 Yuan Zhang
Yuan Zhang Li Shu
Li Shu Qiying Sun
Qiying Sun Xun Zhou1
Xun Zhou1 Jifeng Guo
Jifeng Guo- 1Department of Neurology, Xiangya Hospital, Central South University, Changsha, China
- 2Department of Geriatrics, Xiangya Hospital, Central South University, Changsha, China
- 3National Clinical Research Center for Geriatric Disorders, Changsha, China
- 4Key Laboratory of Hunan Province in Neurodegenerative Disorders, Central South University, Changsha, China
- 5Center for Medical Genetics, School of Life Sciences, Central South University, Changsha, China
- 6Parkinson's Disease Center of Beijing Institute for Brain Disorders, Beijing, China
Background: Numerous studies have indicated that there is a possible relationship between GBA variants and Parkinson's disease (PD), however, most of them focused on a few variants such as L444P, N370S. We performed a comprehensive pooled analysis to clarify the relationship between variations of GBA and the risk of PD in different racial groups.
Methods: Standard meta-analysis was conducted, including generating inclusion and exclusion criteria, searching literature, extracting and analyzing data.
Results: Fifty studies containing 20,267 PD patients and 24,807 controls were included. We found that variants 84insGG, IVS2+1G>A, R120W, H255Q, E326K, T369M, N370S, D409H, L444P, R496H and RecNciI increased the risk of PD in total populations (OR: 1.78–10.49; p: <0.00001, 0.00005, 0.0008, 0.005, <0.00001, 0.004, <0.00001, 0.0003, <0.00001, <0.0001, 0.0001). In subgroup analysis by ethnicity, in AJ populations, variants 84insGG, R496H, N370S increased the risk of PD (OR: 9.26–3.51; p: <0.00001, <0.0001, <0.00001). In total non-AJ populations, variants L444P, R120W, IVS2+1G>A, H255Q, N370S, D409H, RecNciI, E326K, T369M increased the risk of PD (OR: 8.66–1.89; p: <0.00001, 0.0008, 0.02, 0.005, <0.00001, 0.001, 0.0001, <0.00001, 0.002). Among the non-AJ populations, pooled analysis from five different groups were done separately. Variants L444P, N370S, H255Q, D409H, RecNciI, E326K increased risk of PD (OR: 6.52–1.84; p: <0.00001, <0.00001, 0.005, 0.005, 0.04, <0.00001) in European/West Asians while R120W and RecNciI in East Asians (OR: 14.93, 3.56; p: 0.001, 0.003). L444P increased the risk of PD in Hispanics, East Asians and Mixed populations (OR: 15.44, 12.43, 7.33; p: 0.00004, <0.00001, 0.009). Lacking of enough original studies, we failed to conduct quantitative analysis in Africa.
Conclusions: Obvious racial differences were found for GBA variants in PD. 84insGG and R496H exclusively increased PD risks in AJ populations, so did L444P, R120W, IVS2+1G>A, H255Q, D409H, RecNciI, E326K, T369M in non-AJ populations. N370S increased the risk of PD in both ethnics. In non-AJ subgroup populations, N370S, H255Q, D409H, E326K exclusively increased PD risks in European/West Asians, as were R120W in East Asians. L444P increased the risk of PD in all groups in non-AJ ethnicity. These results will contribute to the future genetic screening of GBA gene in PD.
Introduction
Parkinson's disease (PD), one of the most common progressive neurodegenerative disorders, is characterized by motor and non-motor symptoms. Although the etiology of PD is still unclear, it is convincing that the interaction of genetic factor, environmental factor and aging together contribute to the disease (Kalia and Lang, 2015).
GBA encodes lysosomal enzyme glucocerebrosidase (GCase), the homozygous mutations of which can lead to accumulation of glucocerebroside and storage of lysosomal lipid, thus causing Gaucher's disease (GD) (Grabowski, 1993). More than 300 mutations including point mutations, insertions, deletions and frameshift mutations were discovered as the pathogenic variants of GD nowadays (O'Regan et al., 2017). In the year 2004, Lwin et.al firstly identified a higher rate of GBA mutations, either heterozygous or homozygous (N370S, L444P, K198T, and R329C), in brain samples from PD patients, which indicated a possible relationship between GBA mutations and the disease (Lwin et al., 2004). Since then, numbers of studies have confirmed that GBA mutations were associated with PD risks (Aharon-Peretz et al., 2004; Clark et al., 2005).
A prominent genetic feature of GBA mutations in PD was ethnic heterogeneity in different regions. Several meta-analyses (Mao et al., 2013; Chen et al., 2014; Zhao et al., 2016) have demonstrated that GBA variants like L444P and N370S are risk factors for PD. However, pooled analysis of the association between other GBA variants and the risk of PD is still lacking. And the few available original studies investigating associations between GBA variants, such as 84insGG, R120W, and R496H, and the risk of PD yielded inconsistent results. Thus, we performed a comprehensive meta-analysis to clarify the relationship between the potential variations of GBA and the risk of PD in total populations from different ethnicities, which would be essential in genetic screening in PD patients from different populations with different ethnic background.
Materials and Methods
Inclusion Criteria
We performed our meta-analysis based on PICOS (participants, interventions, controls, outcomes, and studies) rules.
Participants: all PD patients were diagnosed with any accepted criteria without the requirement negative of family history.
Interventions: the DNA from peripheral blood of all cases and controls were analyzed using PCR based methods or other accepted methods.
Controls: all controls were reported healthy controls or had no PD or other neurological disorders.
Outcomes: the number of cases or controls carrying homozygous or heterozygous variants of GBA was reported (unless articles pointing out that there were homozygous variants of GBA screened out in PD patients, we considered the variants of GBA as heterozygous).
Studies: types of all studies were case-control study or cohort study.
Literature Search
With the key words of (“parkinson*” or “PD”) and (“GBA” or “glucocerebrosidase”), we searched the electronic databases including Pubmed, embase, web of science (WOS) and the Cochrane library for English publications up to October 20th, 2017. All searched papers were imported into ENDNOTE for further management. One copy of the overlapping articles from different databases was kept with the help of electronic and manual checking. Two researchers performed the search independently. If there were controversial ideas, the third researcher was asked to solve the debates.
