 Meghan G. Aulich1*
Meghan G. Aulich1* Robert D. McCauley1
Robert D. McCauley1 Brian S. Miller2
Brian S. Miller2 Flore Samaran3
Flore Samaran3 Giacomo Giorli4
Giacomo Giorli4 Benjamin J. Saunders5
Benjamin J. Saunders5 Christine Erbe1
Christine Erbe1- 1Curtin University, Centre for Marine Science and Technology, Bentley, WA, Australia
- 2Australian Antarctic Division, Kingston, TAS, Australia
- 3Lab-STICC CNRS UMR 6285, ENSTA Bretagne, Brest, France
- 4National Institute of Water and Atmospheric Research (NIWA), Wellington, New Zealand
- 5Curtin University, School of Molecular and Life Sciences, Bentley, WA, Australia
The fin whale is listed as globally vulnerable, with ongoing threats to their population, yet little is known about the distribution and movements of the Southern Hemisphere sub-species, Balaenoptera physalus quoyi. This study assesses fin whale distribution in the Southern Hemisphere analysing acoustic recordings from 15 locations in Antarctic and Australian waters from 2002 to 2019. A seasonal acoustic presence of fin whales in Antarctic waters from late austral summer to autumn (February to June) with long-term, consistent annual usage areas was identified at the Southern Kerguelen Plateau and Dumont d’Urville sites. In comparison, limited vocal presence of fin whales was observed at the Casey site. In Australian waters, fin whales were seasonally present from austral autumn to mid-spring (May to October) on east and west coasts, with a decadal pattern of acoustic presence observed at Cape Leeuwin, WA. Two migratory pathways are identified, from the Indian sector of Antarctica to the west coast of Australia and from the Pacific sector of Antarctica to the east coast of Australia. The identified seasonal distributions and migratory pathways provide valuable information to aid in monitoring the recovery of this vulnerable sub-species. We suggest the identified distribution and dispersal from the Southern Kerguelen Plateau and Dumont d’Urville sites to the west and east coasts of Australia respectively, as well as the spatial separation between Antarctic sites, provide preliminary evidence of separate sub-populations of the Southern Hemisphere sub-species of fin whale.
Introduction
The Southern Hemisphere sub-species of fin whale (Balaenoptera physalus quoyi) is listed as vulnerable on the IUCN red list (Cooke, 2018) after devastating population decline during the industrial whaling era, with approximately 700,000 fin whales caught (Rocha et al., 2014). This sub-species is under further ongoing threat of population decline due to climate change (Tulloch et al., 2019) and habitat disturbance (Castellote et al., 2012). Like other migratory baleen whale species, the fin whale is thought to occupy high-latitude, polar regions in the summer months and low-latitude, temperate regions in the winter months (Mizroch et al., 1984). Suggested reasons for this seasonal migration include the exploitation of highly productive feeding grounds in polar waters (Lockyer and Brown, 1981) and avoidance of increasing sea-ice (Širović et al., 2004; Simon et al., 2010; Delarue et al., 2013). Calving and breeding are also thought to take place in warmer, lower-latitude waters (Lockyer and Brown, 1981). Identifying the distribution and seasonal presence of the Southern Hemisphere sub-species of fin whale could provide novel insights into the sub-species’ ecology and be useful to aid in monitoring of this vulnerable species at a national and international level.
Passive Acoustic Monitoring (PAM) is a cost effective tool in identifying temporal distribution of cetacean species, as sampling can be conducted over long time periods, in non-optimal habitat and weather conditions such as those found in polar waters (Mellinger et al., 2007). The vocalisations of the fin whale are ideal for PAM as the animals produce highly stereotyped, repetitive sequences of calls (Watkins et al., 1987). The most widely identified and commonly used call type of the fin whale is referred to as the “20 Hz” pulse (Watkins et al., 1987). This 20 Hz call type is characterised by short ~1 s down-sweeping pulses (Watkins et al., 1987), ranging in frequency from 42 to 18 Hz (Watkins et al., 1987; Thompson et al., 1992) and are produced at repeated intervals every 7-26 seconds (Watkins et al., 1987). The fin whale produces bouts of these 20 Hz pulses which can last up to 32 hours (Watkins et al., 1987). Observed variation in the characteristics of the 20 Hz call (Watkins et al., 1987) aligning with the copulation stage of the fin whale (Lockyer and Brown, 1981) and observations of only male fin whales singing (Croll et al., 2002) have led to the hypothesis that this call type is a breeding display produced only by males (Watkins et al., 1987; Croll et al., 2002). Less commonly reported call types of the fin whale include the lower-frequency “back-beat” pulses (13 to 23 Hz) (Thompson et al., 1992; Brodie and Dunn, 2015) which accompany the 20 Hz pulse, 20 Hz pulses that contain a higher-frequency component (89 to 99 Hz) (Širović et al., 2009) and “40 Hz” pulses (Širović et al., 2013) which have also been referred to as higher-frequency “downsweeps” (Gedamke and Robinson, 2010).
Using PAM, the seasonal distribution of fin whales has been widely reported in the Northern Hemisphere over the last two decades. The literature outlines a seasonal migration of fin whales out of high-latitude, polar waters (Stafford et al., 2007; Simon et al., 2010; Delarue et al., 2013; Davis et al., 2020) to lower-latitude, temperate waters for the winter months along the east and west coasts of the North Atlantic (Morano et al., 2012; Harris et al., 2013; Davis et al., 2020) and North Pacific Oceans (Širović et al., 2013; Iwase, 2015). Acoustic studies of fin whales have also reported the presence of fin whales in the winter season in the Bering Sea (Stafford et al., 2007; Oleson et al., 2014; Širović et al., 2015) and in waters off Greenland (Simon et al., 2010), suggesting that not all fin whales may migrate. In addition, populations of non-migratory, resident fin whales have been reported in lower-latitude regions of the North Pacific and Atlantic Oceans (Morano et al., 2012; Širović et al., 2013).
In contrast, in the Southern Hemisphere, an ocean-wide picture of fin whale distribution and migration is not available in the literature. Historical whaling data are nearly a century old, and heavily biased by the behaviour of the fleet (de la Mare, 2014). Visual survey effort in the Southern Hemisphere consists primarily of the IDCR-SOWER circumpolar voyages, with most effort focused almost entirely on Jan-Feb, with heavy emphasis on covering areas near high-latitudes and the ice-edge, so do not provide a full picture (Edwards et al., 2015). Acoustic studies of fin whales in Antarctic waters were limited and largely focused on regions of the Western Antarctic Peninsula, where the animals were found to have a seasonal presence from February to July (Širović et al., 2004; Širović et al., 2007; Širović et al., 2009; Burkhardt et al., 2021). In other regions of Antarctica, identification of fin whale distribution is far more limited. In 2009 the Southern Ocean Research Partnership of the International Whaling Commission (IWC-SORP) commenced a working group to coordinate a long-term acoustic research program to better understand the distribution of this top predator throughout their feeding grounds (Van Opzeeland et al., 2014). While the early efforts of this project focused predominantly on Antarctic blue whales (Balaenoptera musculus), in recent years fin whales have started to become a major focus as well (Bell et al., 2019; Miller et al., 2021a). Vocalisations of the fin whale have been recorded at the Balleny Islands, in the Pacific Sector of Antarctica (Dziak et al., 2017), a low presence of fin whale calls has been observed in deep waters off East Antarctica, in the Indian sector (Širović et al., 2009) and no fin whale presence has been detected in the Ross Sea (Širović et al., 2009). At lower latitudes, a seasonal acoustic presence of fin whale calls has been observed in Australia from as early as April to October (Aulich et al., 2019) and in New Zealand from May to October (McDonald, 2006; Brodie and Dunn, 2015; Constaratas et al., 2021). Fin whale vocalisations have also been observed in regions of the Indian Ocean from April to November (Leroy et al., 2018; Dréo et al., 2019). Off South America, fin whales were acoustically detected in subtropical waters off Chile throughout the austral winter, while vocalisations were rare in summer (Buchan et al., 2019).
There remains a gap in the literature as to how these animals are distributed in Antarctic waters, and whether the low latitude observations are of animals that have migrated out of different areas of Antarctica. The aims of this study are to identify the long-term, broad-scale spatial distribution of the fin whale in regions of Antarctica and the seasonal migration of animals to Australian waters.
Methods
Underwater sound recordings were obtained from 15 locations off Antarctica and Australia between the years 2002 and 2019 (Table 1 and Figure 1) from a range of PAM systems:
1. Moored Acoustic Recorders (MARs) designed and manufactured by the Science Technical Support group of the Australian Antarctic Division (AAD) were deployed at five locations off Eastern Antarctica during the years 2013-2019 (Miller et al., 2021b). These datasets were collected under the International Whaling Commission’s Southern Ocean Research Partnership (IWC-SORP). The MARs had a sampling frequency of 12 kHz and a continuous, year-round recording scheme (duty cycle DC = 1).
2. Customised Underwater Sound Recorders (USRs) (McCauley et al., 2017) developed by Curtin University were deployed in collaboration with the AAD at three locations between Australia and Antarctica. USRs were deployed in the years 2005-2009, had a sampling frequency of 6 kHz and recorded for 13 minutes every hour (DC = 0.22) from start date of deployment to end date.
3. USRs of the Australian Integrated Marine Observing System (IMOS), deployed by Curtin University, recorded at five locations off Australia between 2009 and 2017, with a sampling frequency of 6 kHz and a duty cycle of five minutes every 15 minutes (DC = 0.33).
4. The Comprehensive Test Ban Treaty Organisation (CTBTO) nuclear test monitoring station operates a hydro-acoustic station, HA01, off Cape Leeuwin, WA, Australia. Recordings from this station were obtained from CTBTO through Geoscience Australia, the Australian operator of station HA01, for the years 2002-2011. The recordings had a sampling frequency of 250 Hz and were obtained continuously, year-round (DC = 1).
