 Ahmad I. M. Athamneh
Ahmad I. M. Athamneh Daniel M. Suter
Daniel M. Suter- Bindley Bioscience Center, Birck Nanotechnology Center, Department of Biological Sciences, Purdue University, West Lafayette, IN, USA
Mechanical force plays a fundamental role in neuronal development, physiology, and regeneration. In particular, research has shown that force is involved in growth cone-mediated axonal growth and guidance as well as stretch-induced elongation when an organism increases in size after forming initial synaptic connections. However, much of the details about the exact role of force in these fundamental processes remain unknown. In this review, we highlight: (1) standing questions concerning the role of mechanical force in axonal growth and guidance; and (2) different experimental techniques used to quantify forces in axons and growth cones. We believe that satisfying answers to these questions will require quantitative information about the relationship between elongation, forces, cytoskeletal dynamics, axonal transport, signaling, substrate adhesion, and stiffness contributing to directional growth advance. Furthermore, we address why a wide range of force values have been reported in the literature, and what these values mean in the context of neuronal mechanics. We hope that this review will provide a guide for those interested in studying the role of force in development and regeneration of neuronal networks.
Introduction
The role mechanical forces play in the development and maintenance of neuronal networks has been increasingly recognized and addressed (Bray, 1979; Lamoureux et al., 1989; Suter and Miller, 2011; Franze et al., 2013). Many aspects of axonal growth and development have been examined in the context of mechanical force including cytoskeletal dynamics (Lee and Suter, 2008; Schaefer et al., 2008), axonal transport (Loverde et al., 2011; O’Toole and Miller, 2011; Ahmed and Saif, 2014), growth cone guidance (Suter et al., 1998; Suter and Forscher, 2001; Moore et al., 2009), and molecular motor activity (Bridgman et al., 2001). It has been shown that growth cones generate traction force (Lamoureux et al., 1989; Heidemann et al., 1990) and respond to mechanical stress (Franze et al., 2009) or change in substrate rigidity (Chan and Odde, 2008; Koch et al., 2012). Furthermore, it has been found that mechanical tension induces axonal growth (Bray, 1984; Zheng et al., 1991; Pfister et al., 2004) and that axonal tension is tightly regulated (Lamoureux et al., 1989; Rajagopalan et al., 2010; Hyland et al., 2014). Despite significant advances, many aspects of the mechanical control of axonal growth and guidance as well as maintenance of axons after synapse formation remain unclear.
Standing Questions Pertaining to the Role of Force in Neuronal Processes
There is ample evidence that axonal elongation is influenced by both biochemical and biomechanical factors. The neuronal growth cone controls the direction and rate of axonal growth by navigating the surrounding environment searching for molecular, mechanical, and topographical cues. The machinery responsible for sensing stiffness of the extracellular matrix (ECM) as well as of neighboring cellular surfaces is primarily powered by the actin cytoskeleton, which is highly dynamic and is constantly turning over in the peripheral (P) domain and transition (T) zone (Figure 1). Actin polymerizes at the leading edge and is pulled backward by myosin motors, resulting in retrograde F-actin flow (Lin et al., 1996; Medeiros et al., 2006). Traction force is generated as a result of coupling of the F-actin flow to cellular and ECM substrates through adhesion receptors, such as immunoglobulin superfamily cell adhesion molecules, N-cadherin, and integrins (Suter et al., 1998; Bard et al., 2008; Shimada et al., 2008). Several reviews have discussed this substrate-cytoskeletal coupling model including what is known about the role of the cytoskeleton, molecular motors as well as signaling pathways involved (Suter and Forscher, 2000; Suter and Miller, 2011; Gomez and Letourneau, 2014).
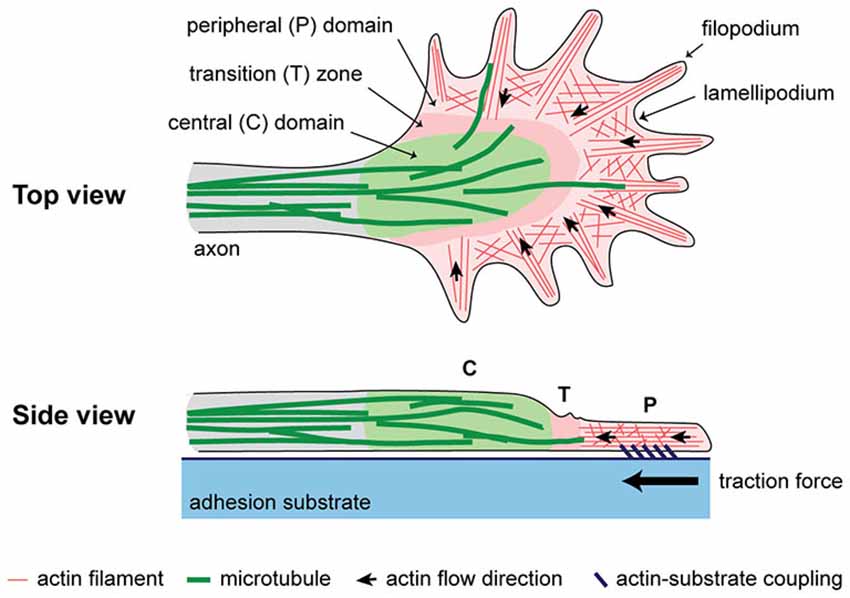
Figure 1. A simplified schematic of the cytoplasmic domains and cytoskeletal structures in the growth cone. Traction force is generated as a result of coupling of F-actin flow in the P domain to an extracellular adhesion substrate.
How Does Substrate Stiffness Affect Axonal Growth?
