 Antonio Fernández1
Antonio Fernández1 Eva Sierra1*
Eva Sierra1* Manuel Arbelo1
Manuel Arbelo1 Ana Gago-Martínez2Jose Manuel Leao Martins2
Ana Gago-Martínez2Jose Manuel Leao Martins2 Natalia García-Álvarez1Yara Bernaldo de Quiros1
Natalia García-Álvarez1Yara Bernaldo de Quiros1 Marina Arregui1Ana Isabel Vela3,4
Marina Arregui1Ana Isabel Vela3,4 Josue Díaz-Delgado1
Josue Díaz-Delgado1
- 1Veterinary Histology and Pathology, Veterinary School, Institute of Animal Health, University of Las Palmas de Gran Canaria, Las Palmas, Spain
- 2Department of Analytical and Food Chemistry, Biomedical Research Center (CINBIO), University of Vigo, Vigo, Spain
- 3Centro de Vigilancia Sanitaria Veterinaria (VISAVET), Complutense University, Madrid, Spain
- 4Department of Animal Health, Faculty of Veterinary, Complutense University of Madrid, Madrid, Spain
Harmful algal blooms (HABs) have been increasingly recorded over the last decades and much work has linked these events to multiple oceanographic and climate disturbances. HABs can affect ecosystems either as events that affect dissolved oxygen, clog fish gills, or smother corals or through the production of biotoxins which affect living marine resources through food web transfers or aerosols. HAB represent a natural driver of decline and potential extinction of aquatic organisms, from invertebrates to mammals, which may offer little evolutionary adaptation particularly in very high and long-lasting exposures. Despite numerous multispecies mass-mortality events linked to HAB-associated biotoxicosis globally, there are no records in cetaceans off the central eastern Atlantic Ocean. Herein, we report the epidemiology, pathologic, microbiologic and toxicologic investigation results attesting to the first documentation of cetacean mass-mortality in European waters associated with brevetoxins. Twelve rough-toothed dolphins (Steno bredanensis) were found dead adrift or beached along the southwestern coast of Gran Canaria from April 28th to May 7th, 2008. Although pathologic examinations were limited by moderate to advanced autolysis and decomposition of the carcasses, consistent findings included multisystemic hemorrhage and undigested ingesta within the gastric compartments, mainly salema porgy (Sarpa salpa). Toxicologic analysis of gastric contents identified PbTx2 and PbTx3 brevetoxins. Our results provide compelling toxicopathologic evidence of fatal brevetoxicosis in a cohort of rough-toothed dolphins. These data add to the limited knowledge on pathology of HAB in cetaceans and provide the first account of brevetoxicosis in European waters. No other mass-mortality or individual fatality of any cetacean species has been linked to brevetoxicosis in the Canary Islands since this event.
Introduction
Harmful algal blooms (HABs) are increasingly frequent worldwide, occurring in lake, riverine, coastal and pelagic waters (Lowenstine, 2007). Different species of microalgae in these blooms, including diatoms and dinoflagellates, produce a variety of biotoxins potentially detrimental for aquatic organisms and human beings. HABs can affect ecosystems either as events that affect dissolved oxygen, clog fish gills, or smother corals or through the production of biotoxins which affect living marine resources through food web transfers or aerosols. HAB represent a natural driver of decline and potential extinction of aquatic organisms, from invertebrates to mammals, which may offer little evolutionary adaptation particularly in very high and long-lasting exposures (Lowenstine, 2007). While morbidity and mortality are among the grave outcomes linked to ingested biotoxin exposures (Geraci et al., 1989; Scholin et al., 2000; Flewelling et al., 2005), more subtle sequela may occur as result of prolonged, non-lethal exposure to different biotoxins (Brodie et al., 2006; Goldstein et al., 2008; Zabka et al., 2009). Massive die-offs involving many different fish species, manatees, sea lions, odontocetes, mysticetes and giant squids have been reported (Lowenstine, 2007). Furthermore, many of these biotoxins pose concerns for public health through ingestion of biotoxins in food or inhalation of biotoxins downwind of the bloom (Gerssen and Gago-Martinez, 2019).
Naturally occurring brevetoxicosis fatalities have involved aquatic invertebrates, fish, sea turtle, seabirds, marine mammals (pinnipeds, cetaceans, and sirenians), terrestrial mammals (coyotes, domestic dogs), green tree frogs, and ground squirrels (Castle et al., 2013; Buttke et al., 2018). Brevetoxins have had the greatest impact in aquatic organisms as they may vector into the food chain, killing fish, invertebrates, birds, sea turtles, and marine mammals (Lowenstine, 2007). Several mortality events in odontocete species, primarily involving common bottlenose dolphins (Tursiops truncatus) have been recorded in the Gulf of Mexico, mostly along the west coast of Florida (Fire et al., 2021). Brevetoxins are produced primarily by dinoflagellates during “red tides,” predominantly by members of genus Karenia. The latter encompasses eight species: K. brevis (formerly known as Gymnodinium breve and Ptychodiscus brevis), K. mikimotoi, K. brevisulcata, K. longicanalis, K. digitata, K. papilionacea, K. selliformis, and K. bidigitata. Reliable identification of these species may require a combination of morphologic features and molecular analysis (Haywood et al., 2004). K. bicuneiformis (syn. K. bidigitata), K. papilionacea, and K. brevis have been recorded in the Canarian archipelago (Ojeda, 2005, 2013). Specifically, K. papilionacea is able to produce neurotoxins and K. brevis produces neurotoxins (designated PbTxs) and hemolytic toxins (Fire et al., 2008a,2015). Other potentially toxic metabolites, such as hemi-brevetoxins, brevenals, brevisamides, brevisins, tamulamides, gymnocins, gymnodimines, brevisucenals, and brevisulcatic acids, can be produced by Karenia species (Caruana and Amzil, 2018; Hort et al., 2021). Intoxication occurs primarily by ingestion of contaminated prey or marine vegetation or by inhalation of toxins in aerosol during algal blooms. Limited clinical signs have been recorded in cetaceans with brevetoxicosis, mainly due to the peracute nature of the neurotoxicity. There are records of low levels of PbTxs in stranded dolphins without specific pathologic findings, and acute neurologic signs have been observed in some dolphins during Karenia blooms (Rowles, T., personal communication). Herein, we report the epidemiology, pathologic, microbiologic and toxicologic investigations results attesting to the first episode of brevetoxicosis-related cetacean mass-mortality event in European waters involving 12 rough-toothed dolphins (Steno bredanensis).
Materials and Methods
Stranding Epidemiology
Twelve rough-toothed dolphins were found adrift or beached along the southeastern coastline of Gran Canaria over a 10 day-period, from April 28th to May 7th, 2008. Carcasses were retrieved and submitted for necropsy to the Institute for Animal Health and Food Safety, University of Las Palmas of Gran Canaria. Biologic data [age class, sex, body condition (BD), conservation status (CS), morphometrics] and stranding conditions for each individual were recorded systematically (Table 1). Age class was based on total body length and gonad development, including fetus/neonate/calf, juvenile/subadult, and adult (Geraci and Lounsbury, 2005; Arbelo et al., 2013; Diaz-Delgado et al., 2018). BD was subjectively classified into good, moderate, poor, and emaciated according to anatomic parameters such as the osseous prominence of the spinous and transverse vertebral processes and ribs, the mass of the epaxial musculature, and the amount of fat deposits, taking into account the species and the age of the animal (Geraci and Lounsbury, 2005; Arbelo et al., 2013; Diaz-Delgado et al., 2018). Carcasses were classified as very fresh, fresh, moderate autolysis, advanced autolysis or very advanced autolysis (Arbelo et al., 2013; Diaz-Delgado et al., 2018). The permission required for the handling of stranded cetaceans within the Canarian archipelago was issued by the environmental department of the Canary Islands’ Government. No experiments were performed on live animals because our work was based on dead stranded cetaceans.
