 Chunlin Yue1
Chunlin Yue1 Qian Yu2
Qian Yu2 Yanjie Zhang2
Yanjie Zhang2 Fabian Herold3
Fabian Herold3 Jian Mei1
Jian Mei1 Zhaowei Kong4
Zhaowei Kong4 Stephane Perrey5Jiao Liu6
Stephane Perrey5Jiao Liu6 Notger G. Müller3
Notger G. Müller3 Zonghao Zhang1Yuliu Tao1
Zonghao Zhang1Yuliu Tao1 Arthur Kramer7,8
Arthur Kramer7,8 Benjamin Becker9
Benjamin Becker9 Liye Zou2,10*
Liye Zou2,10*- 1Department of Physical Education, Soochow University, Suzhou, China
- 2Exercise and Mental Health Laboratory, Shenzhen University, Shenzhen, China
- 3Research Group Neuroprotection, German Center for Neurodegenerative Diseases (DZNE), Leipziger, Germany
- 4Faculty of Education, University of Macau, Macao, China
- 5EuroMov Digital Health in Motion, University of Montpellier, Montpellier, France
- 6National-Local Joint Engineering Research Center of Rehabilitation Medicine Technology, Fujian University of Traditional Chinese Medicine, Fuzhou, China
- 7Department of Psychology, Center for Cognitive and Brain Health, Northeastern University, Boston, MA, United States
- 8Beckman Institute, University of Illinois at Urbana-Champaign, Champaign, IL, United States
- 9Ministry of Education (MOE) Key Laboratory for Neuroinformation, The Clinical Hospital of Chengdu Brain Science Institute, University of Electronic Science and Technology of China, Chengdu, China
- 10Institute of Mental Health, Shenzhen University, Shenzhen, China
Objective: The current study aimed at comparing the effects of Tai Chi (a motor-cognitive exercise) with walking (an exercise without cognitive demands) on cognitive performance, brain structure, and brain function in the elderly.
Methods: This cross-sectional study included 42 healthy elderly women within two groups: Tai Chi (n = 20; mean age = 62.90 ± 2.38 years) and brisk walking exercise (n = 22; mean age = 63.27 ± 3.58 years). All the participants underwent a cognitive assessment via the Montreal Cognitive Assessment and brain structural and resting state functional magnetic resonance imaging (rsfMRI) assessments.
Results: Episodic memory in the Tai Chi group was superior to that of the walking group. Higher gray matter density in the inferior and medial temporal regions (including the hippocampus) and higher ReHo in temporal regions (specifically the fusiform gyrus and hippocampus) were found in the Tai Chi group. Significant partial correlations were found between the gray matter density of the left hippocampus and episodic memory in the whole sample. Significant partial correlations were observed between the ReHo in left hippocampus, left parahippocampal, left fusiform, and delayed memory task, which was observed among all subjects.
Conclusion: The present study suggests that long-term Tai Chi practice may improve memory performance via remodeling the structure and function of the hippocampus.
Introduction
In recent decades, the number of older individuals in societies has increased worldwide. In this population, subjective and objective cognitive decline are highly prevalent and both have been associated with an increased risk for developing dementia, particularly in those over 65 years of age (Qiu and Fratiglioni, 2018). Age-related cognitive impairments are often pronounced in the domain of learning and memory, specifically episodic memory that binds personal experience with the spatio-temporal environment (Tromp et al., 2015). As one of the most important cognitive components, episodic memory is associated with the recollection of personal experiences related to when and where an event takes place (Tulving, 1983), which is necessary for both work performance and daily life activity. Episodic memory represents a critical building block for successful everyday functioning and critically relies on the medial temporal lobe structures, specifically the hippocampal formation (Davachi and Wagner, 2002; Becker et al., 2013). Moreover, episodic memory is particularly sensitive to brain aging (Shing et al., 2010; Nyberg et al., 2012), and is usually the first memory system to decline in both normal and pathological aging (Tromp et al., 2015). Additionally, age-dependent episodic memory decline has been observed to be accompanied by volume reduction in the medial temporal and, to a lesser extent, frontal brain regions (Van Petten et al., 2004; Persson et al., 2006; Rajah et al., 2010).
Given the critical roles of memory dysfunction in everyday functioning, including not only cognitive but also social domains (Allen and Fortin, 2013), timely interventions (i.e., physical activity) are needed to postpone or counteract age-related memory decline. Accumulating evidence from different lines of research suggests that physical activity triggers a multitude of neuroprotective and neurorestorative effects on the brain, which in turn promote cognitive performance (Hillman et al., 2008; Yu et al., 2020). For instance, walking—which represents the most common and favored activity among the elderly (Szanton et al., 2015)—has been shown to be associated with better general cognitive functioning, specifically memory, and a decreased likelihood to develop dementia (Yaffe et al., 2001).
Tai Chi, a multicomponent mind-body exercise, combines slow physical activity with relaxation to serve as a movement meditation (Wang et al., 2016). Prior trials suggested that the beneficial effects of Tai Chi are created by a physical component which capitalizes on the benefits of physical exercise and a mind component which additionally promotes psychological well-being, life satisfaction, and improved perception of health (Black et al., 2014; Wang et al., 2016). With respect to cognitive performance, numerous studies have demonstrated the cognitive benefits (including memory) of Tai Chi on a behavioral level (Ji et al., 2017; Sungkarat et al., 2018; Lim et al., 2019; Zou et al., 2019), however, the potential neurobiological processes and relevant brain mechanisms (structure and function) are still not well-understood (Wei et al., 2013; Cui et al., 2019; Chen et al., 2020; Yue et al., 2020a,b). In addition, it is still unclear whether Tai Chi differentiates from purely physical exercise (e.g., walking) with respect to the beneficial effects on cognition and the underlying neurobiological mechanism. In order to better understand the cognitive benefits of Tai Chi in the elderly, we aimed at identifying cognition, brain functional, and structural profiles differences between regular Tai Chi practitioners and well-matched elderly walkers.
Materials and Methods
Study Participants
Participants were recruited in the Suzhou community and the Suzhou Sports Bureau via word-of-mouth advertisements and posters. A total of 200 older individuals were screened according to the following study criteria to identify eligible participants: (i) right-handed as measured by the Edinburgh Handedness Questionnaire (Oldfield, 1971); (ii) normal visual acuity or corrected vision that allowed them to successfully perform Tai Chi; (iii) aged 60 years or above; (vi) successful completion of primary school; (v) normal cognitive performance indicated by a score of 26 or higher in the Montreal Cognitive Assessment Scale (MOCA); and (vi) more than 6 years of Yang-style Tai Chi training with, at least, 5 × 90-min sessions per week and an award in the Yang-style Instruction Certificate (Tai Chi group) or walking exercise for more than 6 years, at least 5 times weekly with each session lasting 90 min (Walking group) (Yue et al., 2020a,b). Volunteers were excluded if they had: (i) diagnosis of a mental disorder (i.e., schizophrenia and/or depression) and/or physical illness (i.e., osteoarthritis and/or osteosarcomas); (ii) a history of neurodegenerative diseases or brain disorder (i.e., Alzheimer’s disease and/or Parkinson’s disease); (iii) substance addiction according to the Diagnostic and Statistical Manual of Mental Disorders-V (DSM-V); (iv) contraindications for MRI (i.e., metallic implants, claustrophobia, pacemakers, or contrast allergy); and (v) participants who regularly performed other types of physical activity besides Tai Chi and/or walking. Forty-six participants were included for magnetic resonance imaging conducted in the Brain Research Center of the Second Affiliated Hospital of Soochow University. Four individuals were excluded from data analysis due to excessive head movement (n = 3) and claustrophobia (n = 1), leading to a final sample size of 20 and 22 in the Tai Chi group (TCG) and walking group (WG), respectively. Data collection was performed by experts who were not aware of the study design and evaluations were all conducted in the same time slot of each testing day. The study was approved by the ethics committee of the university (Approval No. ECSU-2019000209). All participants provided written informed consent and all study procedures were in accordance with the latest revision of the Declaration of Helsinki.
Assessment of Cognitive Functioning
The MOCA, administrated by trained research assistants, was used to determine group differences on global cognition and its subdomains. This assessment tool is a paper and pencil instrument that takes about 10 min to administer and consists of the following measurements: orientation, short-term memory, long-term memory, visuospatial skills, attention, language, verbal fluency, calculation, and abstraction. The MOCA has excellent internal consistency (r = 0.83) and has been validated in 56 languages to assess cognitive performance globally (Nasreddine et al., 2005).
Data Acquisition
A 3 Tesla whole body magnetic resonance imaging (MRI) system (Philips Ingenia) was employed to collect the data. All volunteers were informed that they should not participate in any physical activity before lab assessment. For each participant, we conducted one functional MRI resting-state assessment using a gradient echo planar imagine (EPI) sequence with the following scan parameters (FOV: 220 mm × 220 mm, TR: 2,000 ms, TE: 30 ms, slices: 36, flip: = 90°, slice thickness: 4 mm, matrix size: 64 × 64, total scan time: 400 s). For the brain structural assessments each participant additionally underwent acquisition of T1-weighted images using a magnetization-prepared rapid gradient echo sequence (MPRAGE) with the scan parameters: voxel size: 0.625 × 0.625 × 1 mm, TR: 7.1 ms, TE: 2.98 ms, flip: 9°, slice thickness: 1.0 mm, FOV: 256 × 256 mm, matrix size: 256 × 256, scanning layer by layer. MRI acquisition was conducted by an experienced radiologist. Each participant was asked to lie comfortably and to think of nothing in particular while moving their head and body as little as possible. Participants were explicitly requested to stay awake during data acquisition.
