 Frederick Toro1,2,3*
Frederick Toro1,2,3* Jaime Alarcón4
Jaime Alarcón4 Bárbara Toro-Barros2Gabriela Mallea2
Bárbara Toro-Barros2Gabriela Mallea2 Juan Capella5,6
Juan Capella5,6 Clara Umaran-Young2,7Paulette Abarca1,8
Clara Umaran-Young2,7Paulette Abarca1,8 Nelly Lakestani3Claudia Peña2
Nelly Lakestani3Claudia Peña2 Mario Alvarado-Rybak3,9Franco Cruz1Yerko Vilina10Jorge Gibbons11
Mario Alvarado-Rybak3,9Franco Cruz1Yerko Vilina10Jorge Gibbons11- 1Escuela de Medicina Veterinaria, Facultad de Recursos Naturales y Medicina Veterinaria, Universidad Santo Tomás, Viña del Mar, Chile
- 2ONG Panthalassa, Red de Estudios de Vertebrados Marinos en Chile, Santiago, Chile
- 3Ph.D. Program in Conservation Medicine, Facultad de Ecologia y Recursos Naturales, Universidad Andrés Bello, Santiago, Chile
- 4Center for Bioinformatics and Integrative Biology, Facultad de Ciencias de la Vida, Universidad Andrés Bello, Santiago, Chile
- 5Whalesound Ltd., Punta Arenas, Chile
- 6Fundación Yubarta, Cali, Colombia
- 7Department of Biology, University of Naples Federico II, Naples, Italy
- 8Programa de Magíster en Ciencias, Mención Biodiversidad y Conservación, Universidad de Valparaíso, Valparaíso, Chile
- 9Centro de Investigación Para la Sustentabilidad, Facultad de Ciencias de la Vida, Universidad Andrés Bello, Santiago, Chile
- 10Facultad de Medicina Veterinaria, Universidad Santo Tomás, Santiago, Chile
- 11Instituto de la Patagonia, Universidad de Magallanes, Punta Arenas, Chile
Standardized measures of behavior can be powerful tools for assessing the impact of whale watching activities on natural populations of cetaceans. To determine the possible impact of tourism on dolphins between a period without whale watching (1989–1992) (T1) and a period with whale watching (2010–2020) (T2), we examined the changes in the rate of surface behaviors, the group size of long-time resident bottlenose dolphins living in the waters of the Humboldt Current off Chile, and for T2 alone, we compared these differences between two localities, the Punta de Choros and Chañaral de Aceituno coves. We observed a significant decrease in the group size of the resident population and in the frequency of surface events associated with the absence and presence of tourism. For T2, we observed significant differences for the frequency of surface events between the Chañaral de Aceituno and Punta de Choros coves and differences in the frequency of surface events at different hours of the day. This was associated with the number of vessels at the time of the encounter. In addition, we observed for T2 that the most observed instantaneous response of the dolphins to the presence of tourist vessels was to avoid the boats, while approaching the boats was the least observed response. The number of vessels present in each dolphin encounter was the most important variable for our model as it explains these differences. These results show that tourism vessels have a significant impact on dolphin behavior and sociability, while the same population of dolphins have different spatial and temporal responses to different impacts of tourism. Further studies are needed to establish whether changes in the rate of surface behaviors are associated with higher levels of stress in dolphins and with effects on their health and reproductive success in the long term.
Introduction
Over the last two decades, commercial boat-based whale watching has exponentially increased in the coastal areas of the world (Hoyt, 2001; Weir and Pierce, 2012; Hoyt and Parsons, 2014; Silva, 2015). Consequently, it has raised concerns about the potential negative impact these activities might have on cetaceans (Richardson et al., 1995; Nowacek et al., 2007). Some studies have shown changes in the behavior of resident dolphin populations depending on the species and the type and number of vessels present (Ford et al., 1996; Bejder et al., 2006; La Manna et al., 2013; Pirotta et al., 2015; New et al., 2020). These changes can be reflected as shifts in distribution (Mattson et al., 2005; Lemon et al., 2006) and feeding sites (Stockin et al., 2008), differences in vocal behavior, and in the frequency of surface behaviors, such as body posture and leaping (Lusseau, 2003a).
In Chile, boat-based whale watching was first developed in the mid-1990s in Chañaral de Aceituno cove, and focused on a school of 40–45 bottlenose dolphins discovered in 1987 next to the west coast of Chañaral Island (29° 01′S, 71° 37′W) (González et al., 1989). These dolphins have been studied with photo-identification comparisons for about 30 years (González et al., 1989; Gibbons, 1992; Capella et al., 1999; Thomas, 2005; Molina, 2006; Cruz, 2011; Toro, 2011). By means of photo-ID, Gibbons (1992) established the residence of these dolphins in this area, where they remained at least until 1995 (Capella et al., 1999). Subsequently, the dolphins moved 28 km to the south coast of Choros Island (29° 15′S, 71° 33′W) (Capella et al., 1999; Sanino and Yáñez, 2001) and between 2000 and 2020 (Perez-Alvarez et al., 2018; Santos-Carvallo et al., 2018), the school has been seen at both sites, keeping the individual composition unchanged for a significant number of members (Thomas, 2005; Molina, 2006; Cruz, 2011).
Even though whale watching was developed in Chañaral de Aceituno cove in the mid-1990s, it extended to Punta de Choros cove at the end of 90s, in response to the shift in distribution of the bottlenose dolphins (Hanshing, 2001). The regular presence of these resident bottlenose dolphins (Gibbons, 1992; Sanino and Yáñez, 2001; Thomas, 2005; Molina, 2006; Cruz, 2011; Toro, 2011) and co-occurrence of a high diversity of marine mammals (Capella et al., 1999, Capella et al., Unpublished data), and also the growth of tourism in the city of La Serena, which is a city 117 km south of Punta de Choros, led to an explosive growth in local whale watching over the last decade. The number of visitors and vessels registered in the area of Punta de Choros–Chañaral de Aceituno coves, shows an increase from just one vessel and a hundred visitors in 1995, 72 vessels and over 51,000 visitors in 2016 (Hanshing, 2001; Toro, 2011, P. Arrospide, pers. comm., March 2017), and up to receiving over 29,000 visitors in the 2020 whale watching season (Hanshing, 2001; Toro, 2011, P. Arrospide, com. Pers.).
In this article, we assess the impact of whale-watching vessels off the Punta de Choros–Chañaral de Aceituno coves on the surface behavior and group size of resident bottlenose dolphins in Chañaral Island and Choros and Damas Island, respectively, between 2010 and 2020 (T2). We compare the rates of surface behavior observed in a first period from 1989 to 1992 (T1) without whale-watching vessels to a second period from 2010 to 2020 (T2) when whale-watching vessels were present and the differences in the rates of surface behavior between the morning and afternoon for both periods. We also compared dolphin behavior and responses to boats during T2 with varying numbers of vessels present during sightings, with a low number of vessels in the morning and a higher number in the afternoon.
Materials and Methods
Study Area
The study area located in the northern limit of the upwelling zone of Coquimbo Bay in the cold waters of the Humboldt Current off a 10 km stretch of mainland coast includes the waters around three coastal islands belonging to the Humboldt Penguin National Reserve (Figure 1). Chañaral Island located further north only offers whale watching from Chañaral de Aceituno cove, and Choros and Damas islands, the other two southern islands offer whale watching from the Punta de Choros cove (29° 15′S).
