 Valérie Duval
Valérie Duval Kimberly Foster
Kimberly Foster Jennifer Brewster
Jennifer Brewster Stuart B. Levy
Stuart B. Levy- Center for Adaptation Genetics and Drug Resistance, Department of Molecular Biology and Microbiology, Tufts University School of Medicine, Boston, MA, United States
In Escherichia coli, OmpF is an important outer membrane protein, which serves as a passive diffusion pore for small compounds including nutrients, antibiotics, and toxic compounds. OmpF expression responds to environmental changes such as temperature, osmolarity, nutrients availability, and toxic compounds via complex regulatory pathways involving transcriptional and post-transcriptional regulation. Our study identified a new regulatory cascade that controls the expression of OmpF porin. This pathway involves BluR, a transcriptional regulator repressing the expression of the ycgZ-ymgABC operon. We showed that BluR was responsible for the temperature-dependent regulation of the ycgZ-ymgABC operon. Furthermore, our results showed that independent expression of YcgZ led to a decreased activity of the ompF promoter, while YmgA, YmgB, and YmgC expression had no effect. We also determined that YcgZ accumulates in the absence of the Lon protease. Thus, mutation in bluR leads to de-repression of ycgZ-ymgABC transcription. With a second mutation in lon, YcgZ protein accumulates to reach levels that do not allow increased expression of OmpF under growth conditions that usually would, i.e., low temperature. With BluR responding to blue-light and temperature, this study sheds a new light on novel signals able to regulate OmpF porin.
Introduction
The outer membrane of Gram-negative bacteria provides a physical barrier to hydrophobic and hydrophilic compounds including many toxic molecules (Pagès et al., 2008). Embedded in the outer membrane, the porins (outer membrane proteins or OMPs) have multiple functions: allowing the diffusion of small molecules, stabilizing the cell envelope, and acting as receptors for phages and bacteriocins or as virulence factors in pathogenic bacteria (Achouak et al., 2001; Nikaido, 2003; Galdiero et al., 2012). Three major porins are found in abundance in the outer membrane of Escherichia coli: OmpA, OmpC, and OmpF (Achouak et al., 2001). With only a small fraction of the porin forming open channels, OmpA seems to be mainly involved in maintaining the shape of the cell (Sugawara and Nikaido, 1994). Conversely, OmpF and OmpC form hydrophilic pores that allow the diffusion of small nutrients and toxic compounds (Chopra and Eccles, 1978; Yoshimura and Nikaido, 1985; Mortimer and Piddock, 1993; Nikaido, 1994, 2003). Although OmpF and OmpC display a similar structure, each porin harbors a unique electrostatic pore potential and consequently a distinct specificity and flow rates for solutes (Cowan et al., 1995; Basle et al., 2003). In this context, OmpF plays a crucial role in the accumulation of small hydrophilic antibiotics such as monoanionic cephalosporins, tetracyclines, and fluoroquinolones (Yoshimura and Nikaido, 1985; Cohen et al., 1989; Mortimer and Piddock, 1993; Duval et al., 2009). In order to respond to changes in environmental conditions, E. coli adjusts OmpF and OmpC expression through a complex regulatory network utilizing both transcriptional and translational regulation (for review, see Forst and Inouye, 1988; Pratt et al., 1996; Vogel and Papenfort, 2006). For instance, the osmolarity-dependent transcriptional control of ompF and ompC is exerted via the EnvZ/OmpR two-component signal transduction system in which EnvZ, an inner membrane histidine protein kinase, senses osmotic signals and transmits them to the transcription factor OmpR (Igo et al., 1989; Egger and Inouye, 1997; Yoshida et al., 2006). High osmolarity leads to lower OmpF levels, while relative expression of OmpC is increased (Forst and Inouye, 1988; Mizuno and Mizushima, 1990; Pratt et al., 1996). In addition, expression of OmpF and OmpC is controlled at the post-transcriptional level by non-translated small RNAs such as MicF, MicC, and RybB (Mizuno et al., 1984; Chen et al., 2004; Gogol et al., 2011).
A major environmental parameter that affects OmpF porin expression is temperature. OmpF is abundant in E. coli outer membrane at ambient temperature, while growth at 37°C leads to decreased amount of OmpF. It is assumed that the downregulation of OmpF at body temperature limits the entry of toxic bile salts into the periplasm while the bacterium is in the host’s intestine (Pratt et al., 1996; Nikaido, 2003). In a previous study, we showed that mutation in two genes, lon and bluR (ycgE), prevented the upregulation of OmpF in E. coli K-12 grown at ambient temperature (Duval et al., 2009). BluR, a regulator harboring a MerR-like N-terminal domain, has been shown to directly repress the transcription of the adjacent ycgZ-ymgABC operon (ZABC operon, Figure 1; Tschowri et al., 2009). The lon locus encodes the Lon protease, an ATP-dependent serine protease involved in the degradation of unstable and misfolded proteins (Tsilibaris et al., 2006; Van Melderen and Aertsen, 2009). Lon also plays a major role in regulating multiple biological processes by controlling the abundance of specific regulatory proteins such as MarA, RcsA, and SulA (Mizusawa and Gottesman, 1983; Torres-Cabassa and Gottesman, 1987; Griffith et al., 2004). With no available studies describing the regulation of OmpF by BluR and Lon, we investigated how BluR and Lon together control the abundance of the OmpF porin. Our study identified the ZABC operon as an intermediate in the regulation of OmpF by BluR and Lon. Precisely, we identified YcgZ as a novel repressor of ompF expression. Using a transcriptional fusion of the ompF promoter with lacZ (PompF-lacZ), we showed that YcgZ acted on ompF promoter. Finally, our study showed that the amount of YcgZ, when expressed from a plasmid, was amplified and highly stable in a lon mutant of E. coli, identifying YcgZ as a novel substrate of the Lon protease.
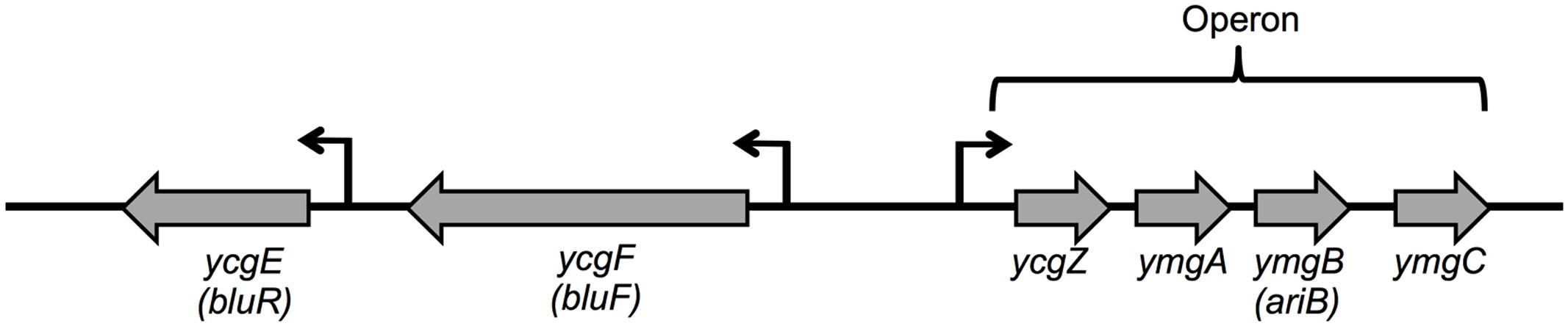
FIGURE 1. Organization of ycgZ-ymgABC, ycgE, and ycgF ORFs in Escherichia coli. The bent arrows indicate the transcription start for each gene. ycgZ, ymgA, ymgB, and ymgC genes are expressed as one transcript. ycgE, ycgF, and ymgB have been renamed bluR, bluF, and ariB, respectively.
Materials and Methods
Bacterial Strains and Growth Conditions
The bacterial strains used in this study are listed in Table 1. E. coli strains were cultured under agitation in LB medium (tryptone 10 g l-1, yeast extract 5 g l-1, NaCl 5 g l-1). The same medium containing 1.5% agar was used for growing bacteria on plates. Selection of E. coli after transformation with pBAD and pMPM vectors was performed using ampicillin 100 μg ml-1 and tetracycline 15 μg ml-1, respectively. Kanamycin 25 μg ml-1 and chloramphenicol 12 μg ml-1 was used for selection of chromosomal insertion of kan and cat genes.

TABLE 1. Bacterial strains and plasmids used in this study.
Plasmids
Expression plasmids used in this study are listed in Table 1 and were constructed as follows. The nucleotide sequence of ycgZ, ymgA, ymgB, and ymgC was amplified by polymerase chain reaction (PCR) using the primers listed in Supplementary Table S1 and E. coli AG100 genomic DNA as template. The fragments were cloned into the pBAD/HisA and pMPM vectors using restriction sites indicated in Table 1. We constructed the pDVMBluR by PCR amplification of bluR nucleotide sequence, the 200 bases upstream of start codon GTG, and the 100 bases downstream of its stop codon TAA using the primers listed in Supplementary Table S1. The PCR fragment was then ligated into the pMPM plasmid using the restriction sites EcoRI and XhoI, resulting in plasmid pDVMBluR. All nucleotide sequences were verified at the Tufts University Core Facility (Tufts University School of Medicine, Boston, MA, United States).
Gene Deletion
Targeted deletion of bluR and ycgZ-ymgABC, and subsequent marker removal were made using the λRed recombinase method previously described (Datsenko and Wanner, 2000). The Flp recombination target (FRT)-flanked chloramphenicol resistance gene (cat) has been amplified by PCR from plasmid pKD3 using primers listed in Supplementary Table S1. bluR-PA/bluR-PB and ycgZ-PA/ymgC-PB primers contain sequences upstream and downstream of bluR and of ycgZ-ymgABC operon, respectively. The PCR product was gel-purified and concentrated by ethanol precipitation. Transformants carrying the Red helper plasmid pKD46 were then grown in LB medium with 100 μg ml-1 ampicillin and 10 mM L-arabinose at 30°C to an optical density at 600 nm (OD600) of 0.6 and then made electro-competent. Electroporation was done using 200 ng of PCR product. Chloramphenicol resistant clones were selected on LB agar plates containing chloramphenicol 12 μg ml-1. Correct integration of the cat gene in the targeted genes was verified by PCR using the primers listed in Supplementary Table S1. Appropriate chloramphenicol resistant clones were subsequently transformed with the pCP20 plasmid and ampicillin resistant clones were selected at 30°C on LB agar plates containing ampicillin 100 μg ml-1. The transformants were then colony-purified non-selectively at 42°C on LB agar and then tested for loss of ampicillin and chloramphenicol resistance. Deletions were further verified by PCR using the primers listed in Supplementary Table S1.
