 Eléonore Frouin1
Eléonore Frouin1 Méline Bes1
Méline Bes1 Bernard Ollivier1
Bernard Ollivier1 Marianne Quéméneur1
Marianne Quéméneur1 Anne Postec1
Anne Postec1 Didier Debroas2
Didier Debroas2 Fabrice Armougom1
Fabrice Armougom1 Gaël Erauso1*
Gaël Erauso1*- 1Aix-Marseille Univ, Université de Toulon, CNRS, IRD, MIO UM 110, Marseille, France
- 2CNRS UMR 6023, Laboratoire “Microorganismes – Génome et Environnement”, Université Clermont Auvergne, Clermont-Ferrand, France
The Bay of Prony, South of New Caledonia, represents a unique serpentinite-hosted hydrothermal field due to its coastal situation. It harbors both submarine and intertidal active sites, discharging hydrogen- and methane-rich alkaline fluids of low salinity and mild temperature through porous carbonate edifices. In this study, we have extensively investigated the bacterial and archaeal communities inhabiting the hydrothermal chimneys from one intertidal and three submarine sites by 16S rRNA gene amplicon sequencing. We show that the bacterial community of the intertidal site is clearly distinct from that of the submarine sites with species distribution patterns driven by only a few abundant populations, affiliated to the Chloroflexi and Proteobacteria phyla. In contrast, the distribution of archaeal taxa seems less site-dependent, as exemplified by the co-occurrence, in both submarine and intertidal sites, of two dominant phylotypes of Methanosarcinales previously thought to be restricted to serpentinizing systems, either marine (Lost City Hydrothermal Field) or terrestrial (The Cedars ultrabasic springs). Over 70% of the phylotypes were rare and included, among others, all those affiliated to candidate divisions. We finally compared the distribution of bacterial and archaeal phylotypes of Prony Hydrothermal Field with those of five previously studied serpentinizing systems of geographically distant sites. Although sensu stricto no core microbial community was identified, a few uncultivated lineages, notably within the archaeal order Methanosarcinales and the bacterial class Dehalococcoidia (the candidate division MSBL5) were exclusively found in a few serpentinizing systems while other operational taxonomic units belonging to the orders Clostridiales, Thermoanaerobacterales, or the genus Hydrogenophaga, were abundantly distributed in several sites. These lineages may represent taxonomic signatures of serpentinizing ecosystems. These findings extend our current knowledge of the microbial diversity inhabiting serpentinizing systems and their biogeography.
Introduction
Serpentinization is the alteration process that abiotically transforms olivine and pyroxene-rich rocks into serpentinites and yields alkaline hot fluids enriched in H2 and CH4 (Chavagnac et al., 2013). Active serpentinization generates physico-chemical conditions that might have prevailed in early Earth and could have been fundamental with regard to the origin of life (Russell et al., 2010; Schrenk et al., 2013). Several serpentinizing systems, particularly terrestrial ones, have recently been studied by molecular approaches using next generation sequencing, to reveal the indigenous microbial community. They included the Cabeço de Vide Aquifer (CVA, Portugal) (Tiago and Veríssimo, 2013) and the ophiolites of Tablelands (Canada) (Brazelton et al., 2013), Santa Elena (Costa Rica) (Sánchez-Murillo et al., 2014; Crespo-Medina et al., 2017), Zambales (Philippines) (Woycheese et al., 2015), Samail (Oman) (Rempfert et al., 2017), Chimaera (Turkey) (Neubeck et al., 2017), Voltri (Italy) (Quéméneur et al., 2015; Brazelton et al., 2016), The Cedars (Placer, CA, United States) (Suzuki et al., 2017) and the nearby site of Coast Range Ophiolite Microbial Observatory (CROMO, Burbank, CA, United States) (Twing et al., 2017). In contrast, only a few submarine serpentinizing fields have been investigated using high-throughput sequencing. Their sampling is indeed complicated due to their remote geographic location and great depths. Among them, the emblematic Lost-City hydrothermal field (LCHF) was the only serpentinizing ecosystem to be subjected to microbiological study targeting the rare biosphere (Brazelton et al., 2010).
The recently described Prony Hydrothermal Field (PHF) (Pelletier et al., 2006; Monnin et al., 2014; Quéméneur et al., 2014; Postec et al., 2015) is a shallow (50 m maximum depth) marine alkaline hydrothermal system, located in the Bay of Prony, South of New Caledonia. The run-off waters from the rocky hills surrounding the Bay percolate through the densely fractured peridotite basement. The resulting end-member fluids discharge into the lagoon and, upon mixing with ambient seawater, precipitate, forming large carbonate edifices resembling Lost-City chimneys (Monnin et al., 2014). Similarly, to those of Lost-City, PHF fluids are characterized by high pH values (between 9.0 and 11.5) and elevated N2, H2, and CH4 concentrations (Monnin et al., 2014). However, the PHF fluids are different in terms of temperatures (maximum 40°C) and salinity (less than 0.5 g/L). The microbial community composition of the inner part of PHF chimneys has been previously characterized by culture-dependent (Ben Aissa et al., 2014, 2015; Mei et al., 2014; Bes et al., 2015), molecular-based techniques such as fingerprinting methods (SSCP and DGGE), clonal Sanger sequencing (Quéméneur et al., 2014; Postec et al., 2015), and Fluorescent In Situ Hybridization (FISH) and micro-imaging (Pisapia et al., 2017). Molecular-based studies revealed that bacteria largely outnumbered the archaea communities. The bacterial communities were composed of diverse populations mostly affiliated to the Chloroflexi and Firmicutes in the submarine sites and to the Proteobacteria in the intertidal sites (Quéméneur et al., 2014). The low-diversity archaeal communities were dominated by Methanosarcinales with phylotypes similar to those of the Lost-City Methanosarcinales (LCMS) and The Cedars Methanosarcinales (TCMS). While previous studies in PHF have identified the most abundant phylotypes, a substantial part of the indigenous microbial diversity was assumed to remain missing from the 16S rRNA genes inventories, due to the low-coverage sequencing (Quéméneur et al., 2014; Postec et al., 2015). Indeed, the coverage of the clone libraries was low, especially for the bacteria, with values ranging from 54 to 79%. Since such approaches underestimate the true biodiversity of environments (Curtis and Sloan, 2005), higher resolution techniques such as deep sequencing are therefore required in order to recover the overall microbial diversity of PHF. A preliminary study was performed with next-generation sequencing in PHF sites, but was only focused on the diversity of potentially hydrogen-producing bacteria and their hydrogenases (Mei et al., 2016).
In this study, we performed 454 pyrosequencing of the V1–V3 regions of the prokaryotic 16S rRNA gene in order to investigate the composition and the diversity of the microbial biosphere inhabiting carbonate chimneys of four sampling sites of the PHF. We showed that the diversity of bacteria (but not archaea) was site-specific and that the overall PHF biosphere was dominated by rare prokaryotic phylotypes. Finally, using publicly available sequences of 16S rRNA genes, we reported the co-occurrence of bacterial and archaeal operational taxonomic units (OTUs) identified in PHF and in other serpentinizing systems.
Materials and Methods
Sample Collection
Description of the sampling sites, sample processing and storage, and physico-chemical characteristics of PHF have been reported previously (Monnin et al., 2014; Quéméneur et al., 2014; Postec et al., 2015). Briefly, four active carbonate chimneys were sampled at four distinct sites of the Bay of Prony, including the “Bain des Japonais” (BdJ), the sites (ST) ST07 (also called “Aiguille de Prony”), ST09 and ST12. The BdJ site is located on the foreshore of the Carenage Bay and is uncovered at low tide. It consists of many fluid outlets building a carbonate plateau. There, small needle-like brittle structures of 10–20 cm high and 3–6 cm in diameter were collected. The submarine sites ST07, ST12, and ST09, located in the Prony Bay, were sampled at 16 meters below sea level (mbsl), 38 mbsl, and 48 mbsl, respectively. They form large carbonate edifices, carrying several chimneys of up to several meters high. The chimneys are porous structures, mainly composed of brucite [Mg(OH)2], aragonite and calcite (CaCO3) and magnesium carbonates (MgCO3) (Pisapia et al., 2017). The external walls of the mature submarine chimneys are colonized by various invertebrates (e.g., ascidia, sponges, corals). The top of such chimneys was collected by scuba-diving in each submarine site. They were kept at 4–8°C during their transportation (∼3 h) to the laboratory, where they were immediately processed. Cross sections (4 cm thick) of these chimneys top were realized aseptically; peripheral parts of the sections (2 cm thick) were cautiously removed using a chisel and sub-samples of the inside parts of the chimneys, bathed with the alkaline fluid, were collected and stored at –80°C until DNA extraction, as detailed previously (Quéméneur et al., 2014; Postec et al., 2015).
Ribosomal RNA Gene Amplification and 454 Pyrosequencing
DNA extraction and quantification procedures have been described previously (Postec et al., 2015). Briefly, about 0.4 g of chimney slurry was transferred into a sterile 2 mL tube containing glass beads (lysing matrix E from MP BioMedicals) and 0.8 mL of lysis buffer (Tris.HCl 100 mM, NaCl 100 mM, EDTA 50 mM, pH 8). Cell lysis was achieved with chemical reactions (adding Sodium Dodecyl Sulfate and Lauryl Sarcosine) and mechanic disruption using a Fast Prep homogenizer. DNA was extracted using phenol:chloroform:isoamyl alcohol and chloroform:isoamyl alcohol. DNA from each sample or dilution was used as template for 16S rRNA amplification. The hypervariable region V1–V3 of the 16S rRNA gene was amplified using the pair of primers 27F (5′-AGAGTTTGATCCTGGCTCAG-3′)/533R (5′-TTACCGCGGCTGCTGGCAC-3′) and 21F (5′-TCYGKTTGATCCYGSCRGA-3′)/529R (5′-TCGCGCCTGCTGCRCCCCGT-3′) for bacteria and archaea, respectively. The forward and reverse primers were barcoded (Supplementary Table S1). PCR reactions were performed in a 50 μl mixture containing PlatiniumTM Taq High Fidelity (Life Technologies) 1 unit, High Fidelity PCR Buffer 1X, dNTP mixture 0.2 mM each, MgSO4 2 mM, primer mix 0.3 μM each, and 2 μl template DNA. The protocol mentioned below was used for amplification: initial denaturation at 95°C for 2 min, followed by 25 cycles of denaturation 95°C for 45 s, primers annealing at 54°C for 45 s, elongation at 72°C for 1 min 30 s, and a final elongation step at 72°C for 7 min. At least three PCR products from each combination of sample and primer pairs were pooled and then purified with QIAquick Gel Extraction Kit (Qiagen). An equimolar amount of each PCR pool was finally used for library construction (addition of the adaptors by ligation) and pyrosequencing using 454 GS FLX Titanium technology (Beckman Coulter Genomics).