Data Extraction
For studies satisfying the aforesaid criteria, two authors independently extracted the following data: year of publication, first author's name, countries, ethnicities, numbers of PD patients and controls, sequencing strategy (all GBA exons or specific variants), the sequencing results of specific variants' genotypes and alleles in PD patients and controls. Complete data were independently extracted by two researchers. Due the prominent ethnic heterogeneity, especially in AJ and non-AJ populations (Sidransky et al., 2009; Sun et al., 2010), of GBA mutations in PD in different regions, all ethnicities information including AJ, non-AJ (Africans, European/West Asians, Hispanics, East Asians, Mixed:composed of at least two different groups) (Risch et al., 2002), were extracted according the original studies. Once confronted with debates or difficulties, the third researcher was asked to make the decision. Newcastle-Ottawa Scale (NOS) (Stang, 2010), a good tool to assess the quality of case-control study from the aspects of selection, comparability and exposure, was used to assess the quality of all included case-control studies.
Statistical Analysis
Revman 5.3 software was used to do all the statistical analysis. Pooled odds ratio (OR) and 95% CI (confidence interval) were calculated to assess the strength of association between the variants and PD. Heterogeneity across each study was identified by a standard Q test. Q statistic (P < 0.1) and I2 statistic (>50%) indicated heterogeneity of the analysis. If the heterogeneity was not significant (P > 0.1, I2 ≤ 50%), a fixed-effects model (FM) was used to do the analysis. Otherwise a random-effects model (RM) was applied. Reporting biases were measured by funnel plot analysis. In order to measure the stability of the analysis, sensitivity analysis was performed by removing each individual study in turn from the total and re-analyzing the results from the remainder.
Results
A total of 3,006 publications were retrieved from all searched databases and finally 50 studies were included for further analysis (Figure 1). The characteristics of all included studies, containing 20,267 PD patients and 24,807 controls, were shown on Table 1. The NOS scores of each study indicated that all of them were of good quality.
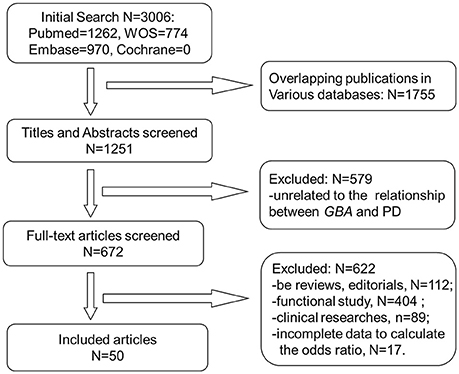
Figure 1. Flow chart of publications included process.
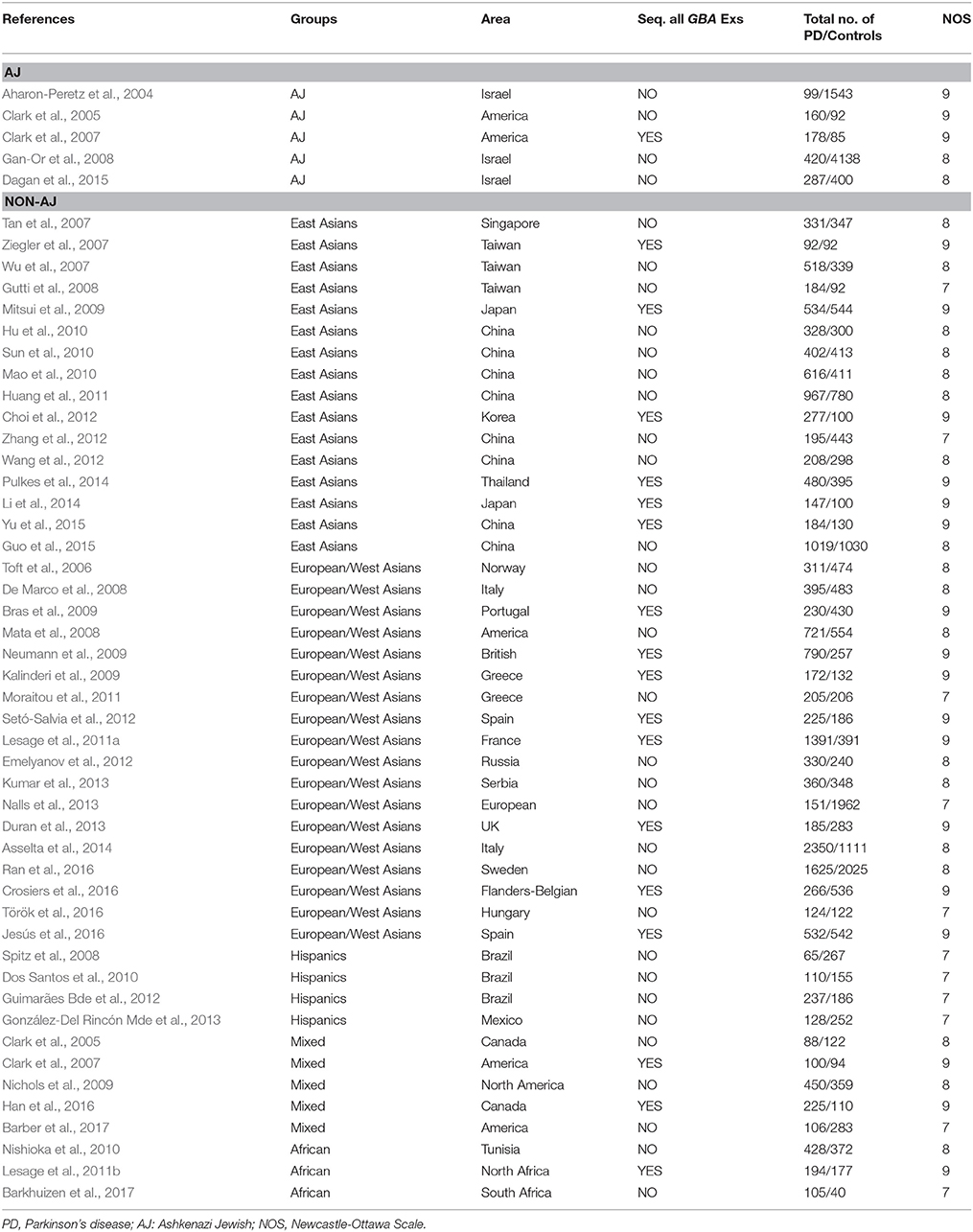
Table 1. The characteristics of all publications included.
