 Josje M. Bootsma1*
Josje M. Bootsma1* Simone R. Caljouw1
Simone R. Caljouw1 Menno P. Veldman2,3
Menno P. Veldman2,3 Natasha M. Maurits4
Natasha M. Maurits4 John C. Rothwell5
John C. Rothwell5 Tibor Hortobágyi1
Tibor Hortobágyi1- 1Department of Human Movement Sciences, University Medical Center Groningen, University of Groningen, Groningen, Netherlands
- 2Movement Control and Neuroplasticity Research Group, Department of Movement Science, KU Leuven, Leuven, Belgium
- 3Leuven Brain Institute, KU Leuven, Leuven, Belgium
- 4Department of Neurology, University Medical Center Groningen, University of Groningen, Groningen, Netherlands
- 5Sobell Department of Motor Neuroscience and Movement Disorders, University College London (UCL) Institute of Neurology, London, United Kingdom
Although a general age-related decline in neural plasticity is evident, the effects of age on neural plasticity after motor practice are inconclusive. Inconsistencies in the literature may be related to between-study differences in task difficulty. Therefore, we aimed to determine the effects of age and task difficulty on motor learning and associated brain activity. We used task-related electroencephalography (EEG) power in the alpha (8–12 Hz) and beta (13–30 Hz) frequency bands to assess neural plasticity before, immediately after, and 24-h after practice of a mirror star tracing task at one of three difficulty levels in healthy younger (19–24 yr) and older (65–86 yr) adults. Results showed an age-related deterioration in motor performance that was more pronounced with increasing task difficulty and was accompanied by a more bilateral activity pattern for older vs. younger adults. Task difficulty affected motor skill retention and neural plasticity specifically in older adults. Older adults that practiced at the low or medium, but not the high, difficulty levels were able to maintain improvements in accuracy at retention and showed modulation of alpha TR-Power after practice. Together, these data indicate that both age and task difficulty affect motor learning, as well as the associated neural plasticity.
Introduction
Unfavorable structural, functional, and biochemical changes in the nervous system affect the speed and accuracy of movements in older adults (Smith et al., 1999; Seidler et al., 2010). However, if and how old age affects the acquisition and retention of new motor skills is still under debate (Voelcker-Rehage, 2008; Coats et al., 2014). Motor practice can modify the structure and function of neural populations involved in motor skill acquisition and retention (Kolb et al., 2003; Pascual-Leone et al., 2005). Although a general age-related decline in such neural plasticity is evident (Reuter-Lorenz and Park, 2010; Pascual-Leone et al., 2011), the age-effects on neural plasticity after a period of motor practice are inconclusive (King et al., 2013; Cai et al., 2014). Understanding the effects of age on motor learning and practice-related plasticity is important to aid functional independence of older adults in increasingly aging societies and to develop novel rehabilitation practice schemes.
When executing the same motor task, older compared with younger adults show greater and more widespread brain activation (Berghuis et al., 2019; Larivière et al., 2019). Two main models have been proposed to characterize the altered age-related activation patterns: a decrease in asymmetry between the hemispheres [Hemispheric Asymmetry Reduction in Older Adults; HAROLD (Cabeza, 2002)] and a shift in activation from posterior to more anterior areas of the brain [Posterior to Anterior Shift in Aging; PASA (Davis et al., 2008)]. Both models are mostly explained as compensatory mechanisms to counteract age-dictated structural and functional changes and minimize the deterioration of motor skills (Seidler et al., 2010). The Compensation-Related Utilization of Neural Circuits Hypothesis (CRUNCH) from the cognitive literature explains age-related overactivation at low levels of cognitive demand as the recruitment of more neural resources to compensate for processing deficiencies (Reuter-Lorenz and Cappell, 2008). However, when cognitive demands increase, compensation strategies in older adults may not be sufficient to maintain young-like task performance. Specifically, as a ceiling level in terms of neural resources is reached, older adults' task performance may consequently deteriorate, causing an increased age-related decline in task performance with increasing task difficulty (Reuter-Lorenz and Cappell, 2008). Evidence showing more pronounced age-related deterioration in motor performance with increasing task difficulty (Smith et al., 1999; Bangert et al., 2010), as well as increased activation in younger adults when the difficulty of a motor task increases (Rietschel et al., 2012; Buetefisch et al., 2014), provide experimental support for the idea that the CRUNCH model is applicable in the motor domain, as well.
While the effects of task difficulty on motor performance are well-described (Fitts, 1954), the effects of task difficulty on motor skill acquisition and retention are less clear. Task difficulty can be defined as the level of challenge to execute a motor task within the current spatial and temporal constraints (Bootsma et al., 2020). When a task becomes increasingly difficult, heightened motor, and cognitive demands might act as a stimulus for motor skill learning until processing capacities are exceeded (Guadagnoli and Lee, 2004). Indeed, retention performance in younger adults was maximized after practice at the highest of four difficulty levels of a keypress sequence (Lee et al., 2016) and the second-highest of four difficulty levels of a postural control task (Akizuki and Ohashi, 2015). In contrast, there are also studies reporting no effects of task difficulty on motor learning in younger adults (Joseph et al., 2013; Ong et al., 2015) or increased learning after practice at lower compared to higher difficulty levels (Maxwell et al., 2001; Chiviacowsky and Harter, 2015). Data on the effects of task difficulty on motor learning in older adults are scarce. In one study, older adults improved more after practice on an easier compared to a more difficult version of a force tracking task (Onushko et al., 2014), which is in line with the CRUNCH model.
To the best of our knowledge, no study to date has systematically examined the effects of age and task difficulty on motor learning, even though such data could provide additional insights into practice-related neural plasticity. Oscillatory activity in the alpha and beta frequency bands has been proposed as a marker of neural plasticity (Jensen et al., 2005; Bavelier et al., 2010). Electroencephalographic (EEG) power in the alpha and beta frequencies is known to be dominant at rest and to be suppressed during movements (Pfurtscheller and Lopes da Silva, 1999). During motor task execution, task-related power is lower in older compared to younger adults, indicative of higher neural activity (Heinrichs-Graham and Wilson, 2016). If and how old age affects changes in task-related power during motor practice is, however, less well-understood. In young adults, decreased task-related power following motor practice has been interpreted as an indication of early neural plasticity (Boonstra et al., 2007; Nakano et al., 2013). Findings in older adults are inconclusive: similar (Espenhahn et al., 2019) as well as reduced (Mary et al., 2015) neural plasticity in comparison with younger adults has been reported after a single practice period. These inconsistencies in the literature may be related to differences in task difficulty between studies. However, if and how age affects the interaction between task difficulty and neural plasticity is unknown.
Therefore, the aim of the current study was to determine the effects of age and task difficulty on motor performance and task-related EEG power before, immediately after, and 24-h after acquisition of a mirror star tracing task. The overarching hypothesis was that both age and task difficulty would affect motor learning and practice-related plasticity. More specifically, we hypothesized an age-related deterioration in motor performance that would be more pronounced with increasing task difficulty. Furthermore, we expected no differences between younger and older adults after practice at a low difficulty level, but lower learning rates for older vs. younger adults after practice at higher difficulty levels. With regard to neural activity, we hypothesized that older compared to younger adults would show more bilateral activity already during motor performance at a low difficulty level and that there would be an increase in bilateral activity with increasing task difficulty in both age groups. Finally, we hypothesized smaller changes in task-related EEG power after the practice period in older compared to younger adults, indicative of an age-related deterioration in neural plasticity. Similar to the age-related deterioration in motor performance, we expected this effect to be more pronounced at higher difficulty levels.
Materials and Methods
Participants
Data from healthy, right-handed (Oldfield, 1971) younger adults (N = 36, age: 21.1 ± 1.3 yr, 16 males) were compared with previously reported data in healthy, right-handed, community-dwelling older adults (N = 36, age: 70.4 ± 4.1 yr, 20 males) (Bootsma et al., 2020). The sample size was based on an a priori power analysis performed in G*Power (version 3.1.9.2) with an alpha of 0.05 and statistical power of 0.8. We determined effect sizes based on behavioral data published previously (Bootsma et al., 2018). To ensure that the sample size in the present study would be sufficient to observe the smallest behavioral effects, we chose the lowest partial eta squared value from those analyses ( = 0.15) as input for the power analysis in the present study. Exclusion criteria were: movement restrictions in the right upper extremity, neurological conditions, and medications affecting neural functioning. In addition, older adults were screened to be cognitively [Mini-Mental State Examination score 29.2 ± 1.1 (Folstein et al., 1975)] and physically [Groningen Activity Restriction Scale score 18.4 ± 1.5 (Kempen et al., 1996)] well-functioning. Written informed consent was obtained before testing and the local ethics committee approved the study protocol, which was carried out in accordance with the Declaration of Helsinki.
Experimental Design
Figure 1 shows the study design and visuomotor task, described in detail previously (Bootsma et al., 2020). Briefly, participants visited the lab on two consecutive days. On day 1, participants practiced a mirror star tracing task at one of three difficulty levels defined by the width of the wall of the star (i.e., the pathway): Practice with Low Difficulty (P-LD, 7 mm), Practice with Medium Difficulty (P-MD, 5 mm), or Practice with High Difficulty (P-HD, 3 mm). Participants were instructed to trace the outline of the star as quickly and accurately as possible with an Apple Pencil held in their right, dominant hand, while they could only see the star and their moving hand through a mirror. Before motor practice, participants performed ten trials on each difficulty level in a block-randomized order to assess baseline performance. Practice consisted of four blocks of twenty trials separated by 5 min rest periods. Immediately after and 24 h after (day 2) motor practice, resting-state EEG, motor performance on the practiced difficulty level, and task-related EEG were assessed. Motor performance was quantified in terms of both speed (movement time) and accuracy (bandwidth error). Movement time was the average time to complete one trial of the star task. Bandwidth error was the average percentage of samples in a trial that was outside of the wall of the star. At the end of day 1, perceived mental workload was measured with the NASA-tlx questionnaire (Hart and Staveland, 1988) and participants rated their perceived fatigue on a visual analog scale ranging from 0 (not fatigued at all) to 10 (completely fatigued).
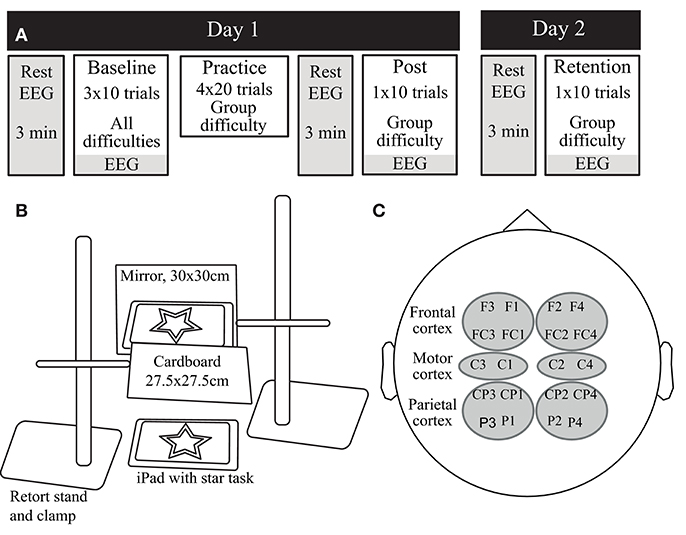
Figure 1. (A) The experimental design of the study. On day 1, participants practiced the visuomotor task at one of three difficulty levels. Before (baseline), immediately after (Post) and 24 h after (Retention) practice, motor performance, and EEG data were acquired. (B) The mirror star-tracing task. Participants were asked to trace the outline of a symmetrical five-point star as quickly and accurately as possible, while only allowed to look at the star and their moving hand through a mirror. (C) The combinations of electrodes used for the regions of interest in the EEG analysis.