16S rRNA Gene Analysis
Pyrosequencing of the V1–V3 region of the microbial 16S rRNA generated 188,058 sequences. Raw nucleotide sequence data were submitted to the European Nucleotide Archive under project accession number PRJEB21795. These sequences were filtered according the following criteria: read length between 150 and 500 bp, no ambiguous bases, average quality score ≥25, and no error allowed in primer and barcode sequences. A total of 74,457 and 95,916 high quality sequences were obtained for the bacteria and archaea, respectively. Alignments and clustering to Operational Taxonomic Units (OTUs) were carried out using the QIIME 1.9.1 workflow (http://www.qiime.org/) (Caporaso et al., 2010b). Chimera sequences were identified and removed using UCHIME (Edgar et al., 2011). Multiple sequence alignments were performed using PyNAST (Caporaso et al., 2010a). OTUs were defined by clustering sequences using UCLUST (Edgar, 2010) with a pairwise distance threshold value of 3%, a cutoff currently used for species demarcation while reducing the potential inflation of the number of OTUs due to pyrosequencing errors. To limit the impact of spurious OTUs, low abundance OTUs accounting for less than 0.005% of total sequences were removed as recommended by Bokulich et al. (2013). Taxonomic assignment of the filtered sequences was performed using the RDP classifier algorithm (Wang et al., 2007) with a minimum bootstrap confidence of 80% against the SILVA database (Quast et al., 2013) release for QIIME (v.123). The OTU table was rarefied (i.e., downsampled to the sample with the smallest set of sequences) to reduce sequencing depth heterogeneity between samples. There is currently no standardized threshold for the rare biosphere. Here, rare OTUs were defined as OTUs comprising 0.005 to 0.2% of sequences per sample, an empirical threshold previously adopted by Hugoni et al. (2013). OTUs comprising 0.2 to 1% of sequences per sample were considered as intermediate and OTUs representing more than 1% of sequences per sample were considered as abundant. The alpha diversity within the four samples was estimated using the Shannon (Shannon and Weaver, 1949) and Simpson (Simpson, 1949) indices, and the beta diversity among the samples was measured using weighted UniFrac (Lozupone and Knight, 2005) metric. Taxonomic composition, and diversity results were visualized using the R package phyloseq v.1.14.0 (McMurdie and Holmes, 2013). BLASTn (Basic Local Alignment Search Tool) searches against the NCBI (National Center for Biotechnology Information) database of nucleotides (nt) and against the RefSeq database were performed for some OTU sequences. To infer the phylogeny of Methanosarcinales, cultivated members of this order and environmental clones were added to the sequences of abundant or ubiquitous PHF OTUs. The V1–V3 regions of these sequences were aligned with MAFFT v7.123b (Katoh et al., 2002) and a maximum likelihood tree was inferred using the software SeaView (Gouy et al., 2010) with 1000 bootstrapped trials.
Taxonomic Comparison of Phylotypes in Serpentinizing Environments
The correctly formatted amplicons and clone libraries of five serpentinizing ecosystems were downloaded from public databases (SRA and Genbank). These data constituted a subset of known serpentinizing systems, including four continental environments: Cabeço de Vide Aquifer (Tiago and Veríssimo, 2013), The Cedars (Suzuki et al., 2013), Voltri ophiolite (Quéméneur et al., 2015), Leka ophiolite (Daae et al., 2013), and one submarine system: the famous Lost-City Hydrothermal Field (Schrenk et al., 2004; Brazelton et al., 2006). A closed-reference OTU picking process was carried out to compare the sequences of amplicons and clone libraries covering different regions of the 16S rRNA gene. The sequences were clustered against a reference sequence database (SILVA database, v123). The OTUs shared among samples were visualized with Anvi’o v2.3.2 (Eren et al., 2015).
Results and Discussion
Microbial Community Diversity
The bacterial species richness of the sampling sites comprised 1,979 distinct OTUs (Supplementary Figure S1A). However, the species richness distribution varied significantly from one site to another, with at least a twofold decrease in OTU abundance for BdJ, ST07, and ST09 compared to the ST12 site (1203 OTUs, Supplementary Table S2). The presence of more than 780 OTUs, specific to the ST12 site (Supplementary Figure S1A) and comprising mainly of rare taxa, might result from a lower in situ pH condition (around 9.2), in comparison with the other studied PHF sites (pH > 9.8) (Monnin et al., 2014). This trend has previously been observed in the Coast Range Ophiolite serpentinizing site, where circumneutral pH wells contained many rare taxa that were absent in the high pH ones (Twing et al., 2017), most likely because these latter drastic conditions selected the few most alkaliphilic members within the prokaryotic community. The overall species richness was considerably lower in the archaea (310 OTUs) than in the bacteria, and also showed an imbalance in OTU distribution across the four sites (Supplementary Figure S1B and Supplementary Table S2). The Shannon and Simpson indexes indicated lower archaeal diversity, as compared to the bacterial one (Supplementary Figure S2 and Supplementary Table S2). This lower richness and diversity of archaea is not specific to PHF as it has been generally observed in various environments globally (Aller and Kemp, 2008), including deep-sea hydrothermal vents (Anderson et al., 2015) and Lost-City (Brazelton et al., 2010). The rarefaction curves had a near-asymptotic shape (Supplementary Figure S3) suggesting that the sequencing effort was suitable to capture the near-full diversity of this serpentinizing environment.
Bacterial Phylogenetic Composition in PHF
The bacterial community composition of the PHF sites encompassed 22 different phyla (including six candidate phyla) reflecting a high biodiversity. The phylum Proteobacteria was preponderant in the intertidal BdJ (78.4% of sequences, 244 OTUs), while the three submarine sites were mainly composed of Chloroflexi (50.7–54.8%, 150–161 OTUs) and, to a lesser extent, of Proteobacteria members (21.1–27.4%, 250–603 OTUs) (Figure 1). Remarkably, all other bacterial phyla shared by all the PHF sites were essentially recovered at much lower abundance. These bacterial phyla comprised Cyanobacteria (0.2–16.0%), Actinobacteria (0.2–4.8%), Bacteroidetes (0.2–3.1%), Firmicutes (0.1–5.3%), Planctomycetes (0.2–1.6%), and the candidate phylum Parcubacteria (formerly candidate division OD1) (0.01–0.6%). By contrast, the Acetothermia, Acidobacteria, Gemmatimonadetes, and Nitrospirae phyla were only retrieved from submarine sites, representing 7.4% of the total sequences. Relative abundances of some bacterial phyla detected in our study were not in agreement with those reported by sequencing clone libraries (Quéméneur et al., 2014; Postec et al., 2015). This was the case for the Firmicutes and the Parcubacteria, which were found in much lower proportion in this study. Nevertheless, significant shifts in phyla proportions have already been observed in the same chimney edifice over a 6 year survey (Postec et al., 2015). Our pyrosequencing approach also allowed the identification of 6 novel phyla that had not been previously detected. Members of all these phyla, i.e., Fibrobacteres, Lentisphaerae, Marinimicrobia, PAUC34f, Saccharibacteria, and Verrucomicrobia, were found among the large diversity of the ST12 site. On the contrary, several phyla previously found in PHF: Atribacteria, Microgeomates, Omnitrophica, and candidate division NPL-UPA2 (Quéméneur et al., 2014; Postec et al., 2015; Mei et al., 2016; Pisapia et al., 2017) were not represented in this study. This discrepancy may be explained by the use of different PCR primers targeting different regions of the 16S rRNA gene of bacteria and archaea. It may also be the consequence of the ongoing evolution of the new generation sequencing technology (i.e., 454 Roche vs. Illumina). That raises a major issue regarding the lack of standards for 16S rRNA gene surveys, which reduces the reproducibility and complicates comparisons between studies (Escobar-Zepeda et al., 2015). Additionally, the sampling area of the chimneys (either central or more peripheral) could also explain these taxonomic discrepancies. Indeed, the highly porous structure of the chimney interiors is exposed to steep physicochemical gradients (in terms of salinity, pH, redox potential, nutrients) along their transverse axis, defining microhabitats colonized by contrasted microbial communities.