While it has been shown that the neuronal growth cone controls the advance of axons in part by sensing and responding to the substrate stiffness, little is known about the physical parameters governing these mechanisms. According to the prevailing model, the level of traction force is determined by the abundance and strength of coupling between F-actin flow and cellular surfaces or ECM. Coupling is influenced by the presence of adhesion substrates as well as the stiffness of the environment. The substrate must be compliant enough to allow adhesion to mature and form strong coupling capable of bearing increasing tension. If the substrate is too stiff, newly formed adhesions abruptly break as lack of compliance in the substrate results in rapid building of tension. This “motor-clutch” model provides a mechanism that inherently senses and responds to substrate stiffness (Chan and Odde, 2008). Consistent with this model, experiments have demonstrated the effect of substrate stiffness on coupling and traction force development in growth cones, but the consequences on neurite outgrowth are not fully clear yet. Furthermore, different types of neuron responded differently to substrate stiffness. For example, softer substrates were shown to promote better neurite growth and branching in spinal cord (Flanagan et al., 2002; Jiang et al., 2008) and hippocampal neurons (Kostic et al., 2007), but not cortical neurons (Norman and Aranda-Espinoza, 2010). Another study found that stiffer substrates significantly increased neurite outgrowth in cortical neurons (Stabenfeldt and LaPlaca, 2011). In a later study, neurite outgrowth of dorsal root ganglion neurons from the peripheral nervous system (PNS) were sensitive to changes in substrate stiffness unlike hippocampal neurons from the central nervous system (CNS; Koch et al., 2012), which seems to contradict earlier observations (Franze et al., 2013). It is important to note that the range of substrate stiffness employed in these studies are not all the same, which makes a direct comparison of the results more challenging, even for the same type of neuron. The apparent discrepancies in the literature, which indicate that different neurons may exhibit different mechanosensitivity, have if anything kept alive the debate about the effect of substrate stiffness on axonal growth, and certainly challenged our understanding of the mechanisms underlying mechanotransduction in neuronal growth cones.
Recently, it has been shown that compliance, necessary to allow time for coupling to mature, is provided by elastic micron-scale deformations in the actomyosin network and the nascent adhesions themselves (Mejean et al., 2013). Adding to the complexity is the observed biphasic dependence of traction force on the substrate stiffness; i.e., traction force generated by nonneuronal and neuronal cell has been shown to increase linearly with the substrate stiffness but then plateau at sufficiently higher substrate stiffness (Saez et al., 2005; Ghibaudo et al., 2008; Koch et al., 2012; Trichet et al., 2012; Yip et al., 2013). This biphasic behavior complicates comparative analysis amongst studies using different experimental techniques or substrates of different stiffness.
Do Growth Cones Respond to Mechanical Force or Deformation?
The lack of understanding of the mechanisms underlying mechanotransduction is manifested in the ongoing debate whether cell respond to force (stress) or deformation (strain) (Saez et al., 2005; Yip et al., 2013). In our lab, we have shown that the absolute level of substrate deformation appears to be a better predictor of adhesion-mediated growth cone advance when compared to the level of traction force (Athamneh et al., in press). We probed Aplysia growth cones with microneedles (MNs) having stiffness ranging from 0.003 to 0.1 N/m, and found that the threshold deformation for a positive growth cone response was about 1 μm of MN deflection regardless of how much force was exerted by the growth cone (Athamneh et al., in press). Therefore, our data highlight the fundamental role of strain vs. stress in controlling the mechanical response during cell migration. Other studies on mechanosensing have also shown evidence of micron-scale deformations involved in sensing and responding to substrate stiffness. For example, rat embryo fibroblasts cultured on a micropillar array caused a constant deformation of 0.84 μm regardless of micropillar stiffness (Trichet et al., 2012). Furthermore, it was found that the mechanical properties of substrate-cytoskeleton linkages in Aplysia growth cones were dominated by elastic structures that undergo micron-sized reversible deformations (Mejean et al., 2013). It is not entirely clear at this time what is the physiological significance of the micron-scale deformation, or how can it be reconciled with our understanding of mechanotransduction. However, these micron-scale deformations imply that mechanotransduction not only involves cell adhesion receptors and coupling proteins (Suter et al., 1998; Bard et al., 2008; Shimada et al., 2008), stretch-induced phosphorylation (Suter and Forscher, 2001; Kostic et al., 2007) and activation of ionic channels (Rajnicek and McCaig, 1997; Gomez et al., 2001; Franze et al., 2009; Kerstein et al., 2013), but also involves cytoskeletal reorganization spanning a significant micron-scale spatial range. Clearly, more work is needed to understand the relationship between events taking place at the nano-scale molecular level and cytoskeletal and cytoplasmic domain rearrangements at the micron-scale level in addition to the signaling/feedback mechanisms spanning the two length scales.
How Does the Growth Cone Build Up Traction Force in Substrate-Mediated Growth?
Related to feedback signaling transcending multiple length scales is the question: how does the growth cone build up traction force in substrate-mediated growth? We have shown that traction force increases gradually over time as the growth cone encounters a new adhesion substrate. The maximum level of the force generated depends on the stiffness of the new substrate, implying continuous strengthening of the clutch and/or active recruitment of molecular motors (Athamneh et al., in press). Similar results were reported for rat embryo fibroblasts cultured on a micropillar array (Trichet et al., 2012). We also know from previous studies that the growth cone response to a physically restrained adhesion substrate includes adhesion formation, Src tyrosine phosphorylation, slowing of retrograde F-actin flow, increased actin assembly, advancing of microtubules, and leading edge advance (Suter et al., 1998, 2004; Suter and Forscher, 2001; Lee and Suter, 2008; Schaefer et al., 2008). Despite all this knowledge, the details of how the different events are orchestrated to gradually build up traction force, up to 102 nN range (Athamneh et al., in press), and guide axonal growth in the direction of the adhesion site are not entirely clear.
What is the Actual Role of Molecular Motors in Growth Cone Steering?
Experimental evidence suggests that in addition to actin and microtubule assembly dynamics, a number of molecular motors are involved in adhesion-mediated growth cone steering. Myosin II is essential for generating retrograde actin flow (Lin et al., 1996; Bridgman et al., 2001), growth cone steering (Turney and Bridgman, 2005), and actin-filament recycling in the T zone (Medeiros et al., 2006). Dynein is important for microtubule forward movement during growth cone steering and axonal elongation (Myers et al., 2006; Grabham et al., 2007; Roossien et al., 2014). Additionally, it has been suggested that kinesin-5 is involved in growth cone steering by opposing the action of cytoplasmic dynein to affect selective polarization of microtubules in the P domain (Nadar et al., 2008). Furthermore, kinesin-1-driven microtubule sliding promotes initial axonal extension in developing Drosophila neurons (Lu et al., 2013). However, the details of motor activity during adhesion-mediated growth, including what structures support these motor activities in order to generate pushing/pulling forces, are not known. Also not clear is how motor activity in growth cones is influenced by different substrates (Lin et al., 1996; Turney and Bridgman, 2005; Medeiros et al., 2006; Ketschek et al., 2007; Rösner et al., 2007; Kollins et al., 2009).