LacZ Transcriptional Fusion and β-Galactosidase Assays
To construct the plasmid pDV415O, amplification of ompF promoter (PompF) was carried out by PCR using chromosomal DNA from strain E. coli AG100 as template and the primers ompF1 and ompF2 listed in Supplementary Table S1. The PompF fragment was 273 bp long (from -273 to +1 relative to the transcription start) and was cloned into the pGEM-T Easy vector (Promega) following the manufacturer instruction. The resulting plasmid was digested with EcoRI and BamHI and the fragment corresponding to PompF was ligated to the similarly cut vector pRS415 yielding the plasmids pDV415O. The sequence of the PompF-lacZ fusion in pDV415O was then verified at the Tufts University Core Facility. Insertion of PompF-lacZ into E. coli chromosome was realized as followed. Recombination between the pDV415O and λRZ5 (Silhavy et al., 1984; Simons et al., 1987) resulted in a lysate bearing λRZ5 (PompF-lacZ). This was used to infect strains BW25113, a λ- and lac- strain of E. coli. AmpR Lac+ lysogens were selected and purified on LB agar containing ampicillin 20 μg ml-1 and X-Gal (5-bromo-4-chloro-3-indolyl-β D-galactopyranoside) 40 μg ml-1. Lysates from these lysogens were then used to infect at low multiplicity of infection (MOI = 0.005) strain BW25113 and derivative mutants. AmpR Lac+ lysogens were again isolated and the resulting strains were confirmed by PCR, as previously described (Powell et al., 1994), to have a single copy of the transcriptional fusion located in the λatt site on the chromosome. To assess the β-galactosidase (LacZ) activity, overnight cultures of fresh colonies of E. coli carrying a chromosomal λPompF-lacZ fusion were grown in LB medium containing ampicillin 20 μg ml-1 and were subsequently diluted to an OD600 of 0.05 in identical medium for growth with no antibiotic added. When the OD600 reached 0.6, β-galactosidase (LacZ) activity was assayed by first rendered the cells permeable with 0.005% sodium dodecyl sulfate and 0.05% chloroform. LacZ activity was expressed in Miller units as previously described (Miller, 1972). All assays were carried out at least in three independent experiments.
RNA Isolation
Fresh colonies of E. coli strains were grown overnight (14–16 h) in 3 ml of LB medium. The cultures were then diluted to an OD600 of 0.05 in 3 ml of identical fresh medium and cells were grown until an OD600 of ∼0.6. The total RNAs were isolated from 0.5 ml of culture using the Trizol Reagent (Thermo Fisher Scientific) following the manufacturer protocol. Directly after the preparation of the RNAs, the integrity of the RNAs was evaluated on a bleach gel stained with ethidium bromide (Aranda et al., 2012). The RNAs amount was quantified by absorbance at 260 nm (A260) and purity was evaluated by a A260/A280 ratio >1.9 and a A260/A230 ratio >1.7 using a Nanodrop ND-1000 spectrophotometer (Thermo Fisher Scientific). RNAs were stored for no more than 3 days at -80°C before DNase treatment and cDNA synthesis (see Reverse Transcription and Quantitative PCR Analysis).
Reverse Transcription and Quantitative PCR Analysis
Two micrograms of purified RNAs were treated using the TurboTM DNase (Thermo Fisher Scientific) following the manufacturer instructions. A total of 500 ng of RNAs was then used to synthesize cDNA by reverse transcription using the Quantitect Reverse Transcription kit (Qiagen) and following the manufacturer instructions. To control for chromosomal DNA contamination, the reverse transcription step was also performed with reaction mixtures containing no reverse transcriptase and was used as a negative control in subsequent quantitative PCR (qPCR) reactions. Primers used for the qPCR were designed using the online PrimerQuest tool from Integrated DNA Technologies and are listed in Supplementary Table S1. Amplification efficiency and specificity for each set of primers are reported in Supplementary Figure S1. All qPCR reactions were performed using a Roche LightCycler 480 instrument II. The following experimental run protocol was used: UDP activation (50°C for 2 min), denaturation (95°C for 2 min), quantification program repeated 40 times (denaturation at 95°C for 5 s, anneal/extend 60°C for 30 s with a single fluorescence measurement), melting curve program (60–95°C with a heating rate of 0.15°C s-1 and a continuous fluorescence measurement). Reactions were carried out in 20 μl with 2 μl of diluted cDNA, 0.4 μl of 10 μM forward primer (0.2 μM final concentration), 0.4 μl of 10 μM reverse primer (0.2 μM final concentration), 7.2 μl H20 and 10 μl of the Power Up SYBR Green Master Mix (Applied Biosystems by Life Technologies). After the reverse transcription step, the cDNA samples were diluted fivefold and used as templates for qPCR amplification of gapA, ompF, and ycgZ. A mix with no cDNA was also prepared (non-template control, NTC) and was run in parallel. NTC wells did either gave a CT > 35 or no CT at all. We quantified the relative expression of ompF and ycgZ transcripts to the reference gene gapA. Using the Pfaffl method (Pfaffl, 2001), we determined the ratio R of a transcript expressed in a sample versus that expressed in the wild type strain grown at 37°C (see CT and calculations in Supplementary Tables S2–S7). gapA encodes the glyceraldehydes 3-phosphate deshydrogenase-A.
Statistical Analysis
We report the average (mean) and the standard deviation (SD) from at least three experimental values. In Figures 2B and 4B, we report the ratio R3 (±SD3) of two numbers R1 ± SD1 and R2 ± SD2, with . The statistical significance of differences between two averages was determined by a Student’s t-test (two independent samples, with two-tailed distribution) using GraphPad Prism software.1
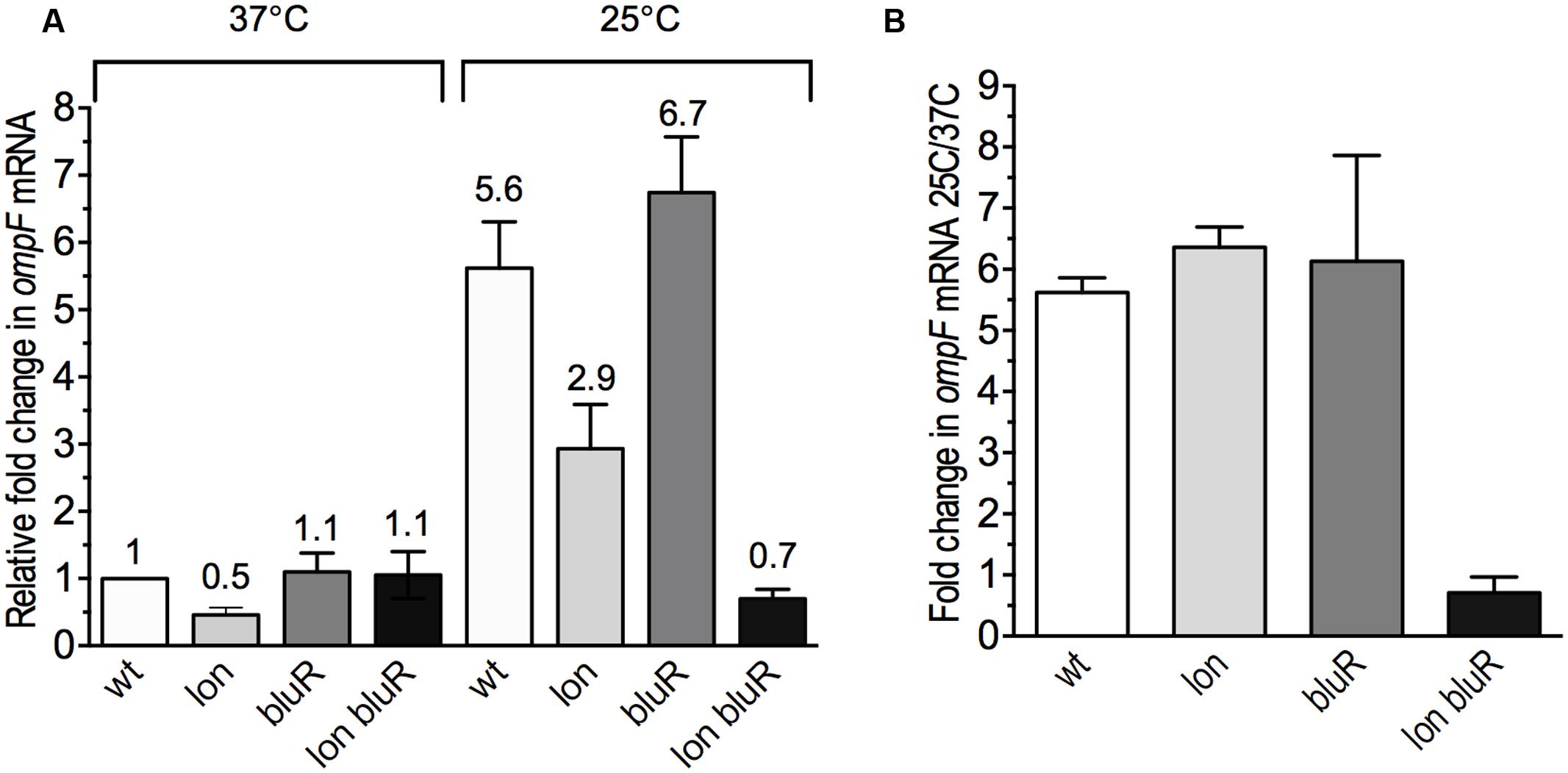
FIGURE 2. Effect of lon and bluR mutations on ompF mRNA levels in E. coli grown at 25 and 37°C. The cells were grown to exponential phase (optical density at 600 nm ∼0.6) in LB medium. Total RNAs were prepared and used to measure the OmpF mRNA levels by RT-qPCR. (A) Fold change in OmpF mRNA levels relative to the wild type strain AG100 grown at 37°C. (B) Fold change in OmpF mRNA levels for all strains grown at 25 versus 37°C. The numbers represent the means and standard deviations of expression levels from three independent experiments performed in duplicate. AG100, wt; M113R, lon; AGEZ3, bluR; M113REZ3, lon bluR. See Supplementary Tables S2, S3 for details.
