 Gen Li1†
Gen Li1† Shishuang Cui1†
Shishuang Cui1† Juanjuan Du1Jin Liu1Pingchen Zhang1Yang Fu1
Juanjuan Du1Jin Liu1Pingchen Zhang1Yang Fu1 Yixi He1Haiyan Zhou1*Jianfang Ma1*Shengdi Chen1,2*
Yixi He1Haiyan Zhou1*Jianfang Ma1*Shengdi Chen1,2*- 1Department of Neurology & Collaborative Innovation Center for Brain Science, Ruijin Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai, China
- 2Co-innovation Center of Neuroregeneration, Nantong University, Nantong, China
Study Objectives: The aim of the study was to investigate the relationship between 22 single nucleotide polymorphisms (SNPs) and Parkinson’s disease (PD) in the Chinese population.
Methods: A total of 250 PD patients and 240 healthy controls were recruited. The SNaPshot technique and the polymer chain reaction were used to detect 22 SNPs.
Results: rs8005172 of GALC, rs9468199 of ZNF184 and rs34043159 of IL1R2, were associated with PD (rs8005172: p = 0.009, OR = 0.69, allele model, p = 0.010, additive model, p = 0.015, OR = 2.17, dominant model; p = 0.020, OR = 2.11, dominant model after adjustment; p = 0.036, OR = 1.47, recessive model after adjustment; rs9468199: p = 0.008, OR = 1.52, allele model, p = 0.008, additive model, p = 0.007, OR = 0.22, recessive model, p = 0.005, OR = 0.20, recessive model after adjustment; rs34043159: p = 0.034, OR = 1.31, allele model, p = 0.036, additive model).
Conclusion: Our study revealed that GALC, ZNF184, and IL1R2 were associated with PD in the southern Chinese population. GALC was also associated with LOPD. ELOVL7 and ZNF184 were associated with EOPD. In addition, trends of association to PD, between SATB1, NMD3, and FGF20, were also found.
Statement of Significance: Genetic play an important role in the pathogenesis factors of Parkinson’s disease (PD). We found that GALC, ZNF184, and IL1R2 were associated with PD. GALC was also associated with late onset of PD, while ELOVL7 and ZNF184 were associated with early onset PD. This study is the first to find an association between GALC, ZNF184, and rs2280104 with PD.
HIGHLIGHTS
- GALC, ZNF184, and IL1R2 were associated with PD.
- GALC was also associated with late onset PD.
- ELOVL7 and ZNF184 were associated with early onset PD.
Introduction
Parkinson’s disease is a debilitating and progressive movement disorder characterized by bradykinesia, resting tremors and rigidity. As the second most common neurodegenerative disorder after Alzheimer’s disease, it causes a high burden on the health-care system (Schrag and Schott, 2006; Ascherio and Schwarzschild, 2012; Olesen et al., 2012). The pathological characteristics of PD include an abnormal α-synuclein aggregation and a loss of dopaminergic neurons in the substantia nigra. In addition, several hypotheses of the pathogenesis of PD, such as autophagy and mitochondrial dysfunction, have been raised previously (Dehay et al., 2013; Wang and Mao, 2014; Wang et al., 2016; Xilouri et al., 2016).
Genetic factors play an important role in the pathogenesis of PD. With the help of GWAS, several PD-associated locus, such as LRRK2, were discovered (Trinh and Farrer, 2013). Recently, using a meta-analysis of GWAS, several PD risk loci such as GALC and IL1R2, etc., have been identified, mainly in Caucasians (Nalls et al., 2014; Chang et al., 2017). However, because of different allele frequencies across ethnicities, we cannot infer distributions of these loci in Asians. Additionally, the association between these loci and PD in the southern Chinese population, was unclear. In this study, we attempted to replicate these loci demonstrated in these two previous articles (Nalls et al., 2014; Chang et al., 2017) and discussed those genes associated with PD in the southern Chinese population.
Materials and Methods
Study Population
Parkinson’s disease was diagnosed by movement disorder specialists, based on diagnostic criteria provided by the MDS (Postuma et al., 2015). Control subjects from a healthy community were enrolled and evaluated by movement disorder specialists for exclusion of PD. Hoehn – Yahr staging and familial history were recorded for patients with PD. When assessing PD patients, secondary causes, such as inflammatory, drug-induced, vascular and toxin-induced parkinsonism were all excluded. Parkinsonism along with other neurodegenerative diseases, such as progressive supranuclear palsy, multiple system atrophy and Wilson’s disease were also excluded. We divided the PD patients into a late onset PD group (LOPD) and an early onset PD group (EOPD). Patients who were above 45 years old when PD was first diagnosed, were regarded as LOPD (Gonzalez-Del Rincon Mde et al., 2013). Mild PD was defined when Hoehn – Yahr staging was below 2.5 after assessment. Patients with relatives who have PD (within the last three generations), were regarded as familial PD patients. All participants signed consent forms and this study was approved by the ethics committee of the Ruijin Hospital affiliated to Shanghai Jiao Tong University School of Medicine.