Data Processing
Voxel-Based Morphometry (VBM)
Structural MRI data was analyzed and processed in Matlab 2013b (The Mathworks®, Natick, MA, United States). SPM12 (Statistical Parametric Mapping)1 and CAT12 (Computational Anatomy Toolbox)2 were used to integrate with Matlab 2013b. The recommended default parameters were employed to preprocess the structural image data. The 3D structural image of each participant was initially segmented into gray matter (GM), white matter (WM), and cerebrospinal fluid (CSF). Voxel resolution was set to 2∗2∗2 mm. Spatial smoothing was performed using a Gaussian kernel with a full width at half maxima (FWHM) of 8 mm. To avoid a partial density effect in the boundary between GM and WM, all voxels with a GM value below 0.2 were excluded. Total intracranial volume (TIV) was calculated and considered as a covariable in all further statistical analysis.
Regional Homogeneity (ReHo) Analysis
The RESTplus software was used to preprocess the resting-state fMRI data in Matlab2013b (Jia et al., 2019). Data preprocessing included standard processing steps, such as: (1) deletion of the first 1-time points, and the data of 190 time points in the resting-state were retained; (2) time layer and head motion correction according to the realign curve. Participants with head motion at x, y, and z-axis translation greater than 3 mm and rotation greater than 3° were excluded; (3) registration of the motion-corrected to the Montreal Neurological Institute (MNI) space via normalization of the functional time-series MNI EPI template to account for interindividual anatomical differences; (4) a linear regression was used to remove linear drift, white matter signals, CSF signals, and 24 head parameters were regressed as covariables to remove physiological influences (Murphy et al., 2009); and (5) temporal band-pass filtering (0.01–0.08 Hz). Based on Kendall’s coefficient of Concordance (KCC), the ReHo value of each voxel was subsequently calculated using RESTplus software to obtain whole brain ReHo maps of each subject. For standardization purposes, each ReHo map was divided by the average ReHo value of the entire brain. A Gaussian kernel of 8 mm FWHM was used for spatial smoothing (Zang et al., 2004).
Statistical Analyses
The SPSS 19.0 software (SPSS Inc., Chicago, IL, United States) was used for data analysis. A T-test was performed to compare differences between measures obtained from the Tai Chi and walking groups, and the level of significance was set at 0.05. Cohen’s d (effect size) was classified as small (d = 0.2), medium (d = 0.5), and large (d ≥ 0.8). The independent-samples T-test in SPM software was used to analyze the differences on GM and ReHo between the Tai Chi and walking groups. Analysis with a threshold of voxel-level p < 0.001 and cluster-level p < 0.01 GRF (Gauss Random Field) correction was applied for the ReHo and VBM analyses. A partial correlation analysis was used to compare the correlation between sub-items of MOCA and GM and ReHo, with age (years), sex, and education (years) as covariates. We used the Bonferroni correction to control for multiple comparisons. The correlation coefficients were defined as follows: 0.00–0.19 no correlation; 0.20–0.39 low correlation; 0.40–0.59 moderate correlation; 0.60–0.79 moderately high correlation; and 0.8 high correlation (Zhu, 2012). The Xjview3 (Xia et al., 2013) and Origin 9 software were used to present the results.
Results
Independent sample T-tests revealed no statistically significant differences between the two groups for age, years of education, or global cognitive performance (operationalized by MOCA scores) (details see Table 1). With respect to the cognitive sub-domains, a significant difference was only observed in the delayed recall test, with better memory function in the Tai Chi group relative to the walking group (details see Table 2).
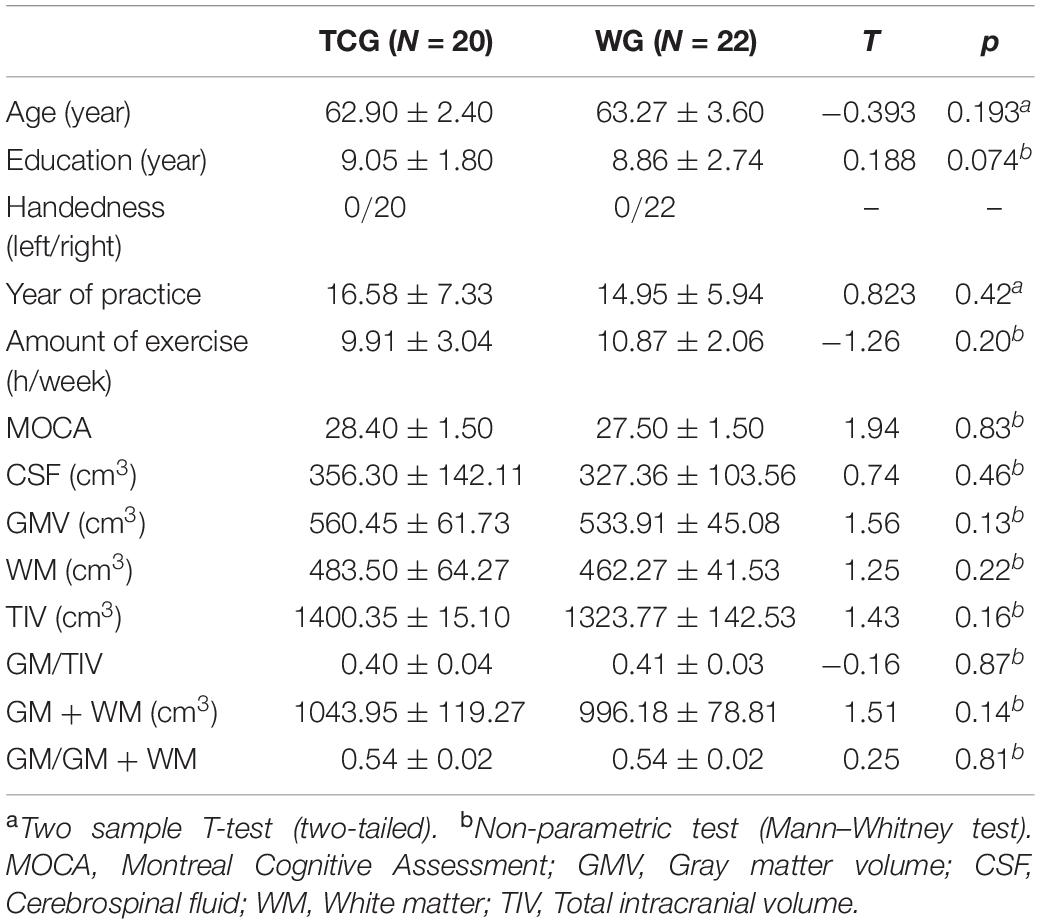
Table 1. Characteristics of the participants in TCG and WG.
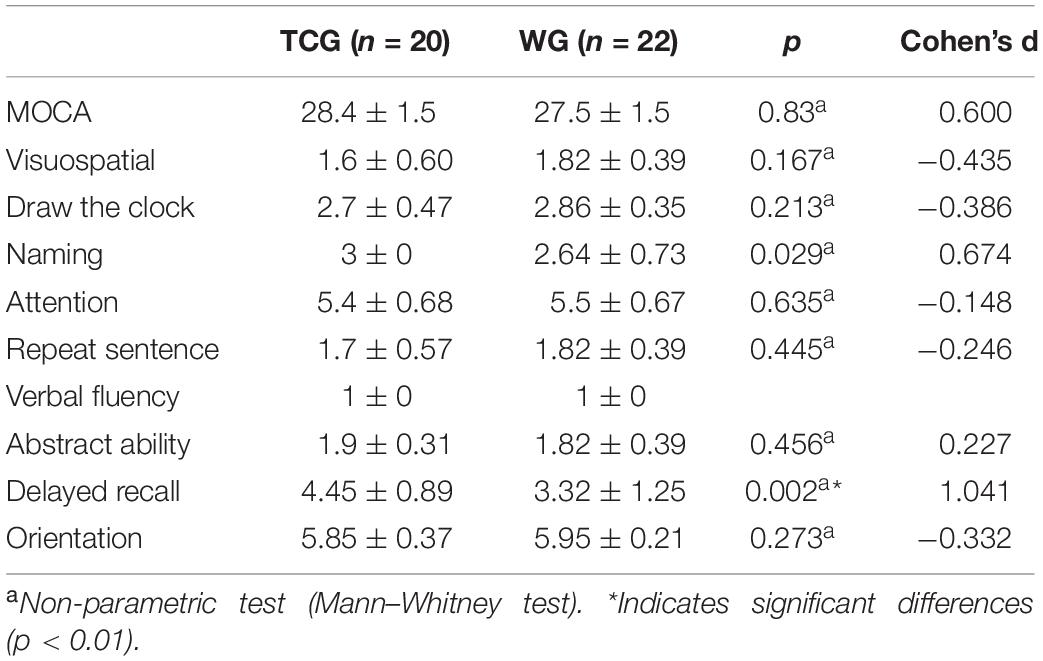
Table 2. Comparisons of cognitive domains of the MOCA between TCG and WG.