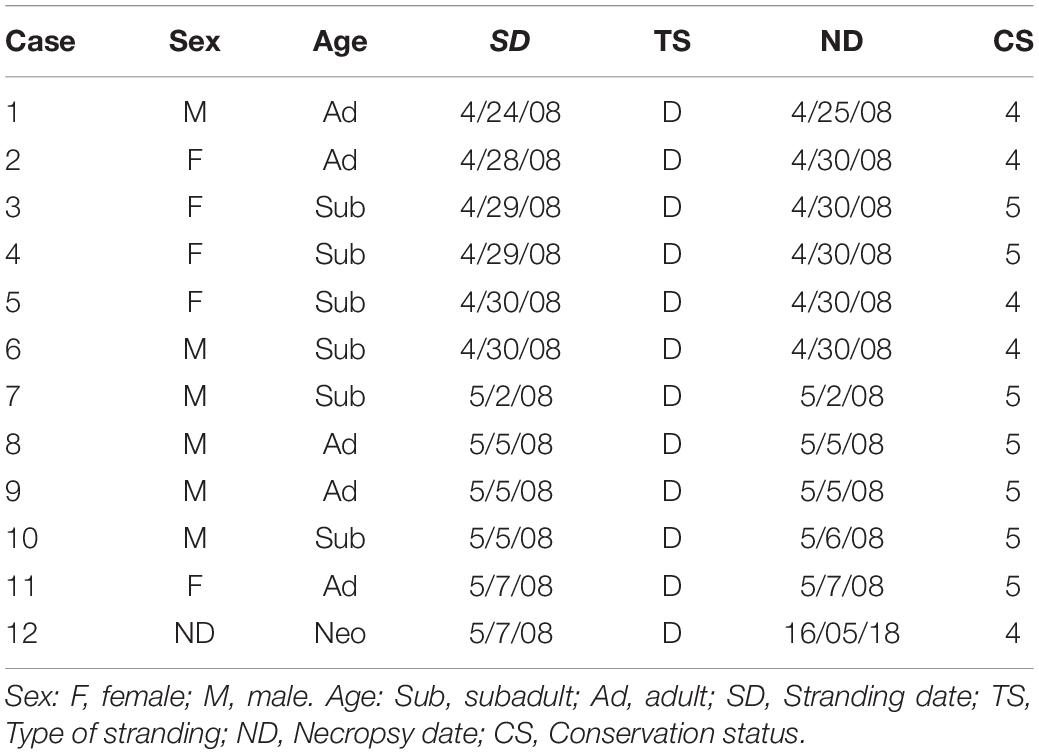
Table 1. Life history data of 11 rough-toothed dolphins (Steno bredanensis) found adrift off or beached along the Southeastern coast of Gran Canaria.
Pathologic Examinations
Necropsies followed standardized protocols (Geraci and Lounsbury, 2005; Arbelo et al., 2013; Diaz-Delgado et al., 2018). Representative tissue samples were collected and fixed in 10% neutral buffered formalin. All these tissues were processed routinely, embedded in paraffin-wax and 5 μm-thick sections were stained with hematoxylin and eosin for microscopic analysis. For histopathologic analysis, formalin-fixed tissues were embedded in paraffin-wax, sectioned at 5 μm, and stained with hematoxylin-eosin. To rule out fat embolism, osmium tetroxide (post-fixation) was used on selected lung tissue sections (Arregui et al., 2020).
Chemical Analysis
Fresh samples (stomach content, skeletal muscle, liver, kidney) from seven animals were collected during necropsy. No water samples were collected. The samples were stored frozen (−80°C) and were submitted to liquid chromatography–mass spectrometry (LC-MS) analysis for the identification of brevetoxins (PbTx). Brevetoxins extraction from homogenized tissue was carried out according to Mcnabb et al. (2012). Briefly, homogenized sample tissue (2.0 ± 0.1 gr) was weighted into 50 mL centrifuge tube, a 9 mL volume of 80% (v/v) aqueous methanol was added and the content was mixed using a vortex during 1 min. The sample was then heated in a water bath at 60 ± 2°C for 20 min. The sample was cooled in an ice bath and centrifuged at 3,000 g for 10 min. The supernatant was pipetted and transferred into another 50 mL centrifuge tube, and the tissue pellet re-extraction was repeated. The final extract volume was adjusted into 20 mL with Milli Q water and the final extract solution was mixed very well; 8 mL of the liver extract was transferred to a 50 mL pyrex tube and partitioned with 10 mL of n-hexane. The hexane layer containing lipid compounds was removed and MilliQ water was added to 5 mL of methanolic extract until achieve 20 mL of final volume extract for purification [solid-phase extraction (SPE) procedure].
PbTx sample cleanup was carried out according to Mcnabb et al. (2012) with slight modifications. Strata-X SPE column (60 mg, 3 mL) was conditioned with 3 mL of methanol and 3 mL of 25% (v/v) aqueous methanol. A total of 20 mL of extract was loaded onto conditioned SPE cartridges at flow rate approximately one to two drops per second followed by 2 mL/min. The stationary phase was subsequently washed with 4.5 mL of 25% (v/v) aqueous methanol at the same flow rate. Graduated tubes (15 mL) were placed inside the manifold for elution volume collection; the retained PbTx in SPE stationary phase was eluted with 4.5 mL of methanol. The final volume was made up to 5.0 mL with MilliQ and mixed; 1 mL of purified extract was filtered with 0.22μM Millipore membrane and injected into LC-MS system.
PbTx analysis was performed by LC-MS using a single Quadropole, Mass Spectrometer under the conditions initially described in Mcnabb et al. (2012) with some slight modifications. A LC/MS Hewlett Packard (HP) System equipped with LC Binary Pump HP Series 100 coupled to Single Quadrupole Mass Spectrometer 1,100 Series MSD using Electrospray (ESI) interface was used. PbTx chromatographic separation was carried out using Luna C18 Reverse Phase LC column (50 × 2 mm, 3 μm, Phenomenex) as stationary phase. PbTx elution was conducted by using mobile phase A (33 mM of ammonium hydroxide and 500 mM ammonium formate in 50% v/v of MeOH) and mobile phase B (33 mM of ammonium hydroxide and 500 mM ammonium formate in 90% v/v of MeOH). The gradient is recorded in Supplementary Table 1. The flow rate was 0.20 mL/min, the injection volume was 20 μL, and the column was set at room temperature. Positive electrospray ionization was used with the following parameters: nitrogen (N2) gas flow 9 L/h, drying gas temperature 350°C, capillary voltage 3,500 V, and fragmentor 120 V. Single Quadrupole mass analyzer was operated in SIM (Single Ion Monitoring) mode. PbTx2 was monitored select m/z (mass/charge) 896 and PbTx3 898, both corresponding to [M + H]+ protonated molecule fragment ion. PbTx2 and PbTX3 standard solutions were kindly provided by Dr. Robert W Dickey (Marine Sciences, University of Texas, United States); 1 μg/mL of individual toxin standard solution was prepared in methanol (grade LC-MS) and used for the identification and quantitation of brevetoxins in dolphin samples.
Diet (Prey Items) Analysis
Gastric contents including intact or fragments of teleost fishes as well as squid beaks and otoliths were collected and submitted for taxonomic identification using national and regional catalogs and identification keys.
Microbiologic Analysis
Fresh tissue samples (liver, kidney, and lung) from six animals were collected routinely during necropsy, were frozen (−80°C) and selectively submitted for bacteriologic analysis. These included routine culture and surface plating on routine media, e.g., Columbia blood agar and preliminary identification of isolates via API® system (API® 20E, API® Rapid 20E, API® Staph, API® 20 Strep, API® Coryne, API® 20A). PCR targeting the 16S rRNA gene coupled with pulsed-field gel electrophoresis were performed on selected isolates.
Virologic Analysis
At necropsy, selected samples (liver, kidney, lymph nodes, brain) were collected for virologic analyses and stored frozen at −80°C until processing for molecular virology testing. Approximately 0.5 gr of fresh-frozen tissue sample from each animal was mechanically macerated in lysis buffer and subsequently centrifuged. DNA/RNA extraction was carried out from each 300 μL macerated sample by pressure filtration method, using a QuickGene R Mini 80 nucleid acid isolation instrument, using the DNA Tissue Kit S (QuickGene, Kurabo, Japan) according to the manufacturer’s instructions with modifications: RNA carrier (Applied Biosystems™, Thermo Fisher Scientific Waltham, Massachusetts, United States.) was added during the lysis step (Sacristan et al., 2015). Herpesvirus DNA was detected by conventional nested PCR using degenerate primers designed to amplify a region of the DNA polymerase gene (VanDevanter et al., 1996). Molecular detection of cetacean morbillivirus (CeMV) was performed by one or more of four different PCR methods: one-step RT-PCR of a 426-bp conserved region of the phosphoprotein (P) gene (Reidarson et al., 1998), one-step real-time RT-PCR that detect the most common CeMV strains known to circulate in the Atlantic Ocean targeting the P gene (Groch et al., 2020), RT-PCR using nested primers targeting the P gene (Sierra et al., 2014), and one-step real-time RT-PCR to detect sequences in a conserved region (192 bp) of the fusion protein (F) gene (Sacristan et al., 2015).
