 Thibault de Garidel-Thoron1*
Thibault de Garidel-Thoron1* Sonia Chaabane1,2,3
Sonia Chaabane1,2,3 Xavier Giraud1
Xavier Giraud1 Julie Meilland4
Julie Meilland4 Lukas Jonkers4
Lukas Jonkers4 Michal Kucera4
Michal Kucera4 Geert-Jan A. Brummer5
Geert-Jan A. Brummer5 Maria Grigoratou6
Maria Grigoratou6 Fanny M. Monteiro7
Fanny M. Monteiro7 Mattia Greco8
Mattia Greco8 P. Graham Mortyn9
P. Graham Mortyn9 Azumi Kuroyanagi10
Azumi Kuroyanagi10 Hélène Howa11
Hélène Howa11 Gregory Beaugrand12
Gregory Beaugrand12 Ralf Schiebel2
Ralf Schiebel2- 1Aix-Marseille Université, CNRS, IRD, Collège de France, INRAE, CEREGE, Aix-en-Provence, France
- 2Department of Climate Geochemistry, Max Planck Institute for Chemistry, Mainz, Germany
- 3Centre de synthèse et d’analyse sur la biodiversité, Fondation pour la recherche sur la biodiversité (FRB), Montpellier, France
- 4Zentrum für Marine Umweltwissenschaften der Universität Bremen (MARUM), Center for Marine Environmental Sciences, University of Bremen, Bremen, Germany
- 5Department of Ocean Systems, Nederlands Instituut voor Onderzoek der Zee (NIOZ), Royal Netherlands Institute for Sea Research, Texel, Netherlands
- 6Gulf of Maine Research Institute, Portland, ME, United States
- 7Bristol Research Initiative for the Dynamic Global Environment (BRIDGE), School of Geographical Sciences, University of Bristol, Bristol, United Kingdom
- 8Institute of Oceanology, Polish Academy of Sciences, Sopot, Poland
- 9Institut de Ciència i Tecnologia Ambientals (ICTA) and Deparment of Geography, Universitat Autonoma de Barcelona, Barcelona, Spain
- 10Tohoku University Museum, Tohoku University, Sendai, Japan
- 11LPG-BIAF, UMR-CNRS 6112, University of Angers, Angers, France
- 12Université Littoral Côte d’Opale, Univ. Lille, CNRS, UMR 8187, LOG, Laboratoire d’Océanologie et de Géosciences, Wimereux, France
Planktonic Foraminifera are ubiquitous marine protozoa inhabiting the upper ocean. During life, they secrete calcareous shells, which accumulate in marine sediments, providing a geological record of past spatial and temporal changes in their community structure. As a result, they provide the opportunity to analyze both current and historical patterns of species distribution and community turnover in this plankton group on a global scale. The FORCIS project aims to unlock this potential by synthesizing a comprehensive global database of abundance and diversity observations of living planktonic Foraminifera in the upper ocean over more than 100 years starting from 1910. The database will allow for unravelling the impact of multiple global-change stressors acting on planktonic Foraminifera in historical times, using an approach that combines statistical analysis of temporal diversity changes in response to environmental changes with numerical modeling of species response based on their ecological traits.
Introduction
Planktonic organisms are subject to a multitude of stressors including anthropogenic effects. Over the last decades, those changes have affected sizably both their biodiversity and distribution at the community level (Poloczanska et al., 2013; Jonkers et al., 2019), but also altered individual biological processes, such as phenology (Ji et al., 2010), and biomineralization (Maas et al., 2018). Concurrent with ongoing global warming, anthropogenic fossil fuel combustion has globally raised the atmospheric CO2 concentration to levels similar to the Pliocene (> 3 million years ago), causing historically unprecedented decrease in pH in the upper ocean (de la Vega et al., 2020). Moreover, the rise of the sea surface temperature (SST) is intensifying ocean thermal stratification and decreasing the nutrient supply from the deeper water column to fertilize the upper ocean (Houghton et al., 2001; Boyd et al., 2019) (Figure 1).
