Universidad Mayor, Santiago, Chile
- 6Instituto de Ecología y Biodiversidad, Facultad de Ciencias, Universidad de Chile, Santiago, Chile
- 7Programa de Postgrado en Oceanografía, Departamento de Oceanografía, Facultad de Ciencias Naturales y Oceanográficas, Universidad de Concepción, Concepción, Chile
- 8Centro de Investigación Oceanográfica COPAS Sur-Austral, Universidad de Concepción, Concepción, Chile
- 9Departamento de Ciencias Ecológicas, Facultad de Ciencias, Universidad de Chile, Santiago, Chile
- 10Instituto de Estadística, Facultad de Ciencias, Universidad de Valparaíso, Valparaíso, Chile
- 11Laboratorio de Ecología de Vida Silvestre, Facultad de Ciencias Forestales y de la Conservación de la Naturaleza, Universidad de Chile, Santiago, Chile
- 12Departamento de Gestión Forestal y su Medio Ambiente, Facultad de Ciencias Forestales y de la Conservación de la Naturaleza, Universidad de Chile, Santiago, Chile
- 13Núcleo Milenio de Salmónidos Invasores (INVASAL), Concepción, Chile
Whale-watching (WW) is an activity which has been increasing worldwide due to the great interest of tourists and the economic benefits it provides to local communities. However, it has been reported that this activity affects the behavioral patterns of some cetaceans, although for some species such as the fin whale (Balaenoptera physalus) this has not been extensively studied. To identify the effects of WW on the behavioral patterns of this species, we studied its traveling and resting behaviors in a locality of north-central Chile from 2015 to 2018. Using a theodolite, we calculated the response variables of swim speed, directness index, and reorientation for each behavior. We used the number of WW boats and the WW scenarios of “before”, “during”, and “after” the presence of boats as possible factors to explain the differences in the response variables of the whales, along with the factors of year, month, group size, and distance from the observation point. Reorientation increased significantly and the directness index decreased significantly for both traveling and resting behaviors from “before” to “during” WW scenarios, indicating more erratic and sinuous movements in the presence of boats. These changes in movement patterns are a commonly reported evasion response of cetaceans to the presence of WW boats. For traveling behavior, the swimming speed significantly increased, and trends showed increased reorientation and a decrease in the directness index in the “after” WW scenario, which suggests perturbation of the whales potentially associated with the speed and the direction in which the boats left. During resting behavior, the trajectories of the fin whales became straighter (decrease in reorientation) as the number of boats increased, thus evasion (more erratic and sinuous movements) is a behavior used less by fin whales as the number of boats increases. Notwithstanding the fact that tourism development in the study area is small in scale, we found that WW generates adverse effects that are reflected in changes in the whales’ movement patterns. This kind of information is valuable to the adjustment and/or design of management strategies for the species, which is fundamental for WW to continue to be a sustainable activity.
Introduction
Whale-watching (WW) is one of the fastest-growing tourism industries in a number of countries in recent decades, providing both economic and socioenvironmental benefits. WW has allowed the tourists who take part in it to gain increased knowledge of the biology and diversity of the species of whale seen and the environments in which they live (Filby et al., 2015; Pacheco et al., 2019). Local communities of artisanal fishers who participate in this economic activity benefit by diversifying from their traditional fishing activities, which allows them to increase their sources of income as fishing resources decrease (Parsons et al., 2003; Garrod and Wilson, 2004; Hoyt and Iñíguez, 2008; Guidino et al., 2020). One consequence of this is greater environmental consciousness, which stimulates interest in the conservation and protection of the marine fauna and their habitat (Higginbottom and Tribe, 2004; Zeppel and Muloin, 2008; Schuler and Pearson, 2019), both on the part of consumers (general public) and those who provide the services (e.g., fishers, researchers, businessmen) (Filby et al., 2015; Schuler and Pearson, 2019).
However, there is currently a strong concern over whether WW is an activity that really promotes the conservation of the subject species (Forestell, 2007). It has been widely reported in the literature that inadequate management of WW is an important source of perturbation for the animals, both in the short- and long-term (e.g., Corkeron, 2004; Bejder et al., 2006a; Argüelles et al., 2016; Sprogis et al., 2020b). Important sources of perturbation include a high number of boats in a confined area, very close proximity to the animals, the time and manner of approaching (and leaving) the animals, and lack of regulations or non-compliance with existing norms and regulations (Hoyt and Parsons, 2014). In the short-term, these sources may induce behavioral changes in the cetaceans, some of which may alter biologically important behavior such as feeding (Arcangeli and Crosti, 2009; Christiansen et al., 2013; Lesage et al., 2017) and resting (Avila et al., 2015; Sprogis et al., 2020a, b). Such behavioral variations may produce additional energy costs for the individuals (Williams et al., 2006; Christiansen et al., 2014), affecting their body condition in the medium and long-term, as well as the state of health and the reproductive success of the animals (Lusseau, 2005; Bejder et al., 2006b), which may eventually represent a threat to the conservation of the species exposed to WW (Corkeron, 2004; Lusseau and Bejder, 2007; Parsons, 2012). The other most frequent short-term effect reported is horizontal evasion, indicated by changes of direction in the movement patterns of the animals (Scheidat et al., 2004; Williams and Ashe, 2007; Williams et al., 2009; Schaffar et al., 2013; Schuler et al., 2019). Thus, in the presence of boats, movement patterns become less predictable as linearity decreases (the straight-line trajectory is lost) and reorientation increases (erratic trajectory) in order to evade the boats (Scheidat et al., 2004; Williams and Ashe, 2007; Schaffar et al., 2013). These evasion tactics vary depending on the number of boats and the approach distance (Williams et al., 2009; Schaffar et al., 2013). It has been shown that humpback whales (Megaptera novaeangliae) change their trajectory continuously when boats are close (Schaffar et al., 2013). It has been shown that the displacement trajectory of orcas (Orcinus orca) becomes more linear as the number of boats increases, indicating that the evasion tactic (increase in reorientation) may not be effective in the presence of a larger number of boats (Williams and Ashe, 2007; Williams et al., 2009). If evasive tactics are not effective, cetaceans must use strategies that are more energetically costly (Morete et al., 2007; Christiansen et al., 2014; Sprogis et al., 2020a, b), such as an increase in the velocity of displacement in the presence of boats (Christiansen et al., 2014; Schuler et al., 2019; Sprogis et al., 2020b). Another energetically expensive tactic used by cetaceans is vertical evasion, in which cetaceans increase their diving time (Stamation et al., 2010; Schaffar et al., 2013) and/or increase the rate of respiration (Christiansen et al., 2014; Schuler et al., 2019).
In Chile, tourism activities involving watching marine fauna and specifically WW are still incipient activities, growing by about 20% per year, with the potential for even greater development (Hoyt and Iñíguez, 2008). There are currently five localities in Chile where this activity is developed formally; Bahía de Mejillones in northern Chile, caleta Chañaral de Aceituno and Punta de Choros in north-central Chile, and caleta Puñihuil and the Francisco Coloane Marine Park in southern Chile (Hoyt and Iñíguez, 2008). Given the growth of the WW industry, the Undersecretary of Fisheries and Aquaculture (Subpesca) of the Chilean government published in 2011 the “General regulations for the observation of aquatic mammals, reptiles and birds and the recording of cetacean sighting” (D.S. No.38/2011; Subpesca, 2011). These regulations established the procedures and general requisites for the observation of these aquatic species, regulating among other aspects, observation distance, the method of approach of boats to the cetaceans and the behavior of tourists during the sightings (D.S. No.38/2011; Subpesca, 2011).
Caleta Chañaral de Aceituno is one of the favorite WW locations of tourists due to the high probability of sighting large cetaceans. This location is visited by a number of large cetacean species in the summer (e.g., blue, fin, humpback, and minke whales; Capella et al., 1999; Pérez et al., 2006; Toro et al., 2016; Sepúlveda et al., 2018). Fin whales are the most commonly observed species in the area (Pérez et al., 2006; Toro et al., 2016; Sepúlveda et al., 2018). The number of tourists visiting this location to participate in WW activities has increased exponentially in the last decade, from approximately 1,200 in the summer of 2010 to approximately 8,000 tourists in the summer of 2020 (Corporación Nacional Forestal, unpublished data). This location contains the Marine Protected Area “Isla Chañaral Marine Reserve” (D.S. No. 150-05; Subpesca, 2005), which has its own regulations for watching marine fauna.
