 Baoling Li1
Baoling Li1 Haizhen Wu
Haizhen Wu Lihua Xiao
Lihua Xiao- 1State Key Laboratory of Bioreactor Engineering, School of Resources and Environmental Engineering, East China University of Science and Technology, Shanghai, China
- 2School of Biotechnology, East China University of Science and Technology, Shanghai, China
- 3College of Veterinary Medicine, South China Agricultural University, Guangzhou, China
- 4Emory Vaccine Center, Yerkes National Primate Research Center, Emory University, Atlanta, GA, United States
- 5Division of Foodborne, Waterborne, and Environmental Diseases, National Center for Emerging and Zoonotic Infectious Diseases, Centers for Disease Control and Prevention, Atlanta, GA, United States
Cryptosporidium spp. are important causes of diarrhea in humans, ruminants, and other mammals. Comparative genomic analysis indicated that genetically related and host-adapted Cryptosporidium species have different numbers of subtelomeric genes encoding the Cryptosporidium-specific MEDLE family of secreted proteins, which could contribute to differences in host specificity. In this study, a Cryptosporidium parvum-specific member of the protein family MEDLE-2 encoded by cgd5_4590 was cloned and expressed in Escherichia coli. Immunofluorescent staining with antibodies generated from the recombinant protein showed the expression of the protein in sporozoites and development stages. In vitro neutralization assay with the antibodies partially blocked the invasion of sporozoites. These results support the potential involvement of MEDLE-2 in the invasion of host cells.
Introduction
Cryptosporidium spp. are important pathogens inhabiting the brush borders of the gastrointestinal epithelium of various vertebrates, causing enterocolitis, diarrhea, and vomiting (Checkley et al., 2015). Of the near 100 recognized Cryptosporidium species and genotypes, most can infect only a few closely related hosts (Ryan et al., 2014). For example, C. andersoni and C. bovis infect mainly cattle, C. hominis and C. viatorum infect mostly humans, and C. canis and C. felis infect dogs and cats, respectively. Some of the species, such as C. parvum, C. meleagridis and C. ubiquitum have a broad host range, thus are considered important zoonotic parasites (Ryan et al., 2014). Among them, C. parvum has attracted public health attention because of its high pathogenicity in ruminants and high infectivity to humans.
The biologic determinants for host specificity in Cryptosporidium spp. are not clear. This is largely caused by our poor knowledge of major components involved in the invasion of host cells by sporozoites (Liu et al., 2016). Efforts have been made in the identification of proteins that are potentially involved in host cell invasion by screening cDNA library using immune sera. A few of these proteins identified are considered potentially involved in Cryptosporidium invasion, such as gp900, gp40/15 (gp60), Cpa135, Cp2, P23, and TRAP-C1 (Bouzid et al., 2013). Most of these proteins are O-glycosylated and many of them are mucins. As the invasion process is considered to be conserved among apicomplexan parasites and these proteins are rarely seen in non-Cryptosporidium apicomplexans, other proteins could also be involved in invasion and host specificity in Cryptosporidium spp.
Comparative genomic analysis of C. parvum and C. hominis, which differ from each at the nucleotide level by merely ∼3% but have very different host range, indicates that these two species have different copies of subtelomeric genes encoding two major protein families (Guo et al., 2015). One of the protein families involved is the Cryptosporidium-specific MEDLE family of secreted proteins at the 3′ end of chromosomes 5 and 6 (Guo et al., 2015). C. parvum has six paralogous genes encoding MEDLE proteins, compared with one in C. hominis. As host-adapted C. parvum IIa (infecting mostly cattle) and IId (infecting mostly sheep and goats) subtype families also differ in the number of MEDLE genes (Feng et al., 2017), it was suggested that MEDLE proteins could contribute to differences in host specificity among Cryptosporidium spp.
In this study, we conducted a preliminary biologic study of the C. parvum-specific MEDLE-2 encoded by the cgd5_4590 gene. The aim of this study was to express the protein in Escherichia coli, assess its expression on sporozoites and developmental stages in culture, and evaluate its potential role in host cell invasion.
Materials and Methods
Cryptosporidium parvum Isolate, Host Cells, E. coli Strains and Plasmid Vectors
Cryptosporidium parvum oocysts (IOWA strain) were purchased from Waterborne, Inc. (New Orleans, LA, United States) and stored in antibiotics at 4°C for less than 2 months prior to use. Before experiments, oocysts were treated with 0.5% sodium hypochlorite on ice for 10 min and washed 3 times with sterile PBS. Human ileocecal adenocarcinoma HCT-8 cells were obtained from Chinese Academy of Sciences Shanghai Branch. Cells were cultured in maintenance medium at 37°C in a humidified atmosphere containing 5% CO2. E. coli strains DH5α and BL21 (DE3) (Tiangen, Beijing, China) were used for plasmid amplification and expression, respectively. The pET28a vector was obtained from Novagen, Inc. (Madison, WI, United States). All restriction enzymes were purchased from Thermo Fisher Scientific, Inc. (Waltham, MA, United States).
Construction of Recombinant Plasmid
The cgd5_4590 gene (XM_625307) was amplified by PCR from C. parvum DNA and cloned into the pET28a vector as an NcoI-XhoI insertion using the following primers: forward 5′-AAATCCATGGAAAATGTAACCGATAATT-3′ (including an NcoI restriction site) and reverse 5′-AAATCTCGAGTTCCAAATCATGAAGAATATC-3′ (including an XhoI restriction site). The 50 μl-PCR reaction contained 1 μl of DNA, 0.25 mM primers, 3 mM MgCl2, 200 μM deoxynucleotide triphosphates, 1× GeneAmp PCR buffer (Applied Biosystems, Foster City, CA, United States), and 1.5 U of Taq polymerase (Promega, Madison, WI, United States). The amplification was performed on a GeneAmp 9700 (Applied Biosystems), consisting of an initial denaturation at 94°C for 5 min; 35 cycles at 94°C for 45 s, 52°C for 45 s, and 72°C for 1 min; and a final extension at 72°C for 7 min. The template DNA was extracted from C. parvum oocysts by using the Qiagen DNeasy Blood & Tissue Kit (Qiagen, Hilden, Germany). The target gene is located in the 3′ subtelomeric region of chromosomes 5, encoding the MEDLE-2 protein. The molecular weight of the expected protein is 21.0 kDa, with no predicted signal peptide, transmembrane domain, or glycosylphosphatidylinositol anchor. It has 16 predicted O-linked glycosylation sites, mostly concentrated between amino acids 48–69 and 104–117.
