 Roberto Arsiè
Roberto Arsiè Gian G. Tartaglia2
Gian G. Tartaglia2 R. Martin Vabulas
R. Martin Vabulas- 1 Charité-Universitätsmedizin Berlin, Institute of Biochemistry, Berlin, Germany
- 2 Centre for Human Technologies (CHT), RNA System Biology Lab, Istituto Italiano di Technologia, Genoa, Italy
Ribonucleotide modifications modulate RNA structure and stability, thereby influencing RNA turnover and a wide range of cellular functions. Recent studies have revealed that specific RNA modifications can also affect the formation, composition, and material properties of biomolecular condensates. This review explores how N1-methyladenine (m1A) and N6-methyladenine (m6A) contribute to RNA-driven phase transitions and the balance between adaptive granulation and pathological protein aggregation. m1A can act as a protective tag: by altering local RNA structure and RNA-protein interactions, it promotes the sequestration of selected transcripts into dynamic stress granules and facilitates the resumption of protein synthesis after stress. During chronic proteotoxicity, m1A helps prevent aberrant RNA-protein entanglement. However, when present on pathogenic repeat RNAs, m1A can recruit aggregation-prone proteins and exacerbate pathology. On the other hand, m6A functions as both a structural switch and a multivalent docking signal. Multiple m6A sites enhance the binding of cognate reader proteins to a transcript, thereby accelerating stress granule assembly. m6A modification has also been implicated in organizing nuclear condensates such as HSATIII lncRNA assemblies. We discuss mechanistic models that aim to reconcile the diverse roles of methyladenine, highlight current experimental challenges, and outline emerging approaches for addressing the remaining open questions.
1 Introduction
Eukaryotic cells employ diverse strategies to achieve spatial and temporal organization. Under specific biochemical conditions, subsets of cellular proteins and nucleic acids undergo liquid-liquid phase separation (LLPS) from the surrounding cytoplasm to form biomolecular condensates (Zacco et al., 2024). These micron-scale assemblies are integral component of eukaryotic cell organization, regulating key processes such as stress adaptation and RNA metabolism (Banani et al., 2017; Shin and Brangwynne, 2017; Sabari et al., 2020; Alberti and Hyman, 2021; Gorsheneva et al., 2024). Unlike membrane-bound organelles, biocondensates are dynamic entities that can assemble and dissemble reversibly in response to environment cues. Their formation depends on the intrinsic biophysical properties of constituent proteins and RNAs, which drive demixing from the surrounding aqueous milieu. These biocondensates function as subcellular hubs that concentrate specific polypeptides and transcripts (Hirose et al., 2023), thereby coordinating numerous cellular processes in a spatiotemporally-controlled, membrane-independent manner. Stress granules (SG) and processing bodies (PB) are two examples of cytosolic ribonucleoprotein assemblies formed by LLPS under physiological conditions or in response to stressors such as heat shock and arsenite exposure (Luo et al., 2018; Protter et al., 2018). In the nucleus, nuclear stress bodies (nSB) represent analogous membraneless organelles that assemble upon thermal stress.
The assembly of biocondensates depends on weak, multivalent interactions among their molecular constituents. In particular, proteins containing intrinsically disordered regions (IDR) or low-complexity domains (LCD) are especially prone to form granular structures, as they serve as scaffolds for intra- and intermolecular interactions (Shin and Brangwynne, 2017; Martin and Mittag, 2018; Vodnala et al., 2021). RNA also plays a central role in biomolecular condensate formation: as a flexible and multivalent polymer, it can engage in numerous interactions with diverse proteins and other RNA molecules (Zhang et al., 2015; Saha et al., 2016; Roden and Gladfelter, 2021; Ripin and Parker, 2023). This intricate network of dynamic interactions enables cells to rapidly assemble and disassemble condensates in response to changing physiological conditions (Cairo et al., 2024; Zhang et al., 2025).
While dynamic and reversible LLPS is essential for cellular homeostasis, its dysregulation can lead to pathological consequences (Zacco et al., 2024). A critical distinction exists between the reversible formation of liquid-like condensates and their conversion into static, irreversible fibrillar aggregates - a hallmark of numerous diseases (Molliex et al., 2015; Alberti and Dormann, 2019; Mathieu et al., 2020). For example, RNA-binding proteins such as TDP-43 and FUS normally form dynamic liquid droplets, but in neurodegenerative conditions like amyotrophic lateral sclerosis (ALS) and frontotemporal dementia (FTD), they accumulate as insoluble fibrillar inclusions (Patel et al., 2015; Carey and Guo, 2022). Therefore, understanding the molecular switches that govern these liquid-to-solid transitions may be crucial for developing potential therapeutic strategies.
Among the various biochemical and environmental factors that drive biocondensate formation, a growing body of evidence indicates the role of certain RNA modifications, specifically, N6-methyladenine (m6A) and N1-mehtyladenine (m1A). These modifications are installed by specific enzymes (“writers”), recognized by cognate “readers”, and removed by dedicated demethylases (“erasers”) - a regulatory machinery that enables dynamic adjustment of methyladenine abundance and its functional activity.
2 Methods for m1A and m6A detection
A wide range of methods have been developed to detect and quantify RNA modifications. One common approach relies on commercially available antibodies that recognize m1A and m6A in RNAs. Typically, these antibodies are used to enrich transcripts carrying the modification, after which the immunoprecipitated RNAs are subjected to next-generation sequencing to map the modified sites (Dominissini et al., 2012; 2016; Meyer et al., 2012; Li et al., 2016). These approaches enable transcriptome-wide mapping of the selected modification but suffer from low resolution (approximately 100 base pairs). Inevitably, the results are influenced by antibody specificity and cross-reactivity (Schwartz, 2018; Grozhik et al., 2019). Cross-linking or reverse-transcription-based variants inducing mutational signatures increase the resolution, but still rely on immunoprecipitation (Linder et al., 2015; Li et al., 2017; Safra et al., 2017; Grozhik et al., 2019; Koh et al., 2019; Roberts et al., 2021).
Chemical or enzymatic conversion approaches do not require antibodies. These methods exploit the fact that certain RNA modifications alter the activity of nucleases, deaminases, or reverse transcriptases, thereby generating characteristic sequence “signatures” that can be detected through high-throughput sequencing. For m6A detection, several antibody-independent protocols have been developed, such as MAZTER-seq, in which the activity of the endoribonuclease MazF is blocked by m6A (Garcia-Campos et al., 2019), and DART-seq, where an engineered APOBEC enzyme edits nucleotides near m6A sites (Meyer, 2019). These methods provide improved resolution without the need for antibodies; however, they require engineered enzymes and yield motif-restricted results. For m1A detection, methods take advantage of its ability to affect the fidelity of reverse transcription, leading to characteristic mutation signatures (Zhou et al., 2019). A major challenge, however, lies in reliably detecting these misincorporation events (Sas-Chen and Schwartz, 2019). Controls often include AlkB-mediated demethylation or the Dimroth rearrangement. The latter–chemical conversion of m1A to m6A under alkaline and heat conditions - poses a significant analytical challenge for accurate RNA modification mapping and quantification. If the rearrangement occurs inadvertently during RNA handling or library preparation, m1A residues may be misclassified as m6A. This artifact is particularly problematic for antibody-based m6A mapping approaches, which cannot distinguish between endogenous m6A and rearranged m1A. As a result, false-positive m6A sites may be reported, distorting modification maps and complicating downstream biological interpretation.