Steady-State Levels of Protein and Stability Assays
Overnight culture of E. coli wild type (BW25113) and lon (JW0419-1) strains carrying plasmids pBAD/HisA and derivative pDVBZ, pDVBZ-XP, pDVBA, pDVBB, and pDVBC were grown in LB medium in the presence of 100 μg ml-1 ampicillin. Fresh identical medium supplemented with L-arabinose was then inoculated to an optical density measured at 600 nm of 0.05 and cells were grown at 37 or 25°C to OD600 of 1. Whole cell extracts were prepared for analysis of the steady-state levels of protein. To assess the intracellular stability of YcgZ, 150 μg ml-1 chloramphenicol was added to stop the proteins synthesis and the cultures were kept at the indicated temperatures. Samples to be used for preparing whole cell extracts were removed at indicated times (0–60 min). Cell extracts were prepared as follows: 1 ml of culture was centrifuged and the cells pellet suspended in 200 μl of lysis buffer per OD600 of 1 [Tris–HCl 10 mM pH 8.0, EDTA 0.5 mM, CaCl2 10 mM, and 1 unit ml-1 of DNase (Promega)]. Cells were sonicated on ice two times 20 pulses using a Branson Sonifier 250 and the following parameters: output control = 1 and a duty cycle = 50%. Protein concentration was determined using the Pierce 660 nm Protein Assay Reagent and bovine serum albumin as a standard (Thermo Fisher Scientific). Eight micrograms of proteins were separated on a 16% acrylamide gel in denaturing condition (100 mM Tris, 100 mM Tricine, and 0.1% SDS). The gels were stained with Coomassie Brilliant Blue R-250 (Sigma-Aldrich) or used for western blot analysis.
Detection of XPress-Tagged YcgZ by Western Blot
After separation on a 16% acrylamide gel, the proteins were electro-transferred to a nitrocellulose membrane (Millipore, Billerica, MA, United States). The membrane was incubated overnight at 4°C in Tris-borate-saline (TBS) buffer supplemented with 3% milk powder. The membrane was then incubated for 2 h at room temperature with monoclonal anti-XPress antibodies (Thermo Fisher Scientific) diluted in TBS (1/6,000). After three 15 min washes with TTBS (TBS supplemented with 0.05% Tween 20), the membrane was incubated for 2 h with alkaline phosphatase-coupled anti-mouse IgG antibodies (Promega) diluted 1/10,000 in TBS followed by three 15 min washes with TTBS and two 5 min washes with TBS. XPress-YcgZ was visualized by adding 5-bromo-4-chloro-3-indolyl phosphate and nitroblue tetrazolium following the manufacturer’s instructions (Promega).
Results
Escherichia coli Carrying Mutations in both lon and bluR Failed to Increased OmpF Expression at 25°C
When grown at 37°C, E. coli expresses low amount of OmpF in the outer membrane, while decreased temperatures lead to increased levels of the porin (Pratt et al., 1996; Nikaido, 2003). A previous study performed in our laboratory demonstrated that E. coli carrying mutations in both lon and bluR loci resulted in significantly lower amounts of the porin when the cells were grown under low temperature conditions (see protein level of OmpF in Duval et al., 2009). To better characterize the effect of lon and bluR on ompF expression, we performed reverse transcription and qPCR (RT-qPCR) and compared the level of OmpF messenger in wild type E. coli AG100 and derivative strains carrying mutations in lon, bluR, and lon bluR. All strains were grown at 37 and 25°C. Our data show that wild type E. coli grown at 25°C induces OmpF expression by approximately sixfold when compared to that of wild type E. coli grown at 37°C (Figures 2A,B). Our data also indicate that a single mutation in bluR has no significant effect on OmpF mRNA level, while a lon mutation slightly decreases the expression of OmpF at both temperatures (Figure 2A). Nevertheless the fold change in OmpF mRNA for both lon and bluR mutants grown at 25 versus 37°C is similar to that of wild type AG100 (Figure 2B). However, a lon bluR mutant carrying both mutations fails to increase ompF expression when grown at 25°C.
To further characterize the role of Lon and BluR in regulating OmpF expression, we constructed a transcriptional reporter fusion of ompF promoter with lacZ (PompF-lacZ). The resulting fusion did not carry ompF 5′-UTR affected by MicF (Andersen and Delihas, 1990), but instead carried lacZ 5′-UTR. In this case, we prevented post-transcriptional effects on ompF 5′-UTR. A single copy of the reporter fusion was then integrated at the att(λ) site of wild type E. coli BW25113, a lac-λ- strain, and of derivative mutants lon, bluR, and lon bluR. LacZ activity was then assayed in cells grown at 37 and 25°C. Our results from the β-galactosidose assays show that transcription of PompF-lacZ increases by approximately threefold in the wild type strain grown under low temperature conditions (Figure 3). In contrast, the lon bluR double mutant fails to increase PompF-lacZ expression at 25°C and displays similar PompF-lacZ activities at 25 and 37°C (Figure 3). Plasmid mediated expression of bluR in the lon bluR mutant restores PompF-lacZ levels similar to that of wild type (Figure 3). Our data also show that a single mutation in either lon or bluR has no effect on PompF-lacZ transcription (Figure 3). We suspect that the decreased OmpF mRNA level observed for the lon mutant (see above) is likely due to a lower stability of the messenger in the absence of the Lon protease since no difference is detected at the transcriptional level.
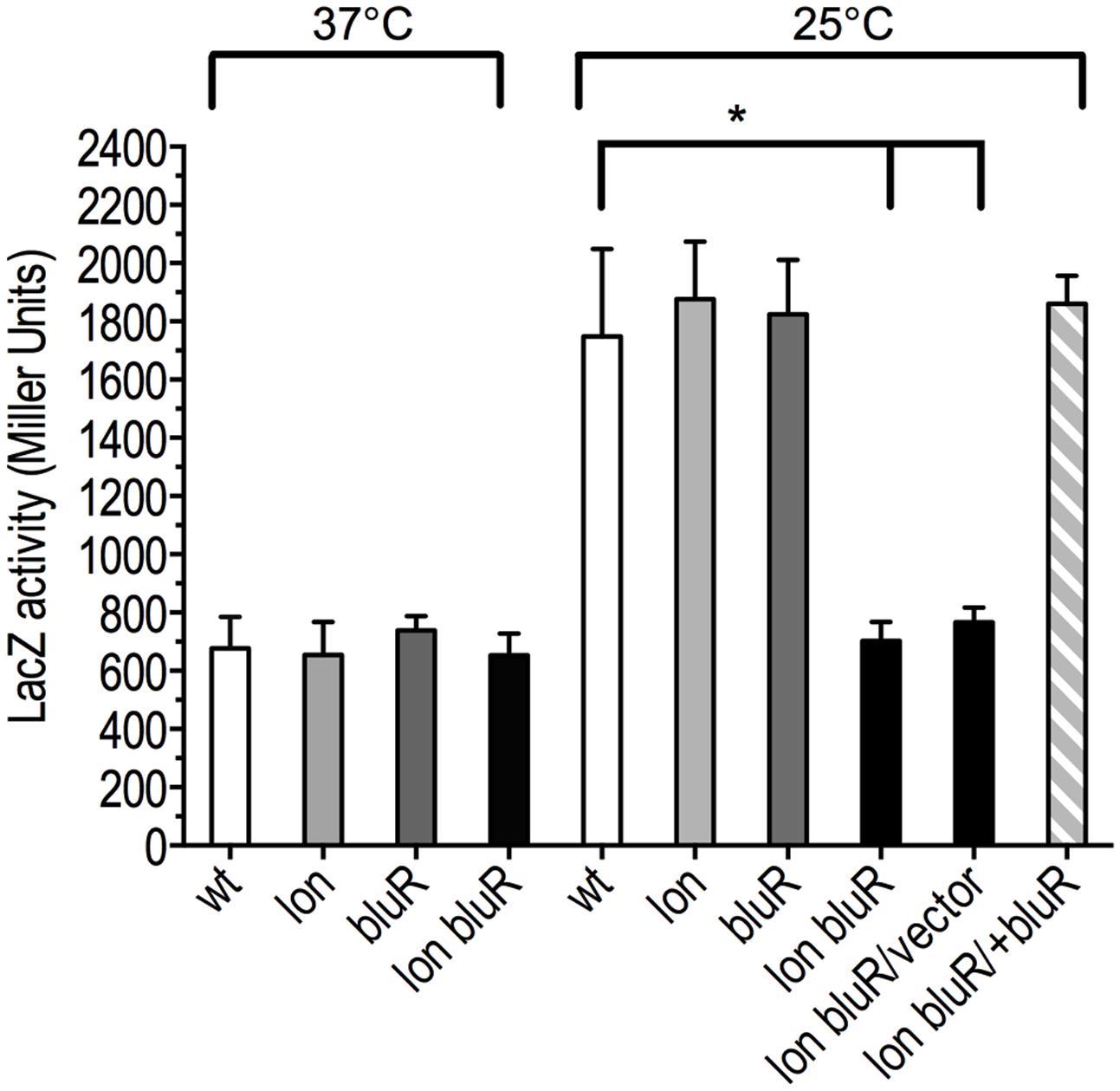
FIGURE 3. lon and bluR mutations lead to decreased ompF transcription at 25°C. The β-galactosidase activity (Miller units) of a single chromosomal copy of PompF-lacZ was measured for cells grown to exponential phase (optical density at 600 nm ∼0.6) in LB medium at 25 and 37°C. Wild type, VDL25113; lon, VDL0419; bluR, VDL101; lon bluR, VDL102; vector, pMPM; +bluR, pDVMbluR. Numbers represent the means and standard deviations from at least three independent experiments. Statistically significant differences for a mutant compared to wild type are shown as asterisk (∗P < 0.05).