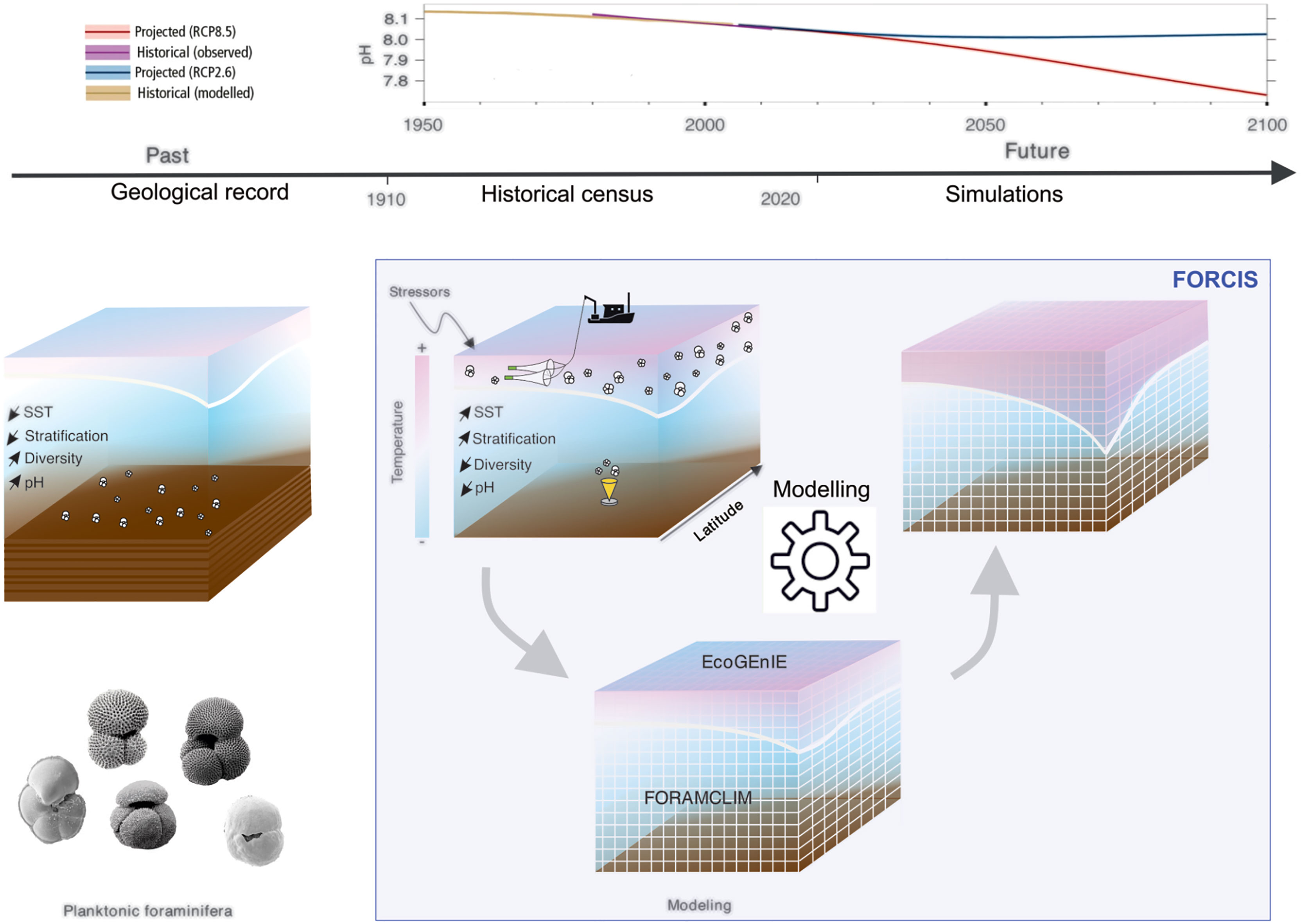
Figure 1 The FORCIS project compiles planktonic Foraminifera census data, analyzes the trends and their links to environmental changes, from historical and future changes in environmental parameters such as pH [top panel, scenario SRES 8.5 & 2.5 – (IPCC, 2019)], and uses those constraints to calibrate models that will be further used for simulations of their past and future distribution patterns.