Results
Epidemiology, Pathologic Investigations, and Diet Analysis
On April 15th, a group of approximately 50 rough-toothed dolphins was sighted in the area where carcasses were later found (27°48′15N; 15°46′46). This group is well known in the area by whale-watching ships as they could be found close to shore (< 2–3 nautical miles). That day the animals were observed as usually, in a very cohesive group including adults and calves. On April 16th, biologists working on board (whale-watching ships) informed about seagrass floating in that area; no alterations on behavior or social structure were noticeable in the group of dolphins. However, on April 17th, the group had dispersed in small groups or individually. One of the dolphins was very quiet for a long time and behaved oddly (as per on board biologist). On April 23rd evening, one dolphin was found dead floating with moderate autolysis around the same location as previous sightings. On April 24th, a group of dolphins (n = 30) was observed (within < 1 nautical mile) displaying normal cohesive behavior including adults and juveniles. On April 27th, a second dolphin was found dead floating. Additional carcasses were recovered on the following days, two on April 29th, two on April 30th, one on May 1st, two on May 4th, and one on May 5th, 7th, and 10th, each, mostly with advanced/very advanced autolysis.
These 12 dolphins were investigated. All the dolphins were in apparent good body condition. Age classes were: neonate (n = 1), adult (n = 5), subadult (n = 6); sex was: male (n = 6), female (n = 5). Nine were found adrift and three beached dead along the coast. Four presented advanced autolysis and eight had very advanced autolysis (Table 1) at necropsy. Pathologic examinations were limited by carcass preservation status; however, varying degrees of multisystemic hemorrhage within the subcutis and axial muscles in occipital, intermandibular, thoracic, and abdominal regions, as well as hemorrhage in the rete mirabile, stomach, esophagus, larynx, and lung, were seen (Figure 1). Most of the animals presented undigested or partially digested prey items within the gastric compartments, primarily salema porgy (Sarpa salpa) (Figure 2). Pathologic findings are recorded in Table 2.
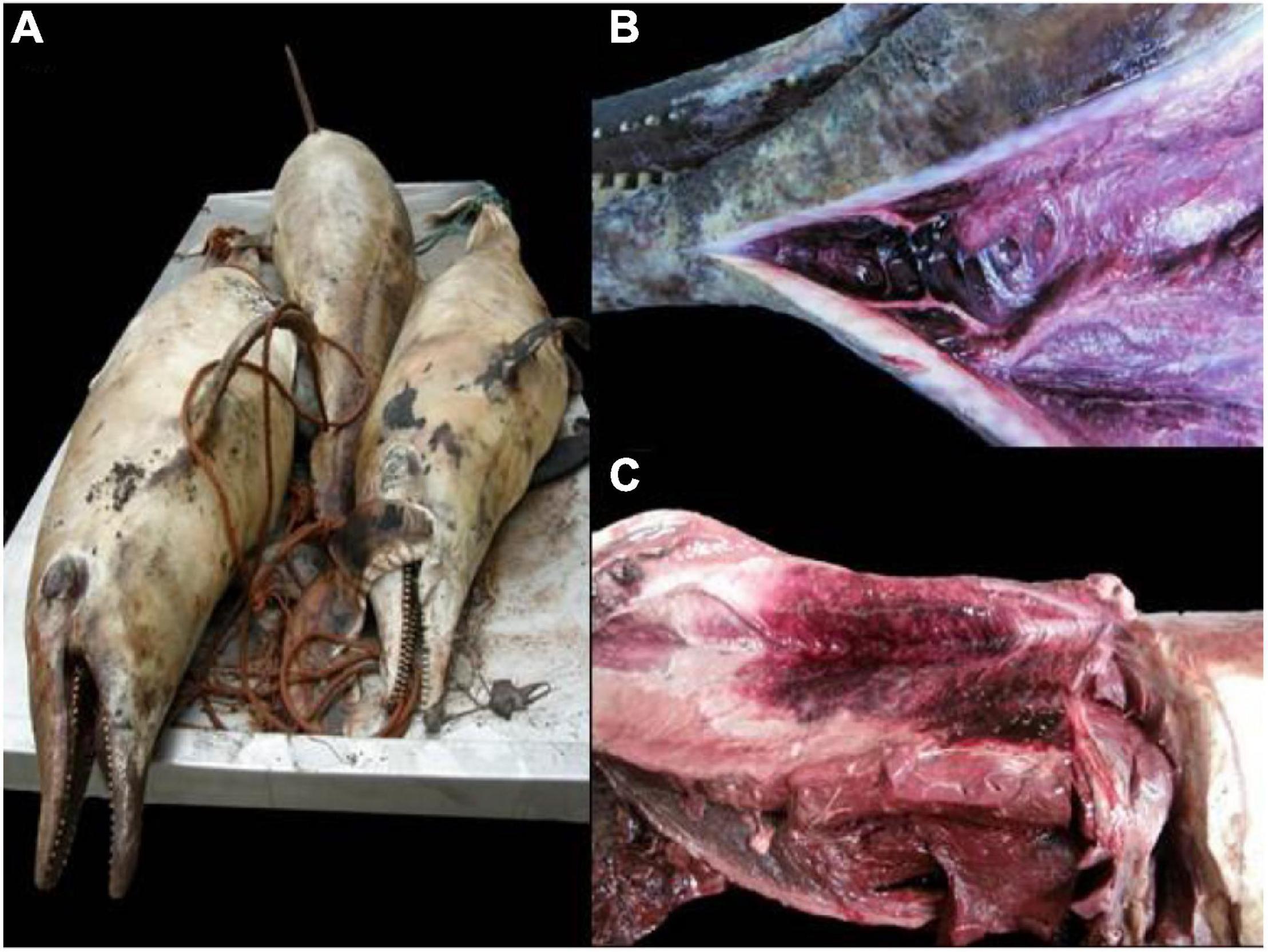
Figure 1. Gross findings in rough-toothed dolphins (Steno bredanensis) included in this study. (A) Carcasses exhibit advanced decomposition and autolysis. (B) Focal subcutaneous intermandibular hemorrhage. (C) Focally extensive hemorrhage in the subcutis and axial skeletal muscle on the left thoracolumbar area.