EEG Data Acquisition and Preprocessing
EEG signals were continuously recorded at 250 Hz in a shielded room using a 64-channel active electrode system placed according to the international 10–10 configuration (Brain Products GmbH, Germany, Chatrian et al., 1985). The ground electrode was placed between Fp1 and Fp2, Fz was used as a recording reference and the impedance of all electrodes was kept below 10 kΩ.
We preprocessed and analyzed the EEG data using the Fieldtrip Toolbox implemented in Matlab (Oostenveld et al., 2011), as detailed previously (Bootsma et al., 2020). Briefly, resting-state and task-related data were low-pass filtered (4th order Butterworth, 70 Hz), band-stop filtered at 50 Hz, re-referenced with an average reference, segmented into non-overlapping 1-s-long epochs, upsampled to 256 Hz using piecewise cubic interpolation, and cleaned from eye-blinks and trials containing artifacts. The first and last 10% of the data of each trial were discarded before epoching to exclude errors related to the start and end of a trial. Power spectra were then calculated from the cleaned EEG data using a Fast Fourier Transform with a 10% Hanning window. Task-related power (TR-Power) spectra were averaged over the alpha (8–12 Hz) and beta (13–30 Hz) frequency bands and expressed as a percentage of power change during task execution relative to the resting-state power, according to the following equation:
Subsequently, we calculated TR-Power for regions of interest (ROIs) over the contra- and ipsilateral frontal cortex (contralateral: average of F3, FC3, F1, and FC1; ipsilateral: average of F4, FC4, F2, and FC2), motor cortex (contralateral: average of C3 and C1; ipsilateral: average of C4 and C2) and parietal cortex (contralateral: average of CP3, P3, CP1, and P1; ipsilateral: average of CP4, P4, CP2, and P2).
Statistical Analysis
All statistical analyses were conducted using R (R Core Team, 2020) in combination with the lme4 (Bates et al., 2015), lmerTest (Kuznetsova et al., 2017), and rstatix (Alboukadel, 2020) packages. Data were checked for normality with the Shapiro-Wilk test and log-transformed when the distribution was not normal. All results are reported untransformed (mean ± SD). From three younger and two older adults, EEG data at one of the three timepoints was missing for technical reasons.
Participant Characteristics
Multivariate Analysis of Variance (MANOVA) was used to check if participant characteristics and perceived fatigue after day 1 differed between age and practice groups. Since NASA-tlx ratings were not normally distributed even after log-transformation, separate Kruskal–Wallis tests were performed for younger and older adults to assess the effect of task difficulty on perceived mental workload. The effect of age on perceived mental workload was assessed by a separate Kruskal–Wallis test averaged over all difficulty levels. For all tests, a false discovery rate (FDR) correction was used to control for multiple comparisons.
Motor Performance and Neuroplasticity Assessments
Motor performance and TR-Power were assessed with multilevel linear random slope and intercept models with repeated measures (level 1) nested within subjects (level 2). First, effects of age and task difficulty on motor performance during the baseline phase were examined using a model that included the experimental variables Difficulty condition (LD, MD, HD), Practice group (P-LD, P-MD, P-HD), Age (young, old) and Block (Block 1, Block 2, Block 3) as fixed factors, as well as the Age*Block and Age*Difficulty condition interactions. Since motor performance is known to vary between individuals and especially between younger and older adults (Seidler et al., 2010), subject was entered as a random effect to allow for variation between subjects and thereby accounting for baseline differences. To assess changes in motor performance over time, multilevel linear random slope and intercept models were fitted with Age (young, old), Practice group (P-LD, P-MD, and P-HD), and Time (pre, post, and retention) as fixed factors and subject as a random effect. A full model was constructed, including the three-way interaction and all constituent terms. Because fast initial improvement took place during the baseline phase of the experiment and to fully capture the effects of task difficulty, pre-test motor performance was estimated from the first ten practice trials instead of the trials during the baseline test. Models were constructed separately for movement time and bandwidth error.
For TR-Power, separate models were constructed for the three ROIs (frontal, motor, parietal cortex) and two frequency bands (alpha, beta). TR-Power during the baseline phase was examined with a model including the fixed factors Difficulty condition (LD, MD, HD), Practice group (P-LD, P-MD, P-HD), Age (young, old), Block (Block 1, Block 2, Block 3), Hemisphere (contralateral, ipsilateral), Block*Age, Difficulty condition*Age, Hemisphere*Age, Difficulty condition*Hemisphere, and Difficulty condition*Hemisphere*Age, as well as a random effect for subject. Changes in TR-Power over time were assessed with a full model including the experimental variables Age (young, old), Practice group (P-LD, P-MD, P-HD), Time (baseline, post, retention) and Hemisphere (contralateral, ipsilateral), and all possible two-, three- and four-way interactions.
Models were fitted using restricted maximum likelihood. Normality of the distribution of the residuals was checked for all models with the Shapiro-Wilk test. In the case of a non-normal distribution, models were re-fitted with a log-transformed dependent variable. In addition, the influence of outliers on model outcomes was assessed using trimmed models excluding all data points with residuals ± 2 SDs from the mean, and robustness of effects was tested with a bootstrap validation (1,000 bootstrap samples, 95% confidence interval). Significance of fixed effect terms was assessed with Satterthwaite's method for degrees of freedom and F-statistic approximation (Luke, 2017). Post-hoc comparisons were tested using estimated marginal means from the models with a Tukey adjustment for multiple comparisons.
Relationship Between Motor Performance and EEG Data
Finally, we performed Pearson's correlation analysis to examine the relationship between motor performance and EEG data. To reduce the number of comparisons, correlation analyses followed a data-driven approach based on the results of the multilevel models. First, data from the baseline phase (i.e., all three difficulty conditions for all participants) were used to examine if there was a relationship between task difficulty and brain activity. Only EEG data for the brain cortices where a significant effect of task difficulty condition was found were correlated with behavior. In addition, if the effect of hemisphere in those cortices was not significant, TR-Power was first averaged across hemispheres. To examine age effects, correlations were calculated separately for younger and older adults. This resulted in four comparisons, which were corrected for multiple comparisons using an FDR correction. Second, the relationship between improvements in motor performance and change in TR-Power was investigated using change scores. Specifically, since the interaction effects with time, age, and practice group showed behavioral differences between the post and retention time-points (see results section), change scores for both behavioral and EEG data were calculated by subtracting post from retention values. Since TR-Power values represent the change in power during task execution relative to resting power (Gerloff et al., 1998) and lower alpha and beta power during task execution relative to a resting state is associated with higher brain activity (Pfurtscheller and Lopes da Silva, 1999), changes in TR-Power between two time-points are indicative of increased/decreased brain activity related to learning. Because we were specifically interested in the effect of task difficulty on learning, only the changes in TR-Power for the brain cortices where a significant Time*Practice group interaction was found were correlated with behavior. In addition, the change in TR-Power was averaged over both hemispheres if the Time*Hemisphere interaction was not significant. Again, an FDR correction was used to control for multiple comparisons (24 in total) and FDR-corrected p-values (pcor) are reported. The alpha level for all statistical tests was set to 0.05.
Results
Table 1 shows the participants' characteristics, which were not different between age and practice groups except that older adults were somewhat shorter and physically more active than younger adults (main effect of age, p < 0.05).
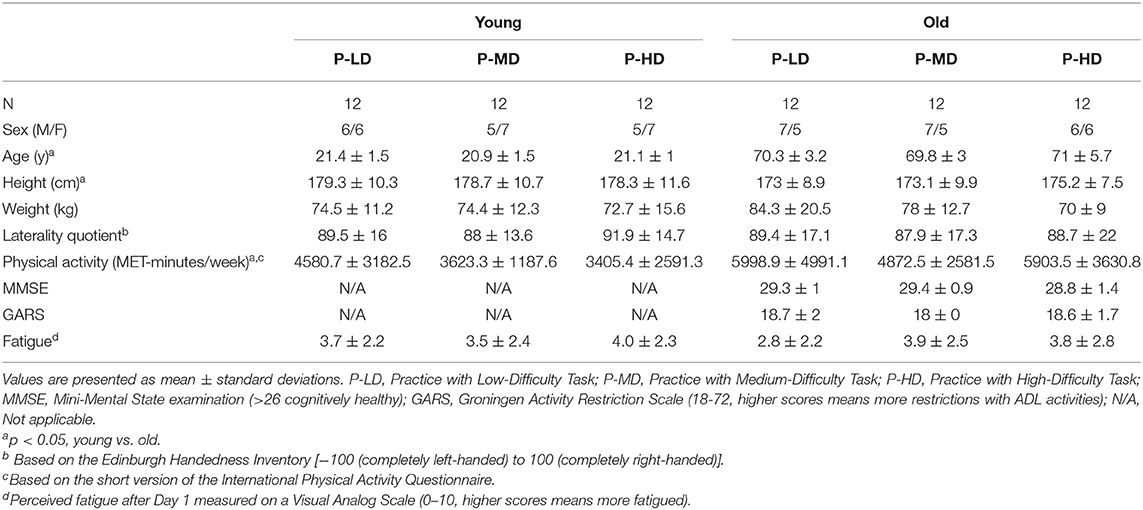
Table 1. Participant characteristics.
Perceived Fatigue and Mental Workload
A 3 (Difficulty condition) by 2 (Age) ANOVA showed that perceived fatigue at the end of day 1 did not differ between age or practice groups (Table 1). With regard to the NASA-tlx, Kruskal-Wallis tests confirmed that task difficulty affected the overall perceived mental workload only in younger adults (Supplementary Table 1 and Supplementary Figure 1) such that it was 49% higher in P-HD compared to P-LD (p = 0.006). Age did not affect the overall perceived mental workload (p = 0.26). The subscales of the NASA-tlx revealed differences between age groups (Supplementary Figure 1). Ratings from younger adults were 165% higher on the temporal demand subscale compared to older adults (p = 0.002), whereas ratings from older adults were 305% higher on the physical demand (p < 0.001) and 112% higher on the effort (p = 0.002) subscales compared to younger adults, suggesting that younger and older adults approached the motor task differently.
Behavioral Data
Table 2 and Figures 2, 3 summarize the motor performance data. The complete multilevel models are presented in Supplementary Table 2.