The impacts of anthropogenic stressors on the current distribution and biodiversity of planktonic organisms are poorly understood. A better knowledge of the role of multiple stressors on the dynamics of planktonic communities today should help to assess future shifts of the marine ecosystem in response to projected further changes of the global marine environment (Hastings et al., 2020). In this context, it is especially important to understand the effect of the multiple, simultaneous global change stressors. For example, the effect of global change on pelagic calcification is hard to predict, because some of the involved stressors act antagonistically: higher temperatures are expected to increase metabolic rates and make calcification energetically less costly, but acidification of surface waters will likely reduce calcification by lowering calcite saturation state (Schiebel and Hemleben, 2017). To understand the effects of global change stressors on marine plankton during historical times requires extensive in-situ observations on a global scale. Understanding these effects is only possible by synthesizing historical datasets of plankton community composition and combining these with records of environmental change/parameters.
Global scale inventories of marine biodiversity based on synoptic genomics and novel in-situ imaging techniques are powerful emergent tools to recover the full spectrum of biodiversity (Lombard et al., 2019). Recent studies on these approaches revealed new challenges for the understanding of the planktonic biodiversity, such as the role of symbiosis in shaping biodiversity patterns (Foster and Zehr, 2019), as well as the importance and limits of the cryptic diversity, and the processes that control its latitudinal gradients (Schiebel et al., 2018). Yet, to capture the processes controlling the pattern and the evolution of biodiversity, a long-term perspective is essential. Only long time-series may reveal the processes, which control the adaptation and acclimation of species to the changing environment. Unfortunately, the genomics era spans less than two decades and access to longer time series will require analyses of observational data with classical taxonomic resolution.
Time-series of abiotic parameters like temperature have been reconstructed over the full instrumental period (i.e., post 1850s), using a combination of modeled and observational data. Using these approaches, it has been demonstrated that the global ocean has warmed by ~1°C over the last century, much of it in response to anthropogenic CO2 release, and concurrent with surface ocean pH decline by about 0.1 unit, i.e., 30%, on average (IPCC, 2019) (Figure 1). Along with the measurements of physical parameters that were used to document environmental changes of the last century, researchers have been sampling and describing the plankton, amassing a unique archive of coeval changes in the plankton community, yet to be exploited. Moreover, historical time-series of the marine biota are spatially scarce and often limited to coastal marine stations affected by a wide range of varying environmental forcing, and/or are limited to the North Atlantic Ocean. No observational open ocean plankton data is available for the preindustrial era, and the sediment record only resolves the pre-industrial evolution of biodiversity in the marine realm, and seldomly for the industrial period (Field et al., 2006).
Planktonic Foraminifera: Ocean Tracers
Among the marine plankton, Foraminifera constitute an ideal tool for documenting those changes, as they have been sampled and counted at the species level since the early 20th century. The taxonomy of planktonic Foraminifera, based on the morphological features of their tests, has been generally stable over the last century, and genetic analyses have confirmed the robustness of the taxonomical framework (Morard et al., 2015). As one of the main oceanic biomineralizing taxa, planktonic Foraminifera secrete an estimated 32 to 80% of the CaCO3 fluxes to the sediments (Schiebel, 2002). Paradoxically, the spatial patterns of species distribution are better known from sedimentary assemblages than from living populations in surface waters, and the interpretation of the fossil record is impeded by a lack of knowledge of the modern population dynamics. Conversely, inferences based solely on the sediment record are overly simplistic as it only provides a mixed account of the long-term evolutionary processes in response to environmental and biological interplays (including archiving overprints).