According to the detecting methods, 17 of 50 publications sequenced all exons of GBA in participants and reported a series of variants either associated with PD or not (Supplementary Table 1). And other 33 included studies just detected specific variants (Supplementary Table 2). Even though more than 130 variants of GBA were researched in PD cohorts of all included publications, most of which were only reported by one or two studies. We further conducted meta-analysis for 18 variants of all included variants as they had sufficient data (effected variant carriers reported in at least four articles) (Figure 2 and Supplementary Table 3).
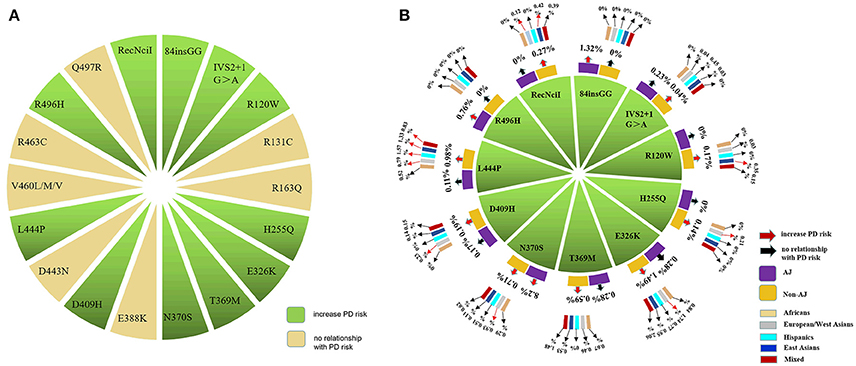
Figure 2. A total of 18 GBA variants were included and conducted in the meta-analysis. (A) 18 GBA variants related to PD were included for meta-analysis according to our inclusion and exclusion criteria. (B) 11 of 18 variants were found to increase the risk for PD. And the MAFs of the 11 risk variants were largely ethnicity dependent.
We found that 11 of the 18 variants, including 84insGG, IVS2+1G>A, R120W, H255Q, E326K, T369M, N370S, D409H, L444P, R496H and RecNciI, increased the risk of PD in total populations (p: < 0.00001, 0.00005, 0.0008, 0.005, < 0.00001, 0.004, < 0.00001, 0.0003, < 0.00001, < 0.0001, 0.0001) with the ORs ranging from 1.78 to 10.49 (Table 2 and Supplementary Figure 1). In subgroup analysis by ethnicities, in AJ populations, three variants of GBA were found to be in statistical difference between PD patients and controls. The variants listed by ORs from the top to the bottom were 84insGG, R496H, N370S (OR: 9.26–3.51; p: < 0.00001, < 0.0001, < 0.00001). In total non-AJ populations, nine variants can increase the risk of PD. Results of variants from the highest OR values to the lowest OR values were L444P, R120W, IVS2+1G>A, H255Q, N370S, D409H, RecNciI, E326K, T369M (OR: 8.66–1.89; p: < 0.00001, 0.0008, 0.02, 0.005, < 0.00001, 0.001, 0.0001, < 0.00001, 0.002). Among the non-AJ populations, pooled analysis from five different groups were done separately. In European/West Asians, six variants were associated with increased risk of PD. Variants having highest OR values to the lowest were L444P, N370S, H255Q, D409H, RecNciI, E326K (OR: 6.52–1.84; p: < 0.00001, < 0.00001, 0.005, 0.005, 0.04, < 0.00001). L444P increased the risk of PD in Hispanics, East Asians and Mixed populations (OR: 15.44, 12.43, 7.33; p: 0.00004, < 0.00001, 0.009). In East Asian populations, besides L444P, R120W and RecNciI can also increase the risk of PD (OR: 14.93, 3.56; p: 0.001, 0.003). For the lack of enough original studies, we were not able to conduct meta-analysis in Africa (Table 2 and Supplementary Figure 2).
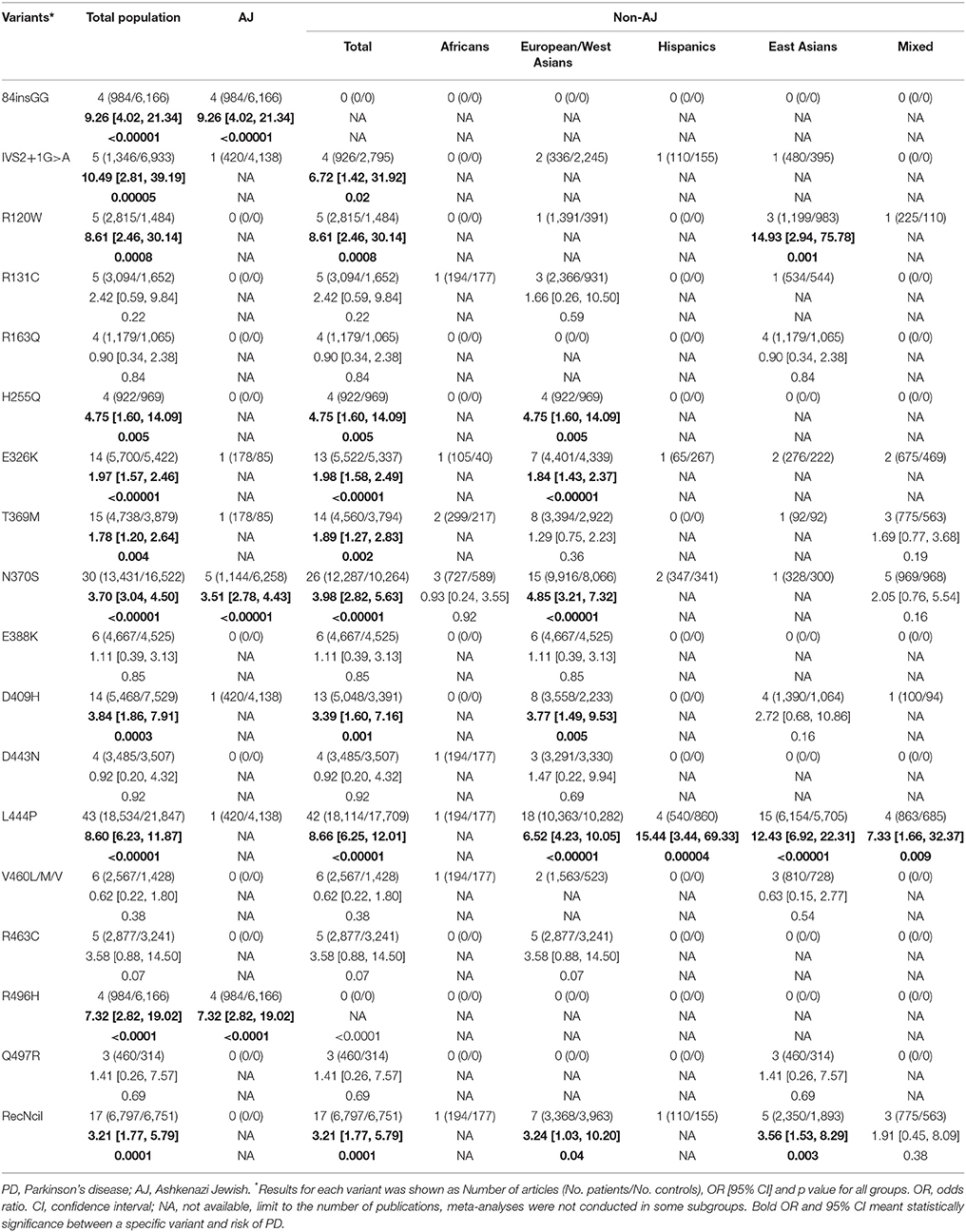
Table 2. The association between variants included in GBA and the risk of PD.
