 Susana Merino
Susana Merino Juan M. Tomás
Juan M. Tomás- Departamento de Microbiología, Facultad de Biología, Universidad de Barcelona, Barcelona, Spain
Initial hydridization tests using Aeromonas salmonicida typical and atypical strains showed the possibility of different lipopolysaccharide (LPS) outer cores among these strains. By chemical structural analysis, LPS-core SDS-PAGE gel migration, and functional and comparative genomics we demonstrated that typical A. salmonicida (subsp. salmonicida) strains and atypical subsp. masoucida and probably smithia strains showed the same LPS outer core. A. salmonicida subsp. achromogenes strains show a similar LPS outer core but lack one of the most external residues (a galactose linked α1-6 to heptose), not affecting the O-antigen LPS linkage. A. salmonicida subsp. pectinolytica strains show a rather changed LPS outer core, which is identical to the LPS outer core from the majority of the A. hydrophila strains studied by genomic analyses. The LPS inner core in all tested A. salmonicida strains, typical and atypical, is well-conserved. Furthermore, the LPS inner core seems to be conserved in all the Aeromonas (psychrophilic or mesophilic) strains studied by genomic analyses.
Introduction
The smooth lipopolysaccharide (LPS) in Gram-negative bacteria consists of large amphiphilic molecules with a hydrophilic polysaccharide and a hydrophobic highly conserved lipid component covalently bound. This lipid, named lipid A, is the bioactive endotoxin subunit. The polysaccharide section is mainly formed by two parts: one more internal and conserved, the core region, and one more external and highly variable, the O-specific chain, named also O-antigen for its immunogenic properties. Smooth LPS molecules show both polysaccharide parts, while rough LPS molecules only the completed or truncated LPS core. The lipid A, LPS-core, and O-antigen LPS have been differentiated and formally classified by their chemical structure, degree of conservation, biosynthetic pathways and genetic determination (see general review Aquilini and Tomás, 2015).
The LPS-core is also subdivided in two regions: inner and outer core. Within a genus or family, the structure of the inner core tends to be well-conserved, and typically consists of unusual sugars, particularly 3-deoxy-D-manno-oct-2-ulosonic acid (Kdo) and heptoses (Hep; Holst, 2002). The outer core shows more structural diversity, is characterized by more common hexose sugars such as glucose (Glc), galactose (Gal), N-acetyl galactosamine (GalNAc) and N-acetyl glucosamine (GlcNAc), and is more variable than the inner core (Holst, 2007).
Aeromonas salmonicida subsp. salmonicida is the typical etiological agent responsible of a systemic fish disease named furunculosis, which converts this bacterium into an important pathogen (Scott, 1968). This pathogen has been subjected to considerable investigation because of its enormous importance in the farmed fish industry. Its major virulence factor is an S-layer (named A-layer), which principally consists of a unique two-dimensional crystalline tetragonal protein (A-protein with a molecular weight of 49 KDa) array (Ishiguro et al., 1981), tethered to the cell by LPS (Belland and Trust, 1985). Some studies chemically characterized the LPS O-antigen polysaccharide and the core oligosaccharide region structures from A. salmonicida strain SJ-15 (Shaw et al., 1983, 1992). Furthermore, recently the functional genomics of the LPS O-antigen and A-layer from typical and atypical strains were described (Merino et al., 2015), and concluded that all the A. salmonicida typical and atypical strains, other than A. salmonicida subsp. pectinolytica strains, shared the same LPS O-antigen and presence of A-layer. A. salmonicida subsp. pectinolytica strains present another type of O-antigen LPS and lack one of the major virulence factors, the A-layer (Merino et al., 2015).
Other studies reported the chemical structure of A. salmonicida subsp. salmonicida LPS core oligosaccharide region from strains A449 and 80204-1 (Wang et al., 2006). Also, we were able to establish the A. salmonicida subsp. salmonicida strain A450 genomics and proteomics for the LPS-core (waasalmo), which is distributed in three different chromosomal gene locations (Jimenez et al., 2009).
Regions 2 and 3 showed identical genes between A. salmonicida subsp. salmonicida strain A450 and A449, and also with A. hydrophila AH-3 (Jimenez et al., 2008, 2009). A. salmonicida subsp. salmonicida strains A450 and A449 region 1 showed seven identical genes, three of them identical to A. hydrophila AH-3, three of them similar but not identical to A. hydrophila AH-3, and one of them totally different that seems to be specific for A. salmonicida subsp. salmonicida (Jimenez et al., 2008, 2009).
Currently, there are five accepted subspecies of Aeromonas salmonicida: A. salmonicida subsp. salmonicida (known as typical), masoucida, achromogenes, pectinolytica, and smithia (Austin and Austin, 2007). Atypical A. salmonicida include subsp. smithia, subsp. masoucida, subsp. achromogenes and subsp. pectinolytica which, with the exception of A. salmonicida subsp. pectinolytica, are found as pathogens in a wide variety of fish species. In this work we studied the three chromosomal regions encoding the LPS core biosynthesis by comparative analysis of published complete genomes of different strains of A. salmonicida subspecies, as well as the chemical LPS core structure for subspecies masoucida, achromogenes, and pectinolytica strains.
Materials and Methods
Bacterial Strains, Plasmids, and Growth Conditions
Bacterial strains, and plasmids used in this study are listed in Table 1. Aeromonas strains were routinely grown on tryptic soy broth (TSB) or tryptic soy agar (TSA) at 20°C (A. salmonicida) and 30°C (A. hydrophila). Escherichia coli strains were grown in Luria-Bertani Miller broth and on the same medium with agar at 37°C. Kanamycin (50 μg ml-1), ampicillin (100 μg ml-1), rifampicin (100 μg ml-1), nalidixic acid (20 μg ml-1), or gentamicin (20 μg ml-1) were added to the different media when required.
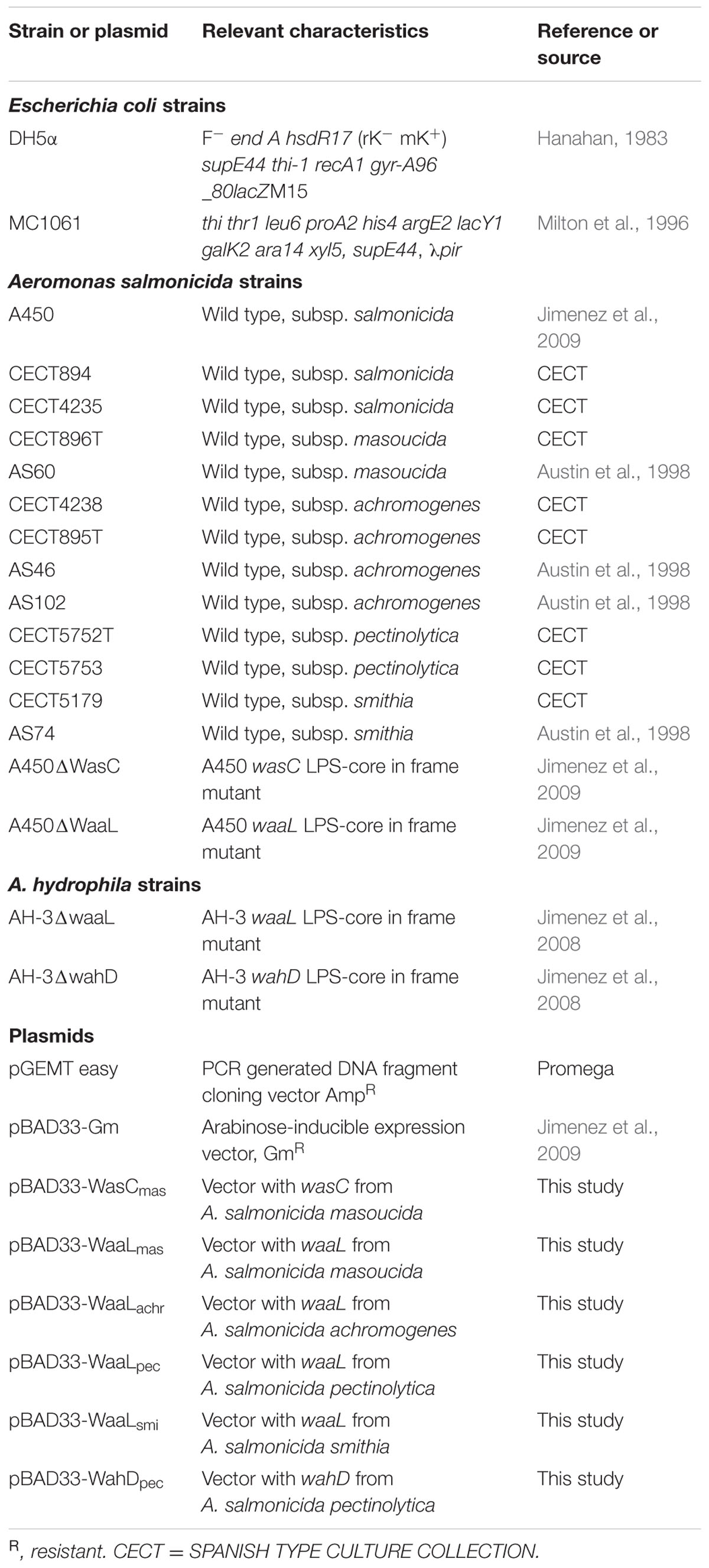
TABLE 1. Bacterial strains and plasmids used.