VBM Analysis
The VBM analysis of the T1-weighted structural images showed that gray matter density in the left cerebellum and right inferior and left medial temporal lobe regions, including the left hippocampus and parahippocampal gyrus, was significantly higher in the TCG group as compared to the WG group (Table 3 and Figure 1). Then, a whole-brain voxel-wise between-subject ANOVA on the VBM maps in two Groups (TCG, WG) was performed to detect regions showing intervention-related differences. The threshold for significant difference was set to p < 0.01 cluster mass-level GFR corrected with a cluster building threshold of uncorrected p = 0.001 on voxel level. In further studies, the regions showing some significant differences were defined as regions of interest (ROIs). We extracted the mean gray matter density signal in each ROI and used partial correlation analysis to examine the correlation between gray matter density signal and delayed memory scores of the two groups.
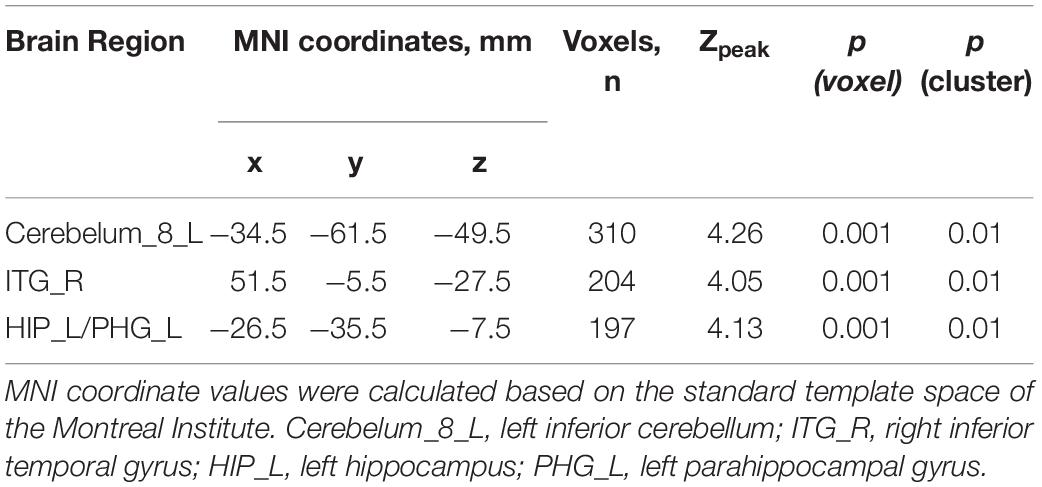
Table 3. Brain regions with higher gray matter density in TCG as compared to WG.
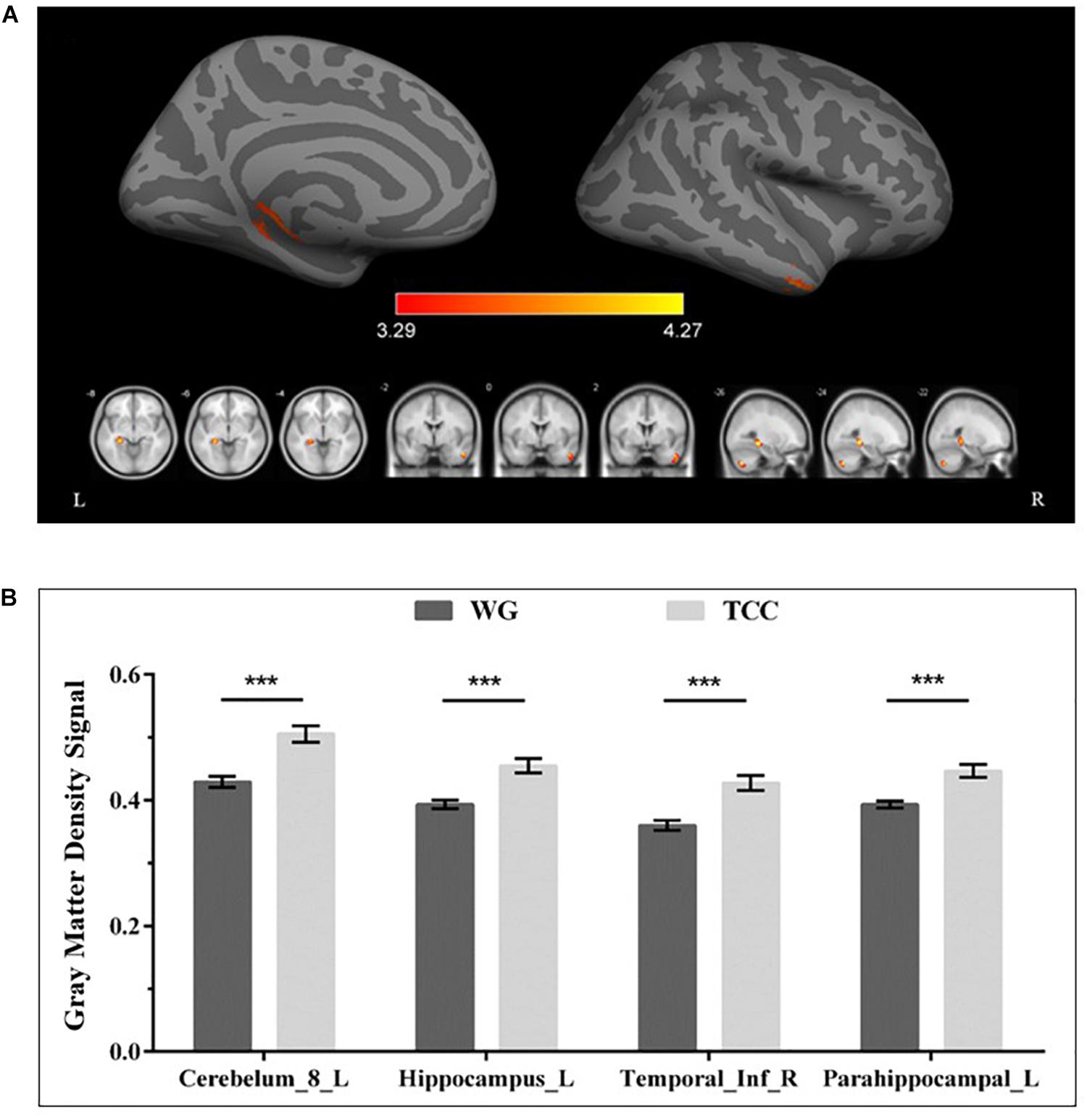
Figure 1. Regional gray matter differences as determined by VBM between the TCG and WG group. (A) The threshold for significant changes was set to p < 0.01 cluster mass-level GFR corrected with a cluster building threshold of p = 0.001 uncorrected on voxel level. The warm color indicates that the GMV of the TCG was greater than that of WG. (B) Bar plots showed the mean gray matter density signal in these ROIs for the TCG and WG group. The independent sample t-test was used to compare the effects of two groups. ***means p < 0.001. Cerebelum_8_L, left inferior cerebellum; ITG_R, right inferior temporal gyrus; HIP_L, left hippocampus; PHG_L, left parahippocampal gyrus.
Reho Analysis
Resting-state fMRI analysis revealed significantly higher regional ReHo activation in the TCG group in the left medial temporal lobe, specifically the hippocampus and parahippocampus, as well as the fusiform gyrus relative to the WG group (Table 4 and Figure 2). Then, a whole-brain voxel-wise between-subject ANOVA on the ReHo maps in two Groups (TCG, WG) was performed to detect regions showing intervention-related differences. The threshold for significant difference was set to p < 0.01 cluster mass-level GFR corrected with a cluster building threshold of uncorrected p = 0.001 on voxel level. In further studies, the regions showing some significant differences were defined as regions of interest (ROIs). We extracted the mean ReHo value in each ROI and used partial correlation analysis to examine the correlation between ReHo and delayed memory scores of the two groups.

Table 4. Brain regions with different ReHo in TCG and WG.
