 Sigrid Lundgren1,2
Sigrid Lundgren1,2 Ganna Petruk1
Ganna Petruk1 Karl Wallblom1,2José F. P. Cardoso1Ann-Charlotte Strömdahl1Fredrik Forsberg1Congyu Luo1
Karl Wallblom1,2José F. P. Cardoso1Ann-Charlotte Strömdahl1Fredrik Forsberg1Congyu Luo1 Bo Nilson3,4Erik Hartman5Jane Fisher1
Bo Nilson3,4Erik Hartman5Jane Fisher1 Manoj Puthia1
Manoj Puthia1 Karim Saleh1,2
Karim Saleh1,2 Artur Schmidtchen1,2*
Artur Schmidtchen1,2*- 1Division of Dermatology and Venereology, Department of Clinical Sciences Lund, Lund University, Lund, Sweden
- 2Department of Dermatology, Skåne University Hospital, Lund, Sweden
- 3Division of Medical Microbiology, Department of Laboratory Medicine, Lund University, Lund, Sweden
- 4Department of Clinical Microbiology, Infection Control and Prevention, Office for Medical Services, Region Skåne, Lund, Sweden
- 5Division of Infection Medicine, Department of Clinical Sciences Lund, Lund University, Lund, Sweden
Introduction: Inflammation is integral to wound healing; yet, its dynamics in normally healing epidermal wounds are not fully characterized. To this end, we analyzed longitudinal wound fluid samples collected from epidermal suction blister wounds in healthy volunteers.
Materials and methods: A total of 48 control wounds were induced by suction blister method in 24 healthy volunteers—1 on each thigh. Immunoassay, MALDI-TOF mass spectrometry, and microbiological methods were applied to analyze cytokine dynamics, neutrophil activity, and bacterial colonization and levels in wound swabs and dressings during epidermal wound healing, focusing on viable cultivable wound bacteria.
Results: Cytokine levels peaked on day 5, followed by a decline on day 8. Wound exudation, measured by protein content, increased from day 2, peaking on day 5. The neutrophil-derived proteins myeloperoxidase, elastase, and heparin-binding protein (HBP) peaked on day 5, correlating with interleukin (IL)-8, a key neutrophil chemoattractant. The identified bacteria consisted primarily of commensal bacteria, including various staphylococci. Levels of such bacteria rose from day 2, peaking on days 5–8, and aligned with increases in the cytokines tumor necrosis factor α, IL-1β, and IL-10 at the peak of inflammation on day 5.
Conclusions: This study uncovers the coordinated dynamics of cytokines, neutrophil activity, and major commensal bacteria in epidermal wound healing and identifies HBP—a marker for neutrophil activation and endothelial leakage—for the first time in normally healing epidermal wounds.
Introduction
Wound healing is a highly regulated biological process characterized by a series of well-orchestrated events involving inflammation, proliferation, and remodeling, all aimed at restoring tissue integrity and function (1–3). Inflammation, exudation, and neutrophil responses are all important aspects of wound defense and repair, collectively ensuring that the wound is cleansed of debris and pathogens, creating a conducive environment for tissue repair and regeneration (3–6). Central to the early phase of wound healing is the release of key pro-inflammatory cytokines, such as tumor necrosis factor (TNF)-α and interleukin (IL)-6, which promote immune cell recruitment and activation. IL-8, a potent neutrophil chemoattractant, plays a critical role in driving neutrophil migration and function at the wound site. Neutrophils, in turn, can release proteolytic enzymes like neutrophil elastase (NE) and myeloperoxidase (MPO), which contribute to bacterial clearance and tissue remodeling (5, 7, 8).
However, certain disease states, including various types of chronic non-healing wounds, are characterized by specific inflammatory and histopathological profiles (9) For instance, excessive neutrophil activity or prolonged cytokine signaling in these states can lead to tissue damage, highlighting the importance of tightly regulated cytokine and protease dynamics in normal wound healing. Moreover, bacteria can significantly influence these processes by enhancing inflammation, leading to prolonged tissue damage and delayed healing (10, 11).
From a physiological perspective, commensal bacteria can promote skin homeostasis and wound healing (12–16). However, recent findings challenge the belief that commensal bacteria solely benefit wound healing. While Staphylococcus epidermidis colonization can be protective against infections, high-dose applications of commensal bacteria, such as S. epidermidis, to abraded skin can delay barrier repair and induce a chronic wound-like inflammatory state (17). This is consistent with reports that barrier disruption generates an inflammatory environment, which itself promotes pathogen colonization and infection, leading to suppression of the protective mechanism of skin commensals. These findings underscore the complex and context-dependent role of commensals in modulating wound healing (18). In this context, we define colonization as the residence or attachment of bacteria in a wound, whereas infection refers to the proliferation of microorganisms in the wound that results in tissue injury and triggers a host inflammatory response (19).
Epidermal wounds, specifically affecting the outermost skin layer, present a controlled environment to study these interactions. The suction blister model offers a standardized approach to create uniform wounds (20–25), providing valuable insights into the dynamics of inflammation and bacterial colonization during healing. This study was conducted to establish a baseline characterization of inflammatory and bacterial dynamics in epidermal wound healing, providing a foundation for future studies employing the suction blister model or related wound models. We performed a secondary analysis of control samples from a randomized, placebo-controlled clinical trial (NCT05378997) (26) utilizing the suction blister wound model. We used multiplex Meso Scale analysis, ELISA, MALDI-TOF mass spectrometry, and microbiological methods focused on viable, metabolically active commensal bacteria to comprehensively examine cytokine dynamics, neutrophil activity, and bacterial colonization and levels in epidermal wound healing. The study reveals the dynamics of cytokines, neutrophil proteins, and commensal bacteria in epidermal wound healing, identifying heparin-binding protein (HBP), a known marker for neutrophil activation and endothelial leakage, for the first time in normally healing epidermal wounds.
Materials and methods
Trial registration
The trial was prospectively entered into the EudraCT database on 16 November 2021 (EudraCT Number: 2021-004728-14). The registration at ClinicalTrials.gov (NCT05378997) was submitted on 25 April 2022 and posted on 18 May 2022. The registration at ClinicalTrials.gov was retrospectively completed by a few days due to logistical delays in registering all relevant parties, as reported in the published study protocol (26). Both registries are recognized by the ICMJE.
Study design
We analyzed samples collected as part of a randomized, controlled trial designed to test the safety and pharmacokinetics of thrombin-derived C-terminal peptide (TCP)-25 in epidermal suction blister wounds. Twenty-four subjects were included in the study, with two placebo-treated suction blister wounds each. We only included samples from control placebo-treated wounds in this study. The study design is described in detail in the published study protocol (26).
Formation of epidermal suction blister wounds
Two blister wounds were created on the medial aspect of each thigh using a Model NP-4 (Electronic Diversities, Finksburg, MD) suctioning device (27) as described in the published study protocol (26). On each thigh, one wound was randomized to receive active drug, and the other was randomized to receive control gel without the drug substance. Since drug-treated wounds were not included in this study, we included 48 control wounds from 24 subjects.
Swab procedure and analysis
The wound was swabbed on day 1 (immediately after blister formation) and on days 2, 3, 5, 8, and 11 using a sterile cotton swab (Selefa, Stockholm, Sweden) per standard methods (28) and extracted as described in the supplement. Swabbing is a common technique for collection of bacteria from wounds and skin, yielding bacterial information that is comparable with other methods (29, 30) (Supplementary Figure 1). Bacterial levels were quantified immediately after extraction as described in the supplement, following standard methods (28, 31).
Dressing extraction procedure and analysis
Wounds were covered with Mepilex dressing (Mölnlycke Healthcare, Gothenburg, Sweden) in between study visits. Dressings were changed on days 2, 3, 5, 8, and 11. At each dressing change, the dressing was removed from the wound and dressing fluid was extracted as described in the supplement. Bacterial levels were quantified immediately after extraction in fresh (never frozen) dressing fluid as described in the supplement. Cytokines (interferon gamma [IFN]-γ, IL-1β, IL-2, IL-4, IL-6, IL-8, IL-10, IL-12p70, IL-13, tumor necrosis factor [TNF]-α), protein content, and neutrophil proteins (neutrophil elastase [NE], myeloperoxidase [MPO], and HBP) were quantified in the stored dressing fluid samples that contained protease inhibitor, as described in the supplement.
Rationale for the chosen bacterial quantification and identification strategy
The scope of this part of the study was limited to evaluating the cultivable bacterial burden on the wound surface and in dressings, with a particular focus on commensals of normally healing wounds. Therefore, to quantify only the viable cultivable bacteria, quantitative bacterial counts was the method of choice for this study (31, 32). With a specific aim to identify and discriminate between the major cultivable aerobic bacteria, including Staphylococcus aureus and various commensal staphylococci, Matrix-Assisted Laser Desorption Ionization—Time Of Flight (MALDI-TOF) mass spectrometry (MS) associated with MALDI Biotyper software was used for the rapid and specific identification at the species level (33, 34).
Identification of major cultivable bacteria
Swab and dressing fluid samples were streaked on a blood agar plate, and six colonies were selected for analysis by MALDI-TOF to identify the major cultivable bacteria, as described in the supplement. Blood agar was selected as the growth medium—as is commonly used in clinical microbiology laboratories—as it is a rich medium that allows growth of most common clinically relevant pathogens (31).
Wound imaging
A disposable centimeter-scale ruler was placed on the skin near the wounds. Two images were taken of each set of wounds on each leg at a distance of 35 cm with a Canfield Twin Flash camera (Canfield Scientific, Parsippany-Troy Hills, NJ, USA) with a modified version of a Canfield close-up scale (35).