What is the Role of Mechanical Force in Regulating Axonal Transport?
On the axon level, it has been show that tension is tightly regulated (Lamoureux et al., 1989; Rajagopalan et al., 2010; Hyland et al., 2014) and induces axonal growth (Bray, 1984; Zheng et al., 1991; Chada et al., 1997; Pfister et al., 2004), although the mechanism is not yet fully understood (Suter and Miller, 2011). For axons to grow, components must be transported. It follows that understanding the effect of tension on axonal transport is key to understanding the mechanisms of tension-induced growth, although very little is known in this area (O’Toole and Miller, 2011). Increased tension has been shown to decrease fast mitochondria transport in DRG neurons (Loverde et al., 2011), but increased fast vesicles transport in Aplysia neurons (Ahmed and Saif, 2014). It was suggested that tension also influences motor activity in the axon, although the details of what motors are affected by tension are not known (Ahmed and Saif, 2014).
In summary, many questions related to the role of force in axonal growth and guidance remain unanswered. We believe that satisfying answers to these questions will require quantitative information about the relationship between elongation, forces, cytoskeletal dynamics, axonal transport, signaling, substrate adhesion, and stiffness contributing to axonal growth and guidance. In the following section, we list experimental techniques used to quantify forces in axons and growth cones and discuss relevant findings.
Techniques for Quantitative Force Measurements in Axons and Growth Cones
Force-Calibrated Microneedles
Bray (1984) was the first to use MNs to demonstrate that neurites grow in response to mechanical tension. Earlier, Bray (1979) used vectorial analysis of the outlines of individually isolated sensory neurons to produce the first evidence that growth cones pull neurites by generating traction force. First quantitative force measurements in axons were performed using force-calibrated MNs by the Heidemann group (Dennerll et al., 1988). A permanent “rest tension” was identified in PC-12 neurites ranging over three orders of magnitude (10−2–100 nN) (Dennerll et al., 1988). In these experiments, the MN was attached to the neurite at a middle point between the soma and the growth cone, and then moved rapidly perpendicular to the neurite’s long axis causing tension in the neurite and needle deflection. The same group later attached a MN to the soma of a cultured chick sensory neuron and raised it so that the cell became attached to the substrate at the growth cone only. Neurite tension was found to be in the 100 nN range and strongly correlated with growth cone advance (Lamoureux et al., 1989). Later, the same group showed that axonal elongation correlated with applied tension using a MN (Zheng et al., 1991). Recently, O’Toole et al. (2015) developed a mathematical model to discern individual subcellular forces within the axon and growth cone by relating theses forces to the net axonal tension measured with a MN. Determination of subcellular forces was enabled by labeling of docked mitochondria to monitor subcellular strain. The mean force generated by the rear of the growth cone and axon was 2.0 and 0.6 nN, respectively, suggesting that contractile forces are generated in microtubule-rich regions at the rear of the growth cone and along the axon. We have used MNs in our laboratory to measure traction force in Aplysia growth cones as they respond to an adhesion substrate (Athamneh et al., in press).
MNs provide a simple, direct, and effective method for applying and measuring forces in neurons with sensitivity in the 10−3 nN range, although the technique can be time-consuming. It does not require specialized instruments or sophisticated analysis procedures. However, the technique is very sensitive to vibration and care must be taken not to cause damage to the cell. Additionally, whereas MNs were effective in measuring traction force in large Aplysia growth cones, they can be too large for measurements in other cell types with smaller growth cones. When cultured on poly-L-lysine, Aplysia growth cones on average cover an area of approximately 1.25 × 103 μm2 (Wu et al., 2008), which is 5–10 times larger than growth cones from other species.
Traction Force Microscopy
In traction force microscopy (TFM), cells are cultured on a deformable substrate, which can be a hydrogel or a nanowire array. Fluorescent beads are embedded as markers within the substrate to facilitate optical detection of the deformation caused by the cell. Force exerted by the cell is calculated using measured deformations and the stiffness of the substrate. Due to the popularity of TFM in the field of cell biomechanics, numerous reviews and method articles have been published describing the theory behind the techniques as well as methods for substrate preparation and fabrication (Sabass et al., 2008; Plotnikov et al., 2014; Style et al., 2014; Schwarz and Soiné, 2015). Bridgman et al. (2001) used the technique to study the role of Myosin IIB in generation of filopodia-mediated traction force in growth cones from mouse superior cervical ganglion neurons. The traction force generated by a single filopodium was found to be in the 100 nN range and partially reduced in Myosin IIB knock-out neurons, suggesting that Myosin IIB was acting in combination with other myosins (Bridgman et al., 2001). Chan and Odde (2008) used TFM to validate their “motor-clutch” model. They identified a 1 kPa threshold for substrate stiffness below which a single filopodium of embryonic-chick—forebrain neurons exhibited oscillatory load-and-fail dynamics, with slower retrograde flow and higher traction forces. On substrates stiffer than 1 kPa, filopodia showed frictional slippage, with fast retrograde flow and low traction forces (Chan and Odde, 2008). Koch et al. (2012) compared traction forces generated by growth cones of rat dorsal root ganglion and hippocampal neurons and found that the growth cones of PNS neurons produce higher traction forces compared to CNS neurons (100 vs. 10−1 nN range). For both neuron types, traction forced increased with increasing stiffness, which is consistent with other studies. Toriyama et al. (2013) observed that netrin-1 positively regulates traction force in the growth cone of rat hippocampal neurons through Pak1-mediated shootin1 phosphorylation, which enhances F-actin-substrate coupling leading to higher force generation. Traction force was found to be highest in the actin-rich P domain of Aplysia growth cones, and although traction force redistributes continuously, the net resulting neurite tension was tightly regulated around 3.1 nN (Hyland et al., 2014). In NG108-15 neuroblastoma cells neurite tension was measured as 0.6 nN (Betz et al., 2011), which is consistent with earlier neurite tension measurements in PC-12 cells using MNs (Dennerll et al., 1988). Finally, using a microfabricated nanowire array TFM revealed a wide range of traction forces (10−2–101 nN) in the growth cones of rat dorsal root ganglion neurons depending on nanowire stiffness (Hällström et al., 2010).