DNA Preparations and Genotyping
Two milliliter blood samples were collected from PD patients and healthy controls. A phenol-chloroform-isopropyl alcohol method was used to extract the DNA. The Primer Premier 5 (version 5.00, PREMIER Biosoft International) was used to design the primers. The following SNPs were tested: rs4784227 of TOX3, rs2280104 of SORBS3/PDLIM2/C8orf58/BIN3, rs11868035 of SREBF/RAI1, rs11343 of COQ7, rs591323 of FGF20, rs7077361 of ITGA8, rs8005172 of GALC, rs34016896 of NMD3, rs2694528 of ELOVL7, rs10906923 of FAM171A1, rs9468199 of ZNF184, rs60298754 of MMP16, rs4073221 of SATB1, rs143918452 of ALAS1/TLR9/DNAH1/BAP1/PHF7/NISCH/STAB1/ITIH3/ITIH4, rs12497850 of NCKIPSD/CDC71, rs4653767 of ITPKB, rs34043159 of IL1R2, rs601999 of ATP6V0A1/PSMC3IP/TUBG2, rs2823357 of USP25, rs2740594 of CTSB and rs78738012 of ANK2/CAMK2D. The SNaPshot technique was used to genotype SNPs. The polymer chain reaction was used to detect the rs13294100 of SH3GL2. Details of primers and the reaction system of the polymer chain reaction are described in Supplementary Table S1.
Statistical Analysis
R (version 3.5.0) was used to perform the statistical analysis. The CATT package (version 2.0), epitools package (version 0.5–10) and the survival package (version 2.41–3) were used in R. A t-test was used to compare the differences in age between PD patients and controls. A chi-square test was used to compare the differences in gender, assessing the HWE and the allele distribution between PD patients and controls. Risk analysis was performed by a logistic regression model, and an OR, with a 95% confidence interval for each SNP, according to the dominant, recessive and overdominant models, was calculated. Age and gender were adjusted accordingly. The Cochran-Armitage trend test was used to calculate additive models. We corrected the p-values using the Bonferroni method. An AFT model was used to assess the relationship between SNPs and PD over time. We assumed the distribution as a logistic in the AFT model. The logLik results were used to model the evaluation. The genetic power was calculated using Power and Sample Size Calculations (version 3.1.2) (Dupont and Plummer, 1990, 1998).