Whale-watching is conducted by artisanal fishers of caleta Chañaral de Aceituno, which has provided an opportunity to expand and diversify their traditional activities (Sepúlveda et al., 2018). Due to the increase in the number of tourists and the importance of the species that visit the area, local authorities have introduced additional regulations in the marine reserve to those of the 2011 regulations, limiting the number of boats and the time they can stay with the animals (Res. Ex. No.655/2020; Sernapesca, 2020). However, in spite of these regulations, and considering the rapid increase of WW activity, to our knowledge, no study has addressed whether the cetaceans at this location are affected by WW and if so, to what degree. This study analyzed the behavioral responses of the fin whale to WW in this locality. The behavioral responses were evaluated using scenarios of WW “before”, “during”, and “after” the presence of WW boats. The number of boats and other factors such as year, month, group size, and distance from the observation point were also recorded. This study will provide an initial insight into the effects of WW on the fin whale in Chile, a species with known conservation issues.
Materials and Methods
Study Area
The study area is located within the Humboldt Current System, in north-central Chile. This is a dynamic and productive coastal environment due to the presence of an important wind-driven coastal upwelling center (Montecino et al., 2006; Thiel et al., 2007). A high diversity of marine fauna has been reported in the area, including birds and marine mammals (Capella et al., 1999; Luna-Jorquera et al., 2003; Pérez et al., 2006; Sepúlveda et al., 2009, 2016), and it has been described as an important feeding area for the fin whale (Balaenoptera physalus) during the austral spring and summer (Pérez et al., 2006; Toro et al., 2016; Sepúlveda et al., 2018). These characteristics are what makes caleta Chañaral de Aceituno one of the favorite tourist places in Chile.
The land-based observation station was located on Chañaral Island (29°01′S, 71°36′W), at 52 m above sea level on the eastern edge of the island (Figure 1). The observation area includes a zone called “the channel,” which is between the island and the continent (Figure 1). The largest concentration of tourist boats occurs in this zone as they see the cetaceans on their way to visit the fauna of the island. A portion of the Isla Chañaral Marine Reserve is also within the observation area (Figure 1); it has its own rules for WW (Res. Ex. No.655/2020; Sernapesca, 2020) in addition to those rules which apply to the whole country (D.S. No.38/2011; Subpesca, 2011).
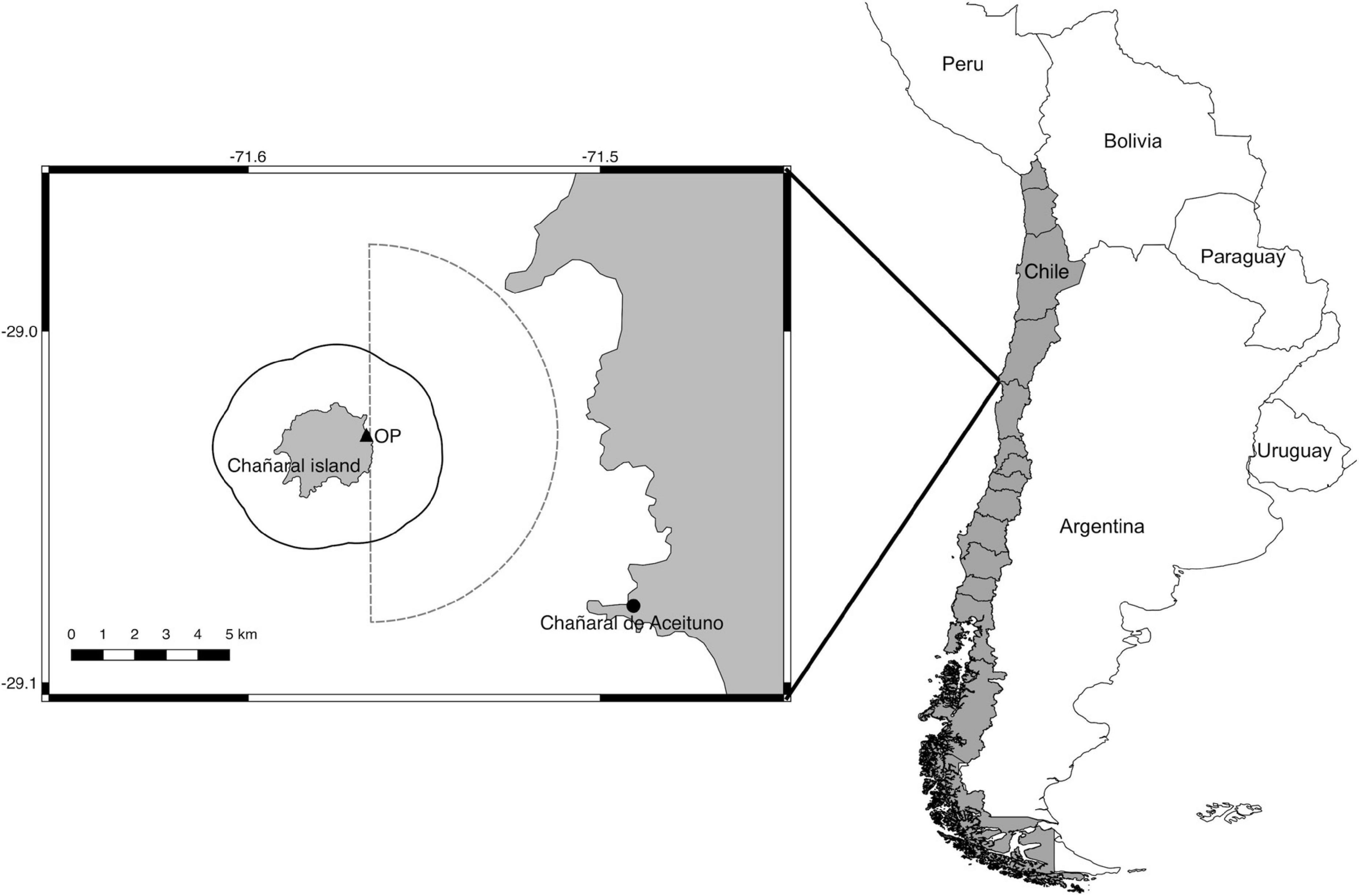
Figure 1. Location of the study area. The black triangle indicates the land-based observation point (OP) and the black circle is the location of the caleta Chañaral de Aceituno. The black line indicates the limit of the Isla Chañaral Marine Reserve and the dashed line indicates the area of observation.
Whale-watching is performed by artisanal fishers of caleta Chañaral de Aceituno, a small locality with about 100 inhabitants, located 9 km from the Chañaral island (Figure 1). Thirty-nine boats have formal permission to perform WW in the Marine Reserve, but in practice less than 20 boats perform these activities. The boats used are up to 10 m in length, with motors of up to 150 hp.
Data Collection
The study was carried out during the months of January and February annually, from 2015 to 2018. Observations were performed from 09:00 to 18:00 when weather conditions provided good visibility to ensure reliable data collection (Beaufort Sea state of 3 or less, with no coastal fog or rain). The viewing area covered approximately 180° and was scanned with either the naked eye or binoculars (10 × 42) (Figure 1).
Spatial movement patterns of fin whales were monitored using a Spectra Precision Model DET-2 digital theodolite with 30-power monocular magnification and 2 s precision. This method has been widely used to follow whales from land stations and has proven to be successful to estimate the position of a whale at a given time (e.g., Würsig et al., 1991; Scheidat et al., 2004; Schaffar et al., 2013; Pirotta et al., 2016). This also allowed the recording of changes in behavior of the animals in the presence and absence of tourist boats without the researchers interfering in the natural behavior of the whales (Würsig et al., 1991; Morete et al., 2018).
When an individual or a group of fin whales was sighted, scanning was suspended and a theodolite tracking session and a focal follow was initiated (i.e., tracking a single individual or group of fin whales at a time) (Altmann, 1974; Mann, 1999), both in the presence or absence of WW boats. A group was defined as two or more individuals that surfaced synchronously within 100 m of one another (Whitehead, 1983; Corkeron, 1995). Individuals or groups of whales were followed and sampled continuously using the description of the focal protocol given here. Focal follows were carried out by a team of three experienced observers: a theodolite operator, a spotter, and a data scorer. During the focal follow activities, the spotter announced the surfacing event of the focal group, the theodolite operator located its position, and the scoring observer recorded the time, the surface event, the behavior (traveling or resting, see below), the vertical and horizontal angles provided by the theodolite, the group size, the presence or absence of WW boats, and the number of WW boats. The whale’s position (vertical and horizontal angles) was recorded every minute (or after the whale came to the surface) (Schaffar et al., 2010). To follow groups, the position of the first whale that came to the surface was recorded. Each whale or group of whales was tracked continuously until the animal was no longer visible, or environmental conditions prevented further tracking. WW boats were considered present and included in the analysis when they approached a whale in a straight line or parallel, according to the direction of the whale, at an estimated distance of 500 m from the animal. These methods are appropriate for fin whales since this species dives for short periods of time (<10 min) (Croll et al., 2001) being easy to follow. Also, in the study area fin whales occurred frequently, both singularly or in small groups (approximately three or four whales) (Toro et al., 2016), which reduce the risk of confusing individuals or groups.
