 Hellen Hiza1,2,3
Hellen Hiza1,2,3 Jerry Hella1,2,3
Jerry Hella1,2,3 Ainhoa Arbués2,3
Ainhoa Arbués2,3 Mohamed Sasamalo1,2,3
Mohamed Sasamalo1,2,3 Veronica Misana1,2,3
Veronica Misana1,2,3 Jacques Fellay4,5,6
Jacques Fellay4,5,6 Sébastien Gagneux2,3Klaus Reither2,3
Sébastien Gagneux2,3Klaus Reither2,3 Damien Portevin2,3*
Damien Portevin2,3*- 1Ifakara Health Institute, Bagamoyo, Tanzania
- 2Swiss Tropical and Public Health Institute, Basel, Switzerland
- 3University of Basel, Basel, Switzerland
- 4School of Life Sciences, Ecole Polytechnique Federale de Lausanne, Lausanne, Switzerland
- 5Swiss Institute of Bioinformatics, Lausanne, Switzerland
- 6Precision Medicine Unit, Lausanne University Hospital, University of Lausanne, Lausanne, Switzerland
T cell activation markers (TAM) expressed by antigen-specific T cells constitute promising candidates to attest the presence of an active infection by Mycobacterium tuberculosis (Mtb). Reciprocally, their modulation may be used to assess antibiotic treatment efficacy and eventually attest disease resolution. We hypothesized that the phenotype of Mtb-specific T cells may be quantitatively impacted by the load of bacteria present in a patient. We recruited 105 Tanzanian adult tuberculosis (TB) patients and obtained blood before and after 5 months of antibiotic treatment. We studied relationships between patients' clinical characteristics of disease severity and microbiological as well as molecular proxies of bacterial load in sputum at the time of diagnosis. Besides, we measured by flow cytometry the expression of CD38 or CD27 on CD4+ T cells producing interferon gamma (IFN-γ) and/or tumor necrosis factor alpha (TNF-α) in response to a synthetic peptide pool covering the sequences of Mtb antigens ESAT-6, CFP-10, and TB10.4. Reflecting the difficulty to extrapolate bacterial burden from a single end-point read-out, we observed statistically significant but weak correlations between Xpert MTB/RIF, molecular bacterial load assay and time to culture positivity. Unlike CD27, the resolution of CD38 expression by antigen-specific T cells was observed readily following 5 months of antibiotic therapy. However, the intensity of CD38-TAM signals measured at diagnosis did not significantly correlate with Mtb 16S RNA or rpoB DNA detected in patients' sputa. Altogether, our data support CD38-TAM as an accurate marker of infection resolution independently of sputum bacterial load.
Introduction
Tuberculosis (TB) is an air-born infection caused by gram-positive bacilli called Mycobacterium tuberculosis (Mtb). In 2020, an estimated 9.9 million people fell ill of TB, of which 4.1 million were not diagnosed and 1.5 million died (1). As such, TB remains today the leading cause of death from a single infectious agent after the SARS-CoV-2 coronavirus (COVID-19). Yet, TB is mostly treatable with a combination of drugs for a minimum of 6 months. Unfortunately, side effects and hepatotoxicity in particular has been observed for up to 28% of patients (2). In addition, lung imaging by tomography coupled to the detection of short-lived mRNA species within sputa demonstrated that nearly a quarter of patients would still harbor active lesions and live mycobacteria in their lungs upon treatment completion (3). This may be linked to Mtb's capacity to respond to immune-induced stresses by switching into a dormant state metabolically associated with drug tolerance, as well as the limited penetration of antibiotics into caseous lung lesions and cavities (4, 5). Thus, despite being apparently cured, many patients may still harbor live bacilli in their lungs at the end of their treatment. TB control strategies should therefore not only rely on improving diagnosis and shortening treatment, but also monitoring bacterial clearance to ensure treatment efficacy and disease resolution (6, 7). One particular problem is that, without accurate monitoring tools, all TB patients have to complete 6 months of treatment despite evidence suggesting cure after 4 months in substantial proportion of patients (8), while others may develop drug-resistant (DR)-TB requiring adjusted treatment regimens. DR-TB treatment are longer, often poorly tolerated and have lower cure rates (9, 10). Shorter TB treatment regimens involving new formulations, new compounds or repurposed drugs are being tested in clinical trials to treat drug-sensitive as well as DR-TB (6). These trials are challenged by the difficulty to accurately quantify and compare treatment responses (11, 12). Sputum-based treatment monitoring tools such as smear microscopy, looking at acid-fast bacilli positivity conversion after 2 months of treatment initiation, have low sensitivity (13, 14). Moreover, this test cannot distinguish between dead and live bacilli and was shown to be poorly sensitive and specific of treatment failure or relapse (15). Molecular-based methods, such as Xpert MTB/RIF, cannot differentiate between live and dead bacteria either (16). Mtb sputum culture results require up to 8 weeks for solid culture and 42 days for liquid culture thus cannot be used to assess the patient's response to treatment in a timely manner (17). Furthermore, collection of sputum samples beyond 2 months of treatment is challenging as patients' coughs have often resolved by then (7). Early bacterial activity (EBA) assessing bacterial decline over 14 days following onset of treatment, has been implemented but this method cannot detect the eventual development of DR-TB (18, 19). Molecular methods such as quantitation of 16S rRNA using molecular bacterial load assay (MBLA) have been employed in EBA studies to reflect bacterial decline as early as 3 days following initiation of treatment, predict relapse, monitor treatment response and are closely correlated with culture-based read-outs (20, 21). However, extrapolating the bacterial burden of a patient from a single sputum sample is inherently inaccurate due to the stochastic release of bacteria across independent coughing episodes and the impossibility to sample bacilli outside the lung compartment in the case of extra-pulmonary TB. In that context, non-sputum-based monitoring tools may be more appropriate. Blood-based biomarker tests constitute promising tools for sputum-independent diagnosis of TB and, by extension, monitor treatment response and predict treatment outcome (7, 8, 22). IFN-γ and/or TNF-α-producing CD4+ T cells expressing activation, differentiation or proliferation markers have been repeatedly explored for TB diagnosis (7, 13, 17, 22–25). In addition to their diagnostic potential, these T cell activation markers (TAM) have been investigated for their ability to monitor bacterial clearance, disease extent and treatment outcome (22, 23, 26, 27). Compared to CD27 and Ki67, CD38 and HLA-DR expressed by Mtb-specific CD4+ T cells declined more rapidly following anti-TB treatment initiation (8, 17, 23). The frequencies of antigen-specific CD4+ T cells expressing HLA-DR or Ki67 have been associated with disease severity and time to culture conversion (23, 24, 28). In that context, we aimed to assess the correlation of CD38 and CD27 biomarkers with measures of disease severity including sputum bacterial load and in response to anti-TB treatment using a standardized whole-blood TAM assay.