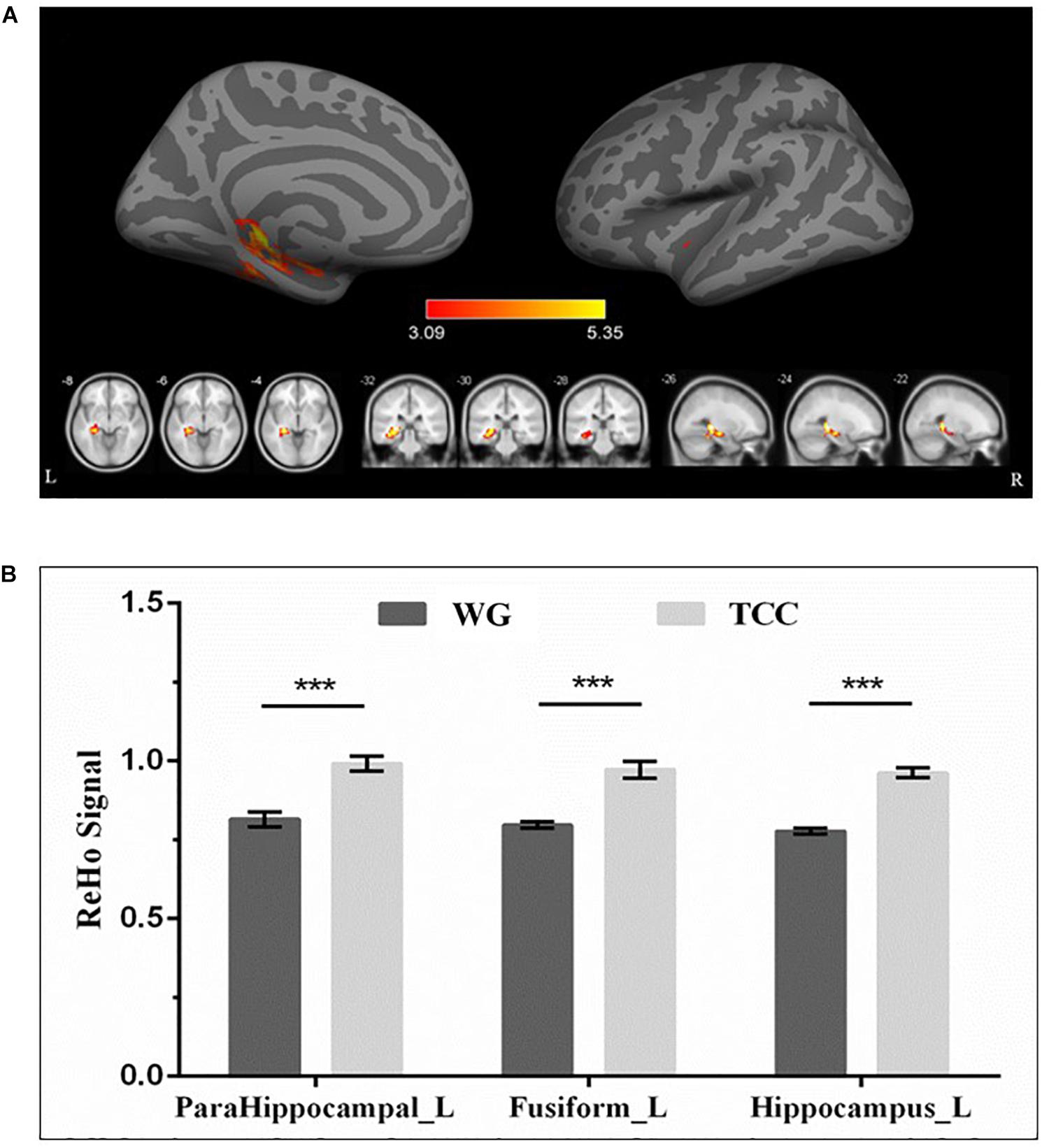
Figure 2. Regional differences in ReHo between TCG and WG. (A) ReHo analysis showed that the ReHo activations of the left hippocampus, parahippocampal gyrus, and fusiform in the participants who practiced Tai Chi were higher than those in the WG. The threshold for significant changes was set to p < 0.01 cluster mass-level GFR corrected with a cluster building threshold of p = 0.001 uncorrected on voxel level. (B) Bar plots showed the mean ReHo signal in these ROIs for the TCG and WG group. The independent sample t-test was used to compare the effects of two groups. ***means p < 0.001. HIP_L, left hippocampus; PHG_L, left parahippocampal gyrus; FG_L, left fusiform.
Correlation Between Brain Structure, Brain Function, and Behavioral Performance
Partial correlation analyses (controlling for the participants’ age and level of education) was conducted on the four regions of interest (ROI) (left inferior cerebellum, right inferior temporal gyrus, left hippocampus, and parahippocampal gyrus) obtained by VBM analysis and eight domains of MOCA (Visuospatial, Draw the clock, Naming, Attention, Repeat sentence, Abstract ability, Delayed recall, and Orientation). As shown in Figure 3, we observed a significant moderate and positive correlation between the gray matter density in the left hippocampus specifically for the delayed memory task (r = 0.547, p < 0.01, Bonferroni correction) at a Bonferroni correction threshold of 0.0014 (0.05/36) when the whole sample was considered (Ludbrook, 1998). In the WG, the correlation between gray matter density in left hippocampus and delayed memory task failed to reach statistical significance at the Bonferroni corrected level (see Statistical analysis) and thus could be considered as only marginally significant (r = 0.613, ps < 0.1, Bonferroni correction; see Figure 3). On the other hand, no significant correlation between the gray matter density changes in the left hippocampus and delayed memory task was found in the TCG (r = 0.241, p = 0.307; see Figure 3; Ludbrook, 1998). Additionally, there was no significant correlation between the gray matter density of the remaining 2 ROI and other sub-items of the MOCA.
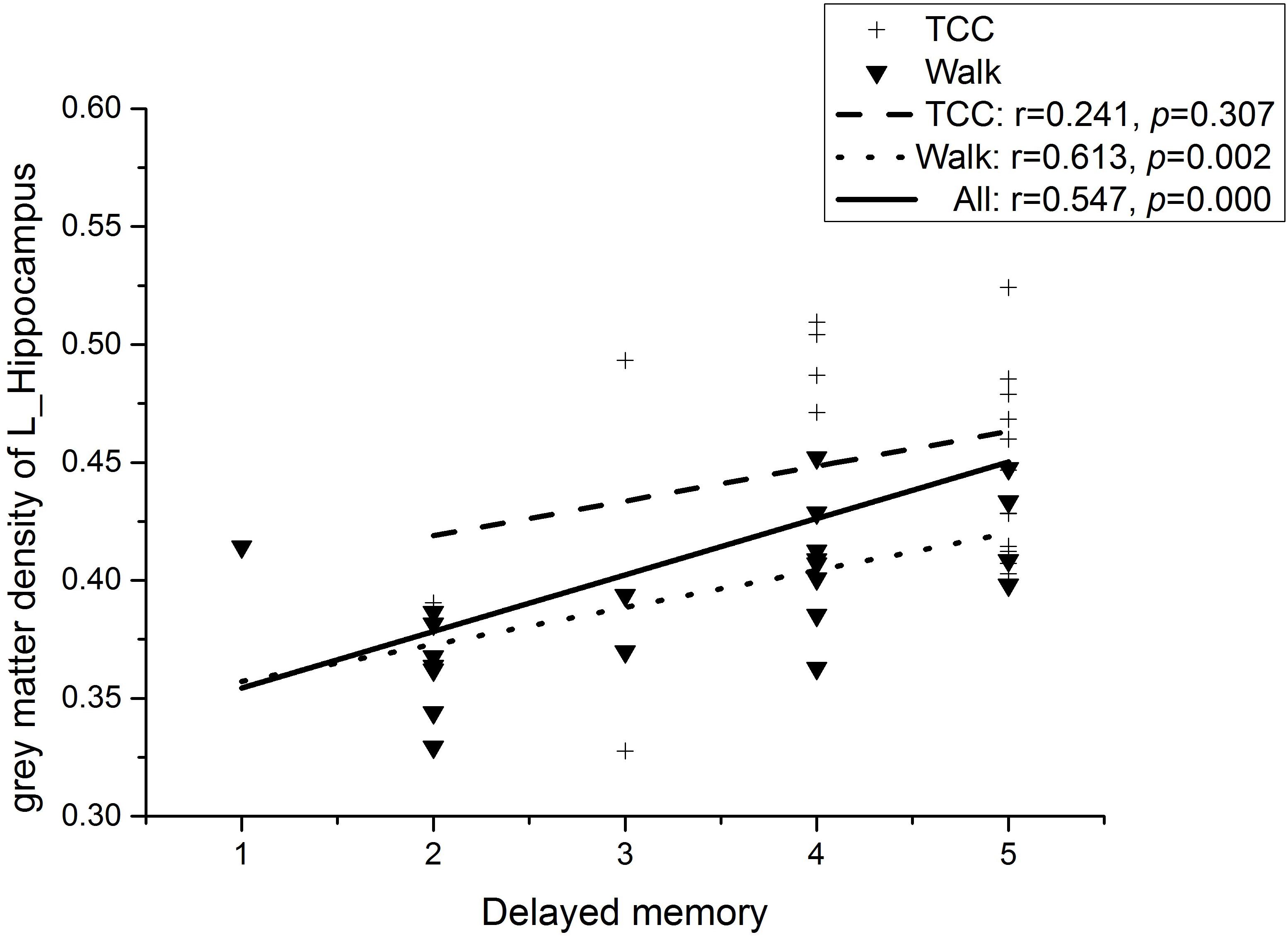
Figure 3. Correlation between gray matter density of the left hippocampal and delayed memory.
Partial correlation analysis was conducted on the three regions of interest (ROI) (the left hippocampus, parahippocampal gyrus, and fusiform) obtained by ReHo analysis and eight sub-items of MOCA (Visuospatial, Draw the clock, Naming, Attention, repeat sentence, Abstract ability, Delayed recall, and Orientation) with controlling the participants’ age and education (Ludbrook, 1998). A moderately high and positive correlation between the ReHo in the left hippocampus (r = 0.679, p < 0.01, Bonferroni correction), left parahippocampal (r = 0.612, p < 0.01, Bonferroni correction), left fusiform (r = 0.570, p < 0.01, Bonferroni correction), and delayed memory task was observed when the whole sample was considered at a Bonferroni correction threshold of 0.0019 (0.05/27). In the WG, a significant moderately high and marginally significant correlation between the ReHo in the left hippocampus and delayed memory task was observed (r = 0.619, ps < 0.1, Bonferroni correction; see Figure 4). On the other hand, no significant correlation between the ReHo changes in the left hippocampus (r = 0.507, p = 0.022; see Figure 4), left parahippocampal (r = 0.448, p = 0.048; see Figure 4), left fusiform (r = 0.450, p = 0.047; see Figure 4), and delayed memory task was found in the TCG (Ludbrook, 1998).