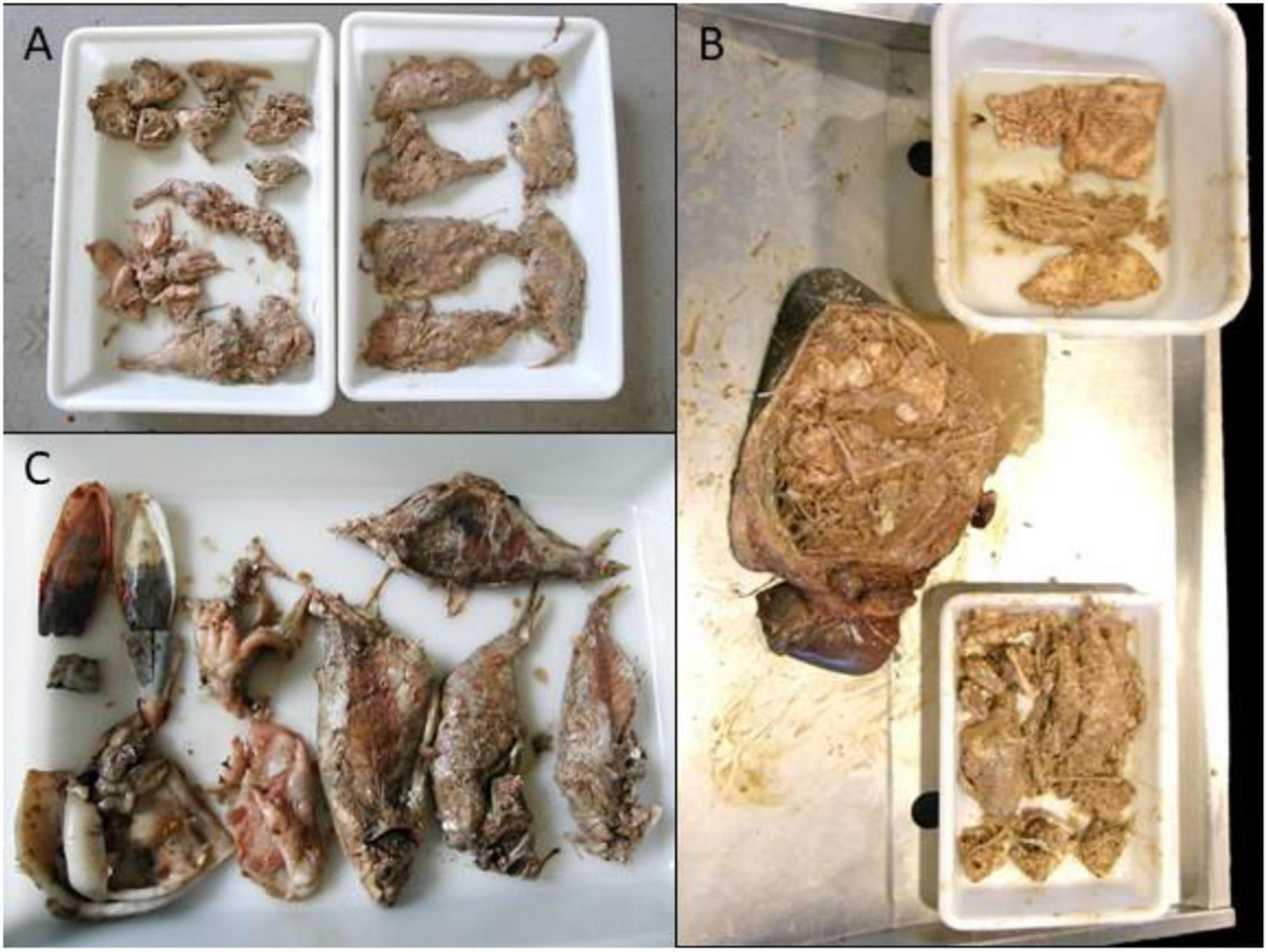
Figure 2. Gastric contents in case 9 (A), case 8 (B) and case 5 (C).
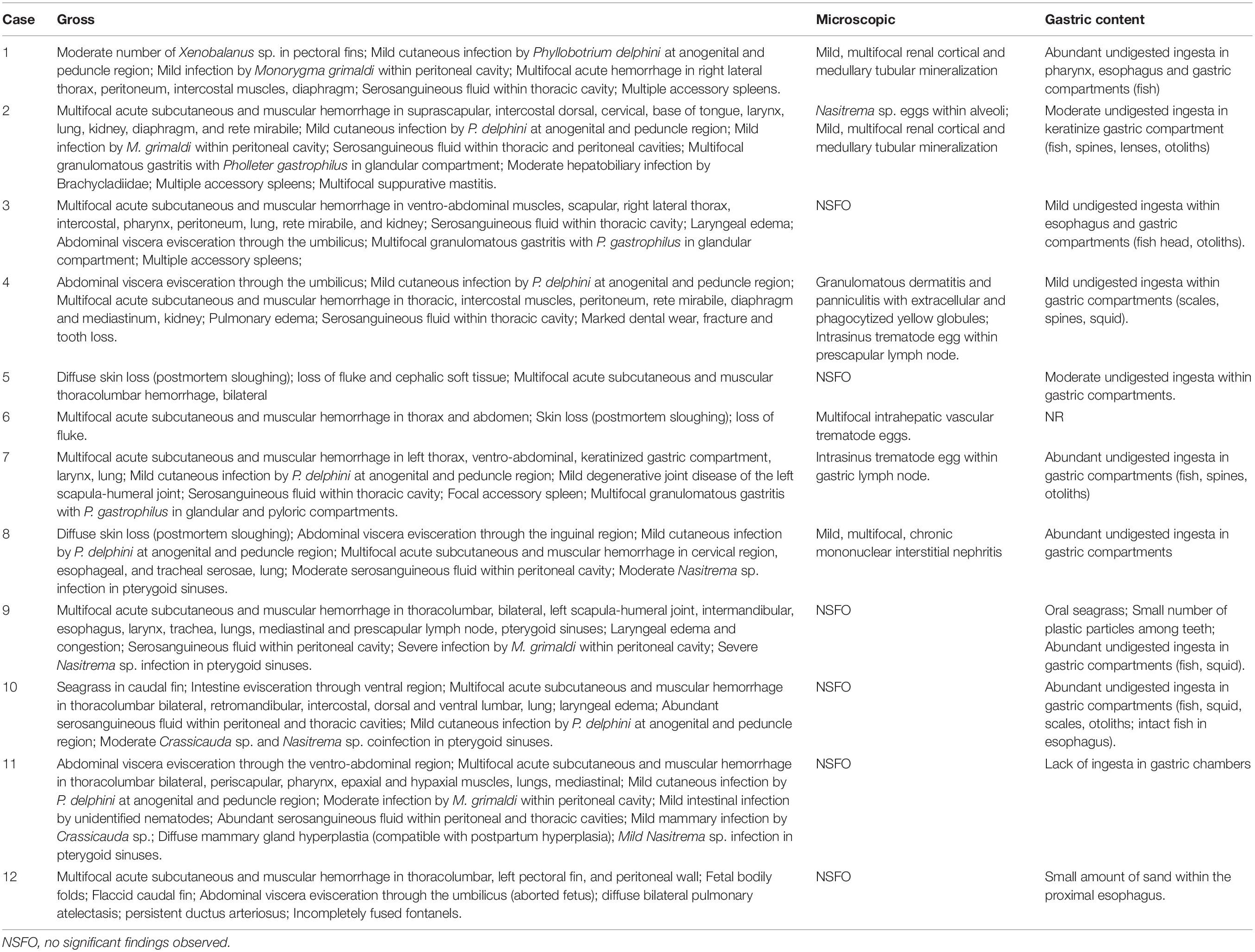
Table 2. Pathologic findings in rough-toothed dolphins (Steno bredanensis) included in this study.
A total of 28 samples from 7 dolphins was examined (Table 3). The retention times observed for each toxin were 13.42 min for PbTx3 and 13.81 min for PbTX2 (Figure 3). Positive samples for PbTx were those in which PbTx2 and PbTx3 were detected; chromatographic data and m/z fragment ion values for these two toxins, for which reference standards were available, were taken into account. Qualitative LC-MS analysis results obtained for seven dolphins tested (cases 190 through 196) are summarized in Table 3. PbTx2 and PbTx3 were detected in gastric contents of 3/4 and 4/4 animals, respectively. An example of LC-MS chromatogram corresponding to the analysis of liver extract from case #196 where PbTx2 and PbTx3 were detected at trace level is recorded in Figure 4.
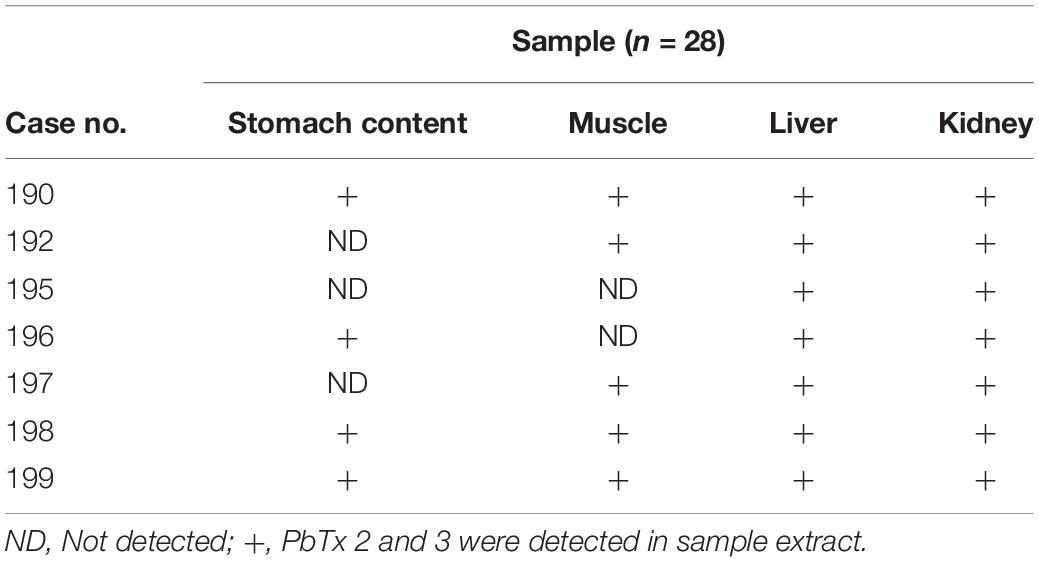
Table 3. Results obtained for brevetoxins screening analysis by LC-MS of dolphin samples collected in Canary Island (Spain).