The simplest model illustrating the effect of both lon and bluR mutations on ompF transcription under the growth conditions used in our experiments implies that BluR represses a locus coding for an intermediate protein that is a substrate of the Lon protease. This intermediate protein would act as a repressor of the ompF promoter. In this model, the inactivation of bluR leads to an increased transcription of the intermediate locus, and only in the absence or with a reduced activity of Lon, can the intermediate protein accumulate and considerably repress ompF at 25°C, a condition that normally permits increased expression of the porin in wild type E. coli. In an otherwise direct activation of ompF promoter by BluR, deletion of bluR alone would lead to decreased ompF transcription.
BluR Controls the Temperature-Dependent Expression of the ycgZ-ymgABC Operon
We subsequently aimed to identify the intermediate locus controlled by BluR and involved in the regulation of ompF expression. BluR has been previously described to directly repress the transcription of the ycgZ-ymgABC operon (ZABC; Tschowri et al., 2012). This operon encodes small proteins of 78–90 amino acid residues involved in biofilm formation through a mechanism that is yet to be determined (Tschowri et al., 2009). Using RT-qPCR, we measured the expression of the ZABC operon in AG100 (wt), AGEZ3 (bluR), and M113REZ3 (lon bluR) derivative strains. When comparing YcgZ mRNA levels at 37 and 25°C for the wild type strain, our results shows a ∼46-fold increased expression of ycgZ when the cells were grown at ambient temperature (Figures 4A,B). Our data also determine that a bluR mutant increases expression of the ZABC operon by ∼100- and ∼4-fold in cells grown at 37 and 25°C, respectively (Figure 4A), indicating a strong repression of the ycgZ promoter by BluR at 37°C. It was previously shown by Tschowri et al. (2009) that BluR–DNA interaction is released in the presence of BluF, a direct antagonist of BluR whose activity is induced by low temperature. We believe that the weaker repression of the ycgZ promoter observed at 25°C likely comes from the BluR inactivation by BluF under these growth conditions. At 37°C, BluR repressor is fully active and its deletion leads to a large de-repression of the ycgZ promoter. Our results in Figure 4A also show that the addition of a lon mutation to a bluR mutation (lon bluR) leads to a slight increased in ycgZ mRNA levels when compared to that of bluR. For AGEZ3 (bluR) and M113REZ3 (lon bluR), the low temperature mediated induction is only approximately twofold, overall indicating that bluR is mainly responsible for the temperature-dependent expression of ycgZ.
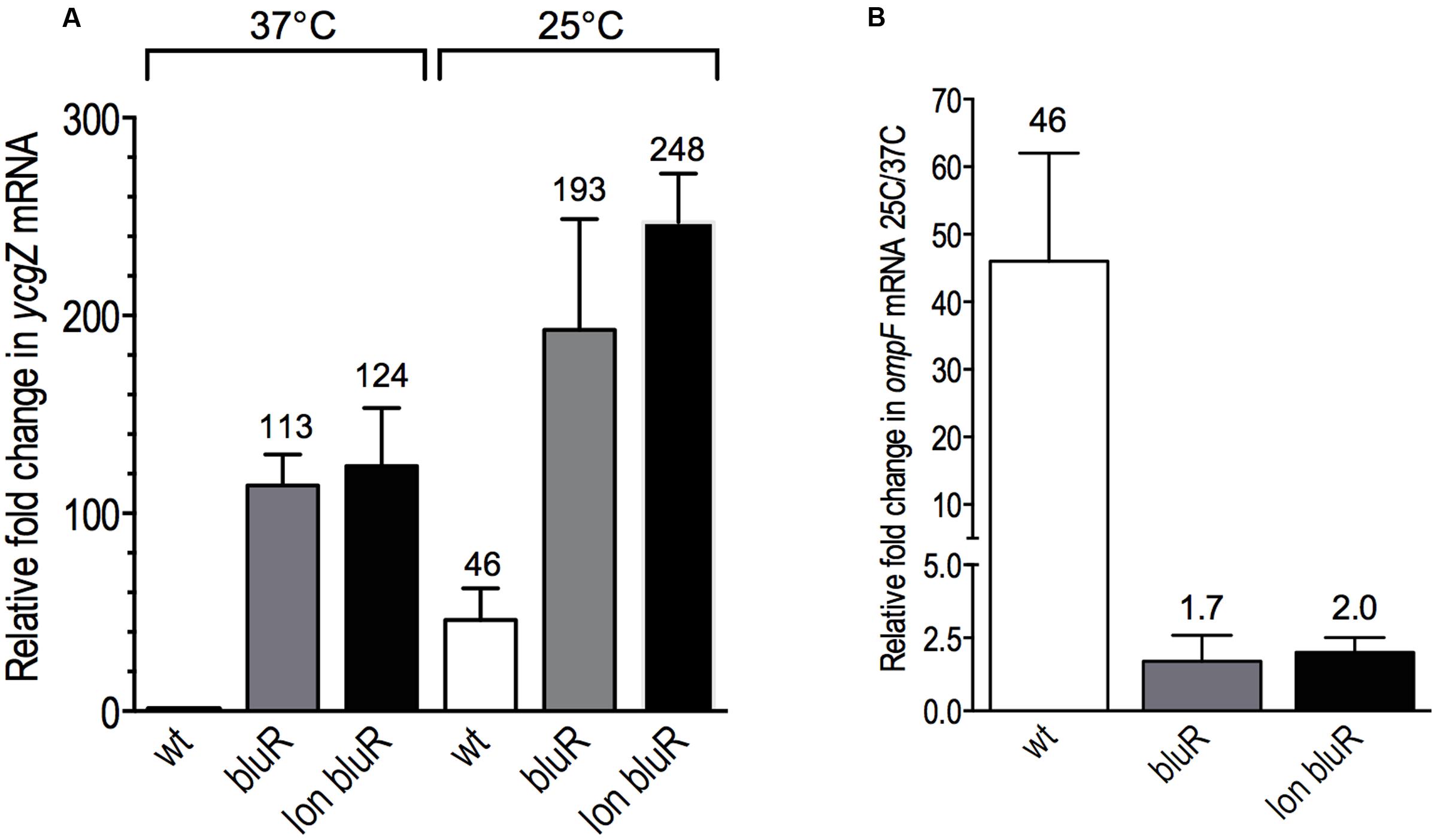
FIGURE 4. BluR is controlling the temperature-dependent expression of ycgZ. (A) Fold change in YcgZ mRNA levels relative to the wild type strain AG100 grown at 37°C. (B) Fold change in YcgZ mRNA levels for all strains grown at 25 versus 37°C. The cells were grown to exponential phase (optical density at 600 nm ∼0.6) in LB medium. Total RNAs were prepared and used to measure the YcgZ mRNA levels by RT-qPCR. Numbers represent the means and standard deviations of expression levels from three independent experiments performed in duplicate. AG100, wt; AGEZ3, bluR; M113REZ3, lon bluR. See Supplementary Tables S4, S5 for details.
YcgZ Represses ompF Transcription
We then investigated whether the ZABC operon, which is de-repressed at 25°C, was involved in the control of ompF expression. In our model, if one of the proteins encoded by the ZABC operon accumulates in a lon bluR strain and represses ompF, we expect to see an increased ompF expression when the ZABC operon is deleted, i.e., in strain lon bluR ZABC. Using RT-qPCR, we compared OmpF mRNA levels in wild type, lon bluR and lon bluR ZABC strains grown at 37 and 25°C. When grown at 37°C, the three strains show similar OmpF mRNA levels (Figure 5A). Under low temperature growth conditions, the lon bluR ZABC strain expressed OmpF mRNA level similar to that of the wild type strain (Figure 5A). We further confirmed that the ZABC operon was involved in the control of ompF transcription using our PompF-lacZ reporter fusion; Figure 5B shows that deletion of the ZABC operon in a lon bluR background restores LacZ activity similar to that of the wild type. Taken together, our results suggest that the ZABC operon encodes a protein able to repress ompF expression. To further identify this repressor, we used a low copy pBAD vector allowing the expression of a target gene from the L-arabinose-dependent promoter araBAD. We independently expressed ycgZ, ymgA, ymgB, and ymgC in the lon bluR ZABC strain and subsequently measured the LacZ activity in the transformants grown at 25°C in the presence of L-arabinose. Our results show that the activity of PompF-lacZ decreases in response to increasing amount of YcgZ, while YmgA, YmgB, and YmgC have no significant effect on PompF-lacZ activity (Table 2 and see also Supplementary Figure S2). While our results identifies YcgZ as a repressor of the ompF promoter and our data show that ycgZ expression is high in a bluR mutant (Figure 4), a bluR mutant does not decrease ompF transcription (Figures 2, 3). However, a lon bluR strain grown at 25°C expresses less ompF transcript than the bluR mutant. These results strongly suggest that YcgZ is unstable in the presence of Lon under the growth conditions used in our experiments. That a lon bluR strain grown at 37°C does not decrease transcription of ompF is probably due to growth conditions that inherently lead to a high repression of ompF expression. In this case, YcgZ amount may be too low to further repress the ompF promoter.