5. The New Zealand National Institute of Water and Atmospheric Research (NIWA) deployed two Autonomous Multichannel Acoustic Recorders (AMARs, JASCO Applied Sciences Pty Ltd) in the Ross Sea for the year 2018. The AMARs had a sampling frequency of 48 kHz and recorded for 5.7 minutes every 13.3 minutes (DC = 0.43).

Table 1 Deployment details of the passive acoustic monitoring systems.
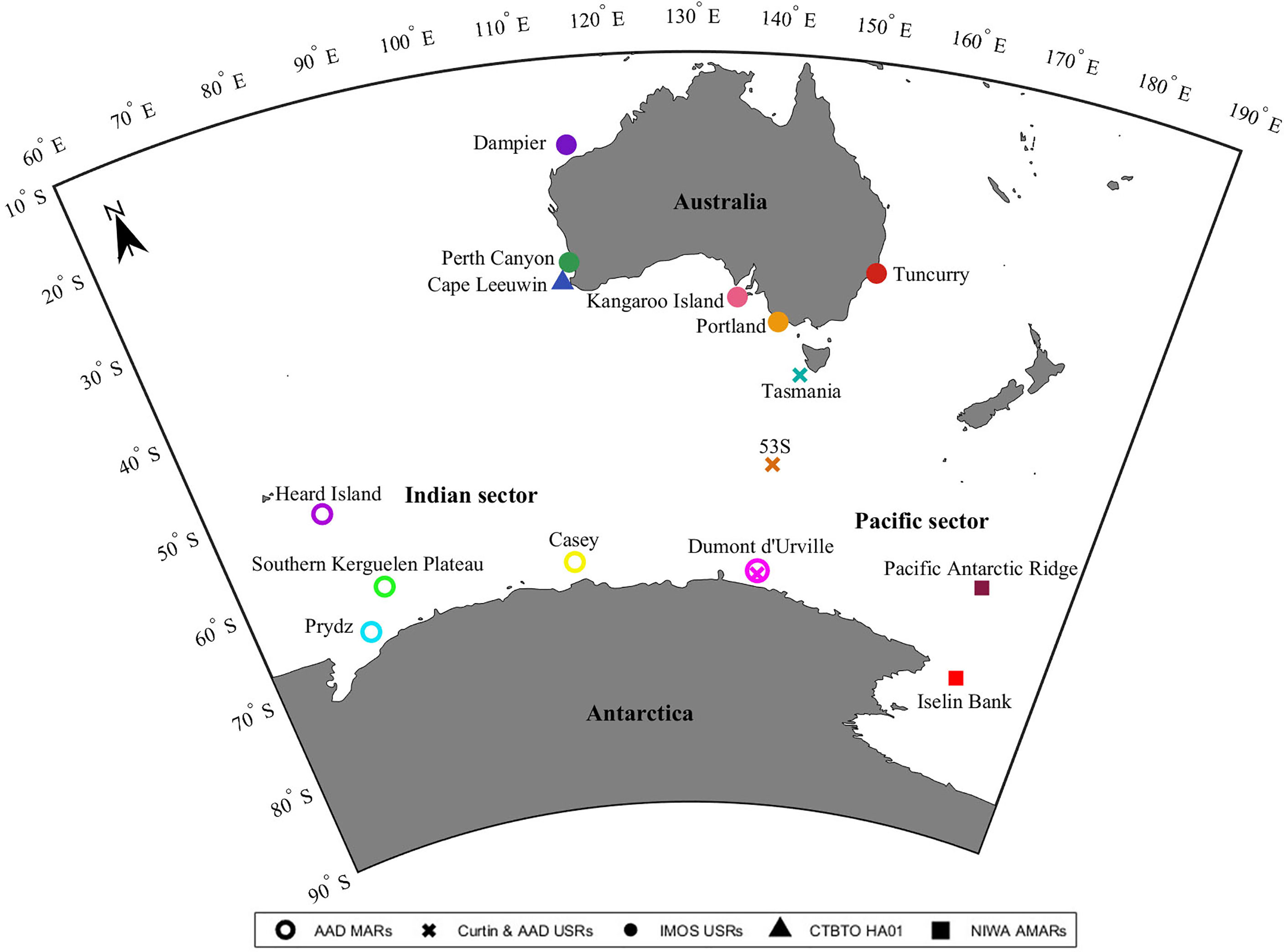
Figure 1 Deployment locations of the passive acoustic monitoring systems used to obtain underwater sounds. Colours indicate sites. Circles indicate Moored Acoustic Recorders (MARs) of the Australian Antarctic Division (AAD). Crosses indicate Customised Underwater Sound Recorders (USRs) of Curtin University and the AAD. Dots indicate USRs of the Australian Integrated Marine Observing System (IMOS). Triangles indicate the hydroacoustic station (HA01) of the Comprehensive Test Ban Treaty Organisation (CTBTO). Squares indicate Autonomous Multichannel Acoustic Recorders (AMARs) of the New Zealand National Institute of Water and Atmospheric Research (NIWA). Equidistant Conic Projection used.
A total of 284,992 hours of recording was collected from 2002 to 2019 across all 15 sites (Table 1). Due to the large quantity of acoustic recordings and the expected large number of fin whale pulses, a process of combined automatic and manual detection was implemented in order to detect all fin whale 20 Hz pulses at each sample site. A brief summary of this detection method is outlined below and more detail is provided in Aulich et al. (2019).
Two automatic detection algorithms were implemented in MATLAB (Version 2019b, The MathWorks Inc, Natick, MA, USA). The first detection algorithm was based on spectrogram cross-correlation (Mellinger and Clark, 2000), using a template of a pre-defined fin whale 20 Hz pulse. The algorithm was run across all recordings (sample lengths:1 hour for AAD recordings, 13 minutes for Curtin/AAD USR recordings, 5 minutes for IMOS USR recordings, 1 hour for CTBTO recordings and 5.7 minutes for NIWA recordings). Automated detections were saved with a time stamp (corresponding to maximum intensity within the pulse). The first algorithm was used to locate time periods (samples) that had fin whale 20 Hz pulses, but did not attempt to precisely identify all pulses. Once the first detection algorithm had been applied, the time periods that contained automated detections were displayed in spectrogram form (maximum display length 300 s) in order to manually verify the sample contained a fin whale 20 Hz pulse (correct detection; true positive) or did not contain a fin whale 20 Hz pulse (false alarm; false positive). Ambient noise in the fin whale call frequency band such as Antarctic blue whale calls and/or sea-ice noise may result in false positive detections. This process of manual verification was carried out across all time periods (samples) containing automated detections, ensuring these false positives were removed. In order to check surrounding time periods (samples) for missed detections (false negatives), a process of ‘bracketing’ was iterated, whereby three samples surrounding verified pulses were manually checked until all surrounding samples were found to not contain fin whale pulses.
Once this process of manual verification and bracketing had been completed, a second, more sensitive detection algorithm was run across all samples with verified fin whale 20 Hz pulses to detect every pulse. This detector was based on cross-correlation in the time domain. The template was the absolute value of the Hilbert transform of an undistorted fin whale pulse at high signal-to-noise ratio (i.e., the positive envelope of the waveform). The template was correlated across the absolute of the Hilbert transform of each recorded sample (after bandpass filtering the recording to the frequency band of interest, 8-45 Hz). The resulting time series of correlation coefficients was thresholded. For times with high correlation, spectral power ratios were computed between the fin whale in-band power (18-25 Hz) and out-of-band power (10-15 Hz and 35-40 Hz). If in-band power was >3 dB above out-of-band power (in each band below and above the fin whale frequency bands), then the detector reported a detection; otherwise the time of high correlation was ignored. This removed non-fin whale broadband pulses. Once this had been completed, samples containing detections were displayed in spectrogram form for final manual verification (accept or reject) and any missed detections were manually added.
In order to evaluate the efficacy of this combined automatic and manual detection process, we calculated the combined error rate of pulse detections (Mellinger and Clark, 2000). The combined error rate is defined as the sum of the false-negative and false-positive error rates (Mellinger and Clark, 2000).The Southern Kerguelen Plateau 2019 dataset (AAS 4102 long term acoustic recordings Kerguelen 2019) was chosen for this analysis as it had strong fin whale vocalisations amid strong confounding ambient noise. A randomised subsample of 200 audio recordings (1 hour each) were analysed and yielded a false positive rate of 0.27% and a false negative rate of 1.62%, with a combined error rate of 1.89%.
Fin whale calling activity was analysed by site and year (site-year), which required splitting some of the datasets (that recorded over New Year’s) into two years at the same site, and merging two datasets that recorded in the same year, albeit with a gap between deployments. The duration of the fin whale season was defined as the number of days between the first and last detection dates at any one site-year. As an indicator of occupancy, the number of days which had fin whale pulses (pulse-days) was divided by the number of days that the recorder was on (recording-days) for each month, season, and calendar year, and presented as a percentage. As an indicator of behaviour and site usage, fin whale pulse rate was computed as the number of pulses detected over a certain period of time (24 h, 1 month, 1 fin whale season and 1 year). To account for the different duty cycles, the number of pulses detected in 1 day (1 month, 1 fin whale season or 1 year) was divided by the cumulative number of recording-seconds during that period. Pulse rates were multiplied by 3600 s/h, yielding pulse rate with a unit of 1/h, which is more meaningful than 1/s, given fin whale pulse duration is >1 s. In addition, the number of pulses detected each month was normalised by DC to allow comparison of the ‘continuous’ number of pulses across sites, as if all recorders had been recording continuously (calculations accounted for different month lengths and leap years). In detail, the number of pulses detected within a month was divided by the recording-seconds that month and multiplied by the number of seconds within that calendar month. A Kruskal-Wallis statistical analysis and post hoc Dunn test using the Bonferroni correction were used to compare mean percentage of call days per year and mean pulse rates per year across Australian sites and across Antarctic sites to discern regional differences within these two areas.