As planktonic Foraminifera are the most important widespread heterotrophic carbonate producers in the open ocean (Schiebel, 2002), a better knowledge of planktonic Foraminifera ecology is critical for three main reasons: (1) They contribute to the global carbon cycle directly by exporting carbonate to the sea floor. (2) Planktonic Foraminifera constitute one of the best paleo-environmental indicators through their wide spatial and vertical distribution in the global ocean and excellent preservation in deep-sea sediments for over 150 million years. They encode a rich record of ambient environmental conditions during shell formation, providing a key tool for paleoceanographic reconstructions and a time-window on past climate change. Both geochemical (isotopic and trace metal) analyses of their calcareous shells and statistical relations between foraminiferal assemblages and the physical state and/or chemical composition of ambient seawater are used to reconstruct paleoceanographic conditions (CLIMAP members, 1976; de Garidel-Thoron et al., 2005; Kucera et al., 2005). This approach is straightforward over the last millions of years, when all modern species existed, and becomes challenging in the more distant past, prior to evolution of some modern species such as Globorotalia truncatulinoides in the Atlantic Ocean around 2.2 Ma (Spencer-Cervato and Thierstein, 1997). Those reconstructions are used to evaluate climate models dedicated to project future climates. However, lingering uncertainties on the ecology of planktonic Foraminifera make climate model benchmarking using those proxies non trivial. For example, changes in the seasonal and/or vertical abundance affect the climate signal contained in the geochemical composition recorded in Foraminifera shells (Jonkers and Kučera, 2019). (3) Finally, planktonic Foraminifera deposits on the sea floor constitute one of the best records of global biodiversity (Rutherford et al., 1999), which is largely used to interpret evolutionary processes leading to the latitudinal gradient in marine biodiversity. Improving our knowledge of the ecology of planktonic Foraminifera is hence one of the major challenges, and great opportunities of the project presented here.
Planktonic Foraminifera are unique in the surface ocean ecosystem, acting more as ubiquitous tracers rather than forcers, presenting a one-of-a-kind tell-tale opportunity from an organism perspective. By this distinction, planktonic Foraminifera are of low enough abundance that they cause little overall effects on other organisms. Instead, they are relatively passive in a community or ecosystem sense, which actually serves as a unique advantage to trace biodiversity and other measures of surface ocean ecosystem health.
Aims and Community Benefits of FORCIS
Planktonic Foraminifera ecology is still understudied in the modern ocean because of (1) low abundance in the water column, (2) limitations in laboratory culture studies (high mortality rates and lack of second generation), (3) the lack of training of planktologists on the classification and diversity of this group, (4) the intermediate size of Foraminifera in the 63-500 µm range between nannoplankton and mesozooplankton groups studied by planktologists, and (5) the historical focus on geological applications in planktonic Foraminifera studies. Altogether, this created a gap of understanding between the marine biologists studying the modern plankton community, micropaleontologists working on fossil assemblages, and paleoceanographers analyzing the biogeochemical signature of fossil tests (Jonkers et al., 2021). However, despite the difficulties in studying living Foraminifera in experimental settings, the richness of species-level census data from the plankton available in the literature, mostly assembled by geologists, yields an opportunity to investigate the spatial and temporal trends of planktonic Foraminifera biodiversity in a coherent ecological framework.
The FORCIS project aims to quantify the planktonic Foraminifera response to Climatic Stress by compiling and analyzing existing data on diversity and species distribution of these organisms in the global ocean that were generated since 1910. In doing so, we aim to bridge the gap between the biology, ecology and geology scientific fields and advance the knowledge base, for the benefit of a more general understanding on the response of calcifying plankton to global change stressors (Figures 1, 2).