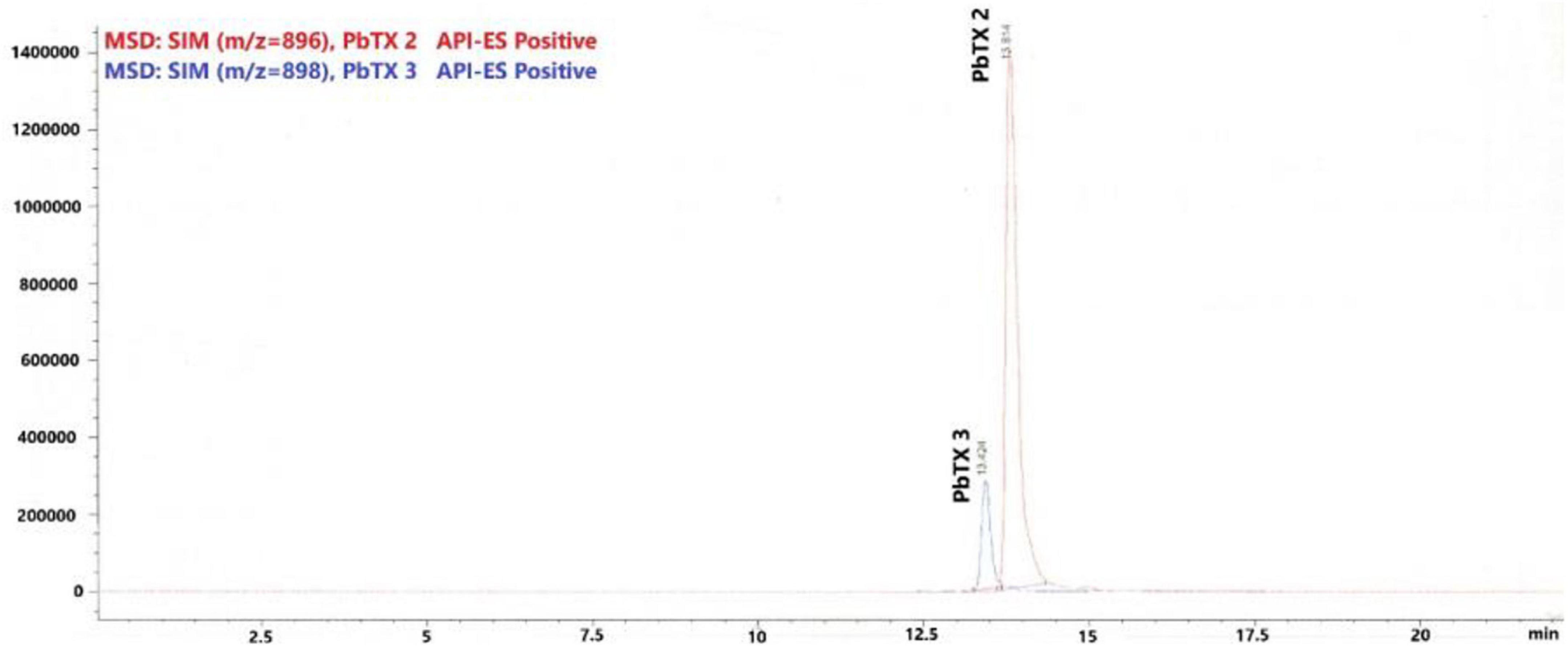
Figure 3. Chromatogram from liquid chromatography–mass spectrometry (LC-MS) analysis of standard solution contains PbTX2 and PbTX3, concentration of each toxin in solution 1μg/mL.
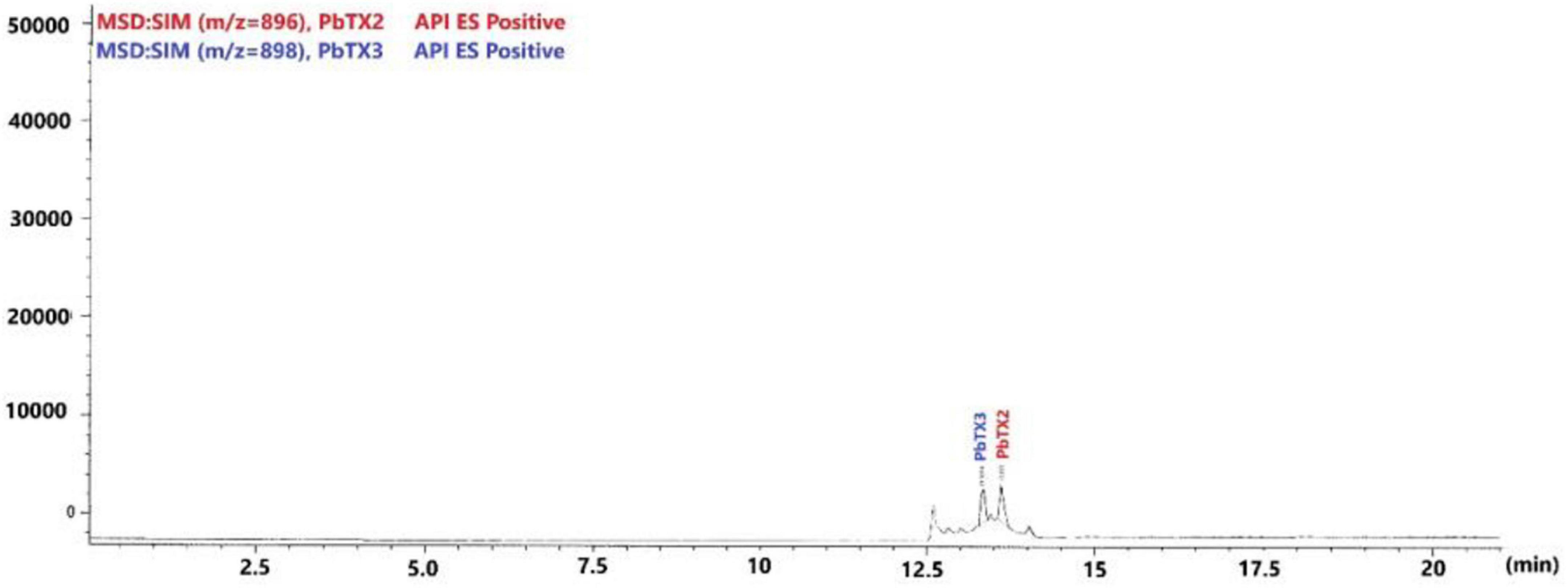
Figure 4. Chromatogram from liquid chromatography–mass spectrometry (LC-MS) analysis of liver extract from case #6.
Microbiologic Analysis
Bacterial culture of liver, kidney, and lung from six animals yielded Clostridium novyi. Animals tested were negative for CeMV and HV.
Discussion
Brevetoxicosis was first suspected and later confirmed to result in cetacean fatalities since 1946, primarily in manatees and to a lesser extent in common bottlenose dolphins in the Gulf of Mexico (Gunter et al., 1948; Van Dolah, 2005). Several UME involving common bottlenose dolphins have been described ever since, primarily in the west coast of Florida and Florida Panhandle regions yet fewer fatalities have been seen in the Atlantic coast of Florida (Mase et al., 2000; Flewelling et al., 2005; Gaydos, 2006; Twiner et al., 2012; Litz et al., 2014; Fire et al., 2015). Mortalities of dolphins with multiple biotoxins including brevetoxins found in the carcasses have been documented (Fire et al., 2011). Little is known about the effects of exposures to multiple biotoxins and whether the toxins are synergistic, additive or neutral. Limited clinical signs have been documented in cetaceans with brevetoxicosis, mainly due to the peracute and fatal nature of the toxicity. Clinical signs may include stranding, seizures, and incoordination, leading to death (Fauquier and Landsberg, 2018). Furthermore, brevetoxins have been shown to interfere with lymphocyte function in vitro (Gebhard et al., 2015), which could be a potential sublethal effect. This remains to be proven in vivo. Pathologic findings in cetaceans known to have succumbed to brevetoxins are largely lacking, often due to the poor preservation status of the carcasses at necropsy or to the peracute neurotoxic death, although even fresher carcasses may lack pathologic findings attributable to PbTx (Mase et al., 2000; Fauquier and Landsberg, 2018).