Results
A total of 812,144 fin whale pulses were detected across all deployment site-years, with Iselin Bank the only site to record no fin whale 20 Hz vocalisations. Though quantifying the proportion of the different call types was beyond the scope of this work, we can report that the 20 Hz pulse was the most commonly observed fin whale call type at every site by a large margin (Figure 2). Other types of fin whale calls that were occasionally observed included the back-beat pulse and higher-frequency components, accompanying 20 Hz pulses (Figure 2).
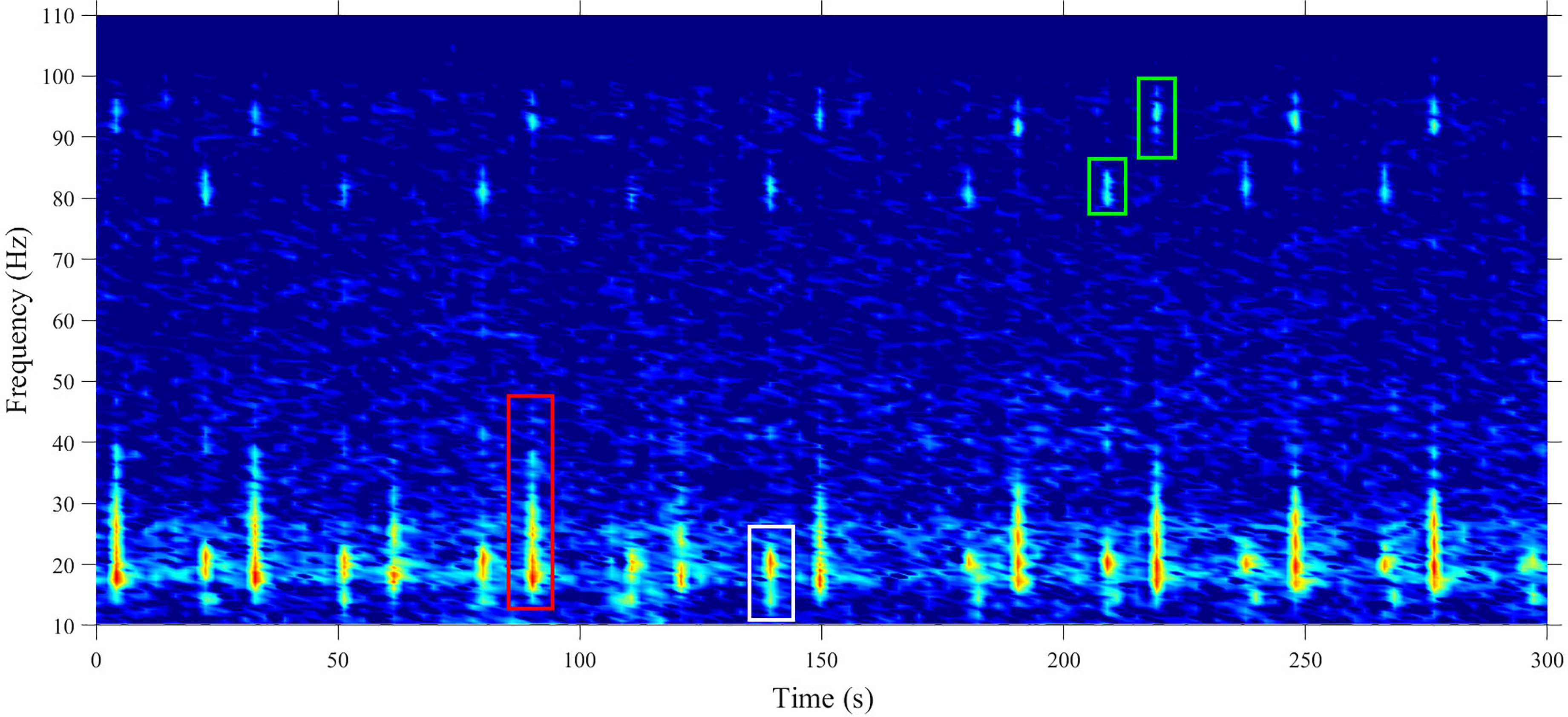
Figure 2 Spectrogram of example vocalisations of the fin whale. The 20 Hz pulse (red box), the back-beat pulse (white box) and the higher-frequency components accompanying both the back-beat and 20 Hz pulses (green boxes). Image was taken from Cape Leeuwin, (06-Aug-2011, 19:00). Spectrograms were calculated in 256-point Hann windows with 0.49 frequency resolution; sampling frequency 250 Hz.
In the Antarctic, fin whales are present early in the year (Feb-Jun) with the earliest detection of a vocalising fin whale occurring in the Indian sector, at Prydz in late January of 2013. At the Southern Kerguelen Plateau, a long-term seasonal pattern of vocal presence was identified from February to June (Figure 3 and Table 2). At the Casey site, an inconsistent pattern of vocal presence was identified over the five years of recording, with no detections in 2018. There was a much greater vocal presence of fin whales in 2019 at Casey, compared to other recording years (Figure 3 and Table 2). In the Pacific sector of Antarctica at Dumont d’Urville, a seasonal pattern of fin whale vocal presence was identified from early February to June (Figure 3 and Table 2). The Pacific Antarctic Ridge site recorded the latest vocalisation of a fin whale across all Antarctic sites, in August 2018 and thereby had the longest season out of all Antarctic sites.
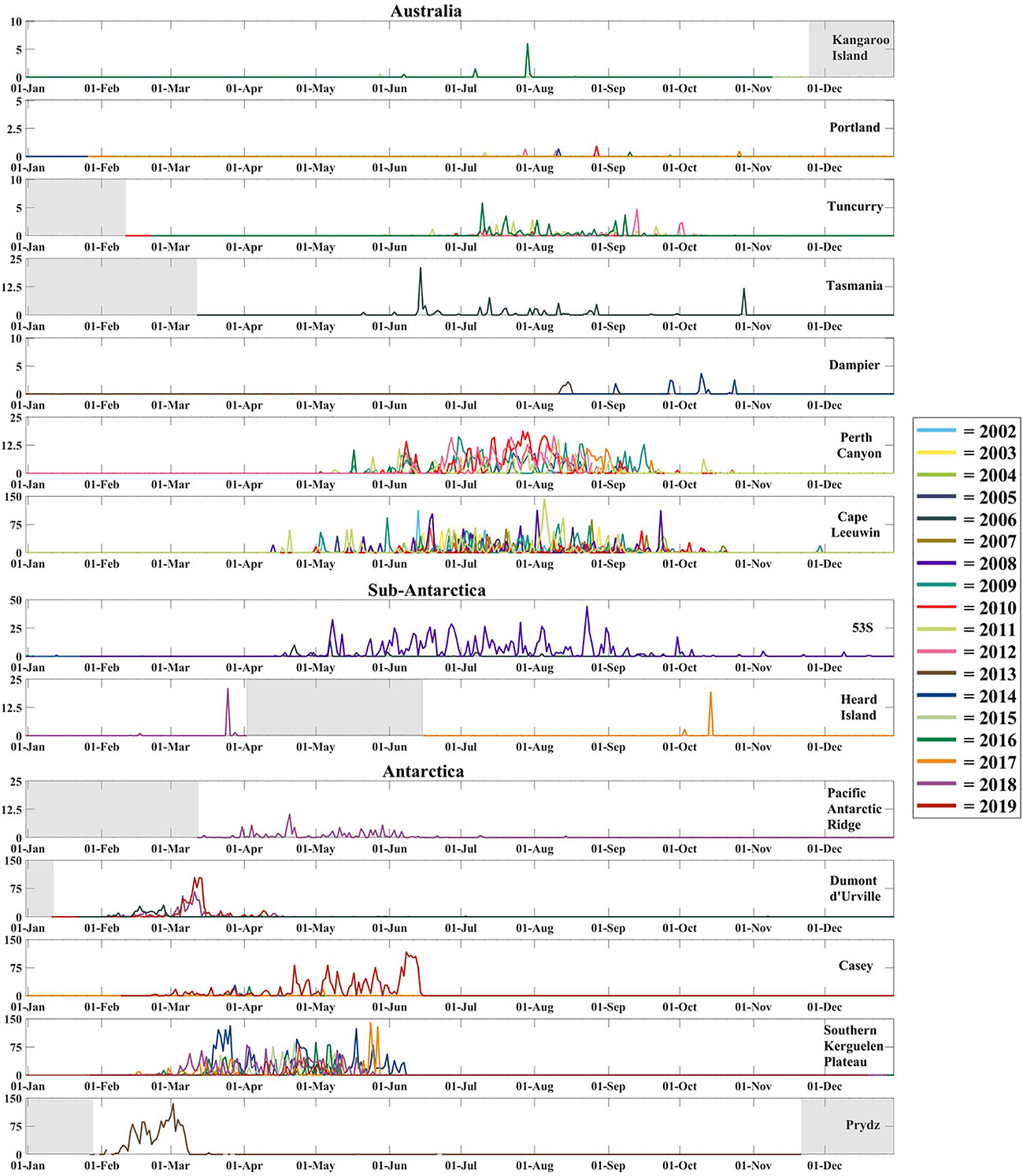
Figure 3 Line-graphs of the fin whale pulse rate [1/h] by day. I.e., number of pulses counted every 24 h, divided by the cumulative number of recording-seconds that day, × 3600 s/h) for all sites and years. Grey-shaded boxes indicate periods with no recording.