While TFM provides high-resolution force measurements at the growth cone and single filopodia levels, the uniformity of the substrate stiffness does not mimic the complex environment in vivo particularly well. The introduction of a stiffness gradient or a defined stiffness micropattern for neurons to interact with remains a technical challenge. Accordingly, force values reported using conventional TFM on a uniform substrate may represent a “quasi” steady state condition and not traction force generation in response to a change in adhesion substrate.
Optical Tweezers
The optical tweezer (OT; or optical trap) setup consists of a highly focused infrared laser beam that can physically hold (i.e., trap) a microbead. The tweezers can be calibrated to know how much force (typically in 10−3–100 nN range) is required to remove the trapped bead from its focal center. Because of its high force sensitivity, this technique has been extensively used to measure molecular forces produced by proteins, especially molecular motors (Elting and Spudich, 2012). Using OTs, it was shown that the forces generated by the filopodia in the growth cones of CNS (hippocampal) neurons were larger when compared with PNS (Dorsal Root Ganglia) neurons (5 × 10−3 vs. 1–2 × 10−3 nN) (Amin et al., 2013). The lamellipodia of both CNS and PNS neurons generated similar lateral forces level up to 20 × 10−3 nN, but exerted larger vertical force in PNS neurons (4 × 10−3 vs. 1–5 × 10−3 nN) (Amin et al., 2013). Mejean et al. (2013) used an optical trap to characterize the mechanics of apCAM-mediated nascent adhesions in Aplysia growth cones and found that for forces in the 10−3 nN scale, nascent adhesions were dominated by an elastic structure, which can be reversibly deformed by up to 1 μm (Mejean et al., 2013). These results suggested a substrate-cytoskeleton interface dominated by a compliant cross-linked network and not a number of stiff molecular springs in parallel. The compliance of the network may provide more time for nascent adhesions to strengthen before larger forces develop. Also using an optical trap, Bard et al. (2008) studied N-cadherin-mediated substrate-cytoskeletal coupling and found that at low forces (<16 × 10−3 nN) slippage of cadherin-cytoskeleton bonds occurred, while at high forces actin accumulated strengthening nascent N-cadherin coupling. Moore et al. (2009) showed that the advancing growth cones of spinal commissural neurons generated traction force greater than 63 × 10−3 nN when confronted with a restrained netrin-1 bead (Moore et al., 2009). Without the bead, the same growth cone generated 9 × 10−3 nN local traction force as determined by TFM.
Using OTs for force measurements can be technically involved. However, once established, the technique can provide superb resolution and spatial selectivity. It also overcomes the issue of substrate uniformity suffered by TFM. A major limitation of OTs is that the level of force that can be applied without causing damage to the cells is in the 10−3 nN range. Thus, OTs typically cannot sustain enough force to induce adhesion-mediated growth cone advance or steering responses (Bard et al., 2008; Shahapure et al., 2010; Amin et al., 2013; Mejean et al., 2013), especially in neurons with large growth cones such as Aplysia.
Atomic Force Microscopy
Atomic force microscopy (AFM) can be used both for applying and measuring forces in neurons (Franze et al., 2009), although it has been mostly used for imaging and measuring cell topography and elasticity (McNally and Borgens, 2004; Grzywa et al., 2006; Xiong et al., 2009; Spedden et al., 2012). Using AFM in the lateral force measurements mode, Fuhs et al. (2013) measured the forward pushing forces of mouse retinal ganglion cell and NG108-15 growth cones and found them to be on the order of 10−1 nN. In the same paper, the authors reported that the total traction force generated by the NG108-15 growth cones as measured by TFM was two orders of magnitude higher. While AFM lateral force measurements may appear to be an ideal solution for measuring traction force in growth cones, significant technical difficulties exist particularly related to calibrating the torsional response of the AFM cantilever (Karhu et al., 2009). Karhu et al. (2009) showed that frictional-force measurement using AFM was possible in longitudinal imaging mode and provided several advantages over lateral imaging mode. In our lab, we have developed a new AFM-based method for measuring retrograde traction force in growth cones that does not require lateral force calibration by following the approach developed by Karhu et al. (2009). We used the new method with an apCAM-coated colloidal cantilever to measure the temporal traction force profile in Aplysia growth cones as they encounter a physically-restrained adhesion substrate (Athamneh et al., in press). In summary, commercially available AFM systems provide high-resolution data over a large force range from 10−3 to 102 nN. However, calibration and data analysis require significant involvement. Additionally, care must be taken to account for noise and instrumental drift, especially in temporal measurements for an extended period of time.
MEMS Force Sensors
Microelectromechanical system (MEMS)-based force sensors are microfabricated from a single silicon crystal and can provide high-resolution quantitative measurements over a large dynamic range (Rajagopalan and Saif, 2011). Using a MEMS sensor, it was found that axons of embryonic Drosophila neurons that have formed neuromuscular junctions maintain a rest tension of 1–13 nN (Siechen et al., 2009; Rajagopalan et al., 2010), which is in good agreement with the in vitro studies determining neurite rest tension with either TFM (Betz et al., 2011; Hyland et al., 2014) or MNs (Dennerll et al., 1988). Axons responded to perturbation of the rest tension by either relaxing or contracting to restore original rest tension (Rajagopalan et al., 2010). The advantages and limitations of MEMS-based force sensors for the study of cell biomechanics have been reviewed by Rajagopalan and Saif (2011).
Summary and Conclusions
Figure 2 and Table 1 show force values reported in the literature for different parts of the neuron using different experimental techniques. The graph illustrates the wide range of force values even for the same experimental technique or the same part of the neuron. For example, values reported for traction force generated by the growth cone ranged over five orders of magnitude. Indeed, different force values can be expected for different types of neurons and different sizes of neuronal areas probed, which will result in different amounts of cytoskeletal structures and motors that are engaged in the process. Furthermore, different experimental techniques may provide slightly different values for the same neuron and area. However, it is possible that some of the variations in force values could be due to calibration problems. If anything, the discrepancy illustrated in Figure 2 suggests that extreme caution must be taken when comparing force values reported in different studies. Setting up standardized calibration methods for each force measurement method might be an approach to reduce some of the variations currently found in the literature.