Totally, the carriers' MAFs among all 11 risk variants of GBA, from the highest to the lowest, were E326K, N370S, L444P, T369M, RecNciI, 84insGG, D409H, R120W, H255Q, R496H, IVS2+1G>A with the range from to 1.46 to 0.06%. Besides, the frequencies were largely ethnicity dependent (Figure 2 and Supplementary Table 4). In AJ populations, the top five common variants by the MAFs were N370S, 84insGG, R496H, E326K, T369M (8.2–0.28%). In non-AJ populations as a whole, the variants with top five MAFs were E326K, L444P, N370S, T369M, RecNciI (1.49–0.27%). 84insGG and R496H were screened out only in AJ populations while RecNciI was exclusively existed in non-AJ populations. In subgroup analysis in non-AJ populations, the variants with high MAFs were similar with some specificities such as the L444P variant ranked the top MAFs in East Asian and Hispanics while E326K had the top MAFs in other three populations. Considering the limitations of the small sample size of case-control studies, we checked the allele frequencies of 16 variants among 18 (88insGG and RecNciI not covered) from the Genome Aggregation Database (gnomAD) (http://gnomad.broadinstitute.org/). The trends of the frequencies' differences between cases and controls reported in gnomAD for the 11 positive variants are comparable with our meta-analysis, and our results were even more significant (Supplementary Table 4).
During the sensitivity analysis, the pooled OR and 95% CI of all variants did not change significantly when deleted one article in turn every time. What's more, there were not significant publication biases from the symmetric shapes of all funnel plots (Supplementary Figures 3, 4).
Discussion
We performed a comprehensive analysis between variants in GBA and the risk of PD, which was an update and a complement to previous meta-analysis (Chen et al., 2014; Zhao et al., 2016) on GBA gene. In order to reach more reliable conclusions, we incorporated as many relevant original studies as possible. Compared to the obvious heterogeneities between previous studies, what we provided here is a more precise assessment focused on the association between GBA variants and risk of PD in total populations with diverse ethnics.
In our analysis of total populations, we provided detailed information about the sequencing methods of each original article. By retrieving information from articles screening specific variants or the full exons of GBA, we included all the published positive results and negative results to reach a more precise conclusion. As can be seen from Supplementary Tables 1, 2, there were more than 130 variants of GBA in the association analysis in PD. They can either increase the risk of PD or have no relationship with the disease. In our meta-analysis, we only included 18 variants with enough original studies as supporting evidence. As a result, we found 11 variants associated with increased risk of PD. There were no quantitative results for other a hundred-odd variant on which further researches are needed to explain their role in PD. It is true that sequencing full exons of GBA could find new potential related variants in PD. These information support that sequencing the entire gene would be more informative than just genotyping the risk variants for PD.
To develop more specific genetic screening strategies, we did subgroup analysis by ethnicities in AJ populations and non-AJ populations based on different features of GBA mutations in these two ethnics. In AJ populations, the frequencies of GBA mutations ranged from 10 to 31% while in non-AJ populations, the frequencies fluctuated between 2.9 and 12% (Sun et al., 2010). Variants such as N370S in GBA was more common in AJ populations with European origin while L444P was more common in non-AJ populations such as China (Sidransky et al., 2009; Sun et al., 2010). Therefore, we conducted our meta-analysis in AJ populations and non-AJ populations separately. As a result, we successfully demonstrated significant ethnic heterogeneities in GBA variants included in our meta-analysis. 84insGG and R496H exclusively increased PD risks in AJ populations (MAF: 1.32%, 0.76%). As what were done by mutations including L444P, R120W, IVS2+1G>A, H255Q, D409H, RecNciI, E326K, T369M in non-AJ populations (MAF: 0.98, 0.17, 0.04, 0.14, 0.19, 0.27, 1.49, 0.59%). N370S was associated with the risk of PD in both ethnics (MAF: 8.20, 0.71%). In non-AJ populations, data from previous reports showed that the frequencies varied in different areas, such as 0.4% in North America, 1.7% in Norway etc. (Neumann et al., 2009). In that case, we did the meta-analysis on different districts in non-AJ populations in PD based on standard racial separations (Risch et al., 2002). From the results of five ethnics in non-AJ populations, N370S, H255Q, D409H, E326K were exclusively related to PD risks in European/West Asians (MAF: 0.93, 0.21, 0.23, 1.74%) while R120W in East Asians (MAF: 0.35%). L444P increased the risk of PD in all populations in non-AJ ethnicity [MAF: 1.33% (East Asians), 0.79% (European/West Asians), 1.57% (Hispanics), 0.52% (African), 0.83% (Mixed)]. In addition, combined with frequency analysis, we found risk variants of GBA in PD mainly located on exons 8, 9, and 10, such as E326K, T369M, N370S, D409H, and L444P. For future genetic analysis of GBA in PD, taking priority to screen the hot exons 8, 9, and 10 and focusing on those ethnicity dependent risk variants may be more time and money saving (Figures 2, 3). In that case, our meta-analysis can be a valuable reference for researchers to choose specific variants in building targeted reseq panel for PD patients according to different ethnics and groups. However, it's worth noticing that although those positive variants of GBA could increase the risk of PD, it does not necessarily mean that the carriers of GBA risk variants would develop PD. Because the etiology of PD is still unclear, genetic background is only one of the interactive factors contributing to the PD pathogenesis.