The PCR product was purified using the SanPrep Column PCR Product Purification Kit (Sangon Biotech, Shanghai, China), digested with NcoI and XhoI and ligated into the expression vector pET28a. The recombinant vector cgd5_4590-pET28a was introduced into competent E. coli DH5α and transformed cells were selected on LB ager with 50 μg/ml of kanamycin. Positive colonies were randomly picked and identified by PCR. The PCR products from these colonies were sequenced to verify their identity and sequence accuracy.
Expression of Recombinant MEDLE-2 Protein
The recombinant plasmid was extracted from E. coli DH5α using the EZ-10 Spin Column Plasmid DNA Minipreps Kit (Sangon Biotech) and introduced into E. coli BL21 (DE3) to express the target protein. The E. coli BL21 (DE3) cells harboring the recombinant plasmid were cultured at 37°C until OD600 reaching 0.6–1.0, by which time 0.1 mM isopropyl b-D-1-thiogalactopyranoside (IPTG) was added to induce protein expression. The induction was conducted at 18, 25, and 37°C for 4 h to select the optimal expression condition. The expression level and solubility of the target protein were compared among induction temperatures by SDS–PAGE and Western blot analyses of the bacterial cells harvested at OD600 = 1.0.
SDS–PAGE and Western Blot Analyses
The bacterial cells were harvested by centrifugation and lysed by boiling in 5× protein loading buffer for 5 min. Proteins in 20 μL lysate were separated by 10% SDS–PAGE and stained with Coomassie Brilliant Blue (Bio-Rad, Hercules, CA, United States).
For Western blot analysis of the recombinant MEDLE-2, proteins resolved by 10% SDS–PAGE were transferred onto a polyvinylidene fluoride (PVDF) membrane using a semi-dry electro-blotting apparatus (Bio-Rad). The transfer was carried out at 400 mA for 1 h. After blocking with TBST containing 5% non-fat milk at room temperature for 2 h, the membrane was incubated overnight with 1:1,000 anti-His tag antibodies (Cell Signaling Technology, Danvers, MA, United States). Afterwards, the PVDF membrane was washed three times with TBST and incubated at room temperature with 1:5,000 goat anti-mouse IgG-A (HRP) antibodies (Yeasen, Shanghai, China) for 2 h. The membrane was finally washed three times with TBST and reactive protein bands in the membrane were detected using the DAB kit (Tiangen Biotech).
For Western blot analysis of native proteins of C. parvum sporozoites, 1 × 108 oocysts were in PBS buffer and Protease Inhibitor Cocktail Set III (Calbiochem, La Jolla, CA, United States) were sonicated on ice for 30 min. The lysis was collected by centrifugation at 6,000 g for 30 min. The soluble native proteins and recombinant protein were mixed with 5× protein loading buffer, heated at 100°C for 5 min, and subjected to Western blot analysis using pre-immune serum, immune serum, and purified polyclonal antibodies to recombinant MEDLE-2.
Purification of MEDEL-2 and Preparation of Polyclonal Antibodies
For the purification of MEDEL-2, E. coli cells were inoculated into 2-L culture with induction of protein expression by 1 mM IPTG at 25°C for 4 h. The cells were harvested by centrifugation, re-suspended in PBS buffer, and lysed by sonication on ice. The soluble fraction of the E. coli lysate was collected by centrifugation at 6,000 g for 20 min and filtered through a 0.45 μm cellulose acetate membrane filter (Millipore, Billerica, MA, United States). The filtrate was loaded onto Ni-NTA beads (Novagen, Madison, WI, United States) at 4°C and 100 rpm for 2 h. The beads were washed with 8 volumes of washing buffer containing 20 mM imidazole and MEDEL-2 was eluted off the beads with buffer containing 100, 200, or 250 mM imidazole. The final product was analyzed by SDS–PAGE. The purified MEDLE-2 protein was used in the production of polyclonal antibodies in rabbits by GL Biochem Ltd. (Shanghai, China). For this, 350 μg of MEDLE-2 protein mixed with Freund’s complete adjuvant was used in the immunization of healthy New Zealand white rabbits on Days 1 and 15. Immunized animals were boosted six times with 150 μg of MEDLE-2 protein in the Freund’s incomplete adjuvant every 7 days. Seven days after the last immunization, sera were collected from the immunized animals and the polyclonal IgG antibodies were purified from the immune sera using protein A sepharose affinity chromatography.
Localization of MEDLE-2 Expression on C. parvum Sporozoites and Developmental Stages
Sporozoites on slides and intracellular stages of C. parvum in HCT-8 cell cultures at 24 and 48 h were fixed at room temperature for 30 min with 4% paraformaldehyde. Afterwards, the slides were washed three times with PBS for 3 min. About 250 μL of DB Blocking Buffer (B100-40, Waterborne, Inc.) was added to the slides. After incubation at 37°C for 30 min and three washes with the buffer, the slides were incubated with affinity-purified anti-MEDLE-2 antibodies (0.3 μg/ml) for 1 h, washed three times with PBS, and incubated with 1:100 Alexa Fluor 488-conjugated AffiniPure Goat Anti-Rabbit lgG (Yeasen) for 1 h (Jakobi and Petry, 2006). After three washes with PBS, 4′, 6-diamidino-2-phenylindole (DAPI, Roche, Basel, Switzerland) was used to counterstain the nuclei of sporozoites and developmental stages. The slides were mounted with No-Fade Mounting Medium (Boster, Wuhan, China) and examined under a BX53 immunofluorescence microscope (Olympus, Tokyo, Japan).