Statistics
Summary data were plotted as median and interquartile range. For correlation between two variables, Spearman correlation coefficients were determined. Although each participant had two control wounds that were included in this study, individual wounds were considered independent biological replicates in all analyses (n = 48 wounds). Due to the non-normal distribution of the data (determined by visual inspection of the data), non-parametric analyses were used throughout. All statistical tests were two-tailed. Data were analyzed and plotted using GraphPad Prism 10 software. When a logarithmic axis was used, zero values were replaced with 1 to enable visualization of all data points on the graph but were not replaced in any statistical analyses.
With regard to the changes in bacterial levels, logarithmic transformation was applied to the data, and a nonparametric Mann-Kendall test was applied to test for trend.
Changes in cytokine and neutrophil protein levels in dressing fluid were analyzed by a matched-pairs Wilcoxon test at selected time points. Due to the exploratory nature of the study, p-values should be considered hypothesis-generating only. P-values adjusted for multiple comparisons by the Holm-Šídák method are presented in the supplement.
Ethical considerations
The clinical trial and collection of specimens for the biobank were approved by the Swedish ethical review authority (Etikprövningsmyndigheten application number 2022-00527-01). Written informed consent was received from all subjects prior to participation.
Results
Inflammation
To determine how inflammation develops over time in normally healing epidermal wounds, we measured the levels of 10 different cytokines in the dressing fluid on days 2, 3, 5, and 8. Because there was very little exudation on day 11, we did not measure cytokine levels on this day as they were likely to be undetectable. Although the individual cytokines had vastly different levels, ranging from about 1 ng/mL (IL-4) to over 10,000 ng/mL (IL-8) at their peak, all cytokines followed similar dynamics over time (Figure 1). Most measured cytokines remained at approximately the same level from day 2 to 3 (IL-1β, IL-2, IL-8, IL-12p70, IL-13, TNF-α), while some declined slightly on day 3 relative to day 2 (IL-4, IL-6, IL-10). IFN-γ, in contrast, increased steadily from day 2 to 5. In spite of the differences in initial dynamics, all measured cytokines reached their peak level on day 5 and then declined on day 8. Most cytokines were significantly higher on day 5 than on day 2 and significantly lower on day 8 than on day 5 (Supplementary Table 1).
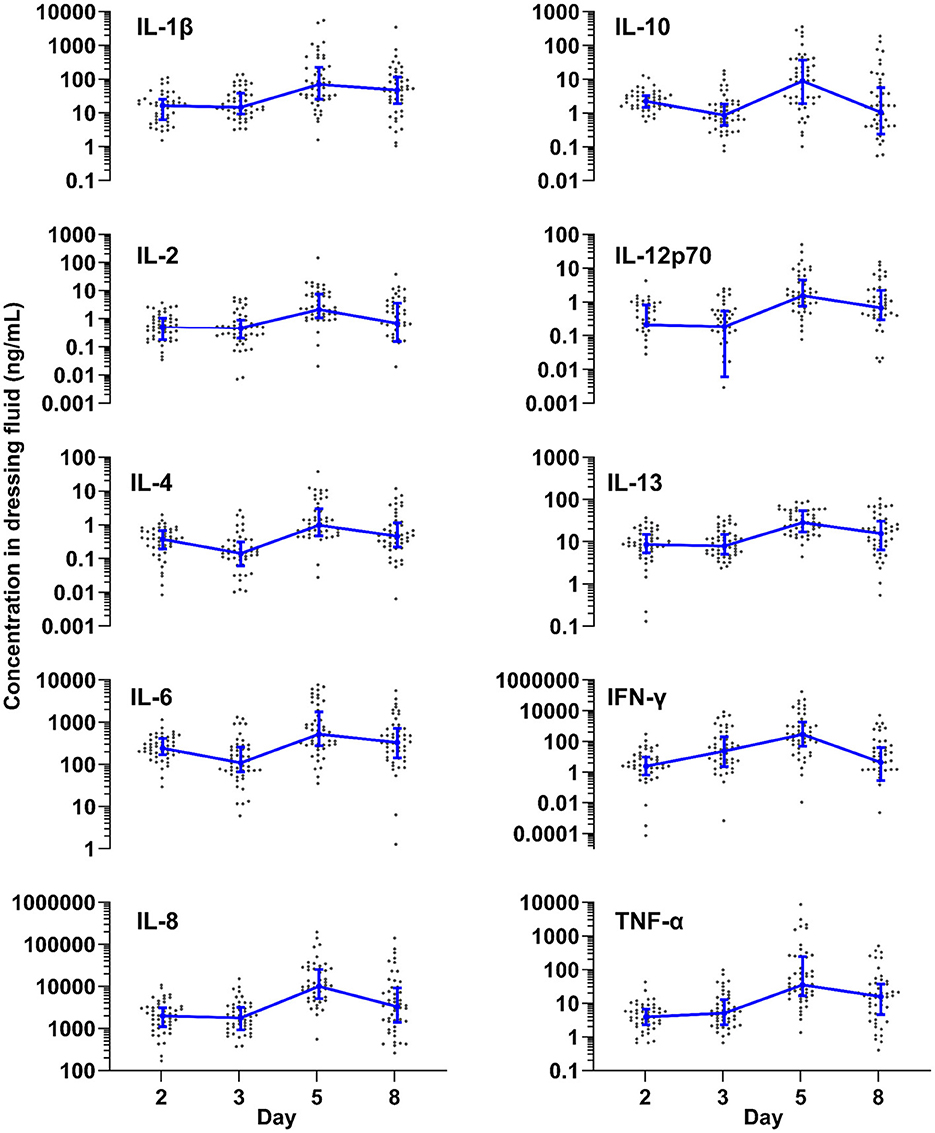
Figure 1. Inflammation over time. Levels of ten different cytokines measured by Meso Scale in dressing fluid over time. Each data point is the wound fluid cytokine concentration in a single wound, while the blue line is the median of all wounds that day (n = 48), and whiskers are the interquartile range.
To confirm that the measured cytokine levels are not dependent on the amount of protein content in the dressing fluid, we normalized the levels of each cytokine to the protein content of that same sample (Supplementary Figure 2A and Supplementary Table 1). When normalized for protein content, the overall dynamics of the cytokines were similar to the non-normalized data at the early time points, reaching their peak at day 5. The normalized level of some cytokines (IL-1 β, IL-4, IL-6, IL-12p70, TNF- α) did not decrease on day 8, unlike un-normalized levels (Supplementary Table 1).
Wound exudation
To determine how the level of wound exudation changes over time, we measured protein content in dressing fluid as a proxy for wound fluid accumulation. Because all dressing fluids were extracted with the same volume of extraction buffer, the amount of protein indicates the accumulated amount of wound fluid in the dressing and is not affected by possible evaporation through the Op-Site film covering the dressing. The results indicated that the level of wound exudate was moderate on days 2 and 3 and then peaked on day 5 before dropping on day 8 (Figure 2A). On day 11, the dressings were very dry and had near-zero levels of protein content, indicating very low exudation on this day.
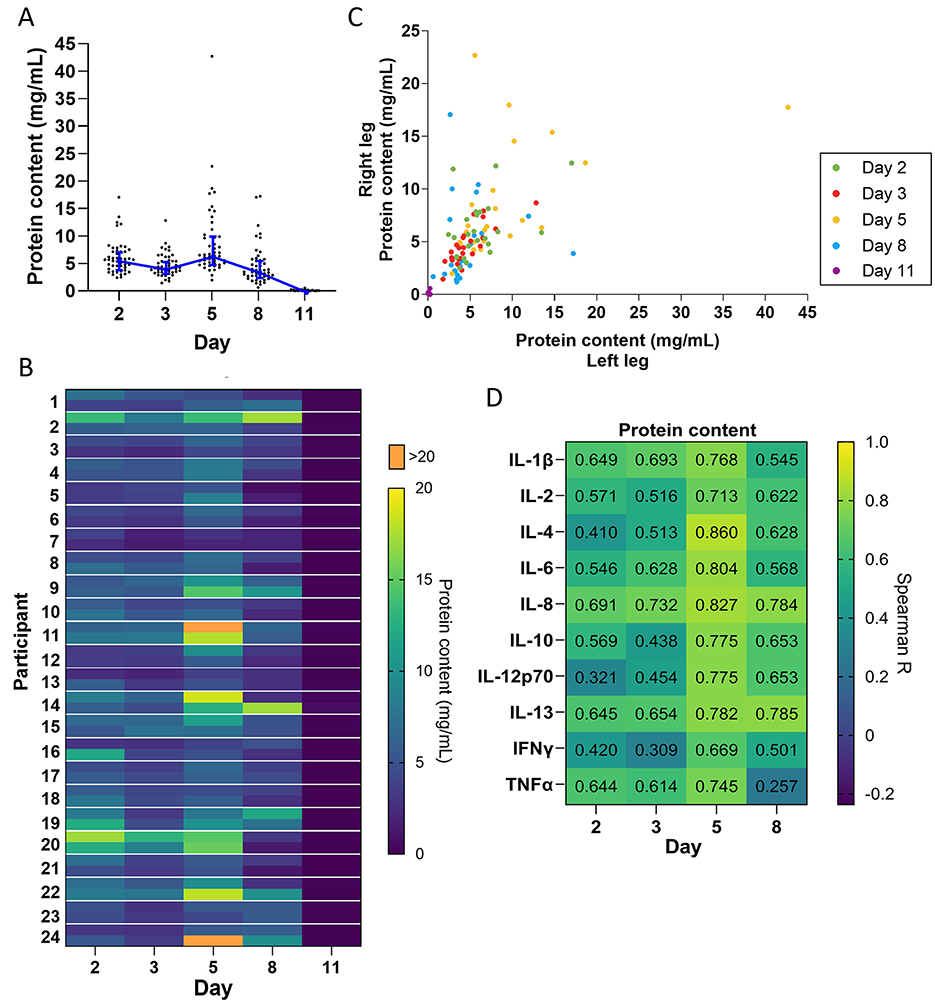
Figure 2. Exudation over time. (A) Protein concentration quantified in dressing fluid from each wound at each time point. Each data point is protein concentration in a single wound, while the blue line is the median of all wounds that day (n = 48), and whiskers are the interquartile range. (B) Heat map of protein concentration quantified in dressing fluid samples from each wound. The left and right wound are each presented as a single cell on the top and bottom of each row, respectively. (C) Scatterplot comparing protein concentration in dressing fluids from wounds on the left leg vs. the right leg in each participant. (D) Spearman correlation coefficients comparing levels of each cytokine with protein content in that wound (n = 48).