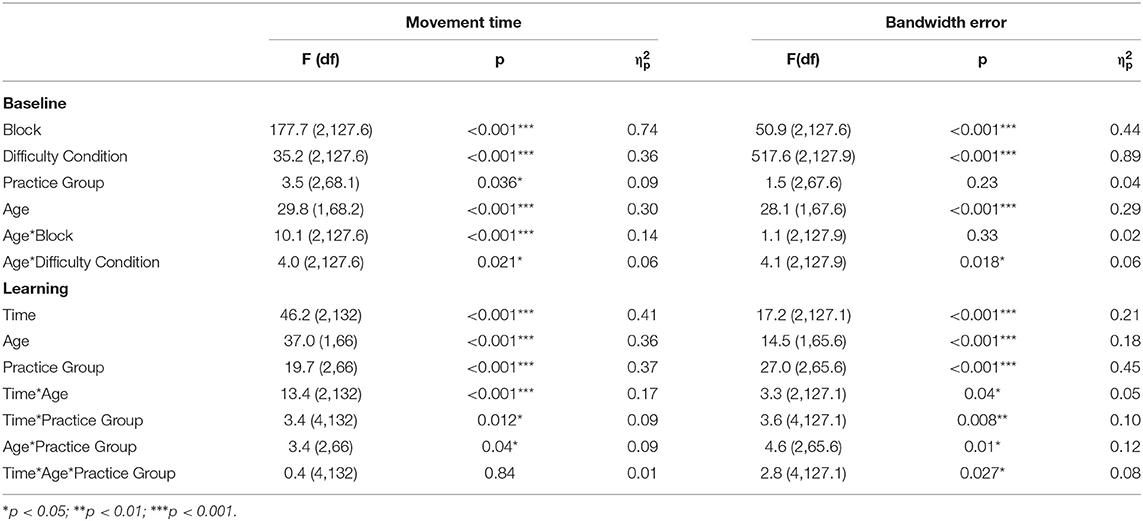
Table 2. F-tests of fixed effects for the behavioral data.
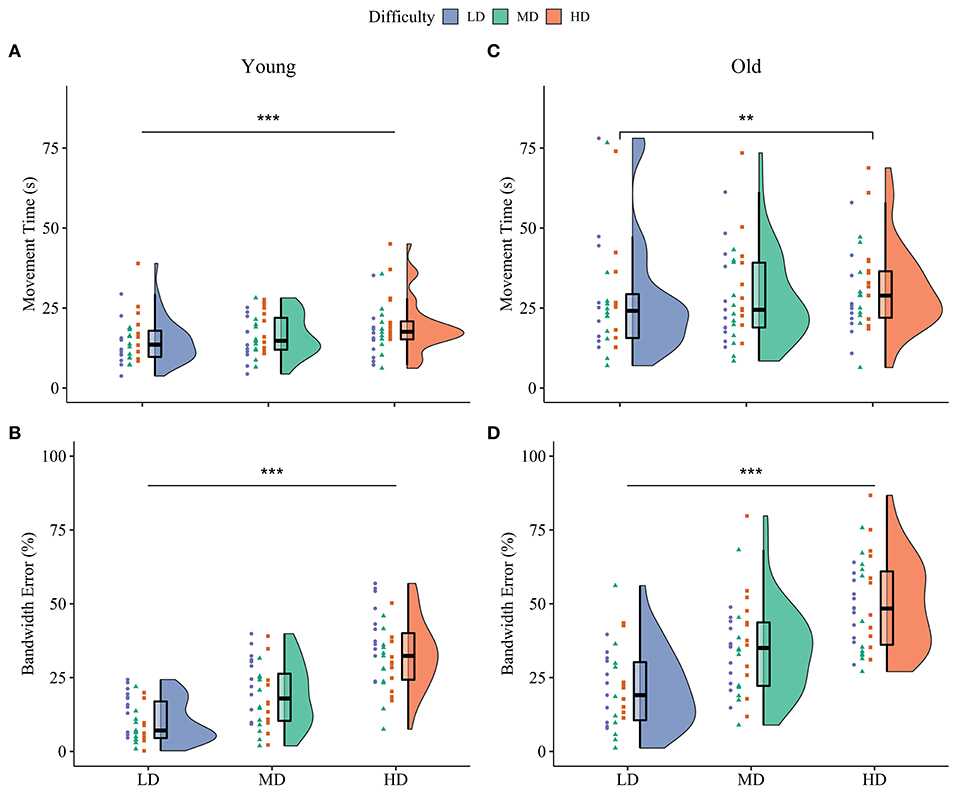
Figure 2. Baseline motor performance on the three difficulty conditions. Both movement time (A) and bandwidth error (B) differed between all three difficulty conditions in younger adults. For older adults, movement time only differed between the low and high difficulty condition (C), while bandwidth error was different for all conditions (D). Violin plots at the right side show the probability density, box-plots in the middle represents the median and interquartile range and dots on the left show individual raw data from the different practice groups. Blue: Low Difficulty (LD), green: Medium Difficulty (MD), orange: High Difficulty (HD). ***p < 0.001, **p < 0.01.
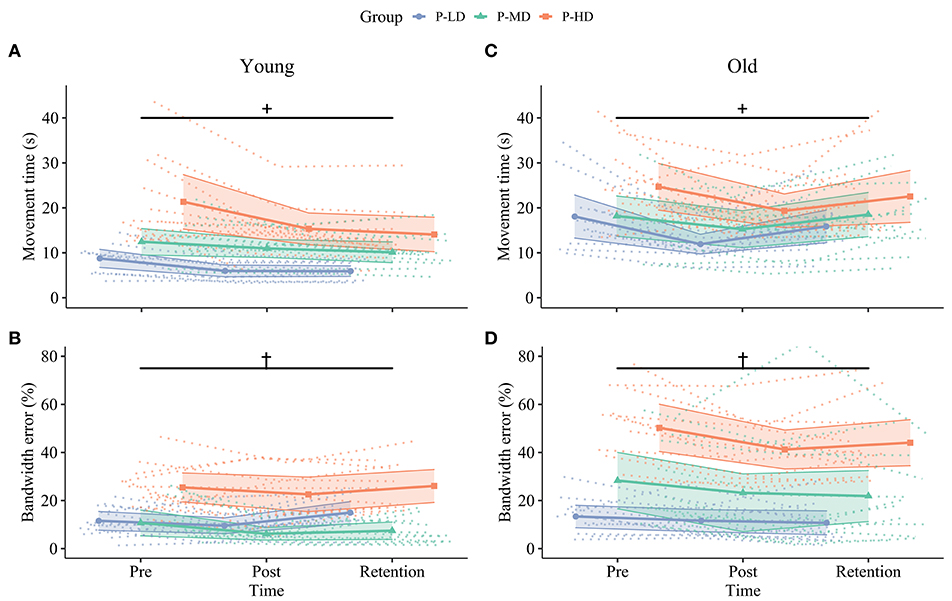
Figure 3. Change in speed (upper row, A,C) and accuracy (bottom row, B,D) over time for younger (left column, A,B) and older (right column, C,D) participants in the different practice groups. Dotted lines in the background show individual trajectories and shaded areas represent the 95% confidence intervals. Improvements in movement time from pre to post were greater in the P-LD and P-HD groups (+, significant Time*Practice group interaction, p = 0.01). Furthermore, younger adults only consolidated improvements in movement time (A), while older adults only consolidated improvements in bandwidth error (D). Practicing at a high difficulty level hindered motor skill retention of bandwidth error in older, but not younger adults (†, significant Time*Age*Practice group interaction, p = 0.03). Blue: Practice with Low Difficulty (P-LD), green: Practice with Medium Difficulty (P-MD), orange: Practice with High Difficulty (P-HD).
Baseline
Movement Time
The multilevel model of motor performance showed that older adults moved 73% slower than younger adults during the baseline phase (Age main effect, p < 0.001, = 0.30). A significant main effect of Block (p < 0.001, = 0.74) indicated fast initial improvement over the baseline trials. Post hoc comparisons of the Age*Difficulty Condition interaction showed that in younger adults, movement time increased between all three difficulty conditions, indicating that younger adults moved slower as task difficulty increased (LD/MD: 11% increase, p = 0.004; LD/HD: 30% increase, p < 0.001; MD/HD: 17% increase, p < 0.001). In older adults, movement time only increased in the high difficulty condition (LD/MD: 2% increase, p = 0.53; LD/HD: 11% increase, p < 0.001; MD/HD: 9% increase, p = 0.016; Figures 2A,C). There was a main effect of Practice group for movement time (p = 0.04, = 0.09), suggesting that participants in P-HD moved slower compared to P-LD and P-MD. However, none of the post-hoc tests reached significance (P-LD/P-HD: p = 0.07; P-MD/P-HD: p = 0.06). An Age*Block interaction for movement time revealed that the improvement from block 1 to block 2 was higher in older (35%) compared to younger (20%) adults (p < 0.001, = 0.14).
Bandwidth Error
Overall, bandwidth error was 71% higher in older than younger adults during the baseline phase (Age main effect, p < 0.001, = 0.29). Bandwidth error differed between all three difficulty conditions in both age groups, indicating the successful manipulation of task difficulty (Difficulty Condition main effect, p < 0.001, = 0.89; Figures 2B,D). Post-hoc tests showed that bandwidth error increased more between the low difficulty condition and the other two difficulty conditions in older compared to younger adults (LD/MD: p = 0.03, LD/HD: p = 0.007). Similar to movement time, a significant main effect of Block indicated fast initial improvement over the baseline trials (p < 0.001, = 0.44).
Learning
Movement Time
The multilevel model assessing learning revealed Time*Age ( = 0.17) and Time*Practice group ( = 0.09) interactions for movement time. While movement time improved in both age groups at similar rates from pre to post across difficulty levels (both 24% improvement, p < 0.001), only the younger adults maintained this improvement at 24 h retention (Time*Age interaction; 7% change post to retention, p = 0.25; Figure 3A), while the movement time from older adults reverted to baseline levels (−22% change post to retention, p < 0.001; Figure 3C). In addition, the Time*Practice group interaction ( = 0.09) showed that across age groups, learning rates from pre to post were higher in P-LD (33% improvement, p < 0.001) and P-HD (25% improvement, p < 0.001) compared to P-MD (14% improvement, p = 0.01).
Bandwidth Error
A significant Time*Age*Practice group interaction indicated that task difficulty affected improvements in bandwidth error differently in younger and older adults (p = 0.03, = 0.08). For younger adults, only the group practicing at the medium difficulty level improved bandwidth error (43% improvement pre-post, p = 0.002; Figure 3B). In contrast, younger adults in the P-LD and P-HD groups did not improve bandwidth error significantly over practice (P-LD: 18% improvement pre-post, p = 0.50; P-HD: 11% improvement pre-post, p = 0.23), and performance from P-LD at retention was even 30% worse compared to the pre-test (p = 0.047). For older adults, improvements from pre to post ranged from 13 to 33% between practice groups. Older P-LD and P-MD were able to maintain this improvement at retention (P-LD: 20% difference pre-retention, p = 0.012; P-MD: 23% difference pre-retention, p = 0.008), while performance from P-HD deteriorated with 7% from post to retention (12% difference pre-retention, p = 0.54; Figure 3D).