The behavior of fin whales was classified into the categories of traveling or resting, based on specific behavioral events observed (Brown et al., 1994). The category of traveling was considered when an individual or group of whales were moving and oriented in the same direction, displaying a quasi-linear trajectory with sub-surface constant swimming without stopping (Brown et al., 1994), while resting was considered when an individual or group of whales was stationary and all members were oriented in the same direction (Brown et al., 1994). Although feeding behavior does occur in the study area (Pérez et al., 2006; Toro et al., 2016; Sepúlveda et al., 2018), this behavior was rarely recorded in the observation area, and therefore was not analyzed.
The information was digitized and entered into the VADAR software (Visual and Acoustic Detection and Ranging, developed by Eric Kniest). This software uses angles from the theodolite, together with the height of the station and the equipment to calculate the position of the focal individual or group. All observational information, including the exact time of each surfacing event and behavior, was imported to VADAR. From this process we obtained the behavioral response variables of: (1) swim speed, (2) reorientation, and (3) directness index (Harcourt et al., 2014). The swim speed (km h–1) of a whale or group of whales was measured as the time (in hours) taken to cross the distance (in kilometers) between two consecutive sightings (Pirotta et al., 2016). Reorientation measures path predictability from one surfacing to the next, and it is defined as the change in the direction of movement of the individual or group (Williams et al., 2002). This measure is the angle between the path taken during a dive and the predicted straight-line path as indicated by the direction of the dive before it, ranging potentially from 0° to 180° (Williams et al., 2009). Low reorientation values indicate a smooth path, while high values indicate an erratic path (Williams et al., 2002). Finally, the directness index is defined as the path predictability of the whales over the length of the tracking session. This index is measured as the straight-line distance between the first and the last fix of a tracking session divided by the cumulative surface distance covered by the group (Williams et al., 2002), and it ranges from 0 (circular path) to 1 (straight line). The three behavioral response variables were given by VADAR for each whale position fixed with the theodolite throughout a tracking session.
For statistical analyses, only theodolite tracking ≤6 km from the station was considered to ensure reliable data for analysis (see Würsig et al., 1991 for height-related errors). A follow event was only considered viable when an individual or group of whales was tracked for at least 15 min and/or at least five surfacing bouts (modified by Scheidat et al., 2004; Schaffar et al., 2010). These tracks allowed a representative sample of the whales’ behavior (Schaffar et al., 2013).
Statistical Analyses
For the behaviors of traveling and resting, we modeled swim speed, reorientation, and directness index in response to the additive effects (i.e., no interactions) of: year (factor; 2015–2018), month (factor; January and February), distance from observation point (km), group size, and number of WW boats. In addition we included three levels of WW scenarios as a predictive variable; (1) “before” the arrival of the boats; (2) “during”, when one or more boats were present with the whale(s); and (3) “after” the boats had left the area in which the whale(s) were located (Scheidat et al., 2004; Avila et al., 2015). For the response variable swim speed we used a normal distribution and for the variables of reorientation (i.e., proportion of 90° of reorientation) and directness index we used a beta distribution with a logit link. For all response variables we modeled only the location parameter of the chosen distribution (mu, i.e., mean). For model selection we used information-theoretic model comparison. Specifically, we carried out the selection of the best model in the set using stepwise model selection based on the generalized Akaike’s information criterion (GAIC) with a penalty of k = 3 (>AIC and <BIC). The GAIC is a generalization of AIC to evaluate parsimony, which penalizes the deviance by a factor k (positive real number) the number of degrees of freedom in the model (i.e., GAIC = k × Df + D), unlike AIC where k is fixed and equals two (Stasinopoulos et al., 2017). In addition, for all best models a Generalized (Pseudo) R-squared test was calculated (Nagelkerke, 1991). Models fitting, selection and diagnoses (based on residuals plots; Dunn and Smyth, 1996) were done using the gamlss package (Rigby and Stasinopoulos, 2005) of R (R Core Team, 2018). For multiple comparisons across factor levels, we used Tukey’s a posteriori HSD test available in the emmeans package of R (Lenth et al., 2018). To avoid co-linearity problems during the modeling process we dropped variables with a variance inflation factor >2 (Zuur et al., 2010). Specifically, for the models of reorientation during traveling behavior and directness index during resting behavior we excluded the variable number of WW boats and for the model of reorientation during resting behavior we excluded the variable group size, due to the high level of collinearity that this variable presented with the variable of greatest interest, i.e., WW scenarios. Marginal effects from final models (i.e., predicted values for certain model terms by holding the non-focal variables constant) were also estimated using the emmeans package. We produced figures using the R package ggplot2 (Wickham, 2016).
Results
Observation Effort
We spent a total of 94 days in the field, with a total of 684 h of observations. During that time, 34 effective focal follow events were usable, 24 for traveling and 10 for resting behaviors. We recorded 12 focal follows for traveling behavior that included the “before” WW scenario, 16 that included “during” WW scenario and eight that included the “after” WW scenario. For resting behavior, we recorded four focal follows “before” WW scenario and seven “during” WW scenario. Since the observed whales were no longer resting after WW, it was not possible to record data for this behavior in this instance.
More than half of the records for traveling and resting (57 and 55%, respectively) had boats present. Most of the observations were with the presence of a single boat (48% traveling, 68% resting), with smaller proportions in the presence of two (42% traveling, 17% resting) or three (10% traveling, 15% resting) boats.
Traveling Behavior
For swim speed during traveling behavior, the most parsimonious model contained the predictive variable of WW scenarios and accounted for 3% of the total variation (Table 1). There was a significant increase in swim speed in the “after” WW scenario (Tukey post hoc; P < 0.05, Figure 2A). The best model for reorientation included WW scenarios, year, and distance from observation point, and accounted for 32% of the total variation (Table 1). There was an increase in reorientation “during” and “after” WW scenarios compared to “before” (Tukey post hoc; P < 0.001, Figure 2B). The greatest values of reorientation were from 2016 (Tukey post hoc; P < 0.05, Figure 2C). Reorientation decreased as the distance from the observation point increased (slope and 95% CI in logit scale = -0.08 [–0.14, –0.01], P < 0.05, Figure 2D). The best model for the directness index included the predictive variable WW scenarios and accounted for 14% of the total variation (Table 1). The directness index decreased “during” and “after” WW compared to “before” WW (Tukey post hoc; P < 0.001, Figure 2E).
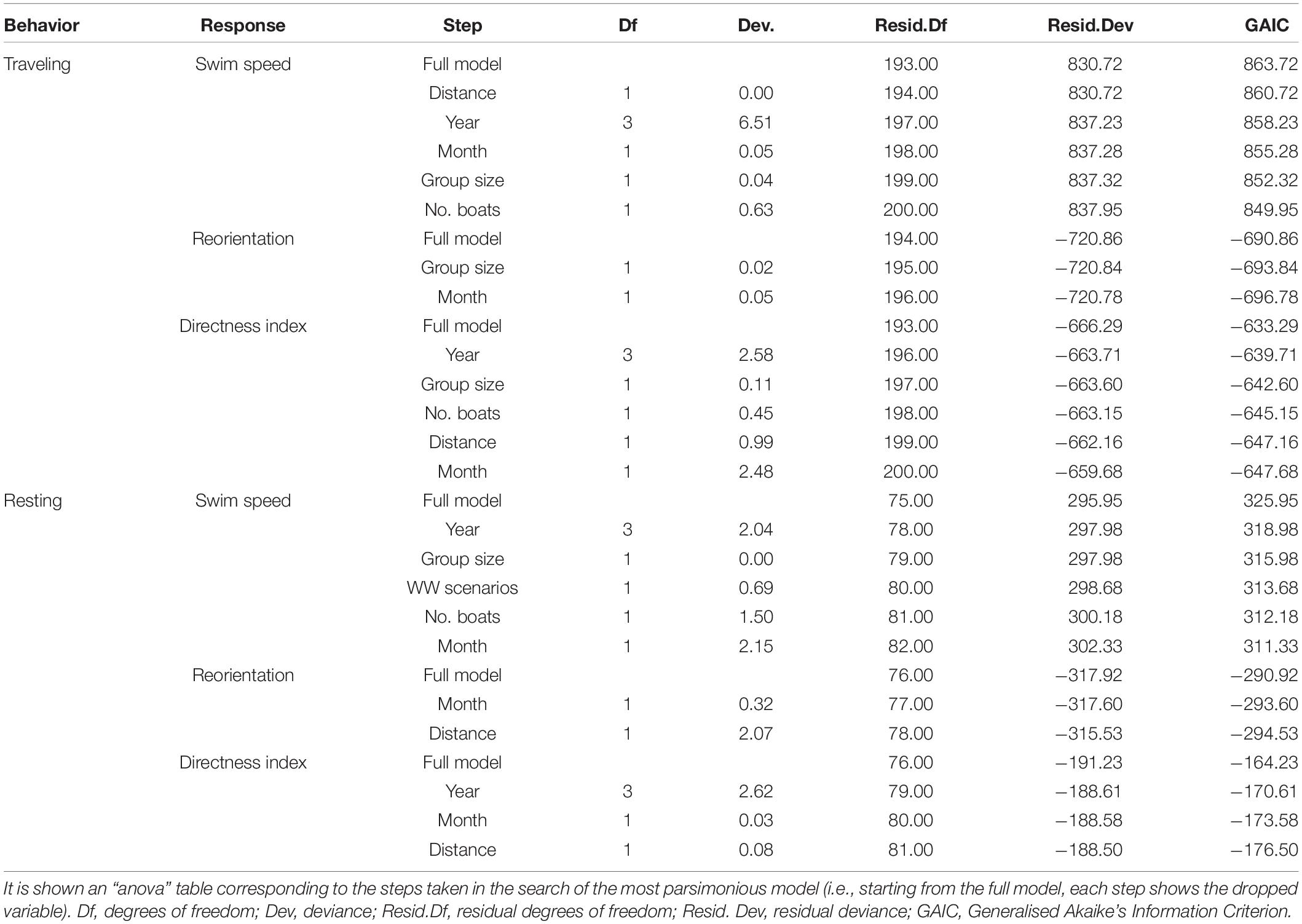
Table 1. Results from the backward stepwise model selection of swim speed, reorientation, and directness index for traveling and resting behaviors of fin whales.
