 Jonne Kotta
Jonne Kotta Kaire Torn
Kaire Torn Tiina Paalme
Tiina Paalme- Estonian Marine Institute, University of Tartu, Tallinn, Estonia
An understanding of the nature of scale-dependence in macroalgal production and to quantify how primary production is incorporated into autotrophic biomass requires an assessment of primary production at multiple scales. In this study we experimentally estimated seasonal variability in individual photosynthesis, community production and growth in biomass of the charophyte Chara aspera in the Baltic Sea together with the key environmental variables known to regulate the production of macroalgae. The experiments suggested that the production and growth of C. aspera was defined largely by light and temperature at all studied scales. However, the algal production at the same light levels was systematically lower at the community than an individual level, suggesting the importance of shelf-shading and below-ground processes in natural plant assemblages. Moreover, the observation scale also defined response types between the environment, plant production and growth. This implies that the patterns of variability of the production and growth of macrophytes should always be interpreted in the context of scale and any multiscale model development should involve experimental validation at all important scales.
Introduction
Primary production is important to all ecosystems as it supplies energy to other living organisms, thereby forming the foundation of food webs. Macroalgae are the dominant primary producers in coastal temperate marine ecosystems, representing some of the highest recorded per-area basis production values (Field et al., 1998; Gattuso et al., 2006). As macroalgae contribute significantly to total global system carbon production (Hill et al., 2015) it is important to understand better how macroalgal primary production links to growth in biomass and how different environmental factors affect these patterns.
Traditionally, the primary production of macroalgae has been quantified using detached pieces of algal thally or whole plants (Littler and Littler, 1980; Gómez et al., 1997) and more recently, the production of macroalgae has been measured directly in natural assemblages (Binzer and Middelboe, 2005; Tait and Schiel, 2010). While the first approach enables quantification of accurate response functions between different environmental variables and primary production (e.g., irradiance–net photosynthesis curves), the latter technique allows the scaling up of primary production in natural conditions. The estimates of primary production are highly dependent on the scale of observation with photosynthetic production being higher at individual than at community levels (Binzer et al., 2006; Pärnoja et al., 2014). Likewise, scale affects how macroalgae respond to changes in ambient conditions. The photosynthesis of individual plants often shows a typical light saturation curve (Lobban et al., 1985), whereas macroalgal photosynthesis in assemblages increases in a roughly linear fashion up to maximum levels of natural irradiance (Middleboe and Binzer, 2004). Changes in biomass integrate the net photosynthetic fixation of inorganic carbon into autotrophic biomass as well as the processes of natural loss of biomass and thereby permits estimation of large-scale variation in realized production over time. Expectedly, when compared to the patterns of photosynthesis, another set of combinations of environmental factors can influence change in macroalgal biomass or the same relationships can have very different functional shapes. In this respect, laboratory experiments have shown species-specific responses of macroalgae to environmental change in which macroalgal growth may increase, decrease or remain unchanged with increasing rates of plant photosynthesis (de los Santos et al., 2000; Israel and Hophy, 2002; Kim et al., 2016). Thus, understanding the nature of scale-dependence in macroalgal production can help to elucidate some of the possible controls by which primary production is incorporated into autotrophic biomass. Despite this importance, knowledge of such controls is limited as few studies have been undertaken to date that quantify the patterns of macroalgal production at different scales with most being performed under laboratory conditions (e.g., de los Santos et al., 2000; Israel and Hophy, 2002; Binzer and Middelboe, 2005; Pärnoja et al., 2014; Kim et al., 2016). Due to artificially induced conditions, as well as the use of separate phytoelements (thallus pieces, leaves), however, laboratory incubations often fail to characterize primary production to that which occurs in nature (Binzer and Middelboe, 2005; Binzer et al., 2006).
Primary production is controlled by both abiotic and biotic factors (Field et al., 1998; Hauxwell et al., 2003) but abiotic constraints are often dominant in natural environments (Hill et al., 2015). The availability of light and nutrients plays a crucial role in regulating primary production of macroalgae (Field et al., 1998; Hauxwell et al., 2003; Binzer et al., 2006). Light limitation may arise from the amount of irradiance arriving at the sea surface, the optical characteristics of water and self-shading within algal assemblages. The first two processes vary over multiple temporal and spatial scales and define the availability of light field above underwater canopies and the maximum photosynthetic rates of macroalgae (Kirk, 1994; Anthony et al., 2004). The third process, shelf-shading, is one of the most critical biological limitations because it sets an actual threshold for realized photosynthesis along vertical light gradient within macroalgal assemblages (Binzer et al., 2006; Tait and Schiel, 2010). The effect is stronger with increasing algal densities (Kotta et al., 2008; Paalme et al., 2013; Pärnoja et al., 2014) and in extreme cases, e.g., within giant kelp forests, only a fraction of light above algal canopy reaches the basal subcanopy species (Reed and Foster, 1984). This has large but mostly unknown affects on photosynthesis and growth of macroalgal assemblages. Even though algal photosynthesis is saturated in the upper layers of algal canopies with increasing irradiance, community photosynthesis will remain unsaturated because photosynthetic tissue in the lower layers of algal canopy has an unused photosynthetic potential (Binzer and Middelboe, 2005; Tait and Schiel, 2010).
Nutrient availability strongly affects the production of macroalgae but the responses are mostly taxon-specific and highly influenced by the local light climate (Raven and Hurd, 2012). Importantly, macroalgae can store nutrients in their tissue in order to overcome temporal disparity between nutrient availability and optimal growth season. For example, the marine Laminariales grow fastest during summer when irradiance is the highest, but when nutrients are depleted above the thermocline. During the peak growth the algae use their nitrogen and phosphorus reserves that were stored during winter when the nutrient availability is greater but light levels are lower (Chapman and Craigie, 1977; Lüning, 1990). In addition, the Charales can effectively transport nutrients from rhizoids (in sediment with high N and P but no light) to shoots (in water with low N and P but high light irradiance) (Vermeer et al., 2003). However, when nutrients are plentiful in the water column and light levels are sufficient to support photosynthesis, direct nutrient uptake from the water through the outer cells of the branchlets and stems is expected (Raven, 2003).
Temperature controls primary productivity of macroalgae to a lesser extent than light and nutrients unless the temperature is above or below the tolerances of the species. Within tolerance limits, macroalgae exhibit relative uniform responses to temperature change (Raven and Smith, 1978; Wiencke and tom Dieck, 1990). The species-specific temperature demands for survival and production relate to different types of temperature stress the macroalgal species have locally adapted (Breeman and Pakker, 1994). Controlled experimental trials further demonstrate the high plasticity of response of macroalgae to specific limiting temperature conditions in which a long-term acclimation of macroalgae at extreme temperatures significantly relaxes this temperature limitation (Nejrup et al., 2013). Thus, estimations of primary production in natural assemblages are increasingly relevant given the growing need for obtaining realistic response patterns to a complex of environmental factors.
Charophytes are found in diverse aquatic habitats, so this group consists of both early colonizers of newly-formed water bodies as well as species preferring more stable environmental conditions and habitats (Blindow and van de Weyer, 2016). The lush charophyte meadows play a underpinning role by providing food and/or refuge to zooplankton (Puche et al., 2020), benthic invertebrates (Kotta et al., 2004), fish and fish larvae (Hargeby et al., 2005), microbes (Kataržytė et al., 2017), and waterfowl (Schmieder et al., 2006). They also reduce water turbidity by stabilizing the sediment with rhizoids (Nõges et al., 2003), immobilizing nutrients (Kufel and Kufel, 2002), and inhibiting phytoplankton biomass (Hilt, 2015). Charophytes are most common in freshwater habitats but are also found in brackish water. The Baltic Sea is one of the best-studied brackish regions with respect to charophytes. In this marginal environment, the shallow low salinity embayments are often dominated by charophytes (Schubert and Blindow, 2003). The most common and widespread species in such habitats is Chara aspera Willd., a species characterized by a broad environmental niche space (Herkül et al., 2018).
To address the importance of scale on the factors regulating macroalgal production, we quantified experimentally the seasonal variability in individual photosynthesis, community production and biomass growth of the charophyte C. aspera in the brackish Baltic Sea. We then analyzed how seasonal change in the assemblage of environmental variables manifested in the production of macroalgae at these different scales. We hypothesized that C. aspera with its well-known broad environmental tolerances can maintain high photosynthetic and growth potential across wide gradients in light, temperature, and water nutrients. We also expected that the relationships between the environment and photosynthesis when assessed on individual plants are strong and simple. At the community level, however, charophytes have an ability to adjust photosynthesis and growth to a different set of environmental conditions. Moreover, below-ground processes may occasionally contribute to the oxygen fluxes. Therefore relationships between light, nutrient concentration, temperature, community photosynthesis and growth are weaker and more complex compared to individual plants.
Materials and Methods
Description of Study Area
The study was carried out in Rame Bay from March 2014 to March 2015. Rame Bay is a shallow and semi-enclosed bay in the northeastern Baltic Sea. Depending on the prevailing wind direction, the bay receives eutrophicated waters from the Gulf of Riga or less eutrophied waters from the northern Baltic Proper (Suursaar et al., 1998). The maximum depth of the bay is 1.5 m. The bottom is composed of fine sand and a thick layer of fine silt (Torn and Martin, 2003). Salinity varies between 2 and 5.6 psu with lower values during heavy rainfall. Rapidly increasing water temperatures during spring, high temperatures in summer, and a long period of subzero temperatures in winter are typical. The ice-cover of shallow bays in the region lasts 4–5 months on average (Jevrejeva and Leppäranta, 2002).
Most of Rame Bay is covered by a rich charophyte community. Five species of charophytes are found in the bay with C. aspera being the most widespread species. The seasonal dynamic of charophytes follows general growth patterns of the northern part of the temperate zone, which is characterized by the highest growth rates at the beginning of summer and degradation of the community in autumn (Torn et al., 2006, 2010). C. aspera hibernate both as detached mats and bulbils in the study area (current study, Torn et al., 2010).
Sampling of Environmental Variables and Communities of C. aspera
The study was carried out at depths of 0.9–1.3 m. The study design combined a traditional sampling of water quality variables and benthic communities with the results from the latest oceanographic technology to achieve an accurate environmental assessment of the Rame Bay area. In addition to these observations, the production of C. aspera was measured experimentally at three organizational scales (for more details see below). C. aspera can be visually determined from other charophytes in the study area as it has clearly distinguishable globular white bulbils on the rhizoids. This feature enabled to run the production of C. aspera without a need for prior determination in the laboratory.
The oceanographic instruments were deployed 20 cm above the seabed to record short-term variability of temperature, light availability, and water velocity in the study site from March 2014 to March 2015. These results provided information on the environment around the experimental site during and in between samplings. Temperature was measured with a General Oceanics thermologger. Light availability was recorded by an Odyssey photosynthetically active radiation (PAR) logger. Temperature and PAR was logged at 60 min intervals. Water velocity was recorded by a JFE Advantech Logger version of the INFINITY 2-D electro-magnetic current meter AEMD-USB. During deployment the instrument recorded current speed once per minute. Water level was measured using a JFE Advantech Logger version of the INFINITY-WH AWH-USB wave height meter. During deployment the instrument recorded pressure once per second. Nutrient concentrations in water were measured triplicate once a month. Immediately after collection, samples were preserved and brought to the laboratory in an insulated ice box. We used standard ISO procedures for measuring total N and total P (ISO 11905 and ISO 15681). We did not use neither filtering nor fractioning for our samples. The analyses were made using a Skalar San + + autoanalyzer.
The charophyte community was sampled for coverage and biomass once in a month. Total coverage of all macrophytes and percentage cover of all visually distinguishable species were recorded by SCUBA diver during each sampling. Biomass samples were collected in three replicates by SCUBA diving using a 20 × 20 cm frame. Samples were stored deep frozen (–18°C) until analysis. In the laboratory all species were identified to the species level. The dry weight of species was obtained after drying the individuals at 60°C for 2 weeks (i.e., the time needed to achievethe same weight for two consecutive measurements). Growth in biomass was measured as the difference in total biomass of the plant community between two consecutive sampling months. The chlorophyll-a concentration was measured spectrophotometrically by Libra S32 after extraction of chlorophyll from algal tissue in ethanol. To determine the phosphorus content of the algae, the material was mineralized by heating with HNO3 and HClO4 (Miller, 1998). The content of the elements in the resulting solution was determined by MP-AES spectrometry. The Dumas method was used to measure the nitrogen and carbon content of the algae (Buckee, 1994).
Photosynthetic Production of Individual Plants
In each study visit the net photosynthetic rate of C. aspera was measured in triplicate from 600 ml glass bottles filled with seawater from the study site. Bottles with about 0.5 g dry weight of C. aspera and without algae (controls) were incubated in situ on special frames at a depth of 1 meter around midday. The hourly net production rates (mg O2 g dry weight –1 h–1) were calculated from the differences in oxygen concentrations, measured over the incubation period (1–3 h). Oxygen concentration and water temperature in incubation bottles was measured with OptiOx optical dissolved oxygen sensor connected to SevenGo DO meter (Mettler Toledo). The dry weight of C. aspera was determined after drying at 60°C for 48 h. During the experiment the photosynthetically active radiation (PAR; μmol m–2 s–1) at the incubation depth was measured using Li 188B (Li-Cor Inc.) quantum meter.
Production in Algal Communities
The production of C. aspera communities was measured in triplicate from transparent plexiglas chambers holding 29 l of water with a surface area 850 cm2. The chambers were placed randomly on the seafloor over the homogeneous monospecific stand of C. aspera. A rubber seal assured an airtight seal between the chamber and the surrounding environment.
Oxygen concentration in the chamber was measured every minute using a calibrated Optode-type oxygen sensor (Aanderaa Instruments) connected to a data logger (data recorder by JFE Advantech Co., Ltd.). This instrument also provides data on water temperature. Changes in dissolved oxygen averaged over minute intervals were used as a proxy of community (algal assemblage + sediment) net production. During deployment irradiance above the charophytes was measured every minute using a calibrated spherical quantum sensor connected to a data logger (ultra-miniature logger for light intensity by JFE Advantech Co., Ltd.). Each production experiment lasted 3 h on average.
After deployment, macroalgae within incubation chambers were harvested and stored in a deep freezer at −20°C. The subsequent sorting and determination of species were performed in the laboratory using a stereomicroscope. The dry weight of each macroalgal community was obtained after drying the individuals at 60°C for 2 weeks. Combining this information with the oxygen flux measurements described above enabled us to express net primary production in mg O2 g dry weight per macrophyte min–1.
Statistical Analyses
The contribution of different environmental variables on the individual, community photosynthesis and growth in biomass of C. aspera was explored using the Boosted Regression Trees technique (BRT). BRT models are capable of handling different types of predictor variables and their predictive performance is superior to most traditional modeling methods [see e.g., comparisons with GLM, GAM and multivariate adaptive regression splines, Elith et al. (2006) and Leathwick et al. (2006)]. Overfitting is often regarded as a problem in statistical modeling, but can be overcome by using independent data sets. The BRT modeling iteratively develops a large ensemble of small regression trees constructed from random subsets of the data. Each successive tree predicts the residuals from the previous tree to gradually boost the predictive performance of the overall model (Elith et al., 2008). Important parameters in building BRT models are the learning rate and tree complexity. The learning rate determines the contribution of each tree to the growing model and tree complexity defines the depth of interactions allowed in a model. A tree complexity of 1 assesses only main effects; A tree complexity >1 includes interactions. Different combinations of these parameters may yield variable predictive performance but generally a lower learning rate and inclusion of interactions gives better results (Elith et al., 2008). In the current study, the model learning rate was kept at 0.001 and tree complexity at 5. In order to avoid potential problems of overfitting, unimportant variables were removed using a simplify tool. In order to eliminate non-informative variables, the simplify tool progressively simplifies the model, then re-fits the model and sequentially repeats the process until a stopping criterion is reached. Such simplification is most useful for small data sets where redundant predictors may degrade performance by increasing variance. Model performance was evaluated using the cross-validation statistics calculated during model fitting (Hastie et al., 2009). The BRT modeling was done in R using the gbm package (Elith et al., 2008). Standard errors for the predictions and pointwise standard errors for the partial dependence curves, produced by R package “pdp” (Greenwell, 2017), were estimated using bootstrap (100 replications). Multicollinearity can be an issue with BRT modeling when assessing if and when environmental variables are of ecological interest. Thus, prior to modeling, the Pearson correlation analysis between all environmental variables was calculated in order to avoid including highly correlated variables into the model. The correlation analysis showed that most variables were only weakly intercorrelated (r < 0.5).
Results
Seasonal Changes in Abiotic Environmental Conditions
Water temperature and light availability above the seabed followed the same seasonal pattern with higher values measured in July and lower values in winter (Figure 1). The thickest ice cover (26 cm) was recorded in March 2014. During the winter 2014/2015 ice formation alternated repeatedly with melting. Winter months were characterized by the highest water velocity and nitrogen concentration. The phosphorus content in the water showed no clear seasonal pattern with the highest values observed in spring and December. August stood out among other summer months with the highest nutrient concentration and variability of water movement.
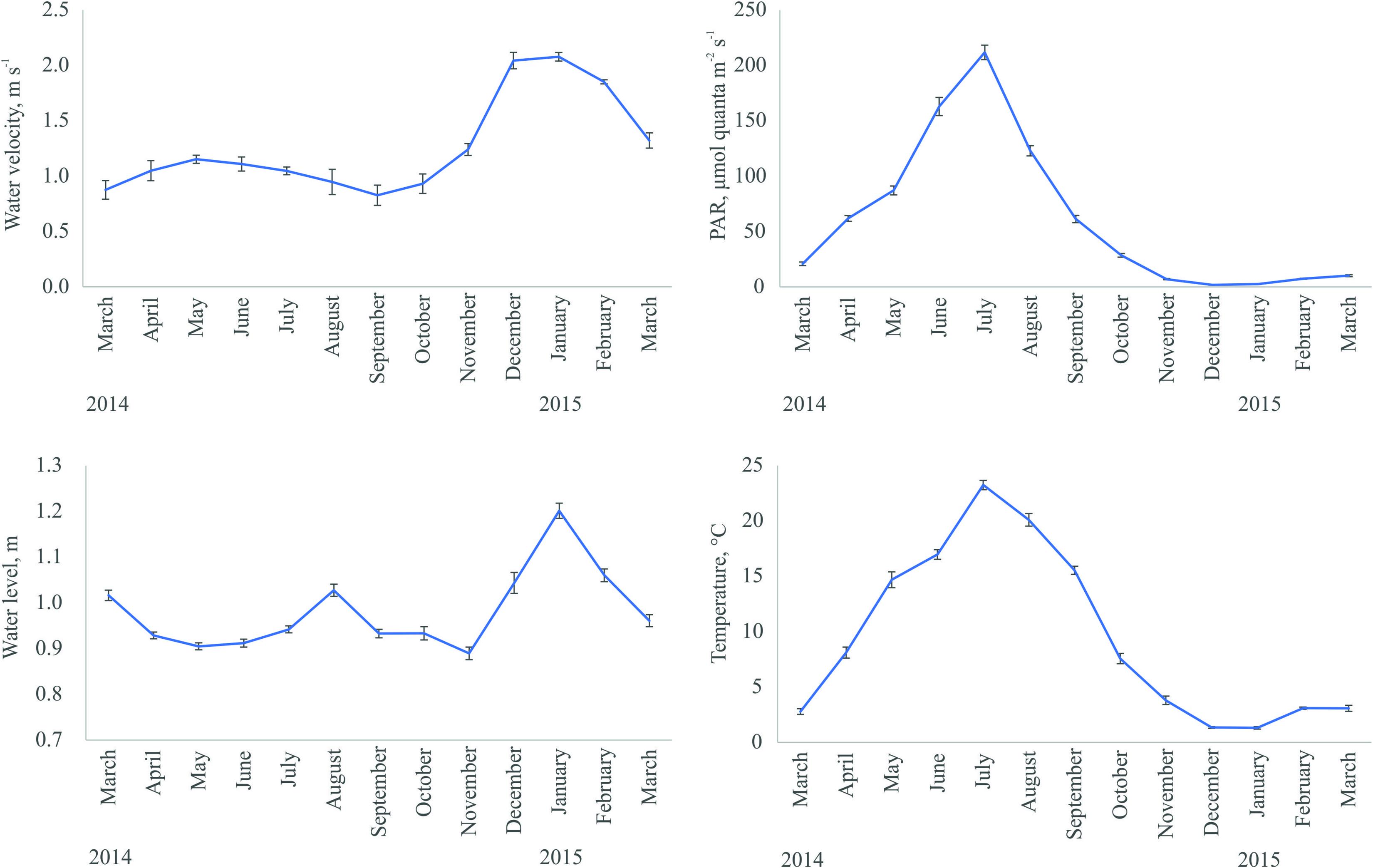
Figure 1. Seasonal dynamics of abiotic environmental variables at near-bottom layer in the study area (means ± Standard Error).
Seasonal Changes in the Community of C. aspera
Charophytes had high biomass in summer and autumn, whereas the algal cover was stably high throughout the year including winter (Figure 2). C. aspera overwintered as a sparse and flat biomass residue. Small fresh shoots were observed only in April and the full community established by July. The rapid decay in biomass occurred during the formation of ice cover. Nutrient content (nitrogen and phosphorus) of C. aspera was high in decomposing algal material in winter and low in summer. The chlorophyll a content of C. aspera plants varied between months with no clear seasonal patterns (somewhat lower values in winter and higher values in summer).
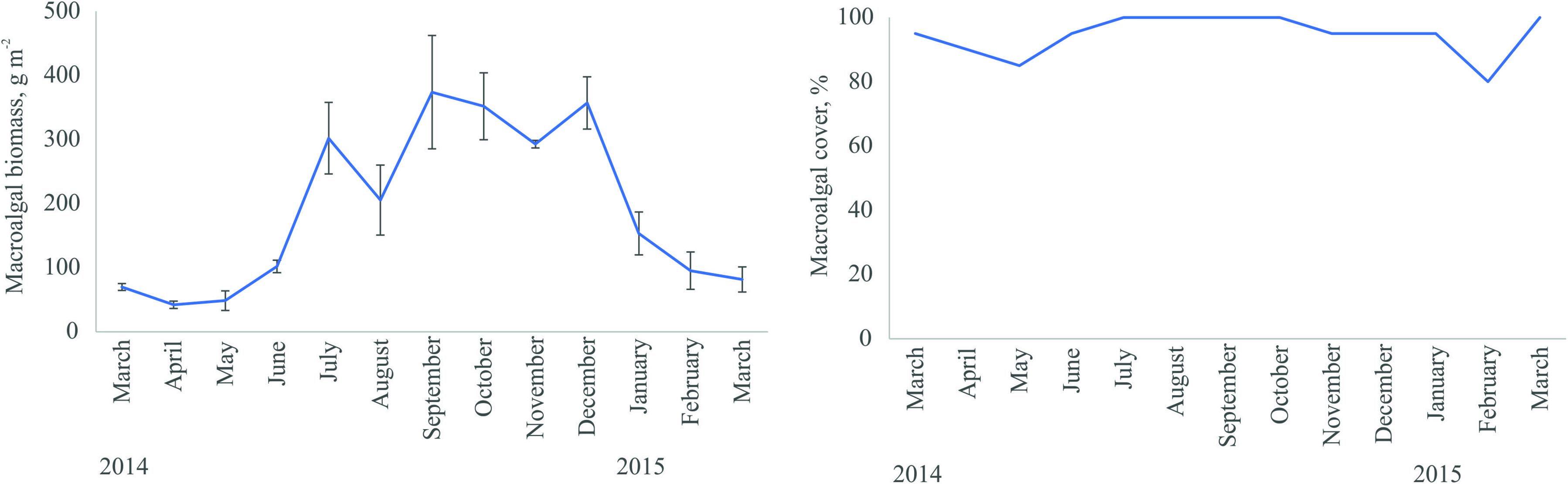
Figure 2. Seasonal dynamics in the biomass and cover of C. aspera community in the study area (means ± Standard Error).
Seasonal Changes in the Individual and Community Production of C. aspera
Net production rates of C. aspera measured at an individual and community level followed similar seasonal patterns over the whole study period with maximum values observed in June and July and lowest values during winter months. The only exceptions were abnormally low community production values in May and August related to the presence of high amounts of organic debris on the sediment. Similarly, the biomass growth of C. aspera was high from June to September and low in other months (Figure 3).
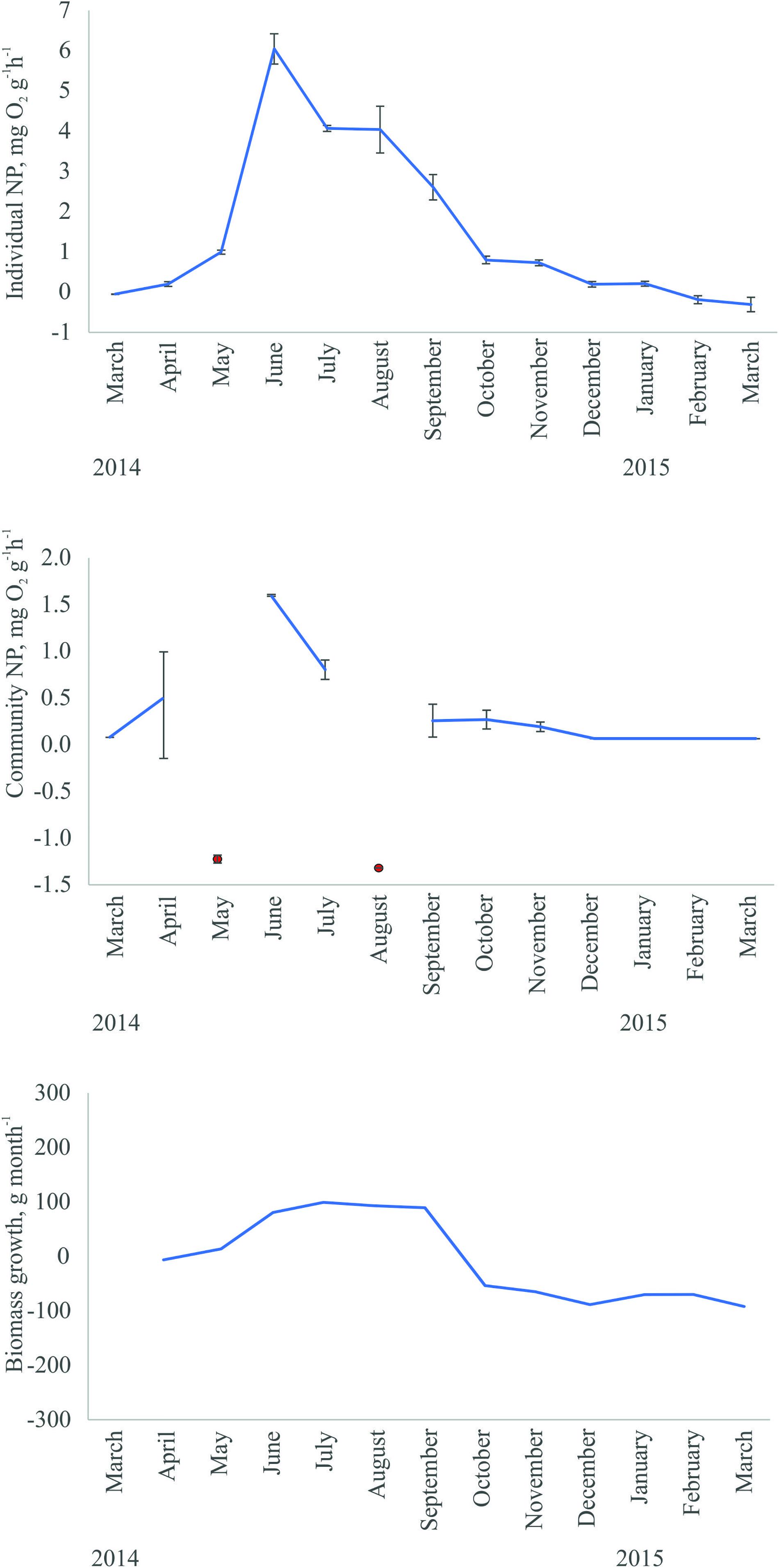
Figure 3. Seasonal dynamics in individual and community net production rates and biomass growth of C. aspera in the study area (means ± Standard Error). Extreme events (negative values) of community net production associated with the presence of high amounts of organic debris on the sediment are shown in red. A moving average smoother was used to visualize the trend in biomass growth.
Relationships Between Environment, Individual, Community Production and Biomass Growth of C. aspera
Boosted regression trees models on the production of C. aspera accounted for a significant proportion of the variability at the studied scales with r2 values as follows: individual photosynthesis 0.855, community production 0.854 and change in algal biomass 0.573 (Table 1). Temperature was the best overall predictor in the models of individual photosynthesis and change in algal biomass and the second-best predictor in the model of community production. However, the thresholds of algal production to increasing temperature varied among the studied scales. Light availability was another generic predictor of the production of C. aspera in all models. Like temperature, the complexity of (functional form) relationships between light and production did not change among models and the differences were due mostly to different light thresholds in different models. Among the studied environmental variables, nutrient concentration in water and chloroplast content in algal tissue were not significant and the variables were not retained in the models.
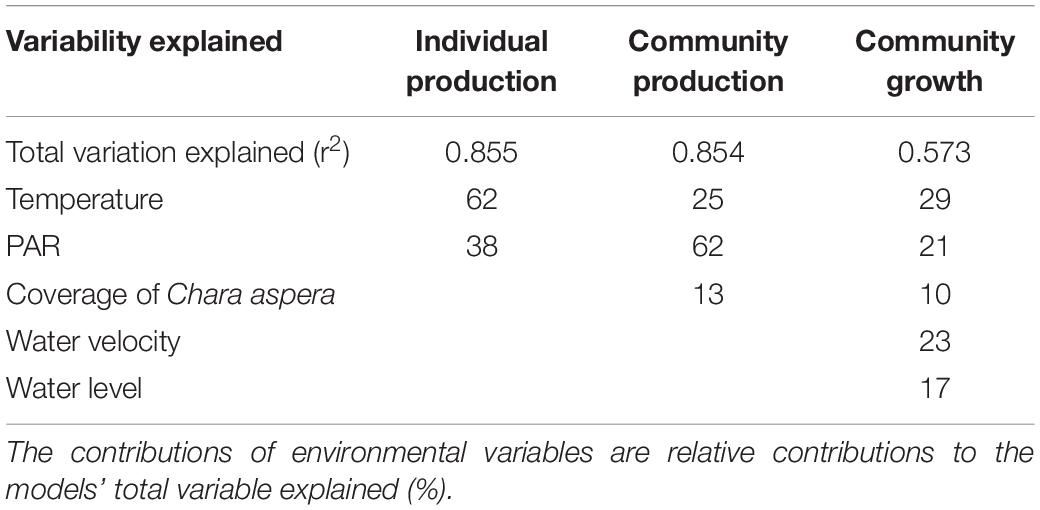
Table 1. Summary statistics of the BRT models on relationships between environment, individual, community production and biomass growth of C. aspera.
Photosynthesis in individual plants was related to temperature and light intensity with these variables accounting for nearly 100% of the predicted variability in the production of C. aspera at this scale. Increasing temperature increased the production of individual plants up to a threshold of 20°C. Above this tipping point, variability in plant production was independent of the ambient water temperature. The production of individual plants had two clear phases: low production at light levels <500 μmol quanta m–2 s–1 and high production at light levels >600 μmol quanta m–2 s–1 with light saturation remaining at 550–600 μmol quanta m–2 s–1 (Figure 4).
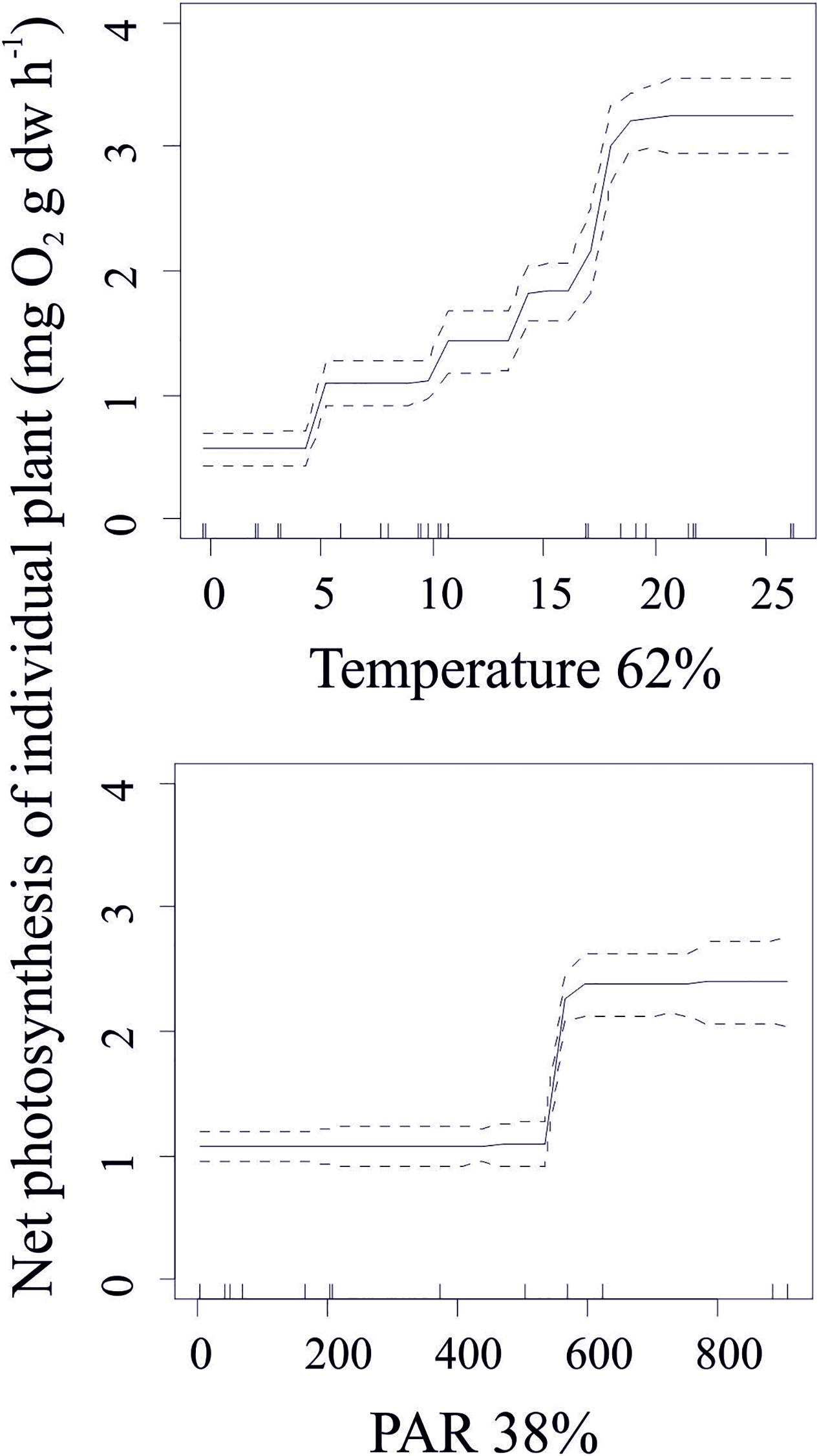
Figure 4. Standardized functional-form relationships (± Standard Error) showing the effect of environmental variables on the net photosynthesis of individual plants of C. aspera, whilst all other variables are held at their means. The variables are ordered by their relative contribution in the BRT model (shown in%). Upward tickmarks on x-axis show the frequency distribution of data along this axis. See the section of methods for further information on environmental variables.
Like photosynthesis in individual plants, the production of benthic communities correlated positively with light intensity. However, the relationship was characterized by a smoother response and a higher light saturation point at 800 μmol quanta m–2 s–1. Importantly, community production decreased when the coverage of C. aspera attained >95%, indicating greater light limitation in denser algal communities. The relationship between temperature and community production exhibited two phases: low production at temperatures <13°C and high production at temperatures >14°C. BRT analysis excluded abnormally low community production values in May and August related to the presence of high amounts of organic debris on the sediment and high levels of belowground respiration. This suggests that when temperature exceeds 21°C, the community net production is expected to decline sharply (Figure 5).
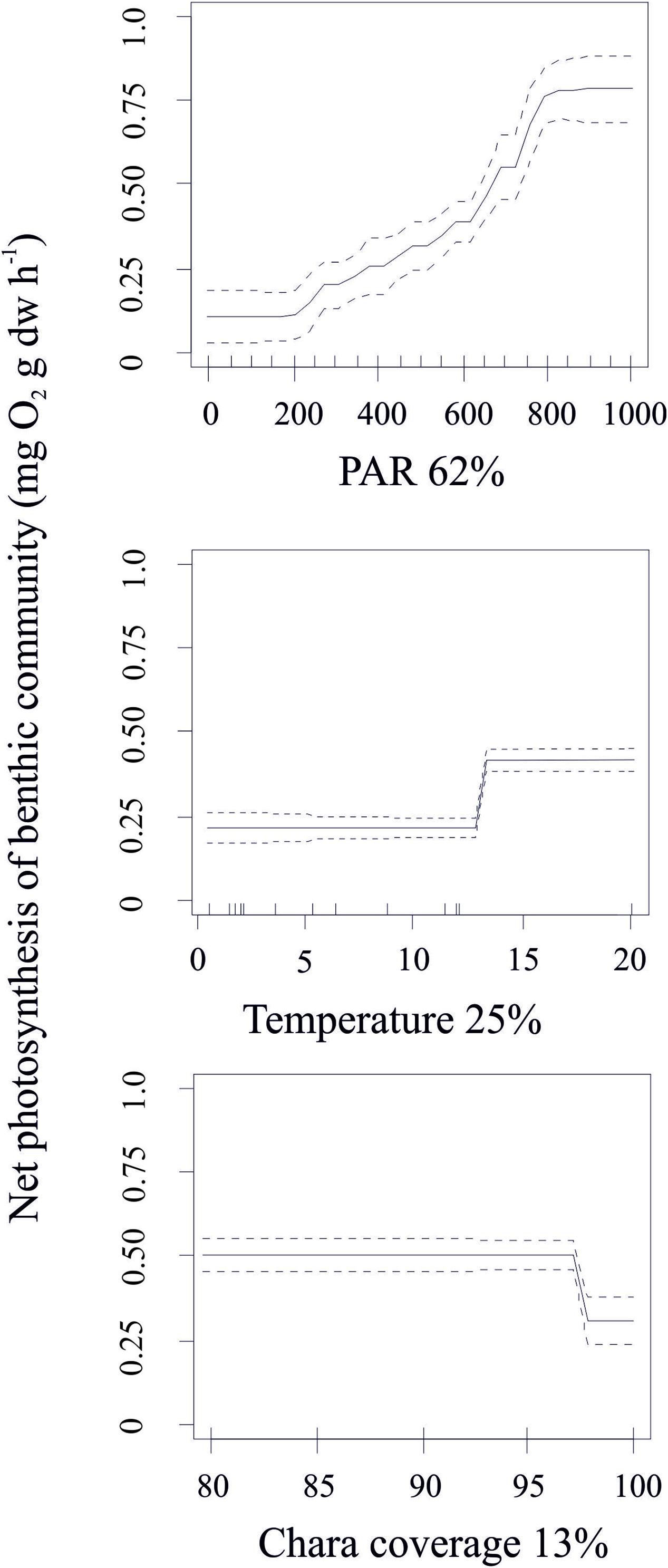
Figure 5. Standardized functional-form relationships (± Standard Error) showing the effect of environmental variables on the community production of C. aspera, whilst all other variables are held at their means. The variables are ordered by their relative contribution in the BRT model (shown in%). Upward tickmarks on x-axis show the frequency distribution of data along this axis. See the section of methods for further information on environmental variables.
Temperature, light climate and underwater currents were the key predictors of change in algal biomass accounting for >70% of the predicted variability in the biomass change. Importantly, high levels of growth were sustained at much lower light intensities, already above 100 μmol quanta m–2 s–1 compared to thresholds of 600 and 800 μmol quanta m–2 s–1 in the models of individual and community production. Similarly, high growth values were predicted only at velocities <1 cm s–1. Moreover, increasing water level was associated with reduced biomass of macroalgal communities (Figure 6).
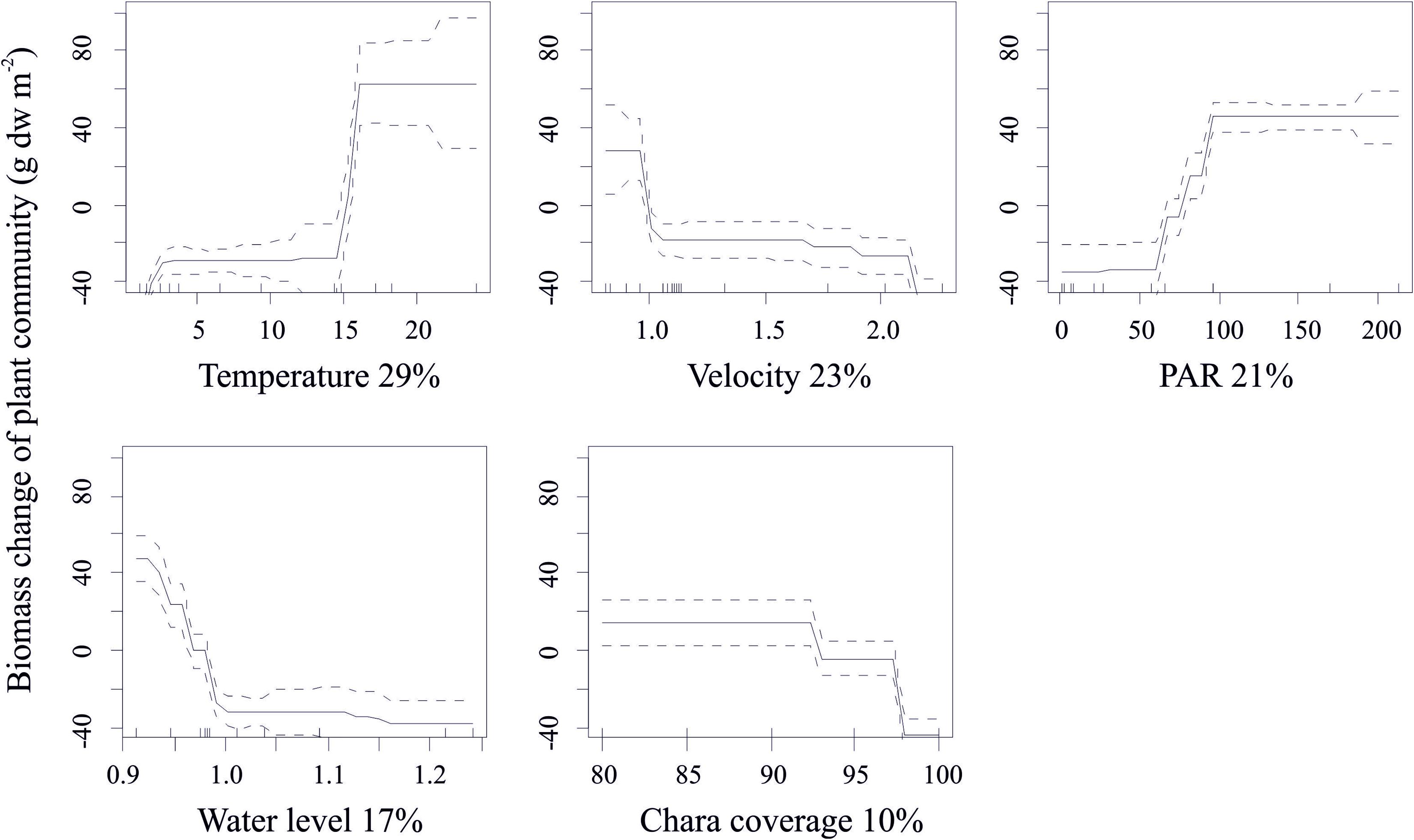
Figure 6. Standardized functional-form relationships (± Standard Error) showing the effect of environmental variables on growth in C. aspera biomass, whilst all other variables are held at their means. The variables are ordered by their relative contribution in the BRT model (shown in%). Upward tickmarks on x-axis show the frequency distribution of data along this axis. See the section of methods for further information on environmental variables.
Discussion
In this study we experimentally quantified seasonal variability in individual photosynthesis, community production and growth in biomass of the charophyte C. aspera in the Baltic Sea along with the key environmental variables that are known to determine the production of macroalgae. We found that temperature and light availability were the best overall predictors of the production of C. aspera, followed by macroalgal coverage, flow velocity and water level. Importantly, the observed scale notably regulated the importance of different predictors, as well as the relationships between the environment and plant responses in the models. This implies that although the physiological responses of C. aspera should always be interpreted in the context of scale, temperature and light availability predict a significant proportion in the variability of charophyte production.
Low light can limit the production potential of temperate macroalgae, particularly during low-light seasons or in turbid waters (Wallentinus, 1978; Littler, 1980); many studies have suggested a direct dependence of charophytes on light availability (Libbert and Walter, 1985; Blindow, 2000; Schneider et al., 2006; Schubert et al., 2018). However, due to high tolerances of C. aspera to low light environments (Blindow, 1992; Sand-Jensen et al., 2000), we hypothesized that the species could maintain positive photosynthesis and growth potential in a wide range of light conditions. In general, this held true for individual photosynthesis, community production and growth in biomass. However, individual photosynthesis was typically higher than community production under the same light conditions. This difference in photosynthesis can be partly attributed to lower light availability (light limitation) in a three-dimensional algal canopy than to individual algae exposed to ambient light levels. For the same reason, the photosynthesis-irradiance curve at community level was smoother than that of individual plants. Specifically, different parts of the algal canopy are expected to receive different amounts of light and therefore are expected to respond differentially. For example, the upper parts may often be oversaturated with light, whereas lower parts are undersaturated. This results in different light adaptation (different light saturation points) along the spatial light gradient and a smooth response of the algal community to light. This also applies for C. aspera, which is known to increase the light saturation point of photosynthesis with increasing irradiance from 100 to 400 μmol quanta m–2 s–1 (Blindow et al., 2003). Similar responses of net photosynthesis along light gradients have been observed for C. aspera in other areas (Kovtun-Kante et al., 2014) and for other charophyte species, such as C. baltica and C. canescens (Küster et al., 2004). Beside shelf-shading, sediment respiration is an additional explanation for the differences between individual and community photosynthesis. At high load of organic debris and/or elevated temperatures algal respiration and sediment decomposition may dominate over benthic production (Carvalho et al., 2005).
Differential responses of photosynthesis and growth to light irradiance is also related to the different approaches used to calculate production at different scales. Specifically, the photosynthesis of individual plants and communities is expressed as relative change per plant biomass, whereas biomass growth is reported on an absolute scale. In spring and early summer with abundant light, photosynthesis of individual plants and communities is enhanced. However, the absolute change in biomass is actually low because the plants themselves are small. Later, when the plants are taller, biomass increases are greater despite moderate light conditions and low photosynthesis of individual plants and communities. This is happening because some inorganic carbon is fixed into autotrophic biomass even at low photosynthesis levels between 100 and 500 μmol quanta m–2 s–1.
In addition to light irradiance reaching the algal canopy, water turbidity affects light availability and the production of charophytes in many habitats (Hellström, 1991; Schneider et al., 2006). Likewise, the sediments in Rame Bay are composed of fine fractions (very fine sand, silt, and clay) and even slow underwater currents (1 cm s–1) can bring silt and mud particles into the water column and thereby modulate the light environment as well as deposit sediment particles on plant thalli. Consequently, less light reaches plant photopigments and the rate of community photosynthesis and biomass growth decreases. This process is supported by experimental evidence in which long exposure to high levels of suspended sediment leads to cessation of growth and ultimately death of charophytes (Henricson et al., 2006). On the other hand, an experimental reduction of light availability to 25% of the natural irradiance does not induce a chronic compensatory reduction of the photosynthesis of charophyte communities (Kovtun-Kante et al., 2014). This suggests that charophytes recover easier from longer-term light reduction than sedimentation and may explain why water velocity (i.e., the intensity of resuspension) was the key predictor in the models of biomass growth. In addition to underwater currents, water level explained a significant part of charophyte growth in biomass. The current charophyte meadows are located in very shallow water (1 m) and a decline in water level of only a few tens of centimeters is expected to improve light conditions in the algal canopy.
It has been suggested that an increased production of photosynthetic pigments and chloroplasts supports more efficient production of charophytes and helps plants to recover when light becomes limiting (Andrews et al., 1984; Küster et al., 2000; Marquardt and Schubert, 2009). Our study did not support this, as variability in photosynthesis and growth along light gradient was independent of the chloroplast content of charophyte tissue. The chloroplast content was measured during fieldwork but due to lack of significance, the variable was not retained in the models and therefore not reported. It has been also suggested that charophytes elongate quickly when light levels are low. This adaptation enables charophytes to reach the water surface where light is more plentiful (Andrews et al., 1984; Henricson et al., 2006). However, the shoot elongation in C. aspera is salinity-dependent and brackish water plants fail to elongate under impoverished light conditions (Blindow et al., 2003). Our study corroborated this as underwater light conditions were not inversely related to plant production throughout the light gradient.
We also hypothesized that C. aspera could maintain positive production and growth potential in a wide range of temperature conditions. This hypothesis held true for individual and community photosynthesis but not for growth in biomass. Interestingly, photosynthesis in individual plants increased with increasing temperature throughout the gradient, whereas this temperature dependence translated into relatively stable positive community photosynthesis with somewhat higher photosynthesis at temperatures >14°C. Nevertheless, at temperatures >22°C benthic production declined substantially due to the dominance of below-ground processes (e.g., sediment decomposition) and algal respiration over algal production (Carvalho et al., 2005; Torn et al., 2006). Thus, C. aspera should sustain positive growth in its Baltic Sea habitats even under relatively extreme temperature conditions, which substantiates earlier studies showing that temperature does not significantly control photosynthesis and growth of charophytes (Raven et al., 1979; Libbert and Walter, 1985; Torn et al., 2006). Due to such broad temperature tolerance, charophytes are expected to benefit from contemporary climate change and broaden their distribution range in the Baltic Sea region (Torn et al., 2020).
Finally, we expected that C. aspera could maintain high photosynthesis and growth potential across wide gradients in water nutrients. This hypothesis was fully supported as nutrient concentrations in water were not significant in all models. Although charophytes are able to acquire nutrients from the water column (Raven, 2003), nutrient concentration in water has been shown to play a marginal effect on the physiological state of charophytes (Blindow, 1988; Munsterhjelm, 2005). Charophytes in our study area seem to rely on nutrients stored in their tissues as shown by the inverse relationship between macroalgal nutrient content and the photosynthesis of charophyte individuals (R2 = 0.15). Such strategy is widely used among long-living macroalgae in order to overcome temporal and/or spatial mismatch between nutrient availability and optimal growth season (Chapman and Craigie, 1977; Lüning, 1990; Raven, 2003).
Prior to the study, we anticipated that models of photosynthesis in individual plants would involve fewer important environmental predictors than the models of photosynthesis and growth of charophyte communities. This prediction also held true. Although water temperature and light irradiance account for most of variability in the photosynthesis of individual plants, this share is less than in models of community photosynthesis and growth. The existence of a weak negative relationship between algal cover and community photosynthesis at the community level suggests that density dependence does play some role in photosynthesis regulation (Binzer and Middelboe, 2005; Tait and Schiel, 2010). Moreover, water properties (water level and flow velocity) were important in regulating charophyte growth. However, as judged by the importance of different predictors in the models, the effects of macroalgal density and water properties were much weaker than light dependence of production and growth of C. aspera.
The availability of different forms of inorganic carbon may limit the photosynthesis of submerged vegetation including C. aspera (van den Berg et al., 2002). Shallow water habitats of our study area are also characterized by a large amplitude of natural variability in pH and pCO2 with a daily range of pH recorded between 8 and 9 (Pajusalu et al., 2013). Among different charophyte communities inhabiting our study area C. aspera is way less sensitive to inorganic carbon limitation compared to C. horrida and C. tomentosa (Pajusalu et al., 2015). Thus, as compared to light and temperature, the limitation of inorganic carbon is likely not a prominent stress to the studied community.
Our study did not focus on interspecific interactions that potentially shape the photosynthesis and growth of C. aspera. A wealth of experimental studies has demonstrated that marine herbivores play key roles in regulating macroalgal communities (Hayward, 1988; Sala and Graham, 2002; Kotta and Witman, 2009). However, previous experiments suggest that herbivory or plant fragmentation induced by herbivores is not crucial in the studied charophyte communities (Kotta et al., 2004). Moreover, C. aspera largely dominates the studied charophyte community and interspecific competition among algae and submerged vegetation is slight. It is plausible that spatial heterogeneity or differential utilization of abiotic resources are behind the pattern in distribution of species (Chambers and Prepas, 1990).
Conclusion
A large set of experimental data is a prerequisite to understand and accurately model the patterns of production of any macroalgal assemblages. In this study we experimentally quantified seasonal variability in individual photosynthesis, community production and growth in biomass of the charophyte C. aspera in the Baltic Sea along with the key environmental variables known to determine the production of macroalgae. The experiments suggested that the production and growth of C. aspera was defined largely by light and partly by temperature at all studied scales. However, algal production was systematically lower at the community than at an individual level under the same light conditions, suggesting the importance of shelf-shading and below-ground processes in natural plant assemblages (e.g., Carvalho et al., 2005; Binzer et al., 2006). Moreover, the observation scale also defined response types between the environment, plant production and growth. This implies that the patterns of variability of the production and growth of macrophytes should always be interpreted in the context of scale and any multiscale model development should involve experimental validation at all important scales.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author Contributions
KT, MR, and JK built the databases. MR and JK analyzed the data. JK, KT, and TP wrote the manuscript. All authors conceived the study, discussed the results, edited the manuscript, and approved the submitted version.
Funding
This study was funded by the Estonian Science Foundation Grant No. 9439 and the Interreg Central Baltic Programme Project MAREA.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Robert Szava-Kovats for proofreading the article.
References
Andrews, M., Box, R., McInroy, S., and Raven, J. A. (1984). Growth of Chara hispida. II. Shade adaptation. J. Ecol. 72, 885–895. doi: 10.2307/2259538
Anthony, K., Ridd, P. V., Orpin, A. R., Larcombe, P., and Lough, J. (2004). Temporal variation of light availability in coastal benthic habitats: effects of clouds, turbidity, and tides. Limnol. Oceanogr. 49, 2201–2211. doi: 10.4319/lo.2004.49.6.2201
Binzer, T., and Middelboe, A. L. (2005). From thallus to communities: scale effects and photosynthetic performance in macroalgae communities. Mar. Ecol. Prog. Ser. 287, 65–75. doi: 10.3354/meps287065
Binzer, T., Sand-Jensen, K., and Middelboe, A. L. (2006). Community photosynthesis of aquatic macrophytes. Limnol. Oceanogr. 51, 2722–2733. doi: 10.4319/lo.2006.51.6.2722
Blindow, I. (1988). Phosphorus toxicity in Chara. Aq. Bot. 32, 393–395. doi: 10.1016/0304-3770(88)90110-6
Blindow, I. (1992). Decline of charophytes during eutrophication – comparison with angiosperms. Freshw. Biol. 28, 9–14. doi: 10.1111/j.1365-2427.1992.tb00557.x
Blindow, I. (2000). Distribution of charophytes along the Swedish coast in relation to salinity and eutrophication. Int. Rev. Hydrobiol. 85, 707–717. doi: 10.1002/1522-2632(200011)85:5/6<707::aid-iroh707>3.0.co;2-w
Blindow, I., Dietrich, J., Möllmann, N., and Schubert, H. (2003). Growth, photosynthesis and fertility of Chara aspera under different light and salinity conditions. Aquat. Bot. 76, 213–234. doi: 10.1016/S0304-3770(03)00053-6
Blindow, I., and van de Weyer, K. (2016). “Ökologie der Characeen,” in Armleuchteralgen: Die Characeen Deutschlands, eds R. Becker, I. Blindow, A. Doege, T. Franke, T. Gregor, U. Hamann, et al. (Berlin: Springer-Verlag), 529–538.
Breeman, A. M., and Pakker, H. (1994). Temperature ecotypes in seaweed – adaptive significance and biogeographic implications. Bot. Mar. 37, 171–180. doi: 10.1515/botm.1994.37.3.171
Buckee, G. K. (1994). Determination of total nitrogen in Barley, Malt and Beer by Kjeldahl procedures and the Dumas combustion method. Collaborative trial. J. Inst. Brew. 100, 57–64. doi: 10.1002/jib.1994.100.2.57
Carvalho, P., Thomaz, S. M., and Bini, L. M. (2005). Effects of temperature on decomposition of a potential nuisance species: the submerged aquatic macrophyte Egeria najas Planchon (Hydrocharitaceae). Braz. J. Biol. 65, 51–60. doi: 10.1590/S1519-69842005000100008
Chambers, P., and Prepas, E. E. (1990). Competition and coexistence in submerged aquatic plant communities: the effects of species interactions versus abiotic factors. Freshw. Biol. 23, 541–550. doi: 10.1111/j.1365-2427.1990.tb00293.x
Chapman, A. R. O., and Craigie, J. S. (1977). Seasonal growth in Laminaria longicruris: relations with dissolved inorganic nutrients and internal reserves of nitrogen. Mar. Biol. 40, 197–205. doi: 10.1007/bf00390875
de los Santos, C. B., Pérez-Lloréns, J. L., and Vergara, J. J. (2000). Photosynthesis and growth in macroalgae: linking functional-form and power-scaling approaches. Mar. Ecol. Prog. Ser. 377, 113–122. doi: 10.3354/meps07844
Elith, J., Graham, C. H., Anderson, R. P., Dudík, M., Ferrier, S., Guisan, A., et al. (2006). Novel methods improve prediction of species’ distributions from occurrence data. Ecography 29, 129–151. doi: 10.1111/j.2006.0906-7590.04596.x
Elith, J., Leathwick, J. R., and Hastie, T. (2008). A working guide to boosted regression trees. J. Anim. Ecol. 77, 802–813. doi: 10.1111/j.1365-2656.2008.01390.x
Field, C. B., Behrenfeld, M. J., Randerson, J. T., and Falkowski, P. (1998). Primary production of the biosphere: integrating terrestrial and oceanic components. Science 281, 237–240. doi: 10.1126/science.281.5374.237
Gattuso, J. P., Gentili, B., Duarte, C. M., Kleypas, J. A., Middelburg, J. J., and Antione, D. (2006). Light availability in the coastal ocean: impact on the distribution of benthic photosynthetic organisms and their contribution to primary productivity. Biogeosciences 3, 489–513. doi: 10.5194/bg-3-489-2006
Gómez, I., Weykam, G., Klöser, H., and Weincke, C. (1997). Photosynthetic light requirements, metabolic carbon balance and zonation of sublittoral macroalgae from King George Island (Antarctica). Mar. Ecol. Prog. Ser. 148, 281–293. doi: 10.3354/meps148281
Greenwell, B. M. (2017). pdp: an R package for constructing partial dependence plots. R. J. 9, 421–436. doi: 10.32614/rj-2017-016
Hargeby, A., Blom, H., Blindow, I., and Andersson, G. (2005). Increased growth and recruitment of piscivorous perch, Perca fluviatilis, during a transient phase of expanding submerged vegetation in a shallow lake. Freshw. Biol. 50, 2053–2062. doi: 10.1111/j.1365-2427.2005.01446.x
Hastie, T., Tibshirani, R., and Friedman, J. (2009). The Elements of Statistical Learning: Data Mining, Inference, and Prediction. New York, NY: Springer.
Hauxwell, J., Cebrián, J., and Valiela, I. (2003). Eelgrass Zostera marina loss in temperate estuaries: relationship to landderived nitrogen loads and effect of light limitation imposed by algae. Mar. Ecol. Prog. Ser. 247, 59–73. doi: 10.3354/meps247059
Hellström, T. (1991). The effect of resuspension on algal production in a shallow lake. Hydrobiology 213, 183–190. doi: 10.1007/BF00016421
Henricson, C., Sandberg-Kilpi, E., and Munsterhjelm, R. (2006). Experimental studies on the impact of turbulence, turbidity and sedimentation on Chara tomentosa L. Cryptogamie Algol. 27, 419–434.
Herkül, K., Torn, K., and Möller, T. (2018). The environmental niche separation between charophytes and angiosperms in the northern Baltic Sea. Bot. Lett. 165, 115–127. doi: 10.1080/23818107.2017.1399824
Hill, R., Bellgrove, A., Macreadie, P. I., Petrou, K., Beardall, J., Steven, A., et al. (2015). Can macroalgae contribute to blue carbon? An Australian perspective. Limnol. Oceanogr. 60, 1689–1706. doi: 10.1002/lno.10128
Hilt, S. (2015). Regime shifts between macrophytes and phytoplankton – concepts beyond shallow lakes, unravelling stabilizing mechanisms and practical consequences. Limnetica 34, 467–480. doi: 10.23818/limn.34.35
Israel, A., and Hophy, M. (2002). Growth, photosynthetic properties and Rubisco activities and amounts of marine macroalgae grown under current and elevated seawater CO2 concentrations. Glob. Change Biol. 8, 831–840. doi: 10.1046/j.1365-2486.2002.00518.x
Jevrejeva, S., and Leppäranta, M. (2002). Ice conditions along the Estonian coast in a statistical view. Nord. Hydrol. 33, 241–262. doi: 10.2166/NH.2002.0025
Kataržytė, M., Vaičiūtė, D., Bučas, M., Gyraitė, G., and Petkuvienė, J. (2017). Microorganisms associated with charophytes under different salinity conditions. Oceanologia 59, 177–186. doi: 10.1016/j.oceano.2016.10.002
Kim, J.-H., Kang, E. J., Edwards, M. S., Lee, K., Jeong, H. J., and Kim, K. Y. (2016). Species-specific responses of temperate macroalgae with different photosynthetic strategies to ocean acidification: a mesocosm study. Algae 31, 243–256. doi: 10.4490/algae.2016.31.8.20
Kirk, J. T. O. (1994). Light and Photosynthesis in Aquatic Ecosystems. Cambridge: Cambridge University Press.
Kotta, J., Paalme, T., Kersen, P., Martin, G., Herkül, K., and Möller, T. (2008). Density dependent growth of the red algae Furcellaria lumbricalis and Coccotylus truncatus in the West-Estonian Archipelago Sea, northern Baltic Sea. Oceanologia 50, 577–585.
Kotta, J., Torn, K., Martin, G., Orav-Kotta, H., and Paalme, T. (2004). Seasonal variation of invertebrate grazing on Chara connivens and C. tomentosa in Kõiguste Bay, NE Baltic Sea. Helgoland Mar. Res. 58, 71–76. doi: 10.1007/s10152-003-0170-2
Kotta, J., and Witman, J. (2009). “Regional-scale patterns,” in Marine Hard Bottom Communities, ed. M. Wahl (Berlin: Springer-Verlag), 89–99. doi: 10.1007/b76710_6
Kovtun-Kante, A., Torn, K., and Kotta, J. (2014). In situ production of charophyte communities under reduced light conditions in a brackish-water ecosystem. Estonian J. Ecol. 63, 28–38. doi: 10.3176/eco.2014.1.03
Kufel, L., and Kufel, I. (2002). Chara beds acting as nutrient sinks in shallow lakes - a review. Aquat. Bot. 72, 249–260. doi: 10.1016/S0304-3770(01)00204-2
Küster, A., Schaible, R., and Schubert, H. (2000). Light acclimation of the charophyte Lamprothamnium papulosum. Aquat. Bot. 68, 205–216. doi: 10.1016/S0304-3770(00)00122-4
Küster, A., Schaible, R., and Schubert, H. (2004). Light acclimation of photosynthesis in three charophyte species. Aquat. Bot. 79, 111–124. doi: 10.1016/j.aquabot.2004.01.010
Leathwick, J. R., Elith, J., Francis, M. P., Hastie, T., and Taylor, P. (2006). Variation in demersal fish species richness in the oceans surrounding New Zealand: an analysis using boosted regression trees. Mar. Ecol. Prog. Ser. 321, 267–281. doi: 10.3354/meps321267
Libbert, E., and Walter, T. (1985). Photosynthetic production of a brackish water community of Chara tomentosa L. and its dependence on environmental conditions. Int. Rev. Ges. Hydrobiol. 70, 359–368. doi: 10.1002/iroh.19850700305
Littler, M. M. (1980). Morphological form and photosynthetic performances of marine macroalgae: tests of a functional/form hypothesis. Bot. Mar. 22, 161–165.
Littler, M. M., and Littler, D. S. (1980). The evolution of thallus form and survival strategies in benthic marine macroalgae: field and laboratory tests of a functional form model. Am. Nat. 116, 25–44. doi: 10.1086/283610
Lobban, C. S., Harrison, P. J., and Duncan, M. J. (1985). The Physiological Ecology of Seaweeds. New York: Cambridge University Press.
Marquardt, R., and Schubert, H. (2009). Photosynthetic characterisation of Chara vulgaris in bioremediation ponds. Charophytes 2, 1–8.
Middleboe, A. L., and Binzer, T. (2004). Importance of canopy structure on photosynthesis in single- and multi-species assemblages of marine macroalgae. Oikos 107, 422–432. doi: 10.1111/j.0030-1299.2004.13345.x
Miller, R. O. (1998). “Nitric-perchloric acid wet digestion in an open vessel,” in Handbook of Reference Methods for Plant Analysis, ed. Y. P. Kalra (Boca Raton, FL: CRC Press), 57–61.
Munsterhjelm, R. (2005). Natural Succession and Human-Induced Changes in the Soft-Bottom Macrovegetation of Shallow Brackish Bays on the Southern Coast of Finland. Helsinki: Walter and Andree de Nottbeck Foundation. PhD Thesis.
Nejrup, L. B., Staehr, P. A., and Thomsen, M. S. (2013). Temperature- and light-dependent growth and metabolism of the invasive red algae Gracilaria vermiculophylla – a comparison with two native macroalgae. Eur. J. Phycol. 48, 295–308. doi: 10.1080/09670262.2013.830778
Nõges, P., Tuvikene, L., Feldmann, T., Tõnno, I., Künnap, H., Luup, H., et al. (2003). The role of charophytes in increasing water transparency: a case study of two shallow lakes in Estonia. Hydrobiol. 506, 567–573. doi: 10.1023/B:HYDR.0000008625.36438.75
Paalme, T., Kotta, J., and Kersen, P. (2013). Does the growth rate of drifting Furcellaria lumbricalis and Coccotylus truncatus depend on their proportion and density. Proc. Estonian Acad. Sci. 62, 141–147. doi: 10.3176/proc.2013.2.08
Pajusalu, L., Martin, G., Põllumäe, A., and Paalme, T. (2013). Results of laboratory and field experiments of the direct effect of increasing CO2 on net primary production of macroalgal species in the brackish water ecosystems. Proc. Estonian Acad. Sci. 62, 148–154. doi: 10.3176/proc.2013.2.09
Pajusalu, L., Martin, G., Põllumäe, A., Torn, K., and Paalme, T. (2015). Direct effects of increased CO2 concentrations in seawater on the net primary production of charophytes in a shallow, coastal, brackish-water ecosystem. Boreal Env. Res. 20, 413–422.
Pärnoja, M., Kotta, J., Orav-Kotta, H., and Paalme, T. (2014). Comparisons of individual and community photosynthetic production indicate light limitation in the shallow water macroalgal communities of the Northern Baltic Sea. Mar. Ecol. 35, 19–27. doi: 10.1111/maec.12074
Puche, E., Rojo, C., Ramos-Jiliberto, R., and Rodrigo, M. A. (2020). Structure and vulnerability of the multi-interaction network in macrophyte-dominated lakes. Oikos 129, 35–48. doi: 10.1111/oik.06694
Raven, J. A. (2003). Long-distance transport in non-vascular plants. Plant Cell Environ. 26, 73–85. doi: 10.1046/j.1365-3040.2003.00920.x
Raven, J. A., and Hurd, C. L. (2012). Ecophysiology of photosynthesis in macroalgae. Photosynth. Res. 113, 105–125. doi: 10.1007/s11120-012-9768-z
Raven, J. A., and Smith, F. A. (1978). Effect of temperature on ion content, ion fluxes and energy metabolism in Chara corallina. Plant Cell Environ. 1, 231–238. doi: 10.1111/j.1365-3040.1978.tb00766.x
Raven, J. A., Smith, F. A., and Glidewell, S. M. (1979). Photosynthetic capacities and biological strategies of giant-celled and small-celled macro-algae. New Phytol. 83, 299–309. doi: 10.1111/j.1469-8137.1979.tb07455.x
Reed, D. C., and Foster, M. S. (1984). The effects of canopy shadings on algal recruitment and growth in a giant kelp forest. Ecology 65, 937–948. doi: 10.2307/1938066
Sala, E., and Graham, M. H. (2002). Community-wide distribution of predator-prey interaction strength in kelp forests. Proc. Natl. Acad. Sci. U.S.A. 99, 3678–3683. doi: 10.1073/pnas.052028499
Sand-Jensen, K., Riis, T., Vestergaard, O., and Larsen, S. E. (2000). Macrophyte decline in Danish lakes and streams over the past 100 years. J. Ecol. 88, 1030–1040. doi: 10.1046/j.1365-2745.2000.00519.x
Schmieder, K., Werner, S., and Bauer, H.-G. (2006). Submersed macrophytes as a food source for wintering waterbirds at Lake Constance. Aquat. Bot. 84, 245–250. doi: 10.1016/j.aquabot.2005.09.006
Schneider, S., Ziegler, C., and Melzer, A. (2006). Growth towards light as an adaptation to high light conditions in Chara branches. New Phytol. 172, 83–91. doi: 10.1111/j.1469-8137.2006.01812.x
Schubert, H., and Blindow, I. (2003). Charophytes of the Baltic Sea. Köningstein: A. R. G. Ganter Verlag Kommanditgesellschaft.
Schubert, H., Blindow, I., Bueno, N. C., Casanova, M. T., Pełechaty, M., and Pukacz, A. (2018). Ecology of charophytes – permanent pioneers and ecosystem engineers. Perspect. Phycol. 5, 61–74. doi: 10.1127/pip/2018/0080
Suursaar, Ü, Otsmann, M., Astok, V., and Kullas, T. (1998). Modelling the water exchange in the Irben, Suur, Soela and Hari Straits in 1995-1996. EMI Rep. Ser. 9, 61–90.
Tait, L. W., and Schiel, D. R. (2010). Primary productivity of intertidal macroalgal assemblages: comparison of laboratory and in situ photorespirometry. Mar. Ecol. Prog. Ser. 416, 115–125. doi: 10.3354/meps08781
Torn, K., and Martin, G. (2003). Changes in the distribution of charophyte species in enclosed seabays of western Estonia. Proc. Estonian Acad. Sci. Biol. Ecol. 52, 134–140.
Torn, K., Martin, G., Kotta, J., and Kupp, M. (2010). Effects of different types of mechanical disturbances on a charophyte dominated macrophyte community. Estuar. Coast. Shelf Sci. 87, 27–32. doi: 10.1016/j.ecss.2009.12.006
Torn, K., Martin, G., and Paalme, T. (2006). Seasonal changes in biomass, elongation growth and primary production rate of Chara tomentosa in the NE Baltic Sea. Ann. Bot. Fenn. 43, 276–283.
Torn, K., Peterson, A., and Herkül, K. (2020). Predicting the impact of climate change on the distribution of the key habitat-forming species in the NE Baltic Sea. J. Coast. Res. SI 95, 177–181. doi: 10.2112/SI95-035.1
van den Berg, M. S., Coops, H., Simons, J., and Pilon, J. (2002). A comparative study of the use of inorganic carbon resources by Chara aspera and Potamogeton pectinatus. Aquat. Bot. 72, 219–233. doi: 10.1016/s0304-3770(01)00202-9
Vermeer, C. P., Escher, M., Portielje, R., and de Klein, J. J. M. (2003). Nitrogen uptake and translocation by Chara. Aquat. Bot. 76, 245–258. doi: 10.1016/s0304-3770(03)00056-1
Keywords: benthic ecology, macroalgae, photosynthesis, primary production, environmental forcing, seasonal variability
Citation: Kotta J, Torn K, Paalme T, Rätsep M, Kaljurand K, Teeveer M and Kotta I (2021) Scale-Specific Patterns of the Production of the Charophyte Chara aspera in the Brackish Baltic Sea: Linking Individual and Community Production and Biomass Growth. Front. Mar. Sci. 8:674014. doi: 10.3389/fmars.2021.674014
Received: 28 February 2021; Accepted: 13 April 2021;
Published: 30 April 2021.
Edited by:
Iñigo Muxika, Technological Center Expert in Marine and Food Innovation (AZTI), SpainReviewed by:
Joao M. Neto, University of Coimbra, PortugalIrmgard Blindow, University of Greifswald, Germany
Copyright © 2021 Kotta, Torn, Paalme, Rätsep, Kaljurand, Teeveer and Kotta. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jonne Kotta, am9ubmVAc2VhLmVl