In summary, both age and task difficulty affected motor skill acquisition and retention. Young participants who practiced at the low and high difficulty levels improved tracing speed, but not accuracy from pre to post. In contrast, young participants who practiced at the medium difficulty level improved tracing accuracy but showed the least improvement in speed. Older participants improved both speed and accuracy across all difficulty levels. At retention, younger adults only consolidated improvements in speed, while older adults only consolidated improvements in accuracy. Motor skill retention was dependent on task difficulty only in older adults. Older adults that practiced at the low or medium, but not the high, difficulty levels were able to maintain improvements in accuracy at retention.
EEG Power Analyses
Table 3 and Figures 4, 5 summarize the TR-Power data. Supplementary Table 3 shows the complete multilevel models.
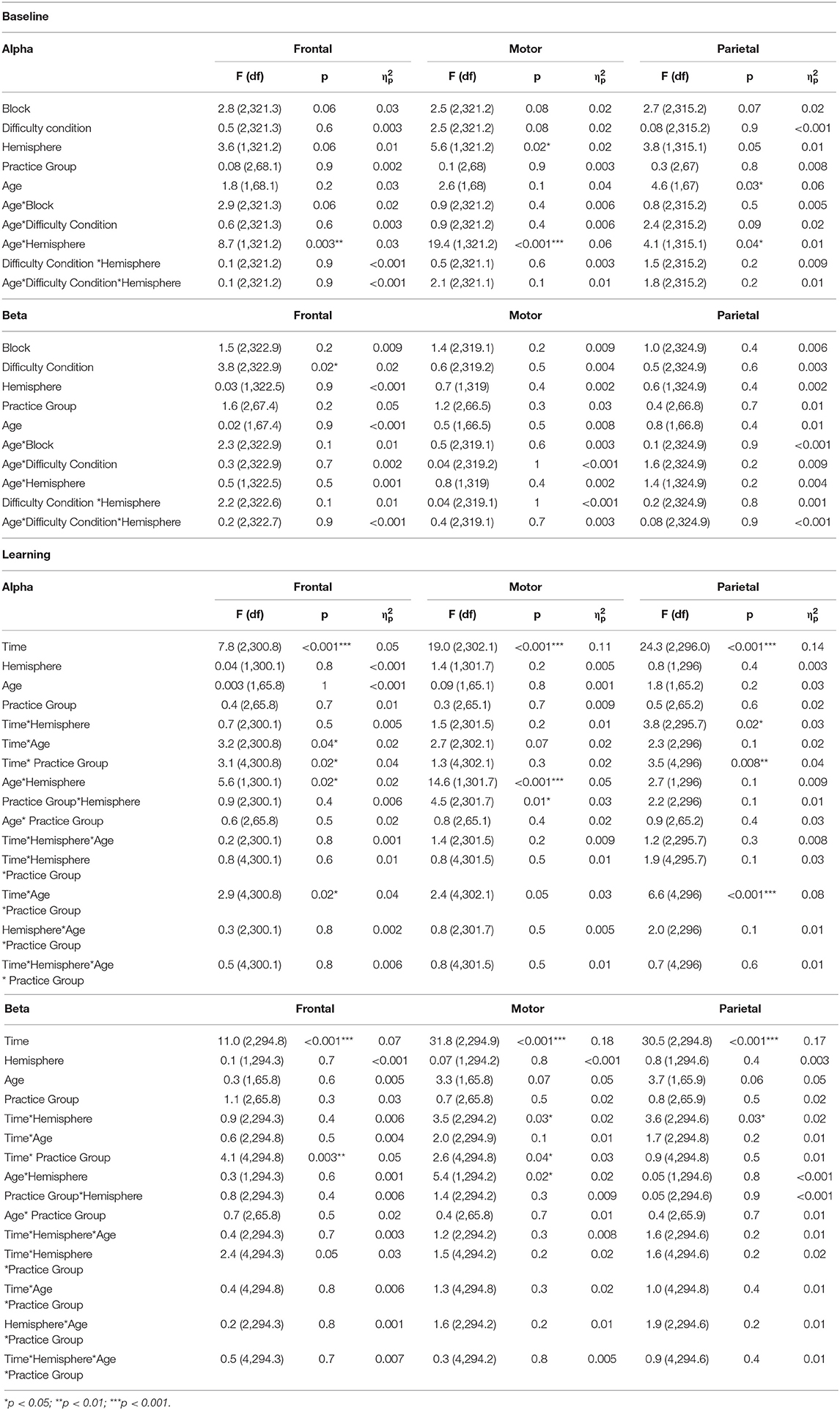
Table 3. F-tests of fixed effects for the TR-Power data.
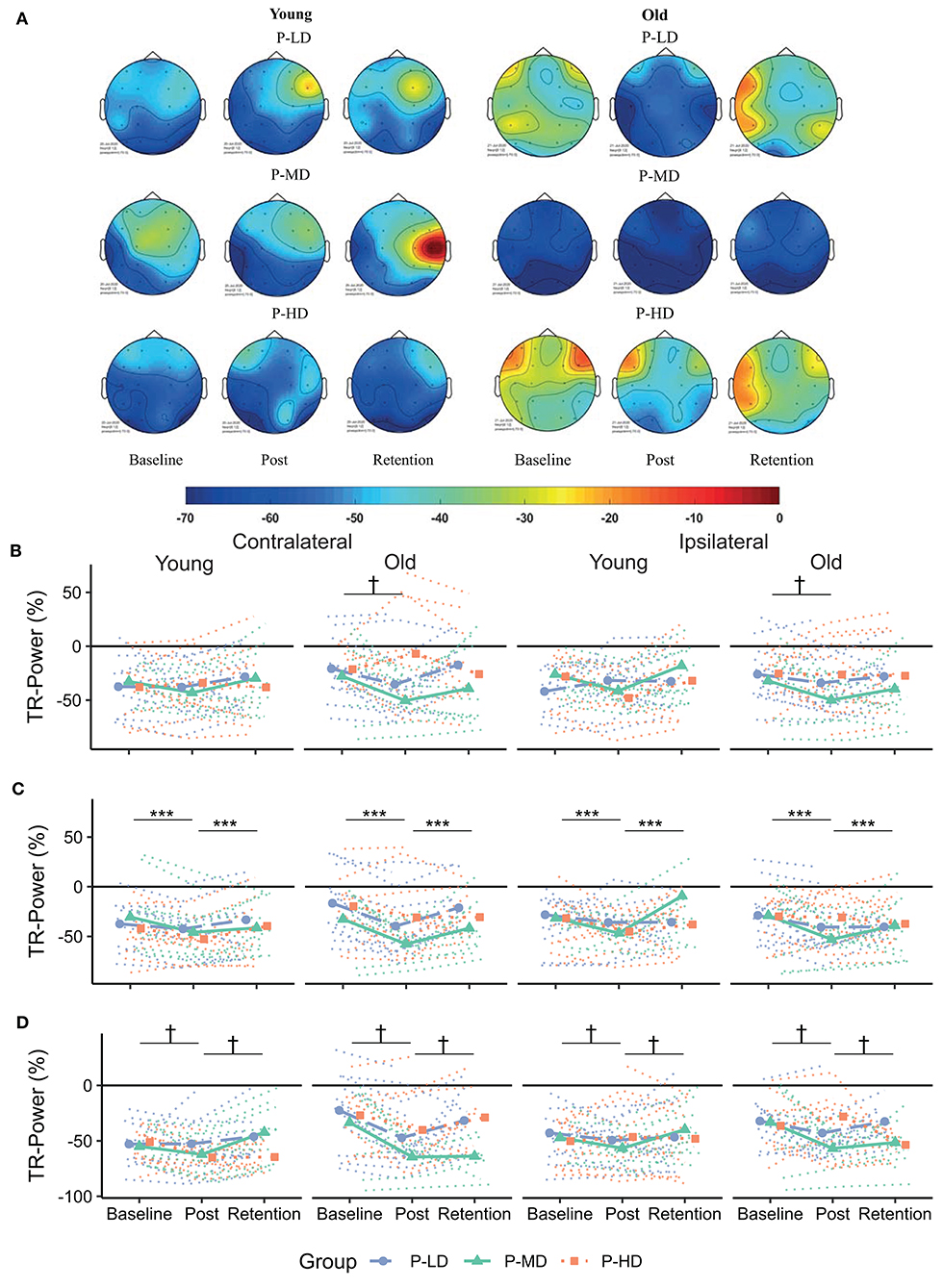
Figure 4. (A) Topographical plots showing the distribution of alpha TR-Power over the ROIs for the different age and practice groups. Cooler colors represent lower TR-Power (i.e., higher activity). Black dots represent the electrodes used to define the ROIs. (B–D) Change in TR-Power over time for the frontal (B), motor (C) and parietal (D) cortex. Dotted lines represent individual trajectories. Overall, alpha power was more lateralized (i.e., more negative) contralateral to the moving hand for younger compared to older adults. Over the frontal and parietal cortices, TR-Power decreased from baseline to the post-test. This decrease was greatest for older adults practicing at a low or medium difficulty level. P-LD, Practice with Low Difficulty (blue dots); P-MD, Practice with Medium Difficulty (green triangles); P-HD, Practice with High Difficulty (orange squares). ***Main effect of time at p < 0.001; †Time*Age*Practice group interaction at p < 0.05.