Results
There was no difference between age and gender in PD patients and control groups. The PD group was predominantly male (58.0%), and the average age was 54.69 ± 13.95 years. 72.0% of the PD patients had mild PD, and 10.80% PD patients had familial PD. 33 (13.20%) of the PD patients had EOPD. In the EOPD patients, 14 (42.42%) were female and 28 (84.84%) of them had mild PD, after assessment, while 4 (12.12%) of them had familial PD. In the LOPD patients, 91 (41.94%) were female, and 152 (69.58%) of them had mild PD, while 23 (10.60%) of them had familial PD. There were no discrepancies in gender between EOPD and LOPD patients, mild PD or familial PD patients (gender: p = 1.000, chi-square test; mild PD: p = 0.1196, chi-square test; familial PD: p = 0.765, fisher test) (Table 1).
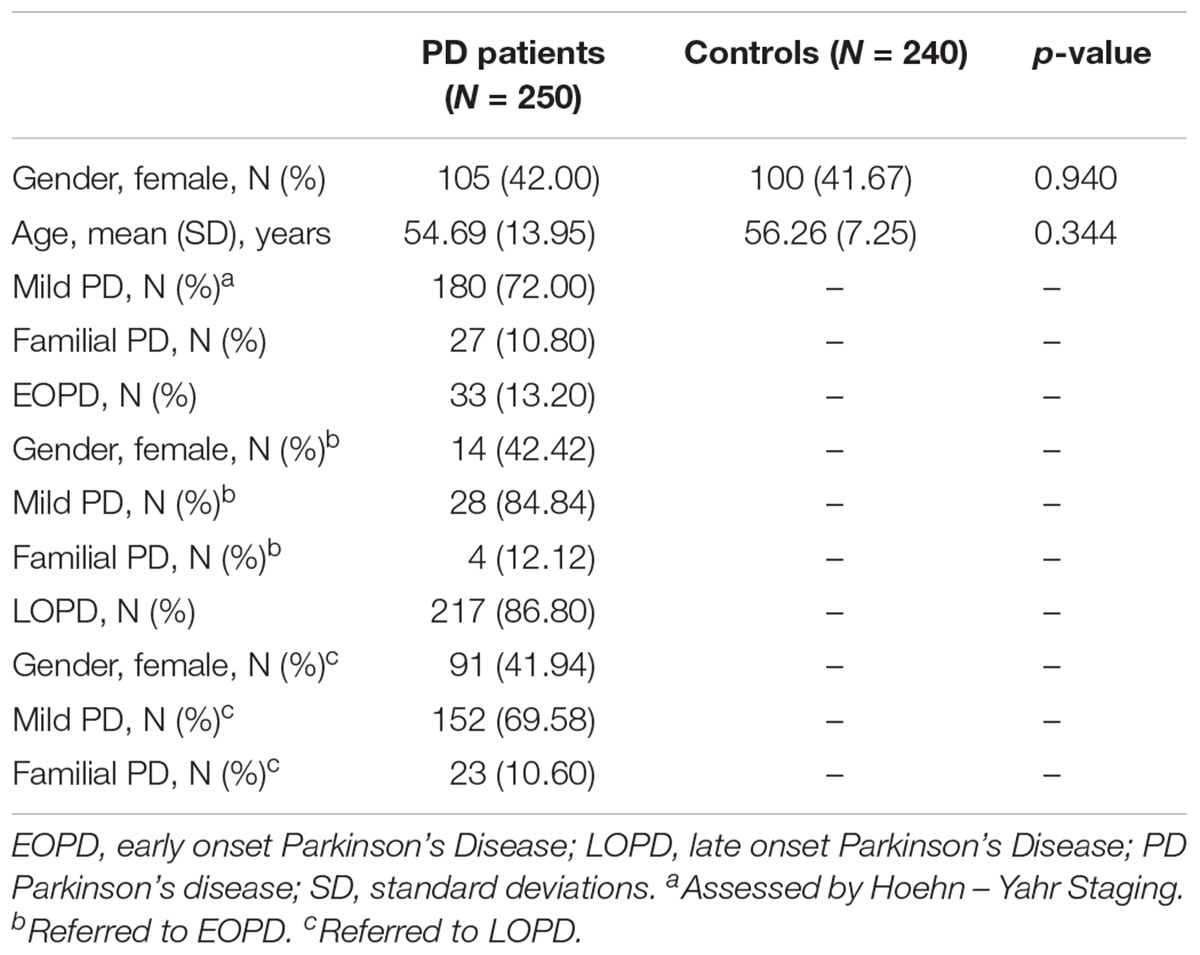
Table 1. Demographic data of cases and controls.
All SNPs were in the HWE except for the rs13294100 of SH3GL2. The rs2740594 of CTSB were all an AA genotype, while the rs7077361 of ITGA8 and rs78738012 of ANK2/CAMK2D were all a TT genotype. The rs8005172 of GALC, rs9468199 of ZNF184 and the rs34043159 of IL1R2 were all associated with PD, in both the allele model as well as the additive model (rs8005172: p = 0.009, OR = 0.69, allele model, p = 0.010, additive model; rs9468199: p = 0.008, OR = 1.52, allele model, p = 0.008, additive model; rs34043159: p = 0.034, OR = 1.31, allele model, p = 0.036, additive model). The dominant model of the rs8005172 of GALC, was associated with PD, with or without adjusting the age and gender (p = 0.015, OR = 2.17; p = 0.020, OR = 2.11, after adjustment). Under recessive models, the rs9468199 of ZNF184 as well as the rs8005172 of GALC, were associated with PD after adjustment (rs9468199: p = 0.007, OR = 0.22, p = 0.005, OR = 0.20, after adjustment; rs8005172: p = 0.036, OR = 1.47, after adjustment). None were statistically significant after the Bonferroni correction (Table 2 and Supplementary Table S2).
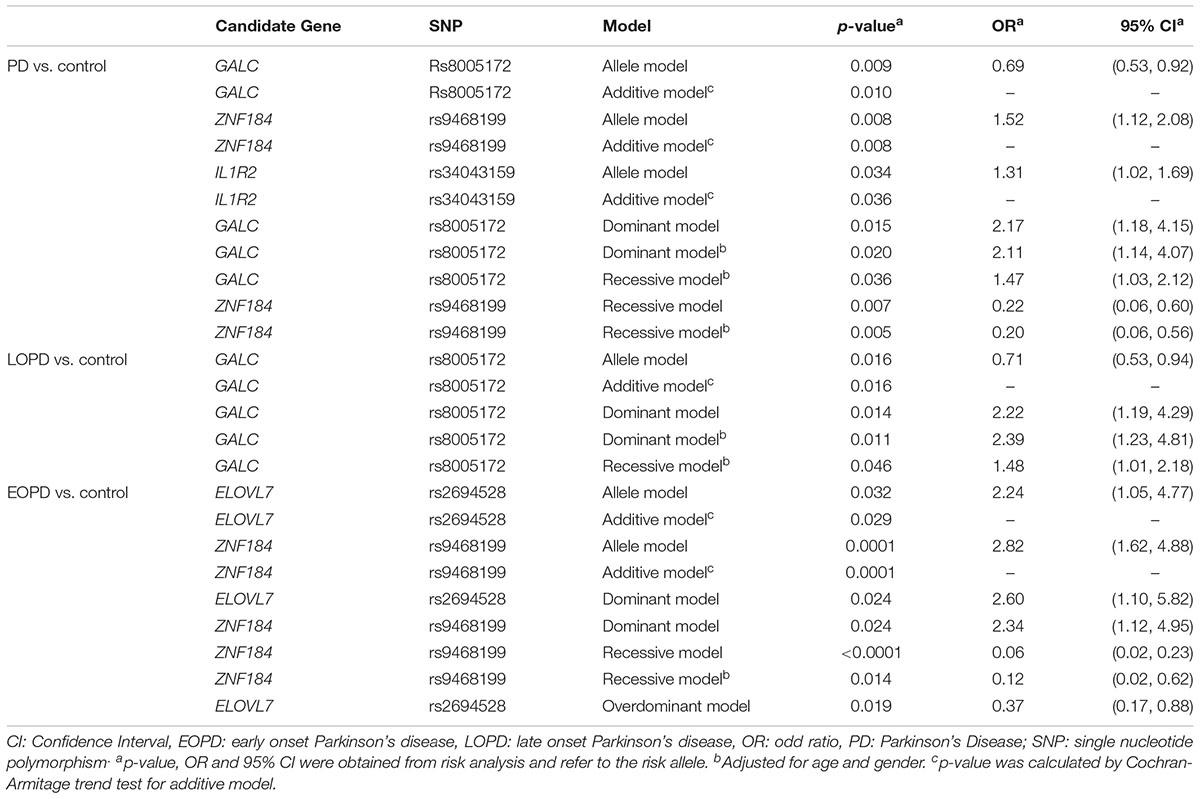
Table 2. Association of SNPs of candidate genes and odds ratio to PD risk.