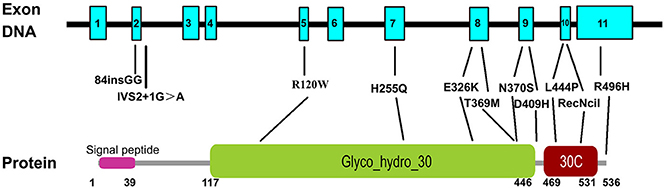
Figure 3. Schematic representation of the GBA gene and protein with the 11 risk variants. All variants are named following the common nomenclature which begins 39 codons downstream from the first ATG, excluding the 39-residue signal peptide. GBA has 11 exons and the protein has three domains, signal peptide, glyco_hydro_30, and glycol_hydro_30C. Numbers below the protein line indicate the boundaries of each domain.
GBA gene encodes GCase. Heterozygous GBA mutations in PD were associated with decreased GCase activities (Ortega et al., 2016). And the activities of GCase in PD patients with GBA homozygotes were even lower than that of heterozygotes (Alcalay et al., 2015). Decreased GCase level could assist α-synuclein accumulation either by interfering protein clearance or by promoting protein aggregation in Lewy bodies. This mechanism were also identified in dementia with Lewy bodies (DLB), which added to the view that GBA is a main genetic risk factor for DLB (Beavan and Schapira, 2013). In addition to the aforementioned mechanism for enzymatic loss of function of GBA leading to PD, the gain of function mechanism was also discovered in GBA related PD where GCase co-localized with α-synuclein (Westbroek et al., 2011). As to the pathological changes underlying specific polymorphisms, although the information was rare, researchers observed reduced activity of GBA enzyme in brains of PD patients with heterozygous N370S mutation. And PD patients with heterozygous L444P mutation were also shown to have presynaptic dopaminergic neuronal dysfunction via neuroimaging examinations (Clark et al., 2007). Thus, whether it is the gain function or loss function of GBA enzyme leads to PD remains unclear.
In clinical use, distinct feature of GBA variants carriers in PD should be paid attention. In previous studies, our group observed unique clinical characteristics of GBA-related PD. Those patients carrying GBA variants were more likely to develop PD at earlier ages, develop bradykinesia as an initial symptom, have a family history and develop non-motor symptoms such as dementia (Zhang et al., 2015). In such case, GBA screening could be conducted in those special PD patients to increase the detection rate. What's more, patients could get more precise treatment in the future when their clinical features and genetic features were combined together.
However, the meta-analysis still has some inevitable limitations. First of all, some original studies without detailed information to calculate OR and 95% CI were not included. Second, the patients and controls of varies clinical features were not separated to do subgroup analysis which may cause biases. Third, due to the limitations of each original study such as the sequencing methods and the sample size, the heterogeneities cannot be totally avoided. For instance, when an original study screened only specific variants, the frequency of other variants may be underestimated.
Conclusion
In conclusion, the contribution of GBA variants to the development of PD is racial dependent. 84insGG and R496H exclusively increased PD risks in AJ populations while L444P, R120W, IVS2+1G>A, H255Q, D409H, RecNciI, E326K, T369M in non-AJ populations. N370S increased the risk of PD in both ethnics. In non-AJ subgroup populations, N370S, H255Q, D409H, E326K exclusively increased PD risks in European/West Asians while R120W in East Asians. L444P increased the risk of PD in all groups in non-AJ ethnicity. Our results will do great help to future GBA screening in clinical use.
Author Contributions
YZ, LS, and BT: Conceived and designed the experiments; YZ, LS, and QS: Performed the experiments; YZ, LS, QS, BT: analyzed the data; YZ, LS, and BT: Wrote the manuscript; XZ, HP, and JG: Reference collection and data management.
Funding
This work was supported by grants from the National Natural Science Foundation of China (No. 81430023, No. 81401059), the National Key Plan for Scientific Research and Development of China (NO. 2016YFC1306000, 2017YFC0909100) and Hunan Provincial Innovation Foundation for Postgraduate (NO. CX2017B066).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnmol.2018.00043/full#supplementary-material
References
Aharon-Peretz, J., Rosenbaum, H., and Gershoni-Baruch, R. (2004). Mutations in the glucocerebrosidase gene and Parkinson's disease in Ashkenazi Jews. N. Engl. J. Med. 351, 1972–1977. doi: 10.1056/NEJMoa033277
Alcalay, R. N., Levy, O. A., Waters, C. C., Fahn, S., Ford, B., Kuo, S. H., et al. (2015). Glucocerebrosidase activity in Parkinson's disease with and without GBA mutations. Brain 138, 2648–2658. doi: 10.1093/brain/awv179
Asselta, R., Rimoldi, V., Siri, C., Cilia, R., Guella, I., Tesei, S., et al. (2014). Glucocerebrosidase mutations in primary Parkinsonism. Parkinson. Relat. Disord. 20, 1215–1220. doi: 10.1016/j.parkreldis.2014.09.003
Barber, T. R., Lawton, M., Rolinski, M., Evetts, S., Baig, F., Ruffmann, C., et al. (2017). Prodromal Parkinsonism and neurodegenerative risk stratification in REM sleep behavior disorder. Sleep 40:zsx071. doi: 10.1093/sleep/zsx071
Barkhuizen, M., Anderson, D. G., van der Westhuizen, F. H., and Grobler, A. F. (2017). A molecular analysis of the GBA gene in Caucasian South Africans with Parkinson's disease. Mol. Genet. Genomic Med. 5, 147–156. doi: 10.1002/mgg3.267