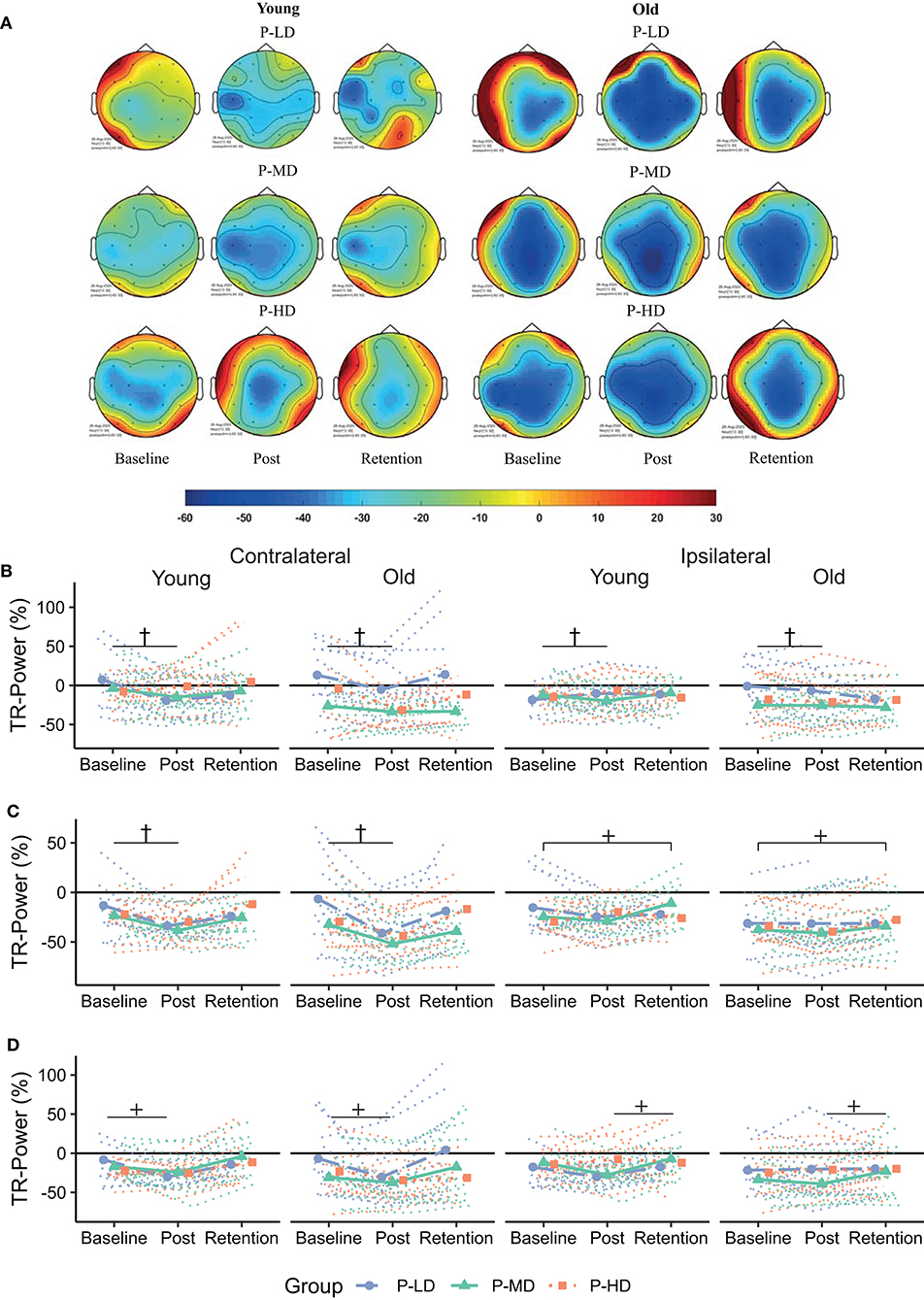
Figure 5. (A) Topographical plots showing the distribution of beta TR-Power over the ROIs for the different age and practice groups. Cooler colors represent lower TR-Power (i.e., higher activity). Black dots represent the electrodes used to define the ROIs. (B–D) Change in TR-Power over time for the frontal (B), motor (C), and parietal (D) cortex. Dotted lines represent individual trajectories. Overall, there was a decrease in beta TR-Power from baseline to the post-test. This decrease was specific for participants who practiced at the low or medium difficulty levels over the frontal and motor cortices and lateralized to the contralateral hemisphere over the motor and parietal cortices. P-LD, Practice with Low Difficulty (blue dots); P-MD, Practice with Medium Difficulty (green triangles), P-HD, Practice with High Difficulty (orange squares). †Time*Practice group interaction at p < 0.05; +Time*Hemsphere interaction at p < 0.05.
Baseline
Alpha Band
During the baseline phase, post-hoc comparisons showed that alpha TR-Power over the frontal and motor cortices was respectively, 14 and 15% higher (i.e., less activity) in the ipsi- compared to the contralateral hemisphere in younger adults (both p < 0.001), while alpha TR-Power did not differ between hemispheres in older adults (Hemisphere*Age interaction, = 0.06; Frontal: p = 0.5; Motor: p = 0.4). An opposite effect was seen over the parietal cortex, where alpha TR-Power was 11% lower in the ipsi- compared to the contralateral hemisphere in older adults (p < 0.001), but did not differ between hemispheres in younger adults (p = 0.9). No significant differences in alpha TR-Power were found between difficulty conditions, practice groups, or baseline blocks (Table 3).
Beta Band
Across age groups, practice groups, and hemispheres, beta TR-Power over the frontal cortex was lower during the high (mean ± SD: −3.25 ± 41.7%), compared to the low (1.67 ± 46.0%) or medium (3.48 ± 55.5%) difficulty conditions (Difficulty Condition main effect: p = 0.02, = 0.02). All other main and interaction terms in the model were not significant, indicating no significant differences in beta TR-Power between practice or age groups, baseline blocks, or hemispheres.
Learning
Alpha Band
Effects of age and task difficulty on changes in alpha TR-Power over the practice period differed by ROI (Figure 4), as revealed by the multilevel model for learning. Over the frontal cortex, TR-Power decreased on average 48 and 58% across hemispheres directly after practice for old P-LD and P-MD (p = 0.03 and p < 0.001, respectively), but did not change for old P-HD (p = 0.9) or any of the young practice groups (Time*Age*Practice group interaction: p = 0.02, = 0.04). At 24 h retention, frontal alpha TR-Power did not significantly differ from baseline levels for any of the groups (all p > 0.05). No effects of age or task difficulty were seen over the motor cortex, where alpha TR-Power decreased from baseline to post (p < 0.001) and increased again from day 1 to day 2 (p < 0.001) across hemispheres in all groups (Time main effect: p < 0.001, = 0.11). Over the parietal cortex, both age and task difficulty affected changes in TR-Power over time (Time*Age*Practice group interaction: p < 0.001, = 0.08). For younger adults, changes in alpha TR-Power over time were only seen in P-MD. For this group, TR-Power decreased 35% across hemispheres directly after learning (p = 0.002) and increased back to baseline levels from day 1 to day 2 (11% difference baseline-retention, p = 0.53). For older adults, TR-Power across hemispheres decreased 84.7% directly after learning for P-LD and 61% for P-MD (both p < 0.001), but did not change for P-HD (p = 0.82). At 24 h retention, TR-Power increased back to baseline levels for P-LD (-19% difference baseline-retention, p = 0.9), while it remained low for P-MD (−55% difference baseline-retention, p < 0.001).
In summary, effects of age and task difficulty on changes in alpha TR-Power over time were seen over the frontal and parietal cortices, where a decrease in TR-Power directly after learning was greatest for older adults practicing the task at a low or medium difficulty level.
Beta Band
Overall, the multilevel model for learning showed that beta TR-Power decreased directly after practice and increased back to baseline levels at 24 h retention (Figure 5). Over the frontal and motor cortices, this practice-related decrease was dependent on task difficulty, but not age. Time*Practice Group interactions in these cortices (Frontal: p = 0.003, = 0.05; Motor: p = 0.04, = 0.03) showed a decrease in TR-Power across hemispheres from baseline to post in P-LD (Frontal: 142% decrease, p < 0.001; Motor: 81% decrease, p < 0.001) and P-MD (Frontal: 195% decrease, p = 0.003; Motor: 41% decrease, p < 0.001), but not in P-HD (Frontal: 101% decrease, p = 0.89; Motor: 43% decrease, p = 0.32). Furthermore, a Time*Hemisphere interaction in the motor and parietal cortices (both p = 0.03, = 0.02) indicated that the practice-related decrease from baseline to post was higher in the contra- compared to the ipsilateral hemisphere (Motor: p = 0.01; Parietal: p = 0.01). In contrast, at day 2 beta TR-Power in the ipsilateral motor cortex was 24% higher than at baseline (p = 0.02), while beta TR-Power in the contralateral motor cortex at retention did not differ from baseline (p = 1.0).
Taken together, there was a decrease in beta TR-Power directly after practice, which was specific for P-LD and P-MD in the frontal and motor cortices and lateralized to the contralateral hemisphere in the motor and parietal cortices. At 24 h retention, TR-Power was higher compared to baseline in the ipsilateral motor cortex for both age groups.
Correlation Analyses
First, correlation analyses were performed with data from the baseline phase to examine if there was a relationship between task difficulty and brain activity. During the baseline phase, an effect of difficulty condition, but not hemisphere, was seen for the frontal beta TR-Power (see section Baseline). Therefore, we correlated average frontal beta TR-Power across hemispheres with behavior. After multiple comparisons correction, no significant correlations were found.
Second, the relationship between improvements in motor performance and change in TR-Power was investigated. Since the interaction effects with time, age, and practice group showed behavioral differences between the post and retention time-points (see section Behavioral Data, Learning), only the change scores between post and retention were used for the correlation analysis. A Time*Practice group interaction was found in four of the six ROIs (frontal alpha, parietal alpha, frontal beta, and motor beta; see section EEG Power Analyses, Learning), two of which also showed a significant Time*Hemisphere interaction (parietal alpha and motor beta). For the remaining two ROIs, data were averaged over the two hemispheres before they were correlated to behavior. None of the correlations reached significance after FDR correction.
Discussion
The present study aimed to determine the effects of age and task difficulty on motor learning and associated neural plasticity. In line with the hypothesis, both age and task difficulty affected motor skill acquisition and retention. Firstly, we found an age-related deterioration in motor performance at baseline that was more pronounced with increasing task difficulty. Secondly, task difficulty differentially affected motor skill acquisition and retention in older compared with younger adults: while motor skill acquisition was only affected by task difficulty in younger, but not older adults, task difficulty affected motor skill retention in older, but not younger adults. Thirdly, at retention, younger adults only consolidated improvements in speed, while older adults only consolidated improvements in accuracy. Fourthly, we found a more bilateral alpha activity pattern for older compared to younger adults during motor performance independent of task difficulty. Lastly, TR-Power decreased from baseline to post and reverted to baseline levels at 24-h retention. The magnitude of this decrease was dependent on age, task difficulty, and the region of interest.
Age-Related Deterioration in Motor Performance Increases With Task Difficulty
As expected, older compared with younger adults executed the star-tracing skill more slowly and less accurately. In addition, the age-related deterioration in movement accuracy was most pronounced in the high difficulty condition (Figure 2D). This is in line with previous studies showing an increased age-related decline in motor performance with increasing task difficulty (Smith et al., 1999; Bangert et al., 2010). Together, these studies provide experimental support for the suggestion that the CRUNCH model is not only applicable in the cognitive but also in the motor domain.
Age and Task Difficulty Affect Motor Skill Acquisition and Retention
Skill acquisition did not differ between younger and older adults, corroborating findings from earlier studies (Cirillo et al., 2011; Berghuis et al., 2019). Interestingly, task difficulty did only impact motor skill acquisition in younger, but not in older adults. Young participants who practiced at the low and high difficulty levels improved tracing speed, but not accuracy from pre to post. In contrast, young participants who practiced at the medium difficulty level improved tracing accuracy but showed the least improvement in speed (Figures 3A,B). Earlier research showed that individuals differ in the strategy they use to achieve the task goal in a redundant motor task, where motor performance can be improved through multiple solutions (i.e., spatial and temporal) (King et al., 2012). The current results extend this finding by suggesting that the strategy in younger adults is not only dependent on individual factors, but also on external factors such as task difficulty. However, since each difficulty group consisted of different individuals, further research with task difficulty as a within-subjects factor is needed to confirm this hypothesis. Furthermore, the observation that motor performance improved irrespective of practice group suggests that theoretical models stating that heightened motor and cognitive demands act as a stimulus for learning when task difficulty increases (Locke and Latham, 2002; Guadagnoli and Lee, 2004) are not sufficient to explain the effect of task difficulty on skill acquisition of a redundant motor task.