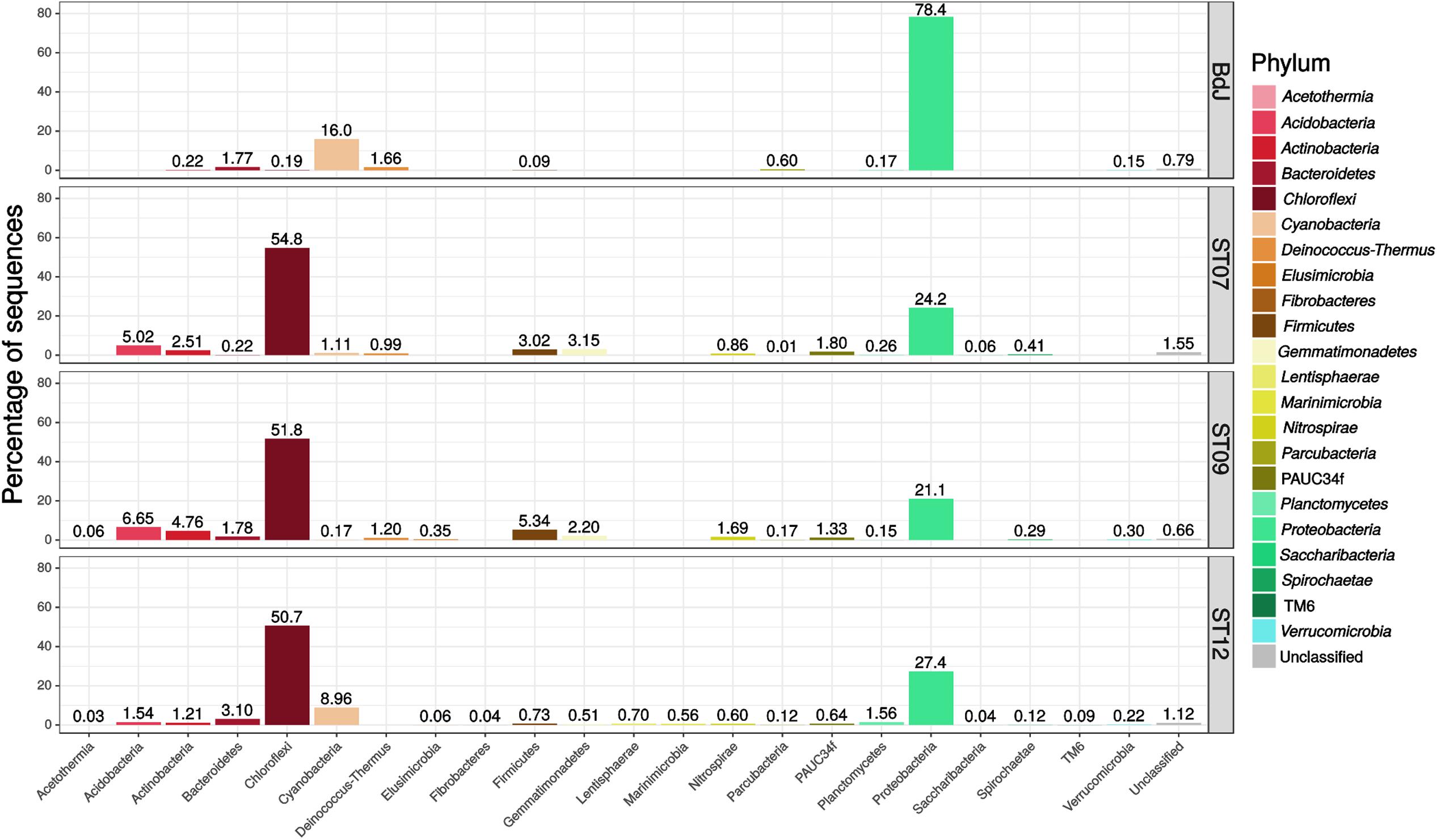
FIGURE 1. Phylum-level bacterial diversity in Prony Hydrothermal Field (PHF) sites based on 16S rRNA gene sequences (V1–V3 region). The relative abundances were estimated for the four chimneys collected in BdJ, ST07, ST09, and ST12 sites.
Archaeal Phylogenetic Composition in PHF
The archaeal community of the PHF sites mainly comprised potential methanogens and autotrophic ammonia-oxidizer representatives among Euryarchaeota and Thaumarchaeota phyla, respectively. The BdJ and ST12 sites were dominated by members of Methanosarcinales (99.8% of sequences, 38 OTUs and 94.6%, 44 OTUs, respectively) within the class Methanomicrobia (Figure 2). All abundant and ubiquitous OTUs of Methanosarcinales were similar to the sequences from other serpentinite-hosted ecosystems, as shown by OTU-based phylogeny (Figure 3). The first set of OTUs constituted a sister group of Lost-City Methanosarcinales (LCMS) (Schrenk et al., 2004) whereas the remaining OTUs were closely associated with the sequence of The Cedars Methanosarcinales (TCMS) (phylotype Ced_A01) (Suzuki et al., 2013). Although the present study reported a more accurate diversity than the previous studies based on clone libraries, no new phylotypes were identified within the Methanosarcinales group. Interestingly, although first detected in a terrestrial serpentinizing system (The Cedars), TCMS-like archaea were detected at lower abundance in the intertidal BdJ site than in the submarine sites of PHF. Moreover, here the overall abundance of the LCMS-like was two-fold higher than the abundance of the TCMS-like (Figure 3). This result is in opposition with the ratio previously observed in clone libraries and thus contradicts the hypothesis that the predominance of TCMS-like archaea in PHF was due to the low salinity of fluids (of meteoric origin such as in The Cedars) (Postec et al., 2015). Finally, these two specific phylotypes of Methanosarcinales were ubiquitous in all PHF sites. In contrast, the ST07 and ST09 sites were dominated by the Thaumarchaeota Marine Group I (69.4%, 179 OTUs and 62.4%, 194 OTUs, respectively), which was detected in all submarine sites despite its low abundance in the ST12 site (Figure 2). The Marine Group I is frequently observed in the global ocean (Auguet et al., 2010; Fernàndez-Guerra and Casamayor, 2012), which may explain why they are found only in submarine sites of PHF. In addition, the Thaumarchaeota Soil Crenarchaeotic Group was also detected in the submarine sites at very low-abundance level (0.1 to 1.9%, 3 to 6 OTUs) and may originate from the surrounding hills having been carried away by run-off waters. Other less represented archaeal classes belonged to the Thermoplasmata (mainly Marine Group II) (0.1–0.7%, 7–18 OTUs), Methanobacteria (0.1%, 2 OTUs at BdJ), Terrestrial Group of Thaumarchaeota (0.1%, 2 OTUs in ST07) and presumed thermophiles of the Thermococci class (0.1%, 1 OTU at ST12) (Figure 2). Finally, the remaining OTUs were affiliated to the Deep Sea Euryarchaeotic Group within the Aenigmarchaeota phylum and to the Miscellaneous Crenarchaeotic Group that were not previously detected in PHF. The archaea of the Miscellaneous Crenarchaeotic Group are widespread in marine sediment and are essentially retrieved from anoxic environments such as deep oceanic subsurface sediments or mud volcanoes (Kubo et al., 2012). Since the temperature of PHF fluids measured at the chimney outlets did not exceed 43°C, the presence of archaea closely related to hyperthermophilic species (e.g., Thermococcus spp.) originating from deep-sea environments was surprising. Despite their low abundance, such representatives of the Thermococcales were, however, previously detected in PHF, even in clone libraries, and the high GC content of their sequence strongly suggested they were bona fide hyperthermophiles (Postec et al., 2015). It is thus likely that they originated from subsurface hyperthermophilic microbial communities fed by serpentinization reactions and carried up to the surface by the fluid as previously suggested (Postec et al., 2015).
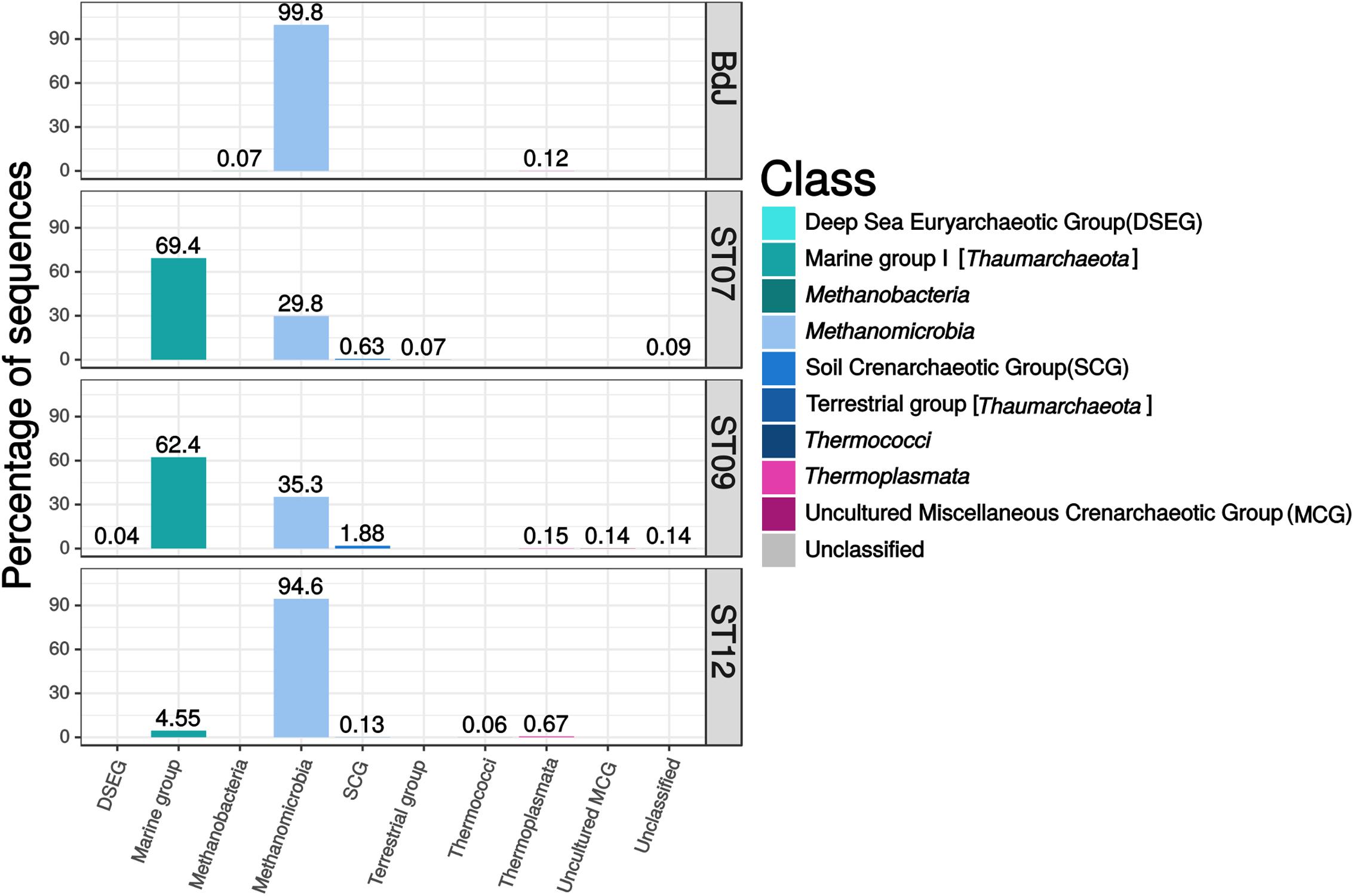
FIGURE 2. Class-level archaeal diversity in PHF sites based on 16S rRNA gene sequences (V1–V3 region). The relative abundances were estimated for the four chimneys collected in BdJ, ST07, ST09, and ST12 sites.