The pathogenic mechanisms of brevetoxins are not fully elucidated (Twiner et al., 2012). Brevetoxins act on voltage-gated sodium channels (Catterall and Gainer, 1985) by binding to neurotoxin receptor site 5, resulting in persistent channel activation (Jeglitsch et al., 1998). This action is particularly prominent in parasympathetic nerves and adrenergic nerve fibers. Bronchospasm ensues when the toxin is inhaled due to release of acetylcholine in postganglionic parasympathetic nerve endings (Smart, 1995). Studies suggest these toxins inhibit neurotransmission to skeletal muscles, yet paralysis has not been documented (Smart, 1995). Also, brevetoxins are potent cardiac toxins (Kirkpatrick et al., 2004). Furthermore, cutaneous exposure to brevetoxins in aerosol may induce irritation, cough, shortness of breath and tearing; ingestion of brevetoxins may result in gastrointestinal and peripheral and central nervous system injury. In our study, it is possible that a presumptive K. brevis-hemolytic toxin (Tatters et al., 2010) could have played a role; we cannot further evaluate this hypothesis because we did not investigate this specific hemolytic toxin. Interestingly, C. novyi was identified in a subset of animals. While C. novyi subtypes may produce several toxins and may be associated with necrotizing and hemorrhagic lesions in muscle and other organs in terrestrial mammals (Uzal et al., 2016), its pathogenic role remains unknown in marine mammals (Li et al., 2019). Typing and toxinotyping analyses were not conducted on these isolates, thus, further conclusions cannot be drawn. It is possible that autolysis and decomposition phenomena (carcasses were probably sunken for several days and then resurfaced) have favored this probable commensal or environmental bacterium to appear in various tissues. Noteworthy, we detected a still birth that could have potentially resulted from abortion secondary to brevetoxicosis. Brevetoxins have been detected in fetal and neonate dolphins, providing evidence of maternal transfer of the toxin in wild populations (Fire et al., 2015).
Diagnosis of brevetoxins may rely on enzyme linked immunosorbent assay, radioimmunoassay, electrochemiluminescence essays, electrochemical immunoanalysis, electrochemical impedance spectroscopy, spectroscopic ellipsometry, liquid chromatography-mass spectrometry, and immunohistochemistry (Bossart et al., 1998). In this study, PbTx2 and PbTx3 were identified and confirmed in gastric contents of 3/4 and 4/4 animals. Studies have reported a wide range of PbTx in bottlenose dolphin gastric contents (174–6,235 ng/g) and livers (37–278 ng/g) (Flewelling et al., 2005; Fire et al., 2008b,2015; Flewelling, 2008; Twiner et al., 2012), however, the lethal dose remains elusive (Hort et al., 2021). Comparatively, experimental studies in mice have shown a median lethal dose value (LD50) of 520 mg/kg bodyweight for PbTx-3 and 66,000 mg/kg bodyweight for PbTx-2 (Baden and Mende, 1982). Furthermore, in humans, it is estimated that an exposure of 2–3 μg/kg bodyweight for PbTx-3 may produce non-lethal toxic effects (Cunningham et al., 2021). While PbTx have been detected and quantified in various organic matrices,
gastric content is regarded as the most reliable matrix for acute brevetoxin exposure determination, yet other matrices allow for diagnosis at lower sensitivities, e.g., feces, urine, liver (Fire et al., 2015). Unfortunately, no quantification analysis was conducted on these samples, therefore, conclusions on this regard cannot be drawn.
Transmission is via inhalation (Bossart et al., 1998) or ingestion (Flewelling et al., 2005; Capper et al., 2013). Toxins remain stable in the environment and within seagrass, and mass mortalities can be seen long after and/or remotely located from a known algal bloom (Flewelling et al., 2005; Capper et al., 2013). PbTx are released upon algal cell lysis and can be aerosolized in the surf zone, by waves, or by wind, and present a risk of exposure by inhalation (Pierce, 1986). Finfish species (Fire et al., 2008a; Twiner et al., 2011) and certain types of seagrass can accumulate or vector brevetoxins and play a primary role in brevetoxin-induced marine mammal mortality events or strandings (Flewelling et al., 2005). In this study, most of the animals presented undigested prey items (in agreement with previous observations) within the gastric compartments wherein salema porgy was overrepresented. The latter is a herbivorous teleost fish that preferentially feeds on the seagrass Posidonia oceanica through the year (Peirano et al., 2001; Prado et al., 2008) and is used for human consumption. It is found over rocky substrates and sandy area with algal growth. Young mainly carnivorous on crustaceans, adults almost exclusively herbivorous, feed on seaweeds. Interestingly, consumption of this fish is inadvisable in certain periods of the year (spring and summer) because of ciguatoxins (Raikhlin-Eisenkraft et al., 1988; Chevaldonne, 1990). As in previous reports (Fire et al., 2021), our data suggest dietary exposure rather than inhalation of brevetoxin was the primary route of intoxication for these cetaceans.
Oceanographic investigations (data not shown) including biogeochemical parameters, surface water temperature and primary production analyses did not identify HAB or specific singularities, except for mildly increased chlorophyll levels. No phytoplankton data from the water body were available for the study period. It is believed that satellite (two)-resolution limitations (1 km2) hampered detection of HAB in this event (González-Ramos, personal communication). There are records of Karenia and other dinoflagellates in neritic and oceanic waters off central eastern Atlantic Ocean, including Canarian waters since 2004 (Ojeda, 2005, 2013), likely conditioned by global climate change, with a slow and progressive warming of the waters that has led to the settlement of species from warmer areas.
In summary, we documented the first cetacean mass-mortality event linked to brevetoxicosis in the eastern Atlantic Ocean. Epidemiologic data, including strandings or retrieval dates as well as carcass decomposition clearly indicated the cluster of deaths occurred at around the same time and area. Pathologic examinations, although limited by carcass preservation status, demonstrated consistent multisystemic hemorrhage and undigested or partially digested prey items, predominantly salema porgy. Toxicologic analyses of gastric contents detected PbTx2 and PbTx3. No other biotoxin related mortality has been recorded in the Canary Islands since this event. Brevetoxins should be considered in the differential diagnosis of cetacean mortality in the eastern Atlantic Ocean. These results raise concern for public health. Therefore, future monitoring for this type of events throughout the stranding programs or observation networks in Europe and/or partnership with the Global Ecology and Oceanography of Harmful Algal Blooms (GEOHAB) would be of great value.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Ethics Statement
Ethical review and approval was not required for the animal study because ethical review and approval were waived for this study, as neither animals were sacrificed nor experiments were performed with live animals. The permission for the management of stranded cetaceans was issued by the Spanish Ministry of Environment.
Author Contributions
AF, ES, MAO, YB, and JDD contributed to the conception and design of the study. AF, ES, MAO, and YB performed the necropsy examination. AF, ES, MAO, and JDD conducted histopathologic examinations. AG-M, JL, NG-Á, and MAI conducted chemical analyses. ES conducted virologic analysis. AV conducted bacteriological investigations. AF and JDD wrote the first draft of the manuscript. All authors contributed to manuscript revision and read and approved the submitted version.