When plotted as a heatmap, protein content showed considerable individual variation and appeared to be similar between left and right wounds (Figure 2B). To confirm this, we plotted protein levels in the left and right wound as a scatterplot (Figure 2C) and determined the Spearman correlation coefficient on each day and overall (Table 1). The data indicated a moderate to high correlation between left and right wounds on all days (R = 0.356–0.803), except for day 11, which had many zero values. The data also indicated a strong correlation overall when all days were included (R = 0.791), indicating that there is some variation in exudation between the left and right wounds in each individual as the wound heals but that overall exudation levels do not differ greatly between wounds on the left and right leg.
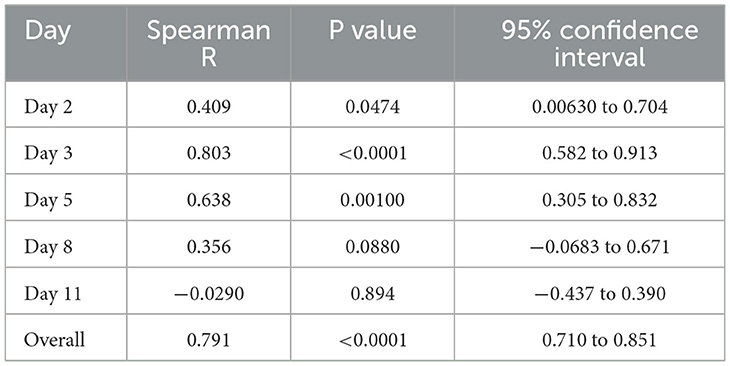
Table 1. Correlation coefficients for protein content in left vs. right leg.
Inflammation is known to induce capillary leakage and subsequent exudation. We determined the correlation coefficient between the level of each cytokine and protein content of the dressing fluid. We found that there was a moderate to strong correlation between cytokine levels and protein content on all days (R = 0.257–0.860), particularly on day 5 (R = 0.669–0.860) (Figure 2D).
Neutrophils
Neutrophils play a key role in wound healing and bacterial control. Therefore, we measured the levels of three different neutrophil proteins (MPO, NE, and HBP) on days 2, 3, 5, and 8 (Figure 3A). Due to the small amount of exudation on day 11, proteins were likely to be undetectable, so we did not measure neutrophil protein levels on this day. MPO and NE remained at approximately the same level from day 2 to 3, peaking on day 5, and then dropping sharply on day 8. In contrast, HBP increased steadily until it peaked on day 5 and then dropped on day 8. The level of NE and HBP was significantly higher on day 5 than on day 2 and significantly lower on day 8 than on day 5 (Supplementary Table 2).
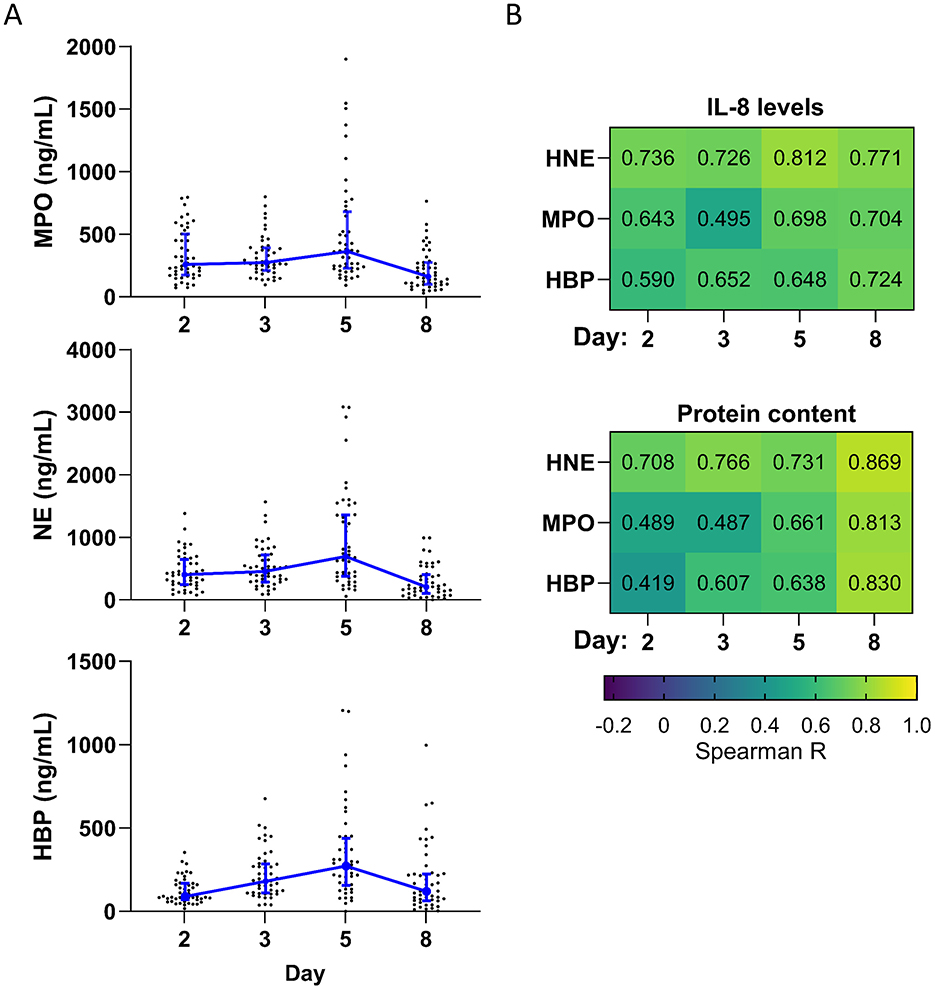
Figure 3. Neutrophil proteins over time. (A) Levels of three different neutrophil proteins measured in dressing fluid over time. Each data point represents the concentration of a specific neutrophil protein in a single wound, while the blue line is the median of all wounds that day (n = 48), and whiskers are the interquartile range. (B) Spearman correlation coefficients comparing levels of each neutrophil protein with IL-8 (top) and protein content (bottom) (n = 48).
The levels of each neutrophil protein were moderately to strongly correlated with levels of IL-8, a potent chemoattractant for neutrophils, on each day (R = 0.495–0.812) (Figure 3B). Neutrophil recruitment and activation are associated with neutrophil extravasation from the blood to the tissues and consequent capillary leakage. We thus determined the correlation coefficient between the level of each neutrophil protein and protein content of the dressing fluid (a proxy for exudation) (Figure 3B) and found that there was a moderate to strong correlation between neutrophil protein levels and protein content on all days (R = 0.419–0.869), particularly on day 8 (R = 0.813–0.867). Notably, this correlation was stronger than it was for most cytokines and occurred later (Figure 2D vs. Figure 3B), suggesting that inflammation and neutrophil recruitment/activation may drive exudation in different stages of wound healing.
To confirm that the measured neutrophil protein levels are not dependent on the amount of protein content in the dressing fluid, we normalized the levels of each neutrophil protein to the protein content of that same sample (Supplementary Figure 2B and Supplementary Table 2). The overall dynamics changed slightly, with normalized neutrophil proteins reaching their peak level on day 3, instead of day 5 (Supplementary Table 2).
Bacterial levels
To explore how levels of aerobic cultivable bacteria in normally healing epidermal wounds change over time, we quantified the number of bacteria in swab and dressing fluid extract samples. Immediately after formation of the wound (day 1), the bacterial levels were zero in most of the swab samples (Figure 4A, Supplementary Table 3). Subsequent samples revealed the rapid reconstitution of cultivable bacteria over time already 1 day after the wounds were formed (day 2), corresponding to a median of 2.9 × 103 CFU per swab and 1.9 × 104 CFU per dressing. Thereafter bacterial levels in the wound increased steadily until day 8, when they reached a peak, corresponding to a median of 1.7 × 106 CFU per swab and 1.6 × 107 CFU per dressing. On day 11, bacterial levels decreased slightly in swabs to a median of 1.3 × 106 CFU per swab. They decreased to a much larger extent in dressings on day 11, to a median of 1.4 × 106 CFU per dressing. By non-parametric Mann-Kendall test, bacterial levels in swab and dressing fluid samples showed a significantly increasing trend over time (Supplementary Table 4).