Examination of CGD5_4590 Gene Expression by QPCR
The expression of the cgd5_4590 gene in parasites developing in HCT-8 cells at 0–72 h was evaluated by qPCR (Mauzy et al., 2012). The expression of 18S rRNA gene was determined in parallel for data normalization (Cai et al., 2005). Each 20 μL-qPCR reaction contained 0.1 mM primers, 4 μL of 1:10 dilution of cDNA synthesized from 2 μg RNA using RevertAid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific) and SsoFast EvaGreen Supermix (Bio-Rad). The qPCR was conducted on a Light Cycler 480 (Roche), with an initial denaturation at 95°C for 1 min and 45 cycles of 95°C for 30 s, 58°C for 30 s, and 68°C for 30 s. The amplification was followed by a melt curve analysis through 95°C for 1 min, ramping down to 57°C in 2 degree intervals, and ramping back to 95°C for 30 s. The primers used were 5′-AGGTAGGGGTGGAGGAGGTA-3′ and 5′-ATGGCTCATCAAATGGGTCT-3′ for the cgd5_4590 gene (amplicon size = 185 bp), and 5′-AGGTAGGGGTGGAGGAGGTA-3′ and 5′-ATGGCTCATCAAATGGGTCT-3′ for the 18S rRNA gene (amplicon size = 256 bp) (Wang et al., 2013; Guo et al., 2016). Threshold cycle (CT) values from qPCR were used in computing the relative levels of expression, in which data were first normalized by computing the ΔCT values between cgd5_4590 mRNA (CT [sample]) and 18S rRNA (CT [Cp18S]) for all samples using the formula:
Next, ΔCT values (relative levels of C. parvum 18S rRNA) in each experiment were further normalized by subtracting the ΔCT0 values (ΔCT at 0 h point):
The relative expression of the cgd5_4590 gene was calculated using the 2-ΔΔCT method (Livak and Schmittgen, 2001). Mean values from at least three independent experiments were obtained.
In Vitro Neutralization of Sporozoite Invasion
A neutralization assay was used to assess the potential involvement of MEDLE-2 in host cell invasion by C. parvum sporozoites in a BSL-2 laboratory (Ludington and Ward, 2016). HCT-8 cells were grown in 12-well plates to 80–90% confluence in RPMI 1640 medium supplemented with 10% fetal bovine serum (FBS), 1 mM sodium pyruvate, 50 U/ml penicillin G, 50 U/ml streptomycin, and 0.25 mg/ml amphotericin B (pH 7.4). Oocysts were treated with 0.5% sodium hypochlorite on ice for 10 min and washed 3 times with cold sterile PBS (Gut and Nelson, 1999). For neutralization assays, 1 × 105 oocysts were incubated with different dilutions of immune serum or pre-immune serum in infection medium in HCT8 cell culture at 37°C for 2 h. Based on results of preliminary evaluations, 1:50, 1:100, and 1:500 dilutions of sera were used in neutralization assays. After the 2-h incubation, the cultures were washed off free sporozoites and incubated for additional 48 h. Cy3-labeled polyclonal antibodies against C. parvum sporozoites (Waterborne, Inc.) were used to determine the infection rate of HCT-8 cells as described (Upton et al., 1995; Deng et al., 2002). Images taken under 200× were digitalized and the relative staining area was quantified by using ImageJ1. The mean parasite load in each culture was calculated based on images from 30 random fields, and compared between treatment groups using the Mann–Whitney U-test. The inhibition rate of sporozoite invasion was calculated based on differences in parasite load between MEDLE-2 antibody treated and control groups.
Results
Cloning of CGD5_4590 Gene and Expression of Recombinant MEDLE-2 Protein
The cgd5_4590 gene was amplified by PCR using specific primers (Figure 1A) and successfully cloned into the pET28a vector using the T4 DNA ligase. After verification of the identity and sequence by DNA sequencing, the recombinant plasmid was introduced into E. coli BL21 (DE3) for MEDLE-2 protein expression. The profile of total proteins from the negative control culture, transformed culture with no induction of MEDLE-2 expression, and transformed culture with induction was analyzed by 12% SDS–PAGE, which showed the presence of two unique protein bands of 21 and 36 kDa size in total proteins from transformed cultures (Figure 1B). This was confirmed using Western blot analysis using anti-His tag antibodies (Figure 1C).