Funding
Part of this research work was supported through the Research National Project “Embolic Pathology (gas/fat) in cetaceans,” which was supported by the Ministerio de Ciencia e Innovación del Gobierno de España (CGL2015-71498-P), Ministerio de Ciencia, Innovación y Universidades (PGC2018-101226-B-100), also MARCET (MAC/1.1b/149) and MARCET II (MAC2/4.6c/392) Projects and the Canary Islands Government, which was funded and provided support to the stranding network.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We would like to thank Teresa Rowles for constructive criticism and suggestions of this manuscript. Furthermore, we thank the collaboration of V. Martin and M. Tejedor (SECAC), A. J. Gonzalez and J. Coca (ULPGC), and Banco Nacional de Algas (ULPGC).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2022.834051/full#supplementary-material
References
Arbelo, M., Los Monteros, A. E., Herraez, P., Andrada, M., Sierra, E., Rodriguez, F., et al. (2013). Pathology and causes of death of stranded cetaceans in the Canary Islands (1999-2005). Dis. Aquat. Organ. 103, 87–99. doi: 10.3354/dao02558
Arregui, M., Fernandez, A., Paz-Sanchez, Y., Santana, A., Sacchini, S., Sierra, E., et al. (2020). Comparison of Three Histological Techniques for Fat Emboli Detection in Lung Cetacean’s Tissue. Sci. Rep. 10:8251. doi: 10.1038/s41598-020-64821-8
Baden, D. G., and Mende, T. J. (1982). Toxicity of two toxins from the Florida red tide marine dinoflagellate. Ptychodiscus Brevis Toxicon 20, 457–461. doi: 10.1016/0041-0101(82)90009-5
Bossart, G. D., Baden, D. G., Ewing, R. Y., Roberts, B., and Wright, S. D. (1998). Brevetoxicosis in manatees (Trichechus manatus latirostris) from the 1996 epizootic: gross, histologic, and immunohistochemical features. Toxicol. Pathol. 26, 276–282. doi: 10.1177/019262339802600214
Brodie, E. C., Gulland, F. M., Greig, D. J., Hunter, M., Jaakola, J., Leger, J. S., et al. (2006). Domoic acid causes reproductive failure in California sea lions (Zalophus californianus). Mar. Mamm. Sci. 22, 700–707. doi: 10.1111/j.1748-7692.2006.00045.x
Buttke, D. E., Walker, A., Huang, I. S., Flewelling, L., Lankton, J., Ballmann, A. E., et al. (2018). Green tree frog (Hyla cinerea) and ground squirrel (Xerospermophilus spilosoma) mortality attributed to inland brevetoxin transport at Padre Island National Seashore. Texas, USA, 2015. J. Wildl. Dis. 54, 142–146. doi: 10.7589/2017-01-018
Capper, A., Flewelling, L. J., and Arthur, K. (2013). Dietary exposure to harmful algal bloom (HAB) toxins in the endangered manatee (Trichechus manatus latirostris) and green sea turtle (Chelonia mydas) in Florida. U.S.A. Harmful Algae 28, 1–9.
Caruana, A. M. N., and Amzil, Z. (2018). “Microalgae and Toxins,” in Microalgae in health and disease prevention, eds I. A. Levine and J. Fleurence (. Cambridge, MA: Academic Press), 263–305.
Castle, K. T., Flewelling, L. J., Bryan, J. II, Kramer, A., Lindsay, J., Nevada, C., et al. (2013). Coyote (Canis latrans) and domestic dog (Canis familiaris) mortality and morbidity due to a Karenia brevis red tide in the Gulf of Mexico. J. Wildl. Dis. 49, 955–964. doi: 10.7589/2012-11-299
Catterall, W. A., and Gainer, M. (1985). Interaction of brevetoxin A with a new receptor site on the sodium channel. Toxicon 23, 497–504. doi: 10.1016/0041-0101(85)90034-0
Chevaldonne, P. (1990). Ciguatera and the saupe, Sarpa salpa (L.), in the Mediterranean: a possible misinterpretation. J. Fish Biol. 37, 503–504.
Cunningham, B. R., Coleman, R. M., Schaefer, A. M., Hamelin, E. I., and Johnson, R. C. (2021). Detection of brevetoxin in human plasma by ELISA. J. Anal. Toxicol. 30:bkab010. doi: 10.1093/jat/bkab010
Diaz-Delgado, J., Fernandez, A., Sierra, E., Sacchini, S., Andrada, M., Vela, A. I., et al. (2018). Pathologic findings and causes of death of stranded cetaceans in the Canary Islands (2006-2012). PLoS One 13:e0204444. doi: 10.1371/journal.pone.0204444
Fauquier, D., and Landsberg, J. H. (2018). “Harmful algae and biotoxins,” in CRC handbook of marine mammal medicine, eds F. M. D. Gulland, L. A. Dierauf, and K. L. Whitman (Boca Raton: CRC Press), 319–328. doi: 10.1016/j.aquatox.2018.03.004
Fire, S. E., Flewelling, L. J., Naar, J., Twiner, M. J., Henry, M. S., Pierce, R. H., et al. (2008a). Prevalence of brevetoxins in prey fish of bottlenose dolphins in Sarasota Bay. Florida Mar. Ecol. Prog. Ser. 368, 283–294.
Fire, S. E., Flewelling, L. J., Wang, Z., Naar, J., Henry, M. S., Pierce, R. H., et al. (2008b). Florida red tide and brevetoxins: association and exposure in live resident bottlenose dolphins (Tursiops truncatus) in the eastern Gulf of Mexico. U.S.A. Mar. Mamm. Sci. 24, 831–844.
Fire, S. E., Flewelling, L. J., Stolen, M., Durden, W. N., De Wit, M., Spellman, A. C., et al. (2015). Brevetoxin-associated mass mortality event of bottlenose dolphins and manatees along the east coast of Florida. U.S.A. Mar. Ecol. Prog. Ser. 526, 241–251. doi: 10.3354/meps11225
Fire, S. E., Miller, G. A., Sabater, E. R., and Wells, R. S. (2021). Utility of red tide (Karenia brevis) monitoring data as a predictive tool to estimate brevetoxin accumulation in live, free-ranging marine mammals. Front. Mar. Sci. 8:284. doi: 10.3389/fmars.2021.611310
Fire, S. E., Wang, Z. H., Byrd, M., Whitehead, H. R., Paternoster, J., and Morton, S. L. (2011). Co-occurrence of multiple classes of harmful algal toxins in bottlenose dolphins (Tursiops truncatus) stranding during an unusual mortality event in Texas. U.S.A. Harmful Algae 10, 330–336. doi: 10.1016/j.hal.2010.12.001
Flewelling, L. J. (2008). Vectors of Brevetoxins to Marine Mammals. Tampa, FL: University of South Florida. [Ph.D.thesis].
Flewelling, L. J., Naar, J. P., Abbott, J. P., Baden, D. G., Barros, N. B., Bossart, G. D., et al. (2005). Brevetoxicosis: red tides and marine mammal mortalities. Nature 435, 755–756. doi: 10.1038/nature435755a
Gaydos, J. K. (2006). Bottlenose Dolphins and Brevetoxins: a Coordinated Research and Response Plan. NOAA Tech. Memo. NMFS OPR 32:46.
Gebhard, E., Levin, M., Bogomolni, A., and De Guise, S. (2015). Immunomodulatory effects of brevetoxin (PbTx-3) upon in vitro exposure in bottlenose dolphins (Tursiops truncatus). Harmful Algae 44, 54–62. doi: 10.1016/j.hal.2015.02.010
Geraci, J. R., Anderson, D. M., Timperi, R. J., St.Aubin, D. J., Early, G. A., Prescott, J. H., et al. (1989). Humpback whales (Megaptera novaeangliae) fatally poisoned by dinoflagellate toxin. Can. J. Fish. Aquat. Sci. 46, 1895–1898.
Geraci, J. R., and Lounsbury, V. J. (2005). Marine Mammals Ashore: a Field Guide for Strandings. Baltimore: National Aquarium in Baltimore.
Gerssen, A., and Gago-Martinez, A. (2019). Emerging Marine Biotoxins. Toxins 11:314. doi: 10.3390/toxins11060314
Goldstein, T., Mazet, J. A., Zabka, T. S., Langlois, G., Colegrove, K. M., Silver, M., et al. (2008). Novel symptomatology and changing epidemiology of domoic acid toxicosis in California sea lions (Zalophus californianus): an increasing risk to marine mammal health. Proc. Biol. Sci. 275, 267–276. doi: 10.1098/rspb.2007.1221
Groch, K. R., Taniwaki, S. A., Favero, C. M., Brandao, P. E., Diaz-Delgado, J., Fernandez, A., et al. (2020). A novel real-time PCR to detect Cetacean morbillivirus in Atlantic cetaceans. J. Virol. Methods 285:113964. doi: 10.1016/j.jviromet.2020.113964
Gunter, G., Williams, R. H., Davis, C. C., and Smith, F. W. (1948). Catastrophic mass mortality of marine animals and coincident phytoplankton bloom on the west coast of Florida, November 1946 to August 1947. Ecol. Monogr. 18, 309–324.