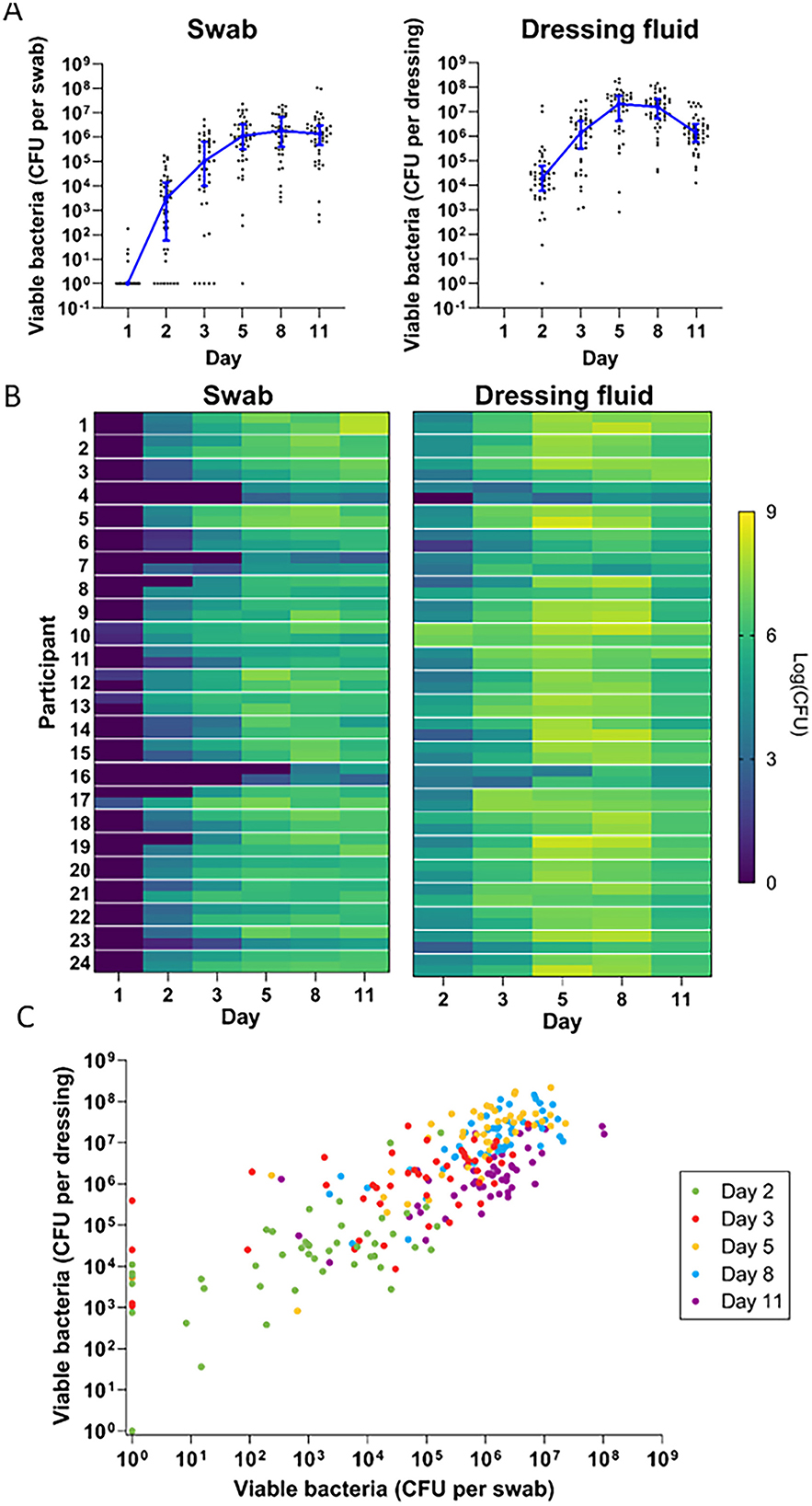
Figure 4. Bacterial development over time. (A) Quantitative bacterial counts in swab samples (left) and dressing fluid samples (right) over time. Each data point is the bacterial level measured in a single wound, while the blue line is the median of all wounds that day (n = 48) and whiskers are the interquartile range. Zero values were replaced with 1 in the graph to enable visualization on the logarithmic axis. (B) Heat map of quantitative bacterial counts in swab samples (left) and dressing fluid samples (right) from each wound. Data from the left and right wound are each presented in separate cells at the top and bottom of each row, respectively. (C) Scatterplot showing quantitative bacterial counts in swabs vs. quantitative bacterial counts in dressing fluid in each wound. Zero values were replaced with 1 in the graph to enable visualization on the logarithmic axis. Each day is indicated in a different color.
Heat maps of bacterial levels in each wound showed considerable individual variation (Figure 4B). Generally, wounds with low bacterial levels in swabs also appeared to have low bacterial levels in dressing fluids, and vice versa. To confirm this, we plotted bacterial levels in swabs and dressings as a scatterplot (Figure 4C) and determined the Spearman correlation coefficient on each day and overall (Table 2). The data indicated a strong correlation between bacterial levels in swab and dressing fluid samples on each day (R = 0.607–0.705) and overall (R = 0.794).
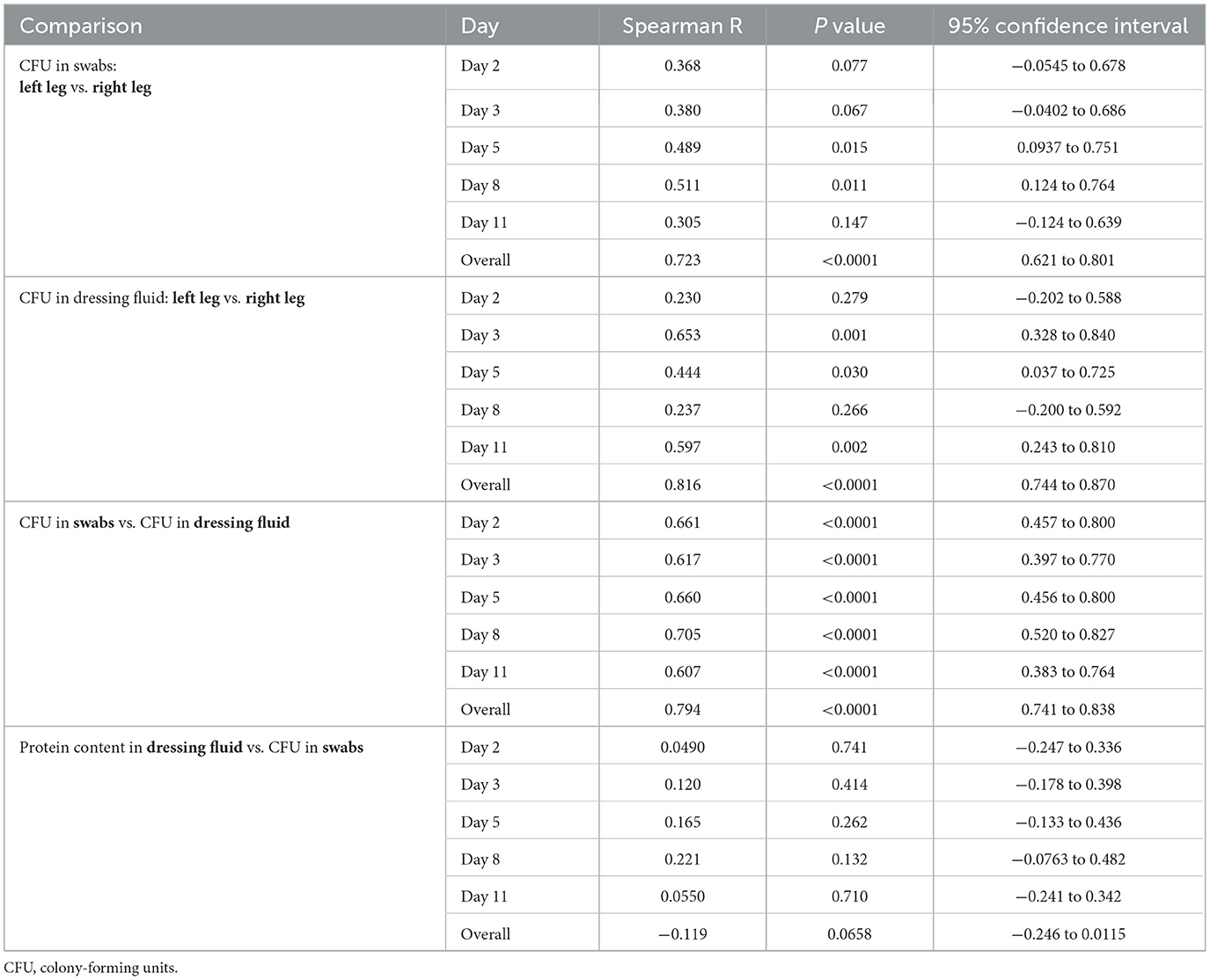
Table 2. Correlation coefficients for bacterial counts in various samples per day and overall.
The bacterial levels that were plotted on the heatmap appeared to be similar between the left and right wounds in most participants but with some variation. To confirm this, we determined the Spearman correlation coefficient on each day and overall (Table 2). The data indicated a weak to moderate correlation between left and right wounds on each day in swabs (R = 0.305–0.511) and dressings (R = 0.230–0.653) but a strong correlation overall in both swabs (R = 0.723) and dressings (R = 0.816) when all days were included, indicating that there is moderate variation between left and right wounds in each individual as the wound heals but that overall bacterial levels do not differ greatly between wounds on the left and right legs, corroborating the data presented in Figure 4C and Table 2, which showed no difference in median bacterial levels between wounds on the left and right legs.
Identity of bacteria
To examine the major cultivable bacterial species that colonize wounds during the healing process, we analyzed bacterial colonies that grew from streaks of each swab and dressing sample on days 3 and 8. Overall, the identified bacterial species were dominated by commensal species, such as S. epidermidis, which was found in swab and dressing samples from the majority of wounds on days 3 and 8 and in swabs and dressings (Table 3; see also Supplementary Figure 3 for a complete representation of the identified bacteria). Some wounds also contained S. aureus and Bacillus sp., which can sometimes be pathogenic, although no signs of infection were observed in these wounds (Supplementary Figure 4). On day 8, we observed an increase in the number of colonies that were identified as S. aureus and the commensal Corynebacterium sp. (Supplementary Figure 3). On examination of images of the wounds over time, we noted that in spite of differences in the cultivable bacteria at the wound surface and in dressings, all wounds healed at roughly the same pace and had a similar appearance at each stage (35).