As for LOPD, all SNPs were in the HWE except for the rs13294100 of SH3GL2. The rs8005172 of GALC was associated with PD in both the allele model as well as the additive model (rs8005172: p = 0.016, OR = 0.71, allele model, p = 0.016, additive model). The dominant model of the rs8005172 of GALC, with or without adjustment as well as the recessive model of the rs8005172 of GALC, after adjustment, were both associated with PD (dominant model: p = 0.014, OR = 2.22; p = 0.011, OR = 2.39, after adjustment; recessive model: p = 0.046, OR = 1.48, after adjustment). None were statistically significant after the Bonferroni correction (Table 2 and Supplementary Table S3).
In the analysis between EOPD and PD, all SNPs were in the HWE except for the rs13294100 of SH3GL2. The genetic power of the rs4784227 of TOX3, was insufficient. In the remaining SNPs, the rs2694528 of ELOVL7 and the rs9468199 of ZNF184 were associated with PD, in both the allele and additive model (rs2694528: p = 0.032, OR = 2.24, allele model, p = 0.029, additive model; rs9468199: p = 0.0001, OR = 2.82, allele model, p = 0.0001, additive model). The dominant models of the rs2694528 of ELOVL7 and the rs9468199 of ZNF184, were associated with PD before adjustment (rs2694528: p = 0.024, OR = 2.60; rs9468199: p = 0.024, OR = 2.34). The recessive model of the rs9468199 of ZNF184, was associated with PD with or without adjustment (p < 0.0001, OR = 0.06; p = 0.014, OR = 0.12, after adjustment). The overdominant model of the rs2694528 of ELOVL7, was associated with PD (p = 0.019, OR = 0.37). After correcting the p-values, the allele model, additive model and the recessive model of the rs9468199 of ZNF184, remained statistically significant (Table 2 and Supplementary Table S4).
Within these models, we found that the combination of the dominant model of the rs8005172 of GALC and the recessive model of the rs9468199 of ZNF184, could better interpret the risk of PD. In LOPD, the dominant model of the rs8005172 of GALC, was better. As for EOPD, the combination of the overdominant model of the rs2694528 of ELOVL7 and the rs9468199 of ZNF184, could over time, interpret the risk better (Table 3 and Supplementary Table S5).