Genetic General Methodology
General DNA manipulations were done essentially as previously described, as well as the DNA sequencing and bioinformatics analysis of sequenced data (Aquilini et al., 2014).
Dot Blot Hybridizations
Total DNA was denatured after 5 min boiling, chilled on ice for 5 min. After, DNA samples were spotted onto prewetted in 2x SSC Hybond N1 (Amersham) nylon membrane and fixed by UV irradiation. Prehybridization was performed in a solution of 5x SSC, 0.1% N-lauroyl sarcosine, 0.02% SDS, 5% blocking reagent (Roche), and 50% formamide for 2 h at 42°C. Hybridization with the correspondent labeled probe (20 ng/ml) with digoxigenin was performed for 18 h at 42°C. The alkaline phosphatase detection system was finally carried out using the enhanced chemiluminescence detection system (Amersham) according to the manufacturer’s instructions.
Plasmid Constructions for Gene Overexpression and Mutant Complementation Studies
For gene complementation studies of previously isolated A. salmonicida A450 and A. hydrophila AH-3 core mutants, the corresponding genes from chromosomal DNA of different A. salmonicida subspecies strains were PCR-amplified using specific primer pairs (Table 2) and ligated to pGEMTeasy plasmid. To generate pBAD33-Gm constructions pGEMT plasmids with the different genes were double digested with XbaI and SmaI and the DNA fragment obtained in each case ligated to pBAD33-Gm double digested with the same enzymes.
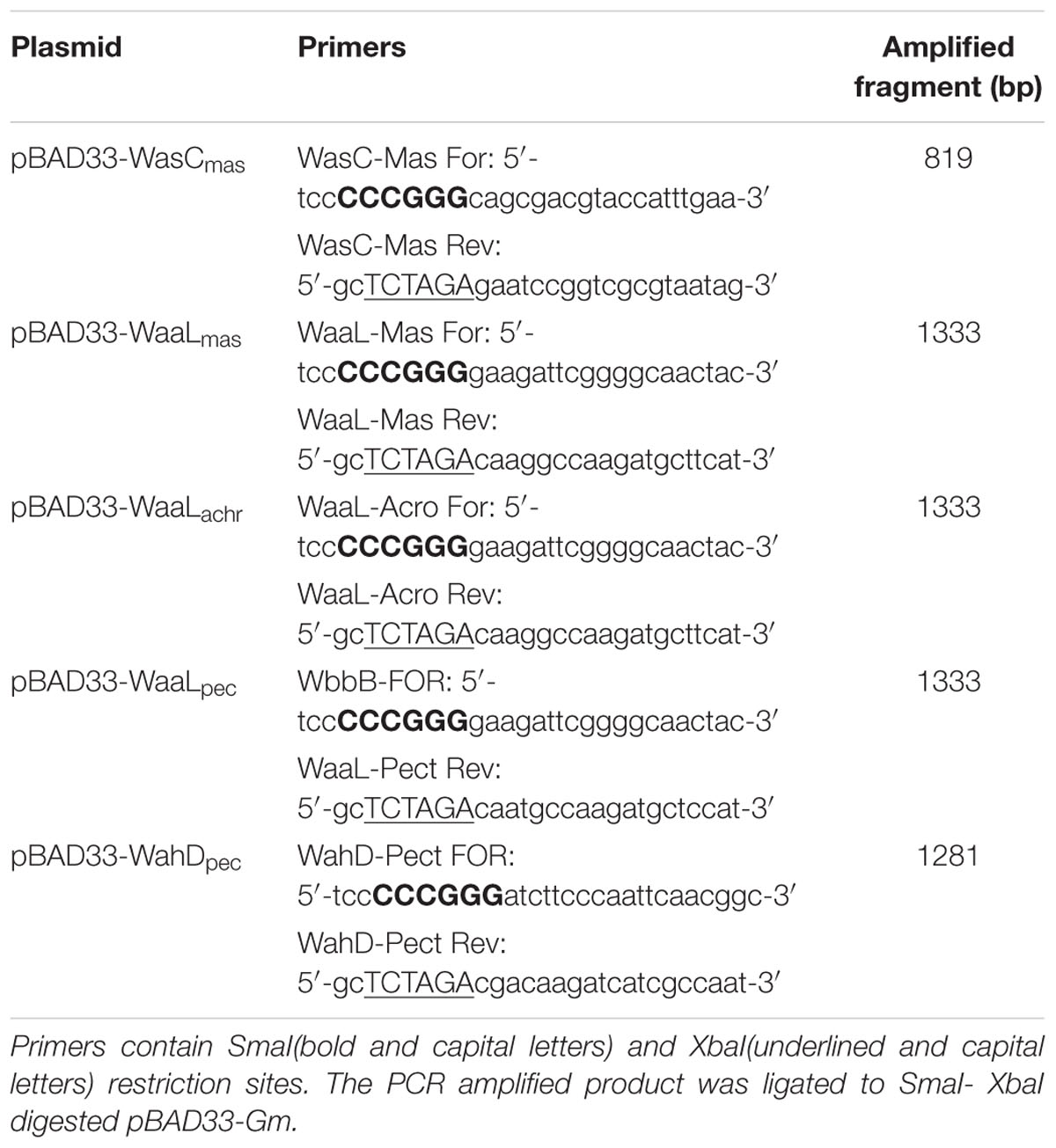
TABLE 2. Primers used for mutant complementation using vector pBAD33-Gm.
Plasmid pBAD33-WaaLsmi was generated using the primers designed for pBAD33-WaaLmas. pBAD33-Gm plasmids into E. coli MC1061 were then transferred into the different mutants by triparental mating using the mobilizing strain HB101/pRK2073 (Jimenez et al., 2009). Mutants were selected on plates containing gentamicin and nalidixic acid for the A450strain, and gentamicin and rifampicin in case of the AH-3 strain. Each gene was expressed from the arabinose-inducible and glucose-repressible pBAD33-Gm promoter.
LPS Isolation and SDS-PAGE
For screening purposes LPS was obtained after proteinase K digestion of whole cells and the LPS samples were separated by SDS-PAGE or SDS-Tricine-PAGE and visualized by silver staining as previously described (Aquilini et al., 2014). Cultures for analysis of LPS were grown in TSB at 20°C. Dried bacterial cells of each strain in 25 mM Tris⋅ HCl buffer containing 2 mM CaCl2 pH 7.63 (10 ml g-1) were treated at 37°C with RNAse, DNAse (24 h, 1 mg g-1 each), and then with proteinase K (36 h, 1 mg g-1). The suspension was dialyzed and lyophilized, and the LPS was extracted by the phenol-water procedure (Westphal and Jann, 1965). A portion of the LPS (∼50 mg) from each strain was heated with aqueous 2% acetic acid (6 ml) at 100°C for 45 min. The precipitate was removed by centrifugation (13,000g × 20 min) and the supernatant fractionated on a column (56 cm × 2.6 cm) of Sephadex G-50 (S) in 0.05 M pyridinium acetate buffer pH 4.5 with monitoring using a differential refractometer. An oligosaccharide fraction was obtained in a yield 9–20 % depending on the strain.
Methylation Analysis and Mass Spectrometry
The methylation analyses were performed as previously described (Jimenez et al., 2009). Positive-ion reflectron time-of-flight mass spectra (MALDI-TOF) were acquired on a Voyager DE-PR instrument (Applied Biosystems) equipped with a delayed extraction ion source and used as previously described (Jimenez et al., 2009).
Comparative Genomics and Reannotation
For each analyzed genome we gathered all CDS and pseudo-CDS information by parsing NCBI GenBank records. When we obtained the UniProt Knowledge Base records for these loci using the cross-reference with Entrez GeneIDs and parsed them for gene names, functional annotations, and associated COG, PFAM, and TIGRFAM protein domains were studied. To annotate orthologs, we wrote custom scripts to analyze reference sequence alignments made to subject genomes with blastn and tblastn via NCBI’s Web application programming interface. Briefly, we manually confirmed contextually accurate alignments, and then the script integrated coordinates and sequence information from both BLAST methods to locate the bounds of the reference gene in the subject genome; if an aligned start or stop codon was not located, we manually inspected the region. The script then analyzed alignments for insertions, deletions, premature stop codons, frameshifts, and changes to the start codon. An alignment in the same genomic context with >95% amino acid identity, excluding gaps and truncations, was our initial cutoff for orthology. The genomes of subsp. salmonicida A449, subsp. masoucida strain NBRC13784, subsp. achromogenes strain AS03 and subsp. pectinolytica strain 34melT are located at the GenBank accession numbers: CP000644, BAWQ00000000, AMQG00000000.2 and ARYZ00000000.2, respectively. The complete nucleotide sequences of the three A. salmonicida A450 chromosomal regions containing the LPS core biosynthetic genes described here have been assigned GenBank accession numbers FJ238464, FJ238465, and FJ238466, respectively. The complete nucleotide sequences of the three A. hydrophila AH-3 chromosomal regions containing LPS core biosynthesis genes described here have been assigned the following GenBank accession numbers: EU296246, EU296247, and EU296248.