Thin-layer chromatography (TLC) and liquid chromatography-tandem mass spectrometry (LC-MS/MS) remain the gold standards for global quantification of RNA modifications (Grosjean et al., 2004; Herbert et al., 2024). These approaches provide absolute stoichiometries and enable the simultaneous detection of multiple modifications. However, they offer little or no positional information and require relatively large amounts of input RNA, making them well suited for studying abundant RNA species (Taoka et al., 2018; Suzuki et al., 2020).
Both m6A and m1A can, in principle, be detected through alterations in the ionic current during Nanopore-direct RNA sequencing (Leger et al., 2021). When combined with machine learning-based models, this approach enables antibody-free, single-molecule detection. While promising, the relatively low signal-to-noise ratio remains a major challenge, and extensive validation is still required.
3 Computational approaches for mapping and predicting m1A and m6A
Alongside experimental mapping, bioinformatics has become central to detecting and interpreting RNA methylation marks. For both m6A and m1A, antibody-based enrichment methods (MeRIP-seq, m1A-seq) generate peaks that require dedicated computational pipelines for analysis. Peak callers such as exomePeak and MACS2 have been widely used to analyse m6A data (Dominissini et al., 2012; Meyer et al., 2012; Linder et al., 2015), while improved frameworks such as exomePeak2 and MeTPeak provide bias correction (Cao et al., 2014; Cui et al., 2016) and statistical modeling. For m1A, computational workflows must also account for potential Dimroth rearrangement to m6A, which complicates the interpretation of sequencing data (Safra et al., 2017; Zhou et al., 2019). Chemical conversion or mutational signature-based approaches provide single-nucleotide resolution but yield heterogeneous misincorporation patterns that require careful computational filtering (Grozhik et al., 2019). More recently, direct RNA Nanopore sequencing combined with machine learning-based classifiers has enabled antibody-free identification of m6A and m1A at single-molecule resolution (Leger et al., 2021). Such computational frameworks are essential for distinguishing true modification signatures from sequencing noise and for integrating orthogonal validation data, such as LC–MS/MS measurements.
In parallel, machine learning and modeling tools can be leveraged to predict methylation sites de novo and assess their structural and functional impacts. For m6A, tools such as SRAMP (Zhou Y. et al., 2016) and DeepM6ASeq (Zhang and Hamada, 2018) combine sequence motifs like DRACH with structural context (e.g., predicted accessibility, pairing propensity). On the m1A front, iRNA-3typeA (Chen et al., 2018) and ISGm1A (Liu et al., 2020) use feature sets derived from k-mer frequencies, nucleotide physicochemical properties, and secondary structure proxies to score candidate adenines. To capture how modifications may alter structure or binding energetics, computational tools such as ViennaRNA have been extended to support folding analyses that include parameters for modified bases, allowing comparison of RNAs containing m6A or m1A (Varenyk et al., 2023). Additionally, catRAPID has incorporated m6A information to estimate how this modification changes RNA–protein binding propensities (Vandelli et al., 2024). Together, these computational tools enable researchers to move from raw sequencing reads to predicted methylomes, and from there to hypotheses about how m6A and m1A influence RNA folding, protein binding, and ultimately condensate dynamics.
4 m6A as a phase separation enhancer
N6-methyladenine (m6A) is the most abundant modification found on eukaryotic mRNA (Desrosiers et al., 1974; Dubin and Taylor, 1975). m6A is installed on RNAs by a multi-protein methyltransferase complex: the catalytic METTL3 subunit pairs with METTL14 to form a stable heterodimer, and WTAP targets this complex to RNAs (Liu et al., 2014; Wang et al., 2016). The m6A mark is reversible. FTO (fat mass and obesity-associated protein) was the first enzyme shown to demethylate m6A (Jia et al., 2011), and ALKBH5 later was identified as a second m6A demethylase affecting mRNA export and metabolism in mouse germ cells (Zheng et al., 2013). Together, these erasers make m6A a dynamic and reversible modification. m6A is recognized by specific reader proteins, most notably member of the YTH-domain family, such as YTHDF1-3 and YTHDC1 (Zhu et al., 2014; Zaccara and Jaffrey, 2020). Biochemically, m6A alters RNA duplex stability and can locally “open” RNA structure, favoring a single-strand conformation, which could influence binding of other factors (Liu et al., 2015; Roost et al., 2015). Functionally, m6A impacts nearly every step of mRNA metabolism. Early transcriptome-wide maps revealed that m6A is enriched near stop codons and within long internal exons, where it influences RNA splicing (Dominissini et al., 2012). Binding of nuclear m6A-marked pre-mRNAs by YTHDC1 was also shown to modulate splicing (Xiao et al., 2016). YTHDF2 binding to m6A generally decreases mRNA stability, thereby shortening the half-life of its targets (Wang et al., 2014; Du et al., 2016). In contrast, YTHDF1 binds m6A-modified mRNAs to enhance their translation efficiency (Wang et al., 2015).
Recent studies have shown that m6A modifications on mRNAs can enhance their capacity for phase separation, together with their associated m6A reader proteins. For example, Ignatova’s research group demonstrated that during oxidative stress induced by arsenite exposure, the total m6A level increased in U2OS cells, with approximately 8% more mRNAs found to be methylated (Anders et al., 2018). The modification colocalized with stress granules also upon heat shock. During stress, the post-transcriptional methylation was enriched in the 5′ UTRs of mRNAs. The m6A reader YTHDF3 bound to these stress-modified RNAs and relocated with them to stress granules. Removed from the translating pool and triaged into stress granules, m6A-labelled mRNAs seem to be dynamically regulated (Figure 1A).