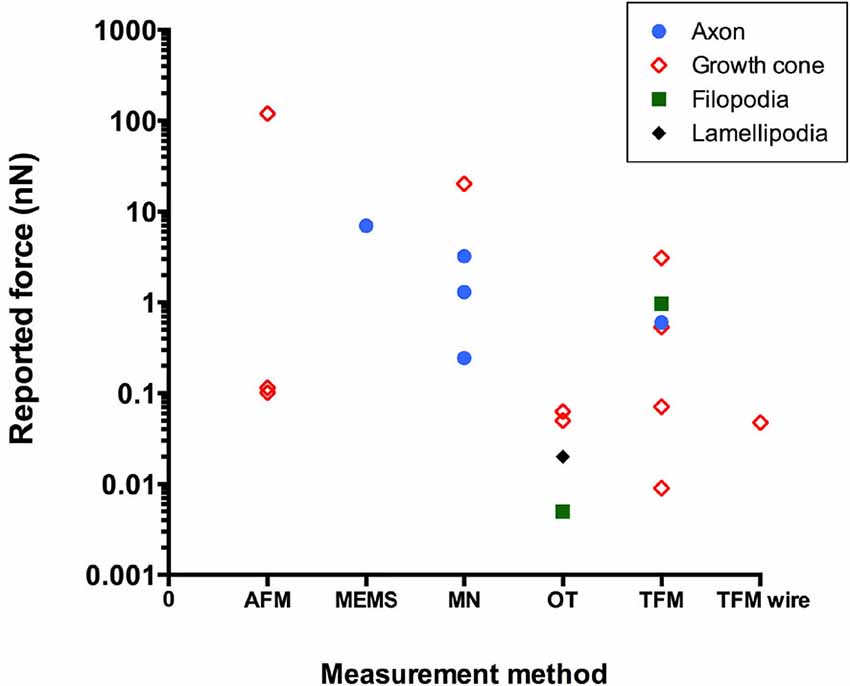
Figure 2. Reported literature values of force in neurons using different experimental techniques (AFM, atomic force microscopy; MEMS, microelectromechanical system-based force sensors; MN, microneedle; OT, optical tweezers; TFM, traction force microscopy).
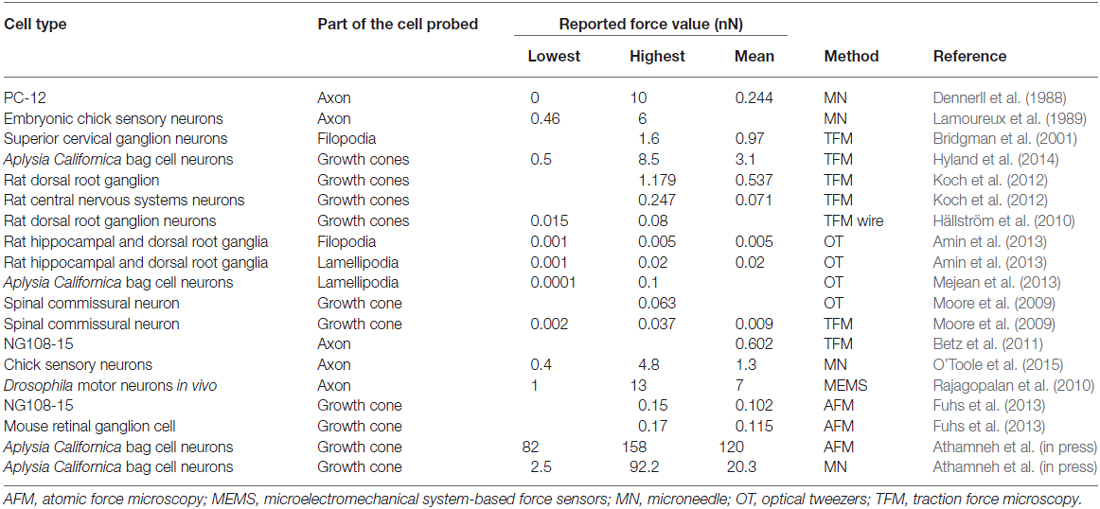
Table 1. Force measurements reported in the literature for different cell types and experimental techniques.
Quantitative studies of mechanical forces in axons and growth cones have yielded significant findings about the critical role of force in growth cone dynamics and axonal elongation. However, many important questions remain unanswered, and apparent discrepancies in the literature, particularly related to the effect of substrate stiffness on neurite outgrowth, need to be addressed. Although the details of the mechanism by which growth cones sense and responds to substrate stiffness are still unclear, it appears to be a highly coordinated mechanism involving a combination of biochemical and biomechanical processes spanning multiple length scales. Understanding the details of this mechanism will undoubtedly require and stimulate more research involving quantitative force measurements.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors are grateful to the National Science Foundation for support of this work under Grant No. 1146944-IOS.
References
Ahmed, W. W., and Saif, T. A. (2014). Active transport of vesicles in neurons is modulated by mechanical tension. Sci. Rep. 4:4481. doi: 10.1038/srep04481
Amin, L., Ercolini, E., Ban, J., and Torre, V. (2013). Comparison of the force exerted by hippocampal and DRG growth cones. PLoS One 8:e73025. doi: 10.1371/journal.pone.0073025
Athamneh, A. I. M., Cartagena-Rivera, A. X., Raman, A., and Suter, D. M. (in press). Substrate deformation predicts neuronal growth cone advance. Biophys. J. (in press).
Bard, L., Boscher, C., Lambert, M., Mège, R.-M., Choquet, D., and Thoumine, O. (2008). A molecular clutch between the actin flow and N-cadherin adhesions drives growth cone migration. J. Neurosci. 28, 5879–5890. doi: 10.1523/JNEUROSCI.5331-07.2008
Betz, T., Koch, D., Lu, Y.-B., Franze, K., and Käs, J. A. (2011). Growth cones as soft and weak force generators. Proc. Natl. Acad. Sci. U S A 108, 13420–13425. doi: 10.1073/pnas.1106145108
Bray, D. (1979). Mechanical tension produced by nerve cells in tissue culture. J. Cell Sci. 37, 391–410.