Results
We previously established the genomics and proteomics of the A. salmonicida subsp. salmonicida A450 strain waa (Jimenez et al., 2009; Figure 1). We studied by Colony Southern blot analysis, using several DNA probes, the waa region of A. salmonicida in subspecies masoucida, achromogenes, pectinolytica, and smithia. The initial selected DNA probes from strain A450 corresponded to complete wasC for chromosomal region 1, complete waaE for region 2, and complete waaC for region 3. WasC is the glycosyltransferase that links Gal to HepV in an α-1,6 linkage, WaaE the glycosyltransferase that links Glc to HepI in a β-1,4 linkage, and WaaC the heptosyltransferase that links HepI to Kdo in an α-1,5 linkage (Figure 1). A positive reaction was obtained with all the subspecies genomic DNA against probes from regions 2 and 3 (Table 3). However, either subspecies pectinolytica or achromogenes showed no reaction against wasC probe from region 1. Subspecies masoucida and smithia rendered a positive reaction against this probe.
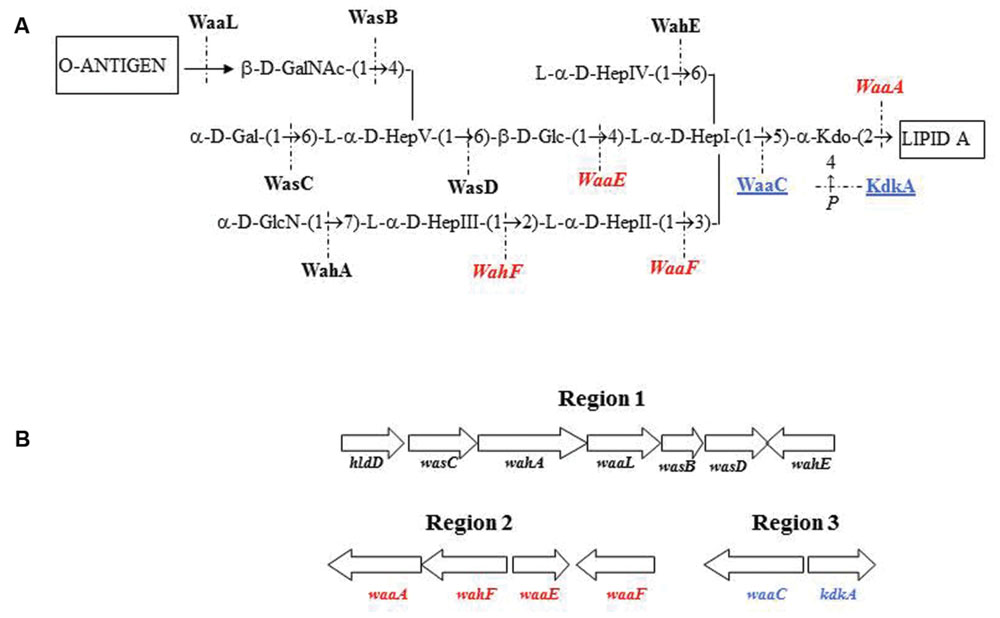
FIGURE 1. Aeromonas salmonicida subsp. salmonicida strain A450 chemical structure (A). Proteins encoded by genes from different chromosomal regions are shown in roman type and black (region 1), in italics and red (region 2), or underlined and blue (region 3). (B) Genomic regions with the genes involved in the A. salmonicida subsp. salmonicida strain A450 waa (LPS-core biosynthesis).
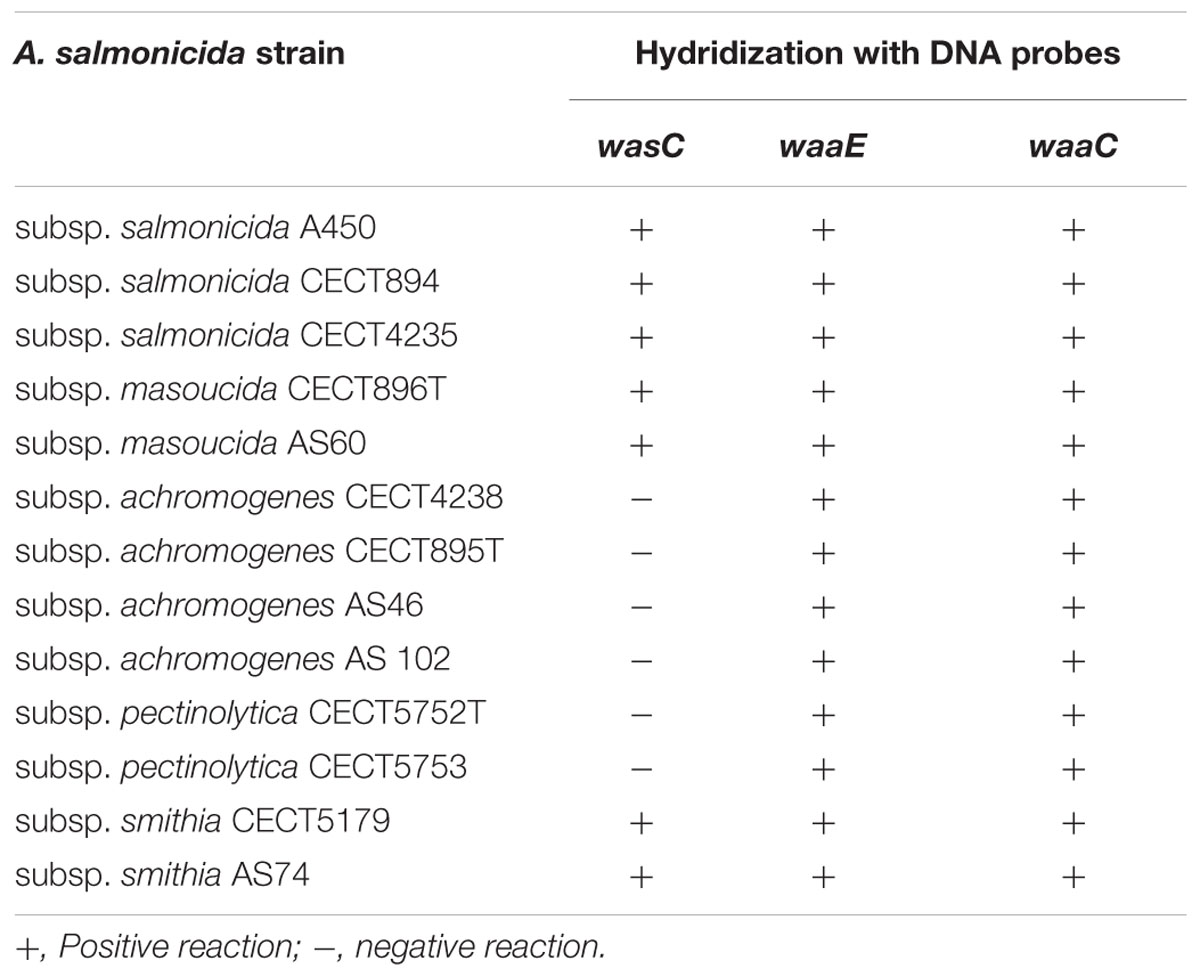
TABLE 3. Hybridization studies with several DNA probes of A. salmonicida strains from different subspecies.
When we used two additional DNA probes from region 1, wahA and wasD (Figure 1), a positive reaction was obtained with either subspecies pectinolytica or achromogenes genomic DNA. These results prompted us to study the LPS-core of the different A. salmonicida subspecies masoucida, pectinolytica, and achromogenes.
A. salmonicida subsp. masoucida
Composition analysis of the strain CECT896T core oligosaccharide from purified LPS by GLC showed the presence of Glc, Gal, GlcN (glucosamine), GalNAc, L,D-Hep, and Kdo in the ratios 1:0.9:0.9:0.8:4.7:0.9, respectively. The mass spectrum from this core oligosaccharide sample showed a major molecular ion peak at m/z 1.888,60 (Figure 2A), corresponding to the full core (calculated molecular mass, 1.887,60 atomic mass units). This molecular mass is essentially similar to those reported for both wild-type A. salmonicida subsp. salmonicida strains A449 and 80204-1 (Wang et al., 2006). Similar to other reported cases, some structural heterogeneity was observed, which was associated with the existence of Kdo in both normal and anhydro forms. The signal observed could be attributed to Kdo1, Hep5, Hex2, HexN1, HexNAc1. Methylation analysis showed that the core oligosaccharide was characterized by containing similar molar ratios of terminal Gal, GlcN, GalNAc, and L,D-Hep. In addition, 6-substituted Glc, 2-substituted Hep, 7-substituted Hep, 4,6-bisubstituted Hep, and 3,4,6-trisubstituted Hep were found. The complete presumptive structure of the LPS from A. salmonicida subsp. masoucida strain CECT896T is shown in Figure 2B.