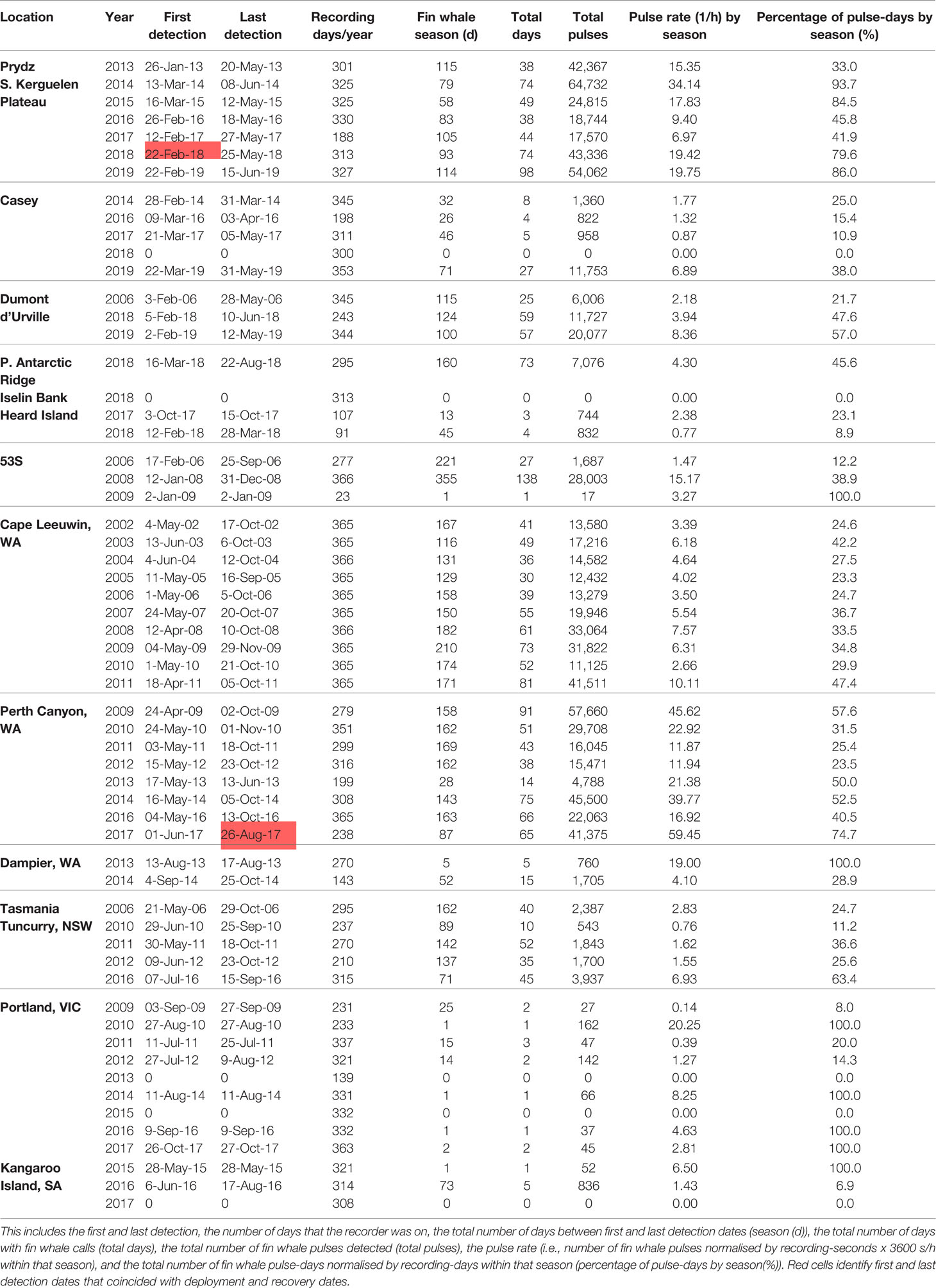
Table 2 Fin whale detection data for all recording site-years.
Sub-Antarctic waters exhibited two seasons: At Heard Island, seasons of vocal presence were very short, with vocalisations identified in October and from February to March, although recording was only conducted from September to April (Figure 3 and Table 2). In comparison, at the other Sub-Antarctic site, 53S, vocal presence was recorded throughout the year, with March being the only month without vocalisations. Therefore, this site had the longest season of 366 days over 2008 and 2009 (Figure 3 and Table 2). Limited vocal presence was detected in January, February and April at 53S, making detections harder to visualise in these months in Figure 3.
Around Australia, fin whale vocal presence occurs in the middle of the year (~May-Oct), with the earliest detection occurring on Australia’s west coast at Cape Leeuwin in April (Table 2). Whale vocalisations at this site were recorded over the decade from 2002 to 2011 with a seasonal presence from April to October with the latest presence in November of 2009 (Figure 3 and Table 2). Cape Leeuwin had the longest Australian season (210 days in 2009). Perth Canyon had long seasons (169 days in 2011) with a general pattern of fin whale vocal presence from May to October, over eight years of recording (Figure 3). Further north of the Perth Canyon, at Dampier, a much later vocal presence was identified from August to late October (Figure 3 and Table 2). On Australia’s east coast the earliest detection of a vocalising fin whale occurred at Tasmania in late May of 2006. A seasonal pattern of presence was identified at Tuncurry from June to October over four years of recording from 2010 to 2016 (Figure 3 and Table 2). On Australia’s south-east coast, an inconsistent and intermittent pattern of vocal presence was identified at Portland and Kangaroo Island, with some years with no vocalisations. Portland had the shortest season (1-25 days) with vocalisations recorded between July and October (Figure 3 and Table 2). In contrast, vocalisations were recorded earlier at Kangaroo Island from May to August with seasons varying in length between 1 and 73 days, depending on year.
The longest season does not imply the greatest pulse rate or pulse-days, as the Perth Canyon featured the greatest pulse rate by season, followed by Southern Kerguelen Plateau (Table 2). Cape Leeuwin has long seasons but lower pulse counts and fewer pulse-days. Pulse counts by season varied greatly at the Southern Kerguelen Plateau between site-years, declining to lowest pulse rates by season and pulse-days by season in 2017 (Table 2). Those sites with 100% pulse-days by season reflects seasons of one calling day (Table 2).
The vocal presence at each site, whether pulse rates or pulse-days varies between years but also on a monthly basis (Figures 4A, B). There are different patterns in pulse rates and percentage of pulse-days by month at Antarctic compared to Australian sites. Peak pulse rates and percentage of pulse-days occur in March and April at Prydz and Southern Kerguelen Plateau respectively. In contrast, peaks in Australia occur in July at the Perth Canyon. Both Antarctic and Australian sites have some overlap with the Sub-Antarctic sites. This is particularly evident at 53S which has a sudden increase in May and remains consistently high before declining in September and throughout the later months (Figures 4A, B).
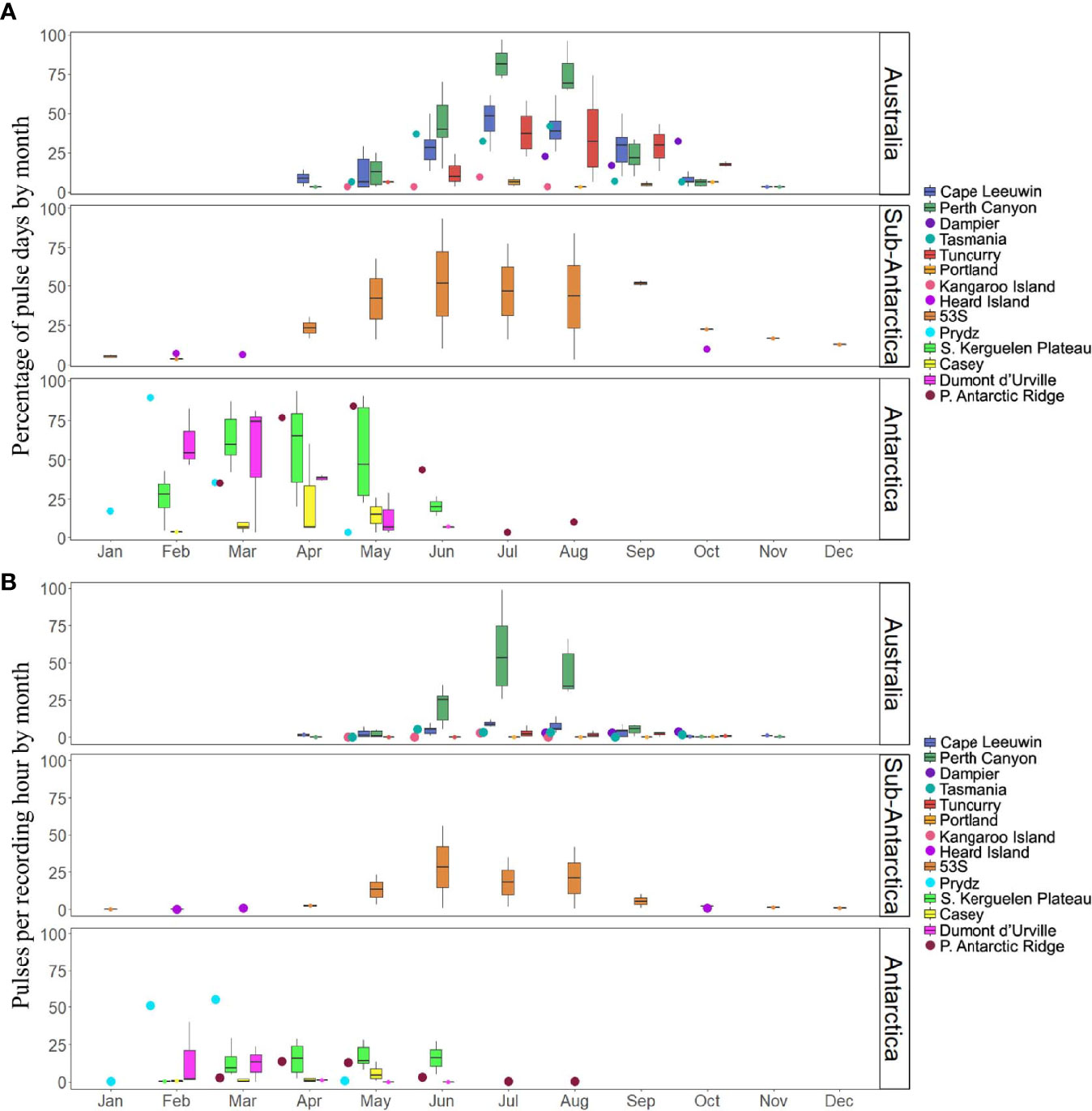
Figure 4 Monthly boxplots of fin whale acoustic presence. (A) The percentage of fin whale pulse-days to recording-days by month at each sample site, combined over sample years and (B) the fin whale pulse rate (i.e., pulses detected per recording-hour) by month at each sample site, combined over sample years. Boxplots display the median, interquartile range, minimum and maximum values. Scatter points display values at sites with only one recording year.