Beavan, M. S., and Schapira, A. H. (2013). Glucocerebrosidase mutations and the pathogenesis of Parkinson disease. Ann. Med. 45, 511–521. doi: 10.3109/07853890.2013.849003
Bras, J., Paisan-Ruiz, C., Guerreiro, R., Ribeiro, M. H., Morgadinho, A., Januario, C., et al. (2009). Complete screening for glucocerebrosidase mutations in Parkinson disease patients from Portugal. Neurobiol. Aging 30, 1515–1517. doi: 10.1016/j.neurobiolaging.2007.11.016
Chen, J., Li, W., Zhang, T., Wang, Y. J., Jiang, X. J., and Xu, Z. Q. (2014). Glucocerebrosidase gene mutations associated with Parkinson's disease: a meta-analysis in a Chinese population. PLoS ONE 9:e115747. doi: 10.1371/journal.pone.0115747
Choi, J. M., Kim, W. C., Lyoo, C. H., Kang, S. Y., Lee, P. H., Baik, J. S., et al. (2012). Association of mutations in the glucocerebrosidase gene with Parkinson disease in a Korean population. Neurosci. Lett. 514, 12–15. doi: 10.1016/j.neulet.2012.02.035
Clark, L. N., Nicolai, A., Afridi, S., Harris, J., Mejia-Santana, H., Strug, L., et al. (2005). Pilot association study of the beta-glucocerebrosidase N370S allele and Parkinson's disease in subjects of Jewish ethnicity. Mov. Disord. 20, 100–103. doi: 10.1002/mds.20320
Clark, L. N., Ross, B. M., Wang, Y., Mejia-Santana, H., Harris, J., Louis, E. D., et al. (2007). Mutations in the glucocerebrosidase gene are associated with early-onset Parkinson disease. Neurology 69, 1270–1277. doi: 10.1212/01.wnl.0000276989.17578.02
Crosiers, D., Verstraeten, A., Wauters, E., Engelborghs, S., Peeters, K., Mattheijssens, M., et al. (2016). Mutations in glucocerebrosidase are a major genetic risk factor for Parkinson's disease and increase susceptibility to dementia in a Flanders-Belgian cohort. Neurosci. Lett. 629, 160–164. doi: 10.1016/j.neulet.2016.07.008
Dagan, E., Schlesinger, I., Ayoub, M., Mory, A., Nassar, M., Kurolap, A., et al. (2015). The contribution of Niemann-Pick SMPD1 mutations to Parkinson disease in Ashkenazi Jews. Parkinsonism Relat. Disord. 21, 1067–1071. doi: 10.1016/j.parkreldis.2015.06.016
De Marco, E. V., Annesi, G., Tarantino, P., Rocca, F. E., Provenzano, G., Civitelli, D., et al. (2008). Glucocerebrosidase gene mutations are associated with Parkinson's disease in Southern Italy. Mov. Disord. 23, 460–463. doi: 10.1002/mds.21892
Dos Santos, A. V., Pestana, C. P., Diniz, K. R., Campos, M., Abdalla-Carvalho, C. B., de Rosso, A. L., et al. (2010). Mutational analysis of GIGYF2, ATP13A2 and GBA genes in Brazilian patients with early-onset Parkinson's disease. Neurosci. Lett. 485, 121–124. doi: 10.1016/j.neulet.2010.08.083
Duran, R., Mencacci, N. E., Angeli, A. V., Shoai, M., Deas, E., Houlden, H., et al. (2013). The glucocerobrosidase E326K variant predisposes to Parkinson's disease, but does not cause Gaucher's disease. Mov. Disord. 28, 232–236. doi: 10.1002/mds.25248
Emelyanov, A., Boukina, T., Yakimovskii, A., Usenko, T., Drosdova, A., Zakharchuk, A., et al. (2012). Glucocerebrosidase gene mutations are associated with Parkinson's disease in Russia. Mov. Disord. 27, 158–159. doi: 10.1002/mds.23950
Gan-Or, Z., Giladi, N., Rozovski, U., Shifrin, C., Rosner, S., Gurevich, T., et al. (2008). Genotype-phenotype correlations between GBA mutations and Parkinson disease risk and onset. Neurology 70, 2277–2283. doi: 10.1212/01.wnl.0000304039.11891.29
González-Del Rincón Mde, L., Monroy Jaramillo, N., Suarez Martinez, A. I., Yescas Gomez, P., Boll Woehrlen, M. C., Lopez Lopez, M., et al. (2013). The L444P GBA mutation is associated with early-onset Parkinson's disease in Mexican Mestizos. Clin. Genet. 84, 386–387. doi: 10.1111/cge.12084
Grabowski, G. A. (1993). Gaucher disease. Enzymology, genetics, and treatment. Adv. Hum. Genet. 21, 377–441. doi: 10.1007/978-1-4615-3010-7_5
Guimarães Bde, C., Pereira, A. C., Rodrigues Fda, C., dos Santos, A. V., Campos, M. Jr., dos Santos, J. M., et al. (2012). Glucocerebrosidase N370S and L444P mutations as risk factors for Parkinson's disease in Brazilian patients. Parkinson. Relat. Disord. 18, 688–689. doi: 10.1016/j.parkreldis.2011.11.028
Guo, J. F., Li, K., Yu, R. L., Sun, Q. Y., Wang, L., Yao, L. Y., et al. (2015). Polygenic determinants of Parkinson's disease in a Chinese population. Neurobiol. Aging 36, 1765.e1–1765.e6. doi: 10.1016/j.neurobiolaging.2014.12.030
Gutti, U., Fung, H. C., Hruska, K. S., Lamarca, M. E., Chen, C. M., Wu, Y. R., et al. (2008). The need for appropriate genotyping strategies for glucocerebrosidase mutations in cohorts with Parkinson disease. Arch. Neurol. 65, 850–851. doi: 10.1001/archneur.65.6.850
Han, F., Grimes, D. A., Li, F., Wang, T., Yu, Z., Song, N., et al. (2016). Mutations in the glucocerebrosidase gene are common in patients with Parkinson's disease from Eastern Canada. Int. J. Neurosci. 126, 415–421. doi: 10.3109/00207454.2015.1023436
Hu, F. Y., Xi, J., Guo, J., Yu, L. H., Liu, L., He, X. H., et al. (2010). Association of the glucocerebrosidase N370S allele with Parkinson's disease in two separate Chinese Han populations of mainland China. Eur. J. Neurol. 17, 1476–1478. doi: 10.1111/j.1468-1331.2010.03097.x