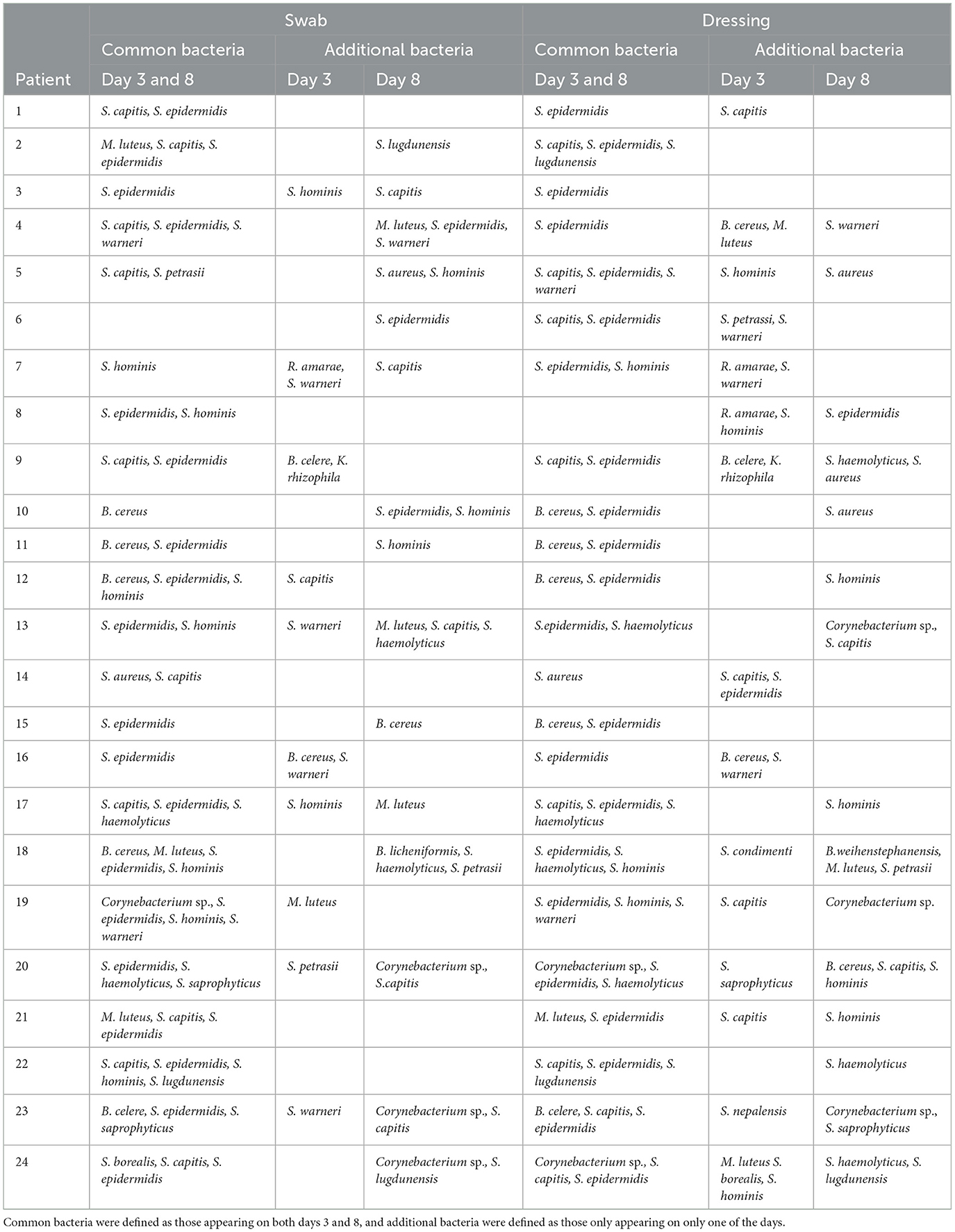
Table 3. Comparison of similar MALDI-TOF-identified bacterial species during study day 3 and day 8 for both swab and dressing fluid samples.
Correlation between inflammation, exudation, neutrophil proteins, and bacterial levels
To determine whether the observed bacterial levels affect inflammation, we determined the correlation coefficient between the level of each cytokine and the bacterial level detected in swabs in that wound and plotted the resulting correlation coefficients as a heatmap (Figure 5A). For most cytokines, correlations with bacterial counts were minimal or absent on days 2, 3, and 8, with only a weak correlation observed on day 5. However, certain cytokines showed moderate correlations with bacterial counts, including TNF-α on days 3 and 5 (R = 0.475 and 0.488, respectively), IL-10 on day 5 (R = 0.531), and IL-1β on days 5 and 8 (R = 0.497 and 0.480, respectively). The correlation coefficients increased slightly for most cytokines when they were normalized to protein content (Supplementary Figure 2C). In contrast, we found little to no correlation between bacterial levels in swabs and neutrophil proteins on each day (R = 0.011–0.353).
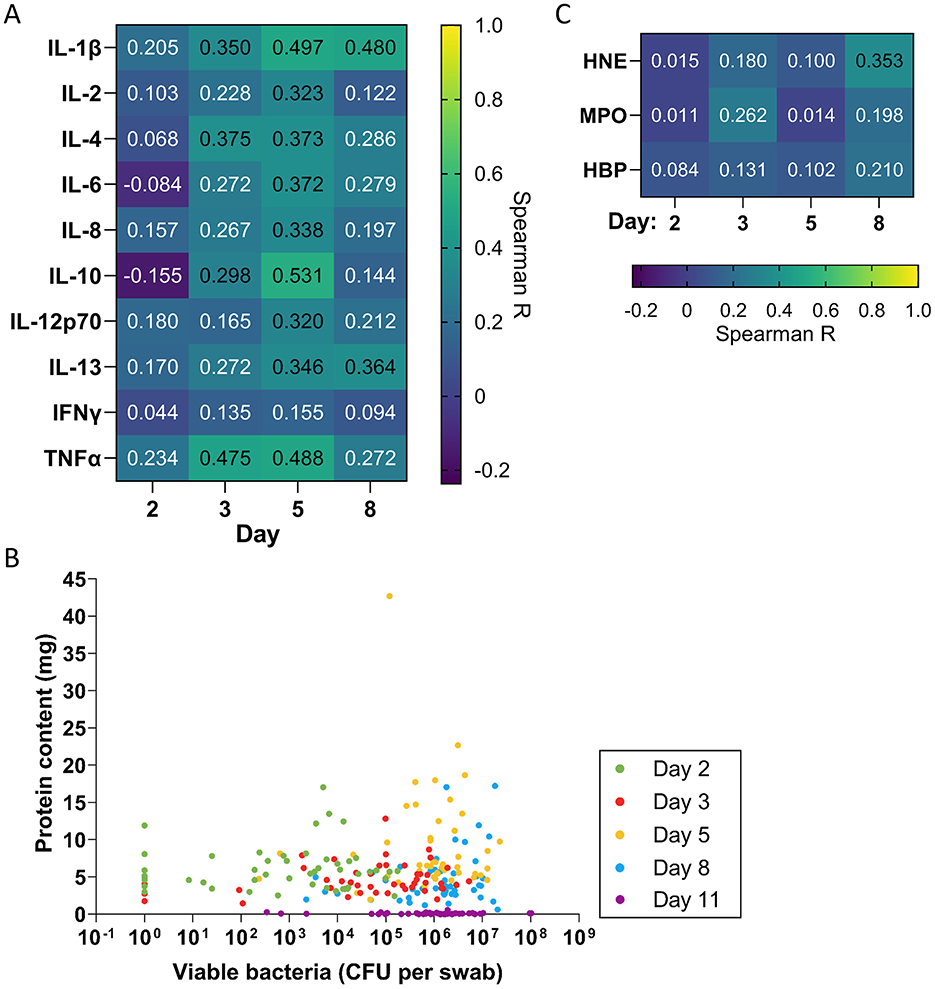
Figure 5. Correlation of inflammation, exudation and neutrophil proteins with bacterial levels. (A) Spearman correlation coefficients comparing levels of each cytokine with quantitative bacterial counts measured in swabs (n = 48). (B) Scatterplot comparing protein concentration in dressing fluids vs. quantitative bacterial counts measured in swabs from each wound. Zero values were replaced with 1 in the graph to enable visualization on the logarithmic axis. (C) Spearman correlation coefficients comparing levels of each neutrophil protein with quantitative bacterial counts measured in swabs (n = 48).
To explore whether the observed level of viable bacteria that were detected on the wound surface affects the level of exudation, we plotted bacterial levels that were measured in swabs against protein content as a scatterplot (Figure 5B) and determined the Spearman correlation coefficient on each day and overall (Table 2). The data indicated that there was weak to no correlation between bacterial levels and protein content on each day (R = 0.049–0.221) and no correlation overall (R = −0.119). In addition to the bacteria directly on the wound surface, bacteria in the dressings could contribute to the induction of inflammation, potentially releasing various proinflammatory products, such as lipoteichoic acid (LTA) and peptidoglycan. Thus, we examined the correlation of cytokine levels and neutrophil proteins with bacterial counts in dressing fluid (Supplementary Figure 5A). In general, the correlations between cytokines and dressing bacterial loads were similar to their correlation with bacterial levels that were measured in swabs, with slightly higher correlations observed for TNF-α, IL-10, and IL-1β at the peak of inflammation on day 5 (Supplementary Figures 5A, B).
Discussion
In this study, we comprehensively describe the dynamics of exudation, inflammation, neutrophil proteins, and bacteria in epidermal suction blister wounds and their interrelatedness during the wound healing process. Inflammation plays a key role in the wound healing process. To assess inflammation over time in normally healing epidermal wounds, we measured levels of 10 cytokines in dressing fluid on days 2, 3, 5, and 8. While cytokine levels varied, they showed similar overall trends: most cytokines stayed constant from days 2 to 3, although some (IL-4, IL-6, IL-10) declined slightly. IFN-γ, however, increased steadily until day 5. All cytokines peaked on day 5 before decreasing on day 8.