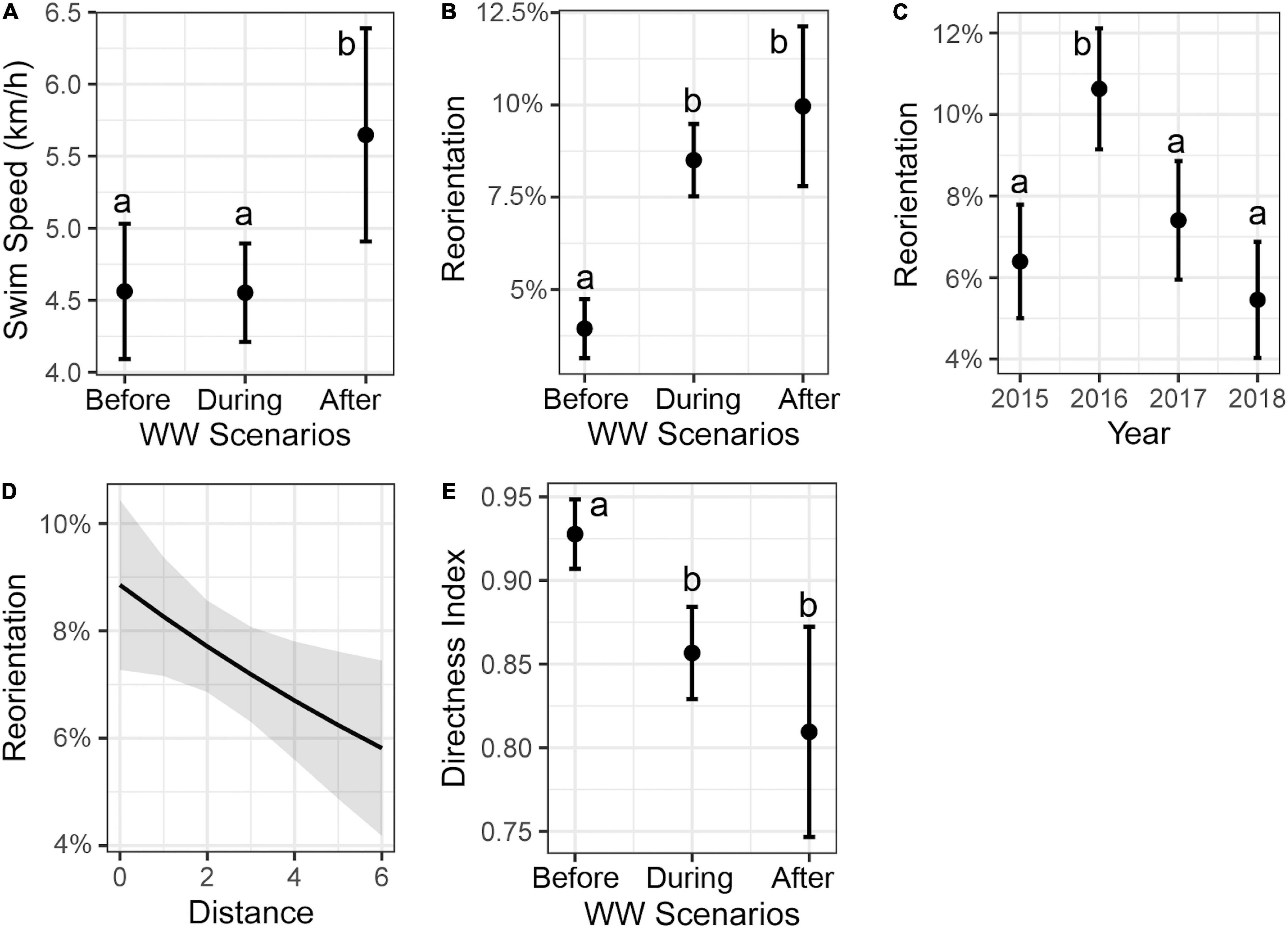
Figure 2. Marginal effects (mean ± 95% confidence interval) from predictors of the best model for (A) swim speed, (B–D) reorientation, and (E) directness index during traveling behavior of fin whales. Different letters indicate significant differences between groups (Tukey post hoc; P < 0.05).
Resting Behavior
For swim speed during resting behavior, the most parsimonious model included distance from observation point and accounted for 8% of the total variance (Table 1). Swim speed increased together with the distance from the observation point (slope and 95% CI = 0.42 [0.12, 0.73], P < 0.05, Figure 3A). The best model for reorientation contained the predictive variables year, WW scenarios and number of WW boats, and accounted for 47% of the total variance (Table 1). The greatest values for reorientation occurred in 2017 (Tukey post hoc; P < 0.05, Figure 3B). There was significant higher reorientation “during” WW scenario than “before” (P < 0.001, Figure 3C). Also, reorientation decreased as the number of WW boats increased (slope and 95% CI in logit scale = –0.32 [–0.49, –0.16], P < 0.001, Figure 3D). The best model for the directness index included WW scenarios and group size, and accounted for 8% of the total variation (Table 1). There was a significant decrease in the directness index “during” WW compared to “before” (P < 0.001, Figure 3E). Directness index increased as the group size increased (slope and 95% CI in logit scale = 0.21 [0.01, 0.42], P < 0.05, Figure 3F).