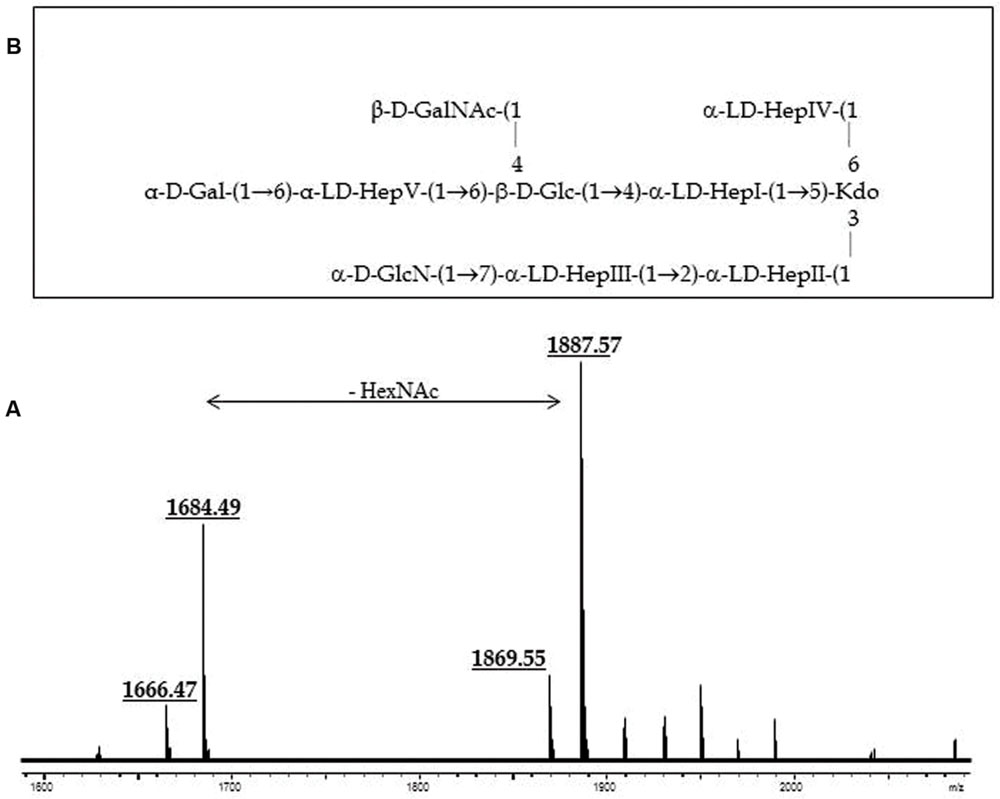
FIGURE 2. (A) Positive-ion MALDI-TOF of purified LPS core from A. salmonicida subspecies masoucida strain CECT8967T. (B) Presumptive LPS-core structure of the same strain according to the previously published A. salmonicida A450 (Jimenez et al., 2009).
The annotation of the waa region in A. salmonicida subsp. masoucida strain NBRC13784 was revised. Comparative genome analysis between the reannotated and the ortholog region in A. salmonicida subsp. salmonicida strain A450 (Jimenez et al., 2009), showed identical genes (Figure 3). The predicted functions encoded by the reannotated waa gene cluster of this A. salmonicida subsp. masoucida were in agreement with the chemical data obtained. Furthermore, the relative mobility of the LPS-core in a silver-stained SDS-PAGE gel from A. salmonicida subsp. masoucida strain NBRC13784 was identical to the mobility of the LPS-core from strain A. salmonicida subspecies salmonicida strain A450 (Figure 4).
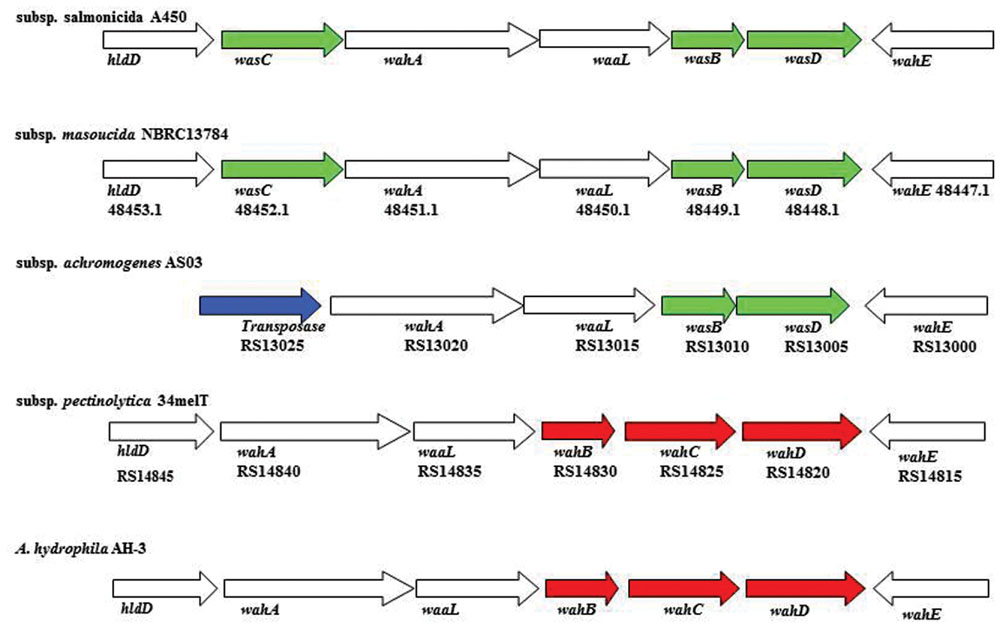
FIGURE 3. The genes in region 1 of A. salmonicida waa from subspecies salmonicida A450; subspecies masoucida strain NBRC13784; subspecies achromogenes strain AS03; subspecies pectinolytica strain 34melT; and A. hydrophila strain AH-3. The genes in green are unique for A. salmonicida strains, in red are initially unique for A. hydrophila strains, no color are shared by both species. The transposase is labeled in blue. The identity percentage of all the genes analyzed is over 97%.
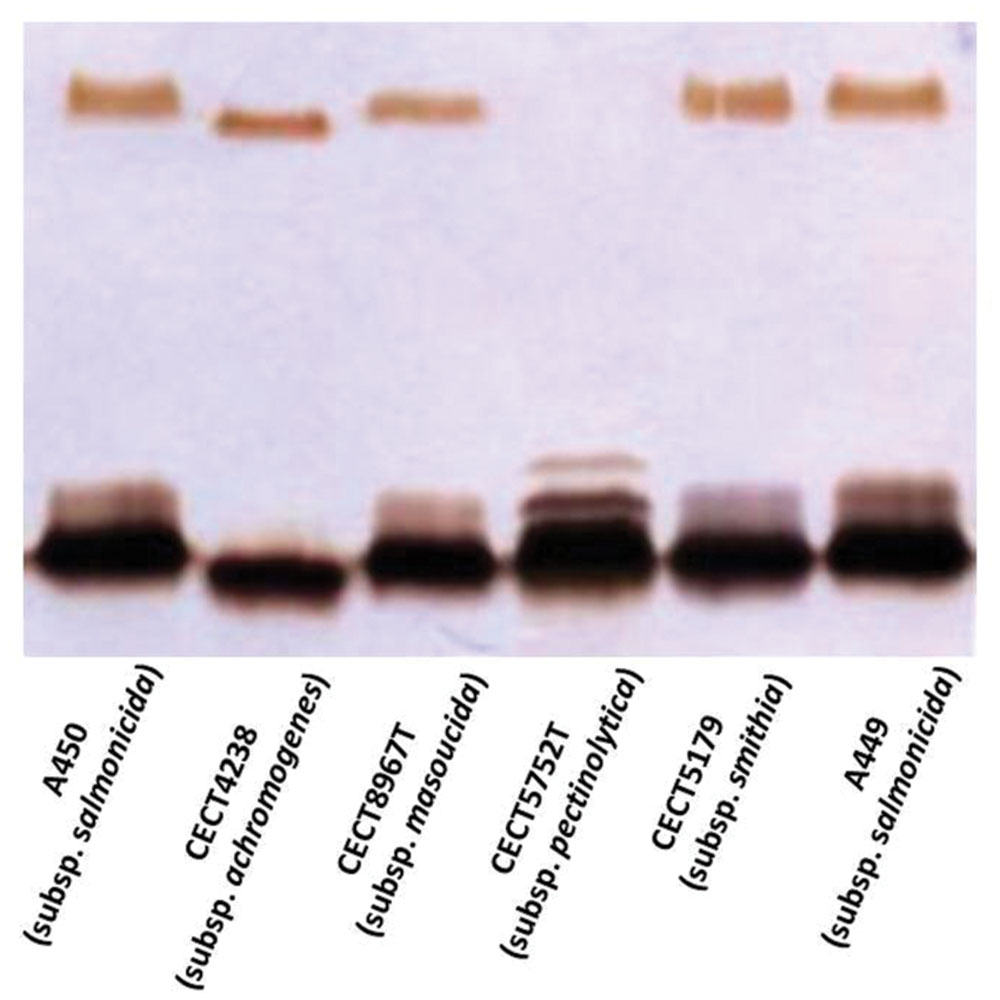
FIGURE 4. Lipopolysaccharide (LPS) analyzed by SDS-Tricine gel and silver stained from the different subsp. of A. salmonicida strains.