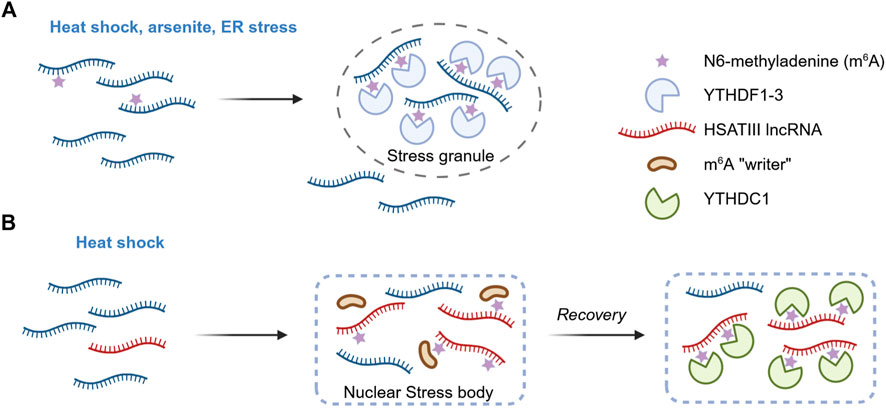
Figure 1. m6A as a regulator of biomolecular condensation. (A) m6A influences stress granule formation. In response to stress, the assembly of stress granules is promoted by multiple m6A modifications on RNAs and the subsequent recruitment of the reader proteins YTHDF1-3. Length-dependent transcript enrichment in SG is also mediated by m6A. However, the contribution of m6A to stress granule dynamics appears to be context-dependent, with some studies reporting a strong effect on granule assembly and others indicating only a modest influence (not shown). (B) m6A in nuclear stress bodies. Following heat shock, the pericentromeric lncRNA HSATIII is transcribed and drives the assembly of nuclear stress bodies. Components of the m6A methyltransferase complex are recruited, leading to post-transcriptional modification of HSATIII. Upon stress recovery, the m6A reader YTHDC1 is sequestered at nuclear stress bodies, leading to its transient depletion from the surrounding nucleoplasm and altering the splicing patterns of target pre-mRNAs.
Several studies have further demonstrated that the m6A reader proteins YTHDF1, YTHDF2, and YTHDF3 undergo liquid-liquid phase separation (LLPS) both in vitro and in vivo, and that this process is markedly enhanced in the presence of m6A-modified RNA (Gao et al., 2019; Ries et al., 2019; Fu and Zhuang, 2020). Multivalence is critical, as multiple m6A modifications were required to cluster sufficient YTHDF molecules to nucleate condensates. Various environmental stressors - such as heat shock, arsenite exposure, endoplasmic reticulum stress - promoted the relocalization of these proteins and the associated m6A-containing RNAs from the cytosol to stress granules. Interestingly, knockout of METTL14 did not affect the formation of stress granules or P-bodies, but the redistribution of YTHDF2 within these compartments was greatly reduced (Ries et al., 2019). This finding indicates that m6A reader proteins must bind modified RNAs in order to move to membraneless compartments (Figure 1A). The LLPS of YTHDF2 was enhanced by m6A under stressful conditions (Man et al., 2025), whereas mutations in its m6A-binding domain impaired granule formation in vivo (Wang et al., 2020). Noteably, m6A has also been shown to govern the preferential enrichment of long mRNAs within stress granules (Ries et al., 2023).
These findings were supported by another study in which the authors implemented a FRET-based method to track m6A-modified RNAs and their interactions with reader proteins (Li et al., 2024). They showed that m6A-marked transcripts rapidly accumulated in stress granules during stress and relocated back to the cytosol during the recovery phase. Knockdown of YTHDF2 accelerated granule disassembly, enabling faster mRNA release and translation recovery.
Challenging the findings described above, the laboratory of Roy Parker questioned the functional significance of m6A in stress granules formation (Khong et al., 2022). In their study, knockout of the m6A writer METTL3 did not alter the recruitment of several poly-m6A-modified mRNAs to arsenite induced stress granules, with m6A status explaining only 6% of the variance in RNA partitioning into these granules. Furthermore, in an artificial system designed to enhance reader protein interaction, 25 copies of YTHDF were tethered to a single transcript, yet no significant increase in RNA recruitment to granules was observed. Aside from the non-physiological nature of tethering 25 proteins to one transcript and correlations drawn from datasets across different studies and cell lines, the apparent discrepancy may also reflect dependencies on stress conditions, cell-types specificities, or yet-to-be-identified compensatory mechanisms.
m6A has also recently been implicated in the regulation of membraneless organelle in the nucleus. During thermal stress, the long non-coding RNA (lncRNA) HSATIII is expressed and contributes to the formation of nuclear stress bodies (nSB) (Jolly et al., 2004; Biamonti and Vourc’h, 2010). Ninomiya and colleagues showed that HSTAIII sequence is highly m6A-modified and that this modification sequestered m6A regulatory factors into the condensate (Ninomiya et al., 2021). Using comprehensive RNA-protein interaction proteomics (m6A-RIP-seq and LC-MS/MS on purified HSATIII fragments), the authors found that approximately 11% of the first adenine in the GGAAU repeat unit of HSATIII is methylated at position 6, and that the m6A writer complex is recruited into nSBs during stress recovery. Furthermore, m6A-modified HSATIII sequestered the nuclear m6A reader YTHDC1 within nSBs during stress recovery, thereby lowering its nucleoplasmic concentration. Since YTHDC1 is known to promote m6A-dependent splicing, its depletion lead to increased intron retention in a subset of m6A-marked pre-mRNAs. This work therefore unveiled a mechanistic link between an m6A-modified structural lncRNA, biocondensation, and genome-wide regulation of temperature-dependent splicing. HSATIII appears to act both as a “reaction crucible” that promotes local biochemical events and as a “molecular sponge” that titrates m6A machinery proteins (Figure 1B).
5 m1A in stress granules and protein aggregates
N1-methyladenine (m1A) was first discovered in non-coding RNA in the 1960s, specifically at position 58 of tRNAs and within the large subunit of ribosomal RNA (Dunn, 1961; Shima and Igarashi, 2020). This post-transcriptional modification is conserved across Bacteria, Archaea and Eukaryotes. In eukaryotes, m1A is installed by several methyltransferases. The TRMT6-TRMT61A complex acts in the cytoplasm, modifying both tRNAs and a subset of mRNAs that contain a conserved GUUCRA motif reminiscent of the tRNA TΨC loop. The homologous enzymes TRMT61B and TRMT10C are active in mitochondria and modify mitochondrial tRNAs and rRNAs there (Chujo and Suzuki, 2012; Vilardo et al., 2012; Bar-Yaacov et al., 2016). In the nucleolus, RRP8/NML creates the m1A1322 mark in 28S rRNA (Waku et al., 2016). m1A is a reversible modification: mammalian FTO and the AlkB homologs ALKBH1, ALKBH3 can demethylate m1A in RNA indicating dynamic regulation. Among these, ALKBH3 is the main demethylase acting on mRNAs.