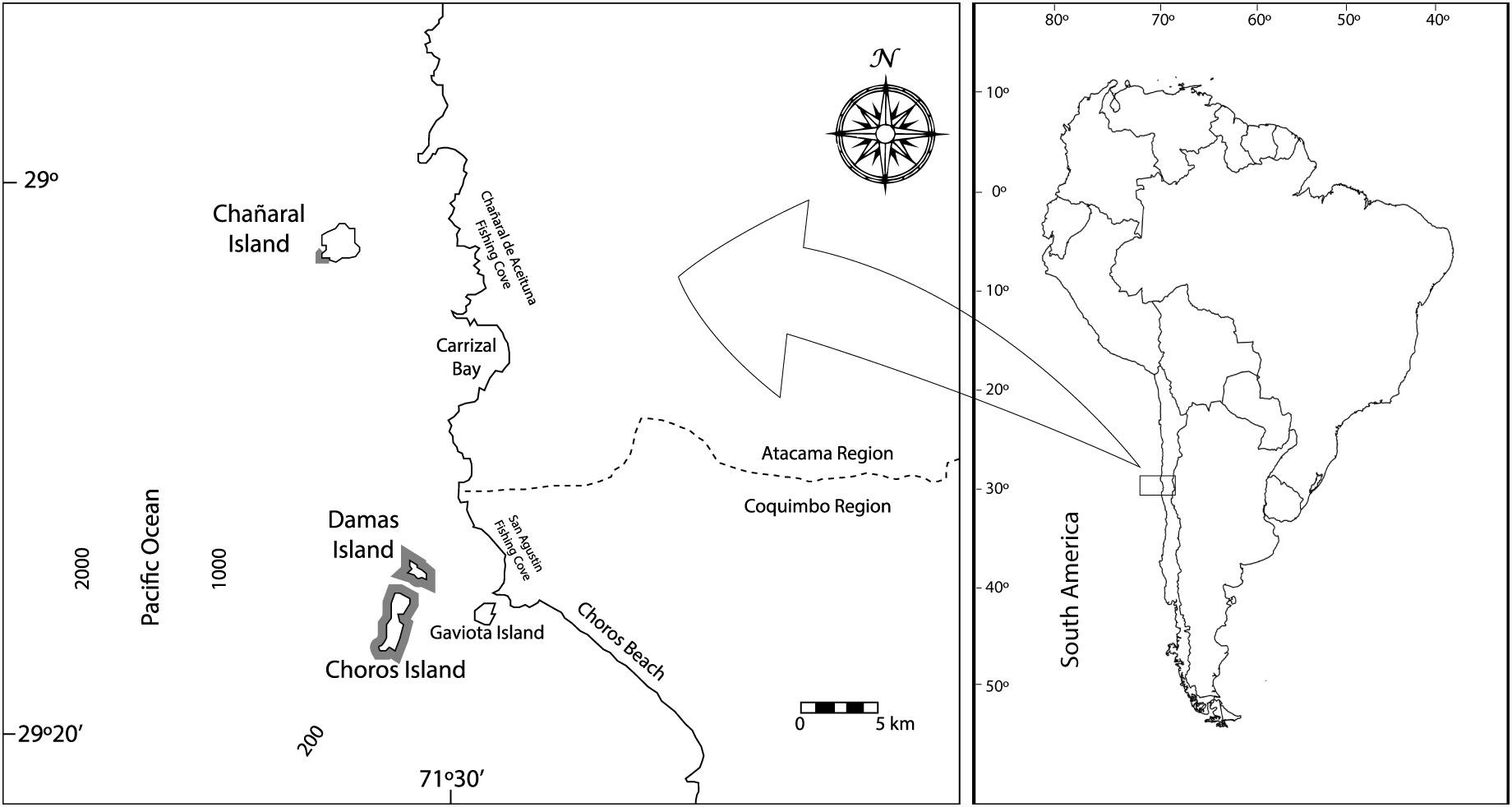
Figure 1. Map of the study area in coastal waters of the Humboldt Current, northern Chile. The gray shaded area at Southwest of Chañaral Island shows the study area for the T1 period without whale watching (1989–1992) and around Chañaral and Damas, Gaviotas, and Choros Islands for T2 with whale watching (2010–2020). In both areas, at least 50% of dolphins were the same individuals.
The focus group studied corresponds to a small resident population of 45–50 bottlenose dolphins. At least 50% of the individuals have been living in the area from 1987 to 2020 (Capella et al., unpublished data; Vilina et al., 1995) and moving among the coastal waters of the three islands.
During the spring–summer seasons of 1989–1990 years (T1), surface behaviors of a single group of bottlenose dolphins were monitored by at least two observers using binoculars (8 × 30) from a cliff 30 masl on the south-west coast of Chañaral Island. A total of 10 surface behaviors (Table 1; Gibbons, 1992; Bearzi et al., 1999; Würsig and Whitehead, 2009) were continuously observed by recording all events (Bearzi et al., 1999; Mann, 1999; Bearzi et al., 2005; Wiliams et al., 2006) from 8:00 to 17:00 h (n = 78 days, 508.2 h) between December and February (Gibbons, 1992). Group size was determined by maximum counts every 5 min.

Table 1. Details of definitions of surface behavior and the response to boat disturbance for bottlenose dolphins (Tursiops truncatus) (Bearzi et al., 1999 and Bearzi et al., 2005 modified).
During 10 successive years between 2011 and 2020 (T2), the dolphins moved in a more scattered and unpredictable way throughout the study area, making it impossible to monitor their behavior and group size systematically from land. Data about the occurrence of surface behaviors and group size were collected during 357 brief encounters (16-min average for each effective encounter) from whale-watching vessels (11 m in length, four-stroke 100 HP engine) in the coastal waters off Chañaral island and the Choros–Damas islands (Figure 1) between 8:00 and 17:00 h (n = 421 days; 3,723 h), on trips lasting 2–3 h between January and February, with two observers.
In addition, during T2, instantaneous responses from each individual dolphin alerted to the presence of tourist vessels from Chañaral de Aceituno cove and Punta de Choros cove were recorded for Chañaral island and the Choros–Damas islands, respectively. The four instantaneous responses are described in Table 1. For all measurements, between 8:00 and 12:00 was classified as AM and between 12:00 and 17:00 was classified as PM.
Analysis
For statistical comparisons of behavioral events, we used a ratio for behavioral events to group sizes for each encounter (number of events/group size). Statistical analyses were performed in R v 4.0.2 and R studio v 1.3.1073 and plotted with the R base options and the ggplot2 package. To determine the normality of the sighting and effort variables, we performed a Shapiro–Wilk normality test. To determine the differences between group size with the presence and absence of tourist vessels, we used a Kruskal–Wallis rank sum test and pairwise comparisons using a Wilcoxon rank sum test with continuity correction.
A generalized linear model (GLM) with normal distribution was built to verify the association between the frequency of surface behaviors and the presence of whale-watching vessels (T1 and T2); time of the day; month, year, and number of vessels. Generalized linear models (GLM) were built using the “glm” function for the package stats, with the argument family = “binomial” and a p-value threshold of <0.05 for significant predictors. All the models were plotted using the package effects and residuals were analyzed for normality and homoscedasticity. One-way ANOVA was used to compare the AM and PM frequency of behaviors to T1 and T2 and for the number of vessels for T2, followed by the post hoc Tukey multiple comparison test to compare the number of surface behaviors in different numbers of vessels, using the package “multcomp” V1.4-10 in the R statistical language (Westfall et al., 1999).
To establish a relationship between the number of vessels and the response of bottlenose dolphins, we made decision trees using the caret and randomForest packages with a split of the observations into 70% training and 30% testing datasets.
Results
In T1, we had a total of 58 days of observation of dolphins from land. Observations were made in morning and afternoon. The observation time varied between 130 and 235 min in the morning and 200 and 355 min in the afternoon. For T2, 357 encounters with groups of resident dolphins were recorded. Of these encounters, 279 were in Punta de Choros and 78 in Chañaral de Aceituno. Of the total number of encounters, 175 correspond to AM hours (146 Punta de Choros and 29 Chañaral de Aceituno) and 182 correspond (133 Punta de Choros and 49 Chañaral de Aceituno) to PM hours. For T2, effective encounters had an average duration of 16 min (max: 31 min; min: 4 min).
The average group size in T1 was 42 individuals (median = 43, range: 40–45) for only one distinguished cohesive group (Gibbons, 1992). During T2, between one to five groups (median = 8, range: 1–15) were seen in both areas during a working day (Figure 2). No significant differences were observed in the group size, neither between the coves and hours of the day nor among encounters with different numbers of whale-watching vessels.