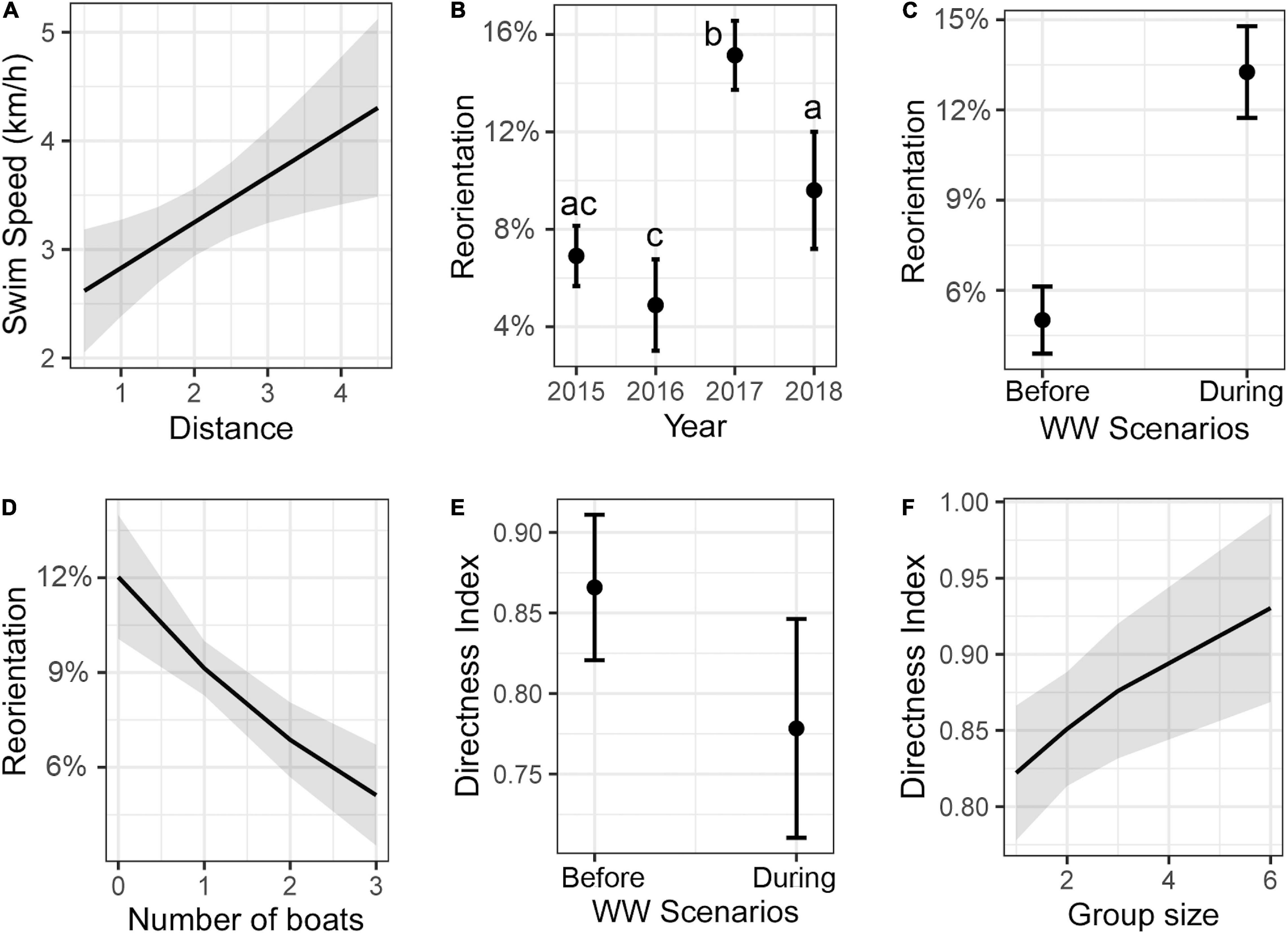
Figure 3. Marginal effects (mean ± 95% confidence interval) from predictors of the best model for (A) swim speed, (B–D) reorientation, and (E,F) directness index during resting behavior of fin whales. Different letters indicate significant differences between groups (Tukey post hoc; P < 0.05).
Discussion
The development and increase in WW has generated extensive discussion on the benefits and disturbance it produces. The economic and socio-environmental benefits have been widely recognized (e.g., Filby et al., 2015; Schuler and Pearson, 2019). However, there is growing evidence of the negative effects of tourism on the conservation of the species (Parsons, 2012; Higham et al., 2016). Considering the sustained increase in WW in the past few decades in a number of countries (Hoyt and Parsons, 2014; Schuler et al., 2019), monitoring of the effects of WW activity on subject species is fundamental to identify potential short-term perturbations with potential medium-and long-term consequences. Our study provides relevant information about the impact of WW on the behavior of the fin whale, a species poorly known in this topic, using a technique that does not interfere in the dynamics of tourism or in the behavioral response of the animals (Würsig et al., 1991; Morete et al., 2018; Piwetz et al., 2018).
Model results indicated that reorientation increased for both traveling and resting behaviors, while the directness index increased between the “before” and “during” WW scenarios. This means that in the presence of boats whales were making constant changes of direction and erratic movements, losing the movement linearity that they displayed before the boats arrived. These changes in the reorientation and directness index have been reported previously as responses of cetaceans to the presence of boats (e.g., Scheidat et al., 2004; Schaffar et al., 2013; Avila et al., 2015; Senigaglia et al., 2016; Sprogis et al., 2020a, b). Frid and Dill (2002) suggested that the alteration in the natural behavior of the animals in the presence of anthropogenic perturbation is produced because the animals perceive these perturbations similarly to a predation risk. For example, the minke whale (Balaenoptera acutorostrata) uses similar evasion tactics in response to the presence of tourist boats as in the presence of its natural predators orcas (Christiansen et al., 2013). This suggests that cetaceans could identify the presence of boats as a threat (Christiansen and Lusseau, 2014), resulting in the onset of avoidance behaviors.
Although the model with the most support for swimming speed had low explicative power, it identified a significant increase in the “after” WW scenario during traveling behavior; i.e., the swim speed of the whales increased after the boats had left the area. An increase in the reorientation and a decrease in the directness index in the “after” WW scenario were also observed. In contrast to these results, different studies have indicated that once the boat visits had finished, the whales returned to their initial behavior after a short time (e.g., Scheidat et al., 2004; Avila et al., 2015). The persistence, and even accentuation of the perturbation of behavior when the boats had left may be related to the behavior of the boat operators after the sighting had finished. Our field observations indicate that the boats would leave the area at high speeds, and sometimes passing in front of the animal. These two factors may be affecting the whales even more than the presence of the boats, since the animals accentuate the evasion strategy, increasing their speed and following less predictable trajectories, after the boats leave. The negative effect of high-speed boats has previously been described, showing that this factor limits the ability of the whales to avoid them (Parsons, 2012). Additionally, considering that the noise of the boats generated by the propeller cavitation produces adverse reactions in whale behavior (Erbe et al., 2019), and that more noise is produced at higher speed (Walker et al., 2019), the behavioral change of the animals may be due to the increase in the boats speed. Although we did not use the speed and path direction of the boats moving away as predictive variables, there is evidence that these variables negatively impact the behavior of large cetaceans (Heckel et al., 2001; Argüelles et al., 2016; Fiori et al., 2019), since this avoidance strategy increases the energetic cost for the animals (Williams et al., 2009; Christiansen et al., 2014; Sprogis et al., 2020b). We recommend that future studies incorporate other variables to describe the boat behavior, such as speed and direction of arriving and departing boats, to identify other potentially relevant factors in the responses of fin whales to WW.
The only model that included the number of boats as a significant variable was reorientation during resting behavior, indicating that the trajectories of the fin whales in this behavior became more direct (decreasing reorientation) as the number of boats increased. The behavior of maximizing linear movement as the number of boats increased was described previously in orcas, indicating that evasion responses (erratic and more sinuous movements) are employed when there are few boats, and by contrast, when the number of boats increases the animals choose a more linear trajectory to move away (Williams et al., 2002, 2009). Although the maximum simultaneous number of boats near a whale or group of whales was considerably smaller in this study compared to the report of Williams et al. (2009) (3 versus 14 boats), our results suggest the effect was similar to that reported for a high number of boats. This suggests that maintaining a low number of boats near an individual or group of whales is crucial to avoid drastic changes in the behavioral responses of the fin whale. This is especially relevant and reinforces the current measures in the Isla Chañaral Marine Reserve, which permit a maximum of two boats per whale or group of whales.
Our study shows that the behavioral responses of the fin whales are directly influenced by the WW activities. However, it must be noted that environmental and/or social factors could also contribute to these responses (Yazvenko et al., 2007; Gailey et al., 2016; Kavanagh et al., 2017), and may be related to the low explicative power (less than 10%) of some of the models. These include environmental factors such as wind speed, depth, time of day, distance to the coast (Yazvenko et al., 2007; Williams et al., 2009; Kavanagh et al., 2017); intraspecific factors such as age, sex and group size (Williams et al., 2009; Kavanagh et al., 2017); and others associated with tourist activity, such as distance and speed of approach and departure and path of approaching (Williams et al., 2009; Schaffar et al., 2013), among others. For instance, our study found that reorientation during traveling behavior decreased as the distance between the observation point and the animals observed increases. This result may not be related to WW, but rather explained by the increase in depth in the study area farther from the coast (Gaymer et al., 2008). More direct movement by the whales may facilitate less energetically costly travel in deeper areas (Gailey et al., 2016). This demonstrates that the behavioral responses of cetaceans are difficult to analyze, since they are influenced by many variables, which are often not linear (Williams et al., 2009). Due to this complexity, it is important to evaluate how environmental and/or social factors affect the behavioral responses of the whales (without anthropogenic factors), in order to identify if these responses could be attributed to natural factors or to anthropogenic disturbances (Kavanagh et al., 2017). Some of the environmental parameters for this specific study would be the depth, swell height, and wind speed that have been reported as relevant variables in other studies of whales (Gailey et al., 2016; Kavanagh et al., 2017).