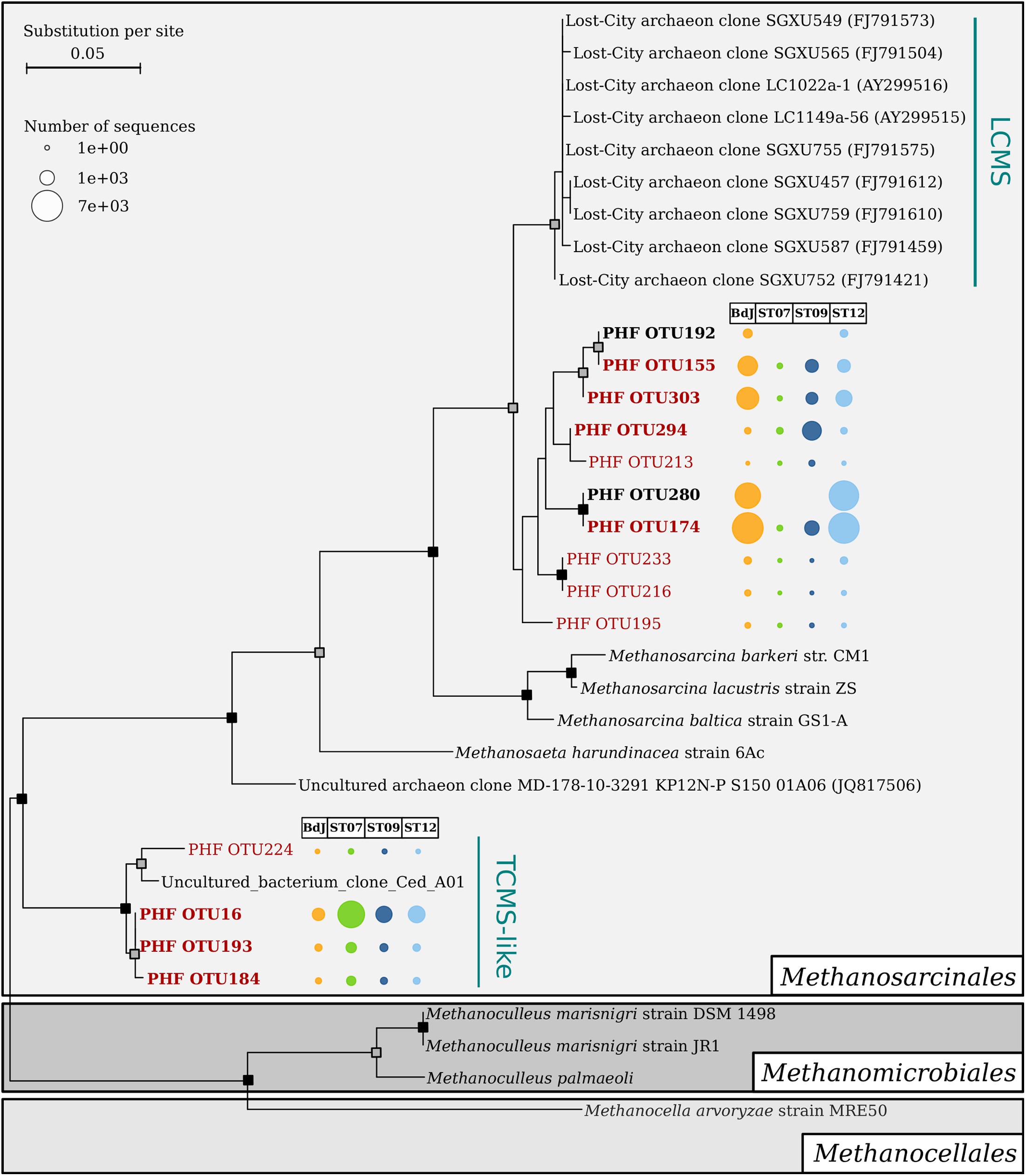
FIGURE 3. Maximum likelihood phylogenetic tree of V1–V3 archaeal 16S rRNA sequences affiliated to the order Methanosarcinales. Bootstrap support values are indicated on nodes with a gray or black square for support greater than 70% or 90%, respectively. Scale bar indicates substitutions per site. The Methanosarcinales sequences are marked in bold if they were abundant in at least one site of PHF, and red font color indicates ubiquitous operational taxonomic units (OTUs; found in the four chimneys of PHF). The bubble chart corresponds to the contingency table of OTUs. Uncultured Methanosarcinales clusters are abbreviated as follows: LCMS, Lost-City Methanosarcinales, and TCMS, The Cedars Methanosarcinales.
Beta-Diversity and Abundant OTUs across the PHF Sites
The bacteria taxonomic profiles observed for the three submarine sites were very similar, while the one observed for the intertidal site (BdJ) differed significantly (Figure 4A). This pattern of distribution, previously observed in a study on PHF (Quéméneur et al., 2014), may be explained by the influence of the marine environment regime (intertidal vs. submarine) variation in the composition of the end-member fluids (Monnin et al., 2014) or the resulting differences observed in the mineralogy of the carbonate chimneys (Pisapia et al., 2017). The relationship between species and sampling sites was mainly driven by a few dominant OTUs in the ordination plot (Figure 4A). Among them, five abundant OTUs in submarine sites were assigned to the class Dehalococcoidia (DEH, phylum Chloroflexi) and were related to clones retrieved from the serpentinizing springs of The Cedars (Suzuki et al., 2013). Two of these dominant OTUs (#915 and #1117) were assigned to the candidate order MSBL5 (Mediterranean Sea Brine Lake group 5) and three (#1432, #1706, and #1947) to the division GIF9 (Groundwater InFlow clone 9) (Supplementary Table S3). The candidate orders MSBL5 and GIF9 were originally detected in a deep-sea anoxic hypersaline lake (Daffonchio et al., 2006) and in a contaminated groundwater-fed bio reactor (Alfreider et al., 2002), respectively. These five dominant OTUs are quite divergent from any cultivated species (the closest similarity is found for OTU1947 with Dehalogenimonas alkenigignens, at 82% identity). The metabolic capabilities of these candidate groups are still largely unknown. Even for the entire class Dehalococcoidia, our knowledge of metabolism is essentially limited to that of the few cultivated representatives, which have the ability to oxidize hydrogen and reduce organohalogenic compounds (Kaster et al., 2014; Biderre-Petit et al., 2016). However, a single cell sequencing of a Dehalococcoidia phylotype, abundant in marine sediments, suggested that this bacterium may replace organohalides by dimethyl sulfoxide as a terminal electron acceptor (Wasmund et al., 2013). In the BdJ site, 4 OTUs (#1298, #153, #1806, #279) representing more than a third of all sequences (Figure 4A) were assigned to the haloalkaliphilic genus Roseibaca (Alphaproteobacteria), a sister genus of the phototrophic anaerobic sulfur-oxidizing Rhodobaca, previously detected as dominant in BdJ (Mei et al., 2016). Representatives of the Roseibaca genus are known to be rose-colored (Labrenz et al., 2009) and could in part explain the dark rose color of the biofilm covering the exteriors of BdJ chimneys. The BdJ site also contained two dominant OTUs (#607 and #763) affiliated to the Comamonadaceae family (Betaproteobacteria) showing 98% identity with Hydrogenophaga defluvii (Supplementary Table S3). Members of the genus Hydrogenophaga are chemoorganotrophic or chemolithoautotrophic, thriving by oxidation of hydrogen under microaerophilic conditions (Willems, 2014). Despite the potential involvement of Firmicutes in the organomineralization of carbonates chimneys in PHF (Pisapia et al., 2017), only one abundant OTU of this phylum was identified in submarine sites and assigned to uncultured anaerobic Syntrophomonadaceae. This OTU, however, differed from the one (KM207235 HPst091-1-1) reported to be dominant in juvenile chimneys of PHF (Pisapia et al., 2017), most likely because of the differences of age of the chimneys studied (i.e. mature vs. juvenile). Members of this family are anaerobes that usually grow in syntrophic association with hydrogenotrophic and formate-utilizing methanogens by thriving energy from oxidation of carboxylic acids (Sobieraj and Boone, 2006). These anaerobic oxidations are thermodynamically feasible only when the H2 or formate produced is concomitantly consumed by methanogens or other syntrophic partner. Recently, syntrophic associations were shown to permit the conversion of organic acids and alcohols to methane under extremely haloalkaline conditions (Sorokin et al., 2016). In PHF, such syntrophic associations between uncultivated members of the Syntrophomonadaceae and the Methanosarcinales could be a relevant strategy to overcome energetic limitations, due to the high pH conditions and the lack of electron acceptors.