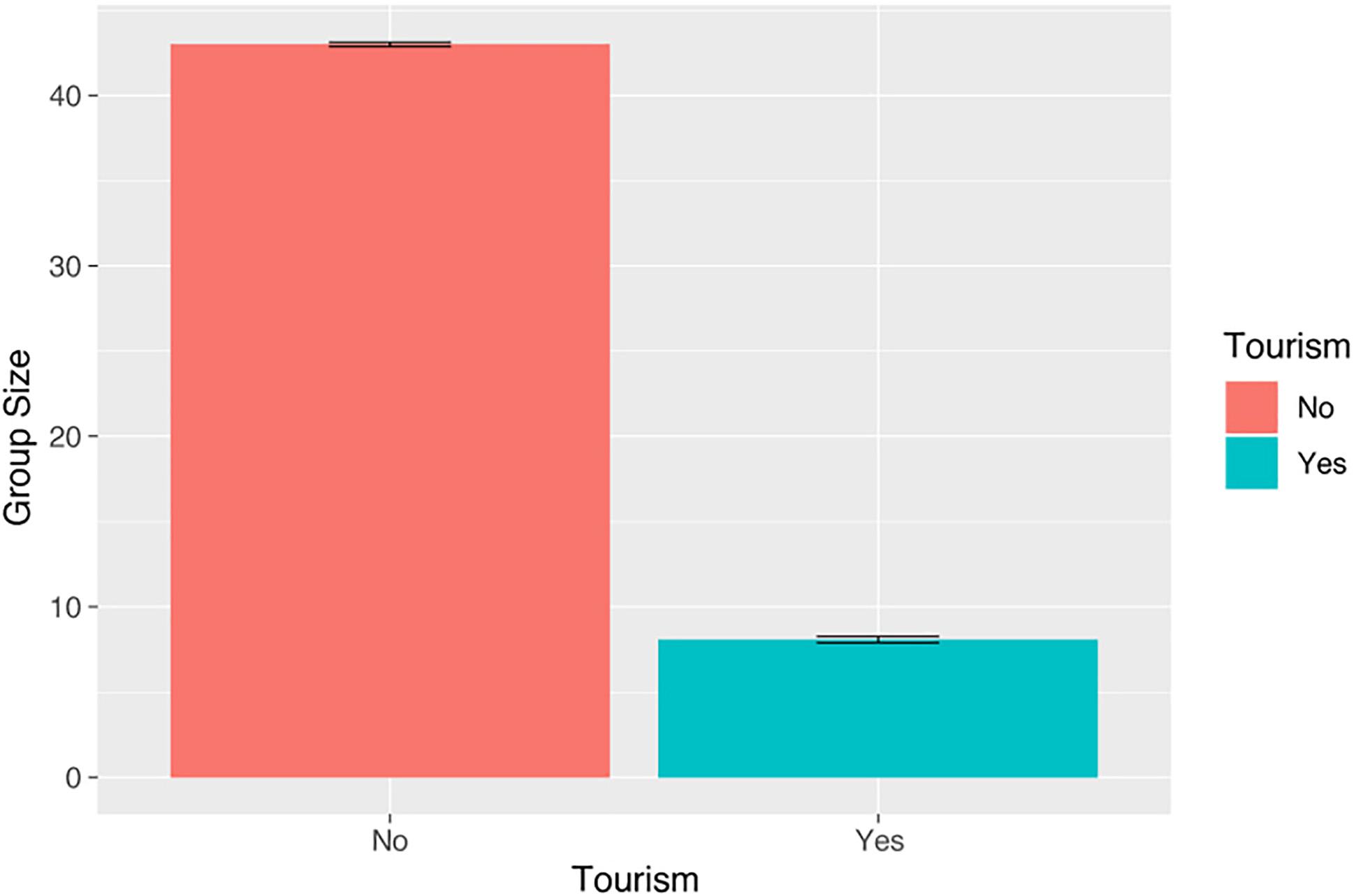
Figure 2. Bottlenose dolphin group sizes in T1 (without whale watching) and T2 (with whale watching) in the study area. The horizontal line in the box represents the mean and the limits in the percentiles 25 and 75. The error bar indicates the maximum and minimum values.
In T1, a total of 5,708 surface behaviors (rate: 4 surface behaviors per individual) were recorded during 6,963 min of dedicated observations at the coast of Chañaral Island. Of these observations, 2,880 min were dedicated to AM hours, registering 2,983 (rate: 2.8 surface behaviors per individual) surface behaviors. In the PM hours, there were 4,083 min of observations and 2,725 (rate: 4.9 surface behaviors per individual) surface behaviors recorded (Figure 3A).
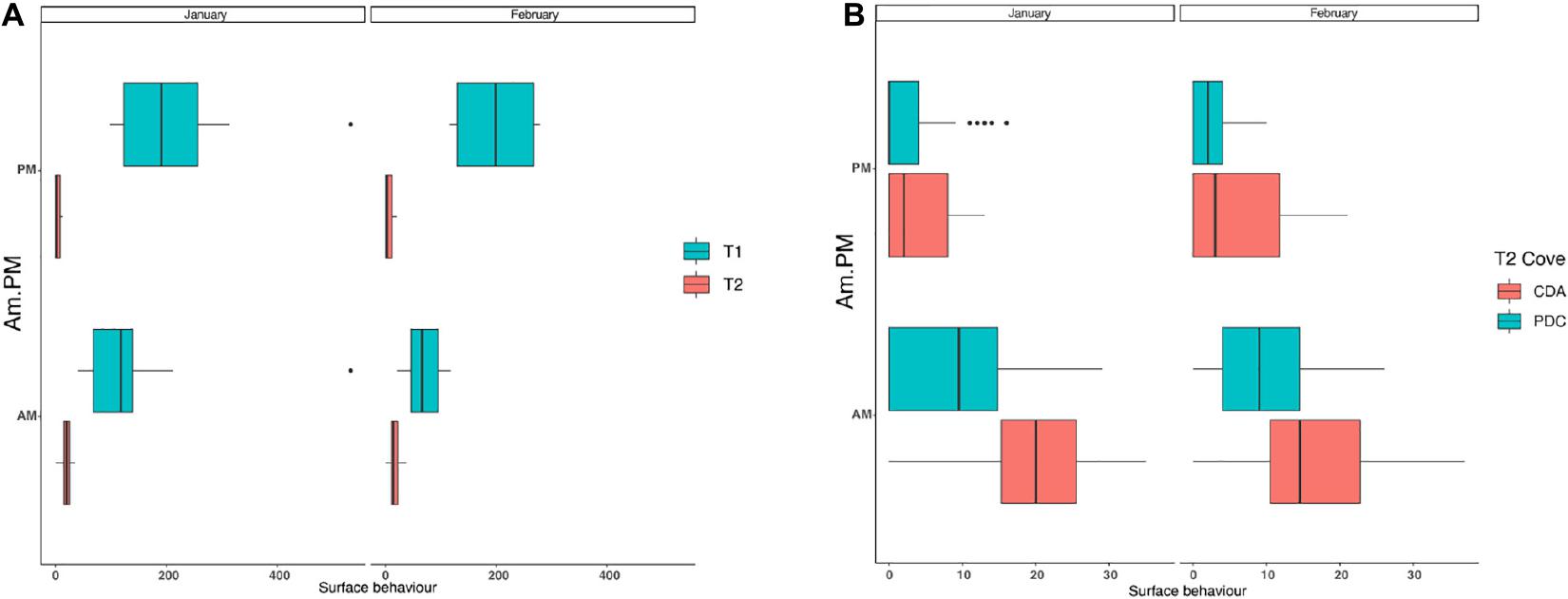
Figure 3. Frequency of occurrence of the ten surface behaviors by bottlenose dolphins in the study area. Without whale watching and with whale watching for the morning (AM) and afternoon (PM) and for January and February (A). Frequency of occurrence of the ten surface behaviors by bottlenose dolphins in the study area in T2 for the Chañaral de Aceituno (CDA) and Punta de Choros (PDC) coves, for the morning (AM) and afternoon (PM) and for January and February (B).