Bray, D. (1984). Axonal growth in response to experimentally applied mechanical tension. Dev. Biol. 102, 379–389. doi: 10.1016/0012-1606(84)90202-1
Bridgman, P. C., Dave, S., Asnes, C. F., Tullio, A. N., and Adelstein, R. S. (2001). Myosin IIB is required for growth cone motility. J. Neurosci. 21, 6159–6169.
Chada, S., Lamoureux, P., Buxbaum, R. E., and Heidemann, S. R. (1997). Cytomechanics of neurite outgrowth from chick brain neurons. J. Cell Sci. 110, 1179–1186.
Chan, C. E., and Odde, D. J. (2008). Traction dynamics of filopodia on compliant substrates. Science 322, 1687–1691. doi: 10.1126/science.1163595
Dennerll, T. J., Joshi, H. C., Steel, V. L., Buxbaum, R. E., and Heidemann, S. R. (1988). Tension and compression in the cytoskeleton of PC-12 neurites. II: quantitative measurements. J. Cell Biol. 107, 665–674. doi: 10.1083/jcb.107.2.665
Elting, M. W., and Spudich, J. A. (2012). Future challenges in single-molecule fluorescence and laser trap approaches to studies of molecular motors. Dev. Cell 23, 1084–1091. doi: 10.1016/j.devcel.2012.10.002
Flanagan, L. A., Ju, Y. E., Marg, B., Osterfield, M., and Janmey, P. A. (2002). Neurite branching on deformable substrates. Neuroreport 13, 2411–2415. doi: 10.1097/00001756-200212200-00007
Franze, K., Gerdelmann, J., Weick, M., Betz, T., Pawlizak, S., Lakadamyali, M., et al. (2009). Neurite branch retraction is caused by a threshold-dependent mechanical impact. Biophys. J. 97, 1883–1890. doi: 10.1016/j.bpj.2009.07.033
Franze, K., Janmey, P. A., and Guck, J. (2013). Mechanics in neuronal development and repair. Annu. Rev. Biomed. Eng. 15, 227–251. doi: 10.1146/annurev-bioeng-071811-150045
Fuhs, T., Reuter, L., Vonderhaid, I., Claudepierre, T., and Käs, J. A. (2013). Inherently slow and weak forward forces of neuronal growth cones measured by a drift-stabilized atomic force microscope. Cytoskeleton (Hoboken) 70, 44–53. doi: 10.1002/cm.21080
Ghibaudo, M., Saez, A., Trichet, L., Xayaphoummine, A., Browaeys, J., Silberzan, P., et al. (2008). Traction forces and rigidity sensing regulate cell functions. Soft Matter 4, 1836–1843. doi: 10.1039/b804103b
Gomez, T. M., and Letourneau, P. C. (2014). Actin dynamics in growth cone motility and navigation. J. Neurochem. 129, 221–234. doi: 10.1111/jnc.12506
Gomez, T. M., Robles, E., Poo, M.-M., and Spitzer, N. C. (2001). Filopodial calcium transients promote substrate-dependent growth cone turning. Science 291, 1983–1987. doi: 10.1126/science.1056490
Grabham, P. W., Seale, G. E., Bennecib, M., Goldberg, D. J., and Vallee, R. B. (2007). Cytoplasmic dynein and LIS1 are required for microtubule advance during growth cone remodeling and fast axonal outgrowth. J. Neurosci. 27, 5823–5834. doi: 10.1523/jneurosci.1135-07.2007
Grzywa, E. L., Lee, A. C., Lee, G. U., and Suter, D. M. (2006). High-resolution analysis of neuronal growth cone morphology by comparative atomic force and optical microscopy. J. Neurobiol. 66, 1529–1543. doi: 10.1002/neu.20318
Hällström, W., Lexholm, M., Suyatin, D. B., Hammarin, G., Hessman, D., Samuelson, L., et al. (2010). Fifteen-piconewton force detection from neural growth cones using nanowire arrays. Nano Lett. 10, 782–787. doi: 10.1021/nl902675h
Heidemann, S. R., Lamoureux, P., and Buxbaum, R. E. (1990). Growth cone behavior and production of traction force. J. Cell Biol. 111, 1949–1957. doi: 10.1083/jcb.111.5.1949
Hyland, C., Mertz, A. F., Forscher, P., and Dufresne, E. (2014). Dynamic peripheral traction forces balance stable neurite tension in regenerating aplysia bag cell neurons. Sci. Rep. 4:4961. doi: 10.1038/srep04961
Jiang, F. X., Yurke, B., Firestein, B. L., and Langrana, N. A. (2008). Neurite outgrowth on a DNA crosslinked hydrogel with tunable stiffnesses. Ann. Biomed. Eng. 36, 1565–1579. doi: 10.1007/s10439-008-9530-z
Karhu, E., Gooyers, M., and Hutter, J. L. (2009). Quantitative friction-force measurements by longitudinal atomic force microscope imaging. Langmuir 25, 6203–6213. doi: 10.1021/la900005z
Kerstein, P. C., Jacques-Fricke, B. T., Rengifo, J., Mogen, B. J., Williams, J. C., Gottlieb, P. A., et al. (2013). Mechanosensitive TRPC1 channels promote calpain proteolysis of talin to regulate spinal axon outgrowth. J. Neurosci. 33, 273–285. doi: 10.1523/JNEUROSCI.2142-12.2013
Ketschek, A. R., Jones, S. L., and Gallo, G. (2007). Axon extension in the fast and slow lanes: substratum-dependent engagement of myosin II functions. Dev. Neurobiol. 67, 1305–1320. doi: 10.1002/dneu.20455
Koch, D., Rosoff, W. J., Jiang, J., Geller, H. M., and Urbach, J. S. (2012). Strength in the periphery: growth cone biomechanics and substrate rigidity response in peripheral and central nervous system neurons. Biophys. J. 102, 452–460. doi: 10.1016/j.bpj.2011.12.025
Kollins, K. M., Hu, J., Bridgman, P. C., Huang, Y. Q., and Gallo, G. (2009). Myosin-II negatively regulates minor process extension and the temporal development of neuronal polarity. Dev. Neurobiol. 69, 279–298. doi: 10.1002/dneu.20704
Kostic, A., Sap, J., and Sheetz, M. P. (2007). RPTPα is required for rigidity-dependent inhibition of extension and differentiation of hippocampal neurons. J. Cell Sci. 120, 3895–3904. doi: 10.1242/jcs.009852