Methods
Study Design and Participants
We collected samples from 105 drug-sensitive TB patients prospectively recruited from the National Tuberculosis and Leprosy Program (NTLP) TB clinic, who were enrolled as a part of an ongoing TB-Dar cohort study (25). All consecutive adult TB patients (age ≥18 years) with a positive sputum smear microscopy (Ziehl-Neelsen staining) and/or M.tuberculosis detected by Xpert MTB/RIF were eligible for the study. Clinical assessment was done at the time of diagnosis as well as after 2, 5, and 12 months to assess patient response to anti-TB treatment and relapses. Blood specimens were collected at the time of TB diagnosis and 5 months after anti-TB treatment initiation. In addition, patients provided sputum samples at the time of study enrolment and, when possible, at month 5 for assessment of treatment success. The study protocol was approved by the institutional review board of the Ifakara Health Institute (IHI; reference no. IHI/IRB/EXT/No: 16-2019) and the Medical Research Coordinating Committee of the National Institute for Medical Research (NIMR; reference no. NIMR/HQ/R.8a/Vol.IX/1641) in Tanzania. All participants provided a signed informed consent to collect clinical data, sputum and blood samples. In case of illiteracy, study information was given in the presence of an impartial, literate witness, who read the information sheet to the participant or witnessed the complete reading of the information sheet to the participant.
TB Score
Excepting tachycardia (data not collected), the TB score was adapted based on 10 of the 11 signs and symptoms of TB disease that were previously reported as a potential measure of treatment outcome in resource-limited settings (29). The TB score used here therefore encompassed cough, hemoptysis, dyspnea, chest pain and night sweat self-reported symptoms as well as pallor anemia, positive finding at lung auscultation, fever (>37°C), body mass index (BMI) <18, BMI <16, middle upper arm circumference (MUAC) <220 and MUAC <200. Each parameter accounted for 1 point adding up to a maximum of 12 points.
Microbiological Procedures
The Xpert MTB/RIF test was performed at the NTLP laboratory and only positive patients were enrolled for this study. On the day of enrolment and prior initiation of treatment, an early morning sputum specimen was collected for culture and bacterial load assessment. The samples were first homogenized for 30 min then treated with cetylpyridinium chloride at 25°C for 4–7 days to increase culture recovery as reported previously (30). A final decontamination step with 1% NaOH (final concentration) was performed before culture on Lowenstein–Jensen media at 37°C. For molecular bacterial load assessment (MBLA), 1 ml of homogenized sputum was mixed with freshly thawed guanidine thiocyanate solution (4M GTC, Promega; V2791) in 1M Tris-HCl pH 7.5 complemented with 1% β-mercaptoethanol shortly before usage. GTC-treated sputa were left standing for 2 h at 25°C before storage at -80°C.
RNA Extraction and Quantitation
Sputum samples in GTC were centrifuged at 3,000 g for 30 min. Pellet was suspended in 500 μl of TRI Reagent® (Zymo; R2050-1-200) and stored at −80°C. TRI samples were spiked with 50 ng of internal control (IC) made of a 1957 bp sequence of the potato gene phyB prepared as described elsewhere (31) and transferred to BeadBug tubes (SIGMA; Z763721). Subsequently, samples were bead-beaten using a FastPrep-24 (Life sciences) for 25 s at 6.5 m/s, cooled on ice for 5 min, bead-beaten for an additional 20 s cycle and placed back on ice. RNA extraction was performed using the Direct-zol™ RNA Microprep kit (Zymo; R2062) according to manufacturer's instructions. DNA contamination was assessed following 16S quantitative PCR (qPCR) using HOT FirePol EvaGreen qPCR mix plus (Solis BioDyne: 08-24-0000S) on a StepOnePlus Real-Time PCR System (ThermoFisher). Samples with a 16S cycle threshold (Ct) value <35 were treated with RQ1 DNase (Promega; M6101) and purified using RNAeasy MinElute® cleanup kit (Qiagen; 74204) according to manufacturer's protocol. Multiplex reverse transcription (RT)- qPCR was performed using SOLIScript 1-step Multiplex Probe Kit (Solis BioDyne; 08-59-0000S) and the primers and probes listed in the Supplementary Table 1. Each sample was assessed in duplicate plus a minus RT control.
MBLA Normalization
To account for potential RNA loss during the extraction and amplification inhibition, we assessed the correlation between RT-qPCR detection of 16S rRNA and the IC across 24 technical replicates of RNA extraction from an H37Rv culture (31). Linear regression was performed and the resulting slope was used for IC-based normalization of the 16S rRNA Ct value obtained for each sample (Supplementary Figure 1).
MBLA Standard Curve
To establish the linearity and the limit of detection of the MBLA assay, triplicates of 10-fold serial dilutions were prepared from an exponentially growing H37Rv culture (OD 0.51). Bacterial concentration was quantified by CFU assessment on 7H11/OADC agar plates. 16S rRNA Ct values were normalized using the equation 16S rRNA Ct – [(IC Ct – 16.00) × slope] and plotted against the bacterial amount determined by CFU (ranging approximately from 5 × 107 to 5 CFU/ml). Best-fit linear equation was subsequently used to extrapolate sputum bacterial load from the respective normalized 16S rRNA Ct value obtained from the sputum specimen (Supplementary Figure 1).
TAM-TB Assay
Freshly collected blood samples were processed for TAM-TB assessment as recently described (25) excepting blood specimens from five patients that were processed retrospectively from cryopreserved PBMCs to circumvent technical errors that prevented the interpretation of the blood-based read-out and as described previously with comparable assay performance (32). None of the PBMC specimen belong to the TAM-TB outliers identified and highlighted in Figure 2. The following reagents were obtained through BEI Resources (NIAID, NIH): Peptide Array, Mycobacterium tuberculosis ESAT-6 Protein, NR-50711, Mycobacterium tuberculosis CFP-10 Protein, NR-50712 and Mycobacterium tuberculosis TB10.4, NR-34826. The following antibody clones were used: anti-CD3-FITC (clone OKT3), anti-CD4-FITC (clone RPA-T4), anti-CD8a-APC (clone HIT8a), anti-IFN-γ-PerCP (clone 4S.B3) and anti-TNF-α-PerCP (clone Mab11). TAM-TB outputs were made of ratios between the frequency of cytokine-producing CD4+ T cells negative for the investigated biomarker (CD27 or CD38) divided by the frequency of cytokine-producing CD4+ T cells expressing the respective biomarker of interest following the gating strategy reported previously (25). For linear regression analysis, TAM-TB outputs were subjected to the identification and removal of outliers using the ROUT method (33).
Statistical Analyses
Proportions and measures of central tendency (mean or median) were used to describe patient characteristics as detailed in Table 1. Mann-Whitney U-test and roc.test (pROC) functions were used under R version 4.0.3 to, respectively, test differences in medians between groups and compare correlated ROC curves. Analysis of variance (ANOVA), linear regression, identification of outliers (ROUT method) and Wilcoxon matched–paired rank tests were performed with GraphPad Prism 8.2.1.