Structurally, the addition of a methyl group at position 1 on the adenine base disrupts its ability to form standard hydrogen bonds and imparts a positive charge on it, if at physiological pH. This alteration affects the secondary and tertiary structures of the modified RNA, strongly influencing its intra- and intermolecular interactions. In mRNA, m1A exerts context- and position-dependent effects on transcripts’ translation and stability. One of the first m1A mapping studies revealed that m1A in 5′UTRs, especially in the proximity to the cap, correlates with increased translation efficiency (Li et al., 2017). In contrast, the same study showed that m1A within coding sequences tends to inhibit translation, likely by blocking codon-anticodon pairing. Several m1A “readers” have been identified among cytoplasmic YTH domain-containing proteins. For instance, YTHDF3 was found to specifically bind m1A sites in IGF1R mRNA and recruit RNA decay machinery, thereby downregulating that transcript (Zheng et al., 2020). Thus, m1A appears to function as a molecular switch that can either promote translation (when located in 5′UTR) or mark transcripts for degradation (through YTHDF binding), linking methylation to mRNA stability, localization, and translation control.
m1A is also a stress-related RNA modification, and growing evidence suggests its involvement in biomolecular condensation. A recent analysis uncovered that stress granule (SG)-enriched transcripts are statistically more likely to contain the m1A consensus motif GUUCRA, indicating a function link between m1A and granule localization (Alriquet et al., 2020). The study further showed that the m1A-installing TRMT6/TRMT61A complex accumulates in stress granules upon heat shock or arsenite treatment in HeLa cells. LC-MS/MS analyses demonstrated that SG-associated RNA contains significantly higher amounts of m1A compared to bulk cytosolic mRNA. The authors engineered a reporter mRNA harboring a single m1A motif in the 5′UTR and compared it with a non-methylatable mutant. Under normal conditions, both transcripts exhibited comparable translation efficiency. However, during heat shock, translation of the wild-type reporter was repressed more rapidly than that of the control. Upon recovery, translation from the m1A-containing reporter resumed more quickly, producing substantially higher protein levels than the non-methylatable counterpart (Figure 2A).
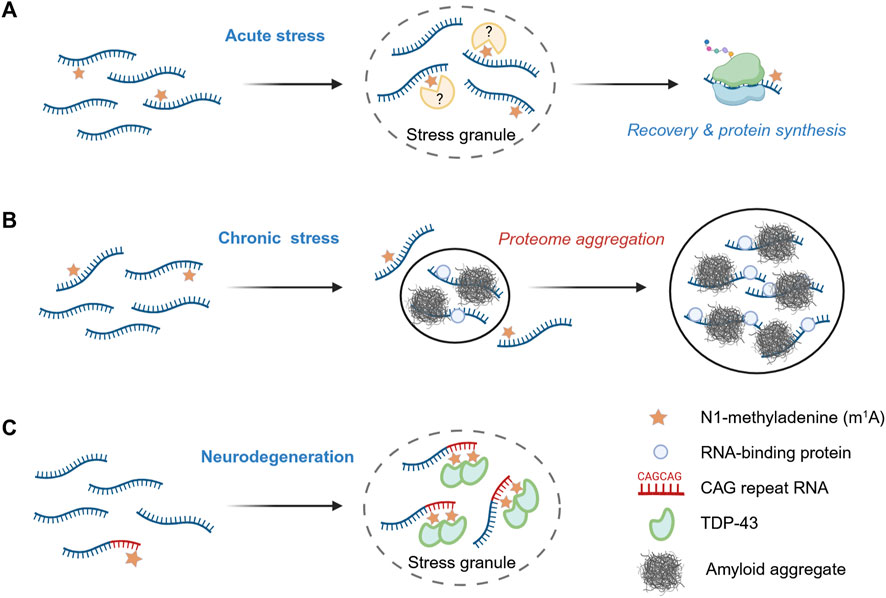
Figure 2. m1A as a regulator of biomolecular condensation. (A) Acute stress response. During acute stress, m1A-modified mRNA are preferentially sequestered into stress granules, and, upon stress release, are translated more efficiently than their non-methylated counterparts. (B) Chronic stress conditions. Under prolonged stress, such as during amyloidogenesis, reduced m1A levels promote protein aggregation and enhance the recruitment of bystander RNA-binding proteins. (C) m1A -modified CAG repeat RNAs and TDP-43 recruitment. Ectopic expression of RNAs containing expanded CAG repeats promotes stress granule assembly and leads to extensive m1A modification. This modification facilitates the relocalization of the RNA-binding protein TDP-43 from the nucleus to cytosolic stress granules, contributing to aberrant aggregation events associated with neurodegeneration.
These results indicate an active safeguarding role of m1A during acute stress. Subsequent work extended this protective function to conditions of chronic proteotoxic stress, suggesting a general role of m1A in maintaining cellular homeostasis (Alriquet et al., 2025). Using a reporter system similar to that described above, the authors demonstrated that protein translation from methylation-deficient transcripts is impaired during amyloidogenesis. Forced reduction of m1A tagging–either by knockdown of the methyltransferase subunit TRMT61A or by orthogonal upregulation of the m1A demethylase ALKBH3 - led to increased amyloidogenesis in HeLa cells. Proteomic analysis of aggregates from m1A-deficient cells revealed a threefold increase in the co-aggregation of amyloid with bystander proteins, particularly with RNA-binding proteins (RBPs). Amyloid was more strongly associated with mRNA under these conditions. Together, these findings support a model in which m1A protects transcripts from entanglement with misfolded proteins, thereby limiting the RNA-driven propagation of protein aggregation during amyloidogenesis (Figure 2B).
A recent study by the Wang laboratory uncovered specific circumstance in which m1A can have the opposite effect, enhancing protein aggregation (Sun et al., 2023). The authors observed an accumulation of m1A in ectopically expressed CAG repeat RNAs, which are characteristic of several hereditary neurodegenerative disorders, including Huntington’s disease, spinocerebellar ataxia, and amyotrophic lateral sclerosis. Longer CAG tracts resulted in a higher level of adenine methylation and correlated with lower expression of the demethylase ALKBH3, suggesting a potential role of N1-methyladenine in the pathogenesis of triplet-repeat expansion diseases. Elevated m1A levels were also associated with enhanced neurodegeneration in C. elegans and Drosophila. The authors showed that m1A is recognized by the two RNA recognition motif (RRM) domains of TDP-43, a protein known to mislocalize and aggregate in the cytosol during neurodegeneration (Dai et al., 2018). Consistently, the authors observed the mislocalization and aggregation of TDP-43 upon ectopic expression of CAG repeat RNA in U2OS cells. In the cytosol, TDP-43 and CAG repeat RNAs co-localized with the stress granule marker G3BP1, and this association was reduced upon ALKBH3 overexpression (Figure 2C). Long CAG repeats with multiple m1A modifications altered the biophysical properties of TDP-43 condensates, shifting their phase behavior toward less soluble, amyloid-like aggregates. These findings highlight the toxic potential of m1A as a pathological enhancer of RNA-protein condensation, whereby this RNA modification is hijacked by expanded CAG repeats to promote aggregate formation and neurodegeneration. Gaining a precise mechanistic understanding of how m1A exerts protective versus toxic effects will require experimental advances to identify the structural changes that precede these divergent cellular outcomes.
6 Discussion
It seems surprising that the addition of a single methyl group can so strongly influence the intracellular behavior of a much larger mRNA molecule. At least two, potentially interconnected, explanations can be proposed: (1) methylation-induced alterations in the global RNA architecture, and (2) localized changes in the bonding capacity of methyladenine. As noted previously, the addition of a methyl group at the N1 position of adenine imparts a positive charge and prevents both Watson–Crick and Hoogsteen base pairing, thereby profoundly influencing intra- and intermolecular associations as well as the formation of structural motifs such as RNA hairpins (Micura et al., 2001; Zhou H. et al., 2016). These structural perturbations may diminish an mRNA’s ability to engage stably with RNA-binding proteins or other RNAs. This view is consistent with the accumulating evidence that nucleic acid structure can modulate, and in some cases promote, protein aggregation both in vitro and in vivo.