In T2, a total of 2,465 (rate 0.8) surface behaviors were counted in 5,282 min of observations. Surface behaviors were observed in 61.2% of all encounters with dolphins; 57.7% of the meetings in Punta de Choros and 74.6% in Chañaral de Aceituno (Figure 3B). For both T1 and T2 periods, the most observed surface behavior was leaping, however, the proportion of this event increased significantly for T2 (p-value = 0.00134).
For T1, significant differences (p-value = 0.001) were observed in the surface behaviors between AM and PM, with PM being the period with more surface events (Figure 3A). At T2, significant differences between AM and PM were observed, with most surface behaviors for both the Punta de Choros and Chañaral de Aceituno coves occurring in the AM hours (Figure 3B). We found a significant difference in the frequency of surface events in T2 with the presence of tourism and T1 with the absence of tourism (p-value < 3.312e-17) for the Kruskal–Wallis test.
As for the results of the GLM analysis, we found a significant effect for the variables presence of tourism, cove, and years (p-value = 0.05), AM–PM (in T1 and T2), and presence and number of vessels (p-value < 0.0001). For the month variable (January–February), no significant effect was observed on surface events (Figure 3). For T2, we observed significant differences between the frequency of surface behaviors and the presence of whale-watching vessels (ANOVA, p > 0.001) for the Chañaral de Aceituno (Figure 4A) and Punta de Choros coves (Figure 4B). For the post hoc Tukey analysis, we found a significant difference when the number of whale-watching vessels was more than two.
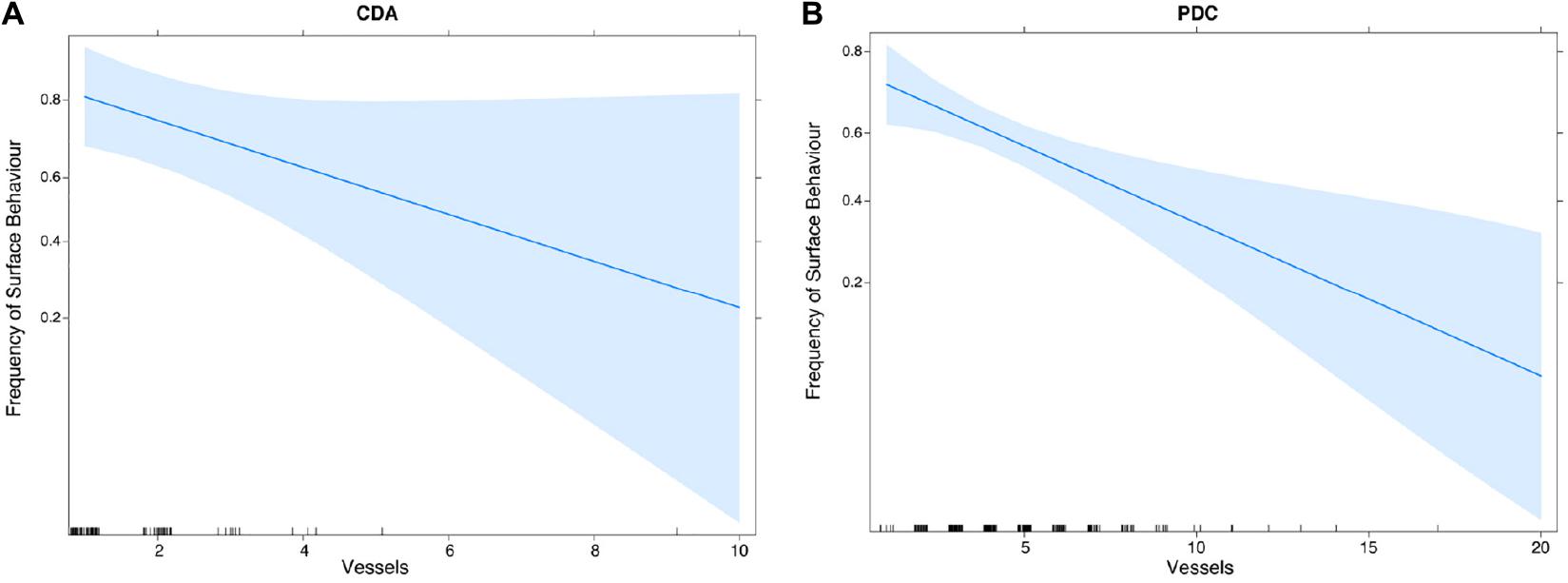
Figure 4. Effect plot of the prediction model based on the data for the frequency of surface behavior and number of vessels present in each encounter with bottlenose dolphins, for the (A) Chañaral de Aceituno (CDA) and (B) Punta de Choros (PDC) coves.
In T2, out of a total 2,883 instantaneous responses to vessels’ disturbance, 51.1% corresponded to avoidance, 30.3% resulted in no response, 13.7% attempted to bowride, and 4.9% approached the boat. From that total, it was found that in Punta de Choros, 63.2% of the responses corresponded to avoidance and it increased for the PM hours, and the least observed response was approaching with 0.3%. Whereas in the Chañaral de Aceituno waters, with fewer whale-watching vessels than Punta de Choros, no response (45.1%) was the most frequent, followed by bowride at 25.8% (Figure 5).
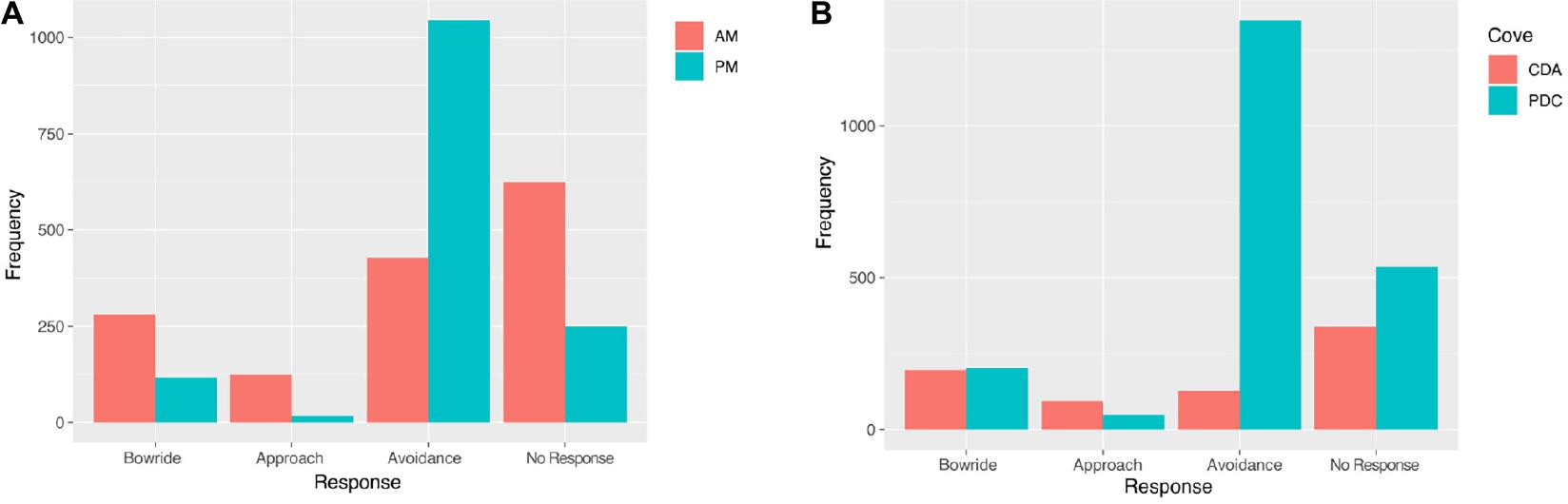
Figure 5. Individual response to the vessels for the morning (AM) and afternoon (PM) (A). Frequency of an individual response to vessel disturbance for the dolphins. Individual responses for the Chañaral de Aceituno (CDA) and Punta de Choros (PDC) cove in T2 (B).