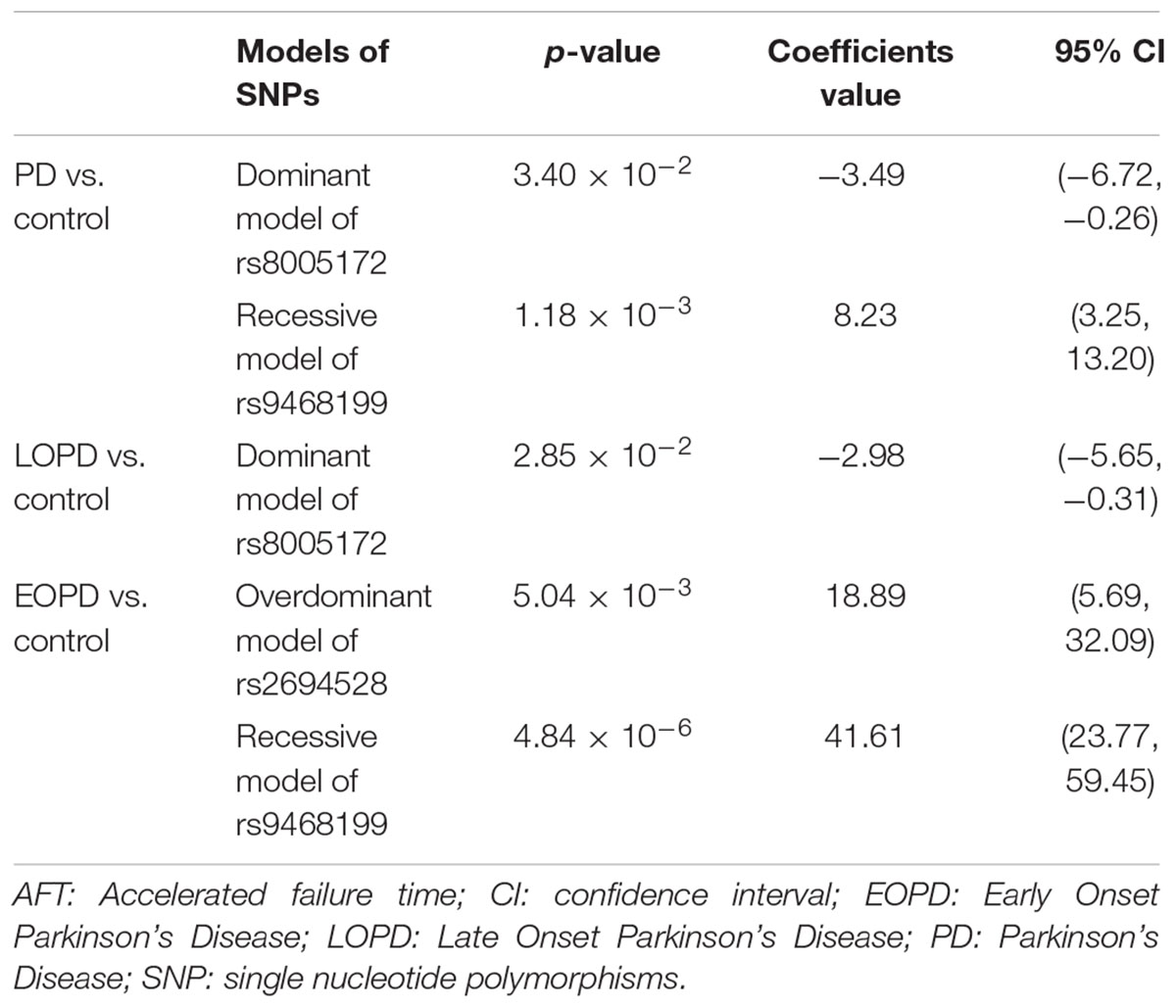
Table 3. Accelerated failure time models of genetic models of SNPs of each comparison.
Discussion
In our study, we found that the following SNPs were associated with PD: the rs8005172 of GALC (allele model, additive model, dominant model, recessive model after adjustment), the rs9468199 of ZNF184 (allele model, additive model, recessive model) and the rs34043159 of IL1R2 (allele model, additive model). The rs8005172 of GALC (allele model, additive model, dominant model, recessive model after adjustment) was associated with LOPD, while the rs2694528 of ELOVL7 (allele model, additive model, dominant model, overdominant model) and the rs9468199 of ZNF184 (allele model, additive model, dominant model, recessive model) were associated with EOPD. We also found that the rs4073221 of SATB1, the rs34016896 of NMD3 and the rs591323 of FGF20 were all associated with PD in the southern Chinese in trend. This research, is the first to replicate the associations of GALC, ZNF184, ELOVL7, and IL1R2 with PD, in the southern Chinese population. We failed to replicate the association of the remaining loci, identified in Caucasians, with PD in the Chinese population. As ethnicities differ (between Caucasians and Chinese) and because of very few interracial marriages, allele frequencies and the genetic biomarkers of PD might also differ.
GALC, which encodes galactocerebrosidase, is the cause of Krabbe disease, a lysosomal storage disorder (Marshall and Bongarzone, 2016). α-synuclein can be degraded and eliminated by autophagy, which could maintain the homeostasis of the dopaminergic neurons (Mishra et al., 2015). Dysfunction of GALC could increase autophagy. In GALC deficient cells, autophagy markers were increased (Ribbens et al., 2014). The deposition of α-synuclein can also be observed in Krabbe disease (Smith et al., 2014). Synaptic function, influenced by psychosine, may be associated with the pathogenesis of PD (Marshall and Bongarzone, 2016). Similarly, Gaucher disease, a lysosomal storage disorder in which α-synuclein was also found, is associated with PD via several links such as genetics, autophagy impairment, and mitochondrial dysfunction (DePaolo et al., 2009; Velayati et al., 2010; Shachar et al., 2011). Our findings provide more proof of the association between lysosomal storage disorder and PD, and support the theory of autophagy in the pathogenesis of PD.