Off Antarctica, the Southern Kerguelen Plateau had the longest dataset (six years) and the greatest inter-annual variability in percentage of pulse-days by month (Figure 4A). Dumont d’Urville also had high inter-annual variability (Figure 4A). In the Sub-Antarctic region, 53S had great inter-annual variability (Figure 4A). In Australian waters, the greatest inter-annual variability in percentage of pulse-days by month occurred at Tuncurry in August (Figure 4A). Perth Canyon and Cape Leeuwin had comparatively less inter-annual variability (Figure 4A).
In Antarctic waters, inter-annual variability of pulse rates by month was greatest at Dumont d’Urville and the Southern Kerguelen Plateau (Figure 4B). In Sub-Antarctic waters, at 53S, inter-annual variability was higher than at the Antarctic sites (Figure 4B). In Australian waters, the Perth Canyon had by far the greatest inter-annual variability in pulse rates by month (Figure 4B).
Kruskal-Wallis and post-hoc Dunn’s tests indicated that most areas were statistically similar to one another in terms of the pulse rates and percentage of pulse-days across all Australian and Antarctic sites (Tables 3, 4). Off Antarctica, Southern Kerguelen Plateau had a greater pulse rate and greater percentage of pulse-days than Casey, while Dumont d’Urville was statistically similar to both of these locations in terms of pulse rate and pulse-days (Table 4 and Figures 4A, B). Off Australia, Perth Canyon, had a greater pulse rate than Kangaroo Island, Portland and Tuncurry (Table 3 and Figure 4B), and a greater percentage of pulse-days than Portland and Tuncurry (Table 3 and Figure 4A).
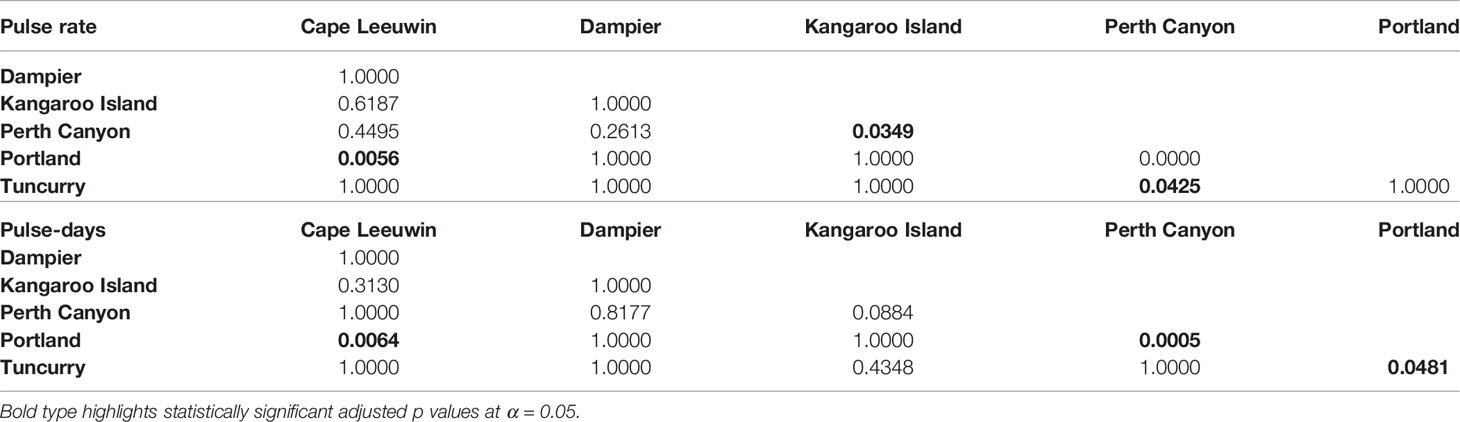
Table 3 Kruskal-Wallis and post hoc Dunn test comparisons for differences in pulse rates per year (KW Chi-squared=28.79, df=5 and p= >0.01) and pulse-days per year (KW Chi-squared=21.76, df = 5, p = >0.01) between Australian sites with p value displayed.
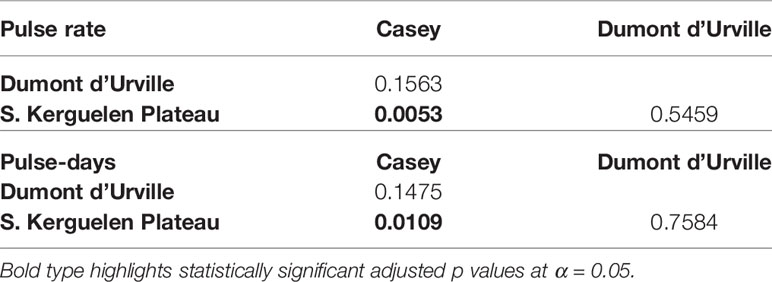
Table 4 Kruskal-Wallis and post hoc Dunn test comparisons for differences in pulse rates per year (KW Chi-squared=8.53, df = 2 and p = 0.01) and pulse-days per year (KW Chi-squared=7.32, df=2, p = 0.02) between Antarctic sites with p value displayed.
Figure 5 shows the monthly calling activity of fin whales at each site on a chart. Fin whale migration between sites is thus easily visualised and compared. Fin whales that are present in the Indian sector of Antarctica at the beginning of the year likely migrate to Western Australia later in the year, while fin whales present in the Pacific sector of Antarctica likely migrate to south-eastern Australia (Figure 5).
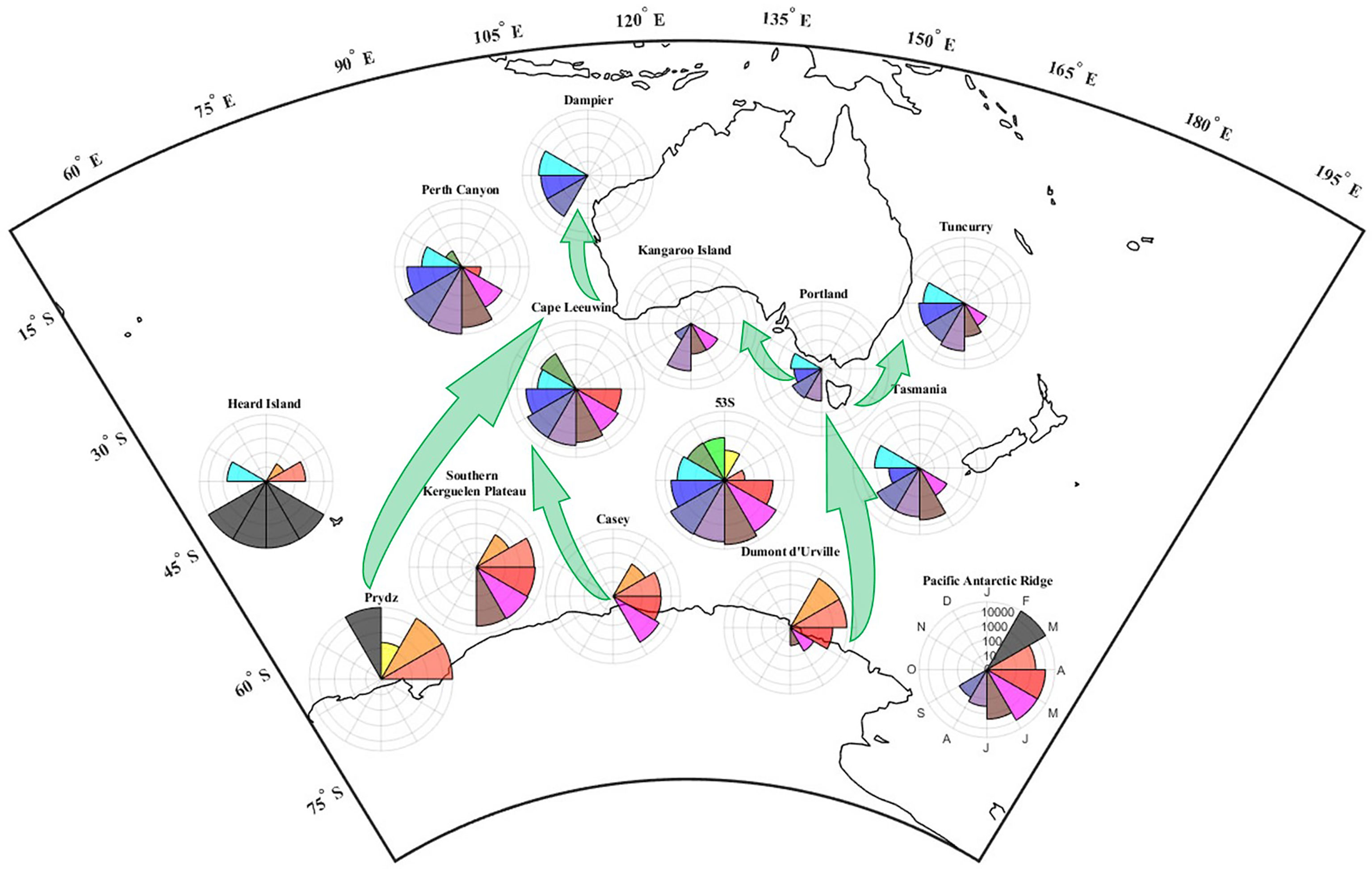
Figure 5 Map of the seasons of the fin whale in Antarctic, Sub-Antarctic and Australian waters. Polar histograms display the mean ‘continuous’ pulse count by month (i.e., number of pulses counted every month, normalised by the DC, and averaged over multiple recording years). Concentric circles correspond to pulse count on a logarithmic scale (10, 100, 1000, 10 000 counts). Polar segments correspond to the months of a year, with January pointing upwards and the following months arranged clock-wise. Grey segments identify months when no recorders were deployed. White segments identify months with recordings but no fin whale pulses. Green arrows highlight the suggested northern migration pathways fin whales take between Antarctic and Australian waters and we assume same paths for the southern migration. For better legibility, polar plots are not positioned right on top of all recorder sites, but have been moved and represent regional trends; see Figure 1 for exact site locations. Equidistant Conic Projection used.