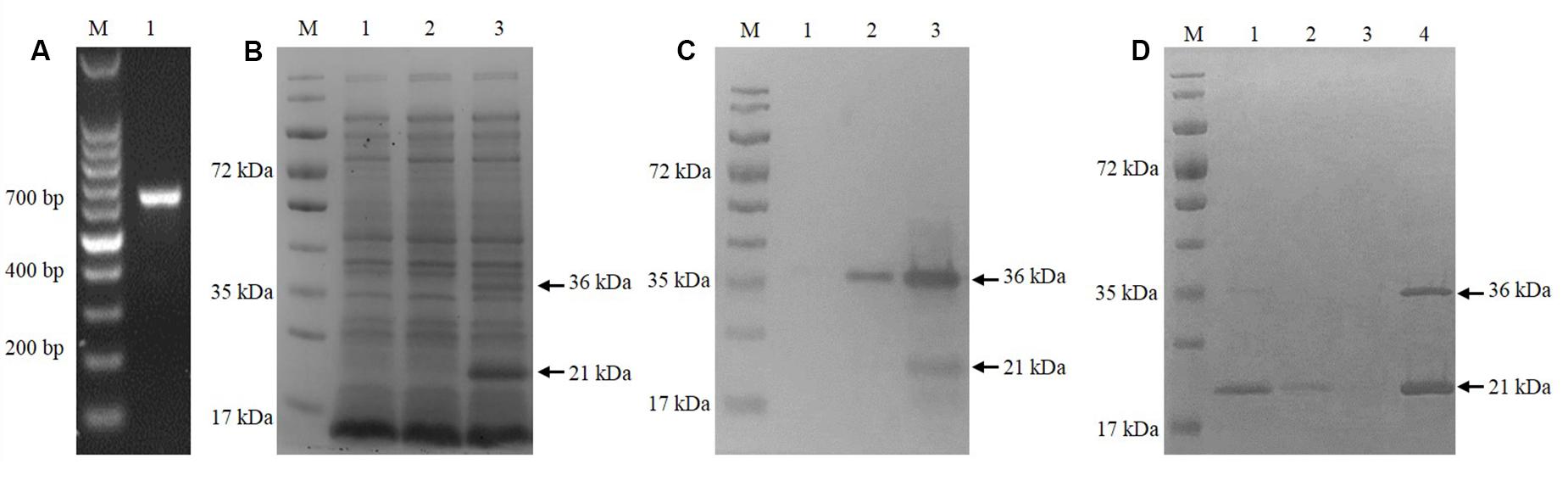
FIGURE 1. Expression of recombinant MEDLE-2 protein in Escherichia coli. (A) PCR amplification of the cgd5_4590 gene in C. parvum. Lane M: 100-bp molecular makers; Lanes 1: cgd5_4590 PCR product. (B) Expression of recombinant MEDLE-2 protein in E. coli BL21 (DE3) revealed by SDS–PAGE analysis. Lane M: protein size makers; Lane 1: lysate from bacterial cells transformed with pET28a vector control; Lane 2: lysate from bacterial cells transformed with pET28a-cgd5_4590 without IPTG induction; Lane 3: lysate from bacterial cells transformed with pET28a-cgd5_4590 with IPTG induction. (C) Expression of recombinant MEDLE-2 protein in E. coli BL21 (DE3) revealed by Western blot analysis using Anti-His tag. Lane M: protein size makers; Lane 1: lysate from bacterial cells transformed with pET28a vector control; Lane 2: lysate from bacterial cells transformed with pET28a-cgd5_4590 without IPTG induction; Lane 3: lysate from bacterial cells transformed with pET28a-cgd5_4590 with IPTG induction. (D) Purity of recombinant MEDLE-2 revealed by SDS–PAGE analysis. Lane M: protein size makers; Lane 1: the first volume of washing solution with 20 mM imidazole from the purification column; Lane 2: the fifth volume of washing solution with 20 mM imidazole from the purification column; Lane 3: the eighth volume of washing solution with 20 mM imidazole from the purification column; Lane 4: target protein eluted from the purification column using 250 mM imidazole.
The Ni-NTA purification method was used to obtain highly pure recombinant MEDLE-2 protein (Figure 1D). The target protein was eluted by using 250 mM imidazole. The MEDLE-2 identity of the purified proteins was established by using the time-of-flight mass spectrometry; both 21 and 36 kDa proteins yielded peptide sequences compatible to the MEDLE-2 sequence (Accession No. XM_625307; data not shown). In western blot analysis, the recombinant MEDLE-2 protein was expectedly recognized by immune serum or purified antibodies from rabbits immunized with MEDLE-2 (Figures 2A,B), but not the pre-immune serum (Figure 2C). The immune serum and purified antibodies also recognized a native protein of approximately 17 kDa from sporozoites.
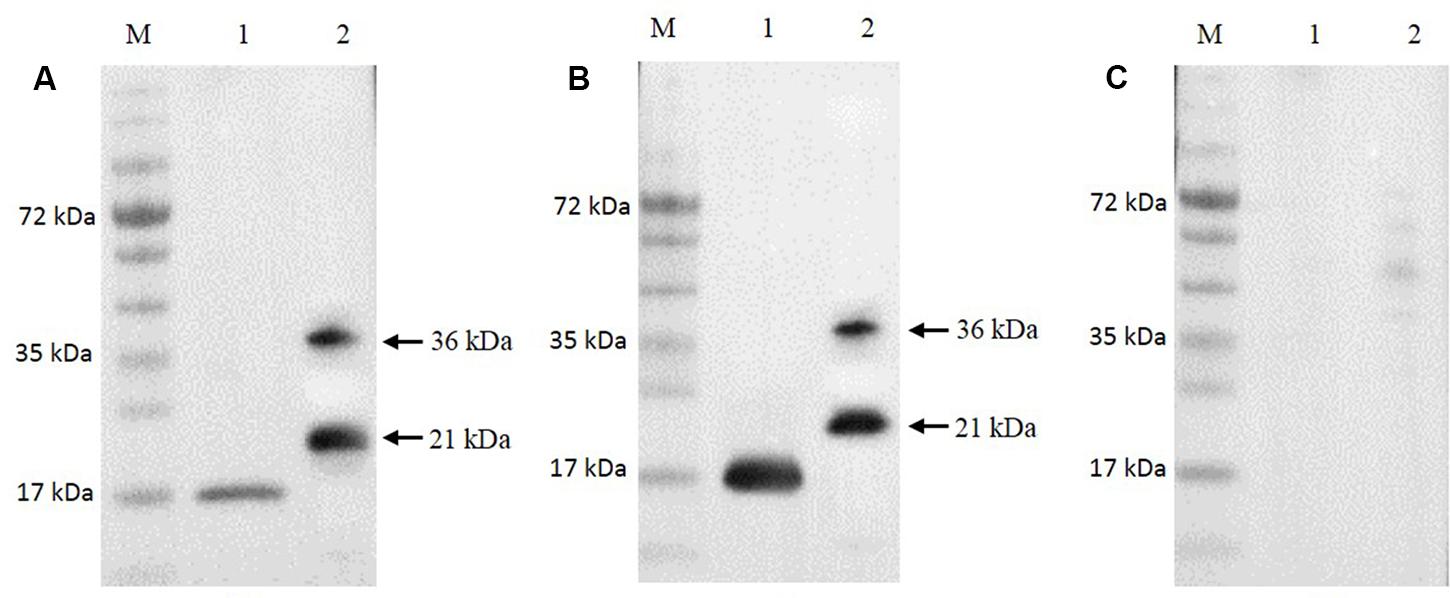
FIGURE 2. Expression of native MEDLE-2 protein in Cryptosporidium parvum sporozoites. Polyclonal IgG antibodies (A), immune serum (B) and pre-immune serum (C) were used in Western blot analysis of total proteins extracted from C. parvum oocysts and recombinant proteins. Lane M: protein size makers; Lane 1: native proteins from C. parvum; Lane 2: recombinant MEDLE-2.