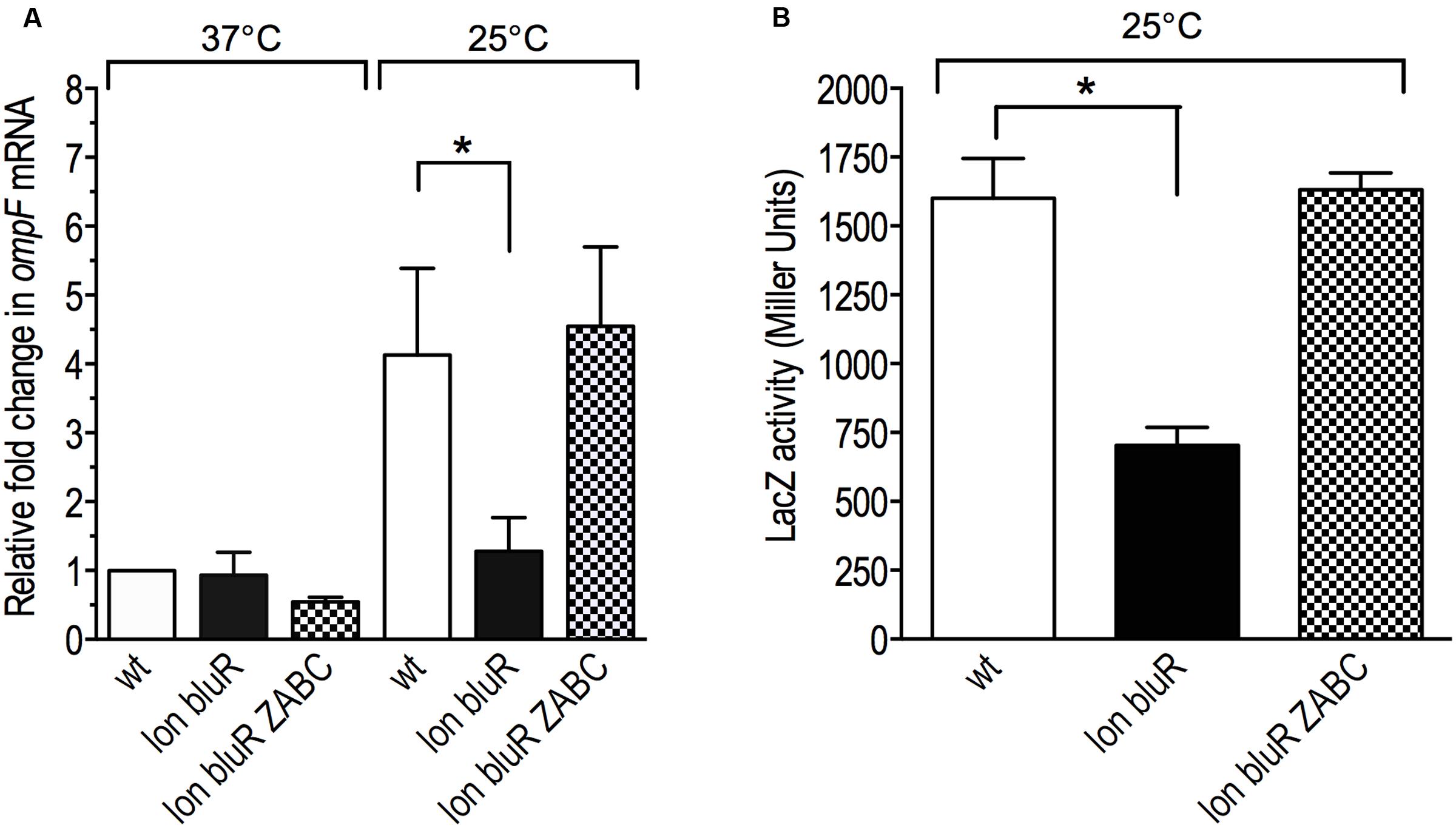
FIGURE 5. lon and bluR mutations decrease ompF transcription via the ycgZ-ymgABC operon. The cells were grown to exponential phase (optical density at 600 nm ∼0.6) in LB medium at 25 and 37°C. Total RNAs were prepared and used to measure the OmpF mRNA levels by RT-qPCR. (A) Fold change in OmpF mRNA levels relative to the wild type strain grown at 37°C. (B) The β-galactosidase activity (Miller units) of a single chromosomal copy of PompF-lacZ was measured for cells grown to exponential phase in LB medium at 25°C. wt, VDL25113; lon bluR, VDL102; lon bluR ycgZ-ymgABC (VDL104). The means and standard deviations of at least three independent experiments are shown. Statistically significant differences for a mutant compared to wild type are shown as asterisk (∗P < 0.05). See Supplementary Table S6, S7 for details.
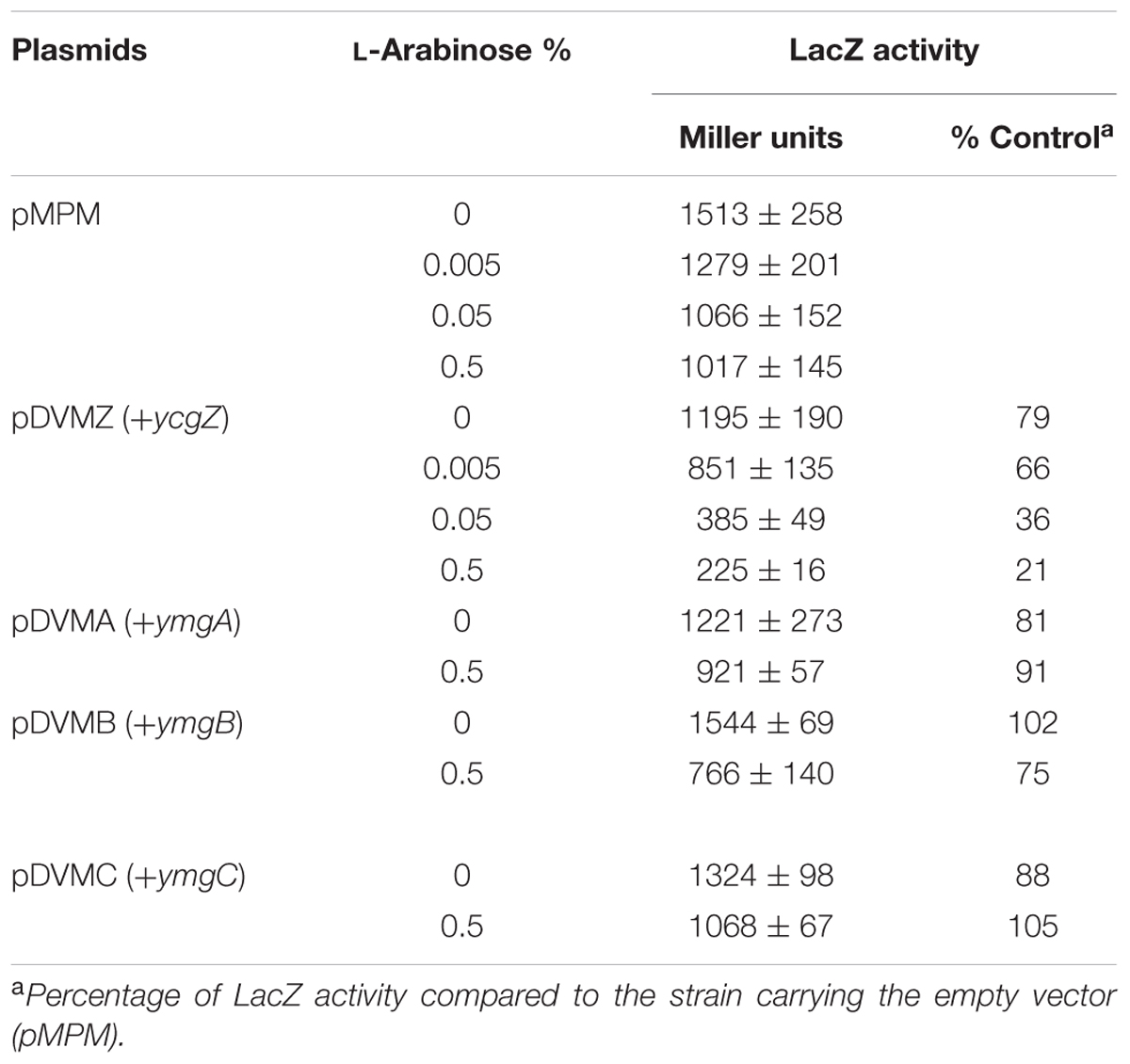
TABLE 2. Expression of PompF-lacZ in E. coli VDL104 (lon bluR ycgZ-ymgABC) grown to exponential phase in LB medium at 25°C.
YcgZ is a Substrate of the Lon Protease
To evaluate whether YcgZ was a substrate of the Lon protease, we compared YcgZ protein amount in E. coli wild type and lon mutant. For completion purpose, we also evaluated the stability of YmgA, YmgB, and YmgC. Lacking antibodies that can specifically interact with the proteins, we decided to evaluate the steady-state level of each native protein when overexpressed from an araBAD promoter using pBAD derivative plasmids (see Table 1). Protein expression was compared between BW25113 (wild type) and JW0419-1 (lon) carrying the plasmids and grown in the presence of L-arabinose. Our experiments indicated that the amount of YcgZ was significantly lower when expressed in the wild type strain, while a lon mutant accumulated a large amount of YcgZ (Figure 6A). Of note, we observed this phenomenon when the cells were grown at both 25 and 37°C. We verified that the lower amount of YcgZ in the wild type was not due to a lesser expression of ycgZ by quantifying ycgZ messenger levels. RT-qPCR experiments showed that ycgZ was similarly expressed in both strains (see Supplementary Table S8), suggesting that the higher level of YcgZ protein observed in the lon mutant is likely due to a higher stability of the protein in the absence of Lon. Our data illustrated in Figure 6A also shows similar amounts of YmgA and YmgC proteins when overexpressed in the wild type and in the lon strains. We could not detect significant amount of YmgB in any of the strains tested even though high level of ymgB messenger was measured by RT-qPCR in both strains (data not shown). Using an XPress-tagged YcgZ (XPYcgZ), we confirmed an L-arabinose-dependent accumulation of YcgZ in the lon strains, while the protein was barely detectable in the wild type (Figure 6B). Additionally, we examined the stability of native YcgZ in the wild type and lon strains after stopping the protein synthesis with chloramphenicol and determining the amount of YcgZ in multiple samples taken over a certain period of time (Figure 6C). We observed a decline in YcgZ quantity in the wild type strain, while a high amount of the protein was maintained over time in the lon mutant, demonstrating a significant stability of YcgZ in the absence of Lon.