Huang, C. L., Wu-Chou, Y. H., Lai, S. C., Chang, H. C., Yeh, T. H., Weng, Y. H., et al. (2011). Contribution of glucocerebrosidase mutation in a large cohort of sporadic Parkinson's disease in Taiwan. Eur. J. Neurol. 18, 1227–1232. doi: 10.1111/j.1468-1331.2011.03362.x
Jesús, S., Huertas, I., Bernal-Bernal, I., Bonilla-Toribio, M., Caceres-Redondo, M. T., Vargas-Gonzalez, L., et al. (2016). GBA variants influence motor and non-motor features of Parkinson's disease. PLoS ONE 11:e0167749. doi: 10.1371/journal.pone.0167749
Kalia, L. V., and Lang, A. E. (2015). Parkinson's disease. Lancet 386, 896–912. doi: 10.1016/s0140-6736(14)61393-3
Kalinderi, K., Bostantjopoulou, S., Paisan-Ruiz, C., Katsarou, Z., Hardy, J., and Fidani, L. (2009). Complete screening for glucocerebrosidase mutations in Parkinson disease patients from Greece. Neurosci. Lett. 452, 87–89. doi: 10.1016/j.neulet.2009.01.029
Kumar, K. R., Ramirez, A., Gobel, A., Kresojevic, N., Svetel, M., Lohmann, K., et al. (2013). Glucocerebrosidase mutations in a Serbian Parkinson's disease population. Eur. J. Neurol. 20, 402–405. doi: 10.1111/j.1468-1331.2012.03817.x
Lesage, S., Anheim, M., Condroyer, C., Pollak, P., Durif, F., Dupuits, C., et al. (2011a). Large-scale screening of the Gaucher's disease-related glucocerebrosidase gene in Europeans with Parkinson's disease. Hum. Mol. Genet. 20, 202–210. doi: 10.1093/hmg/ddq454
Lesage, S., Condroyer, C., Hecham, N., Anheim, M., Belarbi, S., Lohman, E., et al. (2011b). Mutations in the glucocerebrosidase gene confer a risk for Parkinson disease in North Africa. Neurology 76, 301–303. doi: 10.1212/WNL.0b013e318207b01e
Li, Y., Sekine, T., Funayama, M., Li, L., Yoshino, H., Nishioka, K., et al. (2014). Clinicogenetic study of GBA mutations in patients with familial Parkinson's disease. Neurobiol Aging 35, 935.e3–935.e8. doi: 10.1016/j.neurobiolaging.2013.09.019
Lwin, A., Orvisky, E., Goker-Alpan, O., Lamarca, M. E., and Sidransky, E. (2004). Glucocerebrosidase mutations in subjects with parkinsonism. Mol. Genet. Metab. 81, 70–73. doi: 10.1016/j.ymgme.2003.11.004
Mao, X., Wang, T., Peng, R., Chang, X., Li, N., Gu, Y., et al. (2013). Mutations in GBA and risk of Parkinson's disease: a meta-analysis based on 25 case-control studies. Neurol. Res. 35, 873–878. doi: 10.1179/1743132813y.0000000224
Mao, X. Y., Burgunder, J. M., Zhang, Z. J., An, X. K., Zhang, J. H., Yang, Y., et al. (2010). Association between GBA L444P mutation and sporadic Parkinson's disease from Mainland China. Neurosci. Lett. 469, 256–259. doi: 10.1016/j.neulet.2009.12.007
Mata, I. F., Samii, A., Schneer, S. H., Roberts, J. W., Griffith, A., Leis, B. C., et al. (2008). Glucocerebrosidase gene mutations: a risk factor for Lewy body disorders. Arch. Neurol. 65, 379–382. doi: 10.1001/archneurol.2007.68
Mitsui, J., Mizuta, I., Toyoda, A., Ashida, R., Takahashi, Y., Goto, J., et al. (2009). Mutations for Gaucher disease confer high susceptibility to Parkinson disease. Arch. Neurol. 66, 571–576. doi: 10.1001/archneurol.2009.72
Moraitou, M., Hadjigeorgiou, G., Monopolis, I., Dardiotis, E., Bozi, M., Vassilatis, D., et al. (2011). beta-Glucocerebrosidase gene mutations in two cohorts of Greek patients with sporadic Parkinson's disease. Mol. Genet. Metab. 104, 149–152. doi: 10.1016/j.ymgme.2011.06.015
Nalls, M. A., Duran, R., Lopez, G., Kurzawa-Akanbi, M., McKeith, I. G., Chinnery, P. F., et al. (2013). A multicenter study of glucocerebrosidase mutations in dementia with Lewy bodies. JAMA Neurol. 70, 727–735. doi: 10.1001/jamaneurol.2013.1925
Neumann, J., Bras, J., Deas, E., O'sullivan, S. S., Parkkinen, L., Lachmann, R. H., et al. (2009). Glucocerebrosidase mutations in clinical and pathologically proven Parkinson's disease. Brain 132, 1783–1794. doi: 10.1093/brain/awp044
Nichols, W. C., Pankratz, N., Marek, D. K., Pauciulo, M. W., Elsaesser, V. E., Halter, C. A., et al. (2009). Mutations in GBA are associated with familial Parkinson disease susceptibility and age at onset. Neurology 72, 310–316. doi: 10.1212/01.wnl.0000327823.81237.d1
Nishioka, K., Vilarino-Guell, C., Cobb, S. A., Kachergus, J. M., Ross, O. A., Wider, C., et al. (2010). Glucocerebrosidase mutations are not a common risk factor for Parkinson disease in North Africa. Neurosci. Lett. 477, 57–60. doi: 10.1016/j.neulet.2009.11.066
O'Regan, G., deSouza, R. M., Balestrino, R., and Schapira, A. H. (2017). Glucocerebrosidase mutations in Parkinson Disease. J. Parkinsons. Dis. 7, 411–422. doi: 10.3233/jpd-171092
Ortega, R. A., Torres, P. A., Swan, M., Nichols, W., Boschung, S., Raymond, D., et al. (2016). Glucocerebrosidase enzyme activity in GBA mutation Parkinson's disease. J. Clin. Neurosci. 28, 185–186. doi: 10.1016/j.jocn.2015.12.004
Pulkes, T., Choubtum, L., Chitphuk, S., Thakkinstian, A., Pongpakdee, S., Kulkantrakorn, K., et al. (2014). Glucocerebrosidase mutations in Thai patients with Parkinson's disease. Parkinson. Relat. Disord. 20, 986–991. doi: 10.1016/j.parkreldis.2014.06.007
Ran, C., Brodin, L., Forsgren, L., Westerlund, M., Ramezani, M., Gellhaar, S., et al. (2016). Strong association between glucocerebrosidase mutations and Parkinson's disease in Sweden. Neurobiol. Aging 45, 212.e215–212.e211. doi: 10.1016/j.neurobiolaging.2016.04.022
Risch, N., Burchard, E., Ziv, E., and Tang, H. (2002). Categorization of humans in biomedical research: genes, race and disease. Genome Biol. 3:comment2007.1-comment2007.12.