Localization of MEDLE-2 Protein
The expression of MEDLE-2 protein on sporozoites and developmental stages of C. parvum in HCT-8 cell cultures at 24 and 48 h were examined by immunofluorescence. The staining of sporozoites with anti-MEDLE-2 polyclonal antibodies showed a diffused expression of MEDLE-2 within sporozoites, including the anterior end. The highest MEDLE-2 expression was in the area just above the nucleus, which was located at the 1/3 posterior end of sporozoites by DAPI staining (Figure 3, Top). The immunofluorescence staining of developmental stages in 24 and 48 h HCT-8 cultures showed the expression of MEDLE-2 proteins on both the parasitophorus vacuoles of meronts and merozoites within them (Figure 3, Middle, Bottom).
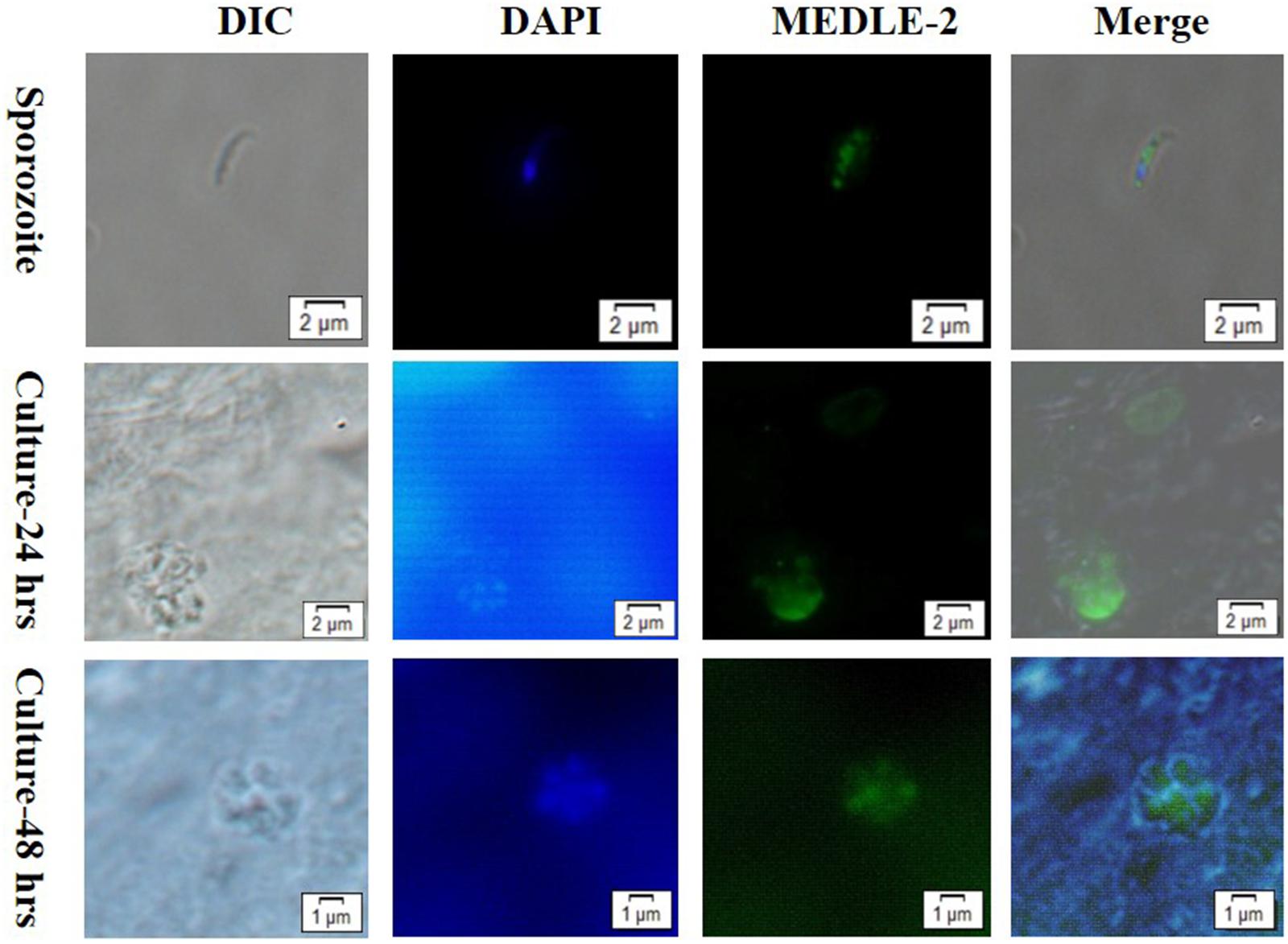
FIGURE 3. Localization of MEDLE-2 protein expression on Cryptosporidium parvum sporozoites (Top) and developmental stages of C. parvum in HCT-8 cell cultures at 24 and 48 h (Middle, Bottom, respectively). Images were taken by differential interference contrast microscopy (DIC), fluorescence microscopy using nuclear stain 4, 6-diamidino-2-phenylindole (DAPI), fluorescence microscopy using polyclonal MEDLE-2 antibodies (MEDLE-2), and superimposition of the three (Merge).