Haywood, A. J., Steidinger, K. A., Truby, E. W., Bergquist, P. R., Bergquist, P. L., Adamson, J., et al. (2004). Comparative morphology and moleculr phylogenetic analysis of three new species of the genus Karenia (Dinophyceae) from New Zealand. J. Phycol. 40, 165–179. doi: 10.1111/j.0022-3646.2004.02-149.x
Hort, V., Abadie, E., Arnich, N., Dechraoui-Bottein, M. Y., and Amzil, Z. (2021). Chemodiversity of brevetoxins and other potentially toxic metabolites produced by Karenia spp. and their metabolic products in marine organisms. Mar. Drugs 19:656. doi: 10.3390/md19120656
Jeglitsch, G., Rein, K., Baden, D. G., and Adams, D. J. (1998). Brevetoxin-3 (PbTx-3) and its derivatives modulate single tetrodotoxin-sensitive sodium channels in rat sensory neurons. J. Pharmacol. Exp. Ther. 284, 516–525.
Kirkpatrick, B., Fleming, L. E., Squicciarini, D., Backer, L. C., Clark, R., Abraham, W., et al. (2004). Literature review of Florida red tide: implications for human health effects. Harmful Algae 3, 99–115. doi: 10.1016/j.hal.2003.08.005
Li, C., Tan, X., Bai, J., Xu, Q., Liu, S., Guo, W., et al. (2019). A survey of the sperm whale (Physeter catodon) commensal microbiome. PeerJ. 7:e7257. doi: 10.7717/peerj.7257
Litz, J. A., Baran, M. A., Bowen-Stevens, S. R., Carmichael, R. H., Colegrove, K. M., Garrison, L. P., et al. (2014). Review of historical unusual mortality events (UMEs) in the Gulf of Mexico (1990-2009): providing context for the multi-year northern Gulf of Mexico cetacean UME declared in 2010. Dis. Aquat. Organ. 112, 161–175. doi: 10.3354/dao02807
Lowenstine, L. J. (2007). “Algal bloom toxicity in marine animals,” in Zoo and Wild Animal Medicine: Current Therapy, eds M. E. Fowler and R. E. Miller (St. Louis, Missouri: Saunders), 341–348. doi: 10.1016/b978-141604047-7.50044-0
Mase, B., Jones, W., Ewing, R., Bossart, G., Vandolah, F., Leighfield, T., et al. (2000). “Epizootic in Bottlenose Dolphins in the Florida Panhandle: 1999-2000. Annual Conference American Association of Zoo Veterinarians”. in Proceedings of the American Association of Zoo Veterans and International Association for Aquatic Animal Medicine eds CK Baer, New Orleans, LA: American Association of Zoo Veterinarians, 522–524.
Mcnabb, P. S., Selwood, A. I., Van Ginkel, R., Boundy, M., and Holland, P. T. (2012). Determination of brevetoxins in shellfish by LC/MS/MS: single-laboratory validation. J. AOAC Int. 95, 1097–1105. doi: 10.5740/jaoacint.11-272
Ojeda, A. (2005). Dinoflagelados de Canarias estudio taxonómico y ecológico. Instituto Estud. Canarios 65:33.
Ojeda, A. (2013). Presencia del género Karenia y nuevos registros de dinoflagelados (Dinoflagellata) en aguas de las Islas Canarias. Instituto Canario Cienc. Mar. 15:37.
Peirano, A., Niccolai, I., Mauro, R., and Bianchi, C. N. (2001). Seasonal grazing and food preference of herbivores in a Posidonia oceanica meadow. Sci. Mar. 65, 367–374. doi: 10.3989/scimar.2001.65n4367
Pierce, R. H. (1986). Red tide (Ptychodiscus brevis) toxin aerosols: a review. Toxicon 24, 955–965. doi: 10.1016/0041-0101(86)90001-2
Prado, P., Farina, S., Tomas, F., Romero, J., and Alcoverro, T. (2008). Marine protection and meadow size alter fish herbivory in seagrass ecosystems. Mar. Ecol. Prog. Ser. 371, 11–21. doi: 10.3354/meps07662
Raikhlin-Eisenkraft, B., Finkelstein, Y., and Spanier, E. (1988). Ciguatera-like poisoning in the Mediterranean. Vet. Hum. Toxicol. 30, 582–583.
Reidarson, T. H., Mcbain, J., House, C., King, D. P., Stott, J. L., Krafft, A., et al. (1998). Morbillivirus infection in stranded common dolphins from the Pacific Ocean. J. Wildl. Dis. 34, 771–776. doi: 10.7589/0090-3558-34.4.771
Sacristan, C., Carballo, M., Munoz, M. J., Belliere, E. N., Neves, E., Nogal, V., et al. (2015). Diagnosis of Cetacean morbillivirus: a sensitive one step real time RT fast-PCR method based on SYBR(R) Green. J. Virol. Meth. 226, 25–30. doi: 10.1016/j.jviromet.2015.10.002
Scholin, C. A., Gulland, F., Doucette, G. J., Benson, S., Busman, M., Chavez, F. P., et al. (2000). Mortality of sea lions along the central California coast linked to a toxic diatom bloom. Nature 403, 80–84. doi: 10.1038/47481
Sierra, E., Sanchez, S., Saliki, J. T., Blas-Machado, U., Arbelo, M., Zucca, D., et al. (2014). Retrospective study of etiologic agents associated with nonsuppurative meningoencephalitis in stranded cetaceans in the canary islands. J. Clin. Microbiol. 52, 2390–2397. doi: 10.1128/JCM.02906-13
Smart, D. (1995). “Clinical toxicology of shellfish poisoning,” in Handbook of Clinical Toxicology of Animal Venoms and Poisons, eds J. White and J. Meier (Boca Raton, Florida: CRC Press).
Tatters, A. O., Muhlstein, H. I., and Tomas, C. R. (2010). The hemolytic activity of Karenia selliformis and two clones of Karenia brevis throughout a growth cycle. J. Appl. Phycol. 22, 435–442. doi: 10.1007/s10811-009-9476-z
Twiner, M. J., Fire, S., Schwacke, L., Davidson, L., Wang, Z., Morton, S., et al. (2011). Concurrent exposure of bottlenose dolphins (Tursiops truncatus) to multiple algal toxins in Sarasota Bay, Florida, USA. PLoS One 6:e17394. doi: 10.1371/journal.pone.0017394
Twiner, M. J., Flewelling, L. J., Fire, S. E., Bowen-Stevens, S. R., Gaydos, J. K., Johnson, C. K., et al. (2012). Comparative analysis of three brevetoxin-associated bottlenose dolphin (Tursiops truncatus) mortality events in the Florida Panhandle region (USA). PLoS One 7:e42974. doi: 10.1371/journal.pone.0042974
Uzal, F. A., Songer, J. G., Prescott, J. F., and Popoff, M. R. (2016). Clostridial Diseases of Animals. New Jersey: John Wiley & Sons.
Van Dolah, F. M. (2005). “Effects of harmful algal blooms,” in Marine Mammal Research: Conservation Beyond Crisis, eds J. E. Reynolds, W. F. Perrin, R. R. Reeves, S. Montgomery, and T. J. Ragen (Baltimore, MD: Johns Hopkins University Press), 85–101.
VanDevanter, D. R., Warrener, P., Bennett, L., Schultz, E. R., Coulter, S., Garber, R. L., et al. (1996). Detection and analysis of diverse herpesviral species by consensus primer PCR. J. Clin. Microbiol. 34, 1666–1671. doi: 10.1128/JCM.34.7.1666-1671.1996
Keywords: biotoxin, Karenia brevis, Harmful algal bloom, climate change, PbTx
Citation: Fernández A, Sierra E, Arbelo M, Gago-Martínez A, Leao Martins JM, García-Álvarez N, Bernaldo de Quiros Y, Arregui M, Vela AI and Díaz-Delgado J (2022) First Case of Brevetoxicosis Linked to Rough-Toothed Dolphin (Steno bredanensis) Mass-Mortality Event in Eastern Central Atlantic Ocean: A Climate Change Effect? Front. Mar. Sci. 9:834051. doi: 10.3389/fmars.2022.834051
Received: 12 December 2021; Accepted: 07 February 2022;
Published: 28 February 2022.
Edited by:
Jeremy Kiszka, Florida International University, United StatesReviewed by:
Annalisa Zaccaroni, University of Bologna, ItalyShauna Murray, University of Technology Sydney, Australia
Copyright © 2022 Fernández, Sierra, Arbelo, Gago-Martínez, Leao Martins, García-Álvarez, Bernaldo de Quiros, Arregui, Vela and Díaz-Delgado. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Eva Sierra, ZXZhLnNpZXJyYUB1bHBnYy5lcw==