Setó-Salvia, N., Pagonabarraga, J., Houlden, H., Pascual-Sedano, B., Dols-Icardo, O., Tucci, A., et al. (2012). Glucocerebrosidase mutations confer a greater risk of dementia during Parkinson's disease course. Mov. Disord. 27, 393–399. doi: 10.1002/mds.24045
Sidransky, E., Nalls, M. A., Aasly, J. O., Aharon-Peretz, J., Annesi, G., Barbosa, E. R., et al. (2009). Multicenter analysis of glucocerebrosidase mutations in Parkinson's disease. N. Engl. J. Med. 361, 1651–1661. doi: 10.1056/NEJMoa0901281
Spitz, M., Rozenberg, R., Pereira Lda, V., and Reis Barbosa, E. (2008). Association between Parkinson's disease and glucocerebrosidase mutations in Brazil. Parkinson. Relat. Disord. 14, 58–62. doi: 10.1016/j.parkreldis.2007.06.010
Stang, A. (2010). Critical evaluation of the Newcastle-Ottawa scale for the assessment of the quality of nonrandomized studies in meta-analyses. Eur. J. Epidemiol. 25, 603–605. doi: 10.1007/s10654-010-9491-z
Sun, Q. Y., Guo, J. F., Wang, L., Yu, R. H., Zuo, X., Yao, L. Y., et al. (2010). Glucocerebrosidase gene L444P mutation is a risk factor for Parkinson's disease in Chinese population. Mov. Disord. 25, 1005–1011. doi: 10.1002/mds.23009
Tan, E. K., Tong, J., Fook-Chong, S., Yih, Y., Wong, M. C., Pavanni, R., et al. (2007). Glucocerebrosidase mutations and risk of Parkinson disease in Chinese patients. Arch. Neurol. 64, 1056–1058. doi: 10.1001/archneur.64.7.1056
Toft, M., Pielsticker, L., Ross, O. A., Aasly, J. O., and Farrer, M. J. (2006). Glucocerebrosidase gene mutations and Parkinson disease in the Norwegian population. Neurology 66, 415–417. doi: 10.1212/01.wnl.0000196492.80676.7c
Török, R., Zádori, D., Török, N., Csility, É., Vécsei, L., and Klivényi, P. (2016). An assessment of the frequency of mutations in the GBA and VPS35 genes in Hungarian patients with sporadic Parkinson's disease. Neurosci. Lett. 610, 135–138. doi: 10.1016/j.neulet.2015.11.001
Wang, Y., Liu, L., Xiong, J., Zhang, X., Chen, Z., Yu, L., et al. (2012). Glucocerebrosidase L444P mutation confers genetic risk for Parkinson's disease in central China. Behav. Brain Funct. 8, 57. doi: 10.1186/1744-9081-8-57
Westbroek, W., Gustafson, A. M., and Sidransky, E. (2011). Exploring the link between glucocerebrosidase mutations and parkinsonism. Trends Mol. Med. 17, 485–493. doi: 10.1016/j.molmed.2011.05.003
Wu, Y. R., Chen, C. M., Chao, C. Y., Ro, L. S., Lyu, R. K., Chang, K. H., et al. (2007). Glucocerebrosidase gene mutation is a risk factor for early onset of Parkinson disease among Taiwanese. J. Neurol. Neurosurg. Psychiatr. 78, 977–979. doi: 10.1136/jnnp.2006.105940
Yu, Z., Wang, T., Xu, J., Wang, W., Wang, G., Chen, C., et al. (2015). Mutations in the glucocerebrosidase gene are responsible for Chinese patients with Parkinson's disease. J. Hum. Genet. 60, 85–90. doi: 10.1038/jhg.2014.110
Zhang, X., Bao, Q. Q., Zhuang, X. S., Gan, S. R., Zhao, D., Liu, Y., et al. (2012). Association of common variants in the glucocerebrosidase gene with high susceptibility to Parkinson's disease among Chinese. Chin. J. Physiol. 55, 398–404. doi: 10.4077/cjp.2011.amm076
Zhang, Y., Sun, Q. Y., Zhao, Y. W., Shu, L., Guo, J. F., Xu, Q., et al. (2015). Effect of GBA mutations on phenotype of Parkinson's disease: a study on Chinese population and a meta-analysis. Parkinson. Dis. 2015:916971. doi: 10.1155/2015/916971
Zhao, F., Bi, L., Wang, W., Wu, X., Li, Y., Gong, F., et al. (2016). Mutations of glucocerebrosidase gene and susceptibility to Parkinson's disease: an updated meta-analysis in a European population. Neuroscience 320, 239–246. doi: 10.1016/j.neuroscience.2016.02.007
Keywords: Parkinson's disease, GBA, AJ, non-AJ, meta-analysis
Citation: Zhang Y, Shu L, Sun Q, Zhou X, Pan H, Guo J and Tang B (2018) Integrated Genetic Analysis of Racial Differences of Common GBA Variants in Parkinson's Disease: A Meta-Analysis. Front. Mol. Neurosci. 11:43. doi: 10.3389/fnmol.2018.00043
Received: 07 November 2017; Accepted: 01 February 2018;
Published: 15 February 2018.
Edited by:
Jean-Marc Taymans, Institut National de la Santé et de la Recherche Médicale (INSERM), FranceReviewed by:
Sulev Kõks, University of Tartu, EstoniaBruno A. Benitez, Washington University in St. Louis, United States
Copyright © 2018 Zhang, Shu, Sun, Zhou, Pan, Guo and Tang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Beisha Tang, YnN0YW5nNzM5OEAxNjMuY29t
†These authors have contributed equally to this work and co-first authors.