Expression of MEDLE-2 Gene in C. parvum Culture
The expression of the cgd5_4590 gene in C. parvum culture was assessed by qPCR, after data normalization using Ct values from qPCR analysis of the 18S rRNA gene of C. parvum. Newly released sporozoites had minimal expression of the cgd5_4590 gene. After the invasion, the expression of the cgd5_4590 gene increased gradually, with the highest expression level detected at 48 h of culture. A very low level of cgd5_4590 gene expression was seen at 72 h of culture (Figure 4).
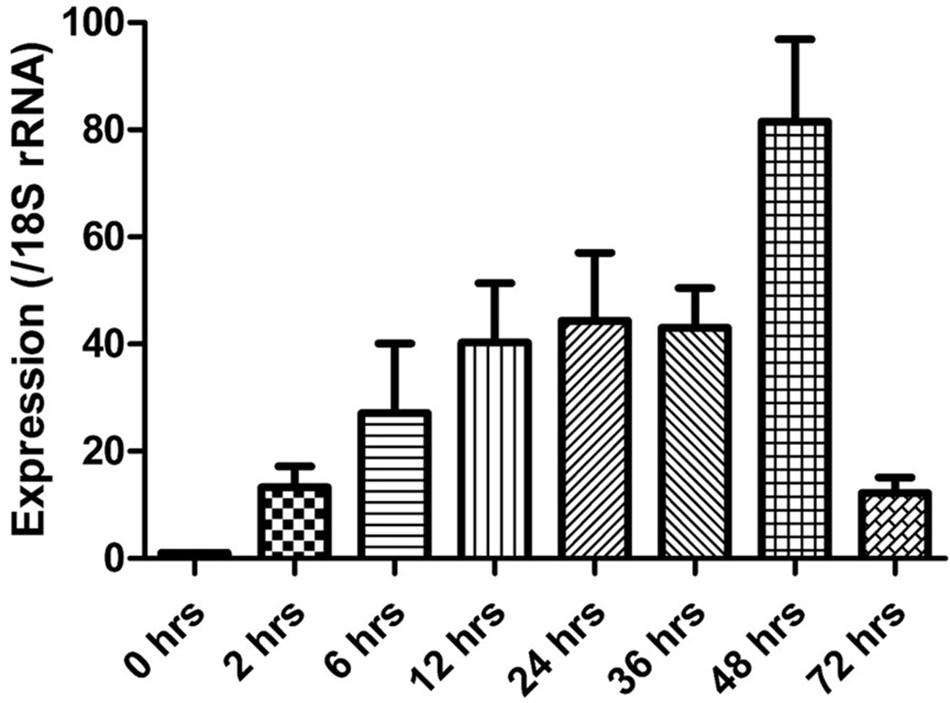
FIGURE 4. Relative expression of the cgd5_4590 gene in HCT-8 cell culture of Cryptosporidium parvum after the normalization of Ct values of qPCR with data from the 18S rRNA gene of C. parvum. Data shown are mean ± SD from three replicate assays.
Neutralization of Sporozoite Invasion by MEDLE-2 Antibodies
We used an invasion neutralization assay to assess the role of MEDLE-2 protein in the infection of intestinal epithelial cells by C. parvum. When HCT-8 cells were infected with sporozoites from 1 × 105 oocysts, comparable parasite loads were obtained 48 h after the invasion between the control cultures and cultures with 1:50, 1:100, and 1:500 dilutions of the pre-immune serum. The mean parasite load in HCT-8 culture without the addition of any serum was 233.6 ± 35.6 per 200 X field. The addition of 1:500 dilution of the immune serum reduced the parasite load in HCT-8 cultures by 33.6% (255.1 ± 57.2 and 169.3 ± 29.2 per 200 X field for the pre-immune serum and immune serum groups, respectively; P = 0.000). At the 1:100 dilution, the inhibition rate of sporozoite invasion by MEDLE-2 immune serum was 41.8% (246.0 ± 46.8 and143.2 ± 38.7 per 200 X field for the pre-immune serum and immune serum groups, respectively; P = 0.000). At the 1:50 dilution, the inhibition rate of sporozoite invasion by MEDLE-2 immune serum was 43.8% (250.1 ± 46.8 and 140.6 ± 35.8 per 200 X field for the pre-immune serum and immune serum groups, respectively; P = 0.000) (Figure 5).