Protein encoded by ZNF184 belong to the Kruppel-like zinc finger family and may be involved in transcriptional regulation. The function of ZNF184 is still unknown. As previously reported, ZNF184 was associated with aberrant cell proliferations, such as lung cancer and choriocarcinoma (Li et al., 2005; ?). A trans-CpG nearest to ZNF184, associated with genetically defined elevated homocysteine (MTHFR C677T), was observed (Mandaviya et al., 2017). Hyperhomocysteinemia is a treatable risk factor of PD. MTHFR C677T was associated with the increased risk of PD (Wu et al., 2013). Higher serum homocysteine could predict higher dopaminergic neurodegeneration in the substantia nigra (Haghdoost-Yazdi et al., 2014). In addition, hyperhomocysteinemia was also associated with the deterioration of several cognitive functions (memory, verbal fluency, etc.) in PD (Licking et al., 2017). The relationship between ZNF184, hyperhomocysteinemia and PD still needs to be investigated further.
The product of ELOVL7 is a member of the elongase family, ELOVLs, catalyzing the elongation of very long-chain fatty acids. The ELOVL7 protein, presented high activity toward acyl-CoAs with a C18 carbon chain length (Naganuma et al., 2011). It is an important lipogenic enzyme that is involved in the pathogenesis of prostate cancer(Tamura et al., 2009). According to the function of ELOVLs, there is a chance thatELOVL7 could be involved in the pathogenesis of oxidative stress. Recent studies also indicated that ELOVL7, with the help of GWAS, was associated with multiple system atrophy (Sailer et al., 2016; Gu et al., 2018). However, the link between ELOVL7 and neurodegeneration is still unknown. In this study, we found that ELOVL7 is associated with EOPD but not with PD. It might indicate that ELOVL7 plays a more important role at the onset of EOPD, rather than in PD. Furthermore, the population that we tested was small. Larger studies are needed to test this association.
IL1R2 expresses the receptor of IL1. The association between IL1 and dopaminergic dysfunction has been proven. In the serum of PD patients, the level of IL1-β was higher than in the control patients (Williams-Gray et al., 2016). The IL1B gene was associated with restless legs syndrome (Hennessy et al., 2014). The increased levels of IL1, in patients suffering from depression, were also observed (Maes et al., 2012). It may indicate that immunological factors are involved in the pathogenesis of PD.
There were some limitations in our study. First, the genetic power of most SNPs in our study was low. As a single centered study, it was difficult to obtain a large population. Similar studies, in the Asian population, are needed to confirm our findings. Second, we did not create subgroups, to analyze differences between other confounders such as gender, PD subtypes, severity of PD, etc., Because of the small sample, validity might be lost, therefore, larger sized cohort studies are warranted.
Our study revealed that GALC, ZNF184, and IL1R2 are associated with PD in the southern Chinese population. GALC is also associated with LOPD, while ELOVL7 and ZNF184 are associated with EOPD. Additionally, trends of the association of SATB1, NMD3, and FGF20 with PD, were also found. Since these genes were identified in both Asian and Caucasian groups, research on the pathogenesis of these genes to PD is needed.
Ethics Statement
This study was carried out in accordance with the recommendations of the ethic committee of Ruijin Hospital affiliated to Shanghai Jiao Tong University School of Medicine with written informed consent from all subjects. All subjects gave written informed consent in accordance with the Declaration of Helsinki. The protocol was approved by the ethic committee of Ruijin Hospital affiliated to Shanghai Jiao Tong University School of Medicine.
Author Contributions
GL and SC collected the PD and control patient data, performed the statistical analysis and drafted the manuscript. JL, JD, PZ, YF, and YH collected the data on PD. SC, JM, and HZ designed the study, supervised the study, double-checked the statistical analysis and revised the manuscript.
Funding
This work was supported by grants from the National Natural Science Foundation of China (Grant Nos. 81430022, 91332107, and 81371407).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank all the patients and healthy controls who participated in this study.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnagi.2018.00402/full#supplementary-material
Abbreviation
AFT, accelerated failure time; EOPD, early onset Parkinson’s disease; GWAS, genome-wide association studies; HWE, Hardy-Weinberg equilibrium; IL1, interleukin 1; LOPD, late onset Parkinson’s disease; MDS, movement disorders society; OR, odds ratio; PD, Parkinson’s disease; SNP, single nucleotide polymorphism.