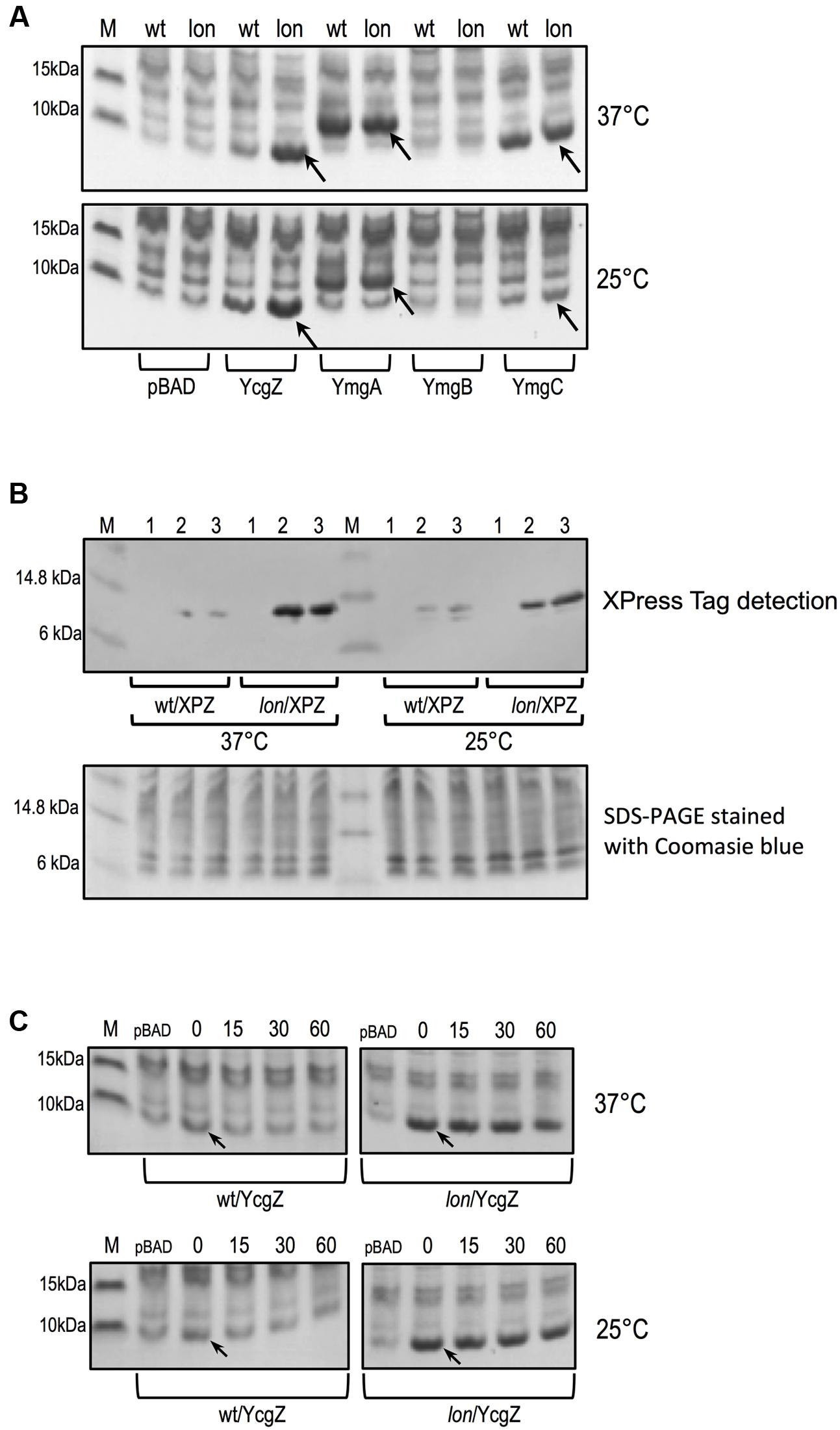
FIGURE 6. Protein levels of YcgZ, YmgA, YmgB, and YmgC in E. coli wild type (BW25113) and lon mutant (JW0419-1). (A) Steady-state level of native YcgZ, YmgA, YmgB, and YmgC expressed with plasmids pBAD (empty plasmid), pDVBZ (YcgZ), pDVBA (YmgA), pDVBB (YmgB), and pDVBC (YmgC), respectively and 0.05% of L-arabinose. (B) Steady-state level of XPress-tagged YcgZ expressed with pDVBZ-XP plasmid and detected by immunoblotting. 1, no L-arabinose; 2, 0.005% L-arabinose; 3, 0.05% L-arabinose. (C) Stability of native YcgZ expressed with plasmid pDVBZ. The numbers indicate the time in minutes after addition of 150 μg ml-1 chloramphenicol. For all experiments, the target gene was expressed from an araBAD promoter using pBAD derivative plasmids. The strains carrying plasmids were grown to an optical density of 1 in the presence of L-arabinose at 37 and 25°C, respectively. Whole cell extracts were then prepared and analyzed by SDS-PAGE and the gels were stained using Coomassie (A,C). The arrow indicates the band corresponding to the expressed protein. Each lane contains 8 μg of proteins. M, Benchmark Protein Ladder.
We further aimed to confirm the lon-dependent repression of ompF promoter by YcgZ. YcgZ was expressed in the ZABC and lon bluR ZABC mutant strains, both carrying a chromosomal PompF-lacZ fusion and providing both lon+/lon- backgrounds. In the absence of lon, expression of YcgZ significantly decreased LacZ activity with addition of 0.0025% of L-arabinose (Figure 7). In a lon+ background, similar repression of ompF promoter was reached by increasing the L-arabinose concentration by at least threefold (0.0075%). Ultimately, when a higher concentration of L-arabinose was used (>0.01%), both strains expressed a similar level of LacZ indicating that under those conditions, an abundant level of YcgZ is sufficient to repress ompF promoter even in the presence of the Lon protease (Figure 7). Of note, the level of ycgZ transcript measured when ycgZ is expressed from its own promoter is much lower than that expressed from the araBAD promoter with concentration of L-arabinose above 0.01% (compared Supplementary Tables S4, S5 with Table S8).
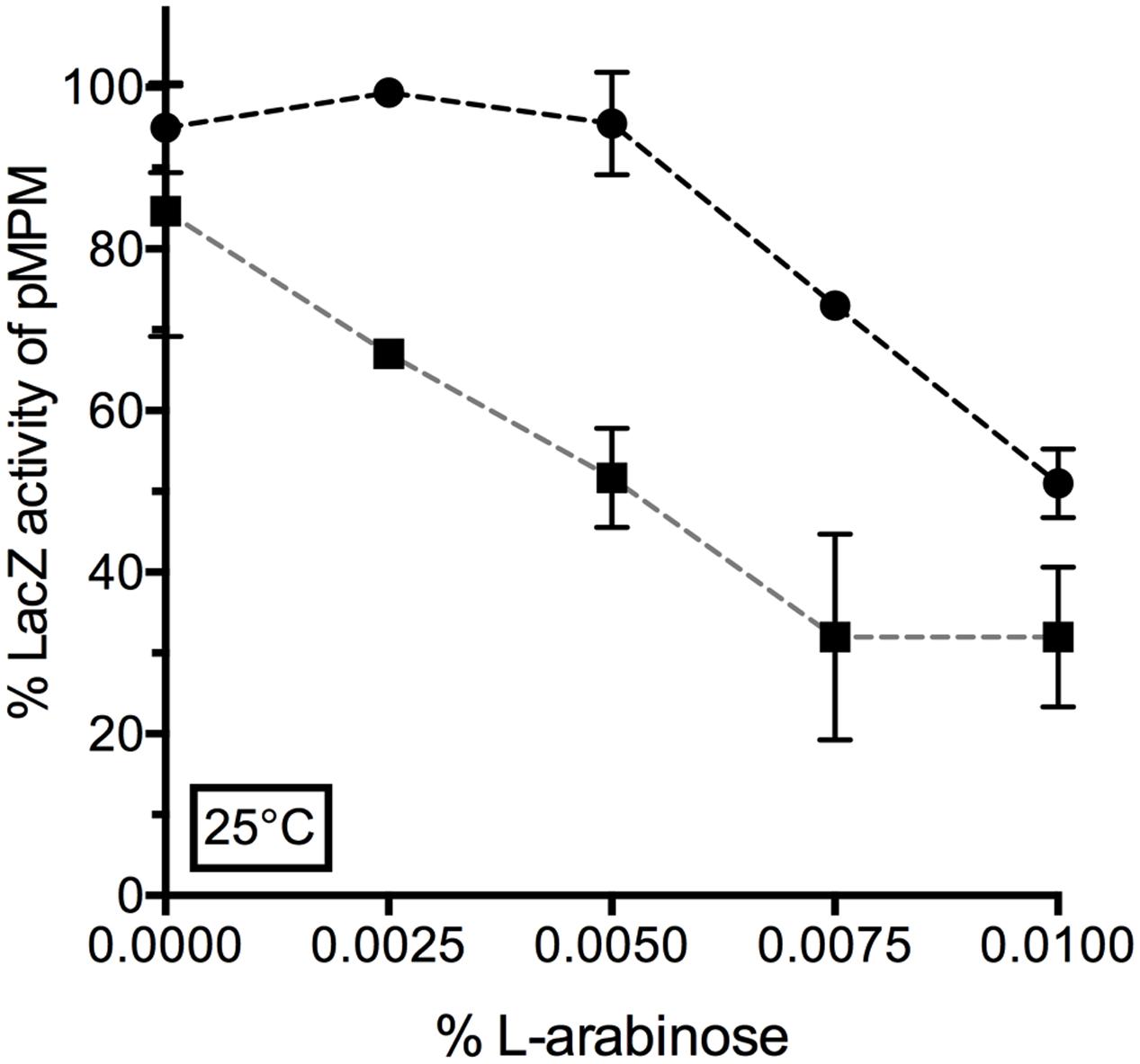
FIGURE 7. Lon-dependent repression of PompF-lacZ by YcgZ. YcgZ was expressed using the pDVMZ vector in VDL103 (ycgZ-ymgABC ●) and VDL104 (lon bluR ycgZ-ymgABC ■) strains. Cells were grown at 25°C in LB medium supplemented with increasing concentration of L-arabinose. LacZ activity was measured when the culture reached exponential phase (optical density at 600 nm ∼0.6) and is expressed as a percentage of the LacZ activity measured for the strain carrying the empty pMPM plasmid. The values represent the mean and the standard deviations of a least three independent experiments.