We made a decision tree between the instantaneous responses to the tourist vessels, considering only the data in which the whole group of dolphins responded to the encounter with the vessels. For Punta de Choros, the group of dolphins had a 65% probability of avoidance of the site in the presence of three or more whale-watching vessels, but when the number of vessels was less than three, the group of dolphins had a 35% probability of no response (Figure 6A). For Chañaral de Aceituno, the group of dolphins had a 17% probability of avoidance of the site in the presence of two tourism vessels (Figure 6B).
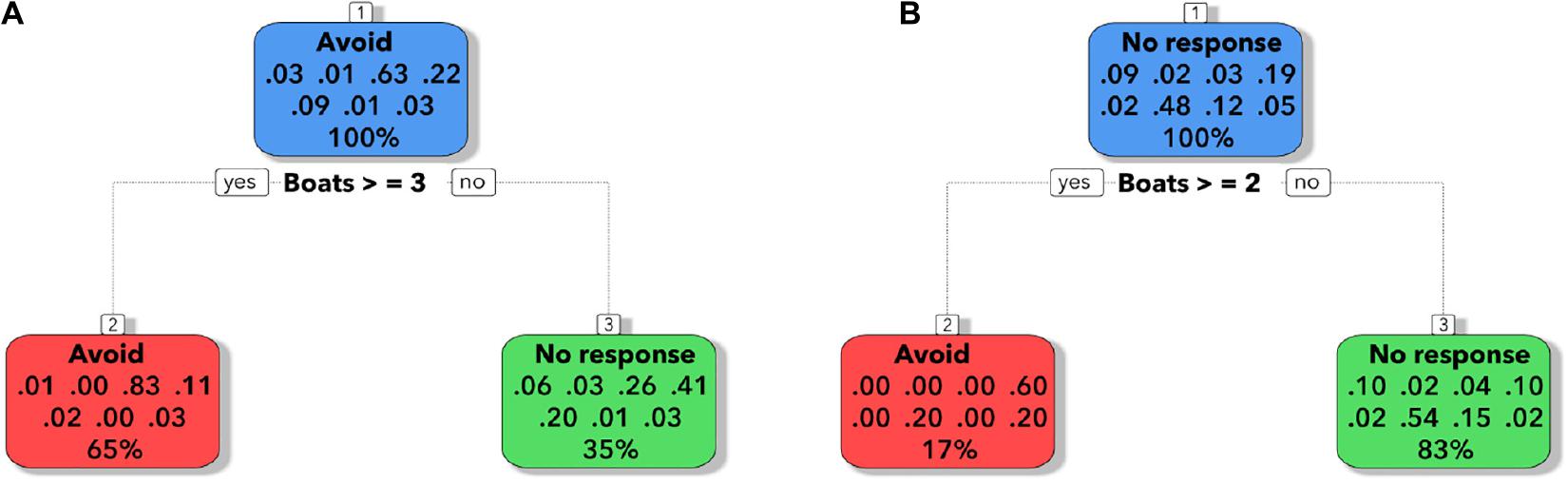
Figure 6. The decision trees show the predicted probability of avoidance or no response for the Punta de Choros dolphin encounters groups (A) and Chañaral de Aceituno dolphin encounters groups (B). Note that the two probabilities add to 100%. The proportion is 100% for the root node.
We found significant differences in the number of vessels at the time between AM and PM for T2 in Punta de Choros (p-value = 0.000234) in the Kruskal–Wallis test. In Punta de Choros, we found an average of five vessels present at each encounter (AM: four vessels; PM: six vessels) and for Chañaral de Aceituno, we found an average of two vessels present per encounter (AM: one vessel; PM: two vessels). Also, it was observed that the high number of vessels present in each encounter was accentuated during the months of February, especially in Punta de Choros cove (Figure 7).
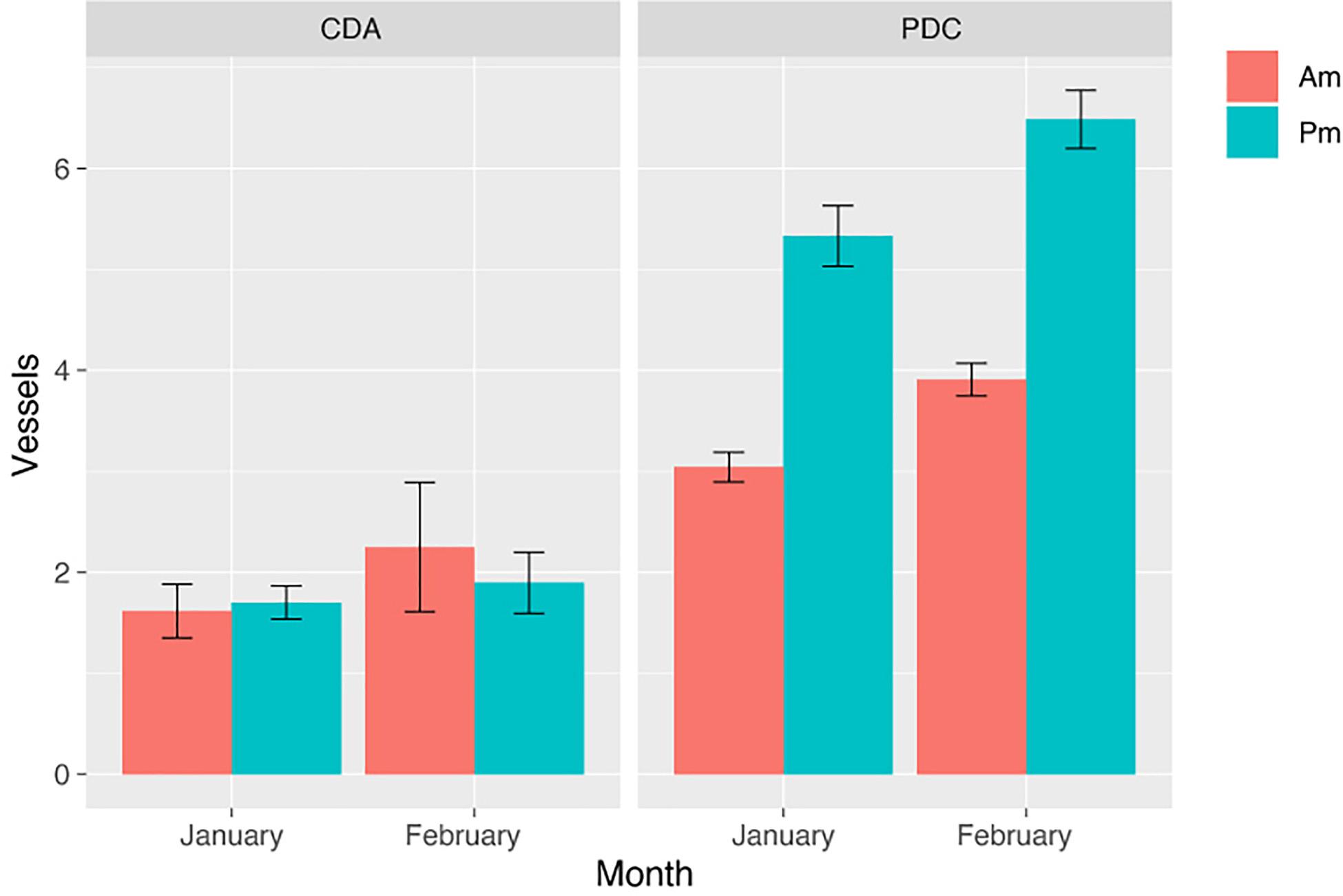
Figure 7. Number of vessels present at each encounter with the dolphins in T2 for both the Chañaral de Aceituno (CDA) and Punta de Choros (PDC) coves, time of day (AM and PM), and January and February. The error bar indicates the standard deviation.