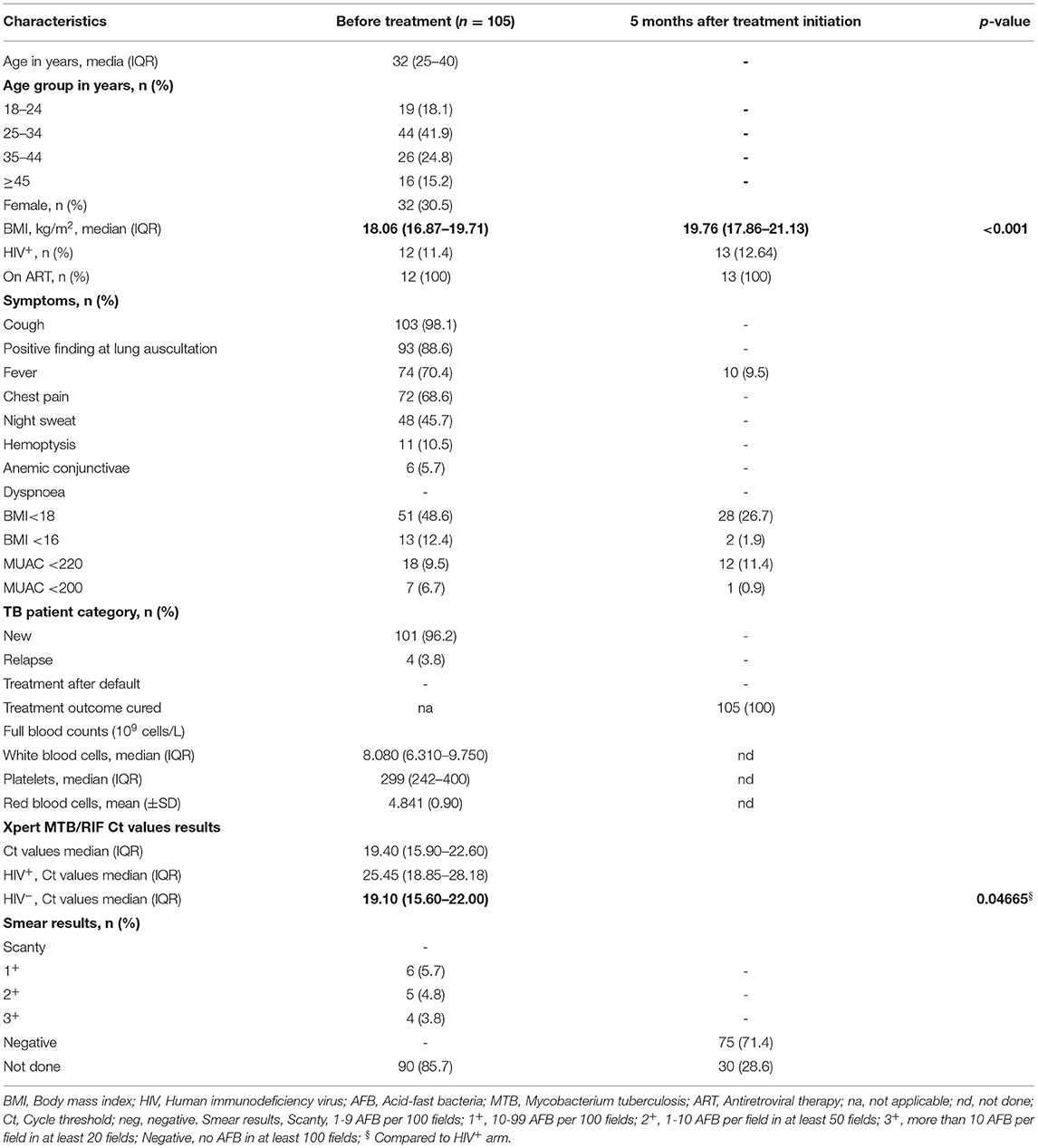
Table 1. Demographics and clinical characteristics before and after 5 months of treatment.
Results
Study Cohort
Clinical data at the time of phlebotomy are summarized in Table 1. All study participants had drug-sensitive TB defined by Xpert MTB/RIF (Xpert) test results with a median age of 32 years [IQR: 25–40]. 12 patients (11.4%) were HIV co-infected, 11 (91.7%) of whom were already on antiretroviral treatment (ART) at the time of TB diagnosis and one initiated ART during the course of antibiotic therapy. Consistent with previous reports (34) and despite ART, HIV co-infected participants presented with a significantly lower bacterial load at the time of TB diagnosis with a reported median Ct value of 25.45 [IQR: 18.85–28.18] compared to 19.40 [IQR: 15.60–22.00] in HIV-negative patients (p = 0.0467). Seventy-five patients (71.4%) were still able to produce sputum specimen after 5 months of anti-TB treatment (ATT) of which all were found smear negative by microscopy. TB clinical severity reflected by TB score showed a significant resolution of disease (p < 0.001) after 5 months of ATT (Figure 1A). Excepting mild fever episodes (<37.8°C) in 9.5% of patients, major TB symptoms had resolved in all participants after 5 months of ATT, yet anti-TB treatment was pursued for another month as per NTLP guidelines (35). Consistent with the complex reverse causality between weight loss and undernutrition in TB pathogenesis and susceptibility (36), we observed a significant increase in median BMI (18.06 kg/m2 [IQR: 16.87–19.71] to 19.76 kg/m2 [IQR: 17.86–21.13], p < 0.001) after 5 months of TB treatment.
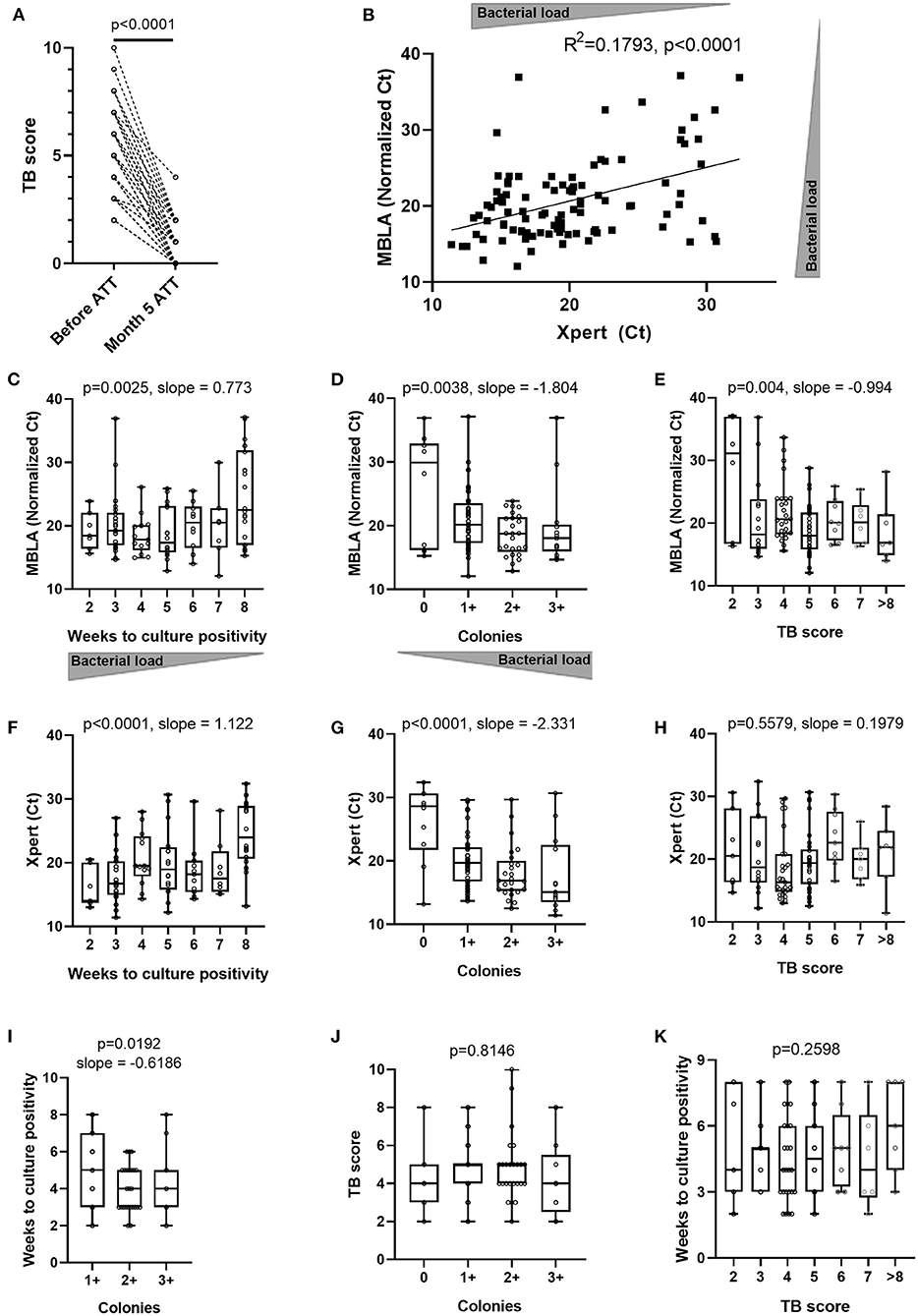
Figure 1. Concordance between various estimates of sputum bacterial load and clinical assessment of disease severity across TB patients at time of diagnosis and after 5 months of antibiotic treatment. (A) Evolution of clinical TB score after 5 months of anti-tuberculous treatment (Month 5 ATT) (Wilcoxon matched-pairs signed rank test p-value). Sputum molecular bacterial load assay (MBLA) results from specimens before initiation of anti-tuberculous treatment (ATT) against: (B) Xpert MTB/RIF Ct values from an independent sputum specimen or (C) time to culture positivity in weeks, or (D) solid culture intensity grades from the same sputum specimen and (E) patient's clinical TB score at time of diagnosis. Xpert MTB/RIF Ct values from sputum specimens before initiation of anti-tuberculous treatment (ATT) against: (F) time to culture positivity results from an independent sputum specimen or (G) solid culture intensity grades from an independent sputum specimen, or (H) patient's clinical TB score at time of diagnosis. (I) Time to culture positivity results against solid culture intensity grades from the same sputum specimen. Patient's clinical TB score at time of diagnosis against (J) sputum solid culture intensity grades or (K) time to culture positivity results. (B) Pearson correlation p-value (two-tailed) and R squared. (C–K) Ordinary one-way ANOVA test for trend p-values with slopes for p < 0.05.
Sputum Bacterial Load and TB Disease Severity