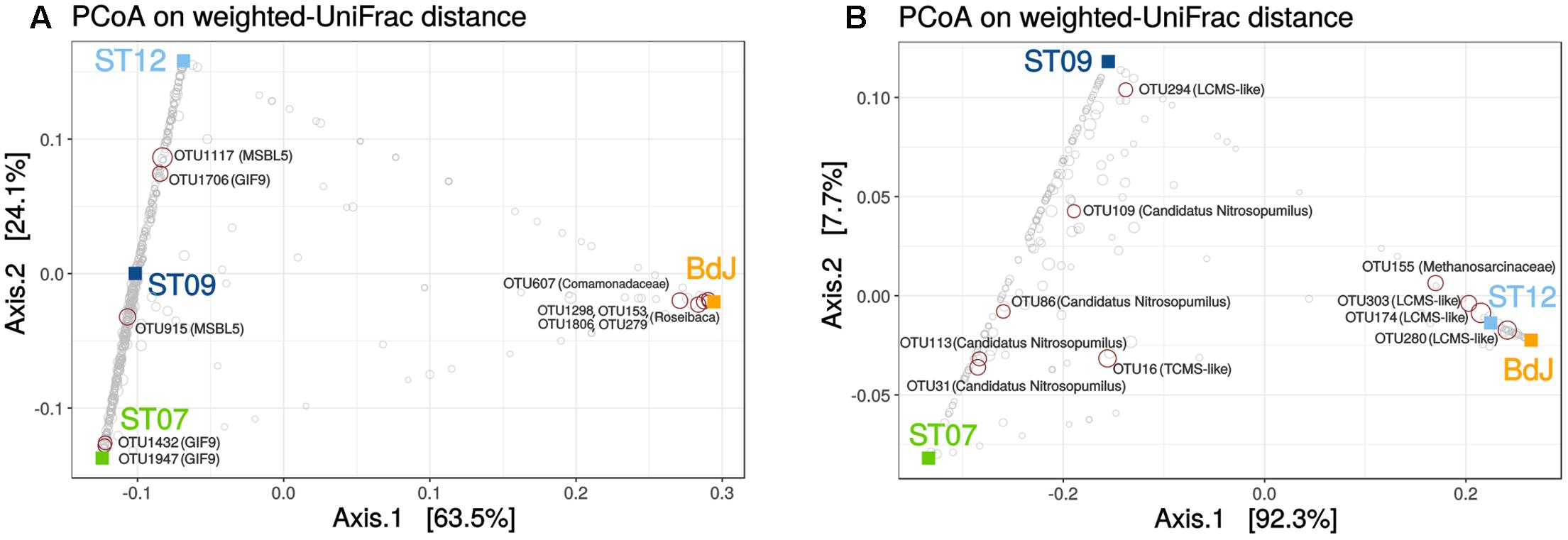
FIGURE 4. Beta diversity of bacterial (A) and archaeal (B) communities of PHF. PCoA analysis was carried out using the weighted UniFrac metric. Squares indicate PHF sites and circles correspond to OTUs. Taxonomic annotation was added to the top ten OTUs explaining this partition (red circles). Lost-City Methanosarcinales and The Cedars Methanosarcinales are abbreviated as LCMS and TCMS, respectively.
For archaea, no differential segregation of OTUs between intertidal and submarine sites could be observed, in contrast to the trend previously reported (Quéméneur et al., 2014). Nevertheless, the beta-diversity analysis linked BdJ to ST12 (Figure 4B), with both sites harboring abundant OTUs related to the LCMS (their annotation was refined through phylogenetic analysis, Figure 3), whereas ST09 clustered with ST07 because they shared abundant OTUs related to Candidatus Nitrosopumilus (Thaumarchaeota) and numerous OTUs of Marine Group I (Figure 4B and Supplementary Table S4). In light of these beta-diversity results, the archaeal community seemed less site-dependent than the bacterial one, as exemplified by the two groups of Methanosarcinales OTUs (LCMS and TCMS) found in both intertidal and submarine serpentinizing systems of PHF sites.
Comparison between Phylotypes of Serpentinizing Environments
A comparison of microbial communities across six geographically distant serpentinizing systems was carried out at the OTU level (at 97% identity threshold) used as a proxy of the prokaryotic species that is the most commonly found unit in microbial biogeographic studies. For practical reasons, this comparison was limited to the fraction of known microorganisms (i.e., those which could be assigned to a taxon in the Silva database), thus the results cannot be extrapolated to the whole prokaryotic community. Despite this limitation, some biogeographic patterns could be observed. The general OTU composition appeared to be quite different from one serpentinizing system to another and no OTU was common to all sites, but several related phylotypes were recovered from more than two systems; among them, a few were detected exclusively in serpentinizing systems. The distribution pattern of PHF phylotypes in other geographic sites and its implications is discussed below.
OTUs Belonging to Abundant Taxa Commonly Detected in Serpentinite-Hosted Ecosystems
It mainly concerns the Betaproteobacteria and the Clostridia (Schrenk et al., 2013). The abundance of Betaproteobacteria often exceeded 20% in the terrestrial systems studied herein (Daae et al., 2013; Quéméneur et al., 2015; Suzuki et al., 2013; Tiago and Veríssimo, 2013). Four Betaproteobacteria OTUs, found in both PHF sites and continental serpentinizing springs, were assigned to the H2-consuming genus Hydrogenophaga (Figure 5). Although not exclusive of serpentinizing systems, these OTUs were otherwise distributed in similar geothermal environments such as deep hydrothermal groundwater and sulphidic springs (Supplementary Table S5). This suggests that similar geochemical settings (presence of H2 rich fluids and oxic-anoxic interfaces) may have selected Hydrogenophaga species which metabolic capabilities are well adapted to these particular conditions. Indeed, 16S rRNA gene and, in some cases, related NiFe hydrogenases genes of Hydrogenophaga spp. were previously shown to be abundant in the serpentinizing-hosted systems listed in the comparison and others such as the springs of the Tablelands Ophiolite (Brazelton et al., 2012) or the Samail Ophiolite (Rempfert et al., 2017). It is noteworthy that no OTU representative of its sister genus Serpentinomonas was identified in PHF. These hyperalkaliphilic hydrogenotrophs isolated from The Cedars (Suzuki et al., 2014) have so far been detected only in terrestrial serpentinizing sites where they can be dominant, e.g., one OTU representing up to 14% of the sequences in Voltri (Quéméneur et al., 2015), or even constituting the unique OTU of Betaproteobacteria, accounting for 50% of the sequences, in the most alkaline well CSW1.1 (pH12.2) of CROMO but absent in the other wells with lower pH values (Twing et al., 2017). Otherwise, the submarine PHF sites shared a Ralstonia OTU (EU030486.1) with Lost-City, emphasizing the diversity of microorganisms that could oxidize hydrogen in the PHF sites.
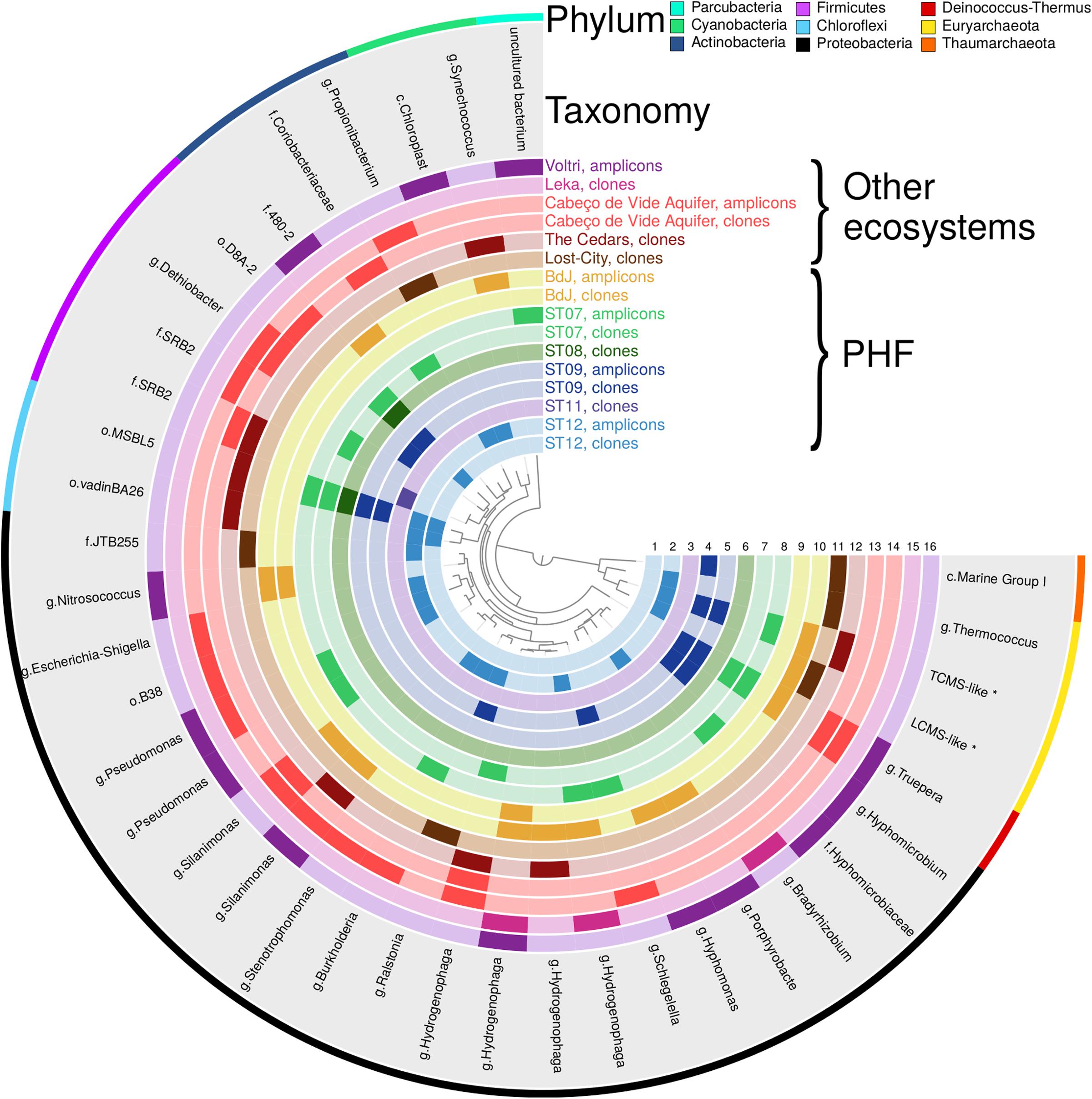
FIGURE 5. Presence-absence plot of OTUs shared at least by one sample of PHF sites and one sample from another serpentinizing ecosystem. The inner circles, numbered from 1 to 16, correspond to the different samples. The 13 samples of PHF include the circles 2, 5, 8, 10 (this study) and 1, 3, 4, 6, 7, 9 (Quéméneur et al., 2014; Postec et al., 2015; Pisapia et al., 2017). The seven other serpentinizing samples comprise the circles 11 from Lost-City Hydrothermal Field (Schrenk et al., 2004; Brazelton et al., 2006), 12 from The Cedars (Suzuki et al., 2013), 13, 14, from Cabeço de Vide (Tiago and Veríssimo, 2013), 15 from Leka ophiolite (Daae et al., 2013), and 16 from Voltri ophiolite (Quéméneur et al., 2015). Each circle color corresponds to a sampling system and the darker cells indicate the presence of OTUs. The taxonomic affiliation of OTUs is indicated in the gray circle at genus level (or at the lowest taxonomic rank). Letters g, f, and o refer to the ranks genus, family and order, respectively. The outer circle specifies the phylum of each OTU.