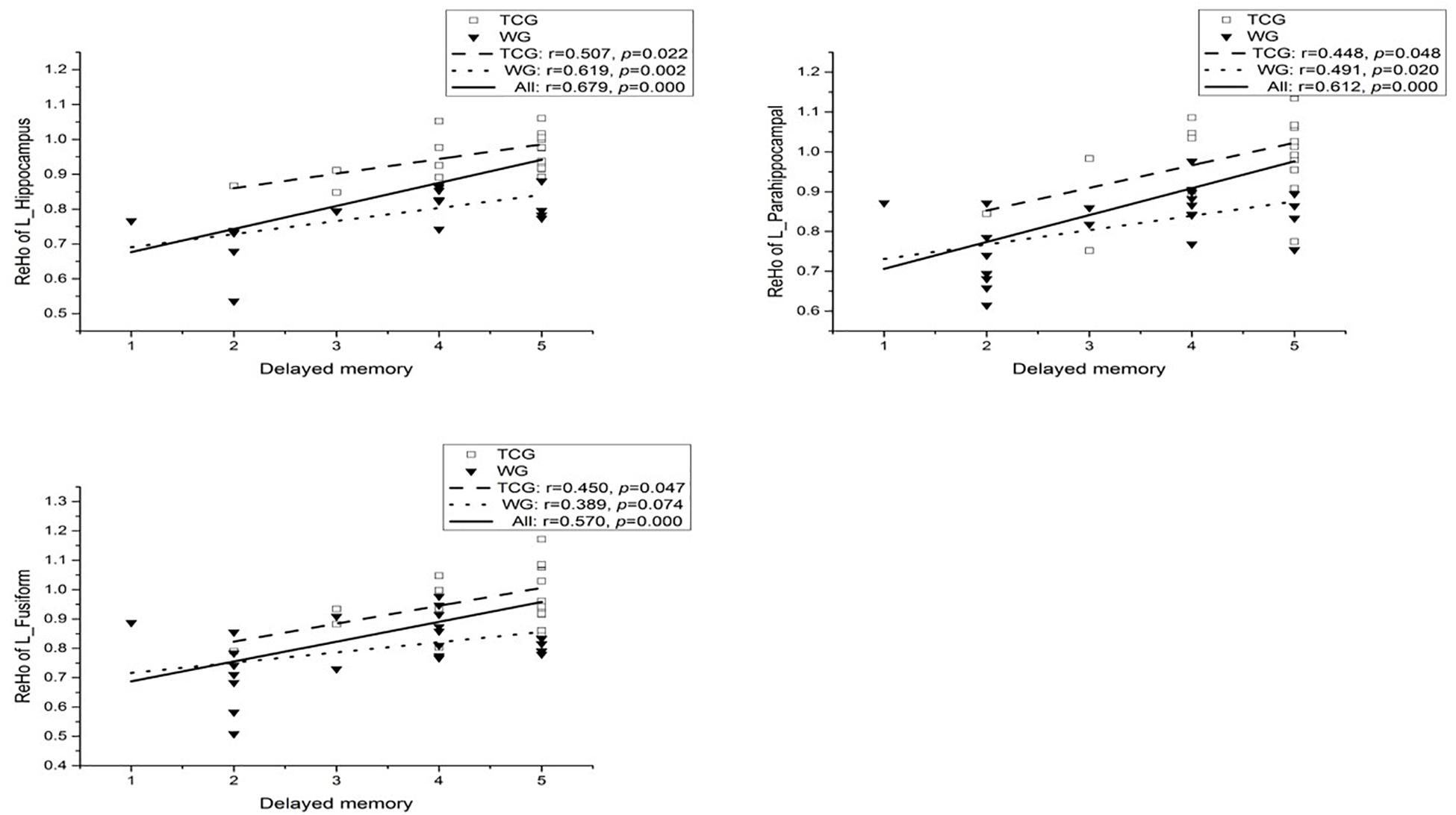
Figure 4. Correlation between ReHo of the left hippocampal and delayed memory.
Discussion
The current study employed a cross-sectional design to determine differences in cognitive performance, brain structure, and functional alterations between long-term elderly practitioners of Tai Chi and walking exercises. Compared with the walking group, Tai Chi practitioners performed better in the domain of delayed memory and additionally exhibited higher gray matter density in temporal regions, including the left hippocampus and the adjacent left parahippocampal gyrus. On the functional level, the Tai Chi practitioners exhibited higher spontaneous ReHo activation in similar temporal regions, including the left hippocampus and parahippocampal gyrus, as well as the fusiform gyrus, as compared to the walking group. Finally, significant associations were observed between the left hippocampal gray matter density and delayed memory performance in the whole sample level, confirming an important contribution of this region to memory performance in the present sample. In particular, imaging findings further support the notion that individuals who regularly engaged in exercise that involves more cognitive loads and demands may gain superior benefits (Tsai et al., 2017; Gu et al., 2019). In the following sections, we will discuss these findings in more detail.
Hippocampus: Structural Changes, Functional Changes, and Relationship With Memory
Long-term Tai Chi practitioners showed higher density and ReHo activation in the hippocampus and parahippocampal gyrus as compared to the WG. Moreover, on a behavioral level, we observed that long-term Tai Chi practice was more effective than walking exercise in enhancing selected aspects of cognitive function (i.e., long-term memory), which is in accordance with previous observations (Ji et al., 2017). This observation fits well with the observations of a positive association between better memory performance and higher density of the hippocampus (Erickson et al., 2011). The episodic memory performance reflects the ability to recollect past experience in the temporal and spatial context, which is highly influenced by the structure of the temporal lobe (especially in hippocampus) (Grilli et al., 2018). More specifically, this brain area is critically engaged in consolidation and retrieval, particularly with associated information. Aging-related hippocampal shrinkage (atrophy) has been increasingly reported and has been associated with memory impairments and a strongly increased risk for the development of dementia and Alzheimer’ disease (Jack et al., 2010; Small et al., 2011). Previous studies consistently indicated that hippocampal neuroplasticity (neurogenesis) could be induced by regular physical exercise, especially physical training (Cotman et al., 2007; de Meireles et al., 2019). Such exercise-induced biochemical changes in the brain region have been observed to associate with: (1) vascular endothelial growth factor (VEGF), which is beneficial for blood vessel survival and growth; (2) insulin-like growth factor (IGF)-1 that can contribute to several neural and angiogenic process; and (3) increased serum levels of brain-derived neurotrophic factor (BDNF) that is conducive to long-term potentiation and proliferation of neurons (Cotman et al., 2007; de Meireles et al., 2019). For example, a randomized controlled trial by Erickson et al. (2011) indicated that 1-year aerobic exercise training effectively contributed to a 2% increase of hippocampal volume that corresponded to improved spatial memory performance. This increased size in this brain region was also positively associated with serum levels of BDNF, whereas older adults in the control group demonstrated nearly 1.5% hippocampal volume loss within this intervention period. Additionally, exercise may attenuate or even improve the aging-induced memory decline via histone modifications and alteration of gene expression (Elsner et al., 2013; Lovatel et al., 2013; de Meireles et al., 2019). Age-related epigenetic imbalances have been observed to associate with declined memory performance (Elsner et al., 2013; de Meireles et al., 2019). For example, decreased H3K9 methylation in the hippocampus have been found in aged rats; BDNF, cFOS, and deoxyribonucleic-acid-methyltransferase-3a (DNMT3A), all of which were associated with cognitive impairment during the aging process, also undergo expression alterations (de Meireles et al., 2019). Notably, it is reported that exercise could improve memory performance through attenuating the process of epigenetic modifications of histones via gene promoters (BDNF, cFos, and DNMT3A) (de Meireles et al., 2019).
Additionally, a seminal study found that a 10 min bout of mild exercise effectively improved pattern separation and functional connectivity between the hippocampal dentate gyrus/CA2 and surrounding brain regions and such objectively measured results were positively associated with memory performance (Suwabe et al., 2018). Although the above-summarized evidence suggests that the observed greater hippocampal density could be triggered by long-term Tai-Chi practice, our cross-sectional study design allows limited direct conclusions with respect to causal relationships and thus further longitudinal studies are required to confirm this assumption empirically. Furthermore, it is widely accepted that ReHo of the resting-state signals play a critical role in cognitive function (including learning and memory) (Zang et al., 2004). Age-associated reduction of ReHo in several brain regions was negatively associated with behavioral outcomes (Diciotti et al., 2017). Recently, a combined program that consisted of cognitive training, Tai Chi, and group counseling led to a reorganization of ReHo patterns in several brain regions (including the superior and middle temporal gyri), which was coincident with improved cognitive function after the 6-week intervention period (Zheng et al., 2015). Given that we observed greater ReHo of the left hippocampal area in the Tai-Chi group and that we noticed a positive neurobehavioral relationship between ReHo patterns and memory performance, it seems to be reasonable to assume that engaging in long-term Tai Chi exercise (in comparison to walking) is a valuable strategy to prevent decline in cognitive performance as it preserves underlying neural correlates.
Other Related Brain Regions in This Study
As compared to the WG, greater gray matter density in the TCG was also observed in other brain regions, including the parahippocampal gyrus, cerebellum, and inferior temporal gyrus; higher ReHo activations were also found in the parahippocampal gyrus and fusiform.