Not only task difficulty but also age affected the performance and learning of the mirror star tracing task. While younger adults only consolidated (i.e., stabilization of performance at retention relative to post) improvements in performance speed, consolidation in older adults was limited to performance accuracy (Figures 3A,D). In addition, older vs. younger adults had lower ratings on the temporal demand subscale of the NASA-tlx. Prioritization of accuracy over speed in older adults has been reported previously and is suggested to reflect a learned strategy to maintain precision of movements and be able to process online feedback (Lamb et al., 2016). Thus, it seems that while learning a mirror star tracing task, younger adults focus more on improving the temporal aspect of the movement, whereas older adults focus more on improving the spatial aspects.
In agreement with the hypothesis, task difficulty affected motor skill retention specifically in older adults. Older adults that practiced at the low or medium, but not the high, difficulty levels were able to maintain improvements in accuracy at retention (Figure 3D). Together with the increased age-related deterioration of motor performance at the high difficulty level, these results support predictions from the CRUNCH model as well as from the optimal challenge point framework, which state that when a task is too difficult, ceiling levels in terms of neural resources are reached and this might hamper learning (Guadagnoli and Lee, 2004; Reuter-Lorenz and Cappell, 2008). This finding also agrees with an earlier study using a force tracking task (Onushko et al., 2014) and confirms that impaired skill retention of tasks with a high level of difficulty is an age-specific phenomenon.
Since sleep is known to affect the overnight retention of motor skills (Robertson et al., 2004; Schmid et al., 2020), altered sleep architecture with increasing age possibly contributes to the impaired skill retention of the high difficulty task in older adults (Ohayon et al., 2004). There is growing consensus that sleep spindles play an important role in overnight memory consolidation (Fogel and Smith, 2011; van Schalkwijk et al., 2020) and it is known that the duration, amplitude, and density of sleep spindles is reduced with increasing age (Martin et al., 2013; Peters et al., 2014). Indeed, the relationship between sleep spindles and motor memory consolidation is different for younger and older adults (Fogel et al., 2014). Furthermore, the difficulty of a declarative memory task affected sleep spindle density in a post-training daytime nap (Schmidt et al., 2006). One might therefore speculate that the density and duration of sleep spindles are also affected by the difficulty of a motor task. Further studies are needed to examine whether motor task difficulty alters the relationship between age, sleep architecture, and motor memory consolidation.
More Bilateral Activity in Older vs. Younger Adults
Consistent with previous data, activity patterns during motor performance were different in older compared to younger adults (Berghuis et al., 2019; Larivière et al., 2019). Specifically, over the frontal and motor cortices, a more bilateral activity pattern was seen for alpha power during task execution for older compared to younger adults. Moreover, alpha activity over the parietal cortex was higher over the contra- compared to the ipsilateral hemisphere in older but not younger adults. Together, these results show a more bilateral and anterior pattern of alpha activity in older compared to younger adults, in line with the HAROLD and PASA models (Cabeza, 2002; Davis et al., 2008).
Bilateral frontal beta TR-Power was lower during the high, compared to the low or medium difficulty conditions in both age groups. Bilateral frontal recruitment has been associated with cognitive control, and an increase in bilateral frontal activity might therefore indicate increased cognitive demand associated with higher task difficulty, in accordance with earlier studies (Manganotti et al., 1998; Verstynen et al., 2005).
Practice-Related Changes in Task-Related Power Are Dependent on Age and Task Difficulty
In general, alpha and beta TR-Power decreased from baseline to post. A decrease of TR-Power after a period of motor practice has been reported earlier and is interpreted as a marker of early processes of neural plasticity (Boonstra et al., 2007; Nakano et al., 2013). Interestingly, while previous studies have reported reduced neural plasticity in older compared to younger adults (Mary et al., 2015; Rueda-Delgado et al., 2019), we found greater changes in alpha TR-Power in older compared to younger adults (Figure 4). A decrease of alpha power during execution of motor tasks relative to resting conditions (i.e., lower TR-Power) has been related to cognitive-motor processing, attention, and effort (Klimesch, 1999; van Wijk et al., 2012). As such, it is possible that the decrease in TR-Power in older adults reflects a compensatory mechanism necessary to cope with the task demand and improve motor performance. The data allude to the possibility that healthy older adults in the present study had a reserve but perhaps unused plasticity in the alpha band, which they were able to exploit, when called for, during motor practice.
In line with the view that heightened activity in older adults reflects a compensatory mechanism, the decrease in alpha TR-Power in older adults was dependent on task difficulty, so that it was only present in P-LD and P-MD, but not in P-HD. Together with the impaired retention, this finding further supports the idea that practicing a motor task at a high difficulty level is not beneficial for motor learning in older adults (Bootsma et al., 2020). Since decreased TR-Power after motor practice can be seen as a marker of early neural plasticity processes, the absence of change for the old P-HD group might indicate a failure to engage neural plasticity, leading to impaired retention of the learned skill.
An alternative explanation for the age-dependent change of alpha TR-Power might be that a decrease of alpha TR-Power is specifically related to an increase in the accuracy of motor performance. While younger adults became faster after practice, older adults became more accurate. Furthermore, young P-MD was the only young group in which parietal alpha TR-Power decreased from baseline to post, while it was also the only young group that improved accuracy over the practice period. As noted before, modulation of alpha power during task relative to resting conditions has been related to attentional processes (van Wijk et al., 2012). In a go/no go task, a greater allocation of visual-spatial attention was found in the group that prioritized accuracy over speed (Perri et al., 2014), providing a possible explanation for the potential relationship between decreased alpha TR-Power and increased accuracy. It should however be noted that we did not find significant correlations between improvements in accuracy and changes in alpha TR-Power and therefore, this hypothesis is merely speculative at this stage and warrants further investigation.
At 24-h retention, beta TR-Power over the ipsilateral motor cortex was higher compared to baseline across all age and practice groups, indicating a shift to a more lateralized activity pattern (Figure 5). A similar finding was found in a previous study using a visuomotor tracking task, where smaller pre-training beta activity over the ipsilateral cortex was related to better performance after practice (Espenhahn et al., 2019). Although the functional role of the ipsilateral cortex in motor control and learning is still under debate (Uehara and Funase, 2014; Barany et al., 2020; Cabibel et al., 2020), these results are consistent with the view that a more lateralized activity pattern is beneficial for motor performance (Takeuchi et al., 2012).
Together, the power data show reduced neural plasticity in older adults specifically after practice at the high difficulty level, while no age-related deterioration in neural plasticity was present after practice at low or medium difficulty levels. Although the changes in task-related power are in line with the changes in motor performance, we did not find any significant correlations between behavioral and neural measures. It is possible that none of the correlations survived the multiple comparison correction due to the relatively small sample size and high between-subjects variability. In line with the view that increased brain activity after motor practice is a compensatory mechanism in older adults, we did observe a trend for a correlation suggesting that older adults who showed the least increase in TR-Power also showed the least deterioration in motor performance from post to retention (data not shown). However, more research with larger samples is needed to verify possible relations between changes in TR-Power and changes in motor performance.
Clinical Applications and Future Directions
Understanding the effects of age on motor learning and practice-related plasticity can aid to develop rehabilitation practice schemes for patients recovering from movement impairments (Ward, 2017). Specifically, the observation that older adults only consolidated improvements in motor performance after practice with low or medium difficulty levels suggests that older adults in rehabilitation settings would benefit more when practicing under conditions with lower difficulty levels. Furthermore, our data imply that age and task difficulty might affect spatial and temporal aspects of a motor skill differently, emphasizing the need to tailor motor task characteristics to the desired rehabilitation outcomes. Task difficulty could potentially be added to the variables used to dose motor learning in clinical rehabilitation (Wang et al., 2020). To apply the current results to clinical settings, further studies need to assess if the improvements after practice with low or medium difficulty levels are also consolidated over longer retention periods. Promising results have been found by several studies showing at least partial retention up to 5 years after motor practice (Rodrigue et al., 2005; Schaefer and Duff, 2015). If and how this long-term retention applies to the current results, as well as how the current results apply to other motor skills, activities of daily living, and clinical populations such as stroke patients, are important targets for future research.
Limitations
Previous studies have suggested that age-related deteriorations of motor performance and practice-related plasticity follow a more gradual decline throughout the age spectrum (Pascual-Leone et al., 2011; King et al., 2018). However, the current sample size and age range was not sufficient to include age as a continuous variable in the analysis. Future studies including a larger age-range are necessary to shed more light on differences in motor performance and practice-related plasticity across the life span. Secondly, while the current sample size was sufficient to observe significant and meaningful effects in both the behavioral and EEG data, further studies with larger samples are needed to replicate and confirm the effects reported here. Furthermore, physical activity levels of the current sample of older adults were high (Table 1). It is shown previously that physical fitness facilitates motor learning (Etnier et al., 2001). Therefore, the current results might not be generalizable to the entire population. However, despite the relatively active sample of older adults, we still found age-related deficits comparable with previous studies. Thirdly, we used fixed values for alpha and beta frequency bands in our EEG analysis. However, aging has been related to a decrease in the peak frequencies in both the alpha and beta bands (Doppelmayr et al., 1998; Ishii et al., 2017). It is, therefore, possible that more subtle age-related changes have been missed, and further research with individualized frequency bands is needed to provide greater insights into the effects of age on alpha and beta task-related power. Lastly, we only measured perceived fatigue at the end of the first testing session. Since peak fatigue levels were presumably reached after the practice period and fatigue may impact motor learning (Branscheidt et al., 2019), further research assessing subjective fatigue at multiple time-points and relating these measures to the objective EEG data is needed to shed more light on the possible confounding effects of fatigue processes on motor learning.
Conclusion
In conclusion, both age and task difficulty affected motor skill acquisition and retention, as well as the associated neural plasticity. We found an age-related deterioration in motor performance which was more pronounced with increasing task difficulty and accompanied by altered patterns of brain activity. Furthermore, both age and task difficulty affected the approach to realize the task goal and improve on either the spatial or temporal dimensions of the mirror star tracing task. While we found no overall age-related deterioration in neural plasticity, task difficulty affected motor skill retention and neural plasticity specifically in older adults. Together, these results imply that the neural mechanisms of motor skill learning are dependent on both age and task difficulty.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found at: DataverseNL: https://doi.org/10.34894/MCN6XM.
Ethics Statement
The studies involving human participants were reviewed and approved by Ethische Commissie Bewegingswetenschappen, Department of Human Movement Sciences, University Medical Center Groningen, University of Groningen, Groningen, The Netherlands. The patients/participants provided their written informed consent to participate in this study.