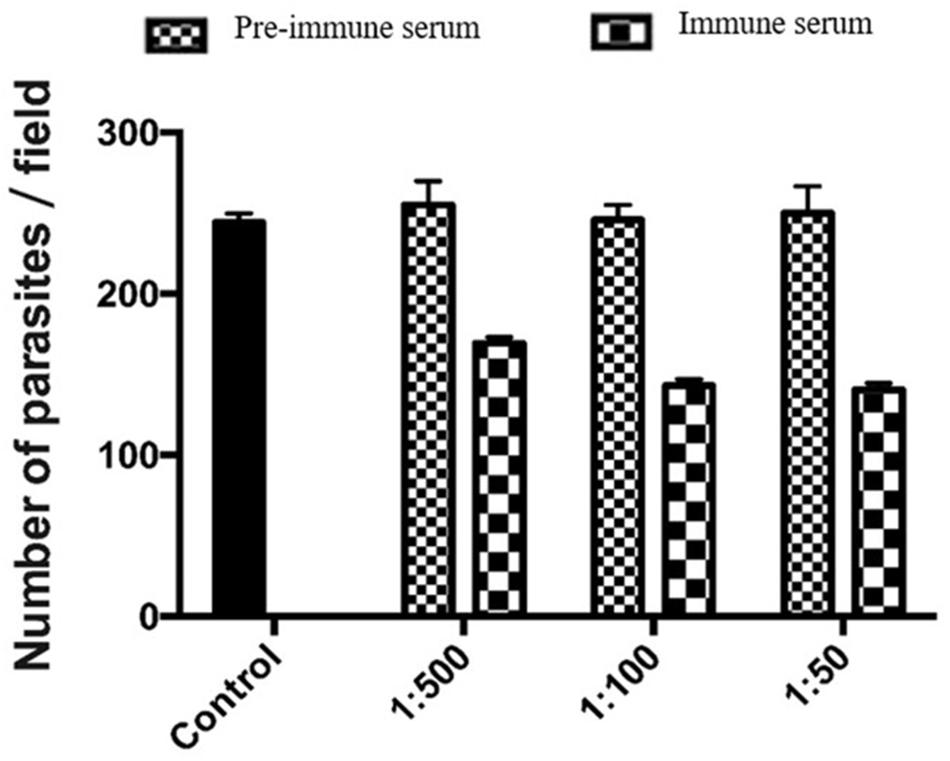
FIGURE 5. Inhibition of Cryptosporidium parvum invasion of HCT-8 cells by polyclonal MEDLE-2 antibodies. Data presented are mean ± SD parasites per 200 X field based on numbers from 30 randomly selected microscopy fields of C. parvum cultures. Control, C. parvum culture maintained in growth medium alone; 1:500, C. parvum culture with the addition of pre-immune or immune serum 1:500 dilution; 1:100, C. parvum culture with the addition of pre-immune or immune serum 1:100 dilution; 1:50, C. parvum culture with the addition of pre-immune or immune serum 1:50 dilution.
Discussion
Results of the study support the previous suggestion on the potential involvement of MEDLE proteins in the invasion of Cryptosporidium spp. This is based on the expression of the MEDLE-2 protein in C. parvum sporozoites and merozoites, the expression of the cgd5-4590 gene in multiple developmental stages of the parasite, and the significant reduction of parasite load by polyclonal MEDLE-2 antibodies in in vitro cultures. As in studies of other invasion-related proteins of Cryptosporidium spp. (Barnes et al., 1998; Cevallos et al., 2000; O’Connor et al., 2009), only a partial blockage of the invasion of sporozoites was produced by anti-MEDLE-2 antibodies. This is expected, as apicomplexans have been long known to use multiple strategies to invade host cells (Sibley, 2004). Like some other Cryptosporidium proteins involved in invasion (Strong et al., 2000), native MEDLE-2 appears to be O-glycosylated, although as not heavily. The neutralization ability of MEDLE-2 antibodies could be enhanced if the recombinant protein is expressed in a eukaryotic system that would produce proteins with similar O-linked glycosylation.
The function of MEDLE proteins is not clear. The MEDLE family of secreted proteins were initially identified through whole genome sequencing of C. parvum (Abrahamsen et al., 2004). They have significant sequence similarity to each other, and most of them end with the amino acid sequence MEDLE. As they have no conserved protein domains and no apparent counterparts in other apicomplexan parasites, MEDLE proteins are considered Cryptosporidium-specific (Abrahamsen et al., 2004). The biologic importance of MEDLE proteins is reflected by their numbers; C. parvum was identified to have six genes encoding the MEDLE proteins but only one copy of genes encoding most other proteins. The potential involvement of MEDLE proteins in host specificity and invasion of Cryptosporidium spp. was suggested by comparative genomic analysis of two genetically related species C. parvum and C. hominis, as the species with a broad host range, C. parvum, has six MEDLE genes, compared with one MEDLE gene in C. hominis, which mainly infects humans and non-human primates (Guo et al., 2015; Liu et al., 2016).
In this preliminary study, we have focused on the characterization of MEDLE-2. This protein was chosen as the first MEDEL protein to study largely because of its divergent sequence compared with other MEDLE proteins. While other MEDLE proteins share 55–75% amino acid sequence similarities, MEDLE-2 has <30% sequence identity to them. This is especially the case in the second half of the protein, which has minimal sequence identity to other MEDLE proteins and ends with LHDLE instead of MEDLE. This makes gene cloning and protein characterization of MEDLE-2 simpler.
The subcellular location of MEDLE-2 is not clear. Although MEDLE proteins are all called secreted proteins, it appears that only MEDEL-5 and MEDLE-6 encoded by subtelomeric genes in chromosome 6 have the predicted signal peptide. The native protein of MEDLE-2 is smaller than expected (17 kDa versus 21 kDa), indicating that it could be processed after its translation. Thus, different MEDLE proteins may have different subcellular locations and function differently. Further immunochemistry and biologic studies are needed to understand the function of MEDLE-2 and other MEDLE proteins. Studies are also needed to determine whether the function of these antigens is dependent on the presence of O-linked alpha-N-acetylgalactosamines.
Author Contributions
BL, JS, RJ, JJ, and NL conducted the experiments. BL, HW, YF, and LX analyzed the data and wrote the paper.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (31630078 and 31425025). The findings and conclusions in this report are those of the authors and do not necessarily represent the views of the U.S. Centers for Disease Control and Prevention.