Inflammation also plays a role in driving wound exudation, with pro-inflammatory cytokines contributing to this process. Therefore, because wound exudate contains plasma-derived proteins like albumin, expressing cytokine levels in dressing fluid relative to total protein can introduce a confounding factor. As exudation increases, the dilution of cytokines by the influx of plasma proteins can obscure the actual amounts of cytokines that are released in the wound. To mitigate this effect, we present cytokine levels in two complementary ways: per mL to reflect their absolute concentration and per mg of total protein to account for variations in protein content due to exudation. Using both approaches provides a nuanced representation of cytokine dynamics and strengthens the conclusions of the analysis. We found that the overall dynamics of the cytokines remained consistent, even after normalization to protein content, indicating that the trends that we observed were not significantly influenced by wound exudate dilution. This suggests that the inflammatory response, as reflected by cytokine levels, is a robust process that correlates with wound healing progression independently of variations in wound exudate volume. Overall, our findings are in line with several studies that have explored different cytokines and their dynamics in wound healing (5, 7, 8, 20).
Wound exudation is a critical aspect of the wound healing process (6). We found that exudation, as indicated by the level of protein content in dressing fluid, was moderately high already on day 2 and then peaked on day 5, before declining on day 8. Inflammation, as indicated by cytokine levels, correlated with protein content in dressing fluid, particularly on day 5, when inflammation was highest, corroborating the link between inflammation and capillary leakage, which in turn leads to exudation (6).
Neutrophils play a crucial role in wound healing and bacterial control, and both neutrophil infiltration and activation are known to increase in the inflammatory stage of wound healing (3–5). We observed that neutrophil protein levels, which could be attributed to increased neutrophil recruitment and activation, followed similar dynamics as inflammation. MPO and NE increased on day 5, while HBP began to increase already on day 3. These dynamics are consistent with the fact that HBP is stored in secretory vesicles, which have a high propensity for release, while MPO and NE are stored exclusively in azurophilic granules, which have a low propensity for release (36). Neutrophil protein levels correlated strongly with IL-8, a potent chemoattractant for neutrophils.
A novel observation in this study is the identification of HBP in epidermal wounds (37). HBP is known to increase vascular permeability, leading to capillary leakage and subsequent exudation. This mechanism has been observed in various conditions, including burn wounds and sepsis. For instance, studies have shown that HBP, released from activated neutrophils, is increased in severe burn injuries (38), as well as in non-healing venous wounds (39). Similarly, in sepsis, elevated levels of HBP are associated with increased endothelial permeability, leading to fluid leakage and organ dysfunction (40). Although its role in vascular permeability is well established, the specific association between HBP and physiological exudation remains underexamined, meriting further study of this potential link in epidermal wound healing.
The understanding that bacteria, particularly various staphylococcal species, can play a role in modulating normal wound healing (16–18, 41–44) guided our focus on viable aerobic bacteria in this study. While this approach naturally limits the scope of detection and potentially underrepresents anaerobes and non-culturable species compared with broader methods like amplicon or metagenomic sequencing—which are capable of identifying anaerobes and non-culturable organisms—it provides a targeted and clinically relevant analysis of the growth of these key commensals. This is particularly important, because unlike bacterial DNA, the presence of viable bacteria reflects bacterial metabolic activity, which could have direct implications for wound healing dynamics. In addition, the skin microbiome is unique in its composition and activity compared with other human microbiomes. Notably, the skin has a lower proportion of viable bacteria, meaning that a significant fraction of bacterial DNA that is present on the skin surface may come from non-viable organisms.
Traditional sequencing methods, such as 16S rRNA gene sequencing, can overestimate the richness and diversity of the skin microbiome by detecting DNA from dead or inactive bacteria (45), which do not influence biological processes of wound healing—for example, by producing various proteases (46). This distinction between live and dead bacterial populations is crucial when studying the impact of commensals like S. epidermidis on healing. Indeed, the repopulation of the skin microbiome after a disturbance is driven by an underlying viable bacterial population (45). In this context, our focus on viable, cultivable bacteria was reasonable and well justified and allowed us to gain insight into the growth kinetics of commensal species like S. epidermidis during normal wound healing.
At the onset of wound formation, minimal amounts of cultivable bacteria were observed on the wound surface, likely due to the removal of the epidermis and prior skin disinfection with ethanol. Shortly after wounding, bacterial colonization was detected, with levels progressively increasing until day 8, followed by a slight decline by day 11. Although there was some variation in the patterns of cultivable bacteria between individuals, S. epidermidis was consistently dominant, and most wounds contained bacteria that are typically classified as skin commensals. This finding aligns with the view that commensals are conducive to normal healing processes (41–44, 47, 48).
Notably, consistent with recent findings on the pro-inflammatory roles of commensal bacteria (17), we observed a moderate correlation between bacterial levels on day 5 and the pro-inflammatory cytokines TNF-α and IL-1β, suggesting that bacteria can influence inflammation even in normally healing epidermal wounds. Interestingly, a similar correlation was found with IL-10, an anti-inflammatory cytokine that is known to promote a regenerative phenotype in experimental dermal wounds (49). It is also important to note that several other cytokines showed little to no correlation with bacterial levels, underscoring the complex, and likely selective, cytokine responses that are induced by bacteria in human epidermal wounds
Interestingly, previous studies on wound complications in skin grafting procedures in patients have indeed shown that elevated bacterial levels, largely consisting of commensals, are associated with wound complications (50, 51), including increased inflammation, as reflected by elevated levels of TNF-α and IL-1β (52). These findings underscore the dual role of commensals in both facilitating and, under certain conditions, potentially hindering wound healing. It should also be noted, however, that some wounds harbored bacteria that sometimes can act as pathogens, such as S. aureus, which often underlies postoperative infections (53). However, it is also worth noting that the presence of S. aureus does not define infection per se, as the bacterium can, like other commensals, also occur in normally healing wounds (52). Moreover, it has been shown that like commensals, S. aureus can enhance skin regeneration through IL1β-keratinocyte-dependent IL1R/MyD88 signaling (13).
Taken together, these findings collectively highlight the highly dynamic nature of the wound healing process. Bacterial colonization, inflammation, neutrophil activity, and exudation appear to be interrelated but not solely dependent on one another. From a methodological perspective, our study also revealed a strong correlation between the observed levels of the studied bacteria in swab and dressing fluid samples on each day and overall. Given this correlation, analysis of dressing fluid bacteria could be an alternative way to collect information about bacteria in the wound environment. Furthermore, while intra-individual variation existed between the left and right wounds in each participant, shown by the poor correlation between bacterial levels in wounds on the left and right legs, overall bacterial dynamics followed the same trend on both legs.
A key strength of this study is the use of a suction blister model, which allowed for the creation of standardized wounds in human subjects and facilitated a detailed, multifaceted analysis of bacterial dynamics and host factors throughout the healing process. However, there are limitations to consider. First, the bacterial levels that were observed here may not be representative of bacterial levels in other wound types, particularly since the occlusive dressings and hydrogel that were used in this study could have enhanced the moisture in the wound environment, potentially influencing bacterial growth (54–56). Further, the absence of a natural healing control group (i.e., wounds without occlusive dressing or hydrogel) merits a discussion on how dressing materials and hydrogels can affect inflammation and the dynamics of bacterial colonization.
Moisture is a well-known factor for bacterial proliferation, as occluded skin displays a significant increase in bacterial numbers after just 1 day (55). It is therefore possible that the moist environment under the occlusive polyurethane-based dressing that was used here promoted bacterial growth compared with intact skin, which naturally harbors fewer bacteria (57, 58). Our choice of dressings may have thus affected the observed bacterial dynamics. As re-epithelialization progressed and exudation diminished, the decrease in bacterial levels by day 11 could possibly reflect both reduced access to nutrients in wound fluid and a shift in the host immune response.
Moreover, the wounds in this study received a hydroxyethyl cellulose (HEC)-based gel, which added additional moisture. HEC is widely recognized for its high biocompatibility and low immunogenicity (59), ensuring that it did not introduce any confounding effects beyond moisturizing the wound, and similar inert hydrogels are often used in the clinic. Thus, these results represent a clinically relevant scenario, in which moisture contributes to the overall dynamics of bacterial colonization, and justify our model's representativeness for normal healing.
A minor limitation regarding the statistical design of our study was our treatment of each wound as an independent biological replicate, given its exploratory nature. Our analyses thus do not consider the inherent dependence of samples from the same individual.
Finally, this study focused solely on culturable aerobic bacteria. As discussed above, culture-based methods can underestimate bacterial presence, meaning that the total bacterial burden (including viable and non-viable bacteria) in these wounds could be higher than reported—the absence of 16S rRNA sequencing or other culture-independent methods thus limits the comprehensiveness of our microbial characterization. Future studies could address these limitations by employing amplicon-based sequencing, shotgun metagenomics, or metatranscriptomics (60) to provide a more complete profile of the microbiome during epidermal wound healing.
Conclusions
In conclusion, this study highlights the complex interplay between inflammation, exudation, neutrophil activity, and bacterial colonization in normal epidermal wound healing. Cytokine levels peaked on day 5, mirroring neutrophil-derived proteins, such as MPO, NE, and HBP. The correlation between the neutrophil chemoattractant IL-8 and neutrophil activity underscores the role of neutrophils in this process. Additionally, the identification of HBP—a key player in promoting vascular permeability and exudation—offers novel insights into its potential role in wound healing. The dynamics of bacteria, such as S. epidermidis, further underline the presence of commensals in normal wound healing, while possible relationships between bacterial levels and the cytokines TNF-α, IL-1β, and IL-10 point to the potential for bacterial influence on inflammation in some contexts.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: http://doi.org/10.5281/zenodo.10283373, reference number 10283373.