Author Contributions
JB, SC, and TH contributed to the design of the experiment. JB collected and analyzed the data. JB, NM, and MV prepared the analysis. JB, SC, and TH wrote the first draft of the manuscript. MV, NM, and JR revised the manuscript. All authors approved the final version of the manuscript.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank P. Gupta, V. Meijers, S.E. Bouwmeester, and M. Nusselder for their assistance during data collection and E. Smid for technical support.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnagi.2021.643132/full#supplementary-material
References
Akizuki, K., and Ohashi, Y. (2015). Measurement of functional task difficulty during motor learning: what level of difficulty corresponds to the optimal challenge point? Hum. Mov. Sci. 43, 107–117. doi: 10.1016/j.humov.2015.07.007
Alboukadel, K. (2020). rstatix: Pipe-Friendly Framework for Basic Statistical Tests. Available online at: https://cran.r-project.org/package=rstatix
Bangert, A. S., Reuter-Lorenz, P. A., Walsh, C. M., Schachter, A. B., and Seidler, R. D. (2010). Bimanual coordination and aging: neurobehavioral implications. Neuropsychologia 48, 1165–1170. doi: 10.1016/j.neuropsychologia.2009.11.013
Barany, D. A., Revill, K. P., Caliban, A., Vernon, I., Shukla, A., Sathian, K., et al. (2020). Primary motor cortical activity during unimanual movements with increasing demand on precision. J. Neurophysiol. 124, 728–739. doi: 10.1152/jn.00546.2019
Bates, D., Mächler, M., Bolker, B., and Walker, S. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67:i01. doi: 10.18637/jss.v067.i01
Bavelier, D., Levi, D. M., Li, R. W., Dan, Y., and Hensch, T. K. (2010). Removing brakes on adult brain plasticity: from molecular to behavioral interventions. J. Neurosci. 30, 14964–14971. doi: 10.1523/JNEUROSCI.4812-10.2010
Berghuis, K. M. M., Fagioli, S., Maurits, N. M., Zijdewind, I., Marsman, J. B. C., Hortobágyi, T., et al. (2019). Age-related changes in brain deactivation but not in activation after motor learning. Neuroimage 186, 358–368. doi: 10.1016/j.neuroimage.2018.11.010
Boonstra, T. W., Daffertshofer, A., Breakspear, M., and Beek, P. J. (2007). Multivariate time-frequency analysis of electromagnetic brain activity during bimanual motor learning. Neuroimage 36, 370–377. doi: 10.1016/j.neuroimage.2007.03.012
Bootsma, J. M., Caljouw, S. R., Veldman, M. P., Maurits, N. M., Rothwell, J. C., and Hortobágyi, T. (2020). Failure to engage neural plasticity through practice of a high-difficulty task is accompanied by reduced motor skill retention in older adults. Neuroscience 451, 22–35. doi: 10.1016/j.neuroscience.2020.10.013
Bootsma, J. M., Hortobágyi, T., Rothwell, J. C., and Caljouw, S. R. (2018). The role of task difficulty in learning a visuomotor skill. Med. Sci. Sports Exerc. 50, 1842–1849. doi: 10.1249/MSS.0000000000001635
Branscheidt, M., Kassavetis, P., Anaya, M., Rogers, D., Huang, H. D., Lindquist, M. A., et al. (2019). Fatigue induces long-lasting detrimental changes in motor-skill learning. eLife 8:e40578. doi: 10.7554/eLife.40578.022
Buetefisch, C. M., Revill, K. P., Shuster, L., Hines, B., and Parsons, M. (2014). Motor demand-dependent activation of ipsilateral motor cortex. J. Neurophysiol. 112, 999–1009. doi: 10.1152/jn.00110.2014
Cabeza, R. (2002). Hemispheric asymmetry reduction in older adults: the HAROLD model. Psychol. Aging 17, 85–100. doi: 10.1037/0882-7974.17.1.85
Cabibel, V., Hordacre, B., and Perrey, S. (2020). Implication of the ipsilateral motor network in unilateral voluntary muscle contraction: the cross-activation phenomenon. J. Neurophysiol. 123, 2090–2098. doi: 10.1152/jn.00064.2020
Cai, L., Chan, J. S. Y., Yan, J. H., and Peng, K. (2014). Brain plasticity and motor practice in cognitive aging. Front. Aging Neurosci. 6:31. doi: 10.3389/fnagi.2014.00031
Chatrian, G. E., Lettich, E., and Nelson, P. L. (1985). Ten percent electrode system for topographic studies of spontaneous and evoked EEG activities. Am. J. EEG Technol. 25, 83–92. doi: 10.1080/00029238.1985.11080163
Chiviacowsky, S., and Harter, N. M. (2015). Perceptions of competence and motor learning: performance criterion resulting in low success experience degrades learning. Brazilian J. Mot. Behav. 9, 1–10. doi: 10.20338/bjmb.v9i1.82
Cirillo, J., Todd, G., and Semmler, J. G. (2011). Corticomotor excitability and plasticity following complex visuomotor training in young and old adults. Eur. J. Neurosci. 34, 1847–1856. doi: 10.1111/j.1460-9568.2011.07870.x
Coats, R. O., Wilson, A. D., Snapp-Childs, W., Fath, A. J., and Bingham, G. P. (2014). The 50s Cliff: perceptuo-motor learning rates across the lifespan. PLoS ONE 9:e85758. doi: 10.1371/journal.pone.0085758
Davis, S. W., Dennis, N. A., Daselaar, S. M., Fleck, M. S., and Cabeza, R. (2008). Qué PASA? The posterior-anterior shift in aging. Cereb. Cortex 18, 1201–1209. doi: 10.1093/cercor/bhm155
Doppelmayr, M., Klimesch, W., Pachinger, T., and Ripper, B. (1998). Individual differences in brain dynamics: important implications for the calculation of event-related band power. Biol. Cybern. 79, 49–57. doi: 10.1007/s004220050457
Espenhahn, S., van Wijk, B. C. M., Rossiter, H. E., de Berker, A. O., Redman, N. D., Rondina, J., et al. (2019). Cortical beta oscillations are associated with motor performance following visuomotor learning. Neuroimage 195, 340–353. doi: 10.1016/j.neuroimage.2019.03.079
Etnier, J. L., Romero, D. H., and Traustadóttir, T. (2001). Acquisition and retention of motor skills as a function of age and aerobic fitness. J. Aging Phys. Act. 9, 425–437. doi: 10.1123/japa.9.4.425
Fitts, P. M. (1954). The information capacity of the human motor system in controlling the amplitude of movement. J. Exp. Psychol. 47, 381–391. doi: 10.1037/h0055392
Fogel, S. M., Albouy, G., Vien, C., Popovicci, R., King, B. R., Hoge, R., et al. (2014). fMRI and sleep correlates of the age-related impairment in motor memory consolidation. Hum. Brain Mapp. 35, 3625–3645. doi: 10.1002/hbm.22426
Fogel, S. M., and Smith, C. T. (2011). The function of the sleep spindle: a physiological index of intelligence and a mechanism for sleep-dependent memory consolidation. Neurosci. Biobehav. Rev. 35, 1154–1165. doi: 10.1016/j.neubiorev.2010.12.003
Folstein, M. F., Folstein, S. E., and McHugh, P. R. (1975). “Mini-mental state”. A practical method for grading the cognitive state of patients for the clinician. J. Psychiatr. Res. 12, 189–198. doi: 10.1016/0022-3956(75)90026-6
Gerloff, C., Corwell, B., Hallett, M., and Cohen, L. G. (1998). The role of the human motor cortex in the control of complex and simple finger movement sequences. Brain 121, 1695–1709. doi: 10.1093/brain/121.9.1695
Guadagnoli, M. A., and Lee, T. D. (2004). Challenge point: a framework for conceptualizing the effects of various practice conditions in motor learning. J. Mot. Behav. 36, 212–224. doi: 10.3200/JMBR.36.2.212-224
Hart, S. G., and Staveland, L. E. (1988). Development of NASA-TLX (Task Load Index): results of empirical and theoretical research. Adv. Psychol. 52, 139–183. doi: 10.1016/S0166-4115(08)62386-9
Heinrichs-Graham, E., and Wilson, T. W. (2016). Is an absolute level of cortical beta suppression required for proper movement? Magnetoencephalographic evidence from healthy aging. Neuroimage 134, 514–521. doi: 10.1016/j.neuroimage.2016.04.032
Ishii, R., Canuet, L., Aoki, Y., Hata, M., Iwase, M., Ikeda, S., et al. (2017). Healthy and pathological brain aging: from the perspective of oscillations, functional connectivity, and signal complexity. Neuropsychobiology 75, 151–161. doi: 10.1159/000486870
Jensen, O., Goel, P., Kopell, N., Pohja, M., Hari, R., and Ermentrout, B. (2005). On the human sensorimotor-cortex beta rhythm: sources and modeling. Neuroimage 26, 347–355. doi: 10.1016/j.neuroimage.2005.02.008
Joseph, M. E., King, A. C., and Newell, K. M. (2013). Task difficulty and the time scales of warm-up and motor learning. J. Mot. Behav. 45, 231–238. doi: 10.1080/00222895.2013.784240
Kempen, G. I. J. M., Miedema, I., Ormel, J., and Molenaar, W. (1996). The assessment of disability with the Groningen Activity Restriction Scale. Conceptual framework and psychometric properties. Soc. Sci. Med. 43, 1601–1610. doi: 10.1016/S0277-9536(96)00057-3
King, A. C., Ranganathan, R., and Newell, K. M. (2012). Individual differences in the exploration of a redundant space-time motor task. Neurosci. Lett. 529, 144–149. doi: 10.1016/j.neulet.2012.08.014
King, B. R., Fogel, S. M., Albouy, G., and Doyon, J. (2013). Neural correlates of the age-related changes in motor sequence learning and motor adaptation in older adults. Front. Hum. Neurosci. 7:142. doi: 10.3389/fnhum.2013.00142
King, B. R., Van Ruitenbeek, P., Leunissen, I., Cuypers, K., Heise, K. F., Santos Monteiro, T., et al. (2018). Age-related declines in motor performance are associated with decreased segregation of large-scale resting state brain networks. Cereb. Cortex 28, 4390–4402. doi: 10.1093/cercor/bhx297
Klimesch, W. (1999). EEG alpha and theta oscillations reflect cognitive and memory performance: a review and analysis. Brain Res. Rev. 29, 169–195. doi: 10.1016/S0165-0173(98)00056-3
Kolb, B., Gibb, R., and Robinson, T. E. (2003). Brain plasticity and behavior. Curr. Dir. Psychol. Sci. 12, 1–5. doi: 10.1111/1467-8721.01210
Kuznetsova, A., Brockhoff, P. B., and Christensen, R. H. B. (2017). lmerTest package: tests in linear mixed effects models. J. Stat. Softw. 82:13. doi: 10.18637/jss.v082.i13