Discussion
Our study identifies the long-term, seasonal vocal presence of the fin whale in Antarctic waters from late austral summer to early winter and in Australian waters from austral autumn to mid-spring. In Sub-Antarctic waters we identified intermediate seasons. Our findings are based on a cumulative 59 years of passive acoustic recordings made over 2002-2019 across 15 locations.
The observed differential seasonality across sites may imply different ecological needs in the different regions. For example, the late austral-winter presence recorded at the Pacific Antarctic Ridge is anomalous in comparison to all other Antarctic sites. Such a late seasonal vocal presence of the animals in high-latitude waters in winter has led to the suggestion of a behavioural change of the Northern Hemisphere sub-species, with breeding taking place in these high-latitude waters (Simon et al., 2010). Similarly, fin whales may be breeding at the Pacific Antarctic Ridge. However, this needs further targeted research. Another important ecological requirement is feeding. In the Antarctic, the Kerguelen Plateau is an integral location for primary production in the Indian sector, supporting dense aggregations of krill, which act as a productive feeding location for many marine species (Tynan, 1997; Nicol et al., 2000; Hindell et al., 2011). Dense aggregations of krill have also been reported in the Prydz region (Higginbottam and Hosie, 1989). In the Pacific sector of Antarctica, the region of Dumont d’Urville is of high krill density and is a productive feeding location for many cetaceans (Nicol et al., 2000). This high primary productivity may be the motivation for long-term, consistent fin whale seasonal presence in the Indian and Pacific sectors of Antarctica, with the animals feeding at these locations. In Australian waters, the Perth Canyon is a probable feeding zone for migratory fin whales (Aulich et al., 2019) due to the high zooplankton density in this region (Rennie et al., 2009).
Further, this defined differential seasonality of fin whales observed between Antarctic, Sub-Antarctic and Australian sites implies that these animals are migratory and not resident in these areas. No sample location observed year-round fin whale calls, giving no evidence of resident, non-migratory fin whales, in contrast to those reported in the Northern Hemisphere. Migratory pathways for the humpback whale (Megaptera novaeangliae) have been identified between Antarctic and Australian waters, from the Kerguelen Plateau to the west coast of Australia (Bestley et al., 2019) and from the Pacific sector of Antarctica to the east coast of Australia (Andrews-Goff et al., 2018). We suggest the defined seasonality of fin whale vocal presence across these regions implies migratory pathways for the species between Antarctic and Australian waters, similar to those of the humpback whale.
The long-term and high seasonal vocal presence of fin whales recorded at the Southern Kerguelen Plateau and Prydz sites indicates that these areas and the Indian sector of Antarctica are a key usage area for the fin whale as it migrates out of Antarctic waters. In comparison, the limited vocal presence of fin whales recorded at the Casey site, indicates this area may be of limited habitat use for the animals as they migrate. The limited vocal presence of fin whales recorded at the Heard Island site may be indicative of a limited number of animals migrating through the Northern Kerguelen Plateau during both their northward and southward migrations. Those fin whales present in the Indian sector of Antarctica may be migrating north to regions of the western Indian Ocean where fin whale seasonal acoustic presence has been reported (Leroy et al., 2018; Dréo et al., 2019) or to Australian waters. The long-term, seasonal presence of fin whales on the west coast of Australia at Cape Leeuwin and the Perth Canyon implies a migratory pathway between Antarctic and Australian waters, with the animals travelling north between the Kerguelen Plateau and the west coast of Australia. The animals first reach Cape Leeuwin which is suggested as a travelling zone (Aulich et al., 2019) due to the pattern of long seasonal vocal presence and limited pulse rates also observed in our study. The animals do not linger at this location, rather travel north to the Perth Canyon for feeding. The limited seasonal presence of fin whales at Dampier may indicate that this region is at or close to the extreme northern extent of their migration on the west coast of Australia.
Due to the identified seasonal vocal presence of fin whales in the Pacific sector of Antarctica, we propose that the Dumont d’Urville and Pacific Antarctic Ridge sites are utilised as migratory pathways for the animals out of Antarctic waters. Those fin whales present at the Pacific Antarctic Ridge may be migrating north to waters around New Zealand, where fin whale seasonal acoustic presence has been reported (McDonald, 2006; Brodie and Dunn, 2015; Constaratas et al., 2021) or to Australian waters. The vocal presence of fin whales recorded at the Sub-Antarctic 53S site supports our suggestion of a migratory pathway between Dumont d’Urville and the east coast of Australia. The long, extended season observed at this site hints at fin whale migration as a slow dispersal, rather than a mass movement (Širović et al., 2004; Širović et al., 2009; Aulich et al., 2019) as they travel through this area on their north and south journey between Antarctic and Australian waters. Migratory fin whales first arrive on the east coast of Australia at Tasmania, travelling through this region as they disperse north to Tuncurry. The Tuncurry site observed significantly lower vocal presence of fin whales to its west coast equivalent, the Perth Canyon. This indicates that this may not be a key habitat usage area, unlike the Perth Canyon. More likely, due to the long season and low pulse rates observed at both these sites, similar to Cape Leeuwin, the Tuncurry and Tasmania sites likely are travelling zones, with the animals not lingering. The low vocal presence recorded at the Portland and Kangaroo Island sites may indicate fewer animals are migrating to the southern coastline of Australia.
Do the Indian Ocean and Pacific Ocean migratory pathways correspond to two separate sub-populations? In the Northern Hemisphere, evidence of limited mixing between fin whales along different migratory pathways has led to suggestions of separate sub-populations of the species (Bérubé et al., 2002; IWC, 2009). The Southern Kerguelen Plateau and Dumont d’Urville deployment sites are separated approximately by 2,600 km. The limited number of calls recorded at the Casey site might indicate that few animals are present or travelling in this region between the Southern Kerguelen Plateau and Dumont d’Urville sites. In Australian waters, fin whales are seasonally present on both the west and east coasts, separated by continental Australia. The limited fin whale vocal presence at the Portland and Kangaroo Island sites might indicate that few animals are present or travelling in this region between west and east coasts of Australia. We propose that the spatial separation between fin whales in the Indian and Pacific sector Antarctic sites (with limited regional overlap) and the seasonal presence of the animals on both east and west coasts of Australia (with limited regional overlap), provides preliminary evidence of two separate sub-populations of fin whale: a population of fin whales in the Indian sector of Antarctica, migrating to the west coast of Australia, and a population of fin whales in the Pacific sector of Antarctica, migrating to the east coast of Australia.
The lack of fin whale vocalisations detected at the Iselin Bank site in 2018 matches that of Širović et al. (2009) who recorded no 20 Hz pulses in 2003 and 2004 at a site in close proximity to Iselin Bank and Miller et al. (2021a) who found no fin whale sounds at a nearby site in the Ross Sea. Distribution of the fin whale in Antarctic waters may be affected by the presence of sea-ice, with the animals absent from areas covered by sea-ice (Širović et al., 2004). The lack of calls detected at these sites may be due to the year-round, perennial sea-ice coverage in the Ross Sea (Comiso et al., 2011) and may be indicative of long-term absence of fin whales in this region. Ongoing acoustic recording at this location would be useful.
Detection range limits of the acoustic receivers, due to the sound propagation environment, are likely an important contributor to the lack of and/or variation in detected fin whale calls across all site-years. The ambient noise in an area may include natural, abiotic noise (e.g., from wind and ice-cracking), biotic noise (e.g., fish and whale choruses), and anthropogenic noise (mostly from vessels) and affect the fin whale signal-to-noise ratio and therefore may reduce detection range (Erbe et al., 2015). Other factors, such as the bathymetry, seafloor geology, water temperature and salinity, also affect propagation loss and thus the received levels of fin whales and (masking) noise, and ultimately detection range (Erbe et al., 2021). The deployment depth of the acoustic receiver may greatly affect fin whale detection, in particular with respect to sound channels, such as the deep sound channel off Australia and the surface duct off Antarctica (e.g., Gavrilov (2018)). Considerations of the deployment depth of acoustic systems in Antarctic waters suggested depth to be greater than 1000 m for consistent sound propagation of fin whale calls (Van Opzeeland et al., 2014). Recorder depths varied greatly across the 15 deployment sites. Detection range of fin whale vocalisations at these Australian sites is likely to range from several 10’s kms to approximately 100 kms (Aulich et al., 2019). In Antarctic waters, Shabangu et al. (2020) estimated a maximum fin whale detection range of 1,700 kms. Another possibility for the variation in detected fin whale calls across site-years is altered acoustic behaviour, such as when the animals are foraging or engaging in breeding displays (Watkins, 1981). Additionally, seasonal vocal presence of fin whales may have been cut short at some sites such as the Perth Canyon in 2017 and the Southern Kerguelen Plateau in 2018 due to deployment times of acoustic systems. Some seasonal presence may have also been missed as not all of the recorders covered a full 365 days/year (e.g. Heard Island 2017). It is important to note that the detection of fin whales at each location only represents vocalising fin whales and a lack of calls does not confirm an absence of animals, rather that there are no vocalising fin whales. Additionally, these findings may represent only male fin whales, as only males are thought to produce the 20 Hz pulse and no information is currently available on the sexes or ages of those fin whales producing the back-beat or higher frequency component call types.