Ethics statement
The studies involving humans were approved by Swedish ethical review authority (Etikprövningsmyndigheten application number 2022-00527-01). The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
SL: Conceptualization, Methodology, Investigation, Validation, Resources, Project administration, Writing – original draft, Writing – review & editing. GP: Conceptualization, Formal analysis, Investigation, Validation, Visualization, Supervision, Project administration, Writing – review & editing. KW: Conceptualization, Methodology, Investigation, Validation, Resources, Supervision, Project administration, Writing – original draft, Writing – review & editing. JFPC: Formal analysis, Investigation, Validation, Data curation, Visualization, Writing – review & editing. A-CS: Investigation, Validation, Writing – review & editing. FF: Investigation, Validation, Writing – review & editing. CL: Formal analysis, Visualization, Writing – review & editing. BN: Investigation, Validation, Writing – review & editing. EH: Visualization, Writing – review & editing. JF: Formal analysis, Data curation, Visualization, Project administration, Writing – original draft, Writing – review & editing. MP: Conceptualization, Methodology, Writing – review & editing. KS: Conceptualization, Resources, Supervision, Writing – review & editing. AS: Conceptualization, Resources, Supervision, Project administration, Funding acquisition, Writing – original draft, Writing – review & editing.
Funding
The authors declare that financial support was received for the research and/or publication of this article. The exploratory study presented here was supported by grants from the Swedish Research Council (project 2017-02341, 2020-02016), Edvard Welanders Stiftelse and Finsenstiftelsen (Hudfonden), the Royal Physiographic Society, the Crafoord and Österlund Foundations, and the Swedish Government Funds for Clinical Research (ALF). Xinnate AB provided funding for the clinical safety study conducted at the Clinical Trial Unit at Skåne University Hospital in Lund, which supplied the control samples used in this study.
Acknowledgments
We would like to thank Dr. Andreas Sonesson for valuable discussions and Susanne Erdmann and Anne Nielsen and other personnel in the Department of Dermatology, Lund. Lastly, we would also like to thank Åsa Håkansson and Kerstin Weber and other personnel at the Clinical Trial Unit at Skåne University Hospital Lund for support and use of their facilities during the clinical study. We thank Magnus Pettersson (senior biostatistician, fil. lic., PStat) and Amanda Ljungberg (statistician) at Statistikkonsulterna Väst AB for their input regarding statistical analyses.
Conflict of interest
AS is a founder of in2cure AB, a parent company of Xinnate AB, which was the sponsor of the clinical trial from which the biobank samples that were used in this study are derived. GP was employed part-time (20%) by Xinnate AB. JF provides consulting services to Xinnate AB. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors declare that this study received funding from Xinnate AB. The funder had the following involvement in the study: providing project management resources, regulatory expertise, clinical study design, and personnel necessary for the execution of the clinical trial, including the collection of clinical samples and related data.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmed.2025.1609347/full#supplementary-material
Abbreviations
BCA, bicinchoninic acid; CFU, colony forming units; ELISA, enzyme-linked immunosorbent assay; HBP, heparin-binding protein; IFN, interferon; IL, interleukin; IL-1R, interleukin-1 receptor; MALDI-TOF, Matrix-Assisted Laser Desorption Ionization - Time Of Flight; MBT, MALDI Biotyper; MPO, myeloperoxidase; MS, mass spectrometry; MyD88, myeloid differentiation primary response 88; NE, neutrophil elastase; PBS, phosphate-buffered saline; TCP, thrombin-derived C-terminal peptide; TNF, tumor necrosis factor.
References
1. Gurtner GC, Werner S, Barrandon Y, Longaker MT. Wound repair and regeneration. Nature. (2008) 453:314–21. doi: 10.1038/nature07039
2. Reinke JM, Sorg H. Wound repair and regeneration. Eur Surg Res. (2012) 49:35–43. doi: 10.1159/000339613
3. Landén NX Li D, Ståhle M. Transition from inflammation to proliferation: a critical step during wound healing. Cell Mol Life Sci. (2016) 73:3861–85. doi: 10.1007/s00018-016-2268-0
4. Schultz GS, Chin GA, Moldawer L, Diegelmann RF. Principles of Wound Healing. In:Fitridge R, Thompson M, , editors Mechanisms of Vascular Disease: A Reference Book for Vascular Specialists Adelaide (AU): University of Adelaide Press. (2011). doi: 10.1017/UPO9781922064004.024
5. Eming SA, Krieg T, Davidson JM. Inflammation in wound repair: molecular and cellular mechanisms. J Invest Dermatol. (2007) 127:514–25. doi: 10.1038/sj.jid.5700701
6. Spear M. Wound exudate–the good, the bad, and the ugly. Plast Surg Nurs. (2012) 32:77–9. doi: 10.1097/PSN.0b013e318256d638
7. Barrientos S, Stojadinovic O, Golinko MS, Brem H, Tomic-Canic M. Growth factors and cytokines in wound healing. Wound Repair Regen. (2008) 16:585–601. doi: 10.1111/j.1524-475X.2008.00410.x
8. Mahmoud NN, Hamad K, Al Shibitini A, Juma S, Sharifi S, Gould L, et al. Investigating inflammatory markers in wound healing: understanding implications and identifying artifacts. ACS Pharmacol Transl Sci. (2024) 7:18–27. doi: 10.1021/acsptsci.3c00336
9. Misciali C, Dika E, Baraldi C, Fanti PA, Mirelli M, Stella A, et al. Vascular leg ulcers: histopathologic study of 293 patients. Am J Dermatopathol. (2014) 36:977–83. doi: 10.1097/DAD.0000000000000141
10. Tuttle MS. Association between microbial bioburden and healing outcomes in venous leg ulcers: a review of the evidence. Adv Wound Care. (2015) 4:1–11. doi: 10.1089/wound.2014.0535
11. Shen S, Miskolci V, Dewey CN, Sauer JD, Huttenlocher A. Infection induced inflammation impairs wound healing through IL-1β signaling. iScience. (2024) 27:109532. doi: 10.1016/j.isci.2024.109532
12. Lee HJ, Kim M. Skin barrier function and the microbiome. Int J Mol Sci. (2022) 23:13071. doi: 10.3390/ijms232113071
13. Wang G, Sweren E, Liu H, Wier E, Alphonse MP, Chen R, et al. Bacteria induce skin regeneration via IL-1beta signaling. Cell Host Microbe. (2021) 29:777–91 e6. doi: 10.1016/j.chom.2021.03.003
14. Naik S, Bouladoux N, Wilhelm C, Molloy MJ, Salcedo R, Kastenmuller W, et al. Compartmentalized control of skin immunity by resident commensals. Science. (2012) 337:1115–9. doi: 10.1126/science.1225152
15. Nakatsuji T, Chen TH, Narala S, Chun KA, Two AM, Yun T, et al. Antimicrobials from human skin commensal bacteria protect against Staphylococcus aureus and are deficient in atopic dermatitis. Sci Transl Med. (2017) 9:eaah4680. doi: 10.1126/scitranslmed.aah4680
16. Uberoi A, Bartow-McKenney C, Zheng Q, Flowers L, Campbell A, Knight SAB, et al. Commensal microbiota regulates skin barrier function and repair via signaling through the aryl hydrocarbon receptor. Cell Host Microbe. (2021) 29:1235–48.e8. doi: 10.1016/j.chom.2021.05.011
17. Khadka VD, Markey L, Boucher M, Lieberman TD. Commensal skin bacteria exacerbate inflammation and delay skin barrier repair. J Invest Dermatol. (2024) 144:2541-2552.e10. doi: 10.1101/2023.12.04.569980
18. Burian M, Bitschar K, Dylus B, Peschel A, Schittek B. The protective effect of microbiota on S. aureus skin colonization depends on the integrity of the epithelial barrier. J Invest Dermatol. (2017) 137:976–9. doi: 10.1016/j.jid.2016.11.024
19. International Wound Infection Institute. Wound infection in clinical practice- Principles of best practice. International Consensus Update (2022). Available online at: https://woundsinternational.com/wp-content/uploads/2023/05/IWII-CD-2022-web.pdf (Accessed June 17, 2025).