Tourist development in caleta Chañaral de Aceituno is still at a low scale compared to other WW areas, both national and international, due to the small number of visitors (although this is increasing), and the use of small boats (Sepúlveda et al., 2016). In spite of this, we showed that even in this stage of development WW generates adverse effects on fin whales. Although this study only considered short-term behavioral changes, it is important to consider that behavioral changes in the whales may also result in long-term negative effects (Parsons, 2012; Schuler et al., 2019). Alterations in essential behaviors such as resting, feeding, continuous changes in direction and increased swimming speed to avoid boats produce an increase in energetic costs (Bain et al., 2014; Christiansen et al., 2014), which may cause deterioration in the physical condition of the animals if prolonged over time (Beale, 2007). This study area is important for the fin whale, since it is a recognized feeding zone on the Chilean coast (Pérez et al., 2006; Toro et al., 2016; Sepúlveda et al., 2018). Photo-identification has shown that some individuals stay in the area for weeks and even months, and some individuals return in different years (Toro et al., 2016), thus the perturbation by WW may not only be momentary, but rather some animals may be exposed to these perturbations frequently and over an extended period of time. Although we do not know the extent of the exposure of individuals to the WW activities (e.g., maximal daily exposure of the whales to the boats, proportion of the fin whale population that is being affected by WW), and the potential long-term effect of the observed changes in the behavior of fin whales, we do encourage the use of precautionary principle for minimizing impacts by adopting the codes of best practices from the beginning. Future studies should evaluate the level of exposure by relating information regarding permanence pattern and habitat use by fin whales, together with data from tourism activity (e.g., area covered by a boat during a tourist trip, number of boats).
The study area of caleta Chañaral de Aceituno has unique characteristics that make it an optimal location for WW. It is a recognized feeding area for the fin whale and other small and large species of cetaceans, several of which face conservation issues (Capella et al., 1999; Pérez et al., 2006; Toro et al., 2016; Sepúlveda et al., 2018). It is a marine protected area which can implement its own regulations, and tourism is at a small scale performed by artisanal fishers who have important knowledge of the local fishing resources and tourism, as well as identity and sense of belonging to the area (Sepúlveda et al., 2016). The results obtained in this study should be considered in the adjustment of the existing management tools and in the design of new complementary conservation strategies, to increase the balance toward the positive aspects of WW, as has been demonstrated in other places (e.g., Península Valdez; Chalcobsky et al., 2017). Artisan fishers are key actors to include in the design of cetacean’s conservation strategies. Given the importance of the conservation of the fin whale for their well-being, increasing the level of understanding is relevant to offer sustainable tourism services in the long term (Mace, 2014). Knowledge of whale behavior and responses to WW are critical to ensure the sustainability of tourism activity in this locality.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
Ethical review and approval was not required for the animal study because the study was carried out through land-based observations, so there was no manipulation or any intervention in natural behavior of whales.
Author Contributions
MS-C and MS wrote the manuscript text, while all authors wrote, reviewed and edited the manuscript text. MS-C, GP, and MS contributed to the project design and funding acquisition. MS-C, FB, and LG contributed to the investigation methodology, field work, and data processing. MP-A contributed to the investigation methodology and ethology. CC contributed to the conservation approach. HA and CA contributed to the formal analysis. All authors listed have made a substantial, direct and intellectual contribution to the work.
Funding
This study was funded by the Fondo de Protección Ambiental (FPA) of Ministerio del Medio Ambiente entitled “Whale-watching en la Reserva Marina Isla Chanþaral: manejo y planificación para una actividad sustentable” and by the Bienes Públicos de Innova-Corfo project “Plan estratégico de desarrollo sustentable para posicionar a la Región de Atacama como un destino turístico de alta calidad para el avistamiento de cetáceos (TAC-AC), otros mamíferos y aves marinas”. MP-A was supported by the Agencia Nacional de Investigación y Desarrollo (ANID) under Grant Program FONDECYT Iniciación 11170182.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank E. Kniest who provided VADAR development and support. We especially thank all the volunteers who helped with data acquisition in the field (A. Colilef, C. Barrios, G. Alosilla, N. Balcazar, C. Calderón, D. Cárcamo, A. Contreras, A. Farfan, E. Gutiérrez, S. Maldonado, R. Manzul, D. Meneses, V. Morales, E. Oda, U. Otxandorena, G. Peña, D. Pillado, M. Rivera, and H. Werner). We thank Turismos Orca (L. González y J. González), the Agrupación de Turismo Caleta Chañaral de Aceituno and Agrupación de Turismo Delfines for assistance with the logistics of the field work. We thank the Corporación Nacional Forestal (CONAF-Atacama) for authorizing the work in Isla Chañaral, and especially M. Maldonado and C. Rivera for their help in the field. We gratefully acknowledge Lafayette Eaton and Kobe Martin for language translation, and the reviewers for their comments that greatly improved the manuscript.
References
Altmann, J. (1974). Observational study of behavior: sampling methods. Behaviour 49, 227–267. doi: 10.1163/156853974X00534
Arcangeli, A., and Crosti, R. (2009). The short-term impact of dolphin-watching on the behaviour of bottlenose dolphins (Tursiops truncatus) in western Australia. J. Mar. Anim. Ecol. 2, 3–9.
Argüelles, M. B., Coscarella, M., Fazio, A., and Bertellotti, M. (2016). Impact of whale-watching on the short-term behavior of Southern right whales (Eubalaena australis) in Patagonia. Argentina. Tour. Manag. Perspect. 18, 118–124. doi: 10.1016/j.tmp.2016.02.002
Avila, I. C., Correa, L. M., and Parsons, E. C. M. (2015). Whale-Watching activity in Bahía Málaga, on the Pacific coast of Colombia, and its effect on Humpback Whale (Megaptera novaeangliae) behavior. Tourism Mar. Environ. 11, 19–32. doi: 10.3727/154427315X14398263718394
Bain, D. E., Williams, R., and Trites, A. W. (2014). “Energetic linkages between short-term and long-term effects of whale-watching disturbance on cetaceans,” in Whale-watching: sustainable tourism and ecological management, eds J. Higham, L. Bejder, and R. Williams (Cambridge: Cambridge University Press), 206–228. doi: 10.1017/CBO9781139018166.018
Beale, C. M. (2007). The behavioral ecology of disturbance responses. Int. J. Comp. Psychol. 20, 111–120.
Bejder, L., Samuels, A., Whitehead, H., and Gales, N. (2006a). Interpreting short-term behavioural responses to disturbance within a longitudinal perspective. Anim. Behav. 72, 1149–1158. doi: 10.1016/j.anbehav.2006.04.003
Bejder, L., Samuels, A., Whitehead, H., Gales, N., Mann, J., Connor, R., et al. (2006b). Decline in relative abundance of bottlenose dolphins exposed to long-term disturbance. Conserv. Biol. 20, 1791–1798. doi: 10.1111/j.1523-1739.2006.00540.x
Brown, M. R., Corkeron, P. J., Hale, P. T., Schultz, K. W., and Bryden, M. M. (1994). Behavioral responses of east Australian humpback whales Megaptera novaeangliae to biopsy sampling. Mar. Mamm. Sci. 10, 391–400. doi: 10.1111/j.1748-7692.1994.tb00496.x
Capella, J., Vilina, Y., and Gibbons, J. (1999). Observación de cetáceos en isla Chañaral y nuevos registros para el área de la Reserva Nacional Pingüino de Humboldt, norte de Chile. Estud. Oceanol. 18, 57–64.