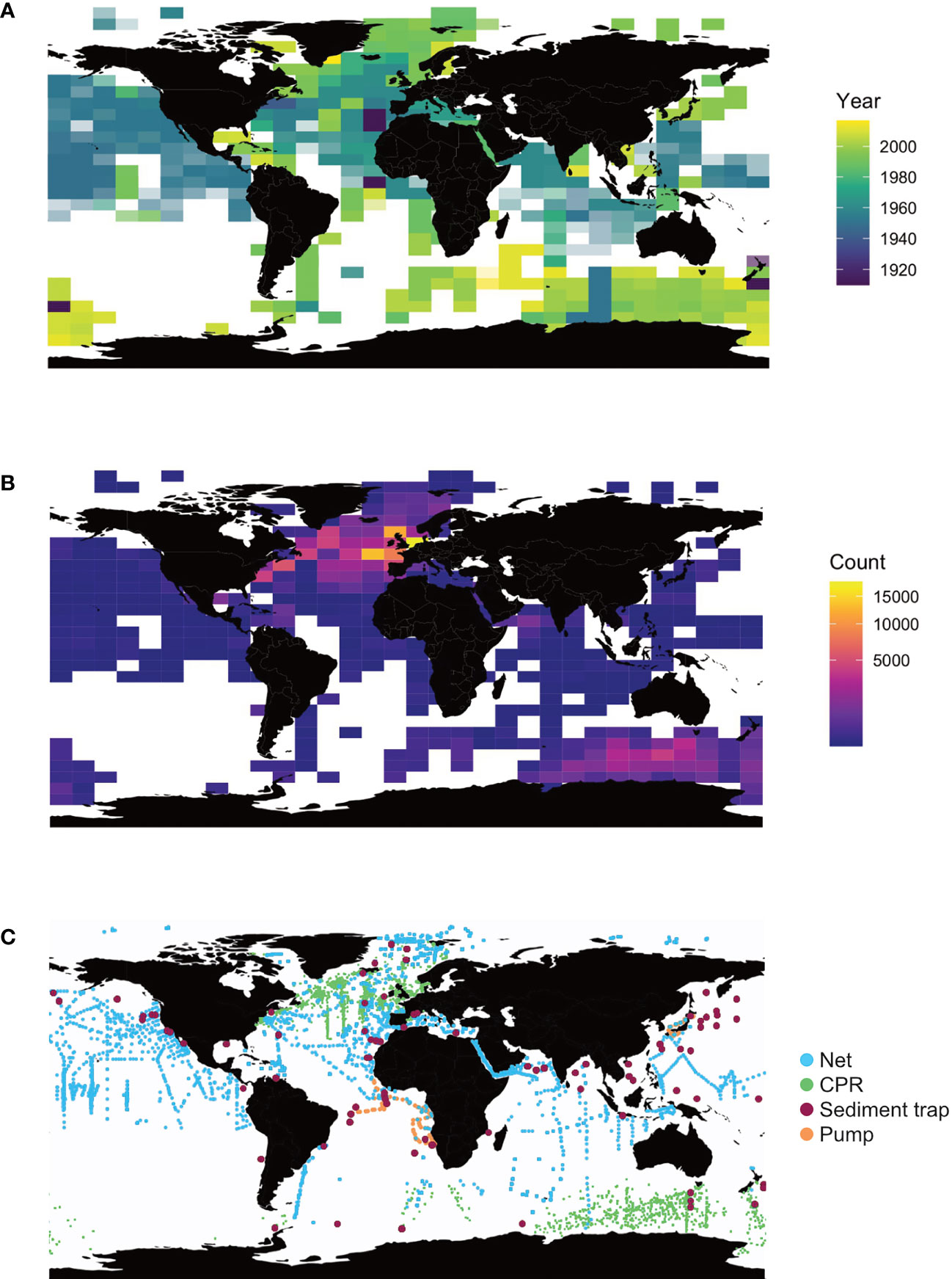
Figure 2 Temporal and spatial coverage of data on planktonic Foraminifera included in the FORCIS database as of November 2020. Maps illustrate (A) the first year of sampling, (B) the number of total observations starting from the first year of sampling, and (C) sampling locations per sampling device in any 5.6° latitude by 11.25° longitude box.
Understanding the modern and predicting the future planktonic Foraminifera response to global environmental change will be achieved by building a global census of planktonic Foraminifera abundances in historical plankton samples (Figure 1). Our strategy towards the FORCIS database has been to assemble data from publications, dissertations, cruise reports, and repositories, often through digitizing older datasets. In FORCIS, census data coming from four types of sampling devices are included in the database: plankton tows, water pump filters, continuous plankton recorder (CPR) (Johns, 2018; Hosie, 2020) and sediment traps (Figure 2), leading to the compilation of more than 180,000 samples (i.e., one single plankton aliquot collected within a depth range, time interval, size fraction, at a single location) including ~157,000 CPR (since 1991), ~16,500 net tow (since 1910), ~7,000 sediment trap (since 1978) and 120 pump (since 1985) samples.