Discussion
OmpF regulation in response to variable growth conditions provides a great example of bacterial adaptive response to the environment. In this work, we characterize a lon bluR mutant of E. coli and we show that transcription of ompF is impaired when the cells are grown at 25°C. Our data reveal that both bluR and lon bluR strains expressed high levels of ycgZ transcript, confirming a transcriptional repression by BluR on the ycgZ promoter. We find that the repression of the ycgZ promoter by BluR is strong when the cells are grown at 37°C, while growth at 25°C led to a weaker repression. We also show a high induction of ycgZ expression at 25°C, which is essentially mediated by bluR. It was previously established that the BluR–DNA interaction is released in the presence of BluF, a direct antagonist of BluR whose activity is induced by low temperature. Inactivation of BluR by BluF consequently led to transcription of the ycgZ-ymgABC operon (Tschowri et al., 2009). This effect is further increased upon exposure of BluF to blue-light irradiation in addition to the temperature downshift as blue-light activates BluF through conformational changes (Hasegawa et al., 2006; Nakasone et al., 2010).
While we do not describe the specific mechanism by which YcgZ regulates OmpF expression, our study clearly establishes that expression of YcgZ represses ompF transcription. Moreover, our analyses demonstrate a high instability of YcgZ in wild type E. coli, while the protein substantially accumulates and remains stable in a lon mutant, identifying YcgZ as a substrate of the Lon protease. The high instability of YcgZ in the presence of Lon explains why, even though ycgZ expression is large in a bluR mutant at 25 and 37°C, ompF transcription is not affected. These growth conditions are detrimental to the stability of YcgZ with the activity of Lon being high enough to keep the concentration of YcgZ below the threshold where it can inhibit the ompF promoter. In other words, when expressed from its own promoter and under the growth conditions used in our experiments, YcgZ will likely have an effect on the ompF promoter only if its stability is increased. Growth at temperatures below 25°C could reduce the proteolytic activity of Lon, leading to a higher level of YcgZ in the cell. For instance, Yersinia pestis YmoA is a substrate of the Lon protease. Jackson et al. (2004) found that YmoA stability increases as the growth temperature decreases to become stable at 17°C (half-life >3 h). Alternatively, the interaction of YcgZ with another yet to be discovered protein could protect YcgZ from degradation by Lon. An example of such phenomenon in E. coli is illustrated with HU-α and HU-β proteins, two homologous proteins encoded by hupA and hupB, respectively and which form heterodimers. In the presence of HU-α, HU-β is fairly stable, while in a hupA mutant HU-β is degraded by Lon (Bonnefoy et al., 1989); HU-α seems to protect HU-β from degradation by Lon.
How YcgZ, a protein of 78 amino acid residues, acts on ompF promoter is still under investigation. YmgB, a small three-helix protein of 88 amino acids encoded by the ZABC operon, was shown to display similarity to protein Hha (Lee et al., 2007) and to downregulate curli expression in an RcsB-dependent pathway (Tschowri et al., 2009). It was then proposed that YmgB acts as “connector” of the Rcs phosphorelay. YcgZ could likely work as a small protein connector as well. Interestingly, YmgA and YmgB proteins have been shown to activate colanic acid expression under ambient temperature conditions (16–28°C), while decreasing curli synthesis (Tschowri et al., 2009). The only known role of YcgZ was to somehow alleviate the activity of both YmgA and YmgB.
Conclusion
Our study identifies OmpF promoter as a new target regulated by YcgZ, a small pleiotropic regulator, which expression is induced by low temperatures via BluR and is destabilized by the Lon protease. We believe our results shed a new light on novel signals able to regulate OmpF porin expression through complex regulatory pathways (see proposed model in Figure 8). The possibility of a more complex regulatory architecture involving other factors not considered in this study is under investigation.
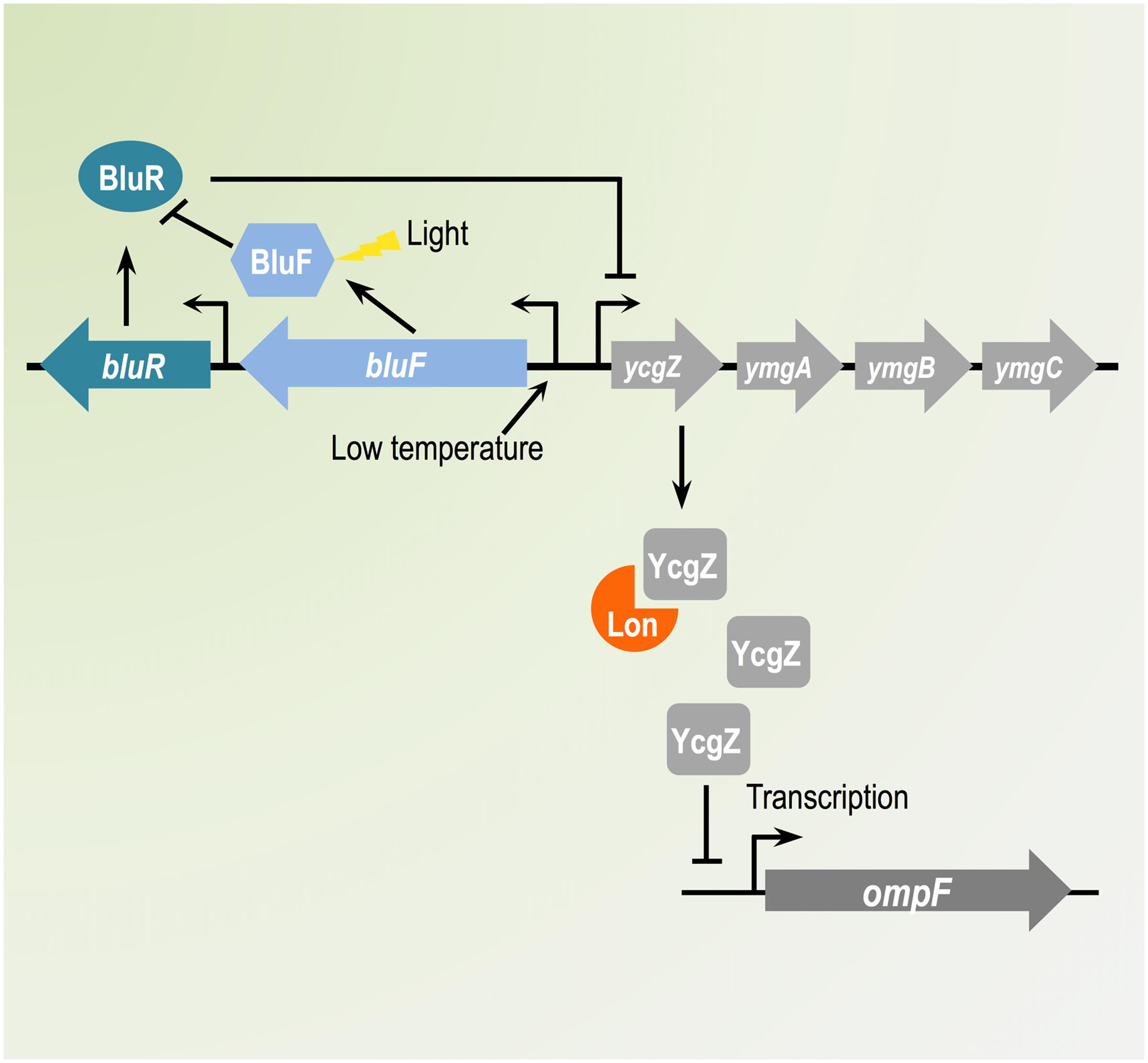
FIGURE 8. Schematic representation of the regulatory cascade employed by BluR and Lon to control OmpF expression. In Escherichia coli, OmpF expression responds to environmental changes via complex regulatory networks. Our report described a novel pathway controlling OmpF synthesis at low temperature. The ycgZ-ymgABC operon, which transcription is controlled by the BluR-BluF system, encodes YcgZ, a protein highly unstable in the presence of the Lon protease. Increased amount of YcgZ leads to reduced transcription of ompF, consequently decreasing the amount of OmpF porin.
Author Contributions
VD and SL conceived the study. VD, KF, and JB performed the experiments. VD analyzed data. VD and SL prepared the manuscript and all the authors contributed to preparing the final version of the manuscript. All authors read and approved the final version of the manuscript.
Funding
The research reported in this article was supported by the National Institute of Allergy and Infectious Diseases of the National Institutes of Health under award number R01 AI056021. The content of this article is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank members of the Sonenshein and Belitsky’s laboratories for helpful suggestions and discussions during the course of this study.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2017.01148/full#supplementary-material
Footnotes
References
Achouak, W., Heulin, T., and Pagès, J. M. (2001). Multiple facets of bacterial porins. FEMS Microbiol. Lett. 199, 1–7. doi: 10.1111/j.1574-6968.2001.tb10642.x
Andersen, J., and Delihas, N. (1990). micF RNA binds to the 5’ end of ompF mRNA and to a protein from Escherichia coli. Biochemistry 29, 9249–9256. doi: 10.1021/bi00491a020
Aranda, P. S., LaJoie, D. M., and Jorcyk, C. L. (2012). Bleach gel: a simple agarose gel for analyzing RNA quality. Electrophoresis 33, 366–369. doi: 10.1002/elps.201100335
Baba, T., Ara, T., Hasegawa, M., Takai, Y., Okumura, Y., Baba, M., et al. (2006). Construction of Escherichia coli K-12 in-frame, single-gene knockout mutants: the Keio collection. Mol. Syst. Biol. 2, 2006.0008. doi: 10.1038/msb4100050
Basle, A. M., Qutub, R., Mehrazin, M., Wibbenmeyer, J. A., and Delcour, A. H. (2003). Functional studies on loop deletion mutants of E. coli OmpF porin. Biophys. J. 84, 533A–533A.
Bonnefoy, E., Almeida, A., and Rouvière-Yaniv, J. (1989). Lon-dependent regulation of the DNA binding protein HU in Escherichia coli. Proc. Natl. Acad. Sci. U.S.A. 86, 7691–7695.