A. salmonicida subsp. achromogenes
GLC analysis of the strain CECT4238 core oligosaccharide from purified LPS showed the presence of Glc, GlcN, GalNAc, L,D-Hep, and Kdo in the ratios 1:1:0.9:4.5:0.9, respectively. The mass spectrum from this core oligosaccharide sample showed a major molecular ion peak at m/z 1.725,43 (Figure 5A), corresponding to the full core (calculated molecular mass, 1.726,10 atomic mass units). The signal observed was attributed to Kdo1, Hep5, Hex1, HexN1, HexNAc1. Similar to previous results some structural heterogeneity was observed due to Kdo in both normal and anhydro forms. Methylation analysis showed that the core oligosaccharide was characterized by containing similar molar ratios of terminal GlcN, GalNAc, and L,D-Hep. In addition, 6-substituted Glc, 2-substituted Hep, 7-substituted Hep, 4-substituted Hep, and 3,4,6-trisubstituted Hep were found. This core fraction was found to be essentially similar to those reported for wild-type A. salmonicida subsp. salmonicida strains, with the lack of the Gal linked in a α1-6 linkage to L,D-HepV (Jimenez et al., 2009). The complete presumptive structure of the LPS from A. salmonicida achromogenes is shown in Figure 5B. Only one complete genome of A. salmonicida subsp. achromogenes is currently available from strain AS03 (Han et al., 2013). When we revised this region by comparative genomics data in other A. salmonicida, we found the genes indicated in Figure 3, with a completely lack of wasC and hldD and the presence of a putative transposase. WasC is the glycosyltransferase that links Gal in an α-1,6 linkage to L,D-HepV in the LPS core of A. salmonicida subsp. salmonicida A450 (Jimenez et al., 2009; Figure 1) and HldD is the epimerase for the L,D-Hep and D,D-Hep (Read et al., 2004). The predicted functions encoded by the genes in this region were in agreement with the chemical data. Furthermore, the relative mobility of the LPS-core from A. salmonicida subsp. achromogenes strain CECT4238 is in a silver-stained SDS-PAGE gel was higher than the mobility of the LPS-core from strain A. salmonicida subsp. salmonicida strain A450 (Figure 4), which was in agreement with the loss of a monosaccharide residue (Jimenez et al., 2009).
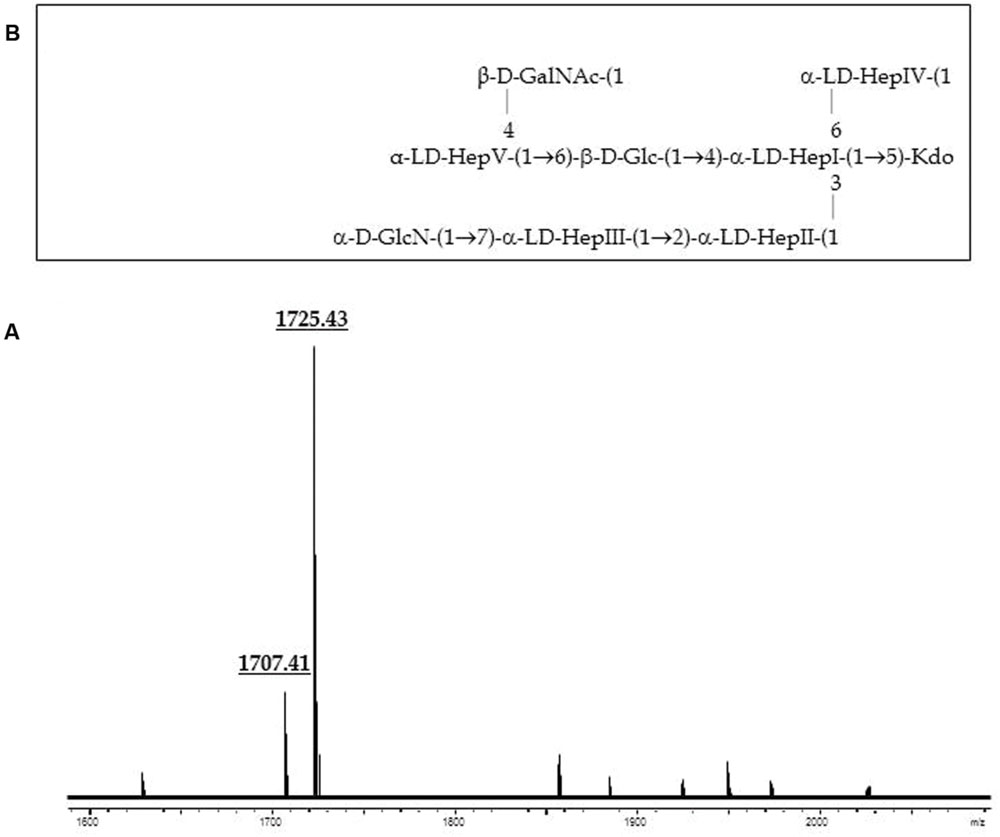
FIGURE 5. (A) Positive-ion MALDI-TOF of purified LPS core from A. salmonicida subspecies achromogenes strain CECT4238. (B) Presumptive LPS-core structure of the same strain according to the previously published A. salmonicida A450 (Jimenez et al., 2009).
A. salmonicida subsp. pectinolytica
Composition analysis of the strain CECT5752T core oligosaccharide from purified LPS by GLC revealed the presence of Glc, Gal, GlcN, D-glycero-D-manno-heptose (D,D-Hep), L-glycero-D-manno-heptose (L,D-Hep), and Kdo in the ratios 1:0.7:0.9:2.1:4.3, respectively. The major molecular ion peak at m/z 1.857,63 in its mass spectrum (Figure 6A) corresponded with calculated molecular mass 1.857,61 atomic mass units. The signal observed was attributed to Kdo1, Hep6, Hex2, HexN1. Methylation analysis resulted in identification of terminal Gal, 6-substituted Glc, terminal GlcN, terminal D,D-Hep, 6-substituted D,D-Hep, 4,6-disubstituted D,D-Hep, terminal L,D-Hep, 2-substituted L,D-Hep, 7-substituted L,D-Hep, and 3,4,6-trisubstituted L,D-Hep. The oligosaccharide sample from the A. salmonicida subsp. pectinolytica strain CECT5752T was found to be essentially identical to that of A. hydrophila AH-3 serogroup O34 (Jimenez et al., 2008), i.e., the same full core LPS. The complete presumptive structure of the LPS from A. salmonicida pectinolytica is shown in Figure 6B.
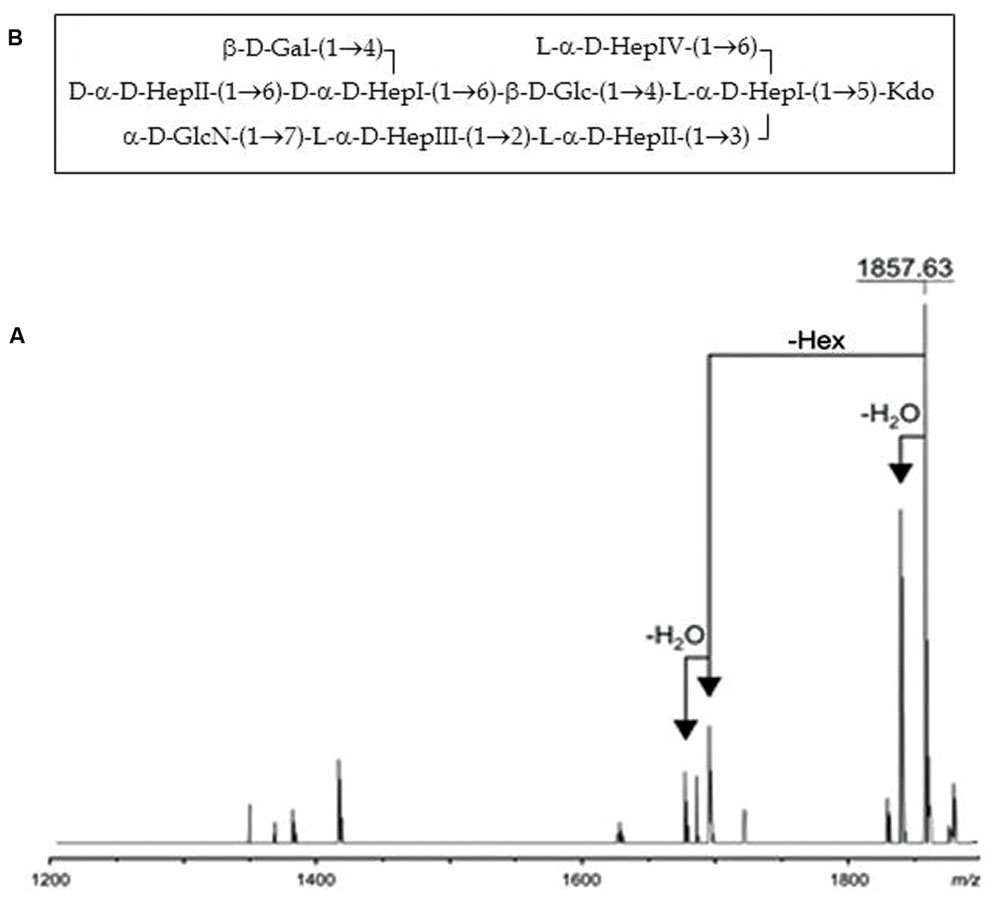
FIGURE 6. (A) Positive-ion MALDI-TOF of purified LPS core from A. salmonicida subspecies pectinolytica strain CECT5752T. (B) Presumptive LPS-core structure of the same strain.