20. Ågren MS, Litman T, Eriksen JO, Schjerling P, Bzorek M, Gjerdrum LMR. Gene expression linked to reepithelialization of human skin wounds. Int J Mol Sci. (2022) 23:15746. doi: 10.3390/ijms232415746
21. Ahlström MG, Gjerdrum LMR, Larsen HF, Fuchs C, Sørensen AL, Forman JL, et al. Suction blister lesions and epithelialization monitored by optical coherence tomography. Skin Res Technol. (2018) 24:65–72. doi: 10.1111/srt.12391
22. Daeschlein G, Alborova J, Patzelt A, Kramer A, Lademann J. Kinetics of physiological skin flora in a suction blister wound model on healthy subjects after treatment with water-filtered infrared-A radiation. Skin Pharmacol Physiol. (2012) 25:73–7. doi: 10.1159/000332753
23. Svedman P, Svedman C, Njalsson T. Epithelialization and blood flow in suction blister wounds on healthy volunteers. J Invest Surg. (1991) 4:175–89. doi: 10.3109/08941939109140778
24. Burian EA, Sabah L, Kirketerp-Møller K, Gundersen G, Ågren MS. Effect of stabilized hypochlorous acid on re-epithelialization and bacterial bioburden in acute wounds: a randomized controlled trial in healthy volunteers. Acta Derm Venereol. (2022) 102:adv00727. doi: 10.2340/actadv.v102.1624
25. Wilhelm KP, Wilhelm D, Bielfeldt S. Models of wound healing: an emphasis on clinical studies. Skin Res Technol. (2017) 23:3–12. doi: 10.1111/srt.12317
26. Lundgren S, Wallblom K, Fisher J, Erdmann S, Schmidtchen A, Saleh K. Study protocol for a phase 1, randomised, double-blind, placebo-controlled study to investigate the safety, tolerability and pharmacokinetics of ascending topical doses of TCP-25 applied to epidermal suction blister wounds in healthy male and female volunteers. BMJ Open. (2023) 13:e064866. doi: 10.1136/bmjopen-2022-064866
27. Larsen HF, Ahlström MG, Gjerdrum LMR, Mogensen M, Ghathian K, Calum H, et al. Noninvasive measurement of reepithelialization and microvascularity of suction-blister wounds with benchmarking to histology. Wound Repair Regen. (2017) 25:984–93. doi: 10.1111/wrr.12605
28. Siddiqui AR, Bernstein JM. Chronic wound infection: facts and controversies. Clin Dermatol. (2010) 28:519–26. doi: 10.1016/j.clindermatol.2010.03.009
29. Grice EA, Kong HH, Conlan S, Deming CB, Davis J, Young AC, et al. Topographical and temporal diversity of the human skin microbiome. Science. (2009) 324:1190–2. doi: 10.1126/science.1171700
30. Ogai K, Nagase S, Mukai K, Iuchi T, Mori Y, Matsue M, et al. A Comparison of Techniques for Collecting Skin Microbiome Samples: Swabbing Versus Tape-Stripping. Front Microbiol. (2018) 9:2362. doi: 10.3389/fmicb.2018.02362
31. Jorgensen JH, Carroll KC, Pfaller MA. American Society for Microbiology. Manual of clinical microbiology. 11 Edn. United States: Washington, DC: ASM Press. (2015)
32. Kallstrom G. Are quantitative bacterial wound cultures useful? J Clin Microbiol. (2014) 52:2753–6. doi: 10.1128/JCM.00522-14
33. Dichtl K, Klugherz I, Greimel H, Luxner J, Koberl J, Friedl S, et al. A head-to-head comparison of three MALDI-TOF mass spectrometry systems with 16S rRNA gene sequencing. J Clin Microbiol. (2023) 61:e0191322. doi: 10.1128/jcm.01913-22
34. Dubois D, Leyssene D, Chacornac JP, Kostrzewa M, Schmit PO, Talon R, et al. Identification of a variety of Staphylococcus species by matrix-assisted laser desorption ionization-time of flight mass spectrometry. J Clin Microbiol. (2010) 48:941–5. doi: 10.1128/JCM.00413-09
35. Wallblom K, Lundgren S, Saleh K, Schmidtchen A, Puthia M. Image-Based Non-invasive assessment of suction blister wounds for clinical safety and efficacy. Wound Repair Regen. (2023) 32:343–359. doi: 10.1111/wrr.13172
36. Borregaard N, Cowland JB. Granules of the human neutrophilic polymorphonuclear leukocyte. Blood. (1997) 89:3503–21. doi: 10.1182/blood.V89.10.3503
37. Fisher J, Linder A. Heparin-binding protein: a key player in the pathophysiology of organ dysfunction in sepsis. J Intern Med. (2017) 281:562–74. doi: 10.1111/joim.12604
38. Johansson J, Lindbom L, Herwald H, Sjöberg F. Neutrophil-derived heparin binding protein–a mediator of increased vascular permeability after burns? Burns. (2009) 35:1185–7. doi: 10.1016/j.burns.2009.02.021
39. Lundqvist K, Herwald H, Sonesson A, Schmidtchen A. Heparin binding protein is increased in chronic leg ulcer fluid and released from granulocytes by secreted products of Pseudomonas aeruginosa. Thromb Haemost. (2004) 92:281–7. doi: 10.1160/TH03-12-0732
40. Bentzer P, Fisher J, Kong HJ, Mörgelin M, Boyd JH, Walley KR, et al. Heparin-binding protein is important for vascular leak in sepsis. Intensive Care Med Exp. (2016) 4:33. doi: 10.1186/s40635-016-0104-3
41. Linehan JL, Harrison OJ, Han SJ, Byrd AL, Vujkovic-Cvijin I, Villarino AV, et al. Non-classical immunity controls microbiota impact on skin immunity and tissue repair. Cell. (2018) 172:784-96 e18. doi: 10.1016/j.cell.2017.12.033
42. Naik S, Bouladoux N, Linehan JL, Han SJ, Harrison OJ, Wilhelm C, et al. Commensal-dendritic-cell interaction specifies a unique protective skin immune signature. Nature. (2015) 520:104–8. doi: 10.1038/nature14052
43. Harrison OJ, Linehan JL, Shih HY, Bouladoux N, Han SJ, Smelkinson M, et al. Commensal-specific T cell plasticity promotes rapid tissue adaptation to injury. Science. (2019) 363:eaat6280. doi: 10.1126/science.aat6280
44. Di Domizio J, Belkhodja C, Chenuet P, Fries A, Murray T, Mondejar PM, et al. The commensal skin microbiota triggers type I IFN-dependent innate repair responses in injured skin. Nat Immunol. (2020) 21:1034–45. doi: 10.1038/s41590-020-0721-6
45. Acosta EM, Little KA, Bratton BP, Lopez JG, Mao X, Payne AS, et al. Bacterial DNA on the skin surface overrepresents the viable skin microbiome. Elife. (2023) 12:RP87192. doi: 10.7554/eLife.87192
46. Lindsay S, Oates A, Bourdillon K. The detrimental impact of extracellular bacterial proteases on wound healing. Int Wound J. (2017) 14:1237–47. doi: 10.1111/iwj.12790
47. Cogen AL, Yamasaki K, Sanchez KM, Dorschner RA, Lai Y, MacLeod DT, et al. Selective antimicrobial action is provided by phenol-soluble modulins derived from Staphylococcus epidermidis, a normal resident of the skin. J Invest Dermatol. (2010) 130:192–200. doi: 10.1038/jid.2009.243
48. Pastar I, O'Neill K, Padula L, Head CR, Burgess JL, Chen V, et al. Staphylococcus epidermidis boosts innate immune response by activation of gamma delta T Cells and induction of perforin-2 in human skin. Front Immunol. (2020) 11:550946. doi: 10.3389/fimmu.2020.550946
49. Short WD, Rae M, Lu T, Padon B, Prajapati TJ, Faruk F, et al. Endogenous interleukin-10 contributes to wound healing and regulates tissue repair. J Surg Res. (2023) 285:26–34. doi: 10.1016/j.jss.2022.12.004
50. Saleh K, Sonesson A, Persson B, Riesbeck K, Schmidtchen A. A descriptive study of bacterial load of full-thickness surgical wounds in dermatologic surgery. Dermatol Surg. (2011) 37:1014–22. doi: 10.1111/j.1524-4725.2011.02040.x
51. Saleh K, Sonesson A, Persson K, Riesbeck K, Schmidtchen A. Can dressings soaked with polyhexanide reduce bacterial loads in full-thickness skin grafting? A randomized controlled trial. J Am Acad Dermatol. (2016) 75:1221-8.e4. doi: 10.1016/j.jaad.2016.07.020
52. Saleh K, Strömdahl AC, Riesbeck K, Schmidtchen A. Inflammation biomarkers and correlation to wound status after full-thickness skin grafting. Front Med. (2019) 6:159. doi: 10.3389/fmed.2019.00159
53. Saleh K, Schmidtchen A. Surgical site infections in dermatologic surgery: etiology, pathogenesis, and current preventative measures. Dermatol Surg. (2015) 41:537–49. doi: 10.1097/DSS.0000000000000364
54. Hutchinson JJ, Lawrence JC. Wound infection under occlusive dressings. J Hosp Infect. (1991) 17:83–94. doi: 10.1016/0195-6701(91)90172-5
55. Kligman AM, Leyden JJ, McGinley KJ. Bacteriology. J Invest Dermatol. (1976) 67:160–8. doi: 10.1111/1523-1747.ep12513007
56. Nayeri F. Occlusive bandaging of wounds with decreased circulation promotes growth of anaerobic bacteria and necrosis: case report. BMC Res Notes. (2016) 9:394. doi: 10.1186/s13104-016-2205-1
57. Grice EA, Kong HH, Renaud G, Young AC, Bouffard GG, Blakesley RW, et al. A diversity profile of the human skin microbiota. Genome Res. (2008) 18:1043–50. doi: 10.1101/gr.075549.107
58. Skowron K, Bauza-Kaszewska J, Kraszewska Z, Wiktorczyk-Kapischke N, Grudlewska-Buda K, Kwiecińska-Piróg J, et al. Human skin microbiome: impact of intrinsic and extrinsic factors on skin microbiota. Microorganisms. (2021) 9:543. doi: 10.3390/microorganisms9030543
59. El Fawal GF, Abu-Serie MM, Hassan MA, Elnouby MS. Hydroxyethyl cellulose hydrogel for wound dressing: Fabrication, characterization and in vitro evaluation. Int J Biol Macromol. (2018) 111:649–59. doi: 10.1016/j.ijbiomac.2018.01.040
Keywords: wound healing, wound bacteria, inflammation, exudation, cytokines, heparin-binding protein
Citation: Lundgren S, Petruk G, Wallblom K, Cardoso JFP, Strömdahl A-C, Forsberg F, Luo C, Nilson B, Hartman E, Fisher J, Puthia M, Saleh K and Schmidtchen A (2025) Temporal dynamics and interrelations of cytokines, neutrophil proteins, exudation, and bacterial colonization in epidermal wound healing. Front. Med. 12:1609347. doi: 10.3389/fmed.2025.1609347
Received: 10 April 2025; Accepted: 30 June 2025;
Published: 21 July 2025.
Edited by:
Emi Dika, University of Bologna, ItalyReviewed by:
Hong-Hui Wang, Hunan University, ChinaElisabetta Magnaterra, University of Bologna, Italy
Copyright © 2025 Lundgren, Petruk, Wallblom, Cardoso, Strömdahl, Forsberg, Luo, Nilson, Hartman, Fisher, Puthia, Saleh and Schmidtchen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Artur Schmidtchen, YXJ0dXIuc2NobWlkdGNoZW5AbWVkLmx1LnNl