Chen, S., Zhang, A., Blyn, L. B., and Storz, G. (2004). MicC, a second small-RNA regulator of Omp protein expression in Escherichia coli. J. Bacteriol. 186, 6689–6697. doi: 10.1128/JB.186.20.6689-6697.2004
Chopra, I., and Eccles, S. J. (1978). Diffusion of tetracycline across the outer membrane of Escherichia coli K-12: involvement of protein Ia. Biochem. Biophys. Res. Commun. 83, 550–557. doi: 10.1016/0006-291X(78)91025-2
Cohen, S. P., McMurry, L. M., Hooper, D. C., Wolfson, J. S., and Levy, S. B. (1989). Cross-resistance to fluoroquinolones in multiple-antibiotic-resistant (Mar) Escherichia coli selected by tetracycline or chloramphenicol: decreased drug accumulation associated with membrane changes in addition to OmpF reduction. Antimicrob. Agents Chemother. 33, 1318–1325. doi: 10.1128/AAC.33.8.1318
Cowan, S. W., Garavito, R. M., Jansonius, J. N., Jenkins, J. A., Karlsson, R., Konig, N., et al. (1995). The structure of Ompf porin in a tetragonal crystal form. Structure 3, 1041–1050. doi: 10.1016/S0969-2126(01)00240-4
Datsenko, K. A., and Wanner, B. L. (2000). One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. U.S.A. 97, 6640–6645. doi: 10.1073/pnas.120163297
Duval, V., Nicoloff, H., and Levy, S. B. (2009). Combined inactivation of lon and ycgE decreases multidrug susceptibility by reducing the amount of OmpF porin in Escherichia coli. Antimicrob. Agents Chemother. 53, 4944–4948. doi: 10.1128/AAC.00787-09
Egger, L. A., and Inouye, M. (1997). Purification and characterization of the periplasmic domain of EnvZ osmosensor in Escherichia coli. Biochem. Biophys. Res. Commun. 231, 68–72. doi: 10.1006/bbrc.1996.6007
Forst, S., and Inouye, M. (1988). Environmentally regulated gene expression for membrane proteins in Escherichia coli. Annu. Rev. Cell Biol. 4, 21–42. doi: 10.1146/annurev.cb.04.110188.000321
Galdiero, S., Falanga, A., Cantisani, M., Tarallo, R., Della Pepa, M. E., D’Oriano, V., et al. (2012). Microbe-host interactions: structure and role of Gram-negative bacterial porins. Curr. Protein Pept. Sci. 13, 843–854. doi: 10.2174/138920312804871120
Gogol, E. B., Rhodius, V. A., Papenfort, K., Vogel, J., and Gross, C. A. (2011). Small RNAs endow a transcriptional activator with essential repressor functions for single-tier control of a global stress regulon. Proc. Natl. Acad. Sci. U.S.A. 108, 12875–12880. doi: 10.1073/pnas.1109379108
Griffith, K. L., Shah, I. M., and Wolf, R. E. (2004). Proteolytic degradation of Escherichia coli transcription activators SoxS and MarA as the mechanism for reversing the induction of the superoxide (SoxRS) and multiple antibiotic resistance (Mar) regulons. Mol. Microbiol. 51, 1801–1816. doi: 10.1046/j.1365-2958.2003.03952.x
Hasegawa, K., Masuda, S., and Ono, T.-A. (2006). Light induced structural changes of a full-length protein and its BLUF domain in YcgF(Blrp), a blue-light sensing protein that uses FAD (BLUF). Biochemistry 45, 3785–3793. doi: 10.1021/bi051820x
Igo, M. M., Ninfa, A. J., and Silhavy, T. J. (1989). A bacterial environmental sensor that functions as a protein kinase and stimulates transcriptional activation. Genes Dev. 3, 598–605. doi: 10.1101/gad.3.5.598
Jackson, M. W., Silva-Herzog, E., and Plano, G. V. (2004). The ATP-dependent ClpXP and Lon proteases regulate expression of the Yersinia pestis type III secretion system via regulated proteolysis of YmoA, a small histone-like protein. Mol. Microbiol. 54, 1364–1378. doi: 10.1111/j.1365-2958.2004.04353.x
Lee, J., Page, R., García-Contreras, R., Palermino, J.-M., Zhang, X.-S., Doshi, O., et al. (2007). Structure and function of the Escherichia coli protein YmgB: a protein critical for biofilm formation and acid-resistance. J. Mol. Biol. 373, 11–26. doi: 10.1016/j.jmb.2007.07.037
Mayer, M. P. (1995). A new set of useful cloning and expression vectors derived from pBlueScript. Gene 163, 41–46. doi: 10.1016/0378-1119(95)00389-N
Miller, J. (1972). Experiments in Molecular Genetics. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Mizuno, T., Chou, M. Y., and Inouye, M. (1984). A unique mechanism regulating gene expression: translational inhibition by a complementary RNA transcript (micRNA). Proc. Natl. Acad. Sci. U.S.A. 81, 1966–1970. doi: 10.1073/pnas.81.7.1966
Mizuno, T., and Mizushima, S. (1990). Signal transduction and gene regulation through the phosphorylation of two regulatory components: the molecular basis for the osmotic regulation of the porin genes. Mol. Microbiol. 4, 1077–1082. doi: 10.1111/j.1365-2958.1990.tb00681.x
Mizusawa, S., and Gottesman, S. (1983). Protein degradation in Escherichia coli: the lon gene controls the stability of SulA protein. Proc. Natl. Acad. Sci. U.S.A. 80, 358–362. doi: 10.1073/pnas.80.2.358
Mortimer, P. G., and Piddock, L. J. (1993). The accumulation of five antibacterial agents in porin-deficient mutants of Escherichia coli. J. Antimicrob. Chemother. 32, 195–213. doi: 10.1093/jac/32.2.195
Nakasone, Y., Ono, T.-A., Ishii, A., Masuda, S., and Terazima, M. (2010). Temperature-sensitive reaction of a photosensor protein YcgF: possibility of a role of temperature sensor. Biochemistry 49, 2288–2296. doi: 10.1021/bi902121z
Nikaido, H. (1994). Porins and specific diffusion channels in bacterial outer membranes. J. Biol. Chem. 269, 3905–3908. doi: 10.1111/j.1365-2958.1992.tb01487.x
Nikaido, H. (2003). Molecular basis of bacterial outer membrane permeability revisited. Microbiol. Mol. Biol. Rev. 67, 593–656. doi: 10.1128/MMBR.67.4.593-656.2003
Oethinger, M., Kern, W. V., Goldman, J. D., and Levy, S. B. (1998). Association of organic solvent tolerance and fluoroquinolone resistance in clinical isolates of Escherichia coli. J. Antimicrob. Chemother. 41, 111–114. doi: 10.1093/jac/41.1.111
Pagès, J.-M., James, C. E., and Winterhalter, M. (2008). The porin and the permeating antibiotic: a selective diffusion barrier in Gram-negative bacteria. Nat. Rev. Microbiol. 6, 893–903. doi: 10.1038/nrmicro1994
Pfaffl, M. W. (2001). A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 29:e45. doi: 10.1093/nar/29.9.e45
Powell, B. S., Rivas, M. P., Court, D. L., Nakamura, Y., and Turnbough, C. L. (1994). Rapid confirmation of single copy lambda prophage integration by PCR. Nucleic Acids Res. 22, 5765–5766. doi: 10.1093/nar/22.25.5765
Pratt, L. A., Hsing, W., Gibson, K. E., and Silhavy, T. J. (1996). From acids to osmZ: multiple factors influence synthesis of the OmpF and OmpC porins in Escherichia coli. Mol. Microbiol. 20, 911–917. doi: 10.1111/j.1365-2958.1996.tb02532.x
Silhavy, T. J., Berman, M. L., Enquist, L. W., and Laboratory, C. S. H. (1984). Experiments with Gene Fusions. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Simons, R. W., Houman, F., and Kleckner, N. (1987). Improved single and multicopy lac-based cloning vectors for protein and operon fusions. Gene 53, 85–96. doi: 10.1016/0378-1119(87)90095-3
Sugawara, E., and Nikaido, H. (1994). OmpA protein of Escherichia coli outer membrane occurs in open and closed channel forms. J. Biol. Chem. 269, 17981–17987.
Torres-Cabassa, A. S., and Gottesman, S. (1987). Capsule synthesis in Escherichia coli K-12 is regulated by proteolysis. J. Bacteriol. 169, 981–989. doi: 10.1128/jb.169.3.981-989.1987
Tschowri, N., Busse, S., and Hengge, R. (2009). The BLUF-EAL protein YcgF acts as a direct anti-repressor in a blue-light response of Escherichia coli. Genes Dev. 23, 522–534. doi: 10.1101/gad.499409
Tschowri, N., Lindenberg, S., and Hengge, R. (2012). Molecular function and potential evolution of the biofilm-modulating blue light-signalling pathway of Escherichia coli. Mol. Microbiol. 85, 893–906. doi: 10.1111/j.1365-2958.2012.08147.x
Tsilibaris, V., Maenhaut-Michel, G., and Van Melderen, L. (2006). Biological roles of the Lon ATP-dependent protease. Res. Microbiol. 157, 701–713. doi: 10.1016/j.resmic.2006.05.004
Van Melderen, L., and Aertsen, A. (2009). Regulation and quality control by Lon-dependent proteolysis. Res. Microbiol. 160, 645–651. doi: 10.1016/j.resmic.2009.08.021
Vogel, J., and Papenfort, K. (2006). Small non-coding RNAs and the bacterial outer membrane. Curr. Opin. Microbiol. 9, 605–611. doi: 10.1016/j.mib.2006.10.006
Yoshida, T., Qin, L., Egger, L. A., and Inouye, M. (2006). Transcription regulation of ompF and ompC by a single transcription factor. OmpR. J. Biol. Chem. 281, 17114–17123. doi: 10.1074/jbc.M602112200
Keywords: regulation, porin, Lon protease, E. coli, blue light, temperature
Citation: Duval V, Foster K, Brewster J and Levy SB (2017) A Novel Regulatory Cascade Involving BluR, YcgZ, and Lon Controls the Expression of Escherichia coli OmpF Porin. Front. Microbiol. 8:1148. doi: 10.3389/fmicb.2017.01148
Received: 16 February 2017; Accepted: 07 June 2017;
Published: 30 June 2017.
Edited by:
Dongsheng Zhou, Beijing Institute of Microbiology and Epidemiology, ChinaReviewed by:
Boris Görke, University of Vienna, AustriaJohannes Geiselmann, Joseph Fourier University, France
Copyright © 2017 Duval, Foster, Brewster and Levy. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Stuart B. Levy, c3R1YXJ0LmxldnlAdHVmdHMuZWR1 Valérie Duval, dmFsZXJpZS5kdXZhbEB0dWZ0cy5lZHU=