Despite significant recent advances in the field, several other important questions remain open. First, combinatorial effects of multiple modifications on the same transcript are still largely unexplored. Do m6A and m1A act synergistically or antagonistically in regulating condensate dynamics? Second, the kinetics of writer and eraser enzyme localization to granules under acute as opposed to chronic stress require real-time analysis, potentially through live-cell Nanopore seqencing (Leger et al., 2021) or super-resolution imaging. Third, methyladenine reader proteins beyond the YTH and YBX families might be involved in interpretation of m1A and m6A within biocondensates and should be systemically identified using proximity-labelling and crosslinking approaches. Fourth, elucidating how these modifications reshape RNA structure in situ and modulate the material properties of granules and aggregates will require advanced techniques such as cryo-electron microscopy (cryo-EM) and single-molecule FRET in future studies.
Therapeutically, manipulating RNA modifications offers a promising avenue to modulate condensate behavior in neurodegenerative and other protein-aggregation diseases. For example, small-molecule inhibitors of TRMT61A or activators of ALKBH3 could mitigate pathological m1A-dependent TDP-43 aggregation in CAG repeat disorders (Sun et al., 2023). Likewise, allosteric regulators of METTL3 or FTO might adjust m6A levels to restore adaptive stress granule dynamics without impairing normal mRNA metabolism. Finally, CRISPR-based epitranscriptomic editing tools could enable locus-specific installation or removal of marks to dissect causality and develop precision therapeutics. As our toolbox expands, combining high-resolution mapping, engineered writer/eraser recruitment, and phase separation biosensors, the field is poised to translate fundamental insights into interventions that restore cellular homeostasis by fine-tuning the epitranscriptomic regulation of phase transitions.
In summary, RNA modifications have emerged as powerful regulators of biomolecular condensates, influencing both physiological granule formation and pathological aggregation. The reversible deposition of m6A and m1A on mRNA fine-tunes multivalent RNA-protein interactions to either promote dynamic liquid-liquid phase separation or, under dysregulated conditions, drive irreversible fibrillar assemblies. Specifically, m6A facilitates stress granule assembly through the multivalent recruitment of YTHDF readers, although its overall contribution to endogenous granule partitioning may be context-dependent (Ries et al., 2019; Khong et al., 2022). On the other side, m1A acts as a protective mark in stress granules, facilitating mRNA triage and translational recovery, while on pathological CAG repeat RNAs it aberrantly recruits TDP-43 into neurotoxic inclusions (Alriquet et al., 2020; Sun et al., 2023). Collectively, these modifications appear to constitute an epitranscriptomic code that enables cells to dynamically regulate ribonucleoprotein condensates under stress and thereby preserve proteostasis.
Author contributions
RA: Writing – review and editing, Writing – original draft. GT: Writing – review and editing, Writing – original draft. RV: Writing – original draft, Writing – review and editing.
Funding
The authors declare that financial support was received for the research and/or publication of this article. We thank the Else Kröner-Fresenius-Stiftung for funding (grant number 2021_EKSE.137).
Acknowledgements
Figures were created using BioRender (BioRender.com).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abbreviations
ALS, amyotrophic lateral sclerosis; EM, Electron Microscopy; FRET, Förster resonance energy transfer; FTD, frontotemporal dementia; IDR, intrinsically disordered regions; LCD, low-complexity domains; LC-MS/MS, liquid chromatography-tandem mass spectrometry; LLPS, liquid-liquid phase separation; lncRNA, long non-coding RNA; m1A, N1-methyladenine; m6A, N6-methyladenine; nSB, nuclear stress body; PB, processing bodies; RBP, RNA-binding protein; RRM, RNA recognition motif; SG, stress granules; UTR, untranslated region.
References
Alberti, S., and Dormann, D. (2019). Liquid–liquid phase separation in disease. Annu. Rev. Genet. 53, 171–194. doi:10.1146/annurev-genet-112618-043527
Alberti, S., and Hyman, A. A. (2021). Biomolecular condensates at the nexus of cellular stress, protein aggregation disease and ageing. Nat. Rev. Mol. Cell Biol. 22, 196–213. doi:10.1038/s41580-020-00326-6
Alriquet, M., Calloni, G., Martínez-Limón, A., Delli Ponti, R., Hanspach, G., Hengesbach, M., et al. (2020). The protective role of m1A during stress-induced granulation. J. Mol. Cell Biol. 12, 870–880. doi:10.1093/jmcb/mjaa023
Alriquet, M., Arsiè, R., Calloni, G., Tartaglia, G. G., and Vabulas, R. M. (2025). Deficiency of N1-Adenine methyltransferase aggravates RNA and protein aggregation. Cells 14, 1370. doi:10.3390/cells14171370
Anders, M., Chelysheva, I., Goebel, I., Trenkner, T., Zhou, J., Mao, Y., et al. (2018). Dynamic m6A methylation facilitates mRNA triaging to stress granules. Life Sci. Alliance 1, e201800113. doi:10.26508/lsa.201800113
Banani, S. F., Lee, H. O., Hyman, A. A., and Rosen, M. K. (2017). Biomolecular condensates: organizers of cellular biochemistry. Nat. Rev. Mol. Cell Biol. 18, 285–298. doi:10.1038/nrm.2017.7
Bar-Yaacov, D., Frumkin, I., Yashiro, Y., Chujo, T., Ishigami, Y., Chemla, Y., et al. (2016). Mitochondrial 16S rRNA is methylated by tRNA methyltransferase TRMT61B in all vertebrates. PLoS Biol. 14, e1002557. doi:10.1371/journal.pbio.1002557
Biamonti, G., and Vourc’h, C. (2010). Nuclear stress bodies. Cold Spring Harb. Perspect. Biol. 2, a000695. doi:10.1101/cshperspect.a000695