The parahippocampal gyrus, located just inferior to the hippocampus, plays an important and distinctive role in the processing of memory (Nemanic et al., 2004). The parahippocampal gyrus is comprised of the entorhinal, perirhinal, and parahippocampal cortices (Burwell, 2000); cognitive information is usually collected by the perirhinal and parahippocampal cortices before being further processed in the entorhinal cortex and hippocampus (Loprinzi, 2019). While the entorhinal cortex and entorhinal cortex process spatial and object-recognition information (Raslau et al., 2015), the parahippocampal cortex has a critical role in episodic memory (Aminoff et al., 2013). Moreover, in previous studies (Nemanic et al., 2004; Siddarth et al., 2018; Loprinzi, 2019), extensive memory-related effects (i.e., increased neural excitability, blood flow, regional glucose metabolism, gray/white matter, and functional connectivity) induced by physical exercise have been reported in the parahippocampal gyrus, which implies that exercise interventions like Tai Chi may facilitate the memory function through regulation of the parahippocampal gyrus at molecular, structural, and functional levels.
The major function of the cerebellum was believed to be efficiently coordinating the timing and forces of multiple muscle groups to initiate voluntary movements, according to both anatomic and clinical evidence before the 1990s. However, later findings have cast doubt on this view. Cerebellar activation has been increasingly investigated using imaging techniques and found to associate with cognitive and affective tasks. Furthermore, among the patients whose injuries place limits on the cerebellum, a variety of non-motor symptoms have been recognized, especially those with cognitive affective syndrome (Schmahmann and Sherman, 1998; Doya, 2000). Evidence from functional mapping of the cerebellum showed that more than half of the cerebellar cortex is connected to the cognition-related areas of the cerebral cortex (Buckner et al., 2011). It is worth noting that the anterior cerebellum (lobules I–IV and V) and anterior part of lobule VI are mainly related with motor functions (Nitschke et al., 1996; Rijntjes et al., 1999; Grodd et al., 2001), while the posterior part plays an evident role in neurocognition and emotion (Eekers et al., 2018). Thus, the cerebellum may contribute to physical and cognitive effects induced by Tai Chi through different regions and connections.
Regarding the fusiform gyrus, a previous study using MRI indicated that the fusiform gyrus area as part of the human brain is responsible for face-specific processing. Furthermore, greater activation in this brain region was observed during facial stimuli as compared to objects (McCarthy et al., 1997). Haxby and colleague proposed a model for face perception and emphasized the distributed human neural system. Specifically, the face-responsive region in the fusiform gyrus was observed to explain more for the unchanging features of a specified facial identity, whereas the face-responsive region in the superior temporal sulcus could be involved in changeable aspects (name) of faces (Haxby et al., 2000). Thus, the fusiform gyrus plays a critical role in social communication or interaction that is associated with cognitive benefits. This assumption is supported by an experimental study suggesting that community-based Tai Chi training facilitates social interaction, which, in turn, might contribute to the observed increase in brain volume and behavioral performance in non-demented older adults (Mortimer et al., 2012).
Regarding the temporal gyrus, its primary function is connected with visual stimuli processing (Lafer-Sousa and Conway, 2013), which is involved with memory, perception, and recognition (Bryan Kolb, 2014). Previous studies believed that the temporal gyrus works in memory processing through cooperation with other brain regions like the hippocampus (key for storing the memory), parahippocampal gyrus (participation in differentiating between scenes and objects), and fusiform gyrus (dealing with facial and body recognition) (Spiridon et al., 2006; Gross, 2008). Additionally, in a study by Makizako et al. (2013), evidence suggested that a higher exercise capacity is associated with better performance in logical and visual memory, and poor memory function was correlated with decreased gray matter volume in the temporal gyrus, hippocampus, and occipital gyrus. These outcomes indicated that exercise (like Tai Chi) may improve visual-stimuli-related memory through maintenance of the gray matter volume of the hippocampus (main region for memory), temporal gyrus, and other brain regions, which is consistent with findings in our study.
Strengths and Limitations
The strengths of this investigation are the high practical relevance of our findings as we used state-of-art neuroimaging to investigate the effects of popular physical exercises (Tai-Chi and walking) in a cohort vulnerable to cognitive decline. Furthermore, we applied rigorous inclusion criteria, allowing us to identify a cohort of long-term practitioners of Tai-Chi and walking who have at least 10 years of experience. One limitation of this investigation, however, is that we did not account for some other potentially important factors (e.g., cardiorespiratory fitness and body mass index) that might bias our findings. For instance, there is evidence in the literature suggesting that cardiorespiratory fitness level is associated with hippocampal volume (Erickson et al., 2009; Szabo et al., 2011) although this finding is not universal (Niemann et al., 2014). Additionally, as only elderly women were included in this study, it is unknown whether our outcomes are applicable to people with different genders.
Conclusion
In conclusion, in this cross-sectional study, we demonstrated that, relative to walking, Tai Chi was more effective in enhancing episodic memory in a sample of healthy older Chinese women. The observed changes in hippocampal structure and function and significant behavioral relationships between hippocampal alteration and memory functions suggest that physical activity like Tai Chi is likely to benefit memory function (extremely episodic memory) via remodeling of the hippocampal structure and function. Furthermore, our findings support the conclusion that engaging in Tai-Chi might be superior in delaying cognitive decline in comparison to walking training. Future cross-sectional and longitudinal research should also consider accounting for a number of moderating factors (e.g., fitness level in cardiorespiratory, muscular and motor dimension, frequency, intensity and duration of relevant activities, whether activities are performed alone or with friends, etc.) and to evaluate additional neurobiological mechanisms (e.g., brain metabolism, muscle energetics, reduced oxidative stress, reduced psychological stress) through which Tai Chi may influence cognitive performance (Gatts, 2008; Goon et al., 2009; Guo et al., 2014; Cheung et al., 2018; Zhou et al., 2018).
Data Availability Statement
All datasets generated for this study are included in the article/supplementary material.
Ethics Statement
The studies involving human participants were reviewed and approved by the Review Board of Soochow University. The patients/participants provided their written informed consent to participate in this study. Written informed consent was obtained from the individual(s) for the publication of any potentially identifiable images or data included in this article.
Author Contributions
CY and LZ designed the study. CY, ZZ, JM, and YT collected and analyzed the data. CY, LZ, YZ, QY, and JM wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Social Science Foundation (Grant No. 13CTY038) and the APC was funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Footnotes
- ^ https://www.fil.ion.ucl.ac.uk/spm/
- ^ http://www.neuro.uni-jena.de/cat12/CAT12
- ^ http://www.nitrc.org/projects/bnv
References
Allen, T. A., and Fortin, N. J. (2013). The evolution of episodic memory. Proc. Natl Acad. Sci. U S A 110(Suppl. 2), 10379–10386. doi: 10.1073/pnas.1301199110
Aminoff, E. M., Kveraga, K., and Bar, M. (2013). The role of the parahippocampal cortex in cognition. Trends Cogn. Sci. 17, 379–390. doi: 10.1016/j.tics.2013.06.009