Discussion
Although there are numerous studies of bottlenose dolphins worldwide, there are a limited number of studies on the introduction of whale-watching vessels on the same individuals across time and space, as we describe here for the Chañaral de Aceituno and Punta de Choros coves. The superficial behaviors of dolphins have been associated with different contexts such as levels of alertness, social behaviors, or collaborative foraging behavior (Constantine et al., 2004; Lusseau, 2006). It is in this context that we wanted to verify if whale watching causes some effect on these behaviors.
We demonstrate the impact of whale-watching vessels on resident bottlenose dolphins in northern Chile by lines of complementary evidence. We found a significant decrease in the rate of dolphin surface behaviors associated with the number of whale-watching vessels visits, for the different sites studied. The differences in the rate of dolphin surface behaviors between islands were associated or coincided with the differences in the numbers of whale-watching vessels that visited them.
The same was true for the differences in the rate of behaviors between the morning and afternoon. We ruled out that this daily pattern of surface activities was a consequence of the natural circadian cycle, because during T1 (without whale-watching vessels), we observed more surface events in the afternoon, which differed from T2, when we found a significant decrease in the rate of the dolphins’ surface activity during afternoons with significantly more vessels present compared to mornings. We also considered a possible cumulative daily effect of whale watching on the dolphins’ behavior from morning to afternoon. More frequencies of surface events were recorded in the afternoon in a population of bottlenose dolphins in North America (Henderson, 2004) as well as that observed in the spinner dolphin (Stenella longirostris) in Hawaii (Norris and Dohl, 1980), these studies were in the absence of tourist vessels.
It complements the abovementioned response to boat disturbance, we found that the avoidance response (51.1%) was the most observed, especially in Punta de Choros increasing toward the afternoon (63.2%), whereas in Chañaral de Aceituno, no response was the most observed (45.1%). Our results match with those observed in Guiana dolphins and river dolphins in the Amazon, where in the presence of more than one vessel, the dolphins tend to avoid the vessel or show no response to its presence (Acosta, 2002). This response to the presence of vessels has also been described in bottlenose dolphins (Constantine, 2001; Steckenreuter et al., 2012; Pérez-Jorge et al., 2017), even observing differences in the ways of escape between males and females in New Zealand bottlenose dolphins (Lusseau, 2003b, 2007). Differences in the relative frequency of “leap forward” behavior with and without the presence of tour vessels has already been described in a resident population of bottlenose dolphins in New Zealand (Lusseau, 2003a). It has been speculated that these behaviors would be cost effective methods for visual and acoustic communication (Whitehead and Waters, 1990; Lusseau, 2003b, 2006, 2007; Wiliams et al., 2006; Lusseau et al., 2009), especially in noisy environments (Erbe, 2002). The frequent withdrawal of dolphins from whale-watching vessels and the increase in the rate of “leap forward” with more vessels could be associated with avoiding the source of disturbance, aggressiveness, or it could be a starting behavior for a set of other responses (Williams et al., 2002; Lusseau, 2006).
The decrease in the surface behavior of bottlenose dolphins associated with the presence of vessels observed in our study is similar to that described for other populations of this species that reside in whale-watching vessel activity areas, such as in Cispatá Bay, Colombia (Ávila, 1995), Sarasota, FL, United States (Nowacek et al., 2001), and Doubtful Sound, New Zealand (Lusseau, 2003a); and also for other dolphin species, such as Hector’s dolphins, Cephalorhynchus hectori, in Porpoise Bay, New Zealand (Bejder et al., 1999).
Our results suggest that whale-watching vessels produce short-term changes in surface behavior. Nevertheless, it is necessary to study whether changes in the rate of surface events are associated with greater stress and whether whale-watching vessel activities could have long-term implications such as health, energy budget, reproduction, and dynamics of this resident population of bottlenose dolphins, and could be addressed by application of the PCoD approach (Williams et al., 1992; New et al., 2015, 2020).
For this resident population of bottlenose dolphins, we have observed that, in the last 3 years, at least one stable group of individuals has decreased (unpublished data). In addition, they have been found in a higher proportion in the same area that we describe for T1. This could be associated with a recolonization of the area in response to the high number of tourism vessels in Punta de Choros cove. This type of response has been described in Panama, Croatia, and Australia, where the high number of tourist vessels causes dolphin populations to avoid these areas in the long term, moving toward areas with less pressure from vessels (Lusseau, 2005; Steckenreuter et al., 2012; Rako et al., 2013).
Comprehensive assessment of the impact of whale-watching vessels on this local bottlenose dolphin population is a basic requirement in the establishment of policies of conservation, environmental education, and regulatory standards. The establishment of these policies is a condition sine qua non for the conservation of the local bottlenose population and also for the economic sustainability of the local community. We hope this work will contribute to these objectives.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
Ethical review and approval was not required for the animal study because it is an observational study, where data were taken from tourist boats, so with or without the study the animals would have the presence of the boats.
Author Contributions
FT, JC, JG, and YV led the project and designed the work. FT, JC, JG, YV, BT-B, GM, CU-Y, and PA collected the data and field work. FT, JA, CP, MA-R, BT-B, GM, JG, and JC wrote the manuscript. FT, JA, NL, and MA-R contributed to the statistic analysis, visualization, and plotting results. CP, NL, and FC helped to improve the manuscript. All authors contributed to the article and approved the submitted version.
Funding
Different phases of this long-term research were funded partially by the Comité Nacional Pro Defensa de la Flora y Fauna (CODEFF), Frankfurt Zoological Society (FZS), Departamento de Investigación y Postgrado and Escuela de Medicina Veterinaria of the Universidad Santo Tomás, Fundación Yubarta, Proyectos Exploratorios–Ministerio de Bienes Nacionales, and Agencia Española de Cooperación Internacional para el Desarrollo (grants D/010828/07 and D/010828/08).
Conflict of Interest
JC was employed by the company Whalesound Ltd.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank especially Antonio Larrea, Alberto Maffei, Juan Carlos Gedda, Manuel Gedda, Agnes Kuester, Paolo Sanino, Francisco Radich, Piera Pizolti, Diego Cortes, Humberto Mieres, Pablo Garrido, Marcela Ramírez, Natacha del Pino, Jacqueline González, Pablo Garrido, Patricia Pereira, Lisell Araya Correa, Xiomara Godoy, Alexis Henríquez, María Francica Del Castillo, Constanza Cifuentes, Pablo Parada, José Ignacio Arriagada, Joaquín Parada, Alejandra Fredes, Clara Agusti and Karen Plaza, to María Paulina Godoy for hospitality. We extend our gratitude to Pedro Alvarez, Undalicio Alvarez, Fernando Alvarez, Luis and Jonathan Gonzalez, Ronny, Fabian and Damiano Pachoco and Patricio Ortiz from Chañaral Cove and to Guillermo “Willy” and Maickool Barrera and J. M. Lobos from Punta de Choros Cove and Pablo Arrospide from Corporación Nacional Forestal (CONAF).
References
Acosta, A. (2002). Efecto Del Tráfico de Botes Sobre los Eventos Superficiales de Los Delfines de Rio Inia Geoffrensis y Sotalia Fluviatilis en el Municipio de Puerto Nariño Durante la Época de Aguas Bajas Amazonas, Colombia. Trabajo de Grado Para Optar al Grado de Bióloga. Facultad de Ciencias. Bogotá: Pontificia Universidad Javeriana.
Ávila, J. M. (1995). Aspectos Biológicos Y Etológicos de Delfines Costeros, Con Énfasis en la Especie Sotalia fluviatilis (DELPHINIDAE), en la Bahía de Cispatá, caribe Colombiano (Biological and ethological aspects of the coastal dolphins; with emphasis in the specie Sotalia fluviatilis (DELPHINIDAE), in the Cispata Bay, Colombian caribe) (Pregraduate thesis). Bogotá: Universidad Jorge Tadeo Lozano.