Lamoureux, P., Buxbaum, R. E., and Heidemann, S. R. (1989). Direct evidence that growth cones pull. Nature 340, 159–162. doi: 10.1038/340159a0
Lee, A. C., and Suter, D. M. (2008). Quantitative analysis of microtubule dynamics during adhesion-mediated growth cone guidance. Dev. Neurobiol. 68, 1363–1377. doi: 10.1002/dneu.20662
Lin, C. H., Espreafico, E. M., Mooseker, M. S., and Forscher, P. (1996). Myosin drives retrograde F-Actin flow in neuronal growth cones. Neuron 16, 769–782. doi: 10.1016/s0896-6273(00)80097-5
Loverde, J. R., Ozoka, V. C., Aquino, R., Lin, L., and Pfister, B. J. (2011). Live imaging of axon stretch growth in embryonic and adult neurons. J. Neurotrauma 28, 2389–2403. doi: 10.1089/neu.2010.1598
Lu, W., Fox, P., Lakonishok, M., Davidson, M. W., and Gelfand, V. I. (2013). Initial neurite outgrowth in drosophila neurons is driven by kinesin-powered microtubule sliding. Curr. Biol. 23, 1018–1023. doi: 10.1016/j.cub.2013.04.050
McNally, H. A., and Borgens, R. B. (2004). Three-dimensional imaging of living and dying neurons with atomic force microscopy. J. Neurocytol. 33, 251–258. doi: 10.1023/b:neur.0000030700.48612.0b
Medeiros, N. A., Burnette, D. T., and Forscher, P. (2006). Myosin II functions in actin-bundle turnover in neuronal growth cones. Nat. Cell Biol. 8, 216–226. doi: 10.1038/ncb1367
Mejean, C. O., Schaefer, A. W., Buck, K. B., Kress, H., Shundrovsky, A., Merrill, J. W., et al. (2013). Elastic coupling of nascent apCAM adhesions to flowing actin networks. PLoS One 8:e73389. doi: 10.1371/journal.pone.0073389
Moore, S. W., Biais, N., and Sheetz, M. P. (2009). Traction on immobilized netrin-1 is sufficient to reorient axons. Science 325:166. doi: 10.1126/science.1173851
Myers, K. A., Tint, I., Nadar, C. V., He, Y., Black, M. M., and Baas, P. W. (2006). Antagonistic forces generated by cytoplasmic dynein and myosin-II during growth cone turning and axonal retraction. Traffic 7, 1333–1351. doi: 10.1111/j.1600-0854.2006.00476.x
Nadar, V. C., Ketschek, A., Myers, K. A., Gallo, G., and Baas, P. W. (2008). Kinesin-5 is essential for growth cone turning. Curr. Biol. 18, 1972–1977. doi: 10.1016/j.cub.2008.11.021
Norman, L., and Aranda-Espinoza, H. (2010). Cortical neuron outgrowth is insensitive to substrate stiffness. Cell. Mol. Bioeng. 3, 398–414. doi: 10.1007/s12195-010-0137-8
O’Toole, M., Lamoureux, P., and Miller, K. E. (2015). Measurement of subcellular force generation in neurons. Biophys. J. 108, 1027–1037. doi: 10.1016/j.bpj.2015.01.021
O’Toole, M., and Miller, K. E. (2011). The role of stretching in slow axonal transport. Biophys. J. 100, 351–360. doi: 10.1016/j.bpj.2010.12.3695
Pfister, B. J., Iwata, A., Meaney, D. F., and Smith, D. H. (2004). Extreme stretch growth of integrated axons. J. Neurosci. 24, 7978–7983. doi: 10.1523/jneurosci.1974-04.2004
Plotnikov, S. V., Sabass, B., Schwarz, U. S., and Waterman, C. M. (2014). “Chapter 20 – high-resolution traction force microscopy,” in Methods in Cell Biology, Vol. 123, eds J. C. Waters and T. Wittman (Atlanta, GA: Elsevier), 367–394. doi: 10.1016/B978-0-12-420138-5.00020-3
Rajagopalan, J., and Saif, M. T. A. (2011). MEMS Sensors and Microsystems for Cell Mechanobiology. J. Micromech. Microeng. 21, 054002–054012. doi: 10.1088/0960-1317/21/5/054002
Rajagopalan, J., Tofangchi, A., and A. Saif, M. T. (2010). Drosophila neurons actively regulate axonal tension in vivo. Biophys. J. 99, 3208–3215. doi: 10.1016/j.bpj.2010.09.029
Rajnicek, A., and McCaig, C. (1997). Guidance of CNS growth cones by substratum grooves and ridges: effects of inhibitors of the cytoskeleton, calcium channels and signal transduction pathways. J. Cell Sci. 110, 2915–2924.
Rösner, H., Möller, W., Wassermann, T., Mihatsch, J., and Blum, M. (2007). Attenuation of actinomyosinII contractile activity in growth cones accelerates filopodia-guided and microtubule-based neurite elongation. Brain Res. 1176, 1–10. doi: 10.1016/j.brainres.2007.07.081
Roossien, D. H., Lamoureux, P., and Miller, K. E. (2014). Cytoplasmic dynein pushes the cytoskeletal meshwork forward during axonal elongation. J. Cell Sci. 127, 3593–3602. doi: 10.1242/jcs.152611
Sabass, B., Gardel, M. L., Waterman, C. M., and Schwarz, U. S. (2008). High resolution traction force microscopy based on experimental and computational advances. Biophys. J. 94, 207–220. doi: 10.1529/biophysj.107.113670
Saez, A., Buguin, A., Silberzan, P., and Ladoux, B. (2005). Is the mechanical activity of epithelial cells controlled by deformations or forces? Biophys. J. 89, L52–L54. doi: 10.1529/biophysj.105.071217
Schaefer, A. W., Schoonderwoert, V. T., Ji, L., Mederios, N., Danuser, G., and Forscher, P. (2008). Cytoskeletal dynamics underlying neurite outgrowth. Dev. Cell 15, 146–162. doi: 10.1016/j.devcel.2008.05.003
Schwarz, U. S., and Soiné, J. R. D. (2015). Traction force microscopy on soft elastic substrates: a guide to recent computational advances. Biochim. Biophys. Acta doi: 10.1016/j.bbamcr.2015.05.028 [Epub ahead of print].