Cairo, L. V., Hong, X., Mueller, M. B. D., Yuste-Checa, P., Jagadeesan, C., Bracher, A., et al. (2024). Stress-dependent condensate formation regulated by the ubiquitin-related modifier Urm1. Cell 187, 4656–4673.e28. doi:10.1016/j.cell.2024.06.009
Cao, M., Pietras, C. M., Feng, X., Doroschak, K. J., Schaffner, T., Park, J., et al. (2014). New directions for diffusion-based network prediction of protein function: incorporating pathways with confidence. Bioinformatics 30, i219–i227. doi:10.1093/bioinformatics/btu263
Carey, J. L., and Guo, L. (2022). Liquid-liquid phase separation of TDP-43 and FUS in physiology and pathology of neurodegenerative diseases. Front. Mol. Biosci. 9, 826719. doi:10.3389/fmolb.2022.826719
Chen, W., Feng, P., Yang, H., Ding, H., Lin, H., and Chou, K.-C. (2018). iRNA-3typeA: identifying three types of modification at RNA’s adenosine sites. Mol. Ther. Nucleic Acids 11, 468–474. doi:10.1016/j.omtn.2018.03.012
Chujo, T., and Suzuki, T. (2012). Trmt61B is a methyltransferase responsible for 1-methyladenosine at position 58 of human mitochondrial tRNAs. RNA 18, 2269–2276. doi:10.1261/rna.035600.112
Cui, X., Meng, J., Zhang, S., Chen, Y., and Huang, Y. (2016). A novel algorithm for calling mRNA m6A peaks by modeling biological variances in MeRIP-seq data. Bioinformatics 32, i378–i385. doi:10.1093/bioinformatics/btw281
Dai, X., Wang, T., Gonzalez, G., and Wang, Y. (2018). Identification of YTH domain-containing proteins as the readers for N1-Methyladenosine in RNA. Anal. Chem. 90, 6380–6384. doi:10.1021/acs.analchem.8b01703
Desrosiers, R., Friderici, K., and Rottman, F. (1974). Identification of methylated nucleosides in messenger RNA from novikoff hepatoma cells. Proc. Natl. Acad. Sci. U. S. A. 71, 3971–3975. doi:10.1073/pnas.71.10.3971
Dominissini, D., Moshitch-Moshkovitz, S., Schwartz, S., Salmon-Divon, M., Ungar, L., Osenberg, S., et al. (2012). Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq. Nature 485, 201–206. doi:10.1038/nature11112
Dominissini, D., Nachtergaele, S., Moshitch-Moshkovitz, S., Peer, E., Kol, N., Ben-Haim, M. S., et al. (2016). The dynamic N1-methyladenosine methylome in eukaryotic messenger RNA. Nature 530, 441–446. doi:10.1038/nature16998
Du, H., Zhao, Y., He, J., Zhang, Y., Xi, H., Liu, M., et al. (2016). YTHDF2 destabilizes m(6)A-containing RNA through direct recruitment of the CCR4-NOT deadenylase complex. Nat. Commun. 7, 12626. doi:10.1038/ncomms12626
Dubin, D. T., and Taylor, R. H. (1975). The methylation state of poly A-containing messenger RNA from cultured hamster cells. Nucleic Acids Res. 2, 1653–1668. doi:10.1093/nar/2.10.1653
Dunn, D. B. (1961). The occurrence of 1-methyladenine in ribonucleic acid. Biochim. Biophys. Acta 46, 198–200. doi:10.1016/0006-3002(61)90668-0
Fu, Y., and Zhuang, X. (2020). m6A-binding YTHDF proteins promote stress granule formation. Nat. Chem. Biol. 16, 955–963. doi:10.1038/s41589-020-0524-y
Gao, Y., Pei, G., Li, D., Li, R., Shao, Y., Zhang, Q. C., et al. (2019). Multivalent m6A motifs promote phase separation of YTHDF proteins. Cell Res. 29, 767–769. doi:10.1038/s41422-019-0210-3
Garcia-Campos, M. A., Edelheit, S., Toth, U., Safra, M., Shachar, R., Viukov, S., et al. (2019). Deciphering the “m6A Code” via antibody-independent quantitative profiling. Cell 178, 731–747.e16. doi:10.1016/j.cell.2019.06.013
Gorsheneva, N. A., Sopova, J. V., Azarov, V. V., Grizel, A. V., and Rubel, A. A. (2024). Biomolecular condensates: structure, functions, methods of research. Biochem.-Moscow 89, S205–S223. doi:10.1134/S0006297924140116
Grosjean, H., Keith, G., and Droogmans, L. (2004). “Detection and quantification of modified nucleotides in RNA using thin-layer chromatography,” in RNA interference, editing, and modification: methods and protocols. Editor J. M. Gott (Totowa, NJ: Humana Press), 357–391. doi:10.1385/1-59259-775-0:357
Grozhik, A. V., Olarerin-George, A. O., Sindelar, M., Li, X., Gross, S. S., and Jaffrey, S. R. (2019). Antibody cross-reactivity accounts for widespread appearance of m1A in 5’UTRs. Nat. Commun. 10, 5126. doi:10.1038/s41467-019-13146-w
Herbert, C., Valesyan, S., Kist, J., and Limbach, P. A. (2024). Analysis of RNA and its modifications. Annu. Rev. Anal. Chem. 17, 47–68. doi:10.1146/annurev-anchem-061622-125954
Hirose, T., Ninomiya, K., Nakagawa, S., and Yamazaki, T. (2023). A guide to membraneless organelles and their various roles in gene regulation. Nat. Rev. Mol. Cell Biol. 24, 288–304. doi:10.1038/s41580-022-00558-8
Jia, G., Fu, Y., Zhao, X., Dai, Q., Zheng, G., Yang, Y., et al. (2011). N6-Methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO. Nat. Chem. Biol. 7, 885–887. doi:10.1038/nchembio.687
Jolly, C., Metz, A., Govin, J., Vigneron, M., Turner, B. M., Khochbin, S., et al. (2004). Stress-induced transcription of satellite III repeats. J. Cell Biol. 164, 25–33. doi:10.1083/jcb.200306104
Khong, A., Matheny, T., Huynh, T. N., Babl, V., and Parker, R. (2022). Limited effects of m6A modification on mRNA partitioning into stress granules. Nat. Commun. 13, 3735. doi:10.1038/s41467-022-31358-5
Koh, C. W. Q., Goh, Y. T., and Goh, W. S. S. (2019). Atlas of quantitative single-base-resolution N6-methyl-adenine methylomes. Nat. Commun. 10, 5636. doi:10.1038/s41467-019-13561-z
Leger, A., Amaral, P. P., Pandolfini, L., Capitanchik, C., Capraro, F., Miano, V., et al. (2021). RNA modifications detection by comparative nanopore direct RNA sequencing. Nat. Commun. 12, 7198. doi:10.1038/s41467-021-27393-3
Li, X., Xiong, X., Wang, K., Wang, L., Shu, X., Ma, S., et al. (2016). Transcriptome-wide mapping reveals reversible and dynamic N1-methyladenosine methylome. Nat. Chem. Biol. 12, 311–316. doi:10.1038/nchembio.2040
Li, X., Xiong, X., Zhang, M., Wang, K., Chen, Y., Zhou, J., et al. (2017). Base-resolution mapping reveals distinct m1A methylome in Nuclear- and mitochondrial-encoded transcripts. Mol. Cell 68, 993–1005.e9. doi:10.1016/j.molcel.2017.10.019