Ecological models coupled to global models offer a unique integrative approach to investigate the links between stressors and the community response. Such models have provided an effective way to conceptualize planktonic Foraminifera ecology at the species level, from simple empirical temperature parameterization of abundance (Zaric et al., 2006; Roche et al., 2018) or growth rate (Lombard et al., 2011), up to more complex trait-based analyses (Fraile et al., 2009; Kretschmer et al., 2018; Grigoratou et al., 2019). These models have provided important insights regarding the interaction between selected planktonic Foraminifera species and their habitat, but their parameterization is based on a limited number of plankton tow and sediment (trap) data. By generating a comprehensive global database of abundance and biomass data, the FORCIS project will allow a much more robust validation and parametrization of the models. Simulations of foraminifer population dynamics constrained by the new data will allow more accurate predictions of future responses of planktonic Foraminifera to different climate scenarios (Roy et al., 2015). We also foresee that the planktonic Foraminifera models will allow quantitative assessments of the total production of foraminifera shell carbonate and organic carbon in climate change scenarios (IPCC, 2022). Over the long term, models addressing the vertical fluxes of planktonic Foraminifera, and the dissolution at the specimen level (Schiebel et al., 2007) might bridge the surface ocean signal recorded in our database with the sedimentary record.
To fully investigate the ecology and global evolution of diversity and abundance of foraminifers, based on the wealth of data assembled within the FORCIS project, and in datasets not yet included, we are open to any community suggestion and are willing to include any scientific dataset or approach able to unravel the processes influencing the evolution of calcifying plankton. In the end, we hope that the FORCIS project will serve as a step forward and will bring plankton ecologists, micropaleontologists and biogeochemists closer together to study planktonic foraminiferal distribution through time and space.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author/s.
Author Contributions
TG-T, SC, XG, JM, LJ, MK, G-JB, MarG, FM, MatG, PM, AK, HH, and RS contributed equally to this work. All authors contributed to the article and approved the submitted version.
Funding
The FORCIS project is funded by the French Foundation for Biodiversity Research (FRB). This research is product of the FORCIS project co-funded by the synthesis center CESAB of the French Foundation for Research on Biodiversity (FRB; www.fondationbiodiversite.fr), French National program LEFE (Les Enveloppes Fluides et l’Environnement), and Max Planck Institute for Chemistry (MPIC).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We appreciate the help of all the modelers and data providers who contributed in designing and building the FORCIS database, especially Marine Courtois, Romain Suarez and Nicolas Casajus.
References
Boyd P. W., Claustre H., Levy M., Siegel D. A., Weber T. (2019). Multi-Faceted Particle Pumps Drive Carbon Sequestration in the Ocean. Nature 568, 327–335. doi: 10.1038/s41586-019-1098-2
CLIMAP members (1976). The Surface of the Ice-Age Earth. Science 191, 1131–1137. doi: 10.1126/science.191.4232.1131
de Garidel-Thoron T., Rosenthal Y., Bassinot F., Beaufort L. (2005). Stable Sea Surface Temperatures in the Western Pacific Warm Pool Over the Past 1.75 Million Years. Nature 433, 294–298. doi: 10.1038/nature03189
de la Vega E., Chalk T. B., Wilson P. A., Bysani R. P., Foster G. L. (2020). Atmospheric CO2 During the Mid-Piacenzian Warm Period and the M2 Glaciation. Sci. Rep. 10, 11002. doi: 10.1038/s41598-020-67154-8
Field D. B., Baumgartner T. R., Charles C. D., Ferreira-Bartrina V., Ohman M. D. (2006). Planktonic Foraminifera of the California Current Reflect 20th-Century Warming. Science 311, 63–66. doi: 10.1126/science.1116220
Foster R. A., Zehr J. P. (2019). Diversity, Genomics, and Distribution of Phytoplankton- Cyanobacterium Single-Cell Symbiotic Associations. Annu. Rev. Microbiol. 73, 435–456. doi: 10.1146/annurev-micro-090817-062650
Fraile I., Mulitza S., Schulz M. (2009). Modeling Planktonic Foraminiferal Seasonality: Implications for Sea-Surface Temperature Reconstructions. Mar. Micropaleontol. 72, 1–9. doi: 10.1016/j.marmicro.2009.01.003
Grigoratou M., Monteiro F. M., Schmidt D. N., Wilson J. D., Ward B. A., Ridgwell A. (2019). A Trait- Based Modelling Approach to Planktonic Foraminifera Ecology. Biogeosciences 16, 1469–1492. doi: 10.5194/bg-16-1469-2019
Hastings R. A., Rutterford L. A., Freer J. J., Collins R. A., Simpson S. D., Genner M. J. (2020). Climate Change Drives Poleward Increases and Equatorward Declines in Marine Species. Curr. Biol. 30, 1572–1577. doi: 10.1016/j.cub.2020.02.043
Hosie G. (2020) Southern Ocean Continuous Plankton Recorder Zooplankton Records, Ver. 8. Australian Antarctic Data Centre (Accessed 2021-01).