Footnotes
References
Abrahamsen, M. S., Templeton, T. J., Enomoto, S., Abrahante, J. E., Zhu, G., Lancto, C. A., et al. (2004). Complete genome sequence of the apicomplexan, Cryptosporidium parvum. Science 304, 441–445. doi: 10.1126/science.1094786
Barnes, D. A., Bonnin, A., Huang, J. X., Gousset, L., Wu, J., Gut, J., et al. (1998). A novel multi-domain mucin-like glycoprotein of Cryptosporidium parvum mediates invasion. Mol. Biochem. Parasitol. 96, 93–110. doi: 10.1016/S0166-6851(98)00119-4
Bouzid, M., Hunter, P. R., Chalmers, R. M., and Tyler, K. M. (2013). Cryptosporidium pathogenicity and virulence. Clin. Microbiol. Rev. 26, 115–134. doi: 10.1128/CMR.00076-12
Cai, X., Woods, K. M., Upton, S. J., and Zhu, G. (2005). Application of quantitative real-time reverse transcription-PCR in assessing drug efficacy against the intracellular pathogen Cryptosporidium parvum in vitro. Antimicrob. Agents Chemother. 49, 4437–4442. doi: 10.1128/AAC.49.11.4437-4442.2005
Cevallos, A. M., Zhang, X., Waldor, M. K., Jaison, S., Zhou, X., Tzipori, S., et al. (2000). Molecular cloning and expression of a gene encoding Cryptosporidium parvum glycoproteins gp40 and gp15. Infect. Immun. 68, 4108–4116. doi: 10.1128/IAI.68.7.4108-4116.2000
Checkley, W., White, A. C., Jaganath, D., Arrowood, M. J., Chalmers, R. M., Chen, X.-M., et al. (2015). A review of the global burden, novel diagnostics, therapeutics, and vaccine targets for Cryptosporidium. Lancet Infect. Dis. 15, 85–94. doi: 10.1016/S1473-3099(14)70772-8
Deng, M., Templeton, T. J., London, N. R., Bauer, C., Schroeder, A. A., and Abrahamsen, M. S. (2002). Cryptosporidium parvum genes containing thrombospondin Type 1 domains. Infect. Immun. 70, 6987–6995. doi: 10.1128/IAI.70.12.6987-6995.2002
Feng, Y., Li, N., Roellig, D. M., Kelley, A., Liu, G., Amer, S., et al. (2017). Comparative genomic analysis of the IId subtype family of Cryptosporidium parvum. Int. J. Parasitol. 47, 281–290. doi: 10.1016/j.ijpara.2016.12.002
Guo, F., Zhang, H., Payne, H. R., and Zhu, G. (2016). Differential gene expression and protein localization of Cryptosporidium parvum fatty acyl-CoA synthetase isoforms. J. Eukaryot. Microbiol. 63, 233–246. doi: 10.1111/jeu.12272
Guo, Y., Tang, K., Rowe, L. A., Li, N., Roellig, D. M., Knipe, K., et al. (2015). Comparative genomic analysis reveals occurrence of genetic recombination in virulent Cryptosporidium hominis subtypes and telomeric gene duplications in Cryptosporidium parvum. BMC Genomics 16:320. doi: 10.1186/s12864-015-1517-1
Gut, J., and Nelson, R. G. (1999). Cryptosporidium parvum: synchronized excystation in vitro and evaluation of sporozoite infectivity with a new lectin-based assay. J. Eukaryot. Microbiol. 46, 56S–57S.
Jakobi, V., and Petry, F. (2006). Differential expression of Cryptosporidium parvum genes encoding sporozoite surface antigens in infected HCT-8 host cells. Microbes Infect. 8, 2186–2194. doi: 10.1016/j.micinf.2006.04.012
Liu, S., Roellig, D. M., Guo, Y., Li, N., Frace, M. A., Tang, K., et al. (2016). Evolution of mitosome metabolism and invasion-related proteins in Cryptosporidium. BMC Genomics 17:1006. doi: 10.1186/s12864-016-3343-5
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Ludington, J. G., and Ward, H. D. (2016). The Cryptosporidium parvum C-type lectin CpClec mediates infection of intestinal epithelial cells via interactions with sulfated proteoglycans. Infect. Immun. 84, 1593–1602. doi: 10.1128/IAI.01410-15
Mauzy, M. J., Enomoto, S., Lancto, C. A., Abrahamsen, M. S., and Rutherford, M. S. (2012). The Cryptosporidium parvum transcriptome during in vitro development. PLoS ONE 7:e31715. doi: 10.1371/journal.pone.0031715
O’Connor, R. M., Burns, P. B., Ha-Ngoc, T., Scarpato, K., Khan, W., Kang, G., et al. (2009). The polymorphic mucin antigens CpMuc4 and CpMuc5 are integral to Cryptosporidium parvum infection in vitro. Eukaryot. Cell 8, 461–469. doi: 10.1128/EC.00305-08
Ryan, U., Fayer, R., and Xiao, L. (2014). Cryptosporidium species in humans and animals: current understanding and research needs. Parasitology 141, 1667–1685. doi: 10.1017/S0031182014001085
Sibley, L. D. (2004). Intracellular parasite invasion strategies. Science 304, 248–253. doi: 10.1126/science.1094717
Strong, W. B., Gut, J., and Nelson, R. G. (2000). Cloning and sequence analysis of a highly polymorphic Cryptosporidium parvum gene encoding a 60-kilodalton glycoprotein and characterization of its 15- and 45-kilodalton zoite surface antigen products. Infect. Immun. 68, 4117–4134. doi: 10.1128/IAI.68.7.4117-4134.2000
Upton, S. J., Tilley, M., and Brillhart, D. B. (1995). Effects of select medium supplements on In Vitro development of Cryptosporidium parvum in HCT-8 Cells†. Clin. Microbiol. 33, 371–375.
Keywords: Cryptosporidium parvum, MEDLE family, invasion, expression, antigen
Citation: Li B, Wu H, Li N, Su J, Jia R, Jiang J, Feng Y and Xiao L (2017) Preliminary Characterization of MEDLE-2, a Protein Potentially Involved in the Invasion of Cryptosporidium parvum. Front. Microbiol. 8:1647. doi: 10.3389/fmicb.2017.01647
Received: 10 May 2017; Accepted: 15 August 2017;
Published: 31 August 2017.
Edited by:
Guan Zhu, Texas A&M University, United StatesReviewed by:
Honorine Ward, Tufts Medical Center, United StatesAlejandro Castellanos-Gonzalez, University of Texas Medical Branch, United States
Copyright © 2017 Li, Wu, Li, Su, Jia, Jiang, Feng and Xiao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yaoyu Feng, eXlmZW5nQGVjdXN0LmVkdS5jbg==