To counteract the poor discriminative value of Xpert likely originating from its capacity to detect cell-free DNA and DNA from dead bacteria (37), we used the 16S rRNA-based molecular bacterial load assay (MBLA) (31) to estimate the number of viable Mtb bacilli present in sputum at baseline. MBLA Ct values plotted against Xpert Ct values revealed a statistically significant, yet moderate, correlation between the two measurements (Figure 1B). We then investigated the relationship between the MBLA or Xpert results and the readouts from solid culture-based methods encompassing time to culture positivity (TTP) (Figures 1C,F) and colony counts (Figures 1D,G). Lower MBLA or Xpert Ct values were both associated with shorter TTP and higher bacterial counts, yet substantial discrepancies remained. For instance, sputum samples producing similar MBLA or Xpert Ct values also displayed different TTP or colony counts. Finally, we sought to explore the relationship between sputum bacterial load and clinical disease severity reflected by the TB score, which accounts for the presence of self-reported symptoms as well as other relevant clinical parameters (29). While we could observe a trend for extremely low or high clinical scores associating with rather high and low MBLA Ct, respectively, patients with a TB score ranging between 3 and 8 displayed highly overlapping bacterial load (Figure 1E). We also observed moderate correlation between time to culture positivity and sputum bacterial load intensity grading on solid culture (Figure 1I). Besides, there was no correlation between Xpert Ct and time to culture positivity or sputum bacterial load intensity grading on solid culture with TB clinical score (Figures 1H,J,K). Together, our data suggest that bacterial load estimates relying on a single end-point sputum specimen are unlikely to accurately reflect the absolute amount of bacteria present in a patient. Hence, we next determined the accuracy of the non-sputum-based TAM-TB assay to discriminate between active and cured TB patients and whether the intensity of the TAM-TB signals correlate with MBLA Ct values.
TAM-TB Signals at Diagnosis Are Independent of Sputum Bacterial Load Proxies
Since TAM-TB signals were found to be actively promoted by ongoing infection, we next studied whether their intensity may be modulated in a quantitative manner by the amount of bacteria detected through the different sputum bacterial load proxies within the same patient. We found that the CD27-based TAM-TB intensities did not decrease with the amount of mycobacterial RNA or DNA recovered in the sputum of the respective patient (Figures 2A,C). In contrast, we observed a rather poor goodness of fit yet, a significant deviation from a non-zero slope between MBLA or Xpert Ct values and the decline of CD38 positive antigen-specific CD4+ T cells that translate into increasing CD38-based TAM-TB results when expressed as a ratio with CD38 negative antigen-specific CD4 T cells (Figures 2B,D). However, the correlation appeared to depend on few very high CD38 TAM-TB results that would stand as outliers following a ROUT-based detection method. Removing these outliers impacted substantially the regression analysis and as a consequence the interpretation of the CD38 TAM-TB data that would no longer correlate significantly with sputum bacterial load (Figures 2B,D). Besides, TAM-TB signals at diagnosis did not correlate with time to culture positivity, nor with colony counts or TB score (Supplementary Figure 2).
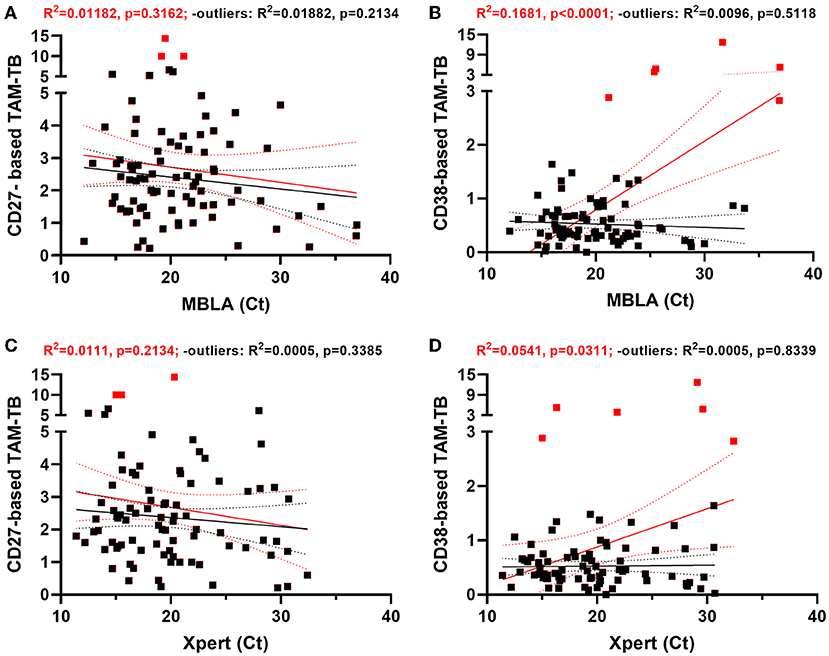
Figure 2. CD38- and CD27-based TAM-TB signals are independent of sputum bacterial load proxies at time of diagnosis. (A–D) Linear regression analysis comparing the strength of the association between CD27 or CD38 biomarker expression by Mtb-specific CD4 T cells (CD27- or CD38-based TAM-TB) and sputum molecular bacterial load assay (MBLA) or Xpert MTB/RIF results from, respectively, blood and sputum specimen of TB patients at time of diagnosis. Regression analysis was performed including (red lines and statistical results) or excluding (black lines and statistical results) potential outliers (red squares) identified using the ROUT method.