A comparative “in silico” analysis of the reannotated region 1 from the A. salmonicida subsp. pectinolytica strain 34melT showed identical genes to A. hydrophila AH-3 serotype O34 but not to any of the A. salmonicida strains. As can be observed in Figure 3, A. salmonicida subsp. pectinolytica strain 34melT shows wahB, wahC, and wahD genes from A. hydrophila AH-3 (in red) and lack the wasB, wasC, and wasD genes characteristic of A. salmonicida strains (in green). Also, Figure 4 shows that this strain lacks the characteristic A. salmonicida O-antigen LPS and present some bands probably from another kind of O-antigen LPS (Merino et al., 2015).
Complementation Studies
In order to confirm some of the gene identity, we initially study complementation in A450ΔWasC (formerly A450ΔORF2) and A450ΔWaaL mutants (Jimenez et al., 2009). As can be observed by SDS-PAGE or SDS-Tricine gels, wasC from A. salmonicida masoucida strain CECT896T (pBAD33-Gm-WasC) was fully able to complement A450ΔWasC mutant (identical migratory profile for LPS-core in gels, see Figure 7A). Figure 7A also showed that waaL from A. salmonicida subsp. masoucida strain CECT896T, from A. salmonicida subsp. achromogenes strain CECT4238, and from A. salmonicida subsp. smithia strain CECT5179 (pBAD33-Gm-WaaLmasoucida, pBAD33-Gm-WaaLachromogenes, and pBAD33-Gm-WaaLsmithia, respectively) were able to complement A450ΔWaaL mutant (recovery of the O-antigen LPS in gel). However, waaL from A. salmonicida subsp. pectinolytica strain CECT5752T (pBAD33-Gm-WaaLpectinolytica) was unable to do it (Figure 7A). Nevertheless, waaL from A. salmonicida subsp. pectinolytica strain CECT5752T (pBAD33-Gm-WaaLpectinolytica) was fully able to complement AH-3ΔWaaL mutant (Jimenez et al., 2008) judged by their LPS profile in gel (recovery of the O-antigen LPS bands), as shown in Figure 7B. Furthermore, wahD from A. salmonicida subsp. pectinolytica strain CECT5752T (pBAD33-Gm-WahDpectinolytica) was fully able to complement AH-3ΔWahD mutant (Jimenez et al., 2008) as can be judged by their LPS profile in gel Figure 7B, recovery of LPS-core mobility as the wild type strain and O-antigen LPS bands.
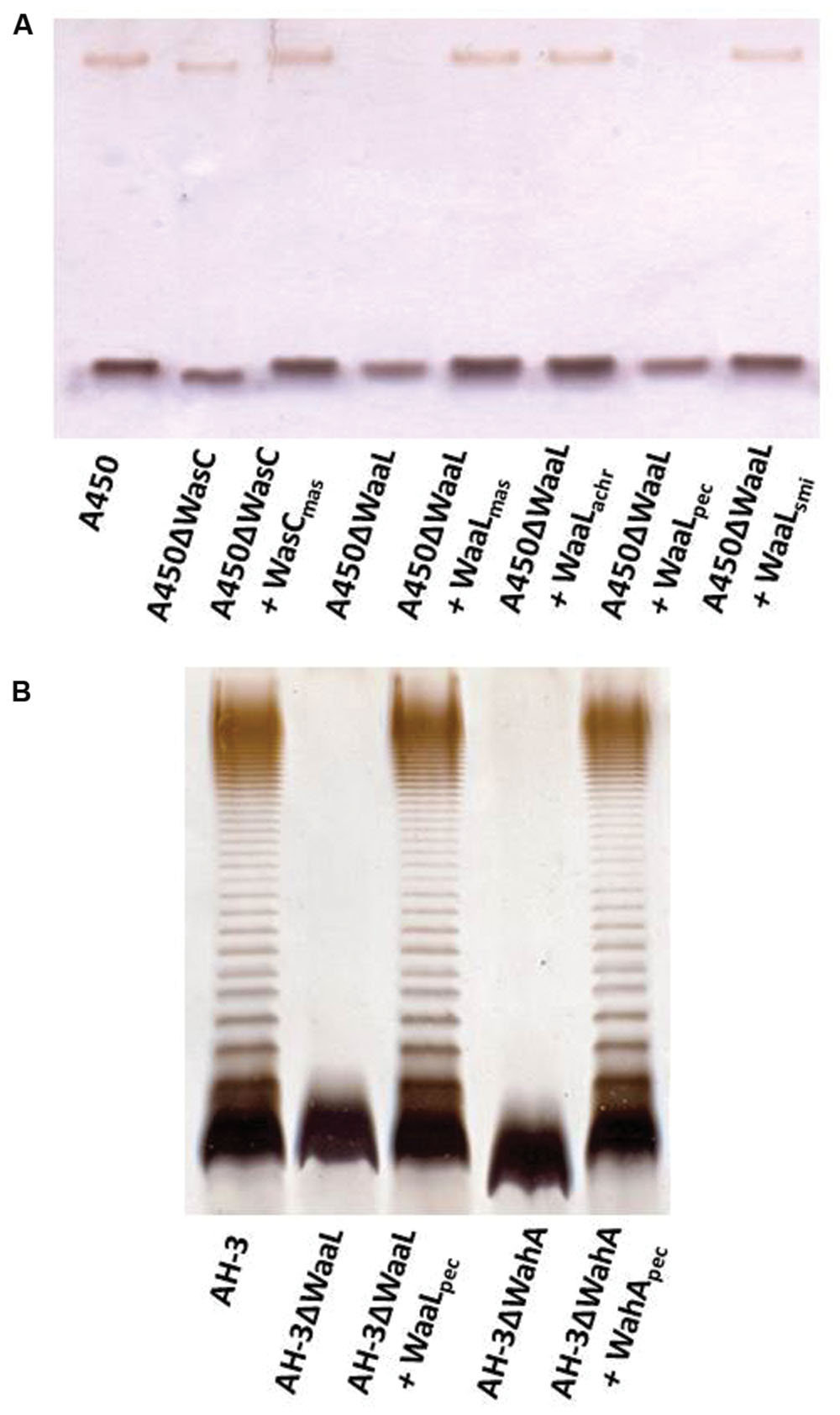
FIGURE 7. (A) Lipopolysaccharide analyzed by SDS-Tricine gel from A. salmonicida A450ΔWasC and A450ΔWaaL mutants and complemented mutant strains with pBAD33 plasmids carrying different single genes. (B) LPS analyzed by SDS-PAGE (12%) from A. hydrophila AH-3ΔWaaL and AH-3ΔWahA mutants and complemented mutant strains with pBAD33 plasmids carrying different single genes.
Discussion
The bacterial species A. salmonicida comprises five subspecies. A. salmonicida subsp. salmonicida is known as typical A. salmonicida, causing furunculosis in salmonid fish (Bernoth, 1997). Atypical A. salmonicida include the other four subspecies: masoucida, achromogenes, smithia, and pectinolytica which, with the exception of A. salmonicida subsp. pectinolytica, are found as pathogens in a wide variety of fish species (Gudmundsdottir and Bjornsdottir, 2007). A. salmonicida subsp. pectinolytica strains are readily distinguished from the other psychrophilic aeromonads using the following phenotypic characteristics: growth at 35°C, melanin production, growth on KCN broth, mannitol and sucrose fermentation with gas from glucose, and indole plus Voges Proskauer assays. Its ability to degrade polypectate is an unusual feature among Aeromonas species (Pavan et al., 2000).