Further studies combining this long-term data analysis with analysis of the call characteristics of these fin whales and genetic sampling may help to define these suggested sub-populations. Additional research investigating environmental variability in these regions may also ascertain drivers for fin whale vocal presence in these regions and the inter-annual variability observed in our study.
Identifying the spatio-temporal pattern of movement of this Southern Hemisphere sub-species of fin whale is vital to inform conservation management of this vulnerable species at a national (Australian) and international level. This study identifies the long-term, seasonal presence of the fin whale in Antarctic waters from late austral summer to early winter and in Australian waters from autumn to mid-spring. We propose two migratory pathways between Antarctic and Australian waters: from the Indian sector of Antarctica to the west coast of Australia and from the Pacific sector of Antarctica to the east coast of Australia. The limited regional mixing and spatial separation of fin whales in Antarctic and Australian waters is preliminary evidence for separate sub-populations between the Indian and Pacific sectors.
Data Availability Statement
Publicly available datasets were analyzed in this study. The data that support the findings of this study are available from the Australian Antarctic Division (https://data.aad.gov.au/metadata/records/AAS_4102_longTermAcousticRecordings), the Integrated Marine Observing System (http://imos.org.au/data/), and the Comprehensive Test Ban Treaty Organisation preparatory commission (https://www.ctbto.org/verification-regime/the-international-data-centre/distribution-of-data-and-data-bulletins-to-member-states/).
Ethics Statement
Ethical review and approval were not required for the animal study because there was no direct involvement with whales or any other marine fauna in this study. All data were collected remotely, through passive, non-invasive ocean observatories.
Author Contributions
MA, RM, BM, and BS conceived the study. MA, with assistance from RM, BM, and CE, designed the experiment. Analysis techniques were developed by RM and CE. BM collected and contributed acoustic data from 5 locations in Antarctica. FS and BM collected and contributed acoustic data from Dumont d’Urville Sea. GG collected and contributed acoustic data from the Ross Sea. All acoustic data analysis was completed by MA with support from RM, CE, BM, and BS. Ecological analysis was completed by MA, CE, and BM. MA wrote the manuscript with input from CE, BM, BS, and RM. All authors approved manuscript submission.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
The Australian Antarctic Division (AAD) provided underwater acoustic recordings made under the Australian Antarctic Science Projects 4101, 4102 and 4600 and the International Whaling Commission’s Southern Ocean Research Partnership (IWC-SORP) Southern Ocean Hydrophone Network (SOHN). The Dumont d’Urville deployments were possible with the support of the Institut Polaire Français Paul Emile Victor under the program SOHN, AREA V (http://dx.doi.org/10.18142/313). Data was sourced from Australia’s Integrated Marine Observing System (IMOS) - IMOS enabled by the National Collaborative Research Infrastructure Strategy (NCRIS). It is operated by a consortium of institutions as an unincorporated joint venture, with the University of Tasmania as Lead Agent. Geoscience Australia provided the acoustic data from the Comprehensive Test Ban Treaty Organisation (CTBTO) hydroacoustic station. Dr Matt Pinkerton at NIWA funded the deployment of the two AMARs in the Ross Sea under the Ross Sea Research and Monitoring Programme of the New Zealand’s Ministry of Business, Innovation and Employment.
References
Andrews-Goff V., Bestley S., Gales N. J., Laverick S. M., Paton D., Polanowski A. M., et al. (2018). Humpback Whale Migrations to Antarctic Summer Foraging Grounds Through the Southwest Pacific Ocean. Sci. Rep. 8 (1), 12333. doi: 10.1038/s41598-018-30748-4
Aulich M. G., McCauley R. D., Saunders B. J., Parsons M. J. G. (2019). Fin Whale (Balaenoptera Physalus) Migration in Australian Waters Using Passive Acoustic Monitoring. Sci. Rep. 9 (1), 8840. doi: 10.1038/s41598-019-45321-w
Bell E., Bedriñana-Romano L., Reyes Reyes M., Marino A., Andrés A. (2019). Annual Report of the Southern Ocean Research Partnership (IWC-SORP) 2018/19. Available at: https://www.researchgate.net/publication/348848538.
Bérubé M., Urbán R. J., Dizon A. E., Brownell R. L., Palsbøll P. J. (2002). Genetic Identification of a Small and Highly Isolated Population of Fin Whales (Balaenoptera Physalus) in the Sea of Cortez, México. Conserv. Genet. 3 (2), 183–190. doi: 10.1023/A:1015224730394
Bestley S., Andrews-Goff V., van Wijk E., Rintoul S. R., Double M. C., How J. (2019). New Insights Into Prime Southern Ocean Forage Grounds for Thriving Western Australian Humpback Whales. Sci. Rep. 9 (1), 13988. doi: 10.1038/s41598-019-50497-2
Brodie D. C., Dunn R. A. (2015). Low Frequency Baleen Whale Calls Detected on Ocean-Bottom Seismometers in the Lau Basin, Southwest Pacific Ocean. J. Acoust. Soc Am. 137 (1), 53–62. doi: 10.1121/1.4904556
Buchan S. J., Gutierrez L., Balcazar-Cabrera N., Stafford K. M. (2019). Seasonal Occurrence of Fin Whale Song Off Juan Fernandez, Chile. Endanger. Species Res. 39, 135–145. doi: 10.3354/esr00956
Burkhardt E., Van Opzeeland I., Cisewski B., Mattmüller R., Meister M., Schall E., et al. (2021). Seasonal and Diel Cycles of Fin Whale Acoustic Occurrence Near Elephant Island, Antarctica. R. Soc Open Sci. 8 (5), 201142. doi: 10.1098/rsos.201142
Castellote M., Clark C. W., Lammers M. O. (2012). Acoustic and Behavioural Changes by Fin Whales (Balaenoptera Physalus) in Response to Shipping and Airgun Noise. Biol. Conserv. 147 (1), 115–122. doi: 10.1016/j.biocon.2011.12.021
Comiso J. C., Kwok R., Martin S., Gordon A. L. (2011). Variability and Trends in Sea Ice Extent and Ice Production in the Ross Sea. J. Geophys. Res. Oceans 116 (C4). doi: 10.1029/2010JC006391
Constaratas A. N., McDonald M. A., Goetz K. T., Giorli G. (2021). Fin Whale Acoustic Populations Present in New Zealand Waters: Description of Song Types, Occurrence and Seasonality Using Passive Acoustic Monitoring. PloS One 16 (7), e0253737. doi: 10.1371/journal.pone.0253737
Cooke J. G. (2018). “Balaenoptera Physalus,” in The IUCN Red List of Threatened Species 2018. Available at: https://dx.doi.org/10.2305/IUCN.UK.2018-2.RLTS.T2478A50349982.en.
Croll D. A., Clark C. W., Acevedo A., Tershy B., Flores S., Gedamke J., et al. (2002). Only Male Fin Whales Sing Loud Songs. Nature 417, 809. doi: 10.1038/417809a
Davis G. E., Baumgartner M. F., Corkeron P. J., Bell J., Berchok C., Bonnell J. M., et al. (2020). Exploring Movement Patterns and Changing Distributions of Baleen Whales in the Western North Atlantic Using a Decade of Passive Acoustic Data. Global Change Biol. 26 (9), 4812–4840. doi: 10.1111/gcb.15191
de la Mare W. K. (2014). Estimating Relative Abundance of Whales From Historical Antarctic Whaling Records. Can. J. Fish Aquat. Sci. 71 (1), 106–119. doi: 10.1139/cjfas-2013-0016
Delarue J., Martin B., Hannay D., Berchok C. L. (2013). Acoustic Occurrence and Affiliation of Fin Whales Detected in the Northeastern Chukchi Sea, July to October 2007-10. Arctic 66 (2), 159–172. doi: 10.14430/arctic4287
Dréo R., Bouffaut L., Leroy E., Barruol G., Samaran F. (2019). Baleen Whale Distribution and Seasonal Occurrence Revealed by an Ocean Bottom Seismometer Network in the Western Indian Ocean. Deep Sea Res. (II Top. Stud. Oceanogr.) 161, 132–144. doi: 10.1016/j.dsr2.2018.04.005
Dziak R. P., Hong J., Kang S., Lau T., Haxel J. H., Matsumoto H. (2017). the Balleny Island Hydrophone Array: Hydro-Acoustic Records of Sea-Ice Dynamics, Seafloor Volcano-Tectonic Activity, and Marine Mammal Vocalizations Off Antarctica. OCEANS 2017 Aberdeen, 1–8. doi: 10.1109/OCEANSE.2017.8084571
Edwards E. F., Hall C., Moore T. J., Sheredy C., Redfern J. V. (2015). Global Distribution of Fin Whales Balaenoptera Physalus in the Post-Whaling Era, (1980–2012). Mamm. Rev. 45 (4), 197–214. doi: 10.1111/mam.12048
Erbe C., Peel D., Smith J. N., Schoeman R. P. (2021). Marine Acoustic Zones of Australia. J. Mar. Sci. Eng. 9 (3), 340. doi: 10.3390/jmse9030340
Erbe C., Verma A., McCauley R., Gavrilov A., Parnum I. (2015). The Marine Soundscape of the Perth Canyon. Prog. Oceanogr. 137, 38–51. doi: 10.1016/j.pocean.2015.05.015
Gavrilov A. (2018). Propagation of Underwater Noise From an Offshore Seismic Survey in Australia to Antarctica: Measurements and Modelling. Acoustics Aust. 46 (1), 143–149. doi: 10.1007/s40857-018-0131-1
Gedamke J., Robinson S. M. (2010). Acoustic Survey for Marine Mammal Occurrence and Distribution Off East Antarctica (30-80°E) in January-February 2006. Deep Sea Res. (II Top. Stud. Oceanogr.) 57 (9), 968–981. doi: 10.1016/j.dsr2.2008.10.042
Harris D., Matias L., Thomas L., Harwood J., Geissler W. H. (2013). Applying Distance Sampling to Fin Whale Calls Recorded by Single Seismic Instruments in the Northeast Atlantic. J. Acoust. Soc Am. 134 (5), 3522–3535. doi: 10.1121/1.4821207
Higginbottam I. R., Hosie G. W. (1989). Biomass and Population Structure of a Large Aggregation of Krill Near Prydz Bay, Antarctica. Mar. Ecol. Prog. Ser. 58 (1), 197–203. doi: 10.3354/meps058197
Hindell M. A., Lea M.-A., Bost C.-A., Charrassin J.-B., Gales N., Goldsworthy S., et al. (2011). Foraging Habitats of Top Predators, and Areas of Ecological Significance, on the Kerguelen Plateau', The Kerguelen Plateau: Marine Ecosystem and Fisheries. Société Française D’Ichtyologie 35, 203–215. doi: 10.26028/cybium/2011-35SP-024
Iwase R. (2015). Fin Whale Vocalizations Observed With Ocean Bottom Seismometers of Cabled Observatories Off East Japan Pacific Ocean. Jap. J. Appl. Phys. 54 (7S1), 07HG03. doi: 10.7567/JJAP.54.07HG03
[IWC] International Whaling Commission (2009). Report of the First Intersessional RMP Workshop on North Atlantic Fin Whales. J. Cetacean Res. Manage.