In the Clostridia class, 4 OTUs representing clone sequences of PHF were also detected at CVA, at The Cedars. However, phylotypes closely related to these four OTUs were more generally distributed in other serpentinizing sites including Voltri and CROMO. As noticed for the Hydrogenophaga OTUs, these 4 OTUs were restricted to serpentinizing systems or to groundwater. The OTUs AM778006.1 and AB476673.1 were close to the candidate group SRB2 found in CROMO (Twing et al., 2017), while AM777965.1 was affiliated to the Dethiobacter genus, of which the only described species, D. alkaliphilus, is strictly anaerobic with a respiration metabolism and able to use acetate and sugars with sulfur, polysulfides, or thiosulfate as an electron acceptor (Sorokin et al., 2008). This bacterium has also been shown to be capable of chemolithoautotrophic growth by disproportion of elemental sulfur (a kind of mineral fermentation), a metabolic process which is more favorable under alkaline conditions (Poser et al., 2013) such those encountered in serpentinizing systems.
Regarding the archaeal community, the present comparison included the uncultivated Methanosarcinales lineages only detected in serpentinizing ecosystems (Quéméneur et al., 2014, 2015; Schrenk et al., 2004; Suzuki et al., 2013). The originality of PHF is provided by the simultaneous presence of the two Methanosarcinales OTUs: the LCMS and TCMS phylotypes (Figure 5 and Supplementary Table S6).
OTUs Belonging to Dominant Taxa in PHF Not Generally Distributed in Other Sites
In the PHF, uncultured Chloroflexi and members of the class Alphaproteobacteria were the main taxa, discriminating the submarine sites from the intertidal one. The large Chloroflexi fraction of the PHF submarine sites was not commonly recovered among other serpentinizing systems, except in less active or extinct chimneys of the Lost-City Hydrothermal Field (Brazelton et al., 2010) and in deep ground water of The Cedars (Suzuki et al., 2013). Additionally, the ecosystem of The Cedars shared two uncultivated Chloroflexi OTU with PHF (Figure 5). The high-abundance level of Alphaproteobacteria (64.7%) in the intertidal BdJ site was also not representative of all serpentinizing systems, as exemplified by their low abundance in CVA and The Cedars ecosystems (Suzuki et al., 2013; Tiago and Veríssimo, 2013). The PHF sites shared, however, five Alphaproteobacteria OTUs assigned to N2-fixing Rhizobiales, chemoheterotrophic Caulobacterales and Sphingomonadales with the Leka ophiolite or Voltri. Thaumarchaeota of the Marine Group I (MG-I) class (Supplementary Tables S4, S6) also represented a dominant taxon in half of the PHF sites as outlined above. One PHF OTU (AY505046.1) was also detected within a group of new MG-I sequences from Lost-City (Brazelton et al., 2006) (Figure 5). These sequences may represent a new ecotype of Thaumarchaeota specifically adapted to serpentinizing hydrothermal system conditions and, in particular, to low ammonium concentrations (Russell et al., 2010) and high pH values (>10). Precisely, pH adaptation has shown evidence to be the major driving force in Thaumarchaeota evolutionary diversification (Gubry-rangin et al., 2015). Thus, serpentinite-hosted systems may represent a niche for specialized alkaliphilic Thaumarchaeota.
OTUs Belonging to Alkaliphilic Cosmopolitan Taxa Shared with Other Serpentinizing Sites
Two Gammaproteobacteria OTUs assigned to the genus Silanimonas (AM778014.1 and KF511881.1) were detected in BdJ and CVA and even in The Cedars and in Voltri for KF511881.1. The genus Silanimonas contains three species (Chun et al., 2017), two of which are slightly thermophilic alkaliphiles with pH optima for growth around 9–10. Although OTU AM778014.1 seems restricted to serpentinizing sites, OTU KF511881.1 is also commonly detected in various types of aquatic habitat such as lake water, marine sediment or hot springs. Considering the presence of Silanimonas in the four serpentinizing systems with a maximum pH around 11.5–12.5 and its absence in the Leka ophiolite (where the maximum in situ pH is 9.6; Daae et al., 2013), this genus might be an indicator of a highly alkaline environment rather than terrestrial serpentinizing ecosystems. A similar conclusion may be drawn for the Truepera OTU (Deinococcus-Thermus phylum) shared by the PHF sites with high pH fluid, Voltri and CVA. Members of the genus Truepera are known to be chemoorganotrophic alkaliphiles, slightly thermophilic and extremely resistant to ionizing radiation.
OTUs Endemic to the PHF Belonging to Candidate Phyla
The OTUs classified into the six bacterial candidate phyla detected in PHF were not recovered in other serpentinizing systems, with the exception of one Parcubacteria OTU (LN561460.1) identified in both the submarine ST07 site and the Voltri terrestrial springs. Members of this candidate phylum could be involved in ‘dark’ anaerobic fermentation (Castelle et al., 2017). Several metagenomic reconstructed genomes of Parcubacteria are very small with reduced metabolic capabilities (e.g., lacking ATP synthase genes), suggesting that they may be intercellular symbionts of other prokaryotes sharing the same habitat (Nelson and Stegen, 2015; Suzuki et al., 2017). Since some Parcubacteria members were found in abundance in the deep groundwater of The Cedars (as well as in Lost-City and in PHF) they were thought to be selected by the geochemical conditions associated with serpentinization reactions (Suzuki et al., 2017). However, no Parcubacteria OTU was shared by PHF and The Cedars (Figure 5) and their closest OTUs indicated less than 90% identity on their overlapping region. Although clone sequencing of The Cedars (Suzuki et al., 2013) may have failed to identify the overall Parcubacteria community, PHF most likely hosted distantly related taxa potentially associated with other metabolisms and different microbial consortia.
Conclusion
In this study, 16S rRNA gene pyrosequencing provided a comprehensive overview of the microbial biosphere of PHF and extended its known diversity by revealing the presence of prokaryotic populations pertaining to six novel phyla not previously detected. Moreover, this study showed that bacterial and archaeal communities in PHF were dominated by rare phylotypes which, for the most part, appeared to be endemic to the PHF ecosystem. This study also included the first exhaustive comparison at OTU scale of microbial taxa across six geographically distant serpentinite-hosted ecosystems. Although at this taxonomic resolution, no stricto sensu core microbial community could be defined, a few phylotypes were shared by several sites. These phylotypes, belonging to generally dominant taxa in serpentinizing systems, seem to be specifically associated with unique niches resulting from the peculiar geochemical conditions linked to serpentinization. Indeed, OTUs representing uncultivated Clostridia of the Syntrophomonaceae family (and related to the Dethiobacter alkaliphilus) or of the Thermoanaerobacterales group SRB2 are hypothesized to live in the anoxic, H2 rich end-member fluids or inside the porous carbonates structures, alike the Methanosarcinales (LCMS and TCMS) (Schrenk et al., 2013). Hydrogenophaga OTUs identified in most of the terrestrial serpentinizing sites probably live at oxic-anoxic interfaces where the mixing of alkaline H2-rich fluids and oxygenated waters creates microaerophilic conditions. Upon mixing, steep gradients of pH, temperature and electron donors and acceptors, also provide a range of microhabitats favorable for endemic microbial populations in these extreme ecosystems. Additional factors, abiotic (geological and hydrological settings, water depths, light, etc.) or biotic (species interactions, viruses), superimposed to the serpentinization reactions most likely contributed significantly in shaping the microbial assemblages, thus explaining the limited similarities between ecosystems at the OTU level. This apparent divergence of microbiome composition in serpentinizing systems raised the question of the existence of common metabolic capabilities that could explain the functioning of these ecosystems better. These issues could be addressed in the future by using new genomic tools, such as comparative metagenomics.
Author Contributions
GE designed the study. AP, GE, and MQ performed the field sampling. The experiments were carried out by MB and GE. EF, MB, and FA performed the bioinformatics analyses and interpretations. EF, MB, FA, and GE wrote the manuscript. AP, MQ, BO, and DD reviewed drafts of the paper.