Interestingly, the structure of the LPS core oligosaccharide from A. salmonicida subsp. pectinolytica is also consistent with the established core structure of A. hydrophila strain AH-3 serotype O34 (Jimenez et al., 2008). Both structures are identical with respect to its inner and outer core regions with D-α-D-Hepp-(1→6)-[β-D-Galp-(1→4)]-D-α-D-Hepp-(1→) trisaccharide outer core fragment being present, while A. salmonicida subsp. salmonicida showed a trisaccharide outer core of α-D-Galp-(1→6)-β-D-GalpNAc-(1→4)-L-α-D-Hepp-(1→). When we inspected and deeply studied the LPS core gene clusters of A. salmonicida subsp. pectinolytica from the published fully sequenced genome, the predicted gene functions were in agreement with the chemical structure. Either by gene analysis or by complementation studies the region 1 of waa from A. salmonicida subsp. pectinolytica corresponds to the A. hydrophila AH-3 waa determined. The genomic analyses of the A. salmonicida subsp. pectinolytica region 1 from strain 34melT versus the Aeromonas whole genomes from mesophilic strains found in Pubmed (http://www.ncbi.nlm.nih.gov/genome/?term=Aeromonas) rendered that approximately 89% of the strains contain the same region 1. Nevertheless, from the 121 whole genomes inspected, 13 of them belonging to the species A. hydrophila, A. veronii, A. caviae, A. media, and Aeromonas sp. showed some different genes (Table 4).
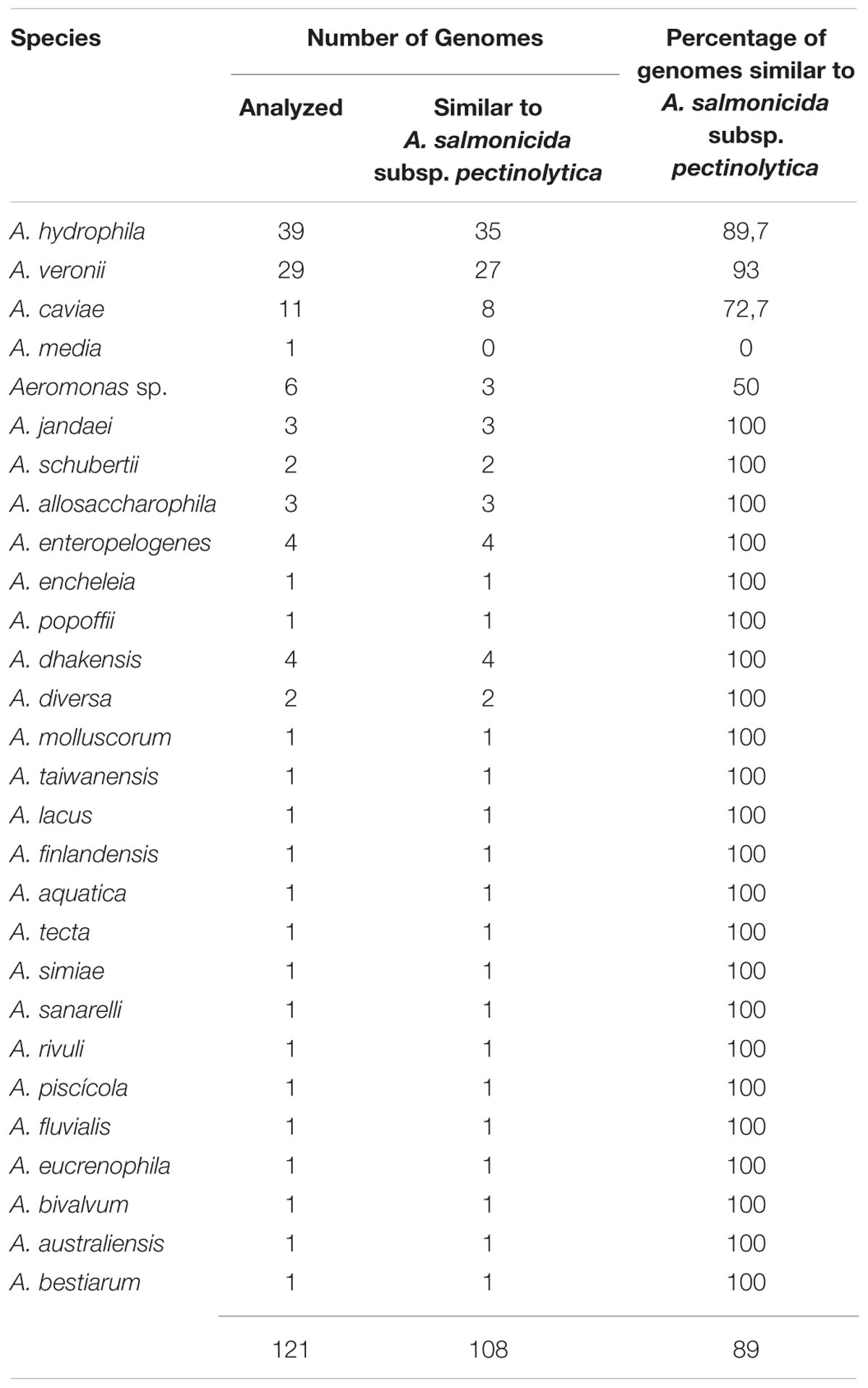
TABLE 4. Genomic analyses of the A. salmonicida subsp. pectinolytica waa region 1 from strain 34melT versus the Aeromonas whole genomes from mesophilic strains.
Aeromonas salmonicida subsp. achromogenes showed a disaccharide in its LPS outer core of β-D-GalpNAc-(1→4)-L-α-D-Hepp-(1→) instead of the previously mentioned A. salmonicida subsp. salmonicida trisaccharide. When we inspected and deeply studied the unique A. salmonicida subsp. achromogenes fully sequenced genome, the analysis and reannotation of the region 1 was in agreement with the biosynthesis of this chemical structure. The wasC and hldD were absent from region 1 of A. salmonicida subsp. achromogenes waa and instead a transposase was present. The transposase DDE found in subsp. achromogenes strain AS03 contains two domains Pfam 13737 and 01609, which are members of the DDE superfamily, which contain three carboxylate residues that are believed to be responsible for coordinating metal ions needed for catalysis. The catalytic activity of this enzyme involves DNA cleavage at a specific site followed by a strand transfer reaction. This family contains transposases for mainly insertion sequence (IS) 4 or 421 (Klaer et al., 1981). WasC is the glycosyl transferase that links Gal in a α1-6 linkage to L,D-HepV in the LPS core (Figure 1) and this monosaccharide residue is missing in the outer core LPS. By genomic analyses we could confirm the complete absence of wasC over the genome and only 126bp are retained between the transposase and wahA genes (11,2% of total gene). No fragment of wasC, was found retained upstream of the transposase gene. Therefore, a complex rearrangement event is probably responsible of the loss of the hldD and wasC genes. HldD (the epimerase for D,D-Hep) is not needed in A. salmonicida subsp. achromogenes LPS-core because D,D-Hep is not found. No hldD gene could be found by genomic analyses in the subsp. achromogenes strain AS03 total genome. Accordingly, the A. salmonicida subsp. achromogenes strains LPS-core migration in SDS-PAGE is faster than the one observed for LPS-core of A. salmonicida subsp. salmonicida strains.
No changes in the outer core trisaccharide (α-D-Galp-(1→6)-β-D-GalpNAc-(1→4)-L-α-D-Hepp-1→) are found in A. salmonicida subsp. masoucida strains, being region 1 of A. salmonicida subsp. salmonicida waa identical to the subspecies masoucida according to chemical structure data, genomic information, LPS-core SDS-PAGE gel migration, and complementation studies. Besides that no full genome is still available for A. salmonicida subsp. smithia strains, the complementation studies and the LPS-core SDS-PAGE gel migration suggest that region 1 of A. salmonicida subsp. smithia is probably identical to the one of A. salmonicida subsp. salmonicida.
No changes were observed in regions 2 and 3 of waa from A. salmonicida subspecies. These data were obtained either by hybridization analysis or by genome study of the different public complete genomes of A. salmonicida strains independently of the subspecies. Furthermore, the genomic analyses of the Aeromonas whole genomes from mesophilic strains found in Pubmed (http://www.ncbi.nlm.nih.gov/genome/?term=Aeromonas) indicate that these genomic regions were identical in all the Aeromonas strains studied, either psychrophilic or mesophilic.
WaaL is the ligase enzyme that links the O-antigen LPS to the lipidA-LPS core, and shows two clear features. The enzyme catalyzes the formation of a glycosidic bond but does not share any protein motif with usual glycosyltransferases, and second the specificity of the reaction is based on the requirement for a specific lipid A-core OS acceptor structure but not the O-antigen LPS or any other undecaprenol-P-linked substrate (Valvano, 2011). According to these features, the WaaL from subsp. salmonicida, subsp. masoucida, and subsp. smithia are identical in amino acid sequence (Figure 8). WaaL subsp. smithia sequence was obtained after sequencing pBAD33-WaaLsmi. WaaL from subsp. achromogenes showed a large similarity (nearly identity only with a few amino acid residues changes) to the previous ones, while WaaL from subsp. pectinolytica showed a clearly decreased similarity versus the rest of the WaaL from other salmonicida subspecies (Figure 8). The A. salmonicida subsp. pectinolytica WaaL from strain 34melT showed more identity with many WaaL from several mesophilic Aeromonas strains belonging to different species than to WaaL from other A. salmonicida subspecies.