CD38-Based TAM-TB Most Accurately Reflects Clinical Resolution Following Antibiotic Treatment
We recently reported the accuracy of CD27- and CD38-based TAM-TB assay to diagnose TB in a cohort of adults with presumptive TB (25). In the current study, we aimed to compare the evolution of CD27 and CD38 upon bacterial clearance following antibiotic treatment by performing the assay on blood collected at the time of TB diagnosis prior treatment initiation and after 5 months of ATT and clinical confirmation of disease resolution. We ran the CD27- and CD38-based TAM assays side-by-side. We intentionally implemented our approach using the most basic 4-color flow cytometer apparatus that has been widely implemented in TB endemic settings for CD4 T cell count purposes. Our gating strategy demonstrated that only CD4+/CD3+ benefit from a double shift in FL-1 staining intensity and that only the double positive cell population was gated and subjected to the phenotypic analysis (25). The frequency of cytokine-producing T cells across the two independent stimulations for all collected specimens is plotted in Figure 3A to demonstrate the robustness of the assay to recall Mtb-specific T cell responses in a reproducible manner (slope = 1.043, R2 = 0.9299). Overall, we observed a significant drop in the frequency of antigen-specific T cells recalled by the synthetic peptide pool after 5 months of treatment (Figure 3B; median response before and 5 months post-ATT: 0.131% and 0.0773%, respectively, p < 0.001). Nonetheless and consistent with the concurrent resolution of TB with the effector phenotype of specific T cells, the frequency of Mtb-specific T cells expressing CD27 and CD38 was significantly increased or decreased, respectively, during ATT (Figures 3C,D). Moreover, a receiver operating characteristic (ROC) curve analysis revealed a substantially superior resolution of CD38 compared to CD27 with an area under the ROC (AUROC) of 0.953 vs. 0.7758 (Bootstrap test for two correlated ROC curves, p = 4.919e-06) (Figure 3E). In comparison, the AUROC analysis of the variation of Mtb-specific CD4 cells frequencies before and after ATT showed much poorer diagnostic accuracy (AUROC: 0.644) further demonstrating the incremental performance of the TAM assays over measuring the percentage of Mtb-specific T cells alone.
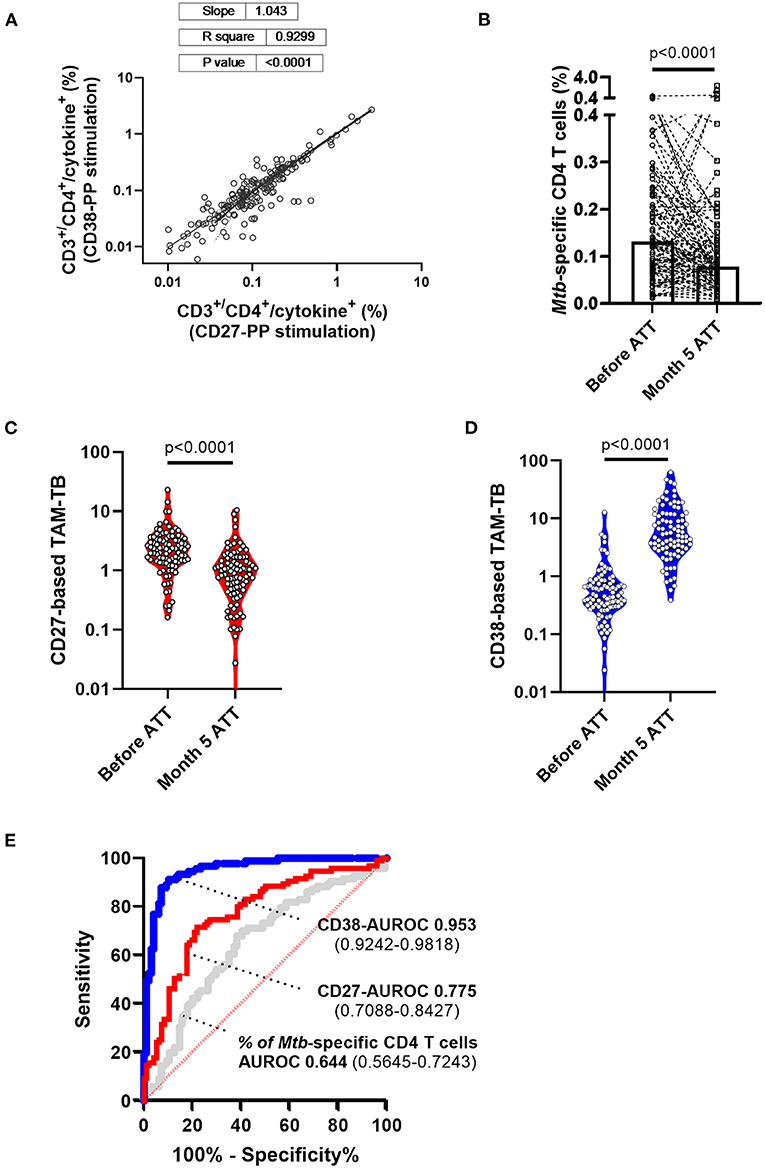
Figure 3. Superiority of CD38- over CD27-based T cell activation marker to attest TB infection resolution following 5 months of anti-tuberculous treatment. (A) Frequencies within CD3+/CD4+ cells of cytokine-producing cells across independent stimulation of peripheral blood mononuclear cells from the same patient and for both study visits i.e., before and after 5 months of anti-tuberculosis treatment (ATT). (B) Patient (lines) and median (box) response of the frequencies of antigen-specific T cells recalled by the synthetic peptide pool before and after 5 months of ATT (Month 5 ATT). Evolution of (C) CD27 and (D) CD38 T cell activation markers expressed by Mtb-specific CD4 T cell before and after 5 months of antibiotic treatment. (E) Receiver operating characteristic (ROC) curve and area under the ROC curve with 95% confidence interval showing the superior discriminatory power (Bootstrap test for two correlated ROC curves, p = 4.919e-06) of CD38- (blue curve) over CD27 (red curve) biomarkers to discriminate Mtb-specific CD4 T cell responses of patients before and after 5 months of chemotherapy. In comparison, the AUROC analysis of the variation of Mtb-specific CD4 T cell frequency alone (gray curve) before and after ATT displays substantially poorer accuracy.