Houghton J. T., Ding Y., Griggs D. J., Noguer M., van der Linden P. J., Xiaosu D. (2001). Climate Change 2001: The Scientific Basis - Contribution of Working Group I to the Third Assessment Report of the Intergovernmental Panel on Climate Change (IPCC). (Cambridge: United Kingdom and New York, NY, USA), 944.
IPCC (2019). “Summary for Policymakers,” in IPCC Special Report on the Ocean and Cryosphere in a Changing Climate. Eds. Pörtner H.-O., Roberts D. C., Masson-Delmotte V., Zhai P., Tignor M., Poloczanska E., Mintenbeck K., Alegría A., Nicolai M., Okem A., Petzold J., Rama B., Weyer N. M.
IPCC (2022). Climate Change 2022: Impacts, Adaptation, and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change Eds. Pörtner H.-O., Roberts D.C., Tignor M., Poloczanska E.S., Mintenbeck K., Alegría A., Craig M., Langsdorf S., Löschke S., Möller V., Okem A., Rama. B. Cambridge University Press. In Press.
Ji R., Edwards M., Mackas D. L., Runge J. A., Thomas A. C., et al (2010). Marine Plankton Phenology and Life History in a Changing Climate: Current Research and Future Directions. J. Plankton Res. 32, 1355–1368. doi: 10.1093/plankt/fbq062
Johns D. (2018). Foraminifera Abundance Per CPR Sample on the North Atlantic 1993 to 2017. The Archive for Marine Species and Habitats Data (DASSH).
Jonkers L., Meilland J., Rillo M. C., de Garidel-Thoron T., Kitchener J. A., Kucera M. (2021). Linking Zooplankton Time Series to the Fossil Record. ICES J. Mar. Sci., fsab123. doi: 10.1093/icesjms/fsab123
Jonkers L., Hillebrand H., Kucera M. (2019). Global Change Drives Modern Plankton Communities Away From the Pre-Industrial State. Nature 570, 372–375. doi: 10.1038/s41586-019-1230-3
Jonkers L., Kučera M. (2019). Sensitivity to Species Selection Indicates the Effect of Nuisance Variables on Marine Microfossil Transfer Functions. Climate Past 15, 881–891. doi: 10.5194/cp-15-881-2019
Kretschmer K., Jonkers L., Kucera M., Schulz M. (2018). Modeling Seasonal and Vertical Habitats of Planktonic Foraminifera on a Global Scale. Biogeosciences 15, 4405–4429. doi: 10.5194/bg-15-4405-2018
Kucera M., Rosell-Mele A., Schneider R., Waelbroeck C., Weinelt M. (2005). Multiproxy Approach for the Reconstruction of the Glacial Ocean Surface (MARGO). Quat. Sci. Rev. 24, 813–819. doi: 10.1016/j.quascirev.2004.07.017
Lombard F., Boss E., Waite A. M., Vogt M., Uitz J., Stemmann L., et al. (2019). Globally Consistent Quantitative Observations of Planktonic Ecosystems. Front. Mar. Sci. 6, 1705–1722. doi: 10.3389/fmars.2019.00196
Lombard F., Labeyrie L., Michel E., Bopp L., Cortijo E., Retailleau S., et al. (2011). Modelling Planktic Foraminifer Growth and Distribution Using an Ecophysiological Multi-Species Approach. Biogeosciences 8, 853–873. doi: 10.5194/bg-8-853-2011
Maas A. E., Lawson G. L., Bergan A. J., Tarrant A. M. (2018). Exposure to CO2 Influences Metabolism, Calcification and Gene Expression of the Thecosome Pteropod Limacina Retroversa. J. Exp. Biol. 221, 1–13. doi: 10.1242/jeb.1644000