Li, Q., Liu, J., Guo, L., Zhang, Y., Chen, Y., Liu, H., et al. (2024). Decoding the interplay between m6A modification and stress granule stability by live-cell imaging. Sci. Adv. 10, eadp5689. doi:10.1126/sciadv.adp5689
Linder, B., Grozhik, A. V., Olarerin-George, A. O., Meydan, C., Mason, C. E., and Jaffrey, S. R. (2015). Single-nucleotide-resolution mapping of m6A and m6Am throughout the transcriptome. Nat. Methods 12, 767–772. doi:10.1038/nmeth.3453
Liu, J., Yue, Y., Han, D., Wang, X., Fu, Y., Zhang, L., et al. (2014). A METTL3–METTL14 complex mediates mammalian nuclear RNA N6-adenosine methylation. Nat. Chem. Biol. 10, 93–95. doi:10.1038/nchembio.1432
Liu, N., Dai, Q., Zheng, G., He, C., Parisien, M., and Pan, T. (2015). N(6)-methyladenosine-dependent RNA structural switches regulate RNA-protein interactions. Nature 518, 560–564. doi:10.1038/nature14234
Liu, L., Lei, X., Meng, J., and Wei, Z. (2020). ISGm1A: integration of sequence features and genomic features to improve the prediction of Human m1A RNA methylation sites. IEEE Access 8, 81971–81977. doi:10.1109/ACCESS.2020.2991070
Luo, Y., Na, Z., and Slavoff, S. A. (2018). P-Bodies: Composition, properties, and functions. Biochemistry 57, 2424–2431. doi:10.1021/acs.biochem.7b01162
Man, J., Zhang, Q., Zhao, T., Sun, D., Long, K., Xiong, W., et al. (2025). YTHDF2 phase separation promotes arsenite-induced oxidative stress by facilitating YTHDF2-mediated PIK3R2 mRNA degradation. Int. J. Biol. Macromol. 318, 144936. doi:10.1016/j.ijbiomac.2025.144936
Martin, E. W., and Mittag, T. (2018). Relationship of sequence and phase separation in protein low-complexity regions. Biochemistry 57, 2478–2487. doi:10.1021/acs.biochem.8b00008
Mathieu, C., Pappu, R. V., and Taylor, J. P. (2020). Beyond aggregation: pathological phase transitions in neurodegenerative disease. Science 370, 56–60. doi:10.1126/science.abb8032
Meyer, K. D. (2019). DART-seq: an antibody-free method for global m6A detection. Nat. Methods 16, 1275–1280. doi:10.1038/s41592-019-0570-0
Meyer, K. D., Saletore, Y., Zumbo, P., Elemento, O., Mason, C. E., and Jaffrey, S. R. (2012). Comprehensive analysis of mRNA methylation reveals enrichment in 3’ UTRs and near stop codons. Cell 149, 1635–1646. doi:10.1016/j.cell.2012.05.003
Micura, R., Pils, W., Höbartner, C., Grubmayr, K., Ebert, M. O., and Jaun, B. (2001). Methylation of the nucleobases in RNA oligonucleotides mediates duplex-hairpin conversion. Nucleic Acids Res. 29, 3997–4005. doi:10.1093/nar/29.19.3997
Molliex, A., Temirov, J., Lee, J., Coughlin, M., Kanagaraj, A. P., Kim, H., et al. (2015). Phase separation by low complexity domains promotes stress granule assembly and drives pathological fibrillization. Cell 163, 123–133. doi:10.1016/j.cell.2015.09.015
Ninomiya, K., Iwakiri, J., Aly, M. K., Sakaguchi, Y., Adachi, S., Natsume, T., et al. (2021). m6 A modification of HSATIII lncRNAs regulates temperature-dependent splicing. EMBO J. 40, e107976. doi:10.15252/embj.2021107976
Patel, A., Lee, H. O., Jawerth, L., Maharana, S., Jahnel, M., Hein, M. Y., et al. (2015). A liquid-to-solid phase transition of the ALS protein FUS accelerated by disease mutation. Cell 162, 1066–1077. doi:10.1016/j.cell.2015.07.047
Protter, D. S. W., Rao, B. S., Treeck, B. V., Lin, Y., Mizoue, L., Rosen, M. K., et al. (2018). Intrinsically disordered regions can contribute promiscuous interactions to RNP granule assembly. Cell Rep. 22, 1401–1412. doi:10.1016/j.celrep.2018.01.036
Ries, R. J., Zaccara, S., Klein, P., Olarerin-George, A., Namkoong, S., Pickering, B. F., et al. (2019). m6A enhances the phase separation potential of mRNA. Nature 571, 424–428. doi:10.1038/s41586-019-1374-1
Ries, R. J., Pickering, B. F., Poh, H. X., Namkoong, S., and Jaffrey, S. R. (2023). m6A governs length-dependent enrichment of mRNAs in stress granules. Nat. Struct. Mol. Biol. 30, 1525–1535. doi:10.1038/s41594-023-01089-2
Ripin, N., and Parker, R. (2023). Formation, function, and pathology of RNP granules. Cell 186, 4737–4756. doi:10.1016/j.cell.2023.09.006
Roberts, J. T., Porman, A. M., and Johnson, A. M. (2021). Identification of m6A residues at single-nucleotide resolution using eCLIP and an accessible custom analysis pipeline. RNA 27, 527–541. doi:10.1261/rna.078543.120
Roden, C., and Gladfelter, A. S. (2021). RNA contributions to the form and function of biomolecular condensates. Nat. Rev. Mol. Cell Biol. 22, 183–195. doi:10.1038/s41580-020-0264-6
Roost, C., Lynch, S. R., Batista, P. J., Qu, K., Chang, H. Y., and Kool, E. T. (2015). Structure and thermodynamics of N6-methyladenosine in RNA: a spring-loaded base modification. J. Am. Chem. Soc. 137, 2107–2115. doi:10.1021/ja513080v
Sabari, B. R., Dall’Agnese, A., and Young, R. A. (2020). Biomolecular condensates in the nucleus. Trends Biochem. Sci. 45, 961–977. doi:10.1016/j.tibs.2020.06.007
Safra, M., Sas-Chen, A., Nir, R., Winkler, R., Nachshon, A., Bar-Yaacov, D., et al. (2017). The m1A landscape on cytosolic and mitochondrial mRNA at single-base resolution. Nature 551, 251–255. doi:10.1038/nature24456
Saha, S., Weber, C. A., Nousch, M., Adame-Arana, O., Hoege, C., Hein, M. Y., et al. (2016). Polar positioning of phase-separated liquid compartments in cells regulated by an mRNA competition mechanism. Cell 166, 1572–1584.e16. doi:10.1016/j.cell.2016.08.006
Sas-Chen, A., and Schwartz, S. (2019). Misincorporation signatures for detecting modifications in mRNA: not as simple as it sounds. Methods 156, 53–59. doi:10.1016/j.ymeth.2018.10.011
Schwartz, S. (2018). m1A within cytoplasmic mRNAs at single nucleotide resolution: a reconciled transcriptome-wide map. RNA 24, 1427–1436. doi:10.1261/rna.067348.118