Discussion
We hypothesized that during an infection, the bacterial antigens drained into secondary lymphoid organs trigger or recall T cell activation in an antigen-specific manner, and that the effector phenotype intensity of the Mtb-specific T cell population that recirculate in the periphery may therefore reflect the bacterial load independently of the infection site. This assumption implies that TAM signals should wane upon bacterial clearance by ATT and that they should also display signs of correlation with alternative read-outs of bacterial load. Side-by-side evaluation of CD27- vs. CD38-based assays to distinguish active from cured TB were previously reported in a pilot study and as a case report in the context of extra-pulmonary TB (8, 17). Compared to previous reports, our study benefited from a significantly bigger sample size and a systematic comparison of the two markers performed in real-time in a clinical research setting of patients seeking care at district hospital level. In addition, attempts to correlate these two specific markers with all possible proxies of sputum bacterial load remained to our knowledge unreported. In that context, we could demonstrate the superiority of CD38 to resolve swiftly after 5 months of ATT and concomitant disease resolution (Figure 3E). This observation is consistent with the studies from Halliday et al. and Streitz et al., that reported the incapacity of CD27 to distinguish between recently and remotely acquired latent TB infection (LTBI) and the very slow reversion of CD27 downregulation in patients up to 1 year after therapy (38, 39). Our results also concur with the excellent performance of CD38 to diagnose TB with no interferences arising from LTBI (13, 25). The superior capacity of CD38 over CD27 to monitor treatment responses independently of the amount of live bacteria recovered from patient sputum is striking and offers a particularly interesting tool to monitor resolution of paucibacillary or extra-pulmonary TB infections. This difference also suggests a differential expression kinetics for each marker upon T cell priming or recall and/or that a differential threshold of antigen concentration may be required to stimulate or maintain the expression of the two markers. Both CD27 and CD38 markers are being classically used as binary markers of antigen-specific T cell differentiation and activation (40). Yet, the phenotypic distinction between naïve T cells and the different clusters alongside human memory and effector T cell differentiation involves the sequential gain or loss of a combination of surface markers encompassing also CD45RO, CCR7, CD28, and CD95 (41). Functionally, CD38 expression may need to be more tightly regulated upon infection resolution notably due to its role in regulating inflammation which activity has been linked to autoimmune disorders (42). In return, the persistence of stem cell memory T cells with a naïve-like (CD27neg) phenotype has been reported and may well-contribute to the poor resolution of CD27-based TAM-TB signals after 5 months of ATT (43). Another T cell marker, CD153, has been recently reported to correlate with bacterial load in non-human primate models as well as in TB patients (22). Conversely, ATT only partially restored CD153 expression in the Mtb-specific CD4 response. In the absence of a gold standard, we cannot attest the superiority of immunodiagnostic tools over microbiological and molecular approaches to assess Mtb bacterial burden most accurately. Previously, sputum bacterial load assessment determined by colony forming units or culture time to positivity correlated only moderately with molecular detection methods based on DNA such as Xpert MTB/RIF assay (37). Consequently, we implemented the recently developed 16S rRNA-based molecular bacterial load assay (MBLA) (31) to estimate viable Mtb bacilli within the collected sputum specimens. As expected, we observed a moderate correlation between DNA- and RNA-based read-outs across the cohort of TB patients. Specimens displaying high MBLA Ct values despite low Xpert Ct values could be readily explained by the detection of cell-free DNA or DNA from dead bacteria. However, the reciprocal scenarios are more difficult to reconcile. Such discrepancies between the two molecular methods could first have arisen from sampling variability. Indeed, Xpert results were provided by the NTLP laboratory while MBLA tests were performed on an independent sputum sample collected on the day of study enrolment. Alternatively and non-exclusively, a high prevalence of dormant bacteria (44) harboring limited transcriptional activity could also fuel such differences. We also found discrepancies between MBLA Ct values and culture-based read-outs of sputum bacterial load. In that regard, differentially cultivable mycobacteria across liquid and solid media, as well as bacterial subpopulations requiring resuscitation factors to grow, may affect bacterial recovery (45, 46). We observed a rather poor relationship between the amount of bacteria recovered from sputum specimens collected at the time of TB diagnosis and clinical disease severity scores (Figure 1E). Since TB score has been found to significantly and inversely correlate with survival estimates, our data suggest that TB disease severity does not always corresponds with greater bacillary burden. Actually, TB in HIV-infected individuals is generally associated with a lower bacterial load in the sputum but also poor prognosis (47). In addition, a single end-point measurement from a unique sputum specimen is unlikely to reflect accurately the absolute amount of bacteria present in the entire lung compartment. Furthermore, sputum-based read-outs may underestimate the bacterial burden of a particular host/strain combination that would favor the dissemination of the bacteria outside the lungs (48).
In conclusion, we observed the superiority of CD38 over CD27 to discriminate active TB patients from cured individuals and so, independently of the amount of live mycobacteria that could be retrieved from the patient's sputa at the time of TB diagnosis. Taken together, our results suggest that the expression of the T cell activation marker CD38 solely warrant the infection status and is not quantitatively induced by the presence of live mycobacteria during TB. Yet, the CD38-based TAM-TB assay constitutes an excellent assay to monitor treatment response. Being not significantly influenced by sputum bacterial load, we also propose that in our efforts to end TB, the CD38-based TAM-TB assay is a promising avenue to diagnose asymptomatic subclinical forms of TB (49).
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found at: BioStudies repository, accession number S-BSST740.
Ethics Statement
The studies involving human participants were reviewed and approved by the Institutional Review Board of the Ifakara Health Institute (IHI; reference no. IHI/IRB/EXT/No: 16-2019) and the Medical Research Coordinating Committee of the National Institute for Medical Research (NIMR; reference no. NIMR/HQ/R.8a/Vol.IX/1641) in Tanzania. The patients/participants provided their written informed consent to participate in this study.
Author Contributions
JF, SG, KR, and DP: study conception and design. HH, JH, AA, MS, VM, and DP: data acquisition and analysis. HH and DP: interpretation of data. HH, SG, KR, and DP drafted the work or substantively revised it. All authors approved the submitted version.