References
Ascherio, A., and Schwarzschild, M. A. (2012). The epidemiology of Parkinson’s disease: risk factors and prevention. Lancet Neurol. 15, 1257–1272. doi: 10.1016/S1474-4422(16)30230-7
Chang, D., Nalls, M. A., Hallgrimsdottir, I. B., Hunkapiller, J., van der Brug, M., Cai, F., et al. (2017). A meta-analysis of genome-wide association studies identifies 17 new Parkinson’s disease risk loci. Nat. Genet. 49, 1511–1516. doi: 10.1038/ng.3955
Dehay, B., Martinez-Vicente, M., Caldwell, G. A., Caldwell, K. A., Yue, Z., Cookson, M. R., et al. (2013). Lysosomal impairment in Parkinson’s disease. Mov. Disord. 28, 725–732. doi: 10.1002/mds.25462
DePaolo, J., Goker-Alpan, O., Samaddar, T., Lopez, G., and Sidransky, E. (2009). The association between mutations in the lysosomal protein glucocerebrosidase and parkinsonism. Mov. Disord. 24, 1571–1578. doi: 10.1002/mds.22538
Dupont, W. D., and Plummer, W. D. (1990). Power and sample size calculations: a review and computer program. Control Clin. Trials 11, 116–128. doi: 10.1016/0197-2456(90)90005-M
Dupont, W. D., and Plummer, W. D. (1998). Power and sample size calculations for studies involving linear regression. Control Clin. Trials 19, 589–601. doi: 10.1016/S0197-2456(98)00037-3
Gonzalez-Del Rincon Mde, L., Monroy Jaramillo, N., Suarez Martinez, A. I., Yescas Gomez, P., Boll Woehrlen, M. C., Lopez Lopez, M., et al. (2013). The L444P GBA mutation is associated with early-onset Parkinson’s disease in Mexican mestizos. Clin. Genet. 84, 386–387. doi: 10.1111/cge.12084
Gu, X., Chen, Y., Zhou, Q., Lu, Y. C., Cao, B., Zhang, L., et al. (2018). Analysis of GWAS-linked variants in multiple system atrophy. Neurobiol. Aging 67, 201.e1–201.e4. doi: 10.1016/j.neurobiolaging.2018.03.018
Haghdoost-Yazdi, H., Sarookhani, M., Faraj, A., Fraidouni, N., Dargahi, T., Yaghoubidoust, M. H., et al. (2014). Evaluation of the association between blood homocysteine concentration and the degree of behavioral symptoms in the 6-hydroxydopamine-induced Parkinsonism in rat. Pharmacol. Biochem. Behav. 124, 297–304. doi: 10.1016/j.pbb.2014.06.020
Hennessy, M. D., Zak, R. S., Gay, C. L., Pullinger, C. R., Lee, K. A., and Aouizerat, B. E. (2014). Polymorphisms of interleukin-1 Beta and interleukin-17Alpha genes are associated with restless legs syndrome. Biol. Res. Nurs. 16, 143–151. doi: 10.1177/1099800413478827
Li, X. N., Li, Y. L., Liu, G. B., and Ding, Y. Q. (2005). Selection of choriocarcinoma-associated genes using bioinformatics. Di Yi Jun Yi Da Xue Xue Bao 25, 1–6.
Licking, N., Murchison, C., Cholerton, B., Zabetian, C. P., Hu, S. C., Montine, T. J., et al. (2017). Homocysteine and cognitive function in Parkinson’s disease. Parkinsonism Relat. Disord. 44, 1–5. doi: 10.1016/j.parkreldis.2017.08.005
Maes, M., Song, C., and Yirmiya, R. (2012). Targeting IL-1 in depression. Expert Opin. Ther. Targets 16, 1097–1112. doi: 10.1517/14728222.2012.718331
Mandaviya, P. R., Joehanes, R., Aïssi, D., Kühnel, B., Marioni, R. E., Truong, V., et al. (2017). Genetically defined elevated homocysteine levels do not result in widespread changes of DNA methylation in leukocytes. PLoS One 12:e0182472. doi: 10.1371/journal.pone.0182472
Marshall, M. S., and Bongarzone, E. R. (2016). Beyond Krabbe’s disease: the potential contribution of galactosylceramidase deficiency to neuronal vulnerability in late-onset synucleinopathies. J. Neurosci. Res. 94, 1328–1332. doi: 10.1002/jnr.23751
Mishra, A. K., ur Rasheed, M. S., Shukla, S., Tripathi, M. K., Dixit, A., and Singh, M. P. (2015). Aberrant autophagy and parkinsonism: does correction rescue from disease progression? Mol. Neurobiol. 51, 893–908. doi: 10.1007/s12035-014-8744-3
Naganuma, T., Sato, Y., Sassa, T., Ohno, Y., and Kihara, A. (2011). Biochemical characterization of the very long-chain fatty acid elongase ELOVL7. FEBS Lett. 585, 3337–3341. doi: 10.1016/j.febslet.2011.09.024
Nalls, M. A., Pankratz, N., Lill, C. M., Do, C. B., Hernandez, D. G., Saad, M., et al. (2014). Large-scale meta-analysis of genome-wide association data identifies six new risk loci for Parkinson’s disease. Nat. Genet. 46, 989–993. doi: 10.1038/ng.3043