Shima, H., and Igarashi, K. (2020). N 1-methyladenosine (m1A) RNA modification: the key to ribosome control. J. Biochem. 167, 535–539. doi:10.1093/jb/mvaa026
Shin, Y., and Brangwynne, C. P. (2017). Liquid phase condensation in cell physiology and disease. Science 357, eaaf4382. doi:10.1126/science.aaf4382
Sun, Y., Dai, H., Dai, X., Yin, J., Cui, Y., Liu, X., et al. (2023). m1A in CAG repeat RNA binds to TDP-43 and induces neurodegeneration. Nature 623, 580–587. doi:10.1038/s41586-023-06701-5
Suzuki, T., Yashiro, Y., Kikuchi, I., Ishigami, Y., Saito, H., Matsuzawa, I., et al. (2020). Complete chemical structures of human mitochondrial tRNAs. Nat. Commun. 11, 4269. doi:10.1038/s41467-020-18068-6
Taoka, M., Nobe, Y., Yamaki, Y., Sato, K., Ishikawa, H., Izumikawa, K., et al. (2018). Landscape of the complete RNA chemical modifications in the human 80S ribosome. Nucleic Acids Res. 46, 9289–9298. doi:10.1093/nar/gky811
Vandelli, A., Broglia, L., Armaos, A., Delli Ponti, R., and Tartaglia, G. G. (2024). Rationalizing the effects of RNA modifications on protein interactions. Mol. Ther. Nucleic Acids 35, 102391. doi:10.1016/j.omtn.2024.102391
Varenyk, Y., Spicher, T., Hofacker, I. L., and Lorenz, R. (2023). Modified RNAs and predictions with the ViennaRNA package. Bioinformatics 39, btad696. doi:10.1093/bioinformatics/btad696
Vilardo, E., Nachbagauer, C., Buzet, A., Taschner, A., Holzmann, J., and Rossmanith, W. (2012). A subcomplex of human mitochondrial RNase P is a bifunctional methyltransferase—extensive moonlighting in mitochondrial tRNA biogenesis. Nucleic Acids Res. 40, 11583–11593. doi:10.1093/nar/gks910
Vodnala, M., Choi, E.-B., and Fong, Y. W. (2021). Low complexity domains, condensates, and stem cell pluripotency. World J. Stem Cells 13, 416–438. doi:10.4252/wjsc.v13.i5.416
Waku, T., Nakajima, Y., Yokoyama, W., Nomura, N., Kako, K., Kobayashi, A., et al. (2016). NML-mediated rRNA base methylation links ribosomal subunit formation to cell proliferation in a p53-dependent manner. J. Cell Sci. 129, 2382–2393. doi:10.1242/jcs.183723
Wang, X., Lu, Z., Gomez, A., Hon, G. C., Yue, Y., Han, D., et al. (2014). N6-methyladenosine-dependent regulation of messenger RNA stability. Nature 505, 117–120. doi:10.1038/nature12730
Wang, X., Zhao, B., Roundtree, I. A., Lu, Z., Han, D., Ma, H., et al. (2015). N6-methyladenosine modulates messenger RNA translation efficiency. Cell 161, 1388–1399. doi:10.1016/j.cell.2015.05.014
Wang, X., Feng, J., Xue, Y., Guan, Z., Zhang, D., Liu, Z., et al. (2016). Structural basis of N6-adenosine methylation by the METTL3–METTL14 complex. Nature 534, 575–578. doi:10.1038/nature18298
Wang, J., Wang, L., Diao, J., Shi, Y. G., Shi, Y., Ma, H., et al. (2020). Binding to m6A RNA promotes YTHDF2-mediated phase separation. Protein Cell 11, 304–307. doi:10.1007/s13238-019-00660-2
Xiao, W., Adhikari, S., Dahal, U., Chen, Y.-S., Hao, Y.-J., Sun, B.-F., et al. (2016). Nuclear m(6)A reader YTHDC1 regulates mRNA splicing. Mol. Cell 61, 507–519. doi:10.1016/j.molcel.2016.01.012
Zaccara, S., and Jaffrey, S. R. (2020). A unified model for the function of YTHDF proteins in regulating m6A-Modified mRNA. Cell 181, 1582–1595.e18. doi:10.1016/j.cell.2020.05.012
Zacco, E., Broglia, L., Kurihara, M., Monti, M., Gustincich, S., Pastore, A., et al. (2024). RNA: the unsuspected conductor in the orchestra of macromolecular crowding. Chem. Rev. 124, 4734–4777. doi:10.1021/acs.chemrev.3c00575
Zhang, Y., and Hamada, M. (2018). DeepM6ASeq: prediction and characterization of m6A-containing sequences using deep learning. BMC Bioinforma. 19, 524. doi:10.1186/s12859-018-2516-4
Zhang, H., Elbaum-Garfinkle, S., Langdon, E. M., Taylor, N., Occhipinti, P., Bridges, A. A., et al. (2015). RNA controls PolyQ protein phase transitions. Mol. Cell 60, 220–230. doi:10.1016/j.molcel.2015.09.017
Zhang, R., Li, S., Feng, W., Qian, S., Chellappa, A. J., and Wang, F. (2025). Rim4-seeded stress granules connect temperature sensing to meiotic regulation. Nat. Commun. 16, 5566. doi:10.1038/s41467-025-60645-0
Zheng, G., Dahl, J. A., Niu, Y., Fedorcsak, P., Huang, C.-M., Li, C. J., et al. (2013). ALKBH5 is a mammalian RNA demethylase that impacts RNA metabolism and mouse fertility. Mol. Cell 49, 18–29. doi:10.1016/j.molcel.2012.10.015
Zheng, Q., Gan, H., Yang, F., Yao, Y., Hao, F., Hong, L., et al. (2020). Cytoplasmic m1A reader YTHDF3 inhibits trophoblast invasion by downregulation of m1A-methylated IGF1R. Cell Discov. 6, 12. doi:10.1038/s41421-020-0144-4
Zhou, H., Kimsey, I. J., Nikolova, E. N., Sathyamoorthy, B., Grazioli, G., McSally, J., et al. (2016). m(1)A and m(1)G disrupt A-RNA structure through the intrinsic instability of hoogsteen base pairs. Nat. Struct. Mol. Biol. 23, 803–810. doi:10.1038/nsmb.3270
Zhou, Y., Zeng, P., Li, Y.-H., Zhang, Z., and Cui, Q. (2016). SRAMP: prediction of Mammalian N6-methyladenosine (m6A) sites based on sequence-derived features. Nucleic Acids Res. 44, e91. doi:10.1093/nar/gkw104
Zhou, H., Rauch, S., Dai, Q., Cui, X., Zhang, Z., Nachtergaele, S., et al. (2019). Evolution of a reverse transcriptase to map N1-methyladenosine in human messenger RNA. Nat. Methods 16, 1281–1288. doi:10.1038/s41592-019-0550-4
Keywords: N1-methyladenine, N6-methyladenine, RNA-binding proteins, granulation, protein aggregation
Citation: Arsiè R, Tartaglia GG and Vabulas RM (2025) Methyladenine as a regulator of biomolecular condensation. Front. Mol. Biosci. 12:1722147. doi: 10.3389/fmolb.2025.1722147
Received: 10 October 2025; Accepted: 17 November 2025;
Published: 26 November 2025.
Edited by:
Matteo Cassandri, Sapienza University of Rome, ItalyReviewed by:
Wenhao Ouyang, Sun Yat-sen University, ChinaGiada Mele, Sapienza University of Rome, Italy
Copyright © 2025 Arsiè, Tartaglia and Vabulas. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: R. Martin Vabulas, bWFydGluLnZhYnVsYXNAY2hhcml0ZS5kZQ==