Morard R., Darling K. F., Mahé F., Audic S., Ujiie Y., Weiner A. K. M., et al. (2015). PFR 2: A Curated Database of Planktonic Foraminifera 18S Ribosomal DNA as a Resource for Studies of Plankton Ecology, Biogeography and Evolution. Mol. Ecol. Resour. 15, 1472–1485. doi: 10.1111/1755-0998.12410
Poloczanska E. S., Brown C. J., Sydeman W. J., Kiessling W., Schoeman D. S., Moore P. J., et al. (2013). Global Imprint of Climate Change on Marine Life. Nat. Climate Change 3, 919–925. doi: 10.1038/nclimate1958
Roche D. M., Waelbroeck C., Metcalfe B., Caley T. (2018). FAME (V1.0): A Simple Module to Simulate the Effect of Planktonic Foraminifer Species-Specific Habitat on Their Oxygen Isotopic Content. Geosci. Model Dev. 11, 3587–3603. doi: 10.5194/gmd-11-3587-2018
Roy T., Lombard F., Bopp L., Gehlen M. (2015). Projected Impacts of Climate Change and Ocean Acidification on the Global Biogeography of Planktonic Foraminifera. Biogeosciences 12, 2873–2889. doi: 10.5194/bg-12-2873-2015
Rutherford S., D’hondt S., Prell W. (1999). Environmental Controls on the Geographic Distribution of Zooplankton Diversity. Nature 400, 749–753. doi: 10.1038/23449
Schiebel R. (2002). Planktic Foraminiferal Sedimentation and the Marine Calcite Budget. Global Biogeochem. Cycles 16, 1065. doi: 10.1029/2001GB001459
Schiebel R., Barker S., Lendt R., Thomas H., Bollmann J. (2007). Planktic Foraminiferal Dissolution in the Twilight Zone. Deep Sea Res. Part II: Topical Stud. Oceanogr. 54, 5–7 : 676-686.
Schiebel R., Hemleben C. (2017). Planktic Foraminifers in the Modern Ocean (Berlin, Heidelberg: Springer Berlin Heidelberg), 358. doi: 10.1016/j.dsr2.2007.01.009
Schiebel R., Smart S. M., Jentzen A., Jonkers L., Morard R. L., Meilland J, et al. (2018). Advances in Planktonic Foraminifer Research: New Perspectives for Paleoceanography. Rev. Micropaléontol. 61, 113–138. doi: 10.1016/j.revmic.2018.10.001
SpencerCervato C., Thierstein H. (1997). First Appearance of Globorotalia Truncatulinoides: Cladogenesis and Immigration. Mar. Micropaleontol. 30, 267–291. doi: 10.1016/S0377-8398(97)00004-2
Keywords: plankton, foraminifera, global warming, ocean acidification, biodiversity
Citation: de Garidel-Thoron T, Chaabane S, Giraud X, Meilland J, Jonkers L, Kucera M, Brummer G-JA, Grigoratou M, Monteiro FM, Greco M, Mortyn PG, Kuroyanagi A, Howa H, Beaugrand G and Schiebel R (2022) The Foraminiferal Response to Climate Stressors Project: Tracking the Community Response of Planktonic Foraminifera to Historical Climate Change. Front. Mar. Sci. 9:827962. doi: 10.3389/fmars.2022.827962
Received: 02 December 2021; Accepted: 08 April 2022;
Published: 06 May 2022.
Edited by:
Sergio M. Vallina, Spanish Institute of Oceanography (IEO), SpainReviewed by:
Katsunori Kimoto, Japan Agency for Marine-Earth Science and Technology (JAMSTEC), JapanCopyright © 2022 de Garidel-Thoron, Chaabane, Giraud, Meilland, Jonkers, Kucera, Brummer, Grigoratou, Monteiro, Greco, Mortyn, Kuroyanagi, Howa, Beaugrand and Schiebel. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Thibault de Garidel-Thoron, Z2FyaWRlbEBjZXJlZ2UuZnI=