Leroy E. C., Samaran F., Stafford K. M., Bonnel J., Royer J. Y. (2018). Broad-Scale Study of the Seasonal and Geographic Occurrence of Blue and Fin Whales in the Southern Indian Ocean. Endanger. Species Res. 37, 289–300. doi: 10.3354/esr00927
Lockyer C., Brown S. (1981). The Migration of Whales. Anim. Migr: Cambridge Univ. Press Binghamton New Y. 13, 105–137.
McCauley R. D., Thomas F., Parsons M. J. G., Erbe C., Cato D. H., Duncan A. J., et al. (2017). Developing an Underwater Sound Recorder: The Long and Short (Time) of It. Acoustics Aust. 45 (2), 301–311. doi: 10.1007/s40857-017-0113-8
McDonald M. A. (2006). An Acoustic Survey of Baleen Whales Off Great Barrier Island, New Zealand. N. Z. J. Mar. Freshwat. Res. 40 (4), 519–529. doi: 10.1080/00288330.2006.9517442
Mellinger D. K., Clark C. W. (2000). Recognizing Transient Low-Frequency Whale Sounds by Spectrogram Correlation. J. Acoust. Soc Am. 107 (6), 3518–3529. doi: 10.1121/1.429434
Mellinger D. K., Stafford K. M., Moore S. E., Dziak R. P., Matsumoto H. (2007). An Overview of Fixed Passive Acoustic Observation Methods for Cetaceans. Oceanography 20 (4), 36–45. doi: 10.5670/oceanog.2007.03
Miller B. S., Balcazar N., Nieukirk S., Leroy E. C., Aulich M. G., Shabangu F. W., et al. (2021a). An Open Access Dataset for Developing Automated Detectors of Antarctic Baleen Whale Sounds and Performance Evaluation of Two Commonly Used Detectors. Sci. Rep. 11 (1), 1–18. doi: 10.1038/s41598-020-78995-8
Miller B. S., Milnes M., Whiteside S. (2021b). Long-Term Underwater Acoustic Recordings 2013-2019. Ver. 4. Aust. Antarctic Data Centre. doi: 10.26179/h7xa-y729
Mizroch S. A., Rice D. W., Breiwick J. M. (1984). The Fin Whale, Balaenoptera Physalus. Mar. Fish Rev. 46 (4), 20–24. doi: 10.1016/B978-0-12-373553-9.00102-4
Morano J. L., Salisbury D. P., Rice A. N., Conklin K. L., Falk K. L., Clark C. W. (2012). Seasonal and Geographical Patterns of Fin Whale Song in the Western North Atlantic Ocean. J. Acoust. Soc Am. 132 (2), 1207–1212. doi: 10.1121/1.4730890
Nicol S., Kitchener J., King R., Hosie G., William K. (2000). Population Structure and Condition of Antarctic Krill (Euphausia Superba) Off East Antarctica (80–150 E) During the Austral Summer of 1995/1996. Deep Sea Res. (II Top. Stud. Oceanogr.) 47 (12-13), 2489–2517. doi: 10.1016/S0967-0645(00)00033-3
Oleson E. M., Širović A., Bayless A. R., Hildebrand J. A. (2014). Synchronous Seasonal Change in Fin Whale Song in the North Pacific. PloS One 9 (12), e115678. doi: 10.1371/journal.pone.0115678
Rennie S., Hanson C., McCauley R., Pattiaratchi C., Burton C., Bannister J., et al. (2009). Physical Properties and Processes in the Perth Canyon, Western Australia: Links to Water Column Production and Seasonal Pygmy Blue Whale Abundance. J. Mar. Syst. 77 (1-2), 21–44. doi: 10.1016/j.jmarsys.2008.11.008
Rocha R. C., Clapham P. J., Ivashchenko Y. V. (2014). Emptying the Oceans: A Summary of Industrial Whaling Catches in the 20th Century. Mar. Fish Rev. 76 (4), 37–48. doi: 10.7755/MFR.76.4.3
Shabangu F. W., Andrew R. K., Yemane D., Findlay K. P. (2020). Acoustic Seasonality, Behaviour and Detection Ranges of Antarctic Blue and Fin Whales Under Different Sea Ice Conditions Off Antarctica. Endanger. Species Res. 43, 21–37. doi: 10.3354/esr01050
Simon M., Stafford K. M., Beedholm K., Lee C. M., Madsen P. T. (2010). Singing Behavior of Fin Whales in the Davis Strait With Implications for Mating, Migration and Foraging. J. Acoust. Soc Am. 128 (5), 3200–3210. doi: 10.1121/1.3495946
Širović A., Hildebrand J. A., Wiggins S. M. (2007). Blue and Fin Whale Call Source Levels and Propagation Range in the Southern Ocean. J. Acoust. Soc Am. 122 (2), 1208–1215. doi: 10.1121/1.2749452
Širović A., Hildebrand J. A., Wiggins S. M., McDonald M. A., Moore S. E., Thiele D. (2004). Seasonality of Blue and Fin Whale Calls and the Influence of Sea Ice in the Western Antarctic Peninsula. Deep Sea Res. (II Top. Stud. Oceanogr.) 51 (17-19), 2327–2344. doi: 10.1016/j.dsr2.2004.08.005
Širović A., Hildebrand J. A., Wiggins S. M., Thiele D. (2009). Blue and Fin Whale Acoustic Presence Around Antarctica During 2003 and 2004. Mar. Mamm. Sci. 25 (1), 125–136. doi: 10.1111/j.1748-7692.2008.00239.x
Širović A., Rice A., Chou E., Hildebrand J. A., Wiggins S. M., Roch M. A. (2015). Seven Years of Blue and Fin Whale Call Abundance in the Southern California Bight. Endanger. Species Res. 28 (1), 61–76. doi: 10.3354/esr00676
Širović A., Williams L. N., Kerosky S. M., Wiggins S. M., Hildebrand J. A. (2013). Temporal Separation of Two Fin Whale Call Types Across the Eastern North Pacific. Mar. Biol. 160 (1), 47–57. doi: 10.1007/s00227-012-2061-z
Stafford K. M., Mellinger D. K., Moore S. E., Fox C. G. (2007). Seasonal Variability and Detection Range Modeling of Baleen Whale Calls in the Gulf of Alaska 1999-2002. J. Acoust. Soc Am. 122 (6), 3378–3390. doi: 10.1121/1.2799905
Thompson P. O., Findley L. T., Vidal O. (1992). 20-Hz Pulses and Other Vocalizations of Fin Whales, Balaenoptera Physalus, in the Gulf of California, Mexico. J. Acoust. Soc Am. 92 (6), 3051–3057. doi: 10.1121/1.404201
Tulloch V. J. D., Plagányi É.E., Brown C., Richardson A. J., Matear R. (2019). Future Recovery of Baleen Whales Is Imperiled by Climate Change. Global Change Biol. 25 (4), 1263–1281. doi: 10.1111/gcb.14573
Tynan C. T. (1997). Cetacean Distributions and Oceanographic Features Near the Kerguelen Plateau. Geophys. Res. Lett. 24(22) pp, 2793–2796. doi: 10.1029/97GL02860
Van Opzeeland I., Samaran F., Stafford K. M., Findlay K., Gedamke J., Harris D., et al. (2014). Towards Collective Circum-Antarctic Passive Acoustic Monitoring: The Southern Ocean Hydrophone Network (SOHN). Polarforschung 83 (2), 47–61. doi: 10.013/epic.44470.d001
Watkins W. A. (1981). Activities and Underwater Sounds of Fin Whales. Sci. Rep. Whales Res. Institute 33, 83–117. doi: 10.1016/0198-0254(82)90294-1
Keywords: Antarctica, Australia, Balaenoptera physalus, fin whale, migratory pathway, passive acoustic monitoring, seasonal distribution
Citation: Aulich MG, McCauley RD, Miller BS, Samaran F, Giorli G, Saunders BJ and Erbe C (2022) Seasonal Distribution of the Fin Whale (Balaenoptera physalus) in Antarctic and Australian Waters Based on Passive Acoustics. Front. Mar. Sci. 9:864153. doi: 10.3389/fmars.2022.864153
Received: 28 January 2022; Accepted: 19 April 2022;
Published: 16 May 2022.
Edited by:
Luis Huckstadt, University of Exeter, United KingdomReviewed by:
Letizia Marsili, University of Siena, ItalyGenevieve Davis, Northeast Fisheries Science Center (NOAA), United States
Simon David Berrow, Galway-Mayo Institute of Technology, Ireland
Copyright © 2022 Aulich, McCauley, Miller, Samaran, Giorli, Saunders and Erbe. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Meghan G. Aulich, TWVnaGFuLmF1bGljaEBob3RtYWlsLmNvbQ==