Chalcobsky, B. A., Crespo, E. A., and Coscarella, M. A. (2017). Whale-watching in Patagonia: What regulation scheme should be implemented when the socio-ecological system is changing? Mar. Policy 75, 165–173. doi: 10.1016/j.marpol.2016.11.010
Christiansen, F., and Lusseau, D. (2014). “Understanding the ecological effects of whale-watching on cetaceans,” in Whale-watching: Sustainable tourism and ecological management, eds J. Higham, L. Bejder, and R. Williams (Cambridge: Cambridge University Press), 177–192. doi: 10.1017/CBO9781139018166.016
Christiansen, F., Rasmussen, M., and Lusseau, D. (2013). Whale watching disrupts feeding activities of minke whales on a feeding ground. Mar. Ecol. Prog. Ser. 478, 239–251. doi: 10.3354/meps10163
Christiansen, F., Rasmussen, M., and Lusseau, D. (2014). Inferring energy expenditure from respiration rates in minke whales to measure the effects of whale watching boat interactions. J. Exp. Mar. Biol. Ecol. 459, 96–104. doi: 10.1016/j.jembe.2014.05.014
Corkeron, P. J. (1995). Humpback whales (Megaptera novaeangliae) in Hervey Bay, Queensland: Behaviour and responses to whale-watching vessels. Can. J. Zool. 73, 1290–1299. doi: 10.1139/z95-153
Corkeron, P. J. (2004). Whale watching, iconography, and marine conservation. Conserv. Biol. 18, 847–849. doi: 10.1111/j.1523-1739.2004.00255.x
Croll, D. A., Acevedo-Gutiérrez, A., Tershy, B. R., and Urbán-Ramírez, J. (2001). The diving behavior of blue and fin whales: is dive duration shorter than expected based on oxygen stores? Comp. Biochem. Physiol. A 129, 797–809. doi: 10.1016/S1095-6433(01)00348-8
Dunn, P. K., and Smyth, G. K. (1996). Randomized quantile residuals. J. Computat. Graphical Statist. 5, 236–244. doi: 10.1080/10618600.1996.10474708
Erbe, C., Marley, S. A., Schoeman, R. P., Smith, J. N., Trigg, L. E., and Embling, C. B. (2019). The effects of ship noise on marine mammals—A review. Front. Mar. Sci. 6:606. doi: 10.3389/fmars.2019.00606
Filby, N. E., Stockin, K. A., and Scarpaci, C. (2015). Social science as a vehicle to improve dolphin-swim tour operation compliance? Mar. Policy. 51, 40–47. doi: 10.1016/j.marpol.2014.07.010
Fiori, L., Martinez, E., Orams, M. B., and Bollard, B. (2019). Effects of whale-based tourism in Vava’u, Kingdom of Tonga: Behavioural responses of humpback whales to vessel and swimming tourism activities. PLoS One 14:e0219364. doi: 10.1371/journal.pone.0219364
Forestell, P. H. (2007). “Protecting the ocean by regulating whalewatching: the sound of one hand clapping,” in Marine Wildlife and Tourism Management: Insights from the Natural and Social Sciences, eds J. Higham and M. Lück (Oxfordshire: CABI Publishing), 272–293. doi: 10.1079/9781845933456.0272
Frid, A., and Dill, L. (2002). Human-caused disturbance stimuli as a form of predation risk. Conserv. Ecol. 6:11. doi: 10.5751/ES-00404-060111
Gailey, G., Sychenko, O., Mcdonald, T., Racca, R., Rutenko, A., and Bröker, K. (2016). Behavioural responses of western gray whales to a 4-D seismic survey off northeastern Sakhalin Island, Russia. Endanger. Species Res. 30, 53–71. doi: 10.3354/esr00713
Garrod, B., and Wilson, J. C. (2004). Nature on the edge? Marine ecotourism in peripheral coastal areas. J. Sustain. Tour. 12, 95–120. doi: 10.1080/09669580408667227
Gaymer, C., Bórquez, R., Dumont, C., Garay, R., Sfeir, R., Stotz, W., et al. (2008). Evaluación de línea base de las reservas marinas “Isla Chañaral” e “Isla Choros-Damas. Informe final proyecto FIP 2006-56. Coquimbo: Universidad Católica del Norte.
Guidino, C., Campbell, E., Alcorta, B., Gonzalez, V., Mangel, J. C., Pacheco, A. S., et al. (2020). Whale Watching in Northern Peru: An Economic Boom? Tourism Mar. Environ. 15, 1–10. doi: 10.3727/154427320X15819596320544
Harcourt, R., Pirotta, V., Heller, G., Peddemors, V., and Slip, D. (2014). A whale alarm fails to deter migrating humpback whales: an empirical test. Endanger. Species Res. 25, 35–42. doi: 10.3354/esr00614
Heckel, G., Reilly, S. B., Sumich, J. L., and Espejel, I. (2001). The influence of whalewatching on the behaviour of migrating gray whales (Eschrictius robustus) in Todos Santos Bay and surrounding waters, Baja California, Mexico. J. Cetacean Res. Manage. 3, 227–238.
Higginbottom, K., and Tribe, A. (2004). “Contributions of wildlife tourism to conservation,” in Wildlife tourism: Impacts, management and planning, ed. K. Higginbottom (Altona, VIC: Common Ground Publishing Pty Ltd).
Higham, J. E., Bejder, L., Allen, S. J., Corkeron, P. J., and Lusseau, D. (2016). Managing whale-watching as a non-lethal consumptive activity. J. Sustain. Tour. 24, 73–90.
Hoyt, E., and Iñíguez, M. (2008). The State of Whale Watching in Latin America. Chippenham: Yarmouth Port.
Hoyt, E., and Parsons, E. C. M. (2014). “The whale-watching industry,” in Whale-watching: Sustainable tourism and ecological management, eds J. Higham, L. Bejder, and R. Williams (Cambridge: Cambridge University Press), 57–70. doi: 10.1017/CBO9781139018166.006
Kavanagh, A. S., Noad, M. J., Blomberg, S. P., Goldizen, A. W., Kniest, E., Cato, D. H., et al. (2017). Factors driving the variability in diving and movement behavior of migrating humpback whales (Megaptera novaeangliae): Implications for anthropogenic disturbance studies. Mar. Mamm. Sci. 33, 413–439. doi: 10.1111/mms.12375
Lenth, R., Singmann, H., Love, J., Buerkner, P., and Herve, M. (2018). Emmeans: Estimated marginal means, aka least-squares means. R Package Ver. 1:3.
Lesage, V., Omrane, A., Doniol-Valcroze, T., and Mosnier, A. (2017). Increased proximity of vessels reduces feeding opportunities of blue whales in the St. Lawrence Estuary, Canada. Endanger. Species Res. 32, 351–361. doi: 10.3354/esr00825
Luna-Jorquera, G., Simeone, A., and Aguilar, R. (2003). “Ecofisiología de Animales Endotermos en un Desierto Cálido y un Mar Frío: el Caso de las Aves Marinas de la Corriente de Humboldt,” in Fisiología Ecológica y Evolutiva: Teoría y casos de estudios en animales, ed. F. Bozinovic (Santiago: Universidad Católica de Chile), 297–316.
Lusseau, D. (2005). Residency pattern of bottlenose dolphins Tursiops spp. in Milford Sound, New Zealand, is related to boat traffic. Mar. Ecol. Prog. Ser. 295, 265–272. doi: 10.3354/meps295265
Lusseau, D., and Bejder, L. (2007). The Long-term Consequences of Short-term Responses to Disturbance Experiences from Whalewatching Impact Assessment. Int. J. Comp. Psychol. 20, 228–236.
Mann, J. (1999). Behavior sampling methods for cetaceans: a review and critique. Mar. Mamm. Sci. 15, 102–122. doi: 10.1111/j.1748-7692.1999.tb00784.x
Montecino, V., Paredes, M., Paolini, P., and Rutllant, J. (2006). Revisiting chlorophyll data along the coast in north-central Chile, considering multiscale environmental variability. Rev. Chil. Hist. Nat. 79, 213–223. doi: 10.4067/S0716-078X2006000200007
Morete, M. E., Abras, D., and Martins, C. C. A. (2018). “Land-Based Station Studies of Aquatic Mammals in Latin America: Understanding Behavior for Conservation,” in Advances in Marine Vertebrate Research in Latin America, eds M. Rossi-Santos and C. Finkl (Cham: Springer), 77–112. doi: 10.1007/978-3-319-56985-7_4
Morete, M., Bisi, T. L., Rosso, S., Morete, M. E., Bisi, T. L., and Rosso, S. (2007). 241 (2007). Mother and calf humpback whale responses to vessels around the Abrolhos Archipelago, Bahia, Brazil. J. Cetacean Res. Manage. 9, 241–248.
Nagelkerke, N. J. D. (1991). A note on a general definition of the coefficient of determination. Biometrika 78, 691–692. doi: 10.1093/biomet/78.3.691
Pacheco, A. S., Silva, S., Alcorta, B., Gubbins, S., Guidino, C., Sanchez-Salazar, F., et al. (2019). Cetacean diversity revealed from whale-watching observations in northern Peru. Aquat. Mamm. 45, 116–123. doi: 10.1578/AM.45.1.2019.116
Parsons, E. C. M. (2012). The negative impacts of whale-watching. J. Mar. Biol. 2012:9. doi: 10.1155/2012/807294
Parsons, E. C. M., Warburton, C. A., Woods-Ballard, A., Hughes, A., and Johnston, P. (2003). The value of conserving whales: the impacts of cetacean-related tourism on the economy of rural West Scotland. Aquat. Conserv. Mar. Freshwat. Ecosyst. 13, 397–415. doi: 10.1002/aqc.582
Pérez, M., Thomas, F., Uribe, F., Sepúlveda, M., Flores, M., and Moraga, R. (2006). Fin Whales (Balaenoptera physalus) Feeding on Euphausia mucronata in Nearshore Waters off North-Central Chile. Aquat. Mamm. 32, 109–113. doi: 10.1578/AM.32.1.2006.109
Pirotta, V., Slip, G., Jonsen, I. D., Peddemors, V. M., Cato, D. H., Ross, G., et al. (2016). Migrating humpback whales show no detectable response to whale alarms off Sydney, Australia. Endanger. Species Res. 29, 201–209. doi: 10.3354/esr00712
Piwetz, S., Gailey, G., Munger, L., Lammers, M. O., Jefferson, T. A., and Würsig, B. (2018). Theodolite tracking in marine mammal research: From Roger Payne to the present. Aquat. Mamm. 44, 683–693. doi: 10.1578/AM.44.6.2018.683
R Core Team (2018). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Rigby, R. A., and Stasinopoulos, D. M. (2005). Generalized additive models for location, scale and shape (with discussion). J. R. Stat. Soc. Ser. C 54, 507–554. doi: 10.1111/j.1467-9876.2005.00510.x
Schaffar, A., Garrigue, C., and Constantine, R. (2010). Exposure of humpback whales to unregulated whalewatching activities in their main reproductive area in New Caledonia. J. Cetacean Res. Manage. 11, 147–152.