Funding
The scientific cruise HYDROPRONY, during which samples were collected, was funded by the French Institute of Research for Development (IRD). This research was supported by the French National Center of Scientific Research (CNRS/INSU) through the National Program EC2CO-Biohefect/Ecodyn/Dril/MicrobiEn to the MicroProny project. This research was also supported by the deepOASES ANR project (ANR-14-CE01-0008-01). EF and MB were awarded a Ph.D. Fellowship from the French Ministry of Education and Scientific Research.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are grateful to Bernard Pelletier (IRD) and Claude Payri (IRD) for the organization of the HYDROPRONY cruise and providing us access to laboratory facilities at IRD Research Center Nouméa. We thank the scuba divers: Régis Hocdé, Eric Folcher, John Butcher, and Bertrand Bourgeois of the US IMAGO team (IRD Nouméa) for collecting the samples and for providing photos of the chimneys. We are grateful to all members of the crew of the R/V ALIS during the HYDROPRONY cruise for their assistance in sea operations. We also thank Jean Le Mer for his assistance in implementing bioinformatics tools on our dedicated server. We finally thank our reviewers for their constructive criticism and helpful suggestions.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.00102/full#supplementary-material
References
Alfreider, A., Vogt, C., and Babel, W. (2002). Microbial diversity in an in situ reactor system treating monochlorobenzene contaminated groundwater as revealed by 16S ribosomal DNA analysis. Syst. Appl. Microbiol. 25, 232–240. doi: 10.1078/0723-2020-00111
Aller, J. Y., and Kemp, P. F. (2008). Are Archaea inherently less diverse than Bacteria in the same environments? FEMS Microbiol. Ecol. 65, 74–87. doi: 10.1111/j.1574-6941.2008.00498.x
Anderson, R. E., Sogin, M. L., and Baross, J. A. (2015). Biogeography and ecology of the rare and abundant microbial lineages in deep-sea hydrothermal vents. FEMS Microbiol. Ecol. 91, 1–11. doi: 10.1093/femsec/fiu016
Auguet, J. C., Barberan, A., and Casamayor, E. O. (2010). Global ecological patterns in uncultured Archaea. ISME J. 4, 182–190. doi: 10.1038/ismej.2009.109
Ben Aissa, F., Postec, A., Erauso, G., Payri, C., Pelletier, B., Hamdi, M., et al. (2014). Vallitalea pronyensis sp. nov., isolated from a marine alkaline hydrothermal chimney. Int. J. Syst. Evol. Microbiol. 64, 1160–1165. doi: 10.1099/ijs.0.055756-0
Ben Aissa, F., Postec, A., Erauso, G., Payri, C., Pelletier, B., Hamdi, M., et al. (2015). Characterization of Alkaliphilus hydrothermalis sp. nov., a novel alkaliphilic anaerobic bacterium, isolated from a carbonaceous chimney of the Prony hydrothermal field, New Caledonia. Extremophiles 19, 183–188. doi: 10.1007/s00792-014-0697-y
Bes, M., Merrouch, M., Joseph, M., Quéméneur, M., Payri, C., Pelletier, B., et al. (2015). Acetoanaerobium pronyense sp. nov., an anaerobic alkaliphilic bacterium isolated from a carbonate chimney of the Prony hydrothermal field (New Caledonia). Int. J. Syst. Evol. Microbiol. 65, 2574–2580. doi: 10.1099/ijs.0.000307
Biderre-Petit, C., Dugat-Bony, E., Mege, M., Parisot, N., Adrian, L., Moné, A., et al. (2016). Distribution of Dehalococcoidia in the anaerobic deep water of a remote meromictic crater lake and detection of Dehalococcoidia-derived reductive dehalogenase homologous genes. PLOS ONE 11:e0145558. doi: 10.1371/journal.pone.0145558
Bokulich, N. A., Subramanian, S., Faith, J. J., Gevers, D., Gordon, I., Knight, R., et al. (2013). Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 10, 57–59. doi: 10.1038/nmeth.2276
Brazelton, W. J., Ludwig, K. A., Sogin, M. L., Andreishcheva, E. N., Kelley, D. S., Shen, C.-C., et al. (2010). Archaea and bacteria with surprising microdiversity show shifts in dominance over 1,000-year time scales in hydrothermal chimneys. Proc. Natl. Acad. Sci. U.S.A. 107, 1612–1617. doi: 10.1073/pnas.0905369107
Brazelton, W. J., Morrill, P. L., Szponar, N., and Schrenk, M. O. (2013). Bacterial communities associated with subsurface geochemical processes in continental serpentinite springs. Appl. Environ. Microbiol. 79, 3906–3916. doi: 10.1128/AEM.00330-13
Brazelton, W. J., Nelson, B., Schrenk, M. O., Biddle, J. F., Campbell, B. J., and Christner, B. C. (2012). Metagenomic evidence for H2 oxidation and H2 production by serpentinite-hosted subsurface microbial communities. Front. Microbiol. 2:268. doi: 10.3389/fmicb.2011.00268
Brazelton, W. J., Schrenk, M. O., Kelley, D. S., and Baross, J. A. (2006). Methane-and sulfur-metabolizing microbial communities dominate the Lost City Hydrothermal Field ecosystem. Appl. Environ. Microbiol. 72, 6257–6270. doi: 10.1128/AEM.00574-06
Brazelton, W. J., Thornton, C. N., Hyer, A., Twing, K. I., Longino, A. A., Lang, S. Q., et al. (2016). Metagenomic identification of active methanogens and methanotrophs in serpentinite springs of the Voltri Massif, Italy. PeerJ 5:e2945. doi: 10.7717/peerj.2945
Caporaso, J. G., Bittinger, K., Bushman, F. D., Desantis, T. Z., Andersen, G. L., and Knight, R. (2010a). PyNAST: a flexible tool for aligning sequences to a template alignment. Bioinformatics 26, 266–267. doi: 10.1093/bioinformatics/btp636
Caporaso, J. G., Kuczynski, J., Stombaugh, J., Bittinger, K., Bushman, F. D., Costello, E. K., et al. (2010b). QIIME allows analysis of high- throughput community sequencing data. Nat. Methods 7, 335–336. doi: 10.1038/nmeth0510-335
Castelle, C. J., Brown, C. T., Thomas, B. C., Williams, K. H., and Banfield, J. F. (2017). Unusual respiratory capacity and nitrogen metabolism in a Parcubacterium (OD1) of the candidate phyla radiation. Sci. Rep. 7:40101. doi: 10.1038/srep40101
Chavagnac, V., Monnin, C., Ceuleneer, G., Boulart, C., and Hoareau, G. (2013). Characterization of hyperalkaline fluids produced by low-temperature serpentinization of mantle peridotites in the Oman and Ligurian ophiolites. Geochem. Geophys. Geosyst. 14, 2496–2522. doi: 10.1002/ggge.20147
Chun, S.-J., Cui, Y., Ko, S.-R., Lee, H.-G., Oh, H.-M., and Ahn, C.-Y. (2017). Silanimonas algicola sp. nov., isolated from laboratory culture of a bloom-forming cyanobacterium, Microcystis. Int. J. Syst. Evol. Microbiol. 67, k3274–3278. doi: 10.1099/ijsem.0.002102
Crespo-Medina, M., Twing, K. I., Sánchez-Murillo, R., Brazelton, W. J., McCollom, T. M., and Schrenk, M. O. (2017). Methane dynamics in a tropical serpentinizing environment: the Santa Elena Ophiolite, Costa Rica. Front. Microbiol. 8:916. doi: 10.3389/fmicb.2017.00916
Curtis, T. P., and Sloan, W. T. (2005). Exploring microbial diversity - a vast below. Science 309, 1331–1333. doi: 10.1126/science.1118176
Daae, F. L., Økland, I., Dahle, H., Jørgensen, S. L., Thorseth, I. H., and Pedersen, R. B. (2013). Microbial life associated with low-temperature alteration of ultramafic rocks in the Leka ophiolite complex. Geobiology 11, 318–339. doi: 10.1111/gbi.12035
Daffonchio, D., Borin, S., Brusa, T., Brusetti, L., van der Wielen, P. W. J. J., Bolhuis, H., et al. (2006). Stratified prokaryote network in the oxic-anoxic transition of a deep-sea halocline. Nature 440, 203–207. doi: 10.1038/nature04418
Edgar, R. C. (2010). Search and clustering orders of magnitude faster than BLAST. Bioinformatics 26, 2460–2461. doi: 10.1093/bioinformatics/btq461
Edgar, R. C., Haas, B. J., Clemente, J. C., Quince, C., and Knight, R. (2011). UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 27, 2194–2200. doi: 10.1093/bioinformatics/btr381
Eren, A. M., Esen,Ö. C., Quince, C., Vineis, J. H., Morrison, H. G., Sogin, M. L., et al. (2015). Anvi’o: an advanced analysis and visualization platform for ‘omics data. PeerJ 3:e1319. doi: 10.7717/peerj.1319
Escobar-Zepeda, A., Vera-Ponce de León, A., and Sanchez-Flores, A. (2015). The road to metagenomics: from microbiology to DNA sequencing technologies and bioinformatics. Front. Genet. 6:348. doi: 10.3389/fgene.2015.00348
Fernàndez-Guerra, A., and Casamayor, E. O. (2012). Habitat-associated phylogenetic community patterns of microbial ammonia oxidizers. PLOS ONE 7:e47330. doi: 10.1371/journal.pone.0047330
Gouy, M., Guindon, S., and Gascuel, O. (2010). SeaView version 4: a multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 27, 221–224. doi: 10.1093/molbev/msp259
Gubry-rangin, C., Kratsch, C., Williams, T. A., Mchardy, A. C., Embley, T. M., and Prosser, J. I. (2015). Coupling of diversification and pH adaptation during the evolution of terrestrial Thaumarchaeota. Proc. Natl. Acad. Sci. U.S.A. 112, 9370–9375. doi: 10.1073/pnas.1419329112
Hugoni, M., Taib, N., Debroas, D., Domaizon, I., Jouan Dufournel, I., Bronner, G., et al. (2013). Structure of the rare archaeal biosphere and seasonal dynamics of active ecotypes in surface coastal waters. Proc. Natl. Acad. Sci. U.S.A. 110, 6004–6009. doi: 10.1073/pnas.1216863110
Kaster, A.-K., Mayer-Blackwell, K., Pasarelli, B., and Spormann, A. M. (2014). Single cell genomic study of Dehalococcoidetes species from deep-sea sediments of the Peruvian Margin. ISME J. 8, 1–12. doi: 10.1038/ismej.2014.24
Katoh, K., Misawa, K., Kuma, K., and Miyata, T. (2002). MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 30, 3059–3066. doi: 10.1093/nar/gkf436
Kubo, K., Lloyd, K. G., Biddle, J., Amann, R., Teske, A., and Knittel, K. (2012). Archaea of the Miscellaneous Crenarchaeotal Group are abundant, diverse and widespread in marine sediments. ISME J. 6, 1949–1965. doi: 10.1038/ismej.2012.37
Labrenz, M., Lawson, P. A., Tindall, B. J., and Hirsch, P. (2009). Roseibaca ekhonensis gen. nov., sp. nov., an alkalitolerant and aerobic bacteriochlorophyll a-producing alphaproteobacterium from hypersaline Ekho Lake. Int. J. Syst. Evol. Microbiol. 59, 1935–1940. doi: 10.1099/ijs.0.016717-0
Lozupone, C., and Knight, R. (2005). UniFrac: a new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 71, 8228–8235. doi: 10.1128/AEM.71.12.8228
McMurdie, P. J., and Holmes, S. (2013). Phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PLOS ONE 8:e61217. doi: 10.1371/journal.pone.0061217
Mei, N., Postec, A., Monnin, C., Pelletier, B., Payri, C. E., Ménez, B., et al. (2016). Metagenomic and PCR-based diversity surveys of [FeFe]-hydrogenases combined with isolation of alkaliphilic hydrogen-producing bacteria from the serpentinite-hosted Prony Hydrothermal Field, New Caledonia. Front. Microbiol. 7:1301. doi: 10.3389/fmicb.2016.01301
Mei, N., Zergane, N., Postec, A., Erauso, G., Ollier, A., Payri, C., et al. (2014). Fermentative hydrogen production by a new alkaliphilic Clostridium sp. (strain PROH2) isolated from a shallow submarine hydrothermal chimney in Prony Bay, New Caledonia. Int. J. Hydrogen Energy 39, 19465–19473. doi: 10.1016/j.ijhydene.2014.09.111
Monnin, C., Chavagnac, V., Boulart, C., Ménez, B., Gérard, M., Gérard, E., et al. (2014). Fluid chemistry of the low temperature hyperalkaline hydrothermal system of Prony bay (New Caledonia). Biogeosciences 11, 5687–5706. doi: 10.5194/bg-11-5687-2014
Nelson, W. C., and Stegen, J. C. (2015). The reduced genomes of Parcubacteria (OD1) contain signatures of a symbiotic lifestyle. Front. Microbiol. 6:713. doi: 10.3389/fmicb.2015.00713
Neubeck, A., Sun, L., Müller, B., Ivarsson, M., Hosgörmez, H., Özcan, D., et al. (2017). Microbial community structure of a serpentine-hosted abiotic gas seepage at the Chimaera ophiolite, Turkey. Appl. Environ. Microbiol. 83:e03430-16. doi: 10.1128/AEM.03430-16
Pelletier, B., Chevillon, C., Menou, J., Butscher, J., Folcher, E., Geoffray, C., et al. (2006). Plongées, forage et cartographie Baie du Prony et Banc Gail, lagon Sud de Nouvelle-Calédonie, campagne 2005-NC-PL du N.O. ALIS 13-17 Juin 2005 et cartographie baie du Prony et canal Woodin N.O. ALIS 25-26 September 2005: Nouméa IRD Sept. 2006. Mission. Sci. Terre Géol. Géophys. 70, 1–44.