Shahapure, R., Difato, F., Laio, A., Bisson, G., Ercolini, E., Amin, L., et al. (2010). Force generation in lamellipodia is a probabilistic process with fast growth and retraction events. Biophys. J. 98, 979–988. doi: 10.1016/j.bpj.2009.11.041
Shimada, T., Toriyama, M., Uemura, K., Kamiguchi, H., Sugiura, T., Watanabe, N., et al. (2008). Shootin1 interacts with actin retrograde flow and L1-CAM to promote axon outgrowth. J. Cell Biol. 181, 817–829. doi: 10.1083/jcb.200712138
Siechen, S., Yang, S., Chiba, A., and Saif, T. (2009). Mechanical tension contributes to clustering of neurotransmitter vesicles at presynaptic terminals. Proc. Natl. Acad. Sci. U S A 106, 12611–12616. doi: 10.1073/pnas.0901867106
Spedden, E., White, J. D., Naumova, E. N., Kaplan, D. L., and Staii, C. (2012). Elasticity maps of living neurons measured by combined fluorescence and atomic force microscopy. Biophys. J. 103, 868–877. doi: 10.1016/j.bpj.2012.08.005
Stabenfeldt, S. E., and LaPlaca, M. C. (2011). Variations in rigidity and ligand density influence neuronal response in methylcellulose-laminin hydrogels. Acta Biomater. 7, 4102–4108. doi: 10.1016/j.actbio.2011.07.026
Style, R. W., Boltyanskiy, R., German, G. K., Hyland, C., MacMinn, C. W., Mertz, A. F., et al. (2014). Traction force microscopy in physics and biology. Soft Matter 10, 4047–4055. doi: 10.1039/c4sm00264d
Suter, D. M., Errante, L. D., Belotserkovsky, V., and Forscher, P. (1998). The ig superfamily cell adhesion molecule, apCAM, mediates growth cone steering by substrate–cytoskeletal coupling. J. Cell Biol. 141, 227–240. doi: 10.1083/jcb.141.1.227
Suter, D. M., and Forscher, P. (2000). Substrate–cytoskeletal coupling as a mechanism for the regulation of growth cone motility and guidance. J. Neurobiol. 44, 97–113. doi: 10.1002/1097-4695(200008)44:2<97::aid-neu2>3.0.co;2-u
Suter, D. M., and Forscher, P. (2001). Transmission of growth cone traction force through apCAM–cytoskeletal linkages is regulated by Src family tyrosine kinase activity. J. Cell Biol. 155, 427–438. doi: 10.1083/jcb.200107063
Suter, D. M., and Miller, K. E. (2011). The emerging role of forces in axonal elongation. Prog. Neurobiol. 94, 91–101. doi: 10.1016/j.pneurobio.2011.04.002
Suter, D. M., Schaefer, A. W., and Forscher, P. (2004). Microtubule dynamics are necessary for Src family kinase-dependent growth cone steering. Curr. Biol. 14, 1194–1199. doi: 10.1016/j.cub.2004.06.049
Toriyama, M., Kozawa, S., Sakumura, Y., and Inagaki, N. (2013). Conversion of a signal into forces for axon outgrowth through pak1-mediated shootin1 phosphorylation. Curr. Biol. 23, 529–534. doi: 10.1016/j.cub.2013.02.017
Trichet, L., Le Digabel, J., Hawkins, R. J., Vedula, S. R. K., Gupta, M., Ribrault, C., et al. (2012). Evidence of a large-scale mechanosensing mechanism for cellular adaptation to substrate stiffness. Proc. Natl. Acad. Sci. U S A 109, 6933–6938. doi: 10.1073/pnas.1117810109
Turney, S. G., and Bridgman, P. C. (2005). Laminin stimulates and guides axonal outgrowth via growth cone myosin II activity. Nat. Neurosci. 8, 717–719. doi: 10.1038/nn1466
Wu, B., Decourt, B., Zabidi, M. A., Wuethrich, L. T., Kim, W. H., Zhou, Z., et al. (2008). Microtubule-mediated Src tyrosine kinase trafficking in neuronal growth cones. Mol. Biol. Cell 19, 4611–4627. doi: 10.1091/mbc.e08-06-0603
Xiong, Y., Lee, A. C., Suter, D. M., and Lee, G. U. (2009). Topography and nanomechanics of live neuronal growth cones analyzed by atomic force microscopy. Biophys. J. 96, 5060–5072. doi: 10.1016/j.bpj.2009.03.032
Yip, A. K., Iwasaki, K., Ursekar, C., Machiyama, H., Saxena, M., Chen, H., et al. (2013). Cellular response to substrate rigidity is governed by either stress or strain. Biophys. J. 104, 19–29. doi: 10.1016/j.bpj.2012.11.3805
Keywords: growth cone biomechanics, axon elongation, mechanotransduction, cytoskeleton, biophysics, traction force
Citation: Athamneh AIM and Suter DM (2015) Quantifying mechanical force in axonal growth and guidance. Front. Cell. Neurosci. 9:359. doi: 10.3389/fncel.2015.00359
Received: 21 May 2015; Accepted: 27 August 2015;
Published: 16 September 2015.
Edited by:
Dieter Wicher, Max Planck Institute for Chemical Ecology, GermanyCopyright © 2015 Athamneh and Suter. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution and reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Daniel M. Suter, Bindley Bioscience Center, Birck Nanotechnology Center, Department of Biological Sciences, Purdue University, 915 West State Street, West Lafayette, IN 47907-2054, USA,ZHN1dGVyQHB1cmR1ZS5lZHU=