Funding
Funds supporting this study were received from the Swiss National Science Foundation (Grant Number 177163).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We would like to express our sincere gratitude to Prof. Timothy D. McHugh and his team at UCL Centre for Clinical Microbiology for providing initial training and the IC plasmid for the implementation of the MBLA in our institutes.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmed.2022.821776/full#supplementary-material
References
2. Tostmann A, Boeree MJ, Aarnoutse RE, de Lange WC, van der Ven AJ, Dekhuijzen R. Antituberculosis drug-induced hepatotoxicity: concise up-to-date review. J Gastroenterol Hepatol. (2008) 23:192–202. doi: 10.1111/j.1440-1746.2007.05207.x
3. Malherbe ST, Shenai S, Ronacher K, Loxton AG, Dolganov G, Kriel M, et al. Persisting positron emission tomography lesion activity and Mycobacterium tuberculosis mRNA after tuberculosis cure. Nat Med. (2016) 22:1094–100. doi: 10.1038/nm.4177
4. Walter ND, Dolganov GM, Garcia BJ, Worodria W, Andama A, Musisi E, et al. Transcriptional adaptation of drug-tolerant Mycobacterium tuberculosis during treatment of human tuberculosis. J Infect Dis. (2015) 212:990–8. doi: 10.1093/infdis/jiv149
5. Strydom N, Gupta SV, Fox WS, Via LE, Bang H, Lee M, et al. Tuberculosis drugs' distribution and emergence of resistance in patient's lung lesions: A mechanistic model and tool for regimen and dose optimization. PLoS Med. (2019) 16:e1002773. doi: 10.1371/journal.pmed.1002773
7. Vickers MA, Darboe F, Muefong CN, Mbayo G, Barry A, Gindeh A, et al. Monitoring anti-tuberculosis treatment response using analysis of whole blood Mycobacterium tuberculosis Specific T cell activation and functional markers. Front Immunol. (2020) 11:572620. doi: 10.3389/fimmu.2020.572620
8. Ahmed MIM, Ntinginya NE, Kibiki G, Mtafya BA, Semvua H, Mpagama S, et al. Phenotypic changes on Mycobacterium Tuberculosis-specific CD4 T cells as surrogate markers for tuberculosis treatment efficacy. Front Immunol. (2018) 9:2247. doi: 10.3389/fimmu.2018.02247
9. Lienhardt C, Raviglione M, Spigelman M, Hafner R, Jaramillo E, Hoelscher M, et al. New drugs for the treatment of tuberculosis: needs, challenges, promise, and prospects for the future. J Infect Dis. (2012) 205 Suppl 2:S241–9. doi: 10.1093/infdis/jis034
10. Schön T, Miotto P, Köser CU, Viveiros M, Böttger E, Cambau E. Mycobacterium tuberculosis drug-resistance testing: challenges, recent developments and perspectives. Clin Microbiol Infect. (2017) 23:154–60. doi: 10.1016/j.cmi.2016.10.022
11. Perrin FM, Lipman MC, McHugh TD, Gillespie SH. Biomarkers of treatment response in clinical trials of novel antituberculosis agents. Lancet Infect Dis. (2007) 7:481–90. doi: 10.1016/S1473-3099(07)70112-3
12. Wallis RS, Kim P, Cole S, Hanna D, Andrade BB, Maeurer M, et al. Tuberculosis biomarkers discovery: developments, needs, and challenges. Lancet Infect Dis. (2013) 13:362–72. doi: 10.1016/S1473-3099(13)70034-3
13. Acharya MP, Pradeep SP, Murthy VS, Chikkannaiah P, Kambar V, Narayanashetty S, et al. CD38+CD27-TNF-α + on Mtb-specific CD4+ T cells is a robust biomarker for tuberculosis diagnosis. Clin Infect Dis. (2021) 73:793–801. doi: 10.1093/cid/ciab144
14. Goletti D, Lindestam Arlehamn CS, Scriba TJ, Anthony R, Cirillo DM, Alonzi T, et al. Can we predict tuberculosis cure? What tools are available? Eur Respir J. (2018) 52:1089. doi: 10.1183/13993003.01089-2018
15. Horne DJ, Royce SE, Gooze L, Narita M, Hopewell PC, Nahid P, et al. Sputum monitoring during tuberculosis treatment for predicting outcome: systematic review and meta-analysis. Lancet Infect Dis. (2010) 10:387–94. doi: 10.1016/S1473-3099(10)70071-2
16. Friedrich SO, Rachow A, Saathoff E, Singh K, Mangu CD, Dawson R, et al. Assessment of the sensitivity and specificity of Xpert MTB/RIF assay as an early sputum biomarker of response to tuberculosis treatment. Lancet Respir Med. (2013) 1:462–70. doi: 10.1016/S2213-2600(13)70119-X
17. Ahmed MIM, Ziegler C, Held K, Dubinski I, Ley-Zaporozhan J, Geldmacher C, et al. The TAM-TB assay-A promising TB immune-diagnostic test with a potential for treatment monitoring. Front Pediatr. (2019) 7:27. doi: 10.3389/fped.2019.00027
18. Honeyborne I, McHugh TD, Kuittinen I, Cichonska A, Evangelopoulos D, Ronacher K, et al. Profiling persistent tubercule bacilli from patient sputa during therapy predicts early drug efficacy. BMC Med. (2016) 14:68. doi: 10.1186/s12916-016-0609-3
19. Dooley KE, Phillips PP, Nahid P, Hoelscher M. Challenges in the clinical assessment of novel tuberculosis drugs. Adv Drug Deliv Rev. (2016) 102:116–22. doi: 10.1016/j.addr.2016.01.014
20. Honeyborne I, Mtafya B, Phillips PP, Hoelscher M, Ntinginya EN, Kohlenberg A, et al. The molecular bacterial load assay replaces solid culture for measuring early bactericidal response to antituberculosis treatment. J Clin Microbiol. (2014) 52:3064–7. doi: 10.1128/JCM.01128-14
21. Sabiiti W, Azam K, Farmer ECW, Kuchaka D, Mtafya B, Bowness R, et al. Tuberculosis bacillary load, an early marker of disease severity: the utility of tuberculosis molecular bacterial load assay. Thorax. (2020) 75:606–8. doi: 10.1136/thoraxjnl-2019-214238
22. Riou C, Du Bruyn E, Ruzive S, Goliath RT, Lindestam Arlehamn CS, Sette A, et al. Disease extent and anti-tubercular treatment response correlates with Mycobacterium tuberculosis-specific CD4 T-cell phenotype regardless of HIV-1 status. Clin Transl Immunol. (2020) 9:e1176. doi: 10.1002/cti2.1176
23. Ferrian S, Ross M, Conradie F, Vally Omar S, Ismail N, Little F, et al. Frequency of circulating CD4(+)Ki67(+)HLA-DR(-) T regulatory cells prior to treatment for multidrug resistant tuberculosis can differentiate the severity of disease and predict time to culture conversion. Front Immunol. (2018) 9:2438. doi: 10.3389/fimmu.2018.02438
24. Adekambi T, Ibegbu CC, Cagle S, Kalokhe AS, Wang YF, Hu Y, et al. Biomarkers on patient T cells diagnose active tuberculosis and monitor treatment response. J Clin Invest. (2015) 125:1827–38. doi: 10.1172/JCI77990
25. Hiza H, Hella J, Arbués A, Magani B, Sasamalo M, Gagneux S, et al. Case-control diagnostic accuracy study of a non-sputum CD38-based TAM-TB test from a single milliliter of blood. Sci Rep. (2021) 11:13190. doi: 10.1038/s41598-021-92596-z