Schaffar, A., Madon, B., Garrigue, C., and Constantine, R. (2013). Behavioural effects of whale-watching activities on an Endangered population of humpback whales wintering in New Caledonia. Endanger. Species Res. 19, 245–254. doi: 10.3354/esr00466
Scheidat, M., Castro, C., Gonzalez, J., and Williams, R. (2004). Behavioural responses of humpback whales (Megaptera novaeangliae) to whalewatching boats near Isla de la Plata, Machalilla National Park, Ecuador. J. Cetacean Res. Manage. 6, 63–68.
Schuler, A. R., and Pearson, H. C. (2019). Conservation benefits of whale watching in Juneau, Alaska. Tourism Mar. Environ. 14, 231–248. doi: 10.3727/154427319X15719404264632
Schuler, A. R., Piwetz, S., Di Clemente, J., Steckler, D., Mueter, F., and Pearson, H. C. (2019). Humpback whale movements and behavior in response to whale-watching vessels in Juneau, AK. Front. Mar. Sci. 6:710. doi: 10.3389/fmars.2019.00710
Senigaglia, V., Christiansen, F., Bejder, L., Gendron, D., Lundquist, D., Noren, D. P., et al. (2016). Meta-analyses of whale-watching impact studies: comparisons of cetacean responses to disturbance. Mar. Ecol. Prog. Ser. 542, 251–263. doi: 10.3354/meps11497
Sepúlveda, M., Inostroza, P., Pérez-Alvarez, M. J., Oliva, D., and Moraga, R. (2009). Seasonal variation in the abundance of South American sea lions Otaria flavescens (Shaw, 1800) in Chañaral Island, Reserva Nacional Pinguüino de Humboldt, Chile. Rev. Biol. Mar. Oceanogr. 44, 685–689.
Sepúlveda, M., Oliva, D., Pavez, G., and Santos-Carvallo, M. (2016). Caleta Chañaral de Aceituno: Destino turístico de alta calidad para el avistamiento de cetáceos, otros mamíferos y aves marinas. Chile: Sernapesca
Sepúlveda, M., Pérez-Álvarez, M. J., Santos-Carvallo, M., Pavez, G., Olavarría, C., Moraga, R., et al. (2018). From whaling to whale watching: Identifying fin whale critical foraging habitats off the Chilean coast. Aquat. Conserv. Mar. Freshwat. Ecosyst. 28, 821– 829. doi: 10.1002/aqc.2899
Sernapesca (2020). Aprueba la regulación de las actividades de buceo recreativo, paseo náutico guiado de observación de flora y fauna, traslado y apoyo de buzos recreativos dentro de la Reserva Marina Isla Chañaral y deja sin efecto la Res.Ex. N° 6248 de 2017 y su modificación. Res.Ex. N°655/2020. Chile: Sernapesca.
Sprogis, K. R., Bejder, L., Hanf, D., and Christiansen, F. (2020a). Behavioural responses of migrating humpback whales to swim-with-whale activities in the Ningaloo Marine Park, Western Australia. J. Exp. Mar. Biol. Ecol. 522:151254. doi: 10.1016/j.jembe.2019.151254
Sprogis, K. R., Videsen, S., and Madsen, P. T. (2020b). Vessel noise levels drive behavioural responses of humpback whales with implications for whale-watching. Elife 9:e56760. doi: 10.7554/eLife.56760
Stamation, K. A., Croft, D. B., Shaughnessy, P., Waples, K. A., and Briggs, S. V. (2010). Behavioral responses of humpback whales (Megaptera novaeangliae) to whale-watching vessels on the southeastern coast of Australia. Mar. Mamm. Sci. 26, 98–122. doi: 10.1111/j.1748-7692.2009.00320.x
Stasinopoulos, D. M., Rigby, R. A., Heller, G. Z., Voudouris, V., and De Bastiani, F. (2017). Flexible regression and smoothing: using GAMLSS in R. London: Chapman & Hall. doi: 10.1201/b21973
Subpesca (2005). Declara Reserva Marina Espacio Marítimo en Torno a Isla Chañaral, III Región, D.S. No. 150-05. Chile: Subpesca.
Subpesca (2011). Reglamento General de Observación de Mamíferos Reptiles y aves Hidrobiológicas y del Registro de Avistamiento de Cetáceos, D.S. N° 38-2011, 8. Chile: Subpesca.
Thiel, M., Macaya, E., Acuña, E., Arntz, W., Bastias, H., Brokordt, K., et al. (2007). The Humboldt current system of Northern and Central Chile. Oceanogr. Mar. Biol. Annu. Rev. 45, 195–344. doi: 10.1201/9781420050943.ch6
Toro, F., Vilina, Y. A., Capella, J. J., and Gibbons, J. (2016). Novel coastal feeding area for eastern south Pacific fin whales (Balaenoptera physalus) in mid-latitude Humboldt current waters off Chile. Aquat. Mamm. 42, 47–55. doi: 10.1578/AM.42.1.2016.47
Walker, T. R., Adebambo, O., Feijoo, M. C. D. A., Elhaimer, E., Hossain, T., Edwards, S. J., et al. (2019). “Environmental effects of marine transportation,” in World Seas: An Environmental Evaluation, ed. C. Sheppard (Cambridge: Academic Press), 505–530. doi: 10.1016/B978-0-12-805052-1.00030-9
Whitehead, H. (1983). Structure and stability of humpback whale groups off Newfoundland. Can. J. Zool. 61, 1391–1397. doi: 10.1139/z83-186
Williams, R., and Ashe, E. (2007). Killer whale evasive tactics vary with boat number. J. Zool. 272, 390–397. doi: 10.1111/j.1469-7998.2006.00280.x
Williams, R., Bain, D. E., Smith, J. C., and Lusseau, D. (2009). Effects of vessels on behaviour patterns of individual southern resident killer whales Orcinus orca. Endanger. Species Res. 6, 199–209. doi: 10.3354/esr00150
Williams, R., Lusseau, D., and Hammond, P. S. (2006). Estimating relative energetic costs of human disturbance to killer whales (Orcinus orca). Biol. Conserv. 133, 301–311. doi: 10.1016/j.biocon.2006.06.010
Williams, R., Trites, A. W., and Bain, D. E. (2002). Behavioural responses of killer whales (Orcinus orca) to whale-watching boats: opportunistic observations and experimental approaches. J. Zool. 256, 255–270. doi: 10.1017/S0952836902000298
Würsig, B., Cipriano, F., and Würsig, M. (1991). “Dolphin movement patterns: information from radio and theodolite tracking studies,” in Dolphin Societies: Discoveries and Puzzles, eds K. Pryor and K. Norris (Berkeley: University of California Press), 79–111.
Yazvenko, S. B., Mcdonald, T. L., Blokhin, S. A., Johnson, S. R., Melton, H. R., Newcomer, M. W., et al. (2007). Feeding of western gray whales during a seismic survey near Sakhalin Island, Russia. Environ. Monit. Assess. 134, 93–106.
Zeppel, H., and Muloin, S. (2008). Conservation benefits of interpretation on marine wildlife tours. Hum. Dimens. Wildl. 13, 280–294. doi: 10.1080/10871200802187105
Keywords: traveling, resting, movement pattern analysis, land-based tracking, tourism effects, Chile
Citation: Santos-Carvallo M, Barilari F, Pérez-Alvarez MJ, Gutiérrez L, Pavez G, Araya H, Anguita C, Cerda C and Sepúlveda M (2021) Impacts of Whale-Watching on the Short-Term Behavior of Fin Whales (Balaenoptera physalus) in a Marine Protected Area in the Southeastern Pacific. Front. Mar. Sci. 8:623954. doi: 10.3389/fmars.2021.623954
Received: 30 October 2020; Accepted: 16 February 2021;
Published: 08 March 2021.
Edited by:
Rebecca Dunlop, The University of Queensland, AustraliaReviewed by:
Vanessa Pirotta, Macquarie University, AustraliaLeslie New, Washington State University Vancouver, United States
Copyright © 2021 Santos-Carvallo, Barilari, Pérez-Alvarez, Gutiérrez, Pavez, Araya, Anguita, Cerda and Sepúlveda. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Macarena Santos-Carvallo, bWFjYXJlbmEuc2FudG9zQHV2LmNs