Bearzi, G., Politi, E., Agazzi, S., Bruno, S., Costa, M., and Bonizzoni, S. (2005). Occurrence and present status of coastal dolphins (Delphinus delphis and Tursiops truncatus) in the eastern Ionian Sea. Aquat. Conserv. Mar. Freshwat. Ecosyst. 15, 243–257. doi: 10.1002/aqc.667
Bearzi, G., Politi, E., and di Sciara, G. N. (1999). Diurnal behavior of free-ranging bottlenose dolphins in the kvarnerić (northern adriatic sea) 1. Mar. Mamm. Sci. 15, 1065–1097. doi: 10.1111/j.1748-7692.1999.tb00878.x
Bejder, L., Dawson, S. M., and Harraway, J. A. (1999). Responses by Hector’s dolphins to boats and swimmers in Porpoise Bay, New Zealand. Mar. Mamm. Sci. 15, 738–750. doi: 10.1111/j.1748-7692.1999.tb00840.x
Bejder, L., Samuels, A. M. Y., Whitehead, H. A. L., Gales, N., Mann, J., Connor, R., et al. (2006). Decline in relative abundance of bottlenose dolphins exposed to long-term disturbance. Conserv. Biol. 20, 1791–1798. doi: 10.1111/j.1523-1739.2006.00540.x
Capella, J., Vilina, Y. A., and Gibbons, J. (1999). Observación de cetáceos en Isla Chañaral y nuevos registros para el área de la Reserva Nacional Pingüino de Humboldt, Norte de Chile [Observation of cetaceans in Chañaral island and new records for the area of the Humboldt Penguin National Reserve, north of Chile]. Estud. Oceanol. 18, 57–64.
Constantine, R. (2001). Increased avoidance of swimmers by wild bottlenose dolphins (Tursiops truncatus) due to long-term exposure to swim-with-dolphin tourism. Mar. Mamm. Sci. 17, 689–702. doi: 10.1111/j.1748-7692.2001.tb01293.x
Constantine, R., Brunton, D. H., and Dennis, T. (2004). Dolphin-watching tour boats change bottlenose dolphin (Tursiops truncatus) behaviour. Biol. Conserv. 117, 299–307. doi: 10.1016/j.biocon.2003.12.009
Cruz, F. (2011). Genetica poblacional de delfines nariz de botella, Tursiops truncatus (Montagu, 1821), grupos simpatricos en aguas de un sistema insular de la Corriente de Humboldt [Population genetics of the Bottlenose dolphins, Tursiops truncatus (Montagu, 1821), simpatric groups in the waters at insular system of the Humboldt Current]. Tesis de Médico Veterinario. Universidad Santo Tomás, Santiago.
Erbe, C. (2002). Underwater noise of whale-watching boats and potential effects on killer whales (Orcinus orca), based on an acoustic impact model. Mar. Mamm. Sci. 18, 394–418. doi: 10.1111/j.1748-7692.2002.tb01045.x
Ford, J. K., Ellis, G. M., and Balcomb, K. C. (1996). Killer Whales: The Natural History and Genealogy of Orcinus orca in British Columbia and Washington. Vancouver: UBC press.
Gibbons, J. (1992). Estudio sobre la conducta y dinámica grupal del tursion (Tursiops truncatus) en isla Chañaral, III Región, Chile [Study on the behavior and grupal dinamycs of bottlenose dolphins on Chañaral island, III Region, Chile]. (Tesis de Magíster en Ciencias Biológicas con mención zoología). Santiago: Universidad de Chile.
González, J., Capella, J., and Gibbons, J. (1989). Delfines nariz de botella Tursiops truncatus (Montagu, 1821) habitando frente a Isla Chañaral, Provincia de Huasco, Chile. [Bottlenose dolphins Tursiops truncatus (Montagu, 1821) living in front of Chañaral island, Huasco province, Chile]. Estudios Oceanol. 8, 67–71.
Hanshing, E. S. (2001). Efectos de las embarcaciones turísticas en la población residente de Tursiops truncatus (Montagu, 1821) (Cetácea: Delphinidae) en Isla Choros IV Región [Effects of tourist boats in the resident population of Tursiops truncatus (Montagu, 1821) (Cetacea: Delphinidae) in Choros Island, IV Region]. Tesis de Biólogo Marino. Universidad Católica del Norte, Coquimbo.
Henderson, E. E. (2004). Behaviour, Association Patterns and Habitat use of a Small Community of Bottlenose Dolphins in San Luis Pass, Texas. M.Sc. thesis, Department of Biology, Texas A&M University, Galveston.
Hoyt, E. (2001). Whale Watching 2001: Worldwide Tourism Numbers, Expenditures, and Expanding Socioeconomic Benefits. Yarmouth Port: International Fund for Animal Welfare.
Hoyt, E., and Parsons, E. C. M. (2014). “The whale-watching industry,” in Whale-watching: Sustainable Tourism and Ecological Management, eds J. Higham and L. Bjeder & R. Williams (Yarmouth Port: International Fund for Animal Welfare).
La Manna, G., Manghi, M., Pavan, G., Lo Mascolo, F., and Sarà, G. (2013). Behavioural strategy of common bottlenose dolphins (Tursiops truncatus) in response to different kinds of boats in the waters of Lampedusa Island (Italy). Aquat. Conserv. Mar. Freshw. Ecosyst. 23, 745–757. doi: 10.1002/aqc.2355
Lemon, M., Lynch, T. P., Cato, D. H., and Harcourt, R. G. (2006). Response of travelling bottlenose dolphins (Tursiops aduncus) to experimental approaches by a powerboat in Jervis Bay, New South Wales, Australia. Biol. Conserv. 127, 363–372. doi: 10.1016/j.biocon.2005.08.016
Lusseau, D. (2003a). Male and female bottlenose dolphins Tursiops spp. have different strategies to avoid interactions with tour boats in Doubtful Sound, New Zealand. Mar. Ecol. Prog. Ser. 257, 267–274. doi: 10.3354/meps257267
Lusseau, D. (2003b). The effects of tour boats on the behavior of bottlenose dolphins (Tursiops spp.): using Maskov chains to model anthropogenic impacts. Conserv. Biol. 17, 1785–1793. doi: 10.1111/j.1523-1739.2003.00054.x
Lusseau, D. (2005). Residency pattern of bottlenose dolphins Tursiops spp. in Milford Sound, New Zealand, is related to boat traffic. Mar. Ecol. Prog. Ser. 295, 265–272. doi: 10.3354/meps295265
Lusseau, D. (2006). Why do dolphins jump? Interpreting the behavioural repertoire of bottlenose dolphins (Tursiops sp.) in Doubtful Sound, New Zealand. Behav. Process. 73, 257–265. doi: 10.1016/j.beproc.2006.06.006
Lusseau, D. (2007). Why are male social relationships complex in the Doubtful Sound bottlenose dolphin population? PLoS One 2:e348. doi: 10.1371/journal.pone.0000348
Lusseau, D., Bain, D. E., Williams, R., and Smith, J. C. (2009). Vessel traffic disrupts the foraging behavior of southern resident killer whales Orcinus orca. Endangered Spec. Res. 6, 211–221. doi: 10.3354/esr00154
Mann, J. (1999). Behavioral sampling methods for cetaceans: a review and critique. Mar. Mammal Sci. 15, 102–122. doi: 10.1111/j.1748-7692.1999.tb00784.x
Mattson, M. C., Thomas, J. A., and St Aubin, D. (2005). Effects of boat activity on the behavior of bottlenose dolphins (Tursiops truncatus) in waters surrounding Hilton Head Island, South Carolina. Aquat. Mamm. 31:133. doi: 10.1578/AM.31.1.2005.133
Molina, C. (2006). Estructura Genética de la Población Residente del Delfin Nariz de Botella (Tursiops truncatus) en la Reserva Nacional Pingüino de Humboldt, Chile [Genetic structure of the resident population of bottlenose dolphin (Tursiops truncatus) at Pingüino de Humboldt National Reserve, Chile]. Tesis de Médico Veterinario, Universidad Santo Tomás, Santiago.