Lamb, D. G., Correa, L. N., Seider, T. R., Mosquera, D. M., Rodriguez, J. A., Salazar, L., et al. (2016). The aging brain: movement speed and spatial control. Brain Cogn. 109, 105–111. doi: 10.1016/j.bandc.2016.07.009
Larivière, S., Xifra-Porxas, A., Kassinopoulos, M., Niso, G., Baillet, S., Mitsis, G. D., et al. (2019). Functional and effective reorganization of the aging brain during unimanual and bimanual hand movements. Hum. Brain Mapp. 40, 3027–3040. doi: 10.1002/hbm.24578
Lee, T. D., Eliasz, K. L., Gonzalez, D., Alguire, K., Ding, K., and Dhaliwal, C. (2016). On the role of error in motor learning. J. Mot. Behav. 48, 99–115. doi: 10.1080/00222895.2015.1046545
Locke, E. A., and Latham, G. P. (2002). Building a practically useful theory of goal setting and task motivation: a 35-year odyssey. Am. Psychol. 57, 705–717. doi: 10.1037/0003-066X.57.9.705
Luke, S. G. (2017). Evaluating significance in linear mixed-effects models in R. Behav. Res. Methods 49, 1494–1502. doi: 10.3758/s13428-016-0809-y
Manganotti, P., Gerloff, C., Toro, C., Katsuta, H., Sadato, N., Zhuang, P., et al. (1998). Task-related coherence and task-related spectral power changes during sequential finger movements. Electroencephalogr. Clin. Neurophysiol. 109, 50–62. doi: 10.1016/S0924-980X(97)00074-X
Martin, N., Lafortune, M., Godbout, J., Barakat, M., Robillard, R., Poirier, G., et al. (2013). Topography of age-related changes in sleep spindles. Neurobiol. Aging. 34, 468–476. doi: 10.1016/j.neurobiolaging.2012.05.020
Mary, A., Bourguignon, M., Wens, V., Op de Beeck, M., Leproult, R., De Tiège, X., et al. (2015). Aging reduces experience-induced sensorimotor plasticity. A magnetoencephalographic study. Neuroimage 104, 59–68. doi: 10.1016/j.neuroimage.2014.10.010
Maxwell, J. P., Masters, R. S., Kerr, E., and Weedon, E. (2001). The implicit benefit of learning without errors. Q. J. Exp. Psychol. Sect. A Hum. Exp. Psychol. 54, 1049–1068. doi: 10.1080/713756014
Nakano, H., Osumi, M., Ueta, K., Kodama, T., and Morioka, S. (2013). Changes in electroencephalographic activity during observation, preparation, and execution of a motor learning task. Int. J. Neurosci. 123, 866–875. doi: 10.3109/00207454.2013.813509
Ohayon, M. M., Carskadon, M. A., Guilleminault, C., and Vitiello, M. V. (2004). Meta-analysis of quantitative sleep parameters from childhood to old age in healthy individuals: developing normative sleep values across the human lifespan. Sleep 27, 1255–1273. doi: 10.1093/sleep/27.7.1255
Oldfield, R. C. (1971). The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia 9, 97–113. doi: 10.1016/0028-3932(71)90067-4
Ong, N. T., Lohse, K. R., and Hodges, N. J. (2015). Manipulating target size influences perceptions of success when learning a dart-throwing skill but does not impact retention. Front. Psychol. 6:1378. doi: 10.3389/fpsyg.2015.01378
Onushko, T., Kim, C., and Christou, E. A. (2014). Reducing task difficulty during practice improves motor learning in older adults. Exp. Gerontol. 57, 168–174. doi: 10.1016/j.exger.2014.06.006
Oostenveld, R., Fries, P., Maris, E., and Schoffelen, J. M. (2011). FieldTrip: open source software for advanced analysis of MEG, EEG, and invasive electrophysiological data. Comput. Intell. Neurosci. 2011:156869. doi: 10.1155/2011/156869
Pascual-Leone, A., Amedi, A., Fregni, F., and Merabet, L. B. (2005). The plastic human brain cortex. Annu. Rev. Neurosci. 28, 377–401. doi: 10.1146/annurev.neuro.27.070203.144216
Pascual-Leone, A., Freitas, C., Oberman, L., Horvath, J. C., Halko, M., Eldaief, M., et al. (2011). Characterizing brain cortical plasticity and network dynamics across the age-span in health and disease with TMS-EEG and TMS-fMRI. Brain Topogr. 24, 302–315. doi: 10.1007/s10548-011-0196-8
Perri, R. L., Berchicci, M., Spinelli, D., and Di Russo, F. (2014). Individual differences in response speed and accuracy are associated to specific brain activities of two interacting systems. Front. Behav. Neurosci. 8:251. doi: 10.3389/fnbeh.2014.00251
Peters, K. R., Ray, L. B., Fogel, S., Smith, V., and Smith, C. T. (2014). Age differences in the variability and distribution of sleep spindle and rapid eye movement densities. PLoS ONE 9:91047. doi: 10.1371/journal.pone.0091047
Pfurtscheller, G., and Lopes da Silva, F. H. (1999). Event-related EEG/MEG synchronization and desynchronization: basic principles. Clin. Neurophysiol. 110, 1842–1857. doi: 10.1016/S.1388-2457(99)00141-8
R Core Team (2020). R: A Language and Environment for Statistical Computing. Available online at: https://www.r-project.org/ (accessed March 16, 2021).
Reuter-Lorenz, P. A., and Cappell, K. A. (2008). Neurocognitive aging and the compensation hypothesis. Curr. Dir. Psychol. Sci. 17, 177–182. doi: 10.1111/j.1467-8721.2008.00570.x
Reuter-Lorenz, P. A., and Park, D. C. (2010). Human neuroscience and the aging mind: a new look at old problems. J. Gerontol. Ser. B Psychol. Sci. Soc. Sci. 65B, 405–415. doi: 10.1093/geronb/gbq035
Rietschel, J. C., Miller, M. W., Gentili, R. J., Goodman, R. N., McDonald, C. G., and Hatfield, B. D. (2012). Cerebral-cortical networking and activation increase as a function of cognitive-motor task difficulty. Biol. Psychol. 90, 127–133. doi: 10.1016/j.biopsycho.2012.02.022
Robertson, E. M., Pascual-Leone, A., and Miall, R. C. (2004). Current concepts in procedural consolidation. Nat. Rev. Neurosci. 5, 576–582. doi: 10.1038/nrn1426
Rodrigue, K. M., Kennedy, K. M., and Raz, N. (2005). Aging and longitudinal change in perceptual-motor skill acquisition in healthy adults. J. Gerontol. B. Psychol. Sci. Soc. Sci. 60, 174–181. doi: 10.1093/geronb/60.4.P174
Rueda-Delgado, L. M., Heise, K. F., Daffertshofer, A., Mantini, D., and Swinnen, S. P. (2019). Age-related differences in neural spectral power during motor learning. Neurobiol. Aging 77, 44–57. doi: 10.1016/j.neurobiolaging.2018.12.013
Schaefer, S. Y., and Duff, K. (2015). Rapid responsiveness to practice predicts longer-term retention of upper extremity motor skill in non-demented older adults. Front. Aging Neurosci. 7:214. doi: 10.3389/fnagi.2015.00214
Schmid, D., Erlacher, D., Klostermann, A., Kredel, R., and Hossner, E. J. (2020). Sleep-dependent motor memory consolidation in healthy adults: a meta-analysis. Neurosci. Biobehav. Rev. 118, 270–281. doi: 10.1016/j.neubiorev.2020.07.028
Schmidt, C., Peigneux, P., Muto, V., Schenkel, M., Knoblauch, V., Münch, M., et al. (2006). Encoding difficulty promotes postlearning changes in sleep spindle activity during napping. J. Neurosci. 26, 8976–8982. doi: 10.1523/JNEUROSCI.2464-06.2006
Seidler, R. D., Bernard, J. A., Burutolu, T. B., Fling, B. W., Gordon, M. T., Gwin, J. T., et al. (2010). motor control and aging: links to age-related brain structural, functional and biomechanical effects. Neurosci. Biobehav. Rev. 34, 721–733. doi: 10.1016/j.neubiorev.2009.10.005
Smith, C. D., Umberger, G. H., Manning, E. L., Slevin, J. T., Wekstein, D. R., Schmitt, F. A., et al. (1999). Critical decline in fine motor hand movements in human aging. Neurology 53, 1458–1458. doi: 10.1212/WNL.53.7.1458
Takeuchi, N., Oouchida, Y., and Izumi, S.-I. (2012). Motor control and neural plasticity through interhemispheric interactions. Neural Plast. 2012, 1–13. doi: 10.1155/2012/823285
Uehara, K., and Funase, K. (2014). Contribution of ipsilateral primary motor cortex activity to the execution of voluntary movements in humans: a review of recent studies. J. Phys. Fit. Sport. Med. 3, 297–306. doi: 10.7600/jpfsm.3.297
van Schalkwijk, F. J., Hauser, T., Hoedlmoser, K., Ameen, M. S., Wilhelm, F. H., Sauter, C., et al. (2020). Procedural memory consolidation is associated with heart rate variability and sleep spindles. J. Sleep Res. 29, 1–8. doi: 10.1111/jsr.12910
van Wijk, B. C. M., Beek, P. J., and Daffertshofer, A. (2012). Neural synchrony within the motor system: what have we learned so far? Front. Hum. Neurosci. 6:252. doi: 10.3389/fnhum.2012.00252
Verstynen, T., Diedrichsen, J., Albert, N., Aparicio, P., and Ivry, R. B. (2005). Ipsilateral motor cortex activity during unimanual hand movements relates to task complexity. J. Neurophysiol. 93, 1209–1222. doi: 10.1152/jn.00720.2004
Voelcker-Rehage, C. (2008). Motor-skill learning in older adults-a review of studies on age-related differences. Eur. Rev. Aging Phys. Act. 5, 5–16. doi: 10.1007/s11556-008-0030-9
Wang, C., Winstein, C., D'Argenio, D. Z., and Schweighofer, N. (2020). The efficiency, efficacy, and retention of task practice in chronic stroke. Neurorehabil. Neural Repair 34, 881–890. doi: 10.1177/1545968320948609
Keywords: aging, electroencephalography, motor learning, plasticity, spectral analysis, task difficulty
Citation: Bootsma JM, Caljouw SR, Veldman MP, Maurits NM, Rothwell JC and Hortobágyi T (2021) Neural Correlates of Motor Skill Learning Are Dependent on Both Age and Task Difficulty. Front. Aging Neurosci. 13:643132. doi: 10.3389/fnagi.2021.643132
Received: 17 December 2020; Accepted: 23 February 2021;
Published: 22 March 2021.
Edited by:
Brian C. Clark, Ohio University, United StatesReviewed by:
Guang H. Yue, Kessler Foundation, United StatesJessica Bernard, Texas A&M University, United States
Copyright © 2021 Bootsma, Caljouw, Veldman, Maurits, Rothwell and Hortobágyi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Josje M. Bootsma, ai5tLmJvb3RzbWFAdW1jZy5ubA==