Olesen, J., Gustavsson, A., Svensson, M., Wittchen, H. U., and Jonsson, B. (2012). The economic cost of brain disorders in Europe. Eur. J. Neurol. 19, 155–162. doi: 10.1111/j.1468-1331.2011.03590.x
Postuma, R. B., Berg, D., Stern, M., Poewe, W., Olanow, C. W., Oertel, W., et al. (2015). MDS clinical diagnostic criteria for Parkinson’s disease. Mov. Disord. 30, 1591–1601. doi: 10.1002/mds.26424
Ribbens, J. J., Moser, A. B., Hubbard, W. C., Bongarzone, E. R., and Maegawa, G. H. (2014). Characterization and application of a disease-cell model for a neurodegenerative lysosomal disease. Mol. Genet. Metab. 111, 172–183. doi: 10.1016/j.ymgme.2013.09.011
Sailer, A., Scholz, S. W., Nalls, M. A., Schulte, C., Federoff, M., Price, T. R., et al. (2016). A genome-wide association study in multiple system atrophy. Neurology 87, 1591–1598. doi: 10.1212/WNL.0000000000003221
Schrag, A., and Schott, J. M. (2006). Epidemiological, clinical, and genetic characteristics of early-onset parkinsonism. Lancet Neurol. 5, 355–363. doi: 10.1016/S1474-4422(06)70411-2
Shachar, T., Lo Bianco, C., Recchia, A., Wiessner, C., Raas-Rothschild, A., and Futerman, A. H. (2011). Lysosomal storage disorders and Parkinson’s disease: gaucher disease and beyond. Mov. Disord. 26, 1593–1604. doi: 10.1002/mds.23774
Smith, B. R., Santos, M. B., Marshall, M. S., Cantuti-Castelvetri, L., Lopez-Rosas, A., Li, G., et al. (2014). Neuronal inclusions of alpha-synuclein contribute to the pathogenesis of Krabbe disease. J. Pathol. 232, 509–521. doi: 10.1002/path.4328
Tamura, K., Makino, A., Hullin-Matsuda, F., Kobayashi, T., Furihata, M., Chung, S., et al. (2009). Novel lipogenic enzyme ELOVL7 is involved in prostate cancer growth through saturated long-chain fatty acid metabolism. Cancer Res. 69, 8133–8140. doi: 10.1158/0008-5472.CAN-09-0775
Trinh, J., and Farrer, M. (2013). Advances in the genetics of Parkinson disease. Nat. Rev Neurol. 9, 445–454. doi: 10.1038/nrneurol.2013.132
Velayati, A., Yu, W. H., and Sidransky, E. (2010). The role of glucocerebrosidase mutations in Parkinson disease and lewy body disorders. Curr. Neurol. Neurosci. Rep. 10, 190–198. doi: 10.1007/s11910-010-0102-x
Wang, B., Abraham, N., Gao, G., and Yang, Q. (2016). Dysregulation of autophagy and mitochondrial function in Parkinson’s disease. Transl. Neurodegener. 5:19. doi: 10.1186/s40035-016-0065-1
Wang, G., and Mao, Z. (2014). Chaperone-mediated autophagy: roles in neurodegeneration. Transl. Neurodegener. 3:20. doi: 10.1186/2047-9158-3-20
Williams-Gray, C. H., Wijeyekoon, R., Yarnall, A. J., Lawson, R. A., Breen, D. P., Evans, J. R., et al. (2016). Serum immune markers and disease progression in an incident Parkinson’s disease cohort (ICICLE-PD). Mov. Disord. 31, 995–1003. doi: 10.1002/mds.26563
Wu, Y. L., Ding, X. X., Sun, Y. H., Yang, H. Y., and Sun, L. (2013). Methylenetetrahydrofolate reductase (MTHFR) C677T/A1298C polymorphisms and susceptibility to Parkinson’s disease: a meta-analysis. J. Neurol. Sci. 335, 14–21. doi: 10.1016/j.jns.2013.09.006
Xilouri, M., Brekk, O. R., and Stefanis, L. (2016). Autophagy and alpha-synuclein: relevance to Parkinson’s disease and related synucleopathies. Mov. Disord. 31, 178–192. doi: 10.1002/mds.26477
Keywords: Parkinson’s disease, single nucleotide polymorphism, GALC, ZNF184, IL1R2, ELOVL7
Citation: Li G, Cui S, Du J, Liu J, Zhang P, Fu Y, He Y, Zhou H, Ma J and Chen S (2018) Association of GALC, ZNF184, IL1R2 and ELOVL7 With Parkinson’s Disease in Southern Chinese. Front. Aging Neurosci. 10:402. doi: 10.3389/fnagi.2018.00402
Received: 13 August 2018; Accepted: 22 November 2018;
Published: 13 December 2018.
Edited by:
Eminy Hsiao-Yuan Lee, Institute of Biomedical Sciences, Academia Sinica, TaiwanReviewed by:
Paul D. Coleman, Arizona State University, United StatesJih-Ing Chuang, National Cheng Kung University, Taiwan
Copyright © 2018 Li, Cui, Du, Liu, Zhang, Fu, He, Zhou, Ma and Chen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Haiyan Zhou, emhhaXlhbi5jb21AMTYzLmNvbQ== Jianfang Ma, bWpmNzRAMTYzLmNvbQ==; bWFqaWZhQGhvdG1haWwuY29t Shengdi Chen, Y2hlbnNkQHJqaC5jb20uY24=
†These authors have contributed equally to this work