FIGURE 8. Alignment of the WaaL aminoacid sequence from A. salmonicida subsp. salmonicida A450, A. salmonicida subsp. masoucida NBRC13784, A. salmonicida subsp. achromogenes strain AS03, A. salmonicida subsp. pectinolytica strain 34melT, and A. salmonicida subsp. smithia CECT5179. Different aminoacids residues among the sequences are labeled in red and bold and inside a square box.
It can be observed that the LPS inner core in A. salmonicida strains is well-conserved; however, there is some structural diversity in the LPS outer core. From the different typical and atypical A. salmonicida strains we can conclude that subsp. salmonicida, subsp. masoucida, and probably subsp. smithia strains shared the same kind of LPS outer core. A. salmonicida subsp. achromogenes strains showed a similar LPS outer core but lacked one branched external residue not affecting the O-antigen LPS linkage. However, A. salmonicida subsp. pectinolytica strains showed a rather changed LPS outer core, identical to many mesophilic Aeromonas strains LPS outer core. However, these LPS-core genes those are structural non-variable genes could be among others of interest for specific phylogenetic analyses.
Author Contributions
All authors listed, have made substantial, direct and intellectual contribution to the work, and approved it for publication.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by Plan Nacional de I + D + i (Ministerio de Economía y Competitividad, Spain) and from Generalitat de Catalunya (Centre de Referència en Biotecnologia). We thank Maite Polo for her technical assistance and the Servicios Científico-Técnicos from University of Barcelona.
References
Aquilini, E., Merino, S., Regué, M., and Tomás, J. M. (2014). Genomic and proteomic studies of Plesiomonas shigelloides lipopolysaccharide core biosynthesis. J. Bacteriol. 196, 556–567. doi: 10.1128/JB.01100-13
Aquilini, E., and Tomás, J. M. (2015). “Lipopolysaccharides (Endotoxins),” Reference Module in Biomedical Science (Amsterdam: Elsevier). doi: 10.1016/B978-0-12-801238-3.07799-0
Austin, B., and Austin, D. A. (2007). “Characteristics of the pathogens: gram-negative bacteria,” in Bacterial Fish Pathogens: Diseases of Farmed and Wild Fish, eds B. Austin and D. A. Austin (Chichester: Springer Praxis Publishing), 81–150.
Austin, B., Austin, D. A., Dalsgaard, I., Gudmundsdottir, B. K., Høie, S., Thornton, J. M., et al. (1998). Characterization of atypical Aeromonas salmonicida by different methods. Syst. Appl. Microbiol. 21, 50–64. doi: 10.1016/S0723-2020(98)80008-8
Belland, R. J., and Trust, T. J. (1985). Synthesis, export, and assembly of Aeromonas salmonicida A-layer analysed by transposon mutagenesis. J. Bacteriol. 163, 877–881.
Bernoth, E. M. (1997). “Furunculosis: the history of the disease and of disease research,” in Furunculosis: Multidisciplinary Fish Disease Research, eds E. M. Bernoth, A. E. Ellis, P. J. Midtlyng, G. Olivier, and P. Smith (London: Academic Press), 1–20.
Gudmundsdottir, B. K., and Bjornsdottir, B. (2007). Vaccination against atypical furunculosis and winter ulcer disease of fish. Vaccine 25, 5512–5523. doi: 10.1016/j.vaccine.2007.02.009
Han, J. E., Kim, J. H., Shin, S. P., Jun, J. W., Chai, J. Y., and Park, S. C. (2013). Draft genome sequence of Aeromonas salmonicida subsp. achromogenes AS03, an atypical strain isolated from crucian carp (Carassius carassius) in the Republic of Korea. Genome Announc. 1:e00791-13. doi: 10.1128/genomeA.00791-13
Hanahan, D. (1983). Studies on transformation of Escherichia coli with plasmids. J. Mol. Biol. 166, 557–580. doi: 10.1016/S0022-2836(83)80284-8
Holst, O. (2002). Chemical structure of the core region of lipopolysaccharides – an update. Trends Glycosci. Glycotechnol. 14, 87–103. doi: 10.4052/tigg.14.87
Holst, O. (2007). The structures of core regions from enterobacterial lipopolysaccharides – an update. FEMS Microbiol. Lett. 271, 3–11. doi: 10.1111/j.1574-6968.2007.00708.x
Ishiguro, E. E., Kay, W. W., Ainsworth, T., Chamberlain, J. B., Buckley, J. T., and Trust, T. J. (1981). Loss of virulence during culture of Aeromonas salmonicida at high temperature. J. Bacteriol. 148, 393–400.
Jimenez, N., Canals, R., Lacasta, A., Kondakova, A. N., Lindner, B., Knirel, Y. A., et al. (2008). Molecular analysis of three Aeromonas hydrophila AH-3 (serotype O34) lipopolysaccharide core biosynthesis gene clusters. J. Bacteriol. 190, 3176–3184. doi: 10.1128/JB.01874-07
Jimenez, N., Lacasta, A., Vilches, S., Reyes, M., Vazquez, J., Aquilini, E., et al. (2009). Genetics and proteomics of Aeromonas salmonicida lipopolysaccharide core biosynthesis. J. Bacteriol. 191, 2228–2236. doi: 10.1128/JB.01395-08
Klaer, R., Kuhn, S., Tillmann, E., Fritz, H. J., and Starlinger, P. (1981). The sequence of IS4. Mol. Gen. Genet. 181, 169–175. doi: 10.1007/BF00268423
Merino, S., de Mendoza, E., Canals, R., and Tomás, J. M. (2015). Functional genomics of the Aeromonas salmonicida lipopolysaccharide O-antigen and A-layer from typical and atypical strains. Mar. Drugs 13, 3791–3808. doi: 10.3390/md13063791
Milton, D. L., O’Toole, R., Horstedt, P., and Wolf-Watz, H. (1996). Flagellin A is essential for the virulence of Vibrio anguillarum. J. Bacteriol. 178, 1310–1319.
Pavan, M. E., Abbott, S. L., Zorzopulos, J., and Janda, J. M. (2000). Aeromonas salmonicida subsp. pectinolytica subsp. nov., a new pectinase-positive subspecies isolated from a heavily polluted river. Int. J. Syst. Evol. Microbiol. 50, 1119–1124. doi: 10.1099/00207713-50-3-1119
Read, J. A., Ahmed, R. A., Morrison, J. P., Coleman, W. G. Jr., and Tanner, M. E. (2004). The mechanism of the reaction catalyzed by ADP-beta-L-glycero-D-manno-heptose 6-epimerase. J. Am. Chem. Soc. 126, 8878–8879. doi: 10.1021/ja0485659
Scott, M. (1968). The pathogenicity of Aeromonas salmonicida (Griffin) in sea and brackish waters. J. Gen. Microbiol. 50, 321–327. doi: 10.1099/00221287-50-2-321
Shaw, D. H., Hart, M. J., and Lüderitz, O. (1992). Structure of the core oligosaccharide in the lipopolysaccharide isolated from Aeromonas salmonicida ssp. salmonicida. Carbohydr. Res. 231, 83–91. doi: 10.1016/0008-6215(92)84010-P
Shaw, D. H., Lee, Y. Z., Squires, M. J., and Lüderitz, O. (1983). Structural studies on the O-antigen of Aeromonas salmonicida. Eur. J. Biochem. 131, 633–638. doi: 10.1111/j.1432-1033.1983.tb07310.x
Valvano, M. A. (2011). Common themes in glycoconjugate assembly using the biogenesis of O-antigen lipopolysaccharide as a model system. Biochemistry (Mosc.) 76, 729–735. doi: 10.1134/S0006297911070029
Wang, Z., Li, J., Vinogradov, E., and Altman, E. (2006). Structural studies of the core region of Aeromonas salmonicida subsp. salmonicida lipopolysaccharide. Carbohydr. Res. 341, 109–117. doi: 10.1016/j.carres.2005.10.017
Keywords: Aeromonas salmonicida, typical and atypical, lipopolysaccharide core, inner and outer core chemical structure, genomics
Citation: Merino S and Tomás JM (2016) The Aeromonas salmonicida Lipopolysaccharide Core from Different Subspecies: The Unusual subsp. pectinolytica. Front. Microbiol. 7:125. doi: 10.3389/fmicb.2016.00125
Received: 06 November 2015; Accepted: 25 January 2016;
Published: 11 February 2016.
Edited by:
Brigitte Lamy, CHU Montpellier, FranceReviewed by:
Steve J. Charette, Université Laval, CanadaEvguenii Vinogradov, National Research Council Canada, Canada
Copyright © 2016 Merino and Tomás. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Juan M. Tomás, jtomas@ub.edu