Becker, B., Wagner, D., Koester, P., Bender, K., Kabbasch, C., Gouzoulis-Mayfrank, E., et al. (2013). Memory-related hippocampal functioning in ecstasy and amphetamine users: a prospective fMRI study. Psychopharmacology 225, 923–934. doi: 10.1007/s00213-012-2873-z
Black, D. S., Irwin, M. R., Olmstead, R., Ji, E., Crabb Breen, E., and Motivala, S. J. (2014). Tai chi meditation effects on nuclear factor-kappaB signaling in lonely older adults: a randomized controlled trial. Psychother. Psychosom. 83, 315–317. doi: 10.1159/000359956
Buckner, R. L., Krienen, F. M., Castellanos, A., Diaz, J. C., and Yeo, B. T. (2011). The organization of the human cerebellum estimated by intrinsic functional connectivity. J. Neurophysiol. 106, 2322–2345. doi: 10.1152/jn.00339.2011
Burwell, R. D. (2000). The parahippocampal region: corticocortical connectivity. Ann. N Y Acad. Sci. 911, 25–42. doi: 10.1111/j.1749-6632.2000.tb06717.x
Chen, L. Z., Yuan, X., Zhang, Y., Zhang, S., Zou, L., Yang, L., et al. (2020). Brain Functional Specialization Is Enhanced Among Tai Chi Chuan Practitioners. Arch. Phys. Med. Rehabil. 101, 1176–1182. doi: 10.1016/j.apmr.2020.02.005
Cheung, T. C. Y., Liu, K. P. Y., Wong, J. Y. H., Bae, Y. H., Hui, S. S., Tsang, W. W. N., et al. (2018). Acute Effects of Tai Chi Training on Cognitive and Cardiovascular Responses in Late Middle-Aged Adults: A Pilot Study. Evid. Based Compl. Alternat Med. 2018, 7575123. doi: 10.1155/2018/7575123
Cotman, C. W., Berchtold, N. C., and Christie, L. A. (2007). Exercise builds brain health: key roles of growth factor cascades and inflammation. Trends Neurosci. 30, 464–472. doi: 10.1016/j.tins.2007.06.011
Cui, L., Yin, H., Lyu, S., Shen, Q., Wang, Y., Li, X., et al. (2019). Tai Chi Chuan vs General Aerobic Exercise in Brain Plasticity: A Multimodal MRI Study. Sci. Rep. 9:17264. doi: 10.1038/s41598-019-53731-z
Davachi, L., and Wagner, A. D. (2002). Hippocampal contributions to episodic encoding: insights from relational and item-based learning. J. Neurophysiol. 88, 982–990. doi: 10.1152/jn.2002.88.2.982
de Meireles, L. C. F., Galvao, F. Jr., Walker, D. M., Cechinel, L. R., de Souza Grefenhagen, A. I., Andrade, G., et al. (2019). Exercise Modalities Improve Aversive Memory and Survival Rate in Aged Rats: Role of Hippocampal Epigenetic Modifications. Mol. Neurobiol. 56, 8408–8419. doi: 10.1007/s12035-019-01675-w
Diciotti, S., Orsolini, S., Salvadori, E., Giorgio, A., Toschi, N., Ciulli, S., et al. (2017). Resting state fMRI regional homogeneity correlates with cognition measures in subcortical vascular cognitive impairment. J. Neurol. Sci. 373, 1–6. doi: 10.1016/j.jns.2016.12.003
Doya, K. (2000). Complementary roles of basal ganglia and cerebellum in learning and motor control. Curr. Opin. Neurobiol. 10, 732–739. doi: 10.1016/s0959-4388(00)00153-7
Eekers, D. B. P., In ’t Ven, L., Deprez, S., Jacobi, L., Roelofs, E., Hoeben, A., et al. (2018). The posterior cerebellum, a new organ at risk? Clin. Transl. Radiat. Oncol. 8, 22–26. doi: 10.1016/j.ctro.2017.11.010
Elsner, V. R., Lovatel, G. A., Moyses, F., Bertoldi, K., Spindler, C., Cechinel, L. R., et al. (2013). Exercise induces age-dependent changes on epigenetic parameters in rat hippocampus: a preliminary study. Exp. Gerontol. 48, 136–139. doi: 10.1016/j.exger.2012.11.011
Erickson, K. I., Prakash, R. S., Voss, M. W., Chaddock, L., Hu, L., Morris, K. S., et al. (2009). Aerobic fitness is associated with hippocampal volume in elderly humans. Hippocampus 19, 1030–1039. doi: 10.1002/hipo.20547
Erickson, K. I., Voss, M. W., Prakash, R. S., Basak, C., Szabo, A., Chaddock, L., et al. (2011). Exercise training increases size of hippocampus and improves memory. Proc. Natl. Acad. Sci. U S A 108, 3017–3022. doi: 10.1073/pnas.1015950108
Gatts, S. (2008). Neural mechanisms underlying balance control in Tai Chi. Med. Sport Sci. 52, 87–103. doi: 10.1159/000134289
Goon, J. A., Aini, A. H., Musalmah, M., Anum, M. Y., Nazaimoon, W. M., and Ngah, W. Z. (2009). Effect of Tai Chi exercise on DNA damage, antioxidant enzymes, and oxidative stress in middle-age adults. J. Phys. Act. Health 6, 43–54. doi: 10.1123/jpah.6.1.43
Grilli, M. D., Wank, A. A., Bercel, J. J., and Ryan, L. (2018). Evidence for Reduced Autobiographical Memory Episodic Specificity in Cognitively Normal Middle-Aged and Older Individuals at Increased Risk for Alzheimer’s Disease Dementia. J. Int. Neuropsychol. Soc. 24, 1073–1083. doi: 10.1017/s1355617718000577
Grodd, W., Hulsmann, E., Lotze, M., Wildgruber, D., and Erb, M. (2001). Sensorimotor mapping of the human cerebellum: fMRI evidence of somatotopic organization. Hum. Brain Mapp. 13, 55–73. doi: 10.1002/hbm.1025
Gross, C. G. (2008). Single neuron studies of inferior temporal cortex. Neuropsychologia 46, 841–852. doi: 10.1016/j.neuropsychologia.2007.11.009
Gu, Q., Zou, L., Loprinzi, P. D., Quan, M., and Huang, T. (2019). Effects of Open Versus Closed Skill Exercise on Cognitive Function: A Systematic Review. Front. Psychol. 10:1707. doi: 10.3389/fpsyg.2019.01707
Guo, L. Y., Yang, C. P., You, Y. L., Chen, S. K., Yang, C. H., Hou, Y. Y., et al. (2014). Underlying mechanisms of Tai-Chi-Chuan training for improving balance ability in the elders. Chin. J. Integr. Med. 20, 409–415. doi: 10.1007/s11655-013-1533-4
Haxby, J. V., Hoffman, E. A., and Gobbini, M. I. (2000). The distributed human neural system for face perception. Trends Cogn. Sci. 4, 223–233. doi: 10.1016/s1364-6613(00)01482-0
Hillman, C. H., Erickson, K. I., and Kramer, A. F. (2008). Be smart, exercise your heart: exercise effects on brain and cognition. Nat. Rev. Neurosci. 9, 58–65. doi: 10.1038/nrn2298
Jack, C. R. Jr., Wiste, H. J., Vemuri, P., Weigand, S. D., Senjem, M. L., Zeng, G., et al. (2010). Brain beta-amyloid measures and magnetic resonance imaging atrophy both predict time-to-progression from mild cognitive impairment to Alzheimer’s disease. Brain 133, 3336–3348. doi: 10.1093/brain/awq277
Ji, Z., Li, A., Feng, T., Liu, X., You, Y., Meng, F., et al. (2017). The benefits of Tai Chi and brisk walking for cognitive function and fitness in older adults. PeerJ. 5:e3943. doi: 10.7717/peerj.3943
Jia, X.-Z., Wang, J., Sun, H.-Y., Zhang, H., Liao, W., Wang, Z., et al. (2019). RESTplus: an improved toolkit for resting-state functional magnetic resonance imaging data processing. Sci. Bull. 64, 953–954. doi: 10.1016/j.scib.2019.05.008
Lafer-Sousa, R., and Conway, B. R. (2013). Parallel, multi-stage processing of colors, faces and shapes in macaque inferior temporal cortex. Nat. Neurosci. 16, 1870–1878. doi: 10.1038/nn.3555
Lim, K. H., Pysklywec, A., Plante, M., and Demers, L. (2019). The effectiveness of Tai Chi for short-term cognitive function improvement in the early stages of dementia in the elderly: a systematic literature review. Clin. Interv. Aging 14, 827–839. doi: 10.2147/cia.S202055
Loprinzi, P. D. (2019). The effects of physical exercise on parahippocampal function. Physiol. Int. 106, 114–127. doi: 10.1556/2060.106.2019.10
Lovatel, G. A., Elsner, V. R., Bertoldi, K., Vanzella, C., Moyses Fdos, S., Vizuete, A., et al. (2013). Treadmill exercise induces age-related changes in aversive memory, neuroinflammatory and epigenetic processes in the rat hippocampus. Neurobiol. Learn. Mem. 101, 94–102. doi: 10.1016/j.nlm.2013.01.007
Ludbrook, J. (1998). Multiple comparison procedures updated. Clin. Exp. Pharmacol. Physiol. 25, 1032–1037. doi: 10.1111/j.1440-1681.1998.tb02179.x
Makizako, H., Shimada, H., Doi, T., Park, H., Yoshida, D., and Suzuki, T. (2013). Six-minute walking distance correlated with memory and brain volume in older adults with mild cognitive impairment: a voxel-based morphometry study. Dement Geriatr. Cogn. Dis. Extra 3, 223–232. doi: 10.1159/000354189
McCarthy, G., Puce, A., Gore, J. C., and Allison, T. (1997). Face-specific processing in the human fusiform gyrus. J. Cogn. Neurosci. 9, 605–610. doi: 10.1162/jocn.1997.9.5.605
Mortimer, J. A., Ding, D., Borenstein, A. R., DeCarli, C., Guo, Q., Wu, Y., et al. (2012). Changes in brain volume and cognition in a randomized trial of exercise and social interaction in a community-based sample of non-demented Chinese elders. J. Alzheimers Dis. 30, 757–766. doi: 10.3233/jad-2012-120079
Murphy, K., Birn, R. M., Handwerker, D. A., Jones, T. B., and Bandettini, P. A. (2009). The impact of global signal regression on resting state correlations: are anti-correlated networks introduced? Neuroimage 44, 893–905. doi: 10.1016/j.neuroimage.2008.09.036