26. Nikitina IY, Kondratuk NA, Kosmiadi GA, Amansahedov RB, Vasilyeva IA, Ganusov VV, et al. Mtb-specific CD27low CD4 T cells as markers of lung tissue destruction during pulmonary tuberculosis in humans. PLoS ONE. (2012) 7:e43733. doi: 10.1371/journal.pone.0043733
27. Jiang J, Wang X, Wang X, Cao Z, Liu Y, Dong M, et al. Reduced CD27 expression on antigen-specific CD4+ T cells correlates with persistent active tuberculosis. J Clin Immunol. (2010) 30:566–73. doi: 10.1007/s10875-010-9418-1
28. Riou C, Gray CM, Lugongolo M, Gwala T, Kiravu A, Deniso P, et al. A subset of circulating blood mycobacteria-specific CD4 T cells can predict the time to Mycobacterium tuberculosis sputum culture conversion. PLoS ONE. (2014) 9:e102178. doi: 10.1371/journal.pone.0102178
29. Wejse C, Gustafson P, Nielsen J, Gomes VF, Aaby P, Andersen PL, et al. TBscore: Signs and symptoms from tuberculosis patients in a low-resource setting have predictive value and may be used to assess clinical course. Scand J Infect Dis. (2008) 40:111–20. doi: 10.1080/00365540701558698
30. Hiza H, Doulla B, Sasamalo M, Hella J, Kamwela L, Mhimbira F, et al. Preservation of sputum samples with cetylpyridinium chloride (CPC) for tuberculosis cultures and Xpert MTB/RIF in a low-income country. BMC Infect Dis. (2017) 17:542. doi: 10.1186/s12879-017-2642-z
31. Honeyborne I, McHugh TD, Phillips PP, Bannoo S, Bateson A, Carroll N, et al. Molecular bacterial load assay, a culture-free biomarker for rapid and accurate quantification of sputum Mycobacterium tuberculosis bacillary load during treatment. J Clin Microbiol. (2011) 49:3905–11. doi: 10.1128/JCM.00547-11
32. Portevin D, Moukambi F, Clowes P, Bauer A, Chachage M, Ntinginya NE, et al. Assessment of the novel T-cell activation marker-tuberculosis assay for diagnosis of active tuberculosis in children: a prospective proof-of-concept study. Lancet Infect Dis. (2014) 14:931–8. doi: 10.1016/S1473-3099(14)70884-9
33. Motulsky HJ, Brown RE. Detecting outliers when fitting data with nonlinear regression - a new method based on robust nonlinear regression and the false discovery rate. BMC Bioinform. (2006) 7:123. doi: 10.1186/1471-2105-7-123
34. Chamie G, Luetkemeyer A, Walusimbi-Nanteza M, Okwera A, Whalen CC, Mugerwa RD, et al. Significant variation in presentation of pulmonary tuberculosis across a high resolution of CD4 strata. Int J Tuberc Lung Dis. (2010) 14:1295–302.
36. Sinha P, Lönnroth K, Bhargava A, Heysell SK, Sarkar S, Salgame P, et al. Food for thought: addressing undernutrition to end tuberculosis. Lancet Infect Dis. (2021) 21:e318–25. doi: 10.1016/S1473-3099(20)30792-1
37. Kayigire XA, Friedrich SO, Venter A, Dawson R, Gillespie SH, Boeree MJ, et al. Direct comparison of Xpert MTB/RIF assay with liquid and solid mycobacterial culture for quantification of early bactericidal activity. J Clin Microbiol. (2013) 51:1894–8. doi: 10.1128/JCM.03290-12
38. Halliday A, Whitworth H, Kottoor SH, Niazi U, Menzies S, Kunst H, et al. Stratification of latent Mycobacterium tuberculosis infection by cellular immune profiling. J Infect Dis. (2017) 215:1480–7. doi: 10.1093/infdis/jix107
39. Streitz M, Tesfa L, Yildirim V, Yahyazadeh A, Ulrichs T, Lenkei R, et al. Loss of receptor on tuberculin-reactive T-cells marks active pulmonary tuberculosis. PLoS ONE. (2007) 2:e735. doi: 10.1371/journal.pone.0000735
40. Morgan J, Muskat K, Tippalagama R, Sette A, Burel J, Lindestam Arlehamn CS. Classical CD4 T cells as the cornerstone of antimycobacterial immunity. Immunol Rev. (2021) 301:10–29. doi: 10.1111/imr.12963
41. Mahnke YD, Brodie TM, Sallusto F, Roederer M, Lugli E. The who's who of T-cell differentiation: human memory T-cell subsets. Eur J Immunol. (2013) 43:2797–809. doi: 10.1002/eji.201343751
42. Piedra-Quintero ZL, Wilson Z, Nava P, Guerau-de-Arellano M. CD38: An immunomodulatory molecule in inflammation and autoimmunity. Front Immunol. (2020) 11:597959. doi: 10.3389/fimmu.2020.597959
43. Tena-Coki NG, Scriba TJ, Peteni N, Eley B, Wilkinson RJ, Andersen P, et al. CD4 and CD8 T-cell responses to mycobacterial antigens in African children. Am J Respir Crit Care Med. (2010) 182:120–9. doi: 10.1164/rccm.200912-1862OC
44. Garton NJ, Waddell SJ, Sherratt AL, Lee SM, Smith RJ, Senner C, et al. Cytological and transcript analyses reveal fat and lazy persister-like bacilli in tuberculous sputum. PLoS Med. (2008) 5:e75. doi: 10.1371/journal.pmed.0050075
45. Mukamolova GV, Turapov O, Malkin J, Woltmann G, Barer MR. Resuscitation-promoting factors reveal an occult population of tubercle Bacilli in Sputum. Am J Respir Crit Care Med. (2010) 181:174–80. doi: 10.1164/rccm.200905-0661OC
46. Chengalroyen MD, Beukes GM, Gordhan BG, Streicher EM, Churchyard G, Hafner R, et al. Detection and quantification of differentially culturable tubercle bacteria in sputum from patients with tuberculosis. Am J Respir Crit Care Med. (2016) 194:1532–40. doi: 10.1164/rccm.201604-0769OC
47. Getahun H, Harrington M, O'Brien R, Nunn P. Diagnosis of smear-negative pulmonary tuberculosis in people with HIV infection or AIDS in resource-constrained settings: informing urgent policy changes. Lancet. (2007) 369:2042–9. doi: 10.1016/S0140-6736(07)60284-0
48. Caws M, Thwaites G, Dunstan S, Hawn TR, Lan NT, Thuong NT, et al. The influence of host and bacterial genotype on the development of disseminated disease with Mycobacterium tuberculosis. PLoS Pathog. (2008) 4:e1000034. doi: 10.1371/journal.ppat.1000034
Keywords: tuberculosis, treatment monitoring, CD38, CD27, TAM-TB, MBLA
Citation: Hiza H, Hella J, Arbués A, Sasamalo M, Misana V, Fellay J, Gagneux S, Reither K and Portevin D (2022) CD38 Expression by Antigen-Specific CD4 T Cells Is Significantly Restored 5 Months After Treatment Initiation Independently of Sputum Bacterial Load at the Time of Tuberculosis Diagnosis. Front. Med. 9:821776. doi: 10.3389/fmed.2022.821776
Received: 24 November 2021; Accepted: 24 March 2022;
Published: 15 April 2022.
Edited by:
Ravnit Grewal, University of the Free State, South AfricaReviewed by:
Tamar Tak, Leiden University Medical Center, NetherlandsElisa Nemes, University of Cape Town, South Africa
Copyright © 2022 Hiza, Hella, Arbués, Sasamalo, Misana, Fellay, Gagneux, Reither and Portevin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Damien Portevin, ZGFtaWVuLnBvcnRldmluQHN3aXNzdHBoLmNo