Pisapia, C., Gérard, E., Gérard, M., Lecourt, L., Lang, S. Q., Pelletier, B., et al. (2017). Mineralizing filamentous bacteria from the Prony Bay Hydrothermal Field give new insights into the functioning of serpentinization-based subseafloor ecosystems. Front. Microbiol. 8:57. doi: 10.3389/fmicb.2017.00057
Poser, A., Lohmayer, R., Vogt, C., Knoeller, K., Planer-Friedrich, B., Sorokin, D., et al. (2013). Disproportionation of elemental sulfur by haloalkaliphilic bacteria from soda lakes. Extremophiles 17, 1003–1012. doi: 10.1007/s00792-013-0582-0
Postec, A., Quéméneur, M., Bes, M., Mei, N., Benaïssa, F., Payri, C., et al. (2015). Microbial diversity in a submarine carbonate edifice from the serpentinizing hydrothermal system of the Prony Bay (New Caledonia) over a 6-year period. Front. Microbiol. 6:857. doi: 10.3389/fmicb.2015.00857
Quast, C., Pruesse, E., Yilmaz, P., Gerken, J., Schweer, T., Yarza, P., et al. (2013). The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. 41, 590–596. doi: 10.1093/nar/gks1219
Quéméneur, M., Bes, M., Postec, A., Mei, N., Hamelin, J., Monnin, C., et al. (2014). Spatial distribution of microbial communities in the shallow submarine alkaline hydrothermal field of the Prony Bay, New Caledonia. Environ. Microbiol. Rep. 6, 665–674. doi: 10.1111/1758-2229.12184
Quéméneur, M., Palvadeau, A., Postec, A., Monnin, C., Chavagnac, V., Ollivier, B., et al. (2015). Endolithic microbial communities in carbonate precipitates from serpentinite-hosted hyperalkaline springs of the Voltri Massif (Ligurian Alps, Northern Italy). Environ. Sci. Pollut. Res. 22, 13613–13624. doi: 10.1007/s11356-015-4113-7
Rempfert, K. R., Miller, H. M., Bompard, N., Nothaft, D., Matte, J. M., Kelemen, P., et al. (2017). Geological and geochemical controls on subsurface microbial life in the Samail Ophiolite, Oman. Front. Microbiol. 8:56. doi: 10.3389/fmicb.2017.00056
Russell, M. J., Hall, A. J., and Martin, W. (2010). Serpentinization as a source of energy at the origin of life. Geobiology 8, 355–371. doi: 10.1111/j.1472-4669.2010.00249.x
Sánchez-Murillo, R., Gazel, E., Schwarzenbach, E. M., Crespo-Medina, M., Schrenk, M. O., Boll, J., et al. (2014). Geochemical evidence for active tropical serpentinization in the Santa Elena Ophiolite, Costa Rica: an analog of a humid early Earth? Geochem. Geophys. Geosyst. 15, 1783–1800. doi: 10.1002/2013GC005213
Schrenk, M. O., Brazelton, W. J., and Lang, S. Q. (2013). Serpentinization, carbon, and deep life. Rev. Mineral. Geochem. 75, 575–606. doi: 10.2138/rmg.2013.75.18
Schrenk, M. O., Kelley, D. S., Bolton, S. A., and Baross, J. A. (2004). Low archaeal diversity linked to subseafloor geochemical processes at the Lost City Hydrothermal Field, Mid-Atlantic Ridge. Environ. Microbiol. 6, 1086–1095. doi: 10.1111/j.1462-2920.2004.00650.x
Shannon, C. E., and Weaver, W. (1949). The Mathematical Theory of Communication. Urbana, IL: University of Illinois Press.
Sobieraj, M., and Boone, D. R. (2006). “Syntrophomonadaceae,” in The Prokaryotes: Bacteria: Firmicutes, Cyanobacteria, Vol. 4, eds M. Dworkin, S. Falkow, E. Rosenberg, K.-H. Schleifer, and E. Stackebrandt (New York, NY: Springer), 1041–1049.
Sorokin, D. Y., Abbas, B., Geleijnse, M., Kolganova, T. V., Kleerebezem, R., and van Loosdrecht, M. C. (2016). Syntrophic associations from hypersaline soda lakes converting organic acids and alcohols to methane at extremely haloalkaline conditions. Environ. Microbiol. 18, 3189–3202. doi: 10.1111/1462-2920.13448
Sorokin, D. Y., Tourova, T. P., Mußmann, M., and Muyzer, G. (2008). Dethiobacter alkaliphilus gen. nov. sp. nov., and Desulfurivibrio alkaliphilus gen. nov. sp. nov.: two novel representatives of reductive sulfur cycle from soda lakes. Extremophiles 12, 431–439. doi: 10.1007/s00792-008-0148-8
Suzuki, S., Gijs Kuenen, J., Schipper, K., Van Der Velde, S., Ishii, S., Wu, A., et al. (2014). Physiological and genomic features of highly alkaliphilic hydrogen-utilizing Betaproteobacteria from a continental serpentinizing site. Nat. Commun. 5:3900. doi: 10.1038/ncomms4900
Suzuki, S., Ishii, S., Hoshino, T., Rietze, A., Tenney, A., Morrill, P. L., et al. (2017). Unusual metabolic diversity of hyperalkaliphilic microbial communities associated with subterranean serpentinization at The Cedars. ISME J. 11, 2584–2598. doi: 10.1038/ismej.2017.111
Suzuki, S., Ishii, S., Wu, A., Cheung, A., Tenney, A., Wanger, G., et al. (2013). Microbial diversity in The Cedars, an ultrabasic, ultrareducing, and low salinity serpentinizing ecosystem. Proc. Natl. Acad. Sci. U.S.A. 110, 15336–15341. doi: 10.1073/pnas.1302426110
Tiago, I., and Veríssimo, A. (2013). Microbial and functional diversity of a subterrestrial high pH groundwater associated to serpentinization. Environ. Microbiol. 15, 1687–1706. doi: 10.1111/1462-2920.12034
Twing, K. I., Brazelton, W. J., Kubo, M. D. Y., Hyer, A. J., Cardace, D., Hoehler, T., et al. (2017). Serpentinization-influenced groundwater harbors extremely low diversity microbial communities adapted to high pH. Front. Microbiol. 8:308. doi: 10.3389/FMICB.2017.00308
Wang, Q., Garrity, G. M., Tiedje, J. M., and Cole, J. R. (2007). Naïve Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 73, 5261–5267. doi: 10.1128/AEM.00062-07
Wasmund, K., Schreiber, L., Lloyd, K. G., Petersen, D. G., Schramm, A., Stepanauskas, R., et al. (2013). Genome sequencing of a single cell of the widely distributed marine subsurface Dehalococcoidia, phylum Chloroflexi. ISME J. 8, 383–397. doi: 10.1038/ismej.2013.143
Willems, A. (2014). “The family Comamonadaceae,” in The Prokaryotes: Alphaproteobacteria and Betaproteobacteria, eds E. Rosenberg, E. F. DeLong, S. Lory, E. Stackebrandt, and F. Thompson (Berlin: Springer), 777–851. doi: 10.1007/978-3-642-30197-1_238
Woycheese, K. M., Meyer-Dombard, D. R., Cardace, D., Argayosa, A. M., and Arcilla, C. A. (2015). Out of the dark: transitional subsurface-to-surface microbial diversity in a terrestrial serpentinizing seep (Manleluag, Pangasinan, the Philippines). Front. Microbiol. 6:44. doi: 10.3389/fmicb.2015.00044
Keywords: microbial communities, prony, shallow hydrothermal field, alkaliphiles, methanosarcinales, serpentinization
Citation: Frouin E, Bes M, Ollivier B, Quéméneur M, Postec A, Debroas D, Armougom F and Erauso G (2018) Diversity of Rare and Abundant Prokaryotic Phylotypes in the Prony Hydrothermal Field and Comparison with Other Serpentinite-Hosted Ecosystems. Front. Microbiol. 9:102. doi: 10.3389/fmicb.2018.00102
Received: 12 October 2017; Accepted: 17 January 2018;
Published: 06 February 2018.
Edited by:
Anna-Louise Reysenbach, Portland State University, United StatesReviewed by:
Donnabella Castillo Lacap-Bugler, Auckland University of Technology, New ZealandWilliam Brazelton, East Carolina University, United States
Copyright © 2018 Frouin, Bes, Ollivier, Quéméneur, Postec, Debroas, Armougom and Erauso. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gaël Erauso, gael.erauso@mio.osupytheas.fr