Nasreddine, Z. S., Phillips, N. A., Bedirian, V., Charbonneau, S., Whitehead, V., Collin, I., et al. (2005). The Montreal Cognitive Assessment, MoCA: a brief screening tool for mild cognitive impairment. J. Am. Geriatr. Soc. 53, 695–699. doi: 10.1111/j.1532-5415.2005.53221.x
Nemanic, S., Alvarado, M. C., and Bachevalier, J. (2004). The hippocampal/parahippocampal regions and recognition memory: insights from visual paired comparison versus object-delayed nonmatching in monkeys. J. Neurosci. 24, 2013–2026. doi: 10.1523/jneurosci.3763-03.2004
Niemann, C., Godde, B., and Voelcker-Rehage, C. (2014). Not only cardiovascular, but also coordinative exercise increases hippocampal volume in older adults. Front. Aging Neurosci. 6:170. doi: 10.3389/fnagi.2014.00170
Nitschke, M. F., Kleinschmidt, A., Wessel, K., and Frahm, J. (1996). Somatotopic motor representation in the human anterior cerebellum. A high-resolution functional MRI study. Brain 119(Pt 3), 1023–1029. doi: 10.1093/brain/119.3.1023
Nyberg, L., Lovden, M., Riklund, K., Lindenberger, U., and Backman, L. (2012). Memory aging and brain maintenance. Trends Cogn. Sci. 16, 292–305. doi: 10.1016/j.tics.2012.04.005
Oldfield, R. (1971). The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 9, 97–113. doi: 10.1016/0028-3932(71)90067-4
Persson, J., Nyberg, L., Lind, J., Larsson, A., Nilsson, L. G., Ingvar, M., et al. (2006). Structure-function correlates of cognitive decline in aging. Cereb Cortex 16, 907–915. doi: 10.1093/cercor/bhj036
Qiu, C., and Fratiglioni, L. (2018). Aging without Dementia is Achievable: Current Evidence from Epidemiological Research. J. Alzheimers Dis. 62, 933–942. doi: 10.3233/jad-171037
Rajah, M. N., Kromas, M., Han, J. E., and Pruessner, J. C. (2010). Group differences in anterior hippocampal volume and in the retrieval of spatial and temporal context memory in healthy young versus older adults. Neuropsychologia 48, 4020–4030. doi: 10.1016/j.neuropsychologia.2010.10.010
Raslau, F. D., Mark, I. T., Klein, A. P., Ulmer, J. L., Mathews, V., and Mark, L. P. (2015). Memory part 2: the role of the medial temporal lobe. AJNR Am. J. Neuroradiol. 36, 846–849. doi: 10.3174/ajnr.A4169
Rijntjes, M., Buechel, C., Kiebel, S., and Weiller, C. (1999). Multiple somatotopic representations in the human cerebellum. Neuroreport 10, 3653–3658. doi: 10.1097/00001756-199911260-00035
Schmahmann, J. D., and Sherman, J. C. (1998). The cerebellar cognitive affective syndrome. Brain 121(Pt 4), 561–579. doi: 10.1093/brain/121.4.561
Shing, Y. L., Werkle-Bergner, M., Brehmer, Y., Muller, V., Li, S. C., and Lindenberger, U. (2010). Episodic memory across the lifespan: the contributions of associative and strategic components. Neurosci. Biobehav. Rev. 34, 1080–1091. doi: 10.1016/j.neubiorev.2009.11.002
Siddarth, P., Rahi, B., Emerson, N. D., Burggren, A. C., Miller, K. J., Bookheimer, S., et al. (2018). Physical Activity and Hippocampal Sub-Region Structure in Older Adults with Memory Complaints. J. Alzheimers Dis. 61, 1089–1096. doi: 10.3233/jad-170586
Small, S. A., Schobel, S. A., Buxton, R. B., Witter, M. P., and Barnes, C. A. (2011). A pathophysiological framework of hippocampal dysfunction in ageing and disease. Nat. Rev. Neurosci. 12, 585–601. doi: 10.1038/nrn3085
Spiridon, M., Fischl, B., and Kanwisher, N. (2006). Location and spatial profile of category-specific regions in human extrastriate cortex. Hum. Brain Mapp. 27, 77–89. doi: 10.1002/hbm.20169
Sungkarat, S., Boripuntakul, S., Kumfu, S., Lord, S. R., and Chattipakorn, N. (2018). Tai Chi Improves Cognition and Plasma BDNF in Older Adults With Mild Cognitive Impairment: A Randomized Controlled Trial. Neurorehabil. Neural. Repair 32, 142–149. doi: 10.1177/1545968317753682
Suwabe, K., Byun, K., Hyodo, K., Reagh, Z. M., Roberts, J. M., Matsushita, A., et al. (2018). Rapid stimulation of human dentate gyrus function with acute mild exercise. Proc. Natl. Acad. Sci. U S A 115, 10487–10492. doi: 10.1073/pnas.1805668115
Szabo, A. N., McAuley, E., Erickson, K. I., Voss, M., Prakash, R. S., Mailey, E. L., et al. (2011). Cardiorespiratory fitness, hippocampal volume, and frequency of forgetting in older adults. Neuropsychology 25, 545–553. doi: 10.1037/a0022733
Szanton, S. L., Walker, R. K., Roberts, L., Thorpe, R. J. Jr., Wolff, J., Agree, E., et al. (2015). Older adults’ favorite activities are resoundingly active: findings from the NHATS study. Geriatr. Nurs. 36, 131–135. doi: 10.1016/j.gerinurse.2014.12.008
Tromp, D., Dufour, A., Lithfous, S., Pebayle, T., and Despres, O. (2015). Episodic memory in normal aging and Alzheimer disease: Insights from imaging and behavioral studies. Ageing Res. Rev. 24, 232–262. doi: 10.1016/j.arr.2015.08.006
Tsai, C. L., Pan, C. Y., Chen, F. C., and Tseng, Y. T. (2017). Open- and Closed-Skill Exercise Interventions Produce Different Neurocognitive Effects on Executive Functions in the Elderly: A 6-Month Randomized, Controlled Trial. Front. Aging Neurosci. 9:294. doi: 10.3389/fnagi.2017.00294
Van Petten, C., Plante, E., Davidson, P. S., Kuo, T. Y., Bajuscak, L., and Glisky, E. L. (2004). Memory and executive function in older adults: relationships with temporal and prefrontal gray matter volumes and white matter hyperintensities. Neuropsychologia 42, 1313–1335. doi: 10.1016/j.neuropsychologia.2004.02.009
Wang, C., Schmid, C. H., Iversen, M. D., Harvey, W. F., Fielding, R. A., Driban, J. B., et al. (2016). Comparative Effectiveness of Tai Chi Versus Physical Therapy for Knee Osteoarthritis: A Randomized Trial. Ann. Intern. Med. 165, 77–86. doi: 10.7326/m15-2143
Wei, G. X., Xu, T., Fan, F. M., Dong, H. M., Jiang, L. L., Li, H. J., et al. (2013). Can Taichi reshape the brain? A brain morphometry study. PLoS One 8:e61038. doi: 10.1371/journal.pone.0061038
Xia, M., Wang, J., and He, Y. (2013). BrainNet Viewer: a network visualization tool for human brain connectomics. PLoS One 8:e68910. doi: 10.1371/journal.pone.0068910
Yaffe, K., Barnes, D., Nevitt, M., Lui, L. Y., and Covinsky, K. (2001). A prospective study of physical activity and cognitive decline in elderly women: women who walk. Arch. Intern. Med. 161, 1703–1708. doi: 10.1001/archinte.161.14.1703
Yu, Q., Herold, F., Becker, B., Klugah-Brown, B., Zhang, Y., Perrey, S., et al. (2020). Cognitive Benefits of Exercise Interventions: An fMRI Activation Likelihood Estimation Meta-Analysis. bioRxiv [Preprint].
Yue, C., Zhang, Y., Jian, M., Herold, F., Yu, Q., Mueller, P., et al. (2020a). Differential Effects of Tai Chi Chuan (Motor-Cognitive Training) and Walking on Brain Networks: A Resting-State fMRI Study in Chinese Women Aged 60. Healthcare 8:67. doi: 10.3390/healthcare8010067
Yue, C., Zou, L., Mei, J., Moore, D., Herold, F., Muller, P., et al. (2020b). Tai Chi Training Evokes Significant Changes in Brain White Matter Network in Older Women. Healthcare 8:57. doi: 10.3390/healthcare8010057
Zang, Y., Jiang, T., Lu, Y., He, Y., and Tian, L. (2004). Regional homogeneity approach to fMRI data analysis. Neuroimage 22, 394–400. doi: 10.1016/j.neuroimage.2003.12.030
Zheng, Z., Zhu, X., Yin, S., Wang, B., Niu, Y., Huang, X., et al. (2015). Combined cognitive-psychological-physical intervention induces reorganization of intrinsic functional brain architecture in older adults. Neural. Plast 2015:713104. doi: 10.1155/2015/713104
Zhou, M., Liao, H., Sreepada, L. P., Ladner, J. R., Balschi, J. A., and Lin, A. P. (2018). Tai Chi Improves Brain Metabolism and Muscle Energetics in Older Adults. J. Neuroimag. 28, 359–364. doi: 10.1111/jon.12515
Zhu, W. (2012). Sadly, the earth is still round (p < 0.05). J. Sport Health Sci. 1, 9–11. doi: 10.1016/j.jshs.2012.02.002
Keywords: Tai Chi, walking, gray matter density, regional homogeneity, delayed memory
Citation: Yue C, Yu Q, Zhang Y, Herold F, Mei J, Kong Z, Perrey S, Liu J, Müller NG, Zhang Z, Tao Y, Kramer A, Becker B and Zou L (2020) Regular Tai Chi Practice Is Associated With Improved Memory as Well as Structural and Functional Alterations of the Hippocampus in the Elderly. Front. Aging Neurosci. 12:586770. doi: 10.3389/fnagi.2020.586770
Received: 24 July 2020; Accepted: 10 September 2020;
Published: 29 October 2020.
Edited by:
Changiz Geula, Northwestern University, United StatesReviewed by:
Viviane Elsner, Centro Universitário Metodista IPA, BrazilPâmela Billig Mello-Carpes, Federal University of Pampa, Brazil
Copyright © 2020 Yue, Yu, Zhang, Herold, Mei, Kong, Perrey, Liu, Müller, Zhang, Tao, Kramer, Becker and Zou. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Liye Zou, bGl5ZXpvdTEyM0BnbWFpbC5jb20=