New, L., Lusseau, D., and Harcourt, R. (2020). Dolphins and boats: when is a disturbance disturbing. Front. Mar. Sci. 7:353. doi: 10.3389/fmars.2020.00353
New, L. F., Hall, A. J., Harcourt, R., Kaufman, G., Parsons, E. C. M., Pearson, H. C., et al. (2015). The modelling and assessment of whale-watching impacts. Ocean Coast. Manag. 30:e7.
Norris, K. S., and Dohl, T. P. (1980). “The structure and function of cetacean schools,” in Cetacean Behavior: Mechanisms and Functions, ed. L. M. Herman (New York: John Wiley & Sons), 211–261.
Nowacek, D. P., Thorne, L. H., Johnson, D. W., and Tyack, P. L. (2007). Responses of cetaceans to anthropogenic noise. Mamm. Rev. 37, 81–115. doi: 10.1111/j.1365-2907.2007.00104.x
Nowacek, S. M., Wells, R. S., and Solow, A. R. (2001). Short-term effects of boat traffic on bottlenose dolphins, Tursiops truncatus, in Sarasota Bay, Florida. Mar. Mamm. Sci. 17, 673–688. doi: 10.1111/j.1748-7692.2001.tb01292.x
Perez-Alvarez, M. J., Vásquez, R. A., Moraga, R., Santos-Carvallo, M., Kraft, S., Sabaj, V., et al. (2018). Home sweet home: social dynamics and genetic variation of a long-term resident bottlenose dolphin population off the Chilean coast. Anim. Behav. 139, 81–89. doi: 10.1016/j.anbehav.2018.03.009
Pérez-Jorge, S., Louzao, M., Oro, D., Pereira, T., Corne, C., Wijtten, Z., et al. (2017). Estimating the cumulative effects of the nature-based tourism in a coastal dolphin population from southern Kenya. Deep Sea Res. II Top. Stud. Oceanogr. 140, 278–289. doi: 10.1016/j.dsr2.2016.08.011
Pirotta, E., Merchant, N. D., Thompson, P. M., Barton, T. R., and Lusseau, D. (2015). Quantifying the effect of boat disturbance on bottlenose dolphin foraging activity. Biol. Conserv. 181, 82–89. doi: 10.1016/j.biocon.2014.11.003
Rako, N., Fortuna, C. M., Holcer, D., Mackelworth, P., Nimak-Wood, M., Pleslić, G., et al. (2013). Leisure boating noise as a trigger for the displacement of the bottlenose dolphins of the Cres–Lošinj archipelago (northern Adriatic Sea, Croatia). Mar. Pollut. Bull. 68, 77–84. doi: 10.1016/j.marpolbul.2012.12.019
Richardson, W. J., Greene, C. R. Jr., Malme, C. I., and Thomson, D. H. (1995). Marine Mammals and Noise. San Diego, CA: Academic Press.
Sanino, G. P., and Yáñez, J. L. (2001). Nueva técnica de video identificación y estimación de tamaño poblacional en cetáceos, aplicada de delfines nariz de botella, Tursiops truncatus, de isla Choros, IV Región de Chile [New technique of video identification and estimation of population size applied to bottlenose dolphins, Tursiops truncatus to Choros island, IV Region of Chile]. Bol. Mus. Nac. Hist. Nat. Chile 50:3763.
Santos-Carvallo, M., Sepúlveda, M., Moraga, R., Landaeta, M. F., Oliva, D., and Pérez-Alvarez, M. J. (2018). Presence, Behavior, and Resighting Pattern of Transient Bottlenose Dolphins (Tursiops truncatus) in the Humboldt Current System off North-Central Chile1. Pac. Sci. 72, 41–56.
Silva, L. (2015). How ecotourism works at the community-level: the case of whale-watching in the Azores. Curr. Issues Tour. 18, 196–211. doi: 10.1080/13683500.2013.78602
Steckenreuter, A., Möller, L., and Harcourt, R. (2012). How does Australia’s largest dolphin-watching industry affect the behaviour of a small and resident population of Indo-Pacific bottlenose dolphins? J. Environ. Manage. 97, 14–21. doi: 10.1016/j.jenvman.2011.11.002
Stockin, K. A., Lusseau, D., Binedell, V., Wiseman, N., and Orams, M. B. (2008). Tourism affects the behavioural budget of the common dolphin Delphinus sp. in the Hauraki Gulf, New Zealand. Mar. Ecol. Prog. Ser. 355, 287–295. doi: 10.3354/meps07386
Thomas, F. (2005). Residencia del delfín nariz de botella Tursiops truncatus (Montagu, 1821) en la Reserva Nacional Pingüino de Humboldt, Chile [Residence of bottlenose dolphins Tursiops truncatus in the Pingüino de Humboldt National Reserve, Chile]. BSc thesis, Universidad de Valparaíso, Viña del Mar.
Toro, F. (2011). Estudio de las respuestas conductuales de los delfines nariz de botella Tursiops truncatus (Montagu, 1821) a la presencia de botes en aguas de un sistema insular de la Corriente de Humboldt [Study of the behavioral responses of the bottlenose dolphins Tursiops truncatus (Montagu, 1821) to the presence of boats in the waters of an island system of the Humboldt Current]. Tesis de Medico Veterinario, Universidad Santo Tomás, Santiago.
Vilina, Y. A., Capella, J. J., González, J., and Gibbons, J. E. (1995). Apuntes para la conservación de las aves de la Reserva Nacional Pingüino de Humboldt. Bol. Chileno Ornithol. 2, 2–6.
Weir, C. R., and Pierce, G. J. (2012). A review of the human activities impacting cetaceans in the eastern tropical A tlantic. Mamm. Rev. 43, 258–274. doi: 10.1111/j.1365-2907.2012.00222.x
Westfall, P. H., Tobias, R. D., Rom, D., Wolfinger, R. D., and Hochberg, Y. (1999). Multiple Comparisons and Multiple Tests Using the SAS System. Cary, NC: SAS Institute Inc.
Whitehead, H., and Waters, S. (1990). Social organization and population structure of sperm whales off the Galapagos Islands, Ecuador (1985 and 1987). Rep. Int. Whal. Comm. Spec. Issue 29, 249–257.
Williams, R., Lusseau, D., and Hammond, P. S. (2006). Estimating relative energetic costs of human disturbance to killer whales (Orcinus orca). Biol. Conserv. 133, 301–311. doi: 10.1016/j.biocon.2006.06.010
Williams, R., Trites, A. W., and Bain, D. (2002). Behavioural responses of killer whales (Orcinus orca) to whale-watching vessels: opportunistic observations and experimental approaches. J. Zool. 256, 255–270. doi: 10.1017/S0952836902000298
Williams, T. M., Friedl, W. A., Fong, M. L., Yamada, R. M., Sedivy, P., and Haun, J. E. (1992). Travel at low energetic cost by swimming and wave-riding bottlenose dolphins. Nature 355, 821–823. doi: 10.1038/355821a0
Keywords: behavior, bottlenose dolphins (Tursiops truncatus), whale-watching, space effect, Chile, time scale
Citation: Toro F, Alarcón J, Toro-Barros B, Mallea G, Capella J, Umaran-Young C, Abarca P, Lakestani N, Peña C, Alvarado-Rybak M, Cruz F, Vilina Y and Gibbons J (2021) Spatial and Temporal Effects of Whale Watching on a Tourism-Naive Resident Population of Bottlenose Dolphins (Tursiops truncatus) in the Humboldt Penguin National Reserve, Chile. Front. Mar. Sci. 8:624974. doi: 10.3389/fmars.2021.624974
Received: 02 November 2020; Accepted: 08 March 2021;
Published: 23 April 2021.
Edited by:
Rob Harcourt, Macquarie University, AustraliaReviewed by:
Marianne Rasmussen, University of Iceland, IcelandSimon David Berrow, Galway-Mayo Institute of Technology, Ireland
Copyright © 2021 Toro, Alarcón, Toro-Barros, Mallea, Capella, Umaran-Young, Abarca, Lakestani, Peña, Alvarado-Rybak, Cruz, Vilina and Gibbons. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Frederick Toro, ZnJlZGVyaWNrLnRvcm8uY0BnbWFpbC5jb20=