 Fannie W. Shabangu1,2*
Fannie W. Shabangu1,2* Katie A. Kowarski3
Katie A. Kowarski3- 1Fisheries Management Branch, Department of Forestry, Fisheries and the Environment, Cape Town, South Africa
- 2Mammal Research Institute Whale Unit, Department of Zoology and Entomology, University of Pretoria, Hatfield, Pretoria, South Africa
- 3JASCO Applied Sciences, Dartmouth, NS, Canada
Little is known of the movements and seasonal occurrence of humpback whales (Megaptera novaeangliae) of South Africa and the Antarctic, populations once brought to near extinction by historic commercial whaling. We investigated the seasonal occurrence and diel-vocalizing pattern of humpback whale songs off the west coast of South Africa (migration route and opportunistic feeding ground) and the Maud Rise, Antarctica (feeding ground), using passive acoustic monitoring data collected between early 2014 and early 2017. Data were collected using acoustic autonomous recorders deployed 200-300 m below the sea surface in waters 855, 1,118 and 4,400 m deep. Acoustic data were manually analyzed for humpback whale vocalizations. While non-song calls were never identified, humpback whale songs were detected from June through December in South African waters, with a peak in percentage of acoustic occurrence around September/October in the austral spring. In Antarctic waters, songs were detected from March through May and in July (with a peak occurrence in April) where acoustic occurrence of humpback whales was negatively correlated to distance to the sea ice extent. Humpback whales were more vocally active at night than in the day at all recording sites. Detection range modelling indicates that humpback whale vocalizations could be detected as far as 18 and 45 km from recorders in South African and Antarctic waters, respectively. This study provides a multi-year description of the offshore acoustic occurrence of humpback whales off the west coast of South Africa and Maud Rise, Antarctica, regions that should continue to be monitored to understand these recovering populations.
Introduction
Southern Hemisphere humpback whales (Megaptera novaeangliae) perform impressive seasonal migrations between their austral summer feeding grounds in Antarctica and their winter breeding and calving grounds in low latitudes. The distribution and migration of humpback whales are believed to be driven by food availability, suitable environmental conditions, and reproductive status (Matthews, 1938; Lockyer, 1984; Best et al., 1995; Brown et al., 1995; Pomilla and Rosenbaum, 2005; Purdon et al., 2020; Dey et al., 2021; El-Gabbas et al., 2021; Reisinger et al., 2021; Schall et al., 2021a; Schall et al., 2021b). Off the west coast of South Africa, current knowledge on humpback whale occurrence is based on whaling data (Olsen, 1914; Mackintosh, 1942; Best, 2007; Best and Allison, 2010), sighting data (e.g., Best et al., 1995; Findlay and Best, 1995; Barendse et al., 2010; Barendse et al., 2011; Barendse et al., 2013; Purdon et al., 2020), and one coastal (i.e., adjacent to the coast and shallower than 100 m water depth) acoustic study (Ross-Marsh et al., 2021). In the Southern Ocean, the known seasonal occurrence of humpback whales is based on whaling data (e.g., Mackintosh, 1942), sighting data (e.g., Thiele et al., 2004) and five acoustic studies (Mckay et al., 2004; Van Opzeeland et al., 2013; Schall et al., 2020; Schall et al., 2021a; Schall et al., 2021b). There is a general knowledge gap in spatio-temporal occurrence of humpback whales in both these regions since whaling data is dated, sighting data are mainly limited to spring and summer, and acoustic studies have been limited to inshore waters or ice edge regions to the marginal ice zone. More research is required in both Antarctic and South African waters to improve our knowledge of humpback whale movements and distribution and how the species adapts to ecological variabilities over time. As the species continues to recover from the impacts of historic whaling, this information is critical to understanding their trends in occurrence that can inform management and mitigations for multiple populations.
Humpback whales are vocally active making them ideal for passive acoustic monitoring (PAM). Their vocalizations can be categorized into non-song and song (Payne and McVay, 1971; Dunlop et al., 2007; Dunlop et al., 2008; Cholewiak et al., 2013). Non-songs are produced by males, females, and calves throughout the year and are used for a wide range of purposes including foraging and social interaction (e.g., Stimpert et al., 2007; Parks et al., 2014; Videsen et al., 2017; Fournet et al., 2018). In addition to non-song calls, male humpback whales produce complex rhythmic songs and singing can last for several hours (e.g., Payne and McVay, 1971; Cholewiak et al., 2013). Songs are made up of units, phrases, and themes, and a series of repeated songs makes up a song session (Cholewiak et al., 2013). Whales within the same breeding ground sing a rendition of the same song, and songs change through time and can spread across different regions via social learning (e.g., Dunlop et al., 2007; Garland et al., 2013; Garland et al., 2017). Songs are a largely seasonal reproductive display produced prolifically on the breeding grounds, as well as during migration, and on feeding grounds pre- and post-migration (Edds-Walton, 1997; Dunlop and Noad, 2016; Gridley et al., 2018; Kowarski et al., 2021; Ross-Marsh et al., 2021). Song notes can range in frequency from approximately 20 Hz to above 24 kHz (Au et al., 2006; Stimpert et al., 2007; Zoidis et al., 2008; Cholewiak et al., 2013).
Here, we use PAM to monitor the acoustic occurrence of humpback whales over long-periods of time in regions difficult to monitor regularly by visual techniques. PAM was employed in two regions of known ecological significance: the west coast of South Africa and the Maud Rise Seamount (Figure 1). Humpback whales off the west coast of South Africa belong to the Breeding Stock B1 (Best, 2011), Breeding Stock B2 (Rosenbaum et al., 2004; Rosenbaum et al., 2009), and possibly Breeding Stock C (Findlay et al., 2017) that breed in Angola, Congo, Gabon, Gulf of Guinea, Namibia or Mozambique (Best et al., 1995; Best and Allison, 2010; Best, 2011). Whales off the Maud Rise belong to Breeding Stocks B, C and D (Amaral et al., 2016; Schall et al., 2021b). Both regions are positioned within the boundaries of the International Whaling Commission (IWC) Southern Hemisphere Management Area III (Donovan, 1991). The west coast of South Africa (Figure 1) is found in the southern Benguela ecosystem, an area which is characterized by the strong wind-driven upwelling regime of the Benguela Current (Shannon, 2009). Cold, nutrient rich waters of the Benguela ecosystem support the occurrence of high numbers of marine mammals (Best, 2007; Shabangu et al., 2019; Shabangu et al., 2020a; Shabangu and Andrew, 2020; Shabangu et al., 2021; Letsheleha et al., 2022). Humpback whales have been observed off the west coast of South Africa feeding opportunistically on euphausiids (possibly Euphausia lucens), amphipods (Themisto gaudichaudi), copepods and clupeid fish such as sardine (Sardinops sagax) and anchovy (Engraulis encrasicolus) (Olsen, 1914; Matthews, 1938; Best et al., 1995; Findlay et al., 2017). The Maud Rise (Figure 1) is a seamount in the eastern Weddell Sea characterized by topographically induced upwelling of relatively warm (>1°C), nutrient-rich deep water, which enhances phytoplankton blooms and breaks sea ice in winter to form polynyas and initiates sea ice melt in spring (e.g., Comiso and Gordon, 1987; Hellmer, 2007; Gordon, 2009). Due to phytoplankton blooms, large swarms of Antarctic krill Euphausia superba are usually present around the Maud Rise (Everson, 2000). As a result, krill-eating predators, such as seals, blue whales, and humpback whales, return to the Maud Rise annually (e.g., Matthews, 1938; Shabangu et al., 2017; Shabangu and Rogers, 2021).
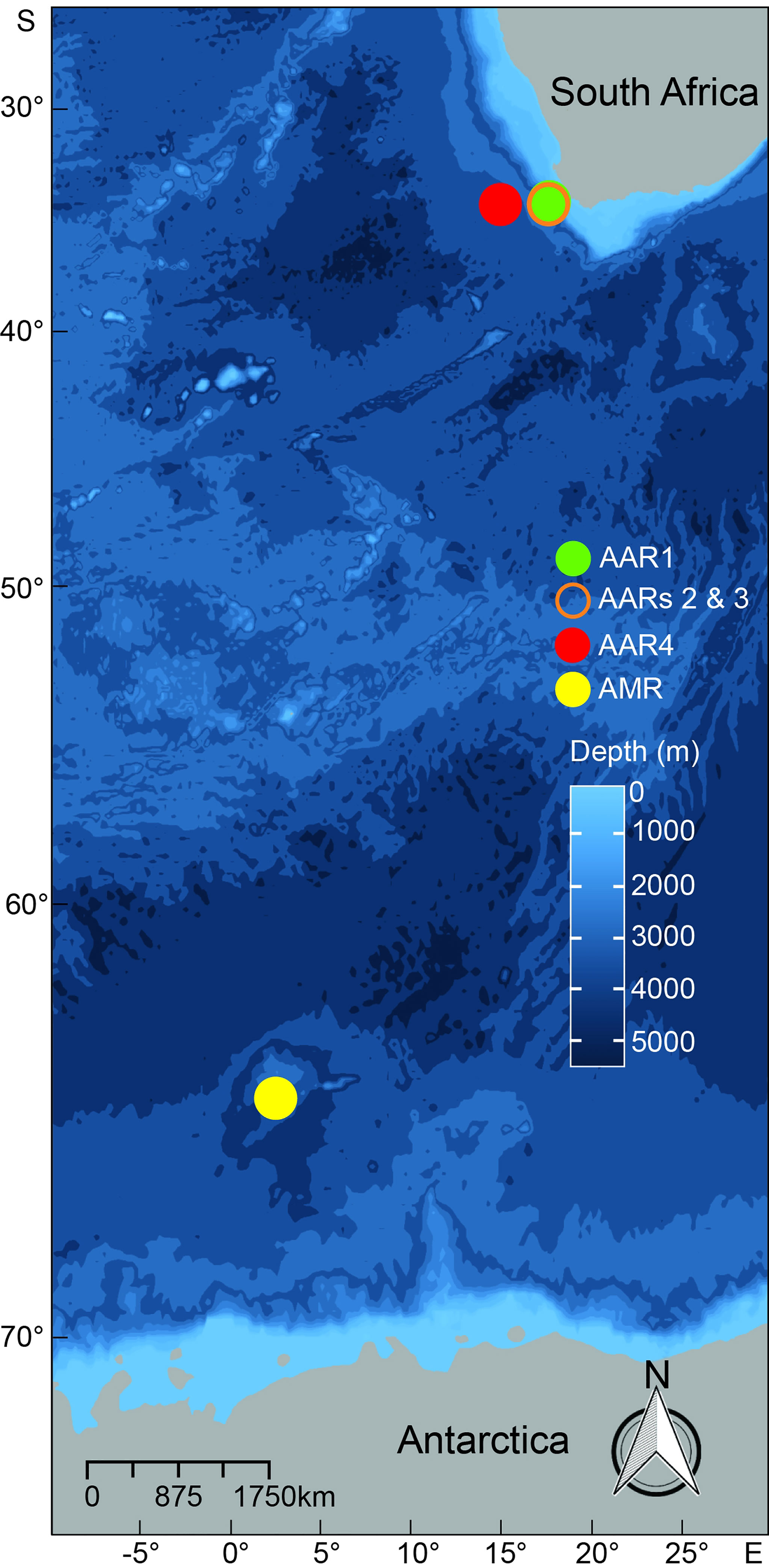
Figure 1 Locations of moorings with autonomous acoustic recorders (AARs) off the west coast of South Africa (AARs 1, 2, 3 and 4) and the Maud Rise, Antarctica (AMR). Shading colour of each circle, as detailed in the key, represents positions of AARs.
No offshore description of the acoustic occurrence of humpback whales exists in South African waters, and likewise, humpback whale occurrence has not previously been studied off the Maud Rise, Antarctica. We aim to fill this knowledge gap by studying PAM data collected between 2014 and 2017. Estimates of detection ranges of humpback whale vocalizations are provided for the recording sites based on acoustic propagation modelling. Results of this study are placed within the context of other baleen whale acoustic occurrence previously described from these same recordings. This information on seasonal occurrence and behavior of humpback whales can inform the assessment of the ecology and the appropriate management of breeding stocks in Antarctic and South African waters.
Materials and Methods
Data Collection
Acoustic recordings were collected between early 2014 and early 2017 from four PAM stations off the west coast of South Africa, and the Maud Rise, Antarctica (Figure 1) as part of the South African Blue Whale Project (SABWP; Shabangu et al., 2019). We used autonomous acoustic recorders (AARs) of Autonomous Underwater Recorder for Acoustic Listening-Model 2 version 04.1.3 (Multi-Électronique Inc., Canada). AARs were deployed on oceanographic moorings (Shabangu et al., 2019) in three locations off the west coast of South Africa, and on an acoustic mooring off the Maud Rise, Antarctica (Figure 1). Settings and sampling protocols for each AAR are detailed in Table 1.
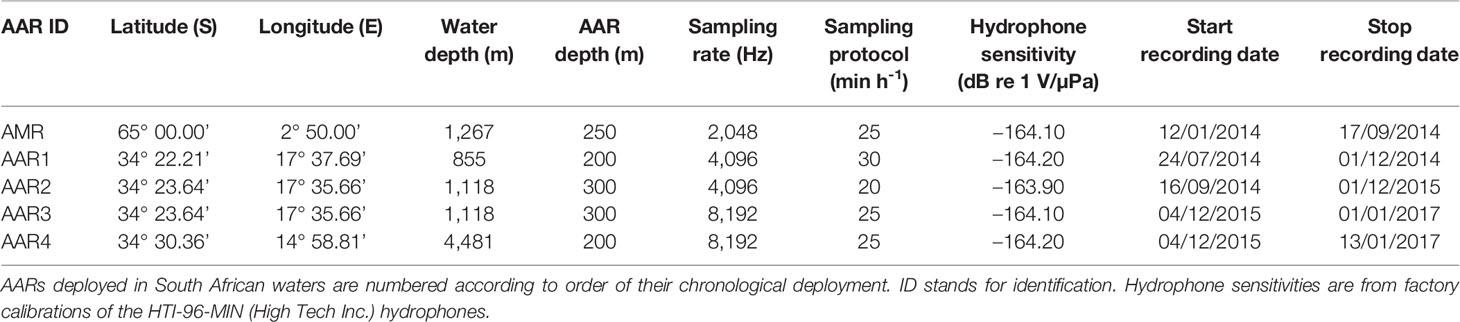
Table 1 Summary of deployment details and recording settings of the five autonomous acoustic recorders (AARs) used in this study.
For diel analysis, hourly sun altitude data were required. Since all AARs off the west coast of South Africa were on the same latitudinal position, we used hourly sun altitudes for each day of the year from 34° 22’ S, 17° 37’ E for all AAR locations. For the Maud Rise, we used the exact location of the seamount (65° 00’ S, 2° 50’ E). We obtained data on sun altitudes from the United States Naval Observatory Astronomical Applications Department (http://aa.usno.navy.mil). Daylight regimes (daytime, night-time, and twilight) were classified according to the geometrical position of the centre of the sun relative to the horizon as detailed in Shabangu et al. (2020a) and Shabangu and Andrew (2020).
For Antarctica, monthly sea ice extensions were sourced from the data set ID: G02135 (Fetterer et al., 2016) at the National Snow and Ice Data Centre data pool server: ftp://sidads.colorado.edu/DATASETS/NOAA/G02135/shapefiles/. The distance of the nearest sea ice edge to the AAR off the Maud Rise (AMR) mooring position was measured from the monthly sea ice extents to determine the effects of sea ice on the acoustic occurrence of humpback whales off the Maud Rise. Daily sea ice concentrations (%) were obtained from the satellite sea ice concentration product of the Advanced Microwave Scanning Radiometer-2 with a 3.1 km grid resolution (Spreen et al., 2008; Beitsch et al., 2014).
Humpback Whale Acoustic Occurrence Analysis
All acoustic files were reviewed by an experienced acoustic analyst (FWS) to the Nyquist frequency of each AAR recordings (1,024 Hz for AMR; 2,048 Hz for AARs 1 and 2; 4,096 Hz for AARs 3 and 4). Acoustic data were inspected via spectrogram analysis in Raven Pro (Center for Conservation Bioacoustics, 2019). The aim of acoustic analysis was to identify humpback whale vocalizations and subsequently categorized them into songs and non-songs using guidelines from published literature (e.g., Payne and McVay, 1971; Stimpert et al., 2012; Cholewiak et al., 2013; Kowarski et al., 2019). Song complexity, in terms of level of hierarchy and repetition of phrases, occurs on a spectrum ranging from the repetition of phrases and sub-phrases over minutes to five or more themes repeated in sequence for hours. To manage this spectrum of complexity, Kowarski et al. (2019) broadly divided songs into full songs, where at least three themes are repeated, and song fragments, which can be as simple as a repeated phrase. Such differentiation of full songs and song fragments can be challenging when working with duty-cycled data or faint acoustic signals and was beyond the scope of the present work. Therefore, the definition of song applied here encompasses both full songs and song fragments where any organization of units into a sequence of phrases or themes was categorized as song. Readers should be mindful of this inclusive definition when interpreting results. Vocalizations not organized into phrases or themes would have been categorized as non-songs, but acoustic signals falling within this definition were never identified. Humpback whale acoustic presence or absence was scored for each acoustic file. The time between which acoustic data were recorded demarcated the sampling session of each AAR (Table 1).
Monthly percentages of humpback whale occurrence for each AAR were calculated as the number of sound files with presence of songs per month divided by the total number of sound files per month. To establish the seasonal diel pattern of humpback whale acoustic occurrence, we divided the number of sampling sessions with humpback whale songs for each hour of the day for that season by the total number of sampling sessions recorded for each hour of the day for that season. Austral seasons were used: summer (December to February), autumn (March to May), winter (June to August), and spring (September to November). Given that time of day is a circular variable, the pattern of diel occurrence of humpback whale songs per season were smoothed through penalized cyclic cubic regression splines (Wood, 2017) in generalized additive models (Guisan et al., 2002). Pearson’s correlation (r) was used to determine the relationship between the percentage of humpback whale occurrence and distance to the sea ice extent.
The SABWP acoustic monitoring program was initiated with the aim to study acoustic occurrence of Antarctic blue whales (Balaenoptera musculus intermedia). Since then, the acoustic occurrence of Antarctic minke (B. bonaerensis), fin (B. physalus), southern right (Eubalaena australis), and sperm (Physeter macrocephalus) whales have been described (Shabangu et al., 2019; Shabangu and Andrew, 2020; Shabangu et al., 2020b; Shabangu et al., 2021; Letsheleha et al., 2022). Humpback whale acoustic occurrence results are interpreted using the current knowledge on both humpback whales and the previously analyzed species in the monitoring areas.
Detection Range Estimation
The objective of detection range estimation was to determine the distance at which humpback whale songs were likely to be detected by our recorders, allowing us to better interpret any variation in occurrence results across recording sites. We used Detection Range Modelling (DRM) software (JASCO) previously described by Warren et al. (2021) and Delarue et al. (In Press) and summarized here. DRM uses the following inputs to model the detection range of humpback whales at different sites: recorded ambient noise, bathymetry, sound speed profiles, geoacoustic properties, and characteristics of humpback whale vocal behaviour. Ambient sound levels were input in the form of sound pressure levels averaged over one min in each decidecade band of the recorded data (International Organization for Standardization, 2017; Ainslie et al., 2018; Martin et al., 2019). Bathymetry information for the modelled sites was collected from the SRTM15+ grid (Smith and Sandwell, 1997; Becker et al., 2009). Sound speed profiles were derived from the U.S. Naval Oceanographic Office’s Generalized Digital Environmental Model V 3.0 (Teague et al., 1990; Carnes, 2009). Geoacoustic properties for the modelled sites were classified as volcanic rock for the Maud Rise (Huang and Jokat, 2016) and sandy mud for the west coast of South Africa (Birch, 1977). For humpback whale vocalization parameters, we input information on songs as that was the only type of acoustic signal identified during manual analysis (as opposed to non-song calls which have different source levels). Song source levels were modelled at 171.5 dB re 1 μPa2 (standard deviation of 5.7; Girola et al., 2019). We modelled from 100-1,000 Hz to capture the predominant frequencies in which humpback whale songs occur and assumed the vocalizing source depth to be between 10 and 30 m. We modelled out to a 50 km distance of the recorders, with the option of extending further if results indicated the full detection range was not captured within 50 km. DRM used the aforementioned parameters and the Marine Operations Noise Model (Matthews and MacGillivray, 2013) to estimate the probability of acoustically detecting humpback whales under different noise conditions.
DRM was performed on AAR2, AAR4, and AMR for the month of May. AARs 1, 2, and 3 were not more than 5 km apart (Figure 1), so these recorders would be expected to have a similar detection range and AAR2 was selected to represent them. One month (May) was selected because 1) it was an achievable timeframe over which modelling could be performed, 2) South Africa does not undergo dramatic seasonal fluctuations, thus a single month is likely representative of the entire year in terms of sound propagation, 3) for Antarctica, May is one of the few months humpback whales were confirmed present and 4) May was a time when there was little interference from ocean current-induced mooring noise across the sites. Low frequency mooring noise was a particular issue for AAR1 and AAR4, with the greatest impact under 100 Hz and spanning to 500 Hz (Shabangu et al., in preparation). Manual analysis of humpback whale songs was still effective above 100 Hz and analysts could make out signals through the noise, therefore, because humpback whale vocalization detection was undergone manually (rather than an automated method) we are confident that mooring induced noise did not significantly impact our humpback whale occurrence results, but it was more of a concern for DRM and therefore mooring noise should be considered when interpreting DRM results.
Results
Seasonal Acoustic Occurrence
A total of 1,567 hours were recorded from AAR1, 3,490 hours from AAR2, 3,982 hours from AAR3, and 4,096 hours from AAR4, which yielded a total of 13,135 hours of acoustic recordings from the South African waters (Table 2). A total of 2,479 hours of acoustic data were recorded off the Maud Rise, Antarctica (Table 2). All detected humpback whale vocalizations were classified as songs (Figure 2) from the 15,614 hours of data analyzed. Non-song calls were never confidently identified, though these signals may have been missed amongst the songs, particularly when there were multiple singers. Off the Maud Rise, the highest number of days and hours with humpback whale songs present were in autumn with few detections in winter (Table 2). In contrast, off the west coast of South Africa, the number of days and hours with detections were high in spring and low in winter with few detections in summer and no detections in autumn (Table 2). With an average of 25% of hours with humpback whale acoustic presence across months, AAR2 had the highest percentage of hours with acoustic occurrence; AARs 3 and 4 had the second highest percentage of hours (20%) with humpback whales while AAR1 had the lowest percentage of hours (15%) with detections (Table 2).
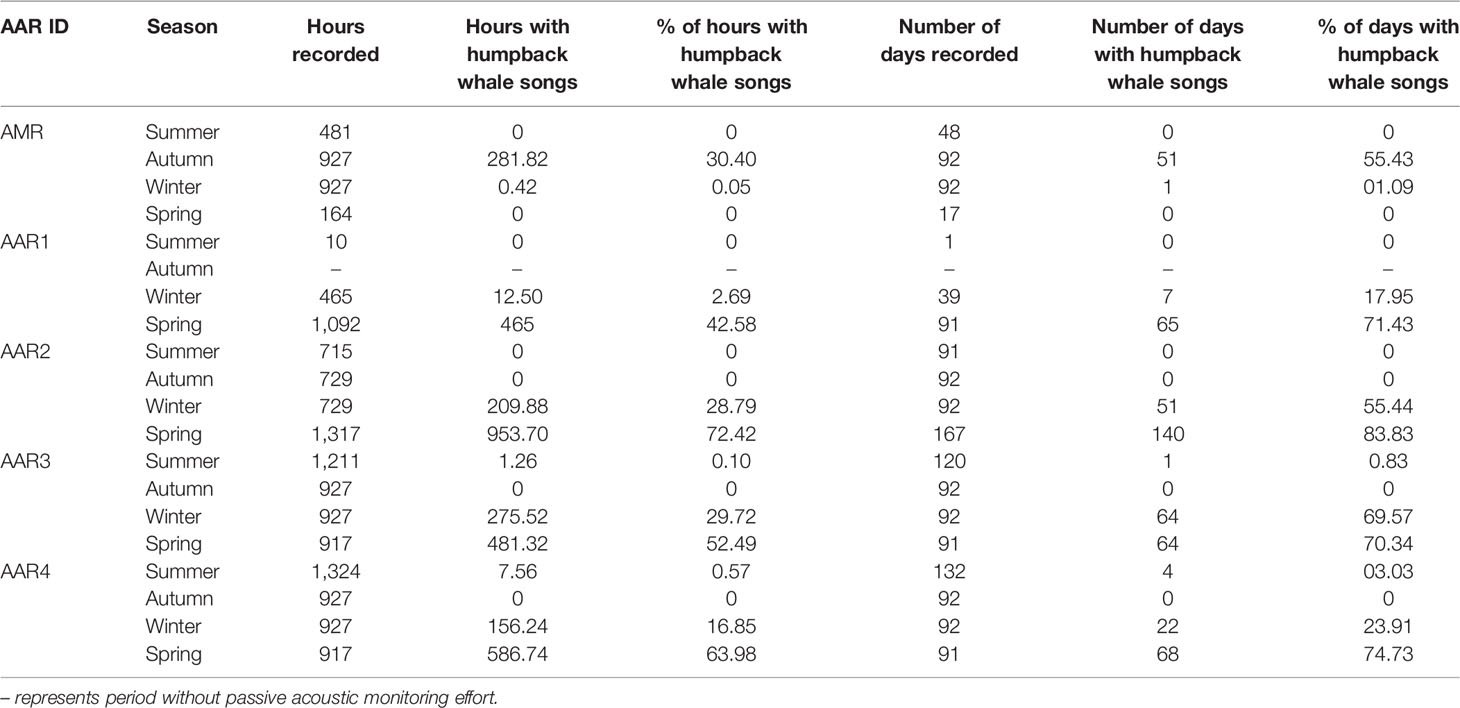
Table 2 Seasonal number and percentage of hours and days containing male humpback whale songs off the west coast of South Africa and the Maud Rise, Antarctica.
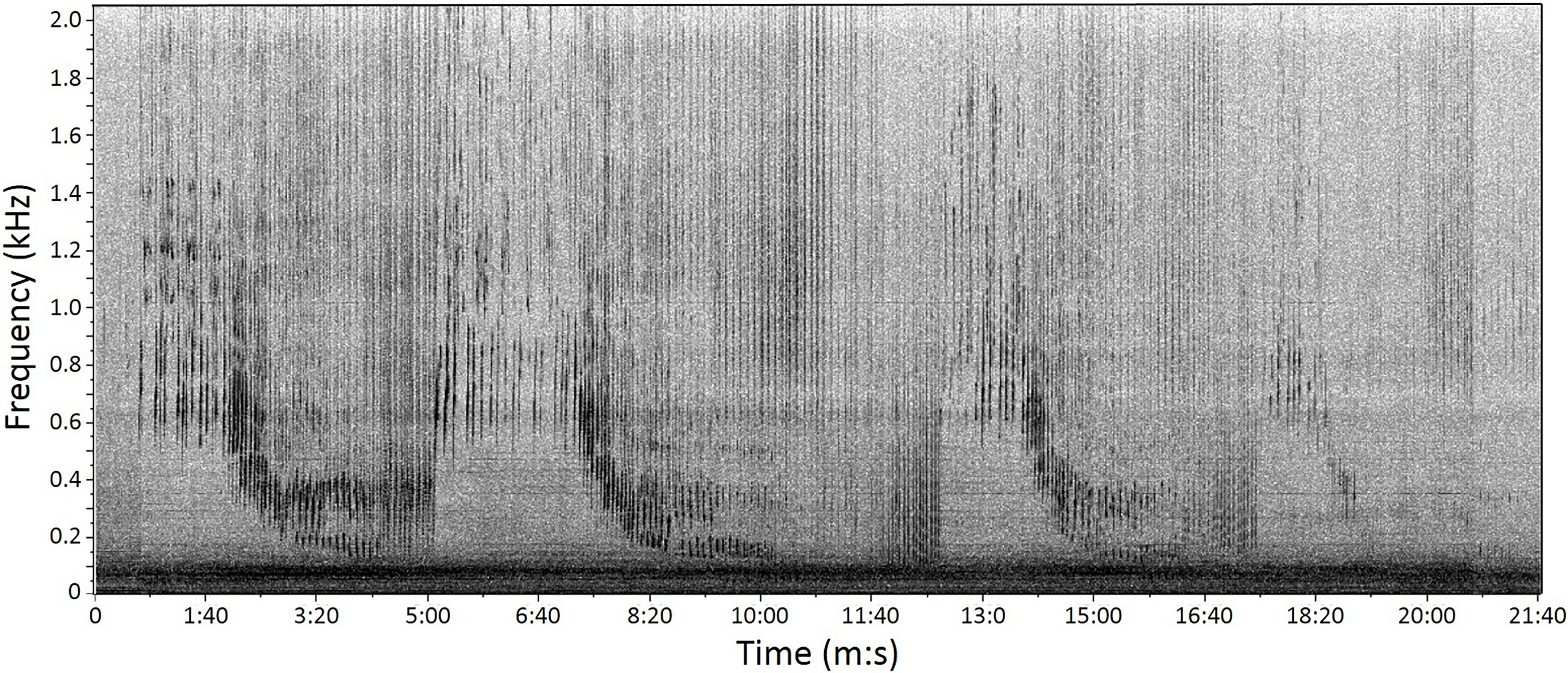
Figure 2 Example of humpback whale song recorded off the west coast of South Africa from AAR2 on 7 July 2015. Spectrogram parameters: frame size 2.32 s, 90% overlap, discrete Fourier transform size 16,384 samples.
Percentage of humpback whale acoustic occurrence from AMR had a single peak of 71.40% in April, and the lowest percentage of occurrence was 0.13% in July (Figure 3). Sea ice concentration was 0% within 3.1 km of the AAR position from 12 January to 20 April 2014 but increased to 50% by the end of April. The AMR mooring position was submerged under sea ice from the beginning of May through mid-September when the recorder stopped working, with sea ice concentrations around 80% at beginning of May and 100% by mid-May through mid-September. Humpback whale songs were first detected in March and continued to be detected even when the AAR mooring position was submerged under sea ice. Percentage of humpback whale occurrence had a weak negative correlation (Pearson’s r= -0.31) with the distance to the sea ice extent, indicating that the acoustic occurrence decreased with increasing distance to the sea ice extent.
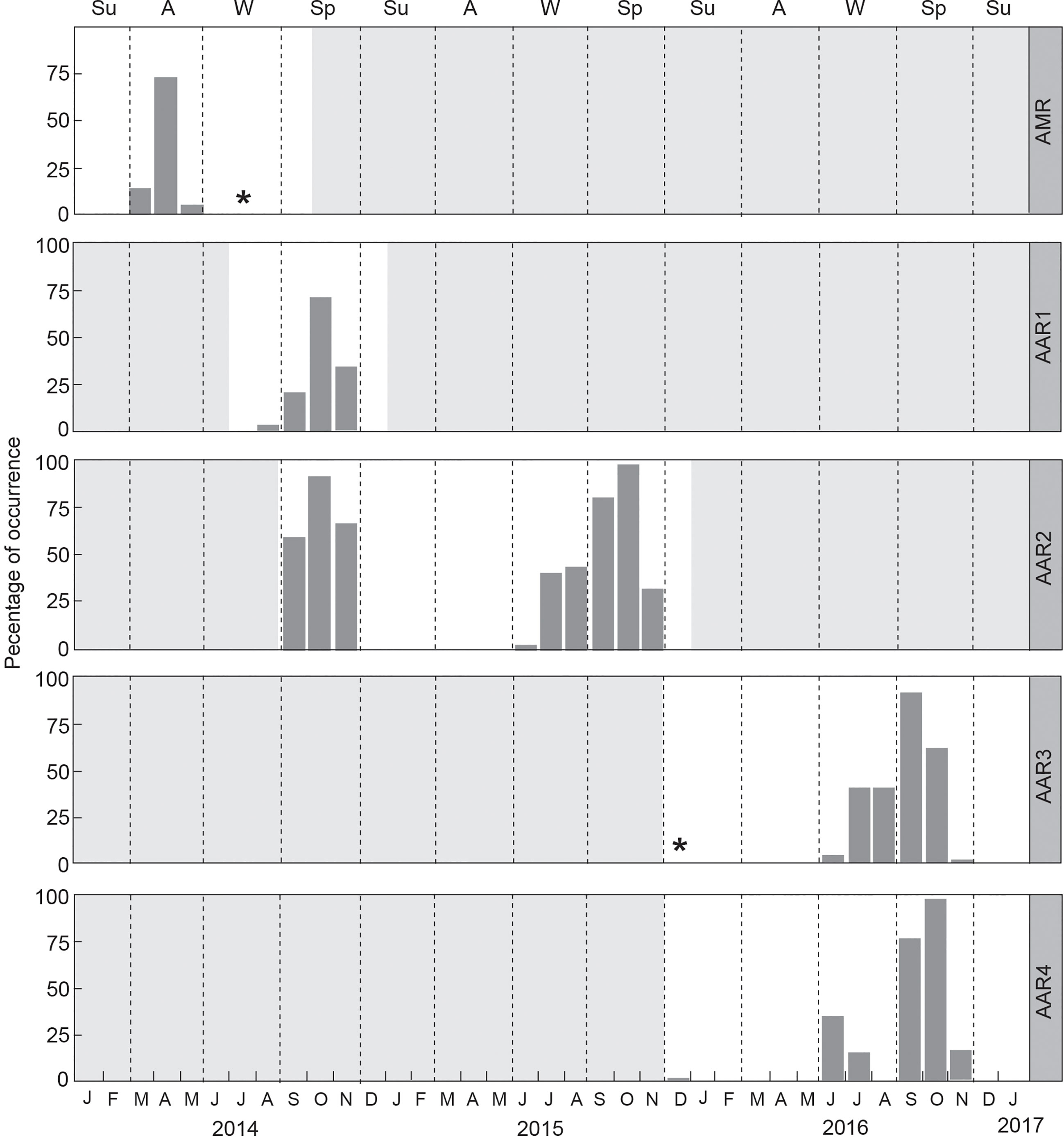
Figure 3 Monthly percentage of acoustic occurrence of male humpback whale songs off the west coast of South Africa and Maud Rise, Antarctica. Asterisks represent cases when the monthly percentage of acoustic occurrence was less than 0.5%. Gray shaded areas designate periods without passive acoustic monitoring effort. Su is summer, A is autumn, W is winter, and Sp is spring are shown on the top axis and defined by dashed lines, and years are stated on the bottom axis.
For AARs 1 and 2, the percentage of humpback whale acoustic occurrence peaked in October and were complementary during periods of recording overlap (from September through November 2014), whereas AAR3 peak in acoustic occurrence was in September 2016 (Figure 3). AAR4 had a bimodal peak of acoustic occurrence in June and October 2016 (Figure 3). Humpback whale songs were detected from late winter through spring in South African waters, and no humpback whale songs were detected from autumn until mid-winter (Figure 3). Songs were detected from August through November for AAR1. For AAR2, songs were detected from September through November in 2014 and from June through November in 2015. Songs were detected in December 2015 and from June through November 2016 for AAR3 (Figure 3). For AAR4, songs were detected in December 2015, between June and July, and from September through November in 2016. The seasonal occurrence of songs did not change inter-annually between AARs 2 and 3 that were deployed on the same location (Figure 3).
Observed Diel-Vocalizing Pattern
Diel percentages of occurrence of humpback whale songs were lowest between 06h00 and 16h00 in winter and spring for South African waters and in autumn for Antarctic waters (Figure 4). Diel percentages of acoustic occurrence were highest between 18h00 and 04h00 for autumn, winter and spring (Figure 4). Due to few hours with songs in summer (Table 3), no diel occurrence pattern was established for all sites (Figure 4). Likewise, no diel occurrence patterns were established for AARs 1-4 in autumn and for AMR in winter and spring due to few hours with songs (Figure 4).
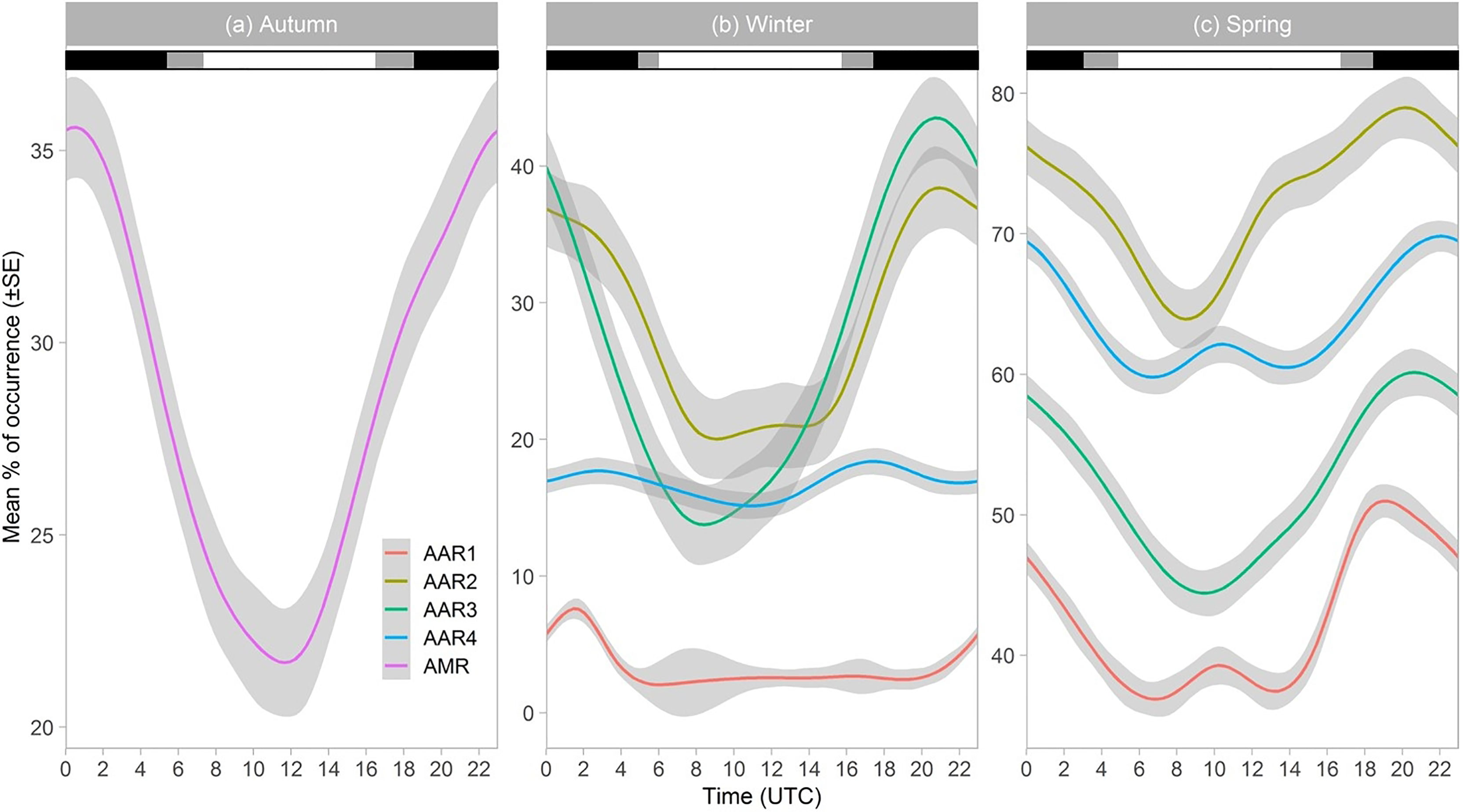
Figure 4 Diel percentage of acoustic occurrence per season for humpback whales off the west coast of South Africa (AARs 1, 2, 3 and 4) and Maud Rise, Antarctica (AMR). Note different y-axes scales. Horizontal bar shadings indicate daylight regime: black represents the average nighttime hours; grey represents average twilight hours; white represents average daytime hours. Grey shading around line plots indicates the standard error (SE). Autumn daylight regime is from the Maud Rise since there was no pattern for South African waters, and remaining daylight regimes are from South African waters since there was no presence data for the Maud Rise. Coordinated Universal Time (UTC) is used.
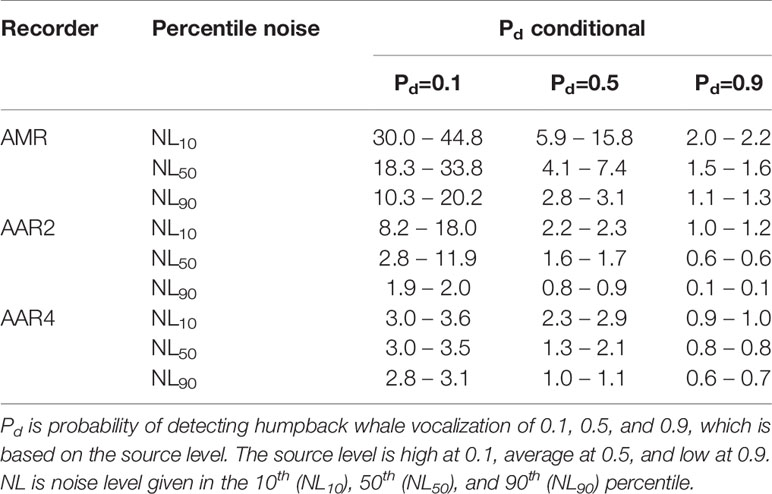
Table 3 Detection ranges (km) of male humpback whale vocalizations in May around AAR2, AAR4 and AMR.
Detection Ranges
Quiet noise conditions (NL10) resulted in longer detection ranges, whereas very noisy conditions (NL90) resulted in shorter detection ranges (Table 3). In average noise conditions (NL50) and detection probability (Pd=0.5), humpback whales were modelled to be detected at a range of approximately 1 to 8 km from the recorder (Table 3). This detection range is likely an underestimate as a portion of the sound levels in the humpback whale vocalization band were high due to mooring noise, but manual analysis of spectrograms allowed for songs to be observed through this noise or in the frequencies above this noise.
Discussion
Here, we provide the offshore seasonal and diel acoustic occurrence of male humpback whales off the west coast of South Africa and the Maud Rise, Antarctica. Humpback whales were acoustically present offshore the west coast of South Africa from June through December and off the Maud Rise from March through May and in July. Given that all acoustic data were analyzed manually, the presence results can be considered a reliable representation of the acoustic occurrence of humpback whales in the data. Though, some considerations should be taken when interpreting the present results.
Detectability
Maximum detection ranges were modelled to be limited for some recording sites, particularly for the more offshore station (AAR4). The short detection ranges are likely a reflection of elevated mooring noise which influenced the modelling outputs. Detection range results indicate that, off South Africa’s west coast, detected humpback whale songs were produced by animals near the recorder and acoustic signals from more distant animals could have been missed. Detections at these sites should therefore be considered a minimum. For the Maud Rise, which had lower ambient and mooring noise, the detection range estimation of 40 km likely captures most vocal animals in the region. Low sampling rate used for AMR, AARs 1 and 2 that yielded recordings with Nyquist frequencies below 2,048 Hz could have, in some cases, negatively biased the detection of some songs and song notes that reside above this frequency. Such instances are expected to be rare given that most humpback whale song notes reside, at least in part, below 2,000 Hz and these lower frequencies would propagate further. Mooring noise below 500 Hz (likely generated by mooring strumming during high ocean current speeds) could have masked some songs below this frequency during strong weather events. The impacts of mooring noise were mitigated by analyzing the data manually, where humans are more sensitive to visually identifying partially masked signals than automated techniques. In addition to the above considerations, it must always be considered that PAM techniques will only capture vocally active animals in the region. We currently do not have a quantitative estimate of the above errors, but these recordings provide a good opportunity to estimate the acoustic occurrence of humpback whales and the overarching trends are certainly accurate.
Offshore South Africa’s West Coast
Acoustic occurrence of humpback whales off the west coast of South Africa was observed from June through December, which is in conjunction with the humpback whale breeding season and seasonal high productivity of the Benguela ecosystem (Best, 2007; Shannon, 2009; Dey et al., 2021). The high humpback whale song detections from early to mid-spring (i.e., September through October) correspond with high sighting rates of the species at this time of the year in coastal waters where whales were observed moving in both southerly and northerly directions (Best et al., 1995; Barendse et al., 2010; Barendse et al., 2011; Barendse et al., 2013; Findlay et al., 2017). Shore based sightings indicate that humpback whales are present year-round off the Saldanha Bay on the west coast of South Africa, designating some form of inshore residency (Barendse et al., 2010; Barendse et al., 2011). Bimodal peaks (June and October) in humpback whale catches were reported from the Hangklip and Dankegrat Whaling Stations off the west coast of South Africa (Best and Allison, 2010). Our acoustic efforts only detected one peak of acoustic occurrence around September/October for most recording sites except for AAR4 that had a small peak in June and a bigger peak in October, suggesting that humpback whales present off the west coast of South Africa around June were less vocally active, were outside the detection ranges of our AARs, or that the whales changed their migration behavior since the end of the whaling era.
The lack of humpback whale songs between January and May from the offshore locations of our South African AARs was somewhat unexpected given recent indications of inshore residency on the west coast of South Africa by Barendse et al. (2011); (2013). The lack of detections could be due to low numbers of whales in the area (Findlay and Best, 1995; Barendse et al., 2011; Barendse et al., 2013), as call numbers (which are used to calculate call rates) of other baleen whales have been found to increase with whale numbers (e.g., Shabangu et al., 2017). Alternatively, whales could have been less frequently vocalizing because it was not the breeding season associated with singing or because the residency observed by Barendse et al. (2011, 2013) was dominated by females. Moreover, whales could have migrated out of the South African waters to higher latitudes if environmental conditions and food availability were limiting.
Where there were recordings at the same location over multiple years (AARs 2 and 3), the seasonal occurrence trends were similar, confirming the region is regularly frequented by this species. Coastal acoustic recordings in False Bay, off the west coast of South Africa, similarly detected humpback whale songs in September and October (Ross-Marsh et al., 2021). The coastal song detections of Ross-Marsh et al. (2021) correspond to the peak acoustic occurrence from AARs 1, 2 and 3; indicating that whales may have passed through the offshore stations on their way to coastal areas or vice versa since these locations are about 70 km apart. Off the Walvis Ridge, Namibia, humpback whale non-songs and songs were detected from May through January (Thomisch et al., 2019), which is comparable to our documented offshore acoustic occurrence of the species.
Humpback whale acoustic occurrence was similar between the closely spaced (5 km apart) AARs 1 and 2 during periods of recording overlap, which is different to that observed for other baleen whales such as Antarctic blue and fin whales (Shabangu et al., 2019) and southern right whales (Shabangu et al., 2021) that have different detections between AARs 1 and 2 even though the recorders were only 5 km apart. The difference in percentage of acoustic occurrence between AARs 1 and 2 for both blue and southern right whales was associated with varying AAR depths relative to the thermocline depth, whale vocalizing depth and noise levels between recording sites (Shabangu et al., 2019; Shabangu et al., 2021; Letsheleha et al., 2022). Humpback whales produce sounds from the sea surface to as deep as 175 m (Stimpert et al., 2012), our recorders deployed at different depths were well positioned to equally record vocalizations of these whales. Thus, the difference in acoustic occurrence of humpback whales from this study and whale species from other studies might be caused by different ecological use of the region and differences in vocalizing depths of different whale species.
In August 2016, humpback whales were not detected at AAR4 but were present 240 km away at AA3. Correspondingly, Antarctic minke and sperm whale acoustic signals were also more common at the more inshore stations and simultaneously detected between AARs 1 and 2 but differently detected between AARs 3 and 4 during periods of overlap (Shabangu et al., 2020a; Shabangu and Andrew, 2020). The difference in humpback whale signal detection between AARs 3 and 4 during the period of overlap is due to the great distance between them (i.e., 240 km) and indicates a preference for the more inshore AAR3 waters. Peak humpback whale acoustic occurrence was in October for AARs 1, 2 and 4, suggesting that most humpback whales arrive in the South African west coast around that month or they sing more often during that month. The peak acoustic occurrence for AAR3 was September (spring), indicating that arrival of whales is slightly early in some years. Barendse et al. (2010) also observed a mid-spring peak of whale occurrence from shore-based observations off Saldanha Bay. Furthermore, difference in peak occurrence between AARs 3 and 4 during time of recording overlap might indicate that whales in the 1,100 and 4,400 m water depths transitioned back and forth between the two sites.
Maud Rise
Off Antarctica, the highest percentage of acoustic occurrence of humpback whales was in April with few detections in May and July, which supports findings of previous studies that showed humpback whales can sometimes inhabit ice-infested areas as they occasionally provide suitable year-round feeding conditions and inhabitable overwintering conditions (e.g., Van Opzeeland et al., 2013; Schall et al., 2020; Schall et al., 2021a; Schall et al., 2021b). The peak occurrence at mid-autumn relates to the high Antarctic krill abundance dominated by large sized zooplankton in late summer (Hewitt et al., 2004), which could indicate that whales were feeding in that region by then. Humpback whales present in Antarctica during winter could be few males that do not migrate to mid- and low latitudes to reproduce (Brown et al., 1995) in efforts to conserve energy associated with migration and reproduction (Lockyer, 1984). Van Opzeeland et al. (2013) detected humpback whale non-songs produced by males and females year-round off the Ekström Ice Shelf, eastern Weddell Sea, and attributed such presence of whales in sea ice-infested waters to food availability and presence of polynyas in that area.
Antarctic blue whales (Shabangu et al., 2020a), Antarctic minke whales (Shabangu et al., 2020b), and crabeater seals (Lobodon carcinophaga) (Shabangu and Charif, 2021) were detected acoustically off the Maud Rise during periods of high sea ice concentration. Humpback whales seems to show a different pattern by having little to no detections when sea ice concentration increased above 80% (hence negative correlation with distance to the sea ice extent), likely to avoid resource competition with Antarctic blue and minke whales that increased their acoustic presence with increasing sea ice concentration (Shabangu et al., 2020a; Shabangu et al., 2020b). Additionally, limited acoustic presence could be due to humpback whales moving on with their migration.
Unfortunately, there was little overlap in recording periods between the Antarctic site and South African sites, making it difficult to interpret movements between the areas. Nonetheless, these acoustic detections provide useful information about acoustic occurrence in these regions. The Maud Rise has been shown to be a useful habitat for other marine mammals such as Antarctic blue and fin whales (Shabangu et al., 2020a), minke whales (Shabangu et al., 2020b), crabeater seals (Shabangu and Charif, 2021), and Ross seals (Ommatophoca rossii) (Shabangu and Rogers, 2021). Thus, the Maud Rise is likely an important summer foraging habitat for humpback whales given their elevated percentage of acoustic occurrence in austral autumn. It is conceivable that songs were produced while feeding (Stimpert et al., 2012; Gridley et al., 2018). Equally, humpback whales have been observed to sing outside of their breeding grounds in the Southern Ocean IWC Management Area V between ~68°S and 166°E (Garland et al., 2013). Furthermore, studies around the world have found humpback whales singing outside of breeding grounds (Au et al., 2000; Parks et al., 2014; Kowarski et al., 2017; Kowarski et al., 2019; Kowarski et al., 2021; Martin et al., 2021), and our results from South Africa and the Maude Rise support the notion that this pattern is true for all migratory humpback whale populations. Humpback whales were observed to sing extensively in IWC Management Area II during the years, 2011-2013, but not in 2015-2016, but again in 2017-2018, most likely in relation to El Niño Southern Oscillation (Schall et al., 2021a). It would therefore be informative to collect more long-term data for the Maud Rise and South Africa to investigate the effects of climate variabilities on humpback whale acoustic ecology.
Diel Trends
The increase in humpback whale song detection during nighttime off the west coast of South Africa and the Maud Rise, Antarctica, suggests that humpbacks are more vocally active at night. Likewise, humpback whale songs were modelled to vocalize more at nighttime in the coastal area of False Bay on the west coast of South Africa (Ross-Marsh et al., 2021). To our best knowledge, the diel pattern of humpback whale songs was previously unknown for the Maud Rise although Schall et al. (2020; 2021a) found no diel vocalizing patterns of humpback whale non-songs and songs from multi-year data for sites close to the Maud Rise. Humpback whales have also been observed elsewhere to vocalize more at night; for example, off Angola (Cerchio et al., 2014) and eastern Canada (Kowarski et al., 2017; Kowarski et al., 2019). Parks et al. (2014) showed that humpback whales tagged with multi-sensor acoustic tags in the Gulf of Maine, western North Atlantic, were more vocally active at nighttime while performing relatively shallower dives. In contrast, bottom dives during daytime were not accompanied by sound production when whales were observed with a conspecific nearby (Parks et al., 2014). Sound production during nighttime dives may allow whales to maintain aural contact with conspecifics when visual contact is limited (Au et al., 2000). Likewise, other baleen whales (Antarctic blue, fin, minke and southern right whales) studied from this data were also more vocally active at night (Shabangu et al., 2019; Shabangu et al., 2020a; Shabangu et al., 2020b; Shabangu et al., 2021; Letsheleha et al., 2022).
Conclusions
We provide the first description of the offshore seasonal occurrence and diel-vocalizing behavior of humpback whales off the west coast of South Africa and the Maud Rise, Antarctica, which both seem to be important regions for this species. High percentages of acoustic occurrence were detected in spring (September/October) off the west coast of South Africa, which corresponds to the breeding season and elevated Benguela Current upwelling activities. Off the Maud Rise, the high percentage of whale acoustic occurrence in mid-autumn (April) matches the high abundance of Antarctic krill at the end of summer. Whales were more vocally active at nighttime in both regions, potentially to maintain acoustic communication with conspecifics when visual contact was not possible. We found the simultaneous acoustic occurrence of humpback whales between two AARs during time of recording overlap to be different to other baleen whale acoustic occurrence previously described from these same recordings given their vocalizing depths and ecological use of the west coast of South Africa. This study contributes essential knowledge towards improving our understanding of the seasonal occurrence and diel-vocalizing behavior of humpback whales in both regions, which is crucial for understanding their occurrence trends which can inform appropriate management of these breeding stocks. Future work should expand upon the present PAM program to observe trends over extended timeframes, understand movements, and identify critical habitats.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Author Contributions
FS conceived the original idea. FS and KAK designed the study. FS processed the acoustic data, performed the statistical data analysis, and lead in writing the manuscript. KAK conducted the sound propagation modelling. KAK provided critical feedback and helped shape the research, analysis and manuscript. All authors contributed to the article and approved the submitted version.
Funding
Project data collection was funded by the South African National Antarctic Programme Grant No. SNA 2011112500003. The funder was not involved in the study analysis, interpretation of data, the writing of this article or the decision to submit it for publication.
Conflict of Interest
KAK is employed by JASCO Applied Sciences (Canada) Ltd.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank the National Research Foundation and South African National Antarctic Programme for funding the SABWP (Grant No. SNA 2011112500003). We are sincerely grateful to Prof Ken Findlay, Meredith Thornton, Marcel van den Berg, Bradley Blows, Chris Wilkinson, numerous colleagues, collaborators, Captains and crew of the RVs SA Agulhas II and Algoa for their assistance with the deployment and recovery of the acoustic instruments used in this study. Thank you to those at JASCO Applied Sciences that contributed to the detection range modelling including Julien Delarue, Marie-Noel Matthews, and Ildar Urazghildiiev. Special thanks to the two reviewers for their thoughtful comments on earlier version of this manuscript.
References
Ainslie M. A., Miksis-Olds J. L., Martin S. B., Heaney K. D., de Jong C. A. F., von Benda-Beckmann A. M., et al. (2018). DEON Underwater Soundscape and Modeling Metadata Standard, Version 1.0. Technical report by JASCO Applied Sciences for ADEON Prime Contract No. M16PC00003 (Halifax: JASCO). doi: 10.6084/m9.figshare.6792359.v2
Amaral A. R., Loo J., Jaris J., Olavarria C., Thiele D., Ensor P., et al. (2016). Population Genetic Structure Among Feeding Aggregations of Humpback Whales in the Southern Ocean. Mar. Biol. 163, 132. doi: 10.1007/s00227-016-2904-0
Au W. W. L., Mobley J., Burgess W. C., Lammers M. O., Nachtigall P. E. (2000). Seasonal and Diurnal Trends of Chorusing Humpback Whales Wintering in Waters Off Western Maui. Mar. Mam. Sci. 16 (3), 530–544. doi: 10.1111/j.1748-7692.2000.tb00949.x
Au W. W. L., Pack A. A., Lammers M. O., Herman L. M., Deakos M. H., Andrews K. (2006). Acoustic Properties of Humpback Whale Songs. J. Acoust. Soc Am. 120, 1103–1110. doi: 10.1121/1.2211547
Barendse J., Best P. B., Carvalho I., Pomilla C. (2013). Mother Knows Best: Occurrence and Associations of Resighted Humpback Whales Suggest Maternally Derived Fidelity to a Southern Hemisphere Coastal Feeding Ground. PloS One 8 (12), e81238. doi: 10.1371/journal.pone.0081238
Barendse J., Best P. B., Thornton M., Elwen S. H., Rosenbaum H. C., Carvalho I., et al. (2011). Transit Station or Destination? Attendance Patterns, Movements and Abundance Estimate of Humpback Whales Off West South Africa From Photographic and Genotypic Matching. Afr. J. Mar. Sci. 33 (3), 353–373. doi: 10.2989/1814232X.2011.637343
Barendse J., Best P. B., Thornton M., Pomilla C., Carvalho I., Rosenbaum H. C. (2010). Migration Redefined? Seasonality, Movements and Group Composition of Humpback WhalesMegaptera Novaeangliae Off the West Coast of South Africa. Afr. J. Mar. Sci. 32 (1), 1–22. doi: 10.2989/18142321003714203
Becker J. J., Sandwell D. T., Smith W. H. F., Braud J., Binder B., Depner J., et al. (2009). Global Bathymetry and Elevation Data at 30 Arc Seconds Resolution: SRTM30_PLUS. Mar. Geod. 32 (4), 355–371. doi: 10.1080/01490410903297766
Beitsch A., Kaleschke L., Kern S. (2014). Investigating High Resolution AMSR2 Sea Ice Concentrations During the February 2013 Fracture Event in the Beaufort Sea. Remote Sens 6, 3841–3856. doi: 10.3390/rs6053841
Best P. B. (2007). Where Is the Breeding Ground for Humpback Whales from Breeding Stock B2? (SC/63/SH17) (Cambridge: International Whaling Commission), p. 18.
Best P. B. (2011). Catch history, seasonal and temporal trends in the migrations of humpback whales along the west coast of southern Africa. (SC/62/SH5). Cambridge: International Whaling Commission, 25.
Best P. B., Allison C. (2010). “Catch History, Seasonal and Temporal Trends in the Migrations of Humpback Whales Along the West Coast of Southern Africa,” in Paper SC/62/SH5 Presented to the IWC Scientific Committee (Agadir, Morocco), p. 25.
Best P. B., Sekiguchi K., Findlay K. P. (1995). A Suspended Migration of Humpback WhalesMegaptera Novaeangliae on the West Coast of South Africa. Mar. Ecol. Prog. Ser. 118, 1–12. doi: 10.3354/meps118001
Birch G. (1977). Surficial Sediments on the Continental Margin Off the West Coast of South Africa. Mar. Geol. 23 (4), 305–337. doi: 10.1016/0025-3227(77)90037-8
Brown M. R., Corkeron P. J., Thale P. T., Schultz K. W., Bryden M. M. (1995). Evidence for a Sex-Segregated Migration in the Humpback Whale (Megaptera Novaeangliae). Proc. R. Soc. B259, 229–234. doi: 10.1098/rspb.1995.0034
Carnes M. R. (2009). Description and Evaluation of GDEM-V 3.0 (Stennis Space Centre: US Naval Research Laboratory), 21 p. Available at: https://apps.dtic.mil/dtic/tr/fulltext/u2/a494306.pdf. MS. NRL Memorandum Report 7330-09-9165.
Center for Conservation Bioacoustics (2019). Raven Pro: Interactive Call Analysis Software (Version 1.6.1) (Ithaca, NY: The Cornell Lab of Ornithology). Available at: http://www.birds.cornell.edu/raven.
Cerchio S., Strindberg S., Collins T., Bennett C., Rosenbaum H. (2014). Seismic Surveys Negatively Affect Humpback Whale Singing Activity Off Northern Angola. PloS One 9 (3), e86464. doi: 10.1371/journal.pone.0086464
Cholewiak D. M., Sousa-Lima R. S., Cerchio S. (2013). Humpback Whale Song Hierarchical Structure: Historical Context and Discussion of Current Classification Issues. Mar. Mamm. Sci. 29, E312–E332. doi: 10.1111/mms.12005
Comiso J. C., Gordon A. L. (1987). Recurring Polynyas Over the Cosmonaut Sea and the Maud Rise. J. Geophys. Res. 92, 2819–2833. doi: 10.1029/JC092iC03p02819
Delarue J. J.-Y., Moors-Murphy H., Kowarski K. A., Davis G. E., Urazghildiiev I. R., Martin S. B. In Press. Co-Acoustic Occurrence of Baleen Whales, Particularly Blue, Fin, and Humpback Whales, Off Eastern Canada 2015–2017. Endanger. Species Res.
Dey S. P., Vichi M., Fearon G., Seyboth E., Findlay K. P., Meynecke J. O., et al. (2021). Oceanographic Anomalies Coinciding With Humpback Whale Super-Group Occurrences in the Southern Benguela. Sci. Rep. 11, 20896. doi: 10.1038/s41598-021-00253-2
Dunlop R. A., Cato D. H., Noad M. J. (2008). Non-Song Acoustic Communication in Migrating Humpback Whales (Megaptera Novaeangliae). Mar. Mamm. Sci. 24, 613–629. doi: 10.1111/j.1748-7692.2008.00208.x
Dunlop R. A., Noad M. J. (2016). The “Risky” Business of Singing: Tactical Use of Song During Joining by Male Humpback Whales. Behav. Ecol. Sociobiol. 70, 2149–2160. doi: 10.1007/s00265-016-2218-8
Dunlop R. A., Noad M. J., Cato D. H., Stokes D. (2007). The Social Vocalization Repertoire of East Australian Migrating Humpback Whales (Megaptera Novaeangliae). J. Acoust. Soc Am. 122 (5), 2893–2905.
Edds-Walton P. L. (1997). Acoustic Communication Signals of Mysticete Whales. Bioacoustics 8, 47–60. doi: 10.1080/09524622.1997.9753353
El-Gabbas A., Van Opzeeland I., Burkhardt E., Boebel O. (2021). Dynamic Species Distribution Models in the Marine Realm: Predicting Year-Round Habitat Suitability of Baleen Whales in the Southern Ocean. Front. Mar. Sci. 8, 2276. doi: 10.3389/fmars.2021.802276
Fetterer F., Knowles K., Meier W., Savoie M. (2016). Sea Ice Index, Version 2 (Boulder, Colorado USA: NSIDC: National Snow and Ice Data Center). doi: 10.7265/N5736NV7
Findlay K. P., Best P. B. (1995). Summer Incidence of Humpback Whales on the West Coast of South Africa. S. Afr. J. Mar. Sci. 15, 279–282. doi: 10.2989/02577619509504851
Findlay K. P., Seakamela S. M., Meÿer M. A., Kirkman S. P., Barendse J., Cade D. E., et al. (2017). Humpback Whale “Super-Groups”- A Novel Low-Latitude Feeding Behaviour of Southern Hemisphere Humpback Whales (Megaptera Novaeangliae) in the Benguela Upwelling System. PloS One 12 (3), e0172002. doi: 10.1371/journal.pone.0172002
Fournet M. E. H., Gabriele C. M., Sharpe F., Straley J. M., Szabo A. (2018). Feeding Calls Produced by Solitary Humpback Whales. Mar. Mamm. Sci. 34 (3), 851–865. doi: 10.1111/mms.12485
Garland E. C., Gedamke J., Rekdahl M. L., Noad M. J., Garrigue C., Gales N. (2013). Humpback Whale Song on the Southern Ocean Feeding Grounds: Implications for Cultural Transmission. PloS One 8 (11), e79422. doi: 10.1371/journal.pone.0079422
Garland E. C., Rendell L., Lamoni L., Poole M. M., Noad M. J. (2017). Song Hybridization Events During Revolutionary Song Change Provide Insights Into Cultural Transmission in Humpback Whales. Proc. Natl. Acad. Sci. U. S. A. 114 (30), 7822–7829. doi: 10.1073/pnas.1621072114
Girola E., Noad M. J., Dunlop R. A., Cato D. H. (2019). Source Levels of Humpback Whales Decrease With Frequency Suggesting an Air-Filled Resonator is Used in Sound Production. J. Acoust. Soc Am. 145 (2), 869–880. doi: 10.1121/1.5090492
Gordon A. L. (2009). “Bottom Water Formation,” in Encyclopedia of Ocean Sciences, 2nd edn, vol. Vol 1 . Eds. Steele J. H., Thorpe S. A., Turekian K. K. (London: Academic Press), 415–421 p.
Gridley T., Silva M., Wilkinson C., Seakamela S., Elwen S. (2018). Song Recorded Near a Super-Group of Humpback Whales on a Mid-Latitude Feeding Ground Off South Africa. J. Acoust. Soc Am. 143 (4), EL298–EL304.
Guisan A., Edwards T. C., Hastie T. (2002). Generalized Linear and Generalized Additive Models in Studies of Species Distributions: Setting the Scene. Ecol. Model. 157, 89–100. doi: 10.1016/S0304-3800(02)00204-1
Hellmer H. (2007). “Oceanography of Weddell Sea,” in Encyclopedia of the Antarctic. Ed. Riffenburgh B. (New York, NY: Routledge), 1053–1056, pp.
Hewitt R. P., Kim S., Naganobu M., Gutierrez M., Kang D., Takao Y., et al. (2004). Variation in the Biomass Density and Demography of Antarctic Krill in the Vicinity of the South Shetland Islands During the 1999/2000 Austral Summer. Deep Sea Res. Part II Top. Stud. Oceanogr 51, 1411–1419.
Huang X., Jokat W. (2016). Sedimentation and Potential Venting on the Rifted Continental Margin of Dronning Maud Land. Mar. Geophys. Res. 37, 313–324.
International Organization for Standardization. (2017). Underwater Acoustics – Measurement of Radiated Underwater Sound From Percussive Pile Driving (Geneva). Available at: https://www.iso.org/obp/ui/#iso:std:iso:18406:ed-1:v1:en. ISO 18406:2017(E).
Kowarski K., Cerchio S., Whitehead H., Moors-Murphy H. (2021). Where, When, and Why do Western North Atlantic Humpback Whales Begin to Sing? Bioacoustics. doi: 10.1080/09524622.2021.1972838
Kowarski K., Evers C., Moors-Murphy H., Martin B., Denes S. L. (2017). Singing Through Winter Nights: Seasonal and Diel Occurrence of Humpback Whale (Megaptera Novaeangliae) Calls in and Around the Gully MPA, Offshore Eastern Canada. Mar. Mamm. Sci. 34 (1), 169–189.
Kowarski K., Moors-Murphy H., Maxner E., Cerchio S. (2019). Western North Atlantic Humpback Whale Fall and Spring Acoustic Repertoire: Insight Into Onset and Cessation of Singing Behavior. J. Acoust. Soc Am. 145 (4), 2305–2316.
Letsheleha I. S., Shabangu F. W., Farrell D., Andrew R. K., la Grange P. L., Findlay K. P. (2022). Year-Round Acoustic Monitoring of Antarctic Blue and in Whales in Relation to Environmental Conditions Off the West Coast of South Africa. Mar. Biol. 169, 41. doi: 10.1007/s00227-022-04026-x
Lockyer C. (1984). Review of Baleen Whale Reproduction and Implications for Management. Rep. Int. Whal. Commn. Spec. Iss. 6, 27–50.
Martin S. C., Aniceto A. S., Ahonen H., Pedersen G., Lindstrøm U. (2021). Humpback Whale (Megaptera Novaeangliae) Song on a Subarctic Feeding Ground. Front. Mar. Sci. 8, 669748. doi: 10.3389/fmars.2021.669748
Martin S. B., Morris C., Bröker K., O'Neill C. (2019). Sound Exposure Level as a Metric for Analyzing and Managing Underwater Soundscapes. J. Acoust. Soc Am. 146 (1), 135–149. doi: 10.1121/1.5113578
Matthews M.-N. R., MacGillivray A. O. (2013). Comparing Modeled and Measured Sound Levels From a Seismic Survey in the Canadian Beaufort Sea. Proc. Meet. Acoust. 19 (1), 1–8. doi: 10.1121/1.4800553
Mckay S., Širović A., Thiele D., Abery N. W., Chester E. T., Hildebrand J., et al. (2004). Combining Acoustic, Sighting Survey and Sea Ice Data to Investigate the Seasonal Presence of Humpback Whales, Megaptera Novaeangliae, in the Western Antarctic Peninsula (SC/56/E26) (Cambridge: International Whaling Commission), 9.
Olsen O. (1914). Whales and Whaling in South Africa (Translated From Norwegian). Bergens Mus Arb. 15 (5), 56.
Parks S., Cusano D., Stimpert A., Weinrich M. T., Friedlaender A. S., Wiley D. N. (2014). Evidence for Acoustic Communication Among Bottom Foraging Humpback Whales. Sci. Rep. 4, 7508. doi: 10.1038/srep07508
Payne R. S., McVay S. (1971). Songs of Humpback Whales. Science 173, 585–597. doi: 10.1126/science.173.3997.585
Pomilla C., Rosenbaum H. C. (2005). Against the Current: An Inter-Oceanic Whale Migration Event. Biol. Lett. 1, 1476–1479. doi: 10.1098/rsbl.2005.0351
Purdon J., Shabangu F. W., Yemane D., Pienaar M., Somers M. J., Findlay K. (2020). Species Distribution Modelling of Bryde’s Whales, Humpback Whales, Southern Right Whales, and Sperm Whales in the Southern African Region to Inform Their Conservation in Expanding Economies. PeerJ 8, e9997.
Reisinger R. R., Friedlaender A. S., Zerbini A. N., Palacios D. M., Andrews-Goff V., Dalla Rosa L., et al. (2021). Combining Regional Habitat Selection Models for Large-Scale Prediction: Circumpolar Habitat Selection of Southern Ocean Humpback Whales. Remote Sens. 13, 13112074. doi: 10.3390/rs13112074
Rosenbaum H. C., Pomilla C., Leslie M., Best P. B., Findlay K. P., Engel M. H., et al. (2004). Mitochondrial DNA Diversity and Population Structure of Humpback Whales From their Wintering Areas in the Indian and South Atlantic Ocean (Breeding Regions A, B, C and X) (SC/56/SH3) (Cambridge: International Whaling Commission), 9.
Rosenbaum H. C., Pomilla C., Mendez M., Leslie M. S., Best P. B., Findlay K. P., et al. (2009). Population Structure of Humpback Whales From Their Breeding Grounds in the South Atlantic and Indian Oceans. PloS One 4 (10), e7318. doi: 10.1371/journal.pone.0007318
Ross-Marsh E. C., Elwen S. H., Prinsloo A. S., James B. S., Gridley T. (2021). Singing in South Africa: Monitoring the Occurrence of Humpback Whale (Megaptera Novaeangliae) Song Near the Western Cape. Bioacoustics 30 (2), 163–179. doi: 10.1080/09524622.2019.1710254
Schall E., Thomisch K., Boebel O., Gerlach G., Spiesecke S., Van Opzeeland I. (2020). Large-Scale Spatial Variabilities in the Humpback Whale Acoustic Presence in the Atlantic Sector of the Southern Ocean. R. Soc Open Sci. 7, 201347. doi: 10.1098/rsos.201347
Schall E., Thomisch K., Boebel O., Gerlach G., Woods S. M., El-Gabbas A., et al. (2021a). Multi-Year Presence of Humpback Whales in the Atlantic Sector of the Southern Ocean But Not During El Niño. Commun. Biol. 4, 790. doi: 10.1038/s42003-021-02332-6
Schall E., Thomisch K., Boebel O., Gerlach G., Woods S. M., Roca I. T., et al. (2021b). Humpback Whale Song Recordings Suggest Common Feeding Ground Occupation by Multiple Populations. Sci. Rep. 11, 18806. doi: 10.1038/s41598-021-98295-z
Shabangu F. W., Andrew R. K. (2020). Clicking Throughout the Year: Sperm Whale Clicks in Relation to Environmental Conditions Off the West Coast of South Africa. Endanger. Species Res. 43, 475–494. doi: 10.3354/esr01089
Shabangu F. W., Andrew R. K., Findlay K. (2021). Acoustic Occurrence, Diel-Vocalizing Pattern and Detection Ranges of Southern Right Whale Gunshot Sounds Off South Africa’s West Coast. Mar. Mamm Sci. 37, 733–750. doi: 10.1111/mms.12760
Shabangu F. W., Andrew R. K., Yemane D., Findlay K. P. (2020a). Acoustic Seasonality, Behaviour and Detection Ranges of Antarctic Blue and Fin Whales Under Different Sea Ice Conditions Off Antarctica. Endang. Species Res. 43, 21–37. doi: 10.3354/esr01050
Shabangu F. W., Charif R. A. (2021). Short Moan Call Reveals Seasonal Occurrence and Diel-Calling Pattern of Crabeater Seals in the Weddell Sea, Antarctica. Bioacoustics 30 (5), 543–563. doi: 10.1080/09524622.2020.1819877
Shabangu F. W., Findlay K., Stafford K. M. (2020b). Seasonal Acoustic Occurrence, Diel-Vocalizing Patterns and Bioduck Call-Type Composition of Antarctic Minke Whales Off the West Coast of South Africa and the Maud Rise, Antarctica. Mar. Mamm. Sci. 36, 658–675. doi: 10.1111/mms.12669
Shabangu F. W., Findlay K. P., Yemane D., Stafford K. M., van den Berg M., Blows B., et al. (2019). Seasonal Occurrence and Diel Calling Behavior of Antarctic Blue Whales and Fin Whales in Relation to Environmental Conditions Off the West Coast of South Africa. J. Mar Syst. 190, 25–39. doi: 10.1016/j.jmarsys.2018.11.002
Shabangu F. W., Rogers T. L. (2021). Summer Circumpolar Acoustic Occurrence and Call Rates of Ross Ommatophoca Rossii, and Leopard, Hydrurga Leptonyx, Seals in the Southern Ocean. Polar Biol. 44, 433–450. doi: 10.1007/s00300-021-02804-9
Shabangu F. W., Yemane D., Stafford K. M., Ensor P., Findlay K. P. (2017). Modelling the Effects of Environmental Conditions on the Acoustic Occurrence and Behavior of Antarctic Blue Whales. PloS One 12 (2), e0172705. doi: 10.1371/journal.pone.0172705
Shannon V. (2009). “Benguela Current,” in Encyclopedia of Ocean Sciences, 2nd edn. Eds. Steele J. H., Thorpe S. A., Turekian K. K. (London: Academic Press), 316–327, p.
Smith W. H. F., Sandwell D. T. (1997). Global Sea Floor Topography From Satellite Altimetry and Ship Depth Soundings. Science 277 (5334), 1956–1962. doi: 10.1126/science.277.5334.1956
Spreen G., Kaleschke L., Heygster G. (2008). Sea Ice Remote Sensing Using AMSR-E 89-GHz Channels. J. Geophys. Res. Oceans 113, C2S03. doi: 10.1029/2005JC003384
Stimpert A. K., Peavey L. E., Friedlaender A. S., Nowacek D. P. (2012). Humpback Whale Song and Foraging Behavior on an Antarctic Feeding Ground. PloS One 7 (12), e51214. doi: 10.1371/journal.pone.0051214
Stimpert A. K., Wiley D. N., Au W. W. L., Johnson M. P., Arsenault R. (2007). “Megapclicks”: Acoustic Click Trains and Buzzes Produced During Nighttime Foraging of Humpback Whales (Megaptera Novaeangliae). Biol. Lett. 3, 467–470. doi: 10.1098/rsbl.2007.0281
Teague W. J., Carronand M. J., Hogan P. J. (1990). A Comparison Between the Generalized Digital Environmental Model and Levitus Climatologies. J. Geophys. Res. 95 (C5), 7167–7183. doi: 10.1029/JC095iC05p07167
Thiele D., Chester E. T., Moore S. E., Širović A., Hildebrand J. A., Friedlaender A. S. (2004). Seasonal Variability in Whale Encounters in the Western Antarctic Peninsula. Deep-Sea Res. II 51, 2311–2325. doi: 10.1016/j.dsr2.2004.07.007
Thomisch K., Boebel O., Bachmann J., Filun D., Neumann S., Spiesecke S., et al. (2019). Temporal Patterns in the Acoustic Presence of Baleen Whale Species in a Presumed Breeding Area Off Namibia. Mar. Ecol. Prog. Ser. 620, 201–214. doi: 10.3354/meps12952
Van Opzeeland I., Van Parijs S., Kindermann L., Burkhardt E., Boebel O. (2013). Calling in the Cold: Pervasive Acoustic Presence of Humpback Whales (Megaptera Novaeangliae) in Antarctic Coastal Waters. PloS One 8 (9), e73007. doi: 10.1371/journal.pone.0073007
Videsen S. K. A., Bejder L., Johnson M., Madsen P. T. (2017). High Suckling Rates and Acoustic Crypsis of Humpback Whale Neonates Maximise Potential for Mother-Calf Energy Transfer. Funct. Ecol. 31, 1561–1573. doi: 10.1111/1365-2435.12871
Warren V. E., Širović A., McPherson C., Goetz K. T., Radford C. A., Constantine R. (2021). Passive Acoustic Monitoring Reveals Spatio-Temporal Distributions of Antarctic and Pygmy Blue Whales Around Central New Zealand. Front. Mar. Sci. 7, 575257. doi: 10.3389/fmars.2020.575257
Wood S. N. (2017). P-Splines With Derivative Based Penalties and Tensor Product Smoothing of Unevenly Distributed Data. Stat. Comput. 27 (4), 985–989. doi: 10.1007/s11222-016-9666-x
Keywords: seasonal occurrence, humpback whales, passive acoustics, diel-vocalizing patterns, Antarctica, South Africa, songs
Citation: Shabangu FW and Kowarski KA (2022) The Beat Goes On: Humpback Whale Song Seasonality in Antarctic and South African Waters. Front. Mar. Sci. 9:827324. doi: 10.3389/fmars.2022.827324
Received: 01 December 2021; Accepted: 21 March 2022;
Published: 26 April 2022.
Edited by:
Ryan Rudolf Reisinger, University of Southampton, United KingdomReviewed by:
Yujiang Hao, Institute of Hydrobiology (CAS), ChinaElena Schall, Alfred Wegener Institute Helmholtz Centre for Polar and Marine Research (AWI), Germany
Copyright © 2022 Shabangu and Kowarski. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Fannie W. Shabangu, ZmFubmllLnNoYWJhbmd